
Submitted:
26 November 2024
Posted:
27 November 2024
You are already at the latest version
Abstract
This work has three main goals. The first is to present a broad historical review of the background of the international research activities dealing with fisheries and climate change, mainly in European waters. Marine environmental variability and climate changes are interconnected, being these, the main causes that determine the fluctuations in ecosystems and cyclically affect fisheries. We have recovered a selection of seminal international scientific publications from 1914-1995, which aroused great interest among the scientific community of that time, although most of these publications fall into oblivion in the 21st century. The second goal is to review the main intergovernmental impulses on climate and marine research from the 1980s to the present, detecting gaps and the lack of unanimity in some guidelines of international organizations. The third is to select proposals for the future, which include the need to promote an international retrospective on fisheries oceanography research, for at least the last hundred years. We also analyze decadal warming/cooling in the Canary Current Upwelling System (extending from NW Iberian Peninsula to Senegal), which may help understand the current rapid tropicalization of pelagic and benthic ecosystems in South European Atlantic seas.
Keywords:
climate change
; fisheries
; ecosystems
; indicator species
; species turnover
; oceanography
; historical research
; anthropogenic warming
; natural warming
; solar variability
1. Introduction
“Knowing the past is vital for developing a vision of the future. The oceans and inshore seascapes of the world are rapidly changing, and understanding the human and marine ecosystem forces, trajectories and responses—sometimes over centuries or millennia— is vital for their informed management” (Ocean Past Initiative, 2018[1]). Therefore, to better understand the future of unexploited marine populations and fisheries in a given region, it is necessary to study their fluctuations in the past (eg. Pitcher, 2001[2]; Gordon et al, 2018[3]; Cooke et al., 2023[4]), preferably during the last 100 years. This is essential to develop a more realistic vision of the future than that based on mathematical models generated with short time series.
Furthermore, documenting the past is necessary to attempt the reconstruction of historical environmental scenarios in meteorology and oceanography, as well as to document the evolution of human interaction with fisheries and marine ecosystems. Following the example of multidisciplinary research efforts that have been carried out internationally during the last decades, aimed at improving the knowledge and understanding of the interaction, from the earliest times between man and the marine environment and, also, the historical variability in the coastal and open sea ecology (Holm et al., 2001[5]; 2010[6]; Starkey et al., 2008[7]; Lotze & McClenachan, 2013[8]). In this contest, the work of the ICES Working Group on the History of Fish and Fisheries (WGHIST): “a forum for interdisciplinary research on social-ecological change in marine and fisheries systems over multi-decadal to centennial and even millennial timescales” (ICES, 2018[9]) is newsworthy. For the above reason the search for and recovery of data series and old scientific literature on specific marine regions and areas has been intensified. This will allow to recognize the historical changes that have occurred in marine populations and submerged ecosystems, derived from cyclical fluctuations in the environment (oceanography and meteorology) and/or increased human pressure on renewable resources (eg. Southward et al. 2005[10], Poulsen et al., 2007[11]; Philippart et al., 2011[12]; Malta et al., 2016[13]; Van Beveren et al., 2016[14]; Fortibuoni et al., 2017[15]; Hernvan & Gascuel, 2019[16]; Sánchez-Garrido et al., 2019[17]; Pérez-Rubín, 2021[18]).
For all these reasons, it would be advisable that the current scales of temporal change of interest to a contemporary fishery or marine biologists (from seasonal, inter-annual variability to decadal changes), should be extended across the centuries of the Anthropocene from c. 1800 (e.g. Thurstan R. H. 2022[19]), looking for connections/relationships between physical variabilities and fisheries, also documented in the long-term temporal scale. Even the research on a geological time-scale (from thousands to million years) and the study of ancient fish remains, an aid in the reconstruction of past marine environments would be advisable (e.g. Finney et al., 2010[20], Barrett, 2019[21]).
It is significant to highlight two shortcomings in the 21st century publications examined changes on climate and marine resources in European waters. On one hand, the recovery of historical information, generally does not begin before 1945-1950, as this is the period for which the most accessible oceanographical and meteorological databases are available (AMO, NAO, EA, SST, FAO fishery statics…). On the other hand, we observe an almost generalized forgetting of the seminal international scientific publications from the period 1914-1995. The information and results provided by these publications can be of great interest and they are an extraordinary source of knowledge, that can provide insights and perspectives, to differentiate between natural environmental/climate variabilities and human-induced changes in the ecosystems and, therefore, can potentially contribute to develop a more realistic vision of the future of different fish and fisheries. Furthermore, in the literature consulted the most frequent situation is that fisheries scientists maintain two methodological biases. In the first place, they mostly continue to accept as baseline references the size of the target population and the composition of species that they knew at the beginning of their respective research careers, and use these specific statuses to evaluate coming changes, errors that were already noticed almost 30 years ago (Pauly, 1995[22]; Pinnegar & Engelhard, 2008[23]; Bonebrake et al., 2010[24]; Soga & Gaston, 2018[25]). And in the second place, researchers are generally interested exclusively in contemporary publications and ignore publications from previous decades.
Therefore, in order to better understand the current situation in relation to the research on fishery science vs climate change and develop a documented proposal for the future, it is necessary to first carry out an extensive review international multidisciplinary bibliography of the period 1914-1995, selecting a sample of seminal publications on the influence of climate variability on regional and global fisheries and marine ecosystems. It is also necessary to review and describe guidelines arising from intergovernmental agreements since the early 1990s, when global research on climate change in the seas and oceans began to be promoted. Although since the 1980s there have been important intergovernmental efforts to promote global meteorological research on climate change, essential global research on the ocean (oceanography, fisheries and ecosystems) was not implemented until the period 1991-1995. Since then, much progress has been made in understanding global long-term physical fluctuations in the atmosphere and ocean, but comparatively less on long-term biological changes in European seas.
1.1. International Impulses for Climate and Marine Research Since the 1980s
In 1979, during the first World Climate Conference, climate change was identified as an urgent global problem and a call was made to governments to address this challenge. The great influence of solar, volcanic and CO2 forcing on recent climatic changes was demonstrated (Gilliland, 1982[26]). Also studies on the relationship of the increase in CO2 content in the atmosphere with the warming caused by climate change were promoted internationally (Carbon Dioxide Review, 1982[27]), including a preferential interest in the coastal regions (especially impacted by a sea level rise) and their fisheries. To the assessment and management of coastal marine resources and their environments, at an international symposium of the American Association for the Advancement of Science (1984), the Large Marine Ecosystem (LME) approach, based on linked ecological criteria, was first introduced “as place-based areas around the globe, to focus the methods of marine science, policy, law, economics and governance on a common strategy for assessing, managing, recovering, and sustaining marine resources and their environments” (Sherman & Hempel, 2008[28]).
The action of the World Meteorological Organization (WMO) during the 1980s was decisive in promoting research on the world climate and climate change on a global scale. The first impulse of the WMO was its sponsorship (together with that of the International Science Council) to establish the World Climate Research Program (WCRP) in 1980. WCRP implemented interest in the ocean in 1993 with the incorporation of the Intergovernmental Oceanographic Commission (IOC) of UNESCO as a sponsor, and in 1995 it began to develop one of the six core projects of the WCRP: Climate and Ocean: Variability, Predictability, and Change (CLIVAR), whose mission is “to understand the dynamics, the interaction, and the predictability of the climate system with emphasis on ocean-atmosphere interactions”[29]. The second decisive impulse of the WMO, along with the essential participation of the United Nations Environment Program (UNEP), has been the creation in 1988 of the Intergovernmental Panel on Climatic Change (IPCC), the leading international body for assessment of climate change. It objective “is to provide governments at all levels with scientific information that they can use to develop climate policies”[30]. In 1987 the Scientific Committee for Oceanic Research (SCOR) initiated the Joint Global Ocean Flux Study (JGOFS), an international research programme on the fluxes of carbon between the atmosphere and ocean. The International Geosphere-Biosphere Programme (IGBP), turn on in 1987 with its primary focus on: “coordinating international research on global-scale and regional-scale interactions between Earth's biological, chemical and physical processes and their interactions with human systems”[31]. The United Nations Framework Convention on Climate Change (UNFCCC) was adopted in 1992 and entered into force in 1994.
The period 1991-1995 was decisive in promoting research on climate change in the aquatic environment and the implementation of oceanographic studies in addition to atmospheric studies. In 1991 the Working Group II of the IPCC, speculated about the possible climate change impacts on fish population and aquatic life, and the following year the Global Environment Monitoring System (GEMS) of UNEP sponsored the publication of the book: Climate variability, climate change and fisheries (Glanz, 1992[32]). This book “addresses the potential implications for fisheries and societies of the regional impacts of a global warming of the atmosphere.” Another very important initiatives were generated in 1995, when the fisheries were included in the IPPC’s 2nd Assessment Report (Everett et al., 1996[33]). It was concluded that “Climate-change impacts are likely to exacerbate existing stresses on fish stocks, notably overfishing, diminishing wetlands and nursery areas, pollution, and UV-B radiation […]. The question of whether overfishing, environmental change, or a combination of the two is responsible for major declines in fish stocks is still a matter for debate and is situation-specific”. Simultaneously, the research program of Global Ocean Ecosystems Dynamics (GLOBEC) was supported by IOC-UNESCO and SCOR, and the first GLOBEC collaboration with the European Atlantic began through the International Council for the Exploration of the Sea (ICES), with a working group on cod and climate change. For the Mediterranean Sea, the year 1995 was also very important, when under the auspices of UNEP/Mediterranean Action Plan (MAP), the Convention for the Protection of the Marine Environment and the Coastal Region of the Mediterranean (Barcelona Convention) was adopted[34].
To European-scale operational oceanography, within the context of the intergovernmental Global Ocean Observing System (GOOS), the international non-profit association EuroGOOS[35] was founded in 1994 and operates in five regional sea areas including the Ireland-Biscay-Iberian area (IBIROOS) and the Mediterranean Sea (MonGOOS). In 1995 the Marine Board, a pan-European platform, was established to facilitate enhanced cooperation between its member organizations to develop a common vision on the research priorities and strategies for advancing marine science in Europe[36].
During 1998, the Year of the Oceans a significant amount of international attention focused on the ocean's role in climate maintenance and change, ocean prediction, and environmental sustainability in the coastal zone emerged. The following year, in 1999, the international ARGO program began to be developed[37]. This program has a set of autonomous profiling floats with sensors for real-time measurement of temperature and salinity through the upper 2000 m of the global ocean, with the aim of contributing to a global description of the seasonal cycle and interannual variability of the upper ocean thermohaline circulation.
Likewise, the agreements of the Kyoto Protocol (in force since 2005) are necessary and complementary, marking the commitments to reduce emissions of greenhouse gases due to their negative influences on climate change. Impacts of climate changes have been observed in marine environments including sea temperature and sea-level rising, ocean acidification, deoxygenation and extreme weather events that imperil ecosystems, economies, and societies worldwide (IPCC, 2023[38]). Pending tasks include extending the record of climate variability to broader time scales, optimizing the reliability of climate predictions, and advancing the prediction of the climate system's response to increases in the concentration of greenhouse gases and aerosols (CLIVAR, 2019[39]). Equally necessary is to understand the multifactorial sun-climate connection. With this objective the National Research Council (NRC, United States) convened in 2011 a workshop, with the interaction of numerous experts (eg. in terrestrial history, solar activity, atmospheric chemistry and fluid dynamics, physics of plasma and energetic particles), to discuss how the Sun's variability over time has affected Earth's climate. They “emphasized that there may be an emerging pattern of paleoclimate change coincident with periods of solar activity and inactivity, but only on long timescales of multiple decades to millennia” (NRC, 2012[40]). In this line, a national research project conducted in Japan to understand the structure and dynamics of the solar–terrestrial environment and to predict the solar variations, was compiled recently (Kusano, 2023[41]), highlighting the need to study the potential risks associated with “space weather” (“such variations in the space environment, which can influence human activities”).
Directly related to the oceans, several sustainable development goals were presented to the United Nations General Assembly in 2014, including the goal to take urgent action to combat climate change and its effects, recognizing the need to “strengthen resilience and the capacity to adapt to climate-related risks and natural disasters in all countries”. In 2015, in the context of the UNFCCC, the need to ensure the integrity of marine ecosystems and advance the understanding of the links between climate and the ocean was recognized. Simultaneously, the 2015 Paris Agreement recognized the need for effective and progressive responses to the urgent threat of climate change, while taking into account the particular vulnerabilities of food production systems. Due to the great nutritional importance of the fishing and aquaculture sector, FAO agreed to have its first workshop of technical experts in Rome (2017). With the conclusions adopted the following year, FAO published the report entitled Impacts of climate change on fisheries and aquaculture. Synthesis of current knowledge, adaptation and mitigation options (Barange et al. 2018[42]). The main objective of this report was “provides the most up-to-date information on the disaggregated impacts of climate change for marine and inland fisheries, and aquaculture, in the context of poverty alleviation and the differential dependency of countries on fish and fishery resources”.
In 2016, the IPCC committed to preparing a special report on the Ocean and Cryosphere in a Changing Climate, which was published three years later (IPCC, 2019[43]). This report included the chapter Changing Ocean, Marine Ecosystems, and Dependent Communities (Arístegui et. al., 2019[44]). At local to regional scales, in 2022 it was confirmed that “climate change worsens the impacts on marine life of non-climate anthropogenic drivers, such as habitat degradation, marine pollution, overfishing and overharvesting, [extreme] nutrient enrichment [eutrophication] and introduction of non-indigenous species” (Cooley et al., 2022[45]). In a complementary way, the world ocean and coasts increase their prominence in the GEMS for the Ocean and Coasts Program (GEMS-Ocean 2022[46]). This allows the scientific community access to global data and assessments on the state of the oceans and coasts with a view to supporting well-informed decision-making, which can be scaled up to drive consensual transformations at national, regional and global levels.
Promoted by the European Union, the Copernicus Marine Environment Monitoring Service (CMEMS, or Copernicus Marine Service)[] was implemented in 2014 by Mercator Ocean International (MOi). CMEMS provides regular and systematic information on the sea ice state and on the physical and biogeochemical ocean processes, on differents scales (global, regional or zonal), in real-time or delayed mode. IOC-UNESCO selected MOi to lead the Ocean Prediction Decade Collaborative Centre in the context of the United Nations Decade of Ocean Science for Sustainable Development or UN Ocean Science Decade[47]. In 2022, six European countries (France, Spain, Portugal, Italy, UK and Norway) signed the “Declaration of Brest” and committed to transform MOi into a Europe-centred Intergovernmental Organisation and invite European coastal countries to join. All these initiatives are completed with the Copernicus Climate Change Service (C3S[48]), which “provides climate monitoring for the globe, Europe and the Arctic. Authoritative information about the past, present and future climate, as well as tools to enable climate change mitigation and adaptation strategies, policy makers and businesses”. C3S together with the WMO both have released a report on the European State of the Climate (ESOTC, 2023[49]). Along with detailed documentation on different marine variables: Sea Surface Temperature (anomalies, ranking), Marine Heatwaves (in Northeastern Atlantic and Western Mediterranean) and Ocean colour (anomalies in chlorophyll a concentration)[50].
Currently UNDOS, with the mission of ensuring sustainable ocean development using transformative ocean science, includes the sustainable development Goal 14 “Life Below Water” and has recently implemented its Challenge 5 (Unlock ocean-based solutions to climate change), “with the goal to enhance understanding of the ocean-climate nexus and to generate knowledge and solutions to mitigate, adapt and build resilience to the effects of climate change across all geographies and at all scales, and to improve services including predictions for the ocean, climate, and weather” (IOC-UNESCO 2024[51]). Numerous programs were developed, focused on observations and co-design of observations, reducing stressors (sustainable fisheries, sound, pH, oxygen and others), emissions management, blue carbon ecosystems and forecasting/prediction. These specific actions are coordinated by it Decade Coordination Unit and published as part of UNESCO's Ocean Decade Series of publications (eg. UNESCO-IOC, 2021[52], 2023[53]). In April 2024, thousands of participants (in-person and virtual) gathered for the 2024 Ocean Decade Conference in Barcelona, which was hosted by Spain and co-organized by IOC/UNESCO. The global state of ocean science has previously been summarized in several publications (UNESCO, 2017[54]; IOC-UNESCO, 2020[55]). A exhaustive review (Potter & Pearson, 2023[56]) analyzed trends in ocean research, specific to the five ocean basins of the world.
In Atlantic African waters, the Canary Current Large Marine Ecosystem (CCLME) project is currently executed by FAO and UNEP, including Spanish waters (Canary Island) and the different coastal African countries from Morocco to Guinea. In that CCLME, IOC-UNESCO implementing the subproject Invasive alien species and other ocean stressors: Furthering the scientific knowledge and capacity basis (see 3.3), which is funded by the Spanish Agency for International Development Cooperation (AECID) and implemented in collaboration with the Spanish Institute of Oceanography[57].
We conclude with the particular case of marine research development in the Mediterranean Sea, a region that faces a growing increase of human activities and an incessant pace of ecosystem transformation. To protect its particular biodiversity, the Strategic Action Program for the Conservation of BIOlogical diversity (SAPBIO) has been proposed and adopted in 2003, posteriorly updated between 2008-2009 on climate change issue. The work of the Mediterranean Science Commission (CIESM) has also been essential. CIESM develops, in the current context of global warming, several research programs initiated since 2002 and its publications on this marginal sea (Schroeder et al., 2010[58]; 2013[59]; 2016[60]; 2017[61]). It also supported the Hydrochanges Program initiated in 2002 to monitor the long-term variability of hydrographic properties of the Mediterranean Sea water masses, including in situ measurements of temperature and salinity of deep waters in four northwestern key areas: Strait of Gibraltar area, Gulf of Lion, Catalan Sea and Ibiza Chanel. Among others, changes in the NW Mediterranean deep water due to the exceptional winter of 2005 (anomalously cold and windly), the heterogeneities of in- and out-flows in the Strait of Gibraltar, the salinification of both flows and about the seasonal and fortnightly variabilities of the Mediterranean out-flows. Among the CIESM's biological research related to climate warming, we highlight a monograph on the related changes in Mediterranean marine biota (CIESM, 2008[62]) and the development of the CIESM Tropical Signals Programme[63]. Those programs study the effects of tropicalization of the Mediterranean Sea, the geographic expansion of native and alien warm-water species and the contraction of native cold-water species, shifts in species depth distribution, intermittent episodes of mass mortalities of marine organisms, etc. With the voluminous documentation generated in the last decade, the CIESM Atlas of Exotic Species in the Mediterranean[64] has been published. It consists of 4 volumes: Fishes (Golani et al., 2021[65]), Crustaceans (Galil, Froglia & Noël, 2002[66]), Molluscs (Zenetos et al, 2003[67]) and Seaweeds (Verlaque et al, 2015[68]). Three main origins of exotic species are distinguished in the Mediterranean: those from the Indo-Pacific (entered through the Suez Canal), those from the tropical Atlantic (entered via the Strait of Gibraltar) and those accidentally introduced by man.
2. Material and Methods
We have extracted our historical scientific references mainly from old publications (papers, books and contributions to symposiums) until 1995. For the subsequent years until 2010, an overview of the literature on the impact of climate change and variability of fish and shellfish populations in the northeast Atlantic Ocean, the Mediterranean and Black seas was included in an ICES report (Rijnsdorp et al., 2010[69]). This report includes a chapter that reviews the knowledge of past climate variability and future climate change in European waters (Drinkwater, 2010[70]), and another with an update of the available information on spatio-temporal changes in fish species distribution (Pinnegar et al., 2010[71]), mainly since the 1970s-1980s. Much documentation has also been generated in the five symposiums devoted to the study of the Effects of Climate Change on the World's Sea, carried out during 2008-2023 (Spain, Korea, Brazil, USA, Norway). These symposiums were mainly sponsored by ICES, IOC and PICES (North Pacific Marine Science Organization). From that last year, above mentioned, there is a valuable review that examines the literature on the influence/relationship of climate on 68 species of great importance in European seas. However, not enough literature was found for most of these species. (Predragovic et al., 2024[72]). Another very important source of documentation are the international conferences on paleoceanography, with a tradition of 40 years (14 conferences during 1983-2022), focused on “reconstructing and simulating the history of the ocean and its role in climate change” (Meckler et al., 2023[73]).
More historical documentation from several European archives (maritime climate, oceanography, fisheries and plankton) is also available on the internet. For maritime climate history research, two interrelated projects are recommended. The Climatological Database for the World's Oceans (CLIWOC[74]), funded by the European Union, can be used to “connecting past climate change to human affairs”. It consists of 287,114 logbooks written aboard Spanish and other countries' sailing ships (mostly between 1750 and 1850), on a global scale. It can be complemented by the Past Global Changes (PAGES) Project, which “provides support for the gathering and synthesis of observations, reconstructions, and the modeling of climate, ecosystem, environmental and societal dynamics in the past”[75]. WISE-Marine, founded by the European Environment Agency (EEA), is a gateway to information on European marine issues, in support of the ecosystem-based management and ocean governance[76]. This program periodically updates the European Marine Climate Change Index (EMCCI[77]), which provides a graphic indication (maps) of the relative magnitude of the effects of climate change stressors on marine ecosystems. In addition, the Marine Board has published synthesis of European research on the effects of climate change on marine environments (Heip et al., 2011[78]). Also, the European Centre for Medium-Range Weather Forecasts (ECMWF[79]), a key player in Copernicus Marine Service (CMEMS), produce global numerical weather predictions, annual averages of sea surface temperatures (see Figure 1), monthly anomalies surface air temperature (range -6ºC to 6 ºC). Other maps generated by the EEA show, as biological indicators, the changes in fish distribution in Europe's seas during 1982/2022 (see a sample in Figure 2).
In the Mediterranean Science Commission, digital archive contains the scientific communications presented to its congresses since 1919[80]. Likewise, for the Northeast Atlantic, the database of the International Council for the Exploration of the Sea contains information since 1898[81], in addition to its publications on the fish landings in Atlantic Europe. FAO's world fishery landing statistics were revised for the period 1950-1994, globally and by oceans (Grainger & García, 1996[82]). The respective Yearbook of Fishery Statistics (catches and landings) are published annually. For retrospective research in marine ecology in the North Atlantic and North Sea it is recommendable to consult the largest multi-decadal plankton monitoring program in the world, operated by the Marine Biological Association (Plymouth, UK): the Continuous Plankton Recorder (CPR) database, on the abundance of phyto and zooplankton recorded monthly from 1958 to present[83] (Richardson et al., 2006[84]).
In Spain, searches for historical publications from the two oldest multidisciplinary oceanographic and fisheries research institutes in the country are available. The centenary Instituto Español de Oceanografía (IEO, Spanish Institute of Oceanography) since 1914[85] and the Instituto de Investigaciones Pesqueras (Fisheries Research Institute) from its foundation to its disintegration into independent institutes (1949-1979)[86]. For the period 1912-1970 the numerous publications, by three IEO researchers (from the Lozano family), on fish and fisheries in Spanish and African waters are of great interest (Pérez-Rubín, 2020[87]). The IEO maintains databases on its oceanographic surveys since 1914 and its tide gauges since the 1940s (Tel, 2023[88]), and a multidisciplinary monitoring program since 1992, carrying out periodic cruises along the coasts of the Iberian Peninsula, the Balearic archipelago and the Canary Islands (Valdés et al., 2002[89]). For the period 1950-1990, the IEO also documented 16 isoline maps for the surface marine waters of the Iberian Peninsula, showing the seasonal variation of average salinity and temperature (maximum, minimum and average) (IGN, 1991[90]).
On the other hand, the Spanish Agencia Estatal de Meteorología (National Meteorological Agency) has the institutional repository 'Arcimis' with publications since 1859[91]; and since 1981 climate maps of Spain and the Maritime Climate Atlas, which includes monthly maps of waves, winds and sea surface temperature[92]. IPCC Reports available since 1990[93] and CLIVAR-Spain reports since 2006[94].
On a global scale, with the enormous volume of international information generated in recent decades on the influence of climatic changes on seas and oceans, it is essential to consult various databases on academic research and on specific changes in marine biodiversity/biogeography. Updating peer-reviewed research literature requires consulting databases such as Web of Science (WoS) and SCOPUS (with more than 22,000 titles). On the other hand, the Ocean Biodiversity Information System (OBIS[95]) provides the world's largest scientific knowledge base on the diversity, distribution and abundance of nearly 120,000 marine species. Information provided by 500 institutions from 56 countries, and that can be integrated with physical and chemical environmental data.
3. Results and Discussion
3.1. Some Background to the International Study of Climate Changes and Fluctuations in Fisheries (1914-1995)
In the 21st century we are aware of the numerous irregular alternations of cold and warm periods of uncertain duration recorded throughout Earth’s history. There is a general consensus on the definition of climate changes: “is any systematic change in the long-term statistics of climate elements from one state to another where the new state is sustained over several decades or longer” (Drinkwater, 2010).
The Spanish ecologist and oceanographer Ramón Margalef (1919-2004) recognized in 1960 that the fluctuations of populations and ecosystems are periodic and are their “very essence”: “Ecosystems results from the integration of populations of different species in a common environment. They rarely remain steady for long, and fluctuations lie in the very essence of the ecosystems and of every one of the […] populations” (In: Smith, 1994[96]).
The transition from the end of the 19th century to the 20th presented dramatic alterations in the polar oceans that had abnormal effects in other latitudes, such as off the coast of New England with the exceptional intrusion of cold Subartic water that produced the almost total destruction of the tilefish (Lopholatilus chamaeleonticeps) in spring 1882 (natural mass mortality). According to Bigelow (1931[97]), the expansion of the ice at the South Pole was produced between 1892 and 1897 with “an outburst of the ice from the Antarctic, sending many floes and icebergs northward into the Southern Ocean.” At the antipodes a similar outburst occurred in 1901 with the Arctic ice, when the Barents Sea was full of pack ice up to May.
In international fisheries, since the end of the 19th century, the “impoverishment” of the sea was recognized (Garstang, 1900-1903[98]) and interest in the study of fisheries in fishing grounds shared by several nations intensified. In 1899 seven northern European countries met to plan research in the Glacial, Baltic and North Seas, an initiative that was institutionalized years later with the creation of the International Council for the Exploration of the Sea (ICES), in a context of social and scientific concern for the fluctuations in fisheries on decades time-scales and the overfishing.
Since the 1890s different European investigations were published intermittently on the spatio-temporal variability of the Gulf Stream current and various species of fish, such as the secular periodicity of the herring (Clupea harengus) fisheries, the news that fishermen exploited Icelandic waters all year round, the disappearance of the sardine (Sardina pilchardus) in the north of the Iberian Peninsula and its alarming scarcity on the coasts of France (its catches of over 50,000,000 kilos in 1898, sunk in 1902 to less than 9,000,000 kilos), with the surprising expansion of anchovy (Engraulis encrasicolus) towards the North reaching the coasts of Scotland. The pioneering publications of Scandinavian authors with the application of marine biological and environmental research to the study of Fluctuations in the great fisheries of Northern Europe (Hjort, 1914[99]), mainly the Norwegian cod (Gadus morhua) and herring fisheries, were a milestone 110 years ago; complemented by Climatic variations in historic and prehistoric time (Petterson, 1914[100]). This last author described a series of violent catastrophic and synchronous events with dramatic economic, political and social repercussions, which caused serious consequences for coastal communities, such as the lucrative Hanseatic herring fishery ceased altogether, frequent volcanic eruptions in Iceland and the surrounding seas, and violent storm-floods devastated the coast of the North Sea and Baltic.
The first comprehensive estimates of global mean temperature (in both hemispheres) between 1861-1984 were published, showing marked warming to 1940, relatively steady conditions to the mid-1970s and a subsequent rapid warming in the 1980s (Jones et al., 1986[101]). Glanz (1992) identified on a global scale a cyclical alternation of warm and cold decades, roughly: 1920s-1940s (warming), 1955-1965 (cooling), late 1960s (warming), and the 1982-1983 El Niño-Southern Oscillation (ENSO, cooling), which had global consequences. With our perspective from the 21st century (eg. Johannessen et al., 2004[102]; Hughes, 2012[103]; Hodson et al., 2014[104]), in general for the European Seas the patterns of sea temperature modified rapidly between 1900-1920 (cooled), 1925-1950s (warmed) and between 1960s-1970s (with two cooling phases: in 1962-1965 and 1970-1975), and since the 1980s warming has already affected global marine ecosystems (see 3.3). Between 1976-1977, a change occurred at a planetary level that could be observed in many series of data: oceanographic, atmospheric and biological (both in exploited populations and in those not subject to anthropogenic influence) (Lluch-Belda et al., 1999[105]). In the pelagic fish assemblage of the European continental shelf ichthyological/environmental changes were demonstrated: shifting from cold-water fish species (the 1960s-1980s) to warmer-water assemblages from the 1990s onwards (see 3.3.1). And also the collapse of the spawners bluefin tuna (Thunnus thynnus) fisheries of northern Europe during the 1960s, related to the overfishing of its juveniles since 1950 in the Bay of Biscay and Atlantic Morocco (Cort & Abaunza, 2015[106], 2019[107]). During approximately 1980-2005, sea surface temperature (SST) warmings were recorded in different regions for the NE Atlantic and Mediterranean (Philippart et al., 2007[108]; Belkin, 2009[109]; Nykjaer, 2009[110]). In the Northern Hemisphere this latest warming is considered greater than at any other time during the last 2000 years (Mann and Jones, 2003[111]; Moberg et al., 2005[112]).
In the World Ocean Large Marine Ecosystems (LMEs) during 1957–2006 several strong regional variations in the rate of sea surface temperature (SST) change has been confirmed (Belkin, 2009). For this author almost all LMEs in the North Atlantic and North Pacific experienced a prolonged cooling in the 1950s–1970s, followed by a rapid and accelerated global warming in 1982–2006 confined to the Subarctic Gyre (in this case likely caused by natural variability related to the NAO), European Seas and East Asian Seas (in both cases the rapid warming may in part be of direct anthropogenic origin). Bakun's proposal that climate change can intensify coastal ocean upwelling (Bakun, 1990[113]) could explain the simultaneous exceptional cooling recorded in the Eastern Pacific upwelling areas (the California Current and Humboldt Current), although it contradicts the situation detected in the coastal upwelling off Northwest Africa (see 3.3.3 and 4.).
It is necessary to advance in understanding the effects of past cyclical changes in sea temperatures on different marine ecosystems at a regional scale in the 20th century. During the warming/cooling phases of the period 1921-1971, ecosystem changes were detected in the English Channel off Plymouth (Russell et al., 1971[114]), with several episodes of alternation between herring and pilchard. In the Plymouth area during 1930-1968 changes were detected in the oceanic/neritic zooplanktonic community, with a first phase of local cooling of the surface water until 1938 (with the presence of arctic-boreal species) and its subsequent warming indicated by neritic-temperate species (Southward, 1984[115]). However in the northernmost northern hemisphere the mid 20th century warm period has been considered a broad 35-year period of warming (1925-1960), which we can correlate with the return of the cod to Greenland (Hansen, 1954[116]) and the relaunch of its very productive fishery. Approximately during that same warm period, the trophic migration of bluefin tuna to the Norwegian Sea and the North Sea was favored, and a large fishery species developed in that region (since the 1920s, with a peak in the 1950s), until its almost disappearance after 1967. This increase in the abundance of bluefin tuna was related to the arrival of the Atlantic saltier water and the abundance of herring and other small pelagics, while its decline at the end of the 1960s coincided with a period of cooling and the decline of Atlantic influence, causing the disappearance of its prey species (Binet & Leroy, 1986[117]). Other more recent authors also did not find direct evidence that the aforementioned increase in tuna was due to a habitat change related to the increase in temperature in the region (MacKenzie & Myers, 2009[118]). Forced variations (due to CO2, solar forcing and volcanism) were involved in this mid-20th century warm period and the temperature increase is comparable to the most recent period of pre-industrial climatic warming in the northern hemisphere: the Medieval Warm Period or now usually referred to as the Medieval Climate Anomaly, this one with three relatively short-lived warming intervals during 1010-1190 (Crowley, 2000[119]). Although it has been stated that the very strong early 20th century warming of the first decades in the Arctic and Greenland are not yet well understood (Hughes, 2012); other authors have associated the warming in the Arctic and along Greenland with the earlier increase in the Atlantic Multidecadal Oscillation (AMO climatic index) during 1920s-1930s (Drinkwater, 2009[120]; Polyakov, 2010[121]). However, it was shown (Yamanouchi, 2011[122]) that the significant warming of the 1920s-1940s in the Arctic is comparable to that of the 1980s-2010, although that was mainly concentrated at high latitudes and the current one at all latitudes.
In the first warming decades of the 1920s-1930s, marked faunal changes were also recorded in Iceland (Fridiksson, 1949[123]), which also affected lower latitudes, such as European waters (Johansen, 1926[124]) and the North American waters of California (Walford, 1931[125]) and Nova Scotia (McKenzie & Homans, 1938[126]). There was an immigration of different species of fishes of temperate thermal affinities into high latitudes, coinciding with an expansion of warm Atlantic water reported in the Barents Sea (Bigelow, 1931). It is suggested that all these are associated with a general warming up of the Atlantic and a greater northerly flow of Atlantic water of southern origin. However locally the low temperatures recorded at the beginning of the year 1929 in the North Sea produced more extreme biological changes, noted by bottom trawler fishermen, such as the large number of dead and decomposing fish caught, coinciding with an extraordinary drop in water temperature along the continental coast (below -1° C) and the arctic conditions that prevailed on the Continent (Lumby & Atkinson, 1929[127]). On the other hand, the consequences of the intense Atlantic warming in the 1930s and early 1940s were also temporally and geographically extensive, with the arrival of southern species to Iceland, Nova Scotia and into British waters. In the bibliography of those decades, in the arctic and subartic regions contemporary climatic, biological and geophysical changes were already detected from Greenland to Eurasia (Jensen, 1939[128]). At the W. English Channel, the first profound changes in ecosystems were documented in 5-year cycles: 1925-26, 1930-31 and 1934-35 (Cushing & Dickson, 1976a[129]). In the western European waters, coinciding with the unusual increase in atmospheric temperature during the years 1932-1934, “evidence that species of zooplankton and fishes usually found in more southern waters were entering British seas, and whales, dolphins and sharks were more than usually common” (Stephen, 1938[130]). Southern species also appeared in French waters in the early 1930s (Desbrosses, 1932[131], 1935[132], 1936[133]). In Spain, the intense sardine crisis of the period 1924-1928 (Buen, 1929[134]), was followed by the very productive years 1929-1938 for the sardine in Galician waters and at the same time the sprat decreased until its virtual disappearance in 1938 (Anadón, 1950[135]). For the Atlantic cod stocks, the different responses verified during the warming periods of the 1920s-1930s and 1990s-2000s, in particular the reduced cod production during the recent warm event, are mainly attributed to the effects of intense fishing pressure (Drinkwater, 2009).
In this context of great environmental and marine fauna variability during 1920s-1930s, several European initiatives were promoted. Since 1936, for species of commercial interest, the ICES collected comparative studies of the fluctuations in the stocks of principal food fishes in the seas of North and West Europe (ICES, 1936[136]), and the need to intensify multidisciplinary oceanographic studies (including more information on the environmental factors during the critical period of larval development) was recognized to understand the great fluctuations in the abundance and distribution of marine fauna (Kemp, 1938[137]). The influence of particular meteorological variables was demonstrated, such as the relevant wind conditions for herring and haddock (Carruthers, 1938[138]); and the especially cold winters of 1946-1947 in the southern North Sea, which caused a negative influence on plankton and certain species of fish, which suffered multiple changes in behavior or mortality (Simpson, 1953[139]; Woodhead, 1964[140]). After an ICES meeting in 1947, in which it was recognized that the problems of climatic change (in distribution, migrations, spawning condition and density of marine animals) is one of the most pressing problems in the area, an Inquiry into the Problem of Climatic and Ecological Changes in Northern Waters was drawn up by G. Rollefsen & Å. V. Tåning the following year (see 4.). Subsequently, in 1949, the first ICES's meeting on climate changes (in the Artic) was held (ICES, 1949[141]).
More articles and the influential book Climate through the ages. A study of the climatic factors and their variations were published (Brooks, 1949[142]), which demonstrated the cyclic changes in the oceanic circulation and in the conditions of the Atlantic, summarized the 11 theories of climatic changes described in the scientific literature until that year and delved into the reversal of deepsea circulation. In the 1950s, research on biological responses to climate changes was resumed with more intensity, with the publication of several articles on various species of fish, both pelagic and demersal, from European waters and the North American Pacific (Hubbs, 1948[143]; Fitch, 1952[144]; Ketchen, 1956[145]) and Asia (Uda, 1952[146]). Other authors warned of Climatic temperature changes and commercial yields of some marine fishes in Canada (Bell & Pruter, 1958[147]) and the first “El Niño events” began to be described (Schweigger, 1959[148]) which offers revealing details of fish, mammal and bird mortality.
In different specific European areas, the unusually large biological variability was also detected: cod and herring fluctuations in the NE Atlantic waters (Rollefsen, 1949[149]; Hansen, 1949[150]; Rasmussen, 1959[151]), as well as in the Southern North Sea (Cushing & Burd, 1957[152]). Also in the 1950s, in the English Channel the unusual abundance of pilchards (Cushing, 1957[153]), the common octopus (Octopus vulgaris) plague of 1950 (Rees, 1951[154]; Rees & Lumby, 1954[155]) and alterations of the bottom fauna were recorded (Holme, 1961[156]). Changes in the benthic fauna were also detected in the Baltic (Shurin, 1961[157]), coinciding with anomalous oceanographic situations such as increases in salinity in the Ostsee in 1951 (Meyer & Kalle, 1950[158]; Wyrtki, 1954[159]) responsible for their particular registered biological alterations. Drastic environmental changes that reached the Spanish northern coasts and were responsible for the displacement of the intertidal biogeographic boundaries studied in 1954-1955 (Fischer-Piette, 1955[160], 1956[161], 1957a[162],1957b[163],1958[164],1963[165]).
In the north of Iberia, in the waters of Galicia and the Bay of Biscay, José Mª Navaz (1897-1975), researcher at the Instituto Español de Oceanografía (Spanish Institute of Oceanography), made important contributions. Particularly his scientific publications on the alternation between different species in Galicia (sardine, sprat –Sprattus sprattus– horse mackerel – Trachurus trachurus– and anchovy) during 1929-1943 (Navaz, 1946a[166]), the appearance of exotic southern fish species also in Galicia (Navaz, 1946b[167]) and in the Cantabrian Sea (Navaz, 1961[168]), along with popular articles in the daily press in relation to the detected warming in the sea water. This important informative work for society in the mid-20th century was also practiced by two pioneering women, who in their popular divulgative books included several conclusions from scientific publications from the period 1920-1950. The North American Rachel Carson (1907-1964) with her synthesis on the climatic changes of the especially alarming decades of 1924-1944 when the floating ice of the Russian Arctic ended up decreasing by a million square kilometers and the profound environmental changes allowed the arrival in Iceland of species of birds and fish typical of warmer and southern waters (Carson, 1952[169]). Abnormal warming reached the productive southern fishing grounds of Newfoundland in 1948 and cod disappeared from its bottoms, as denounced by the Frenchwoman Anita Conti (1899-1997) (Conti, 1953[170]).
The 1960s began with the symposium on Changes of Climate in Rome, Italy, organized in 1961 by UNESCO and WMO. With the difficulty of understanding complex fish population fluctuations, the need to undertake long-term research with historical records of meteorological, oceanographic and biological data was recognized (Sette, 1960[171], 1961[172]). Particularly in the United Kingdom, a succession of unusual weather events since the mid-1940s (an abundance of wet summers and cold winters) and the extremely cold conditions during 1962-1963 and its negative effects on marine life (Crisp, 1964[173]), with its probable ominous implications for the future on food resources, promoted the holding of the symposium The Biological significance of Climatic Changes in Britain in 1964 at the Royal Geographical Society in London. In those years, the anomalies in the sea-surface temperatures of the North Atlantic in the 1950s were also described (Rodewald, 1963[174]), a secular warming trend peaking in the early 1950s was demonstrated along with and a slight tendency towards cooling in certain small areas (Stearns, 1965[175]) and the mean sea level related to solar radiation during the last 20,000 years (Fairbridge, 1963[176]).
Regarding fish and fisheries, we highlight also an integrative vision on hydrodynamic fluctuations and his biological consequences (Beverton & Lee, 1965[177]). Also noteworthy is a study of great temporal and spatial amplitude, which compared the catches during 55 years of several species of pilchard-sardine during 1905-1960 (Zupanovitch, 1968[178]), demonstrating similarities between the populations off California (Sardinops sagax), off Spain (Vigo) and in the Adriatic; as well as linked the catches (low/high) of the Adriatic sardine since the sixteenth century with changes in the landings of the Japanese sardine (Sardinops melanostictus). Subsequent reviews of the Atlantic sardine fishing crises in Spain and Portugal were published (eg. Wyatt & Porteiro, 2002[179]; Orestes, 2002[180]), stating the latter author “that most of the crises have been due to environmental factors, and only during the second half of the 20th century can the effects of fishing be viewed as having added to the recruitment failures observed in the populations”. Indeed, a dramatic increase in fishing pressure worldwide since 1950 has been confirmed and in the Celtic Sea there was a large fishing-induced decline in biomass, mainly before 1970 (Hernvann & Gascuel, 2020[181]).
In three monographs from the 1970s, numerous international multidisciplinary publications from previous years were reviewed. It was confirmed that there is some evidence of a correlation between climatic changes in the Northeast Atlantic region and the global climate regime (Lamb, 1972[182]). A synthesis on the effects of past climate changes on marine resources (Cushing, 1975[183]), with the biological response in the sea and the role of man in the regulation of fish populations, introducing the concept of recruitment overfishing and pointing out a plan of studies for fisheries oceanography. Another study (Cushing & Dickson, 1976b[184]) includes a complete review for areas bordering the Bay of Biscay: southern North Sea and western English Channel, with three events common to the two changes in the 1930s and the 1970s, and the extreme winters occurs in north-west Europe (one every 10 or 15 years), considered as an index of climate change, which usually cause damage to the intertidal fauna and death of fish species. Ending with a description of the links between different scales of climatic changes (seasonal, every 5 or 10 years and a period about a century), the associated biological events and the forms of biological response (indicator species, colonization, alteration of stock size and structural changes in the ecosystem).
The following decade of the 1980s was decisive in consolidating global research on climate variability and its influence on marine populations and ecosystems, with the exchange of information between global researchers in several specialized symposiums and the publication of many seminal articles and books. The multidisciplinary book Climatic changes on a yearly to millennial basis: geological, historical, and instrumental records was published (Mörner & Karlén, 1984[185]), with some subsequent statements: “Major short-term climatic changes during the last 20.000-30.000 years are found to have a duration of the order of 50-150 years, duration that reveals the oceanographic origin” (Mörner, 1986[186]); and that in NW Europe can see a direct link between climatic change and variations in Gulf Stream activity. Other integrative book (Cushing, 1982[187]) examined the connection between climate and changes in fish stocks, describing among other issues: changes in fisheries during historical times (particularly the period of warming from 1880 to 1945), the influence of cold winters in western Europe, the long-term changes in the Baltic Sea and North Sea, reviewing the biological mechanisms and investigating the effects of climatic factors on recruiting year classes, ending with a discussion on the management of fisheries. Other authors (Caddy & Gulland, 1983[188]) warned that for fisheries research it should be taken into account at least three basic prior criteria. i)That fish stocks can be divided into four classes according to their natural patterns of variation: steady, cyclical, irregular and spasmodic; this last pattern is frequent in some anchovy and sardine stocks, characterized by alternating periods of high abundance and scarcity or collapse, with potential fluctuations of 10 to 20 years. ii)The causes of variations in fish landings fall into two main headings which are not mutually exclusive: fluctuations in the marine environment and variations in fishing intensity, and clarifying their relative impacts can help predict upcoming events. iii)In the researchers' scientific career it is difficult for more than one longer-term periodic and catastrophic phenomenon to occur, useful as an indicator of past climate, such as volcanic eruptions (that had a cooling impact on the climate), and certain intense changes in precipitation and their effects on drought and river runoff, which can follow cycles for long periods of the order of 70-100 years.
More interesting multidisciplinary articles were also published in the 1980s. On the influence of the global trend in temperature on the fluctuations in the three major sardine stocks in the Pacific (Kawasaki & Omori, 1988[189]). The great salinity anomaly of the 1970s (Dickson et al., 1988b[190]), traced from Greenland all around the North Atlantic (from 1968 to 1982), was discovered, which caused changes in planktonic communities and reduced the yields of 15 deepwater stocks during its passage (Cushing, 1995[191]). The studies on the four major upwelling areas (California, Peru, Canary and Benguela) and the fluctuations of the 16 common species in those characteristic physical environments (genera Engraulis, Sardina, Sardinops, Trachurus, Scomber, Sarda and Merluccius) (Bakun & Parrish, 1980[192]). For the tropical multispecies fisheries off West Africa, a series of dramatic changes since the mid-1970s in species composition and distribution were described for several species of fish and cephalopods (see 3.3.3). The fluctuations in the herring and pilchard fisheries off southwest Britain were linked to changes in climate since the 16th century (Southward et al., 1988[193]). The first numerical models for the North Sea herring were published, recognizing the need to incorporate details of behavior of plankton and fish larvae to convert this information into ecological forecasts (Backhaus, 1985[194]; Bartsch et al., 1989[195]). A wide thematic variety was generated, among others: on climate and aquatic food production (Bardach & Santerre, 1981[196]), on fisheries management in the climate change context (Frye, 1983[197]); a background paper on relationships of fisheries to climate change and marine environmental factors (Sibley & Strickland, 1985[198]); variations of fish stocks with climate (Shepherd et al., 1984[199]). On the Spanish Mediterranean coasts of the Alboran Sea, four species of fish have been identified that are useful as indicators of cold, temperate and warm waters. Catches of two of those species, of small coastal pelagic fish, common in its waters throughout the year, showed drastic changes since the mid-1960s: during the cooling period of 1966-1975 sardine catches almost tripled those of anchovy, and vice versa in the warming period of 1977-1990 (Pérez-Rubín, 1996[200]). A third species in the years of maximum warming in the pelagic ecosystem, the round sardinella or gilt sardine (Sardinella aurita), opportunistic species, appeared and entered into competition with the anchovy (Pérez-Rubín & Mafalda, 2004[201]). Also the boarfish (Capros aper), a subtropical species, exponentially increases its presence in the region during decades of oceanic warming (Pérez-Rubín & Abad, 1994[202]).
The line of research on long-term marine biological changes and the cyclic fluctuations was promoted and several meetings of world experts in fisheries and oceanography were held in the 1980s, who have been working together to determine what effect human activity has on the marine environment and the effects of environmental factors such as climate, hydrography and fertility of the oceans. The global research collaboration on fisheries was initiated by an international symposium organized in 1983 by FAO and IOC-UNESCO in Costa Rica to examine changes in abundance and species composition of neritic fish resources (Sharp & Csirke, eds., 1983[203]; Csirke & Sharp, eds.,1984[204]). This was followed by the 1986 symposium on Long Term Changes/Variability in the Spanish city of Vigo, on the effects on marine fish populations (Wyatt & Larrañeta, eds. 1988[205]), which included, for Spanish Atlantic waters, five important studies with information since the early 1950s. In Asturias, annual and seasonal changes were described during 1952-1985 in the catches of a total of 10 species of fish, both pelagic and demersal (López-Areta & Villegas, 1986[206]; Villegas & López-Areta, 1986[207]). During the mid 1960s-1970s the Bay of Biscay anchovy withdrew from the Galician coast and the western Cantabrian Sea and retreated to the eastern Bay of Biscay (Junquera, 1988[208]). For the period 1976-1985, the variations in the sardine stock of the Western Iberia were described, mainly due to variations in recruitment (Porteiro, Álvarez & Pérez, 1986[209]) and suggesting the role of Ekman transport in larval recruitment in the coastal waters of Portugal and W. Galicia (Wyatt & Pérez-Gándaras, 1986[210]). In the Channel and the Bay of Biscay, a comparative study on the opposing fluctuations in French sardine and herring fisheries demonstrated that they have followed longterm changes in climate for three centuries (Binet, 1988b[211]). A third symposium, the American on Effects of Climate Change on Fish held in Toronto, Canada, during 1988 (Regier et al., 1990[212]), was followed in 1989 by the Japanese symposium hosted in Sendai on the Long-term variability of pelagic fish populations and their environment (Kawasaki et al., 1991[213]), including this work a communication on interrelations between the environmental change and fluctuating European sardine populations (Wyatt, Cushing & Junquera, 1991[214]). Furthermore, research on the impacts of climate change on marine ecosystems that occurred in previous millennia was also incorporated into the literature. The renewed Paleoecological studies on variability in marine fish populations focused on studies based on fossil fish and in the deposits of their scales in the sediment, as indicators of past sardine and/or anchovy populations in California (Souter & Isaacs, 1974[215]), off Peru over the last 2,000 to 2,700 years (DeVries & Pearcy, 1982[216]), off Namibia (Shackleton, 1988[217]) and a comparative study on the results found for three upwelling regions (Shackleton, 1987[218]).
In the 1990s, more seminal interdisciplinary books were published containing contributions from scientist around the globe on the potential impacts of global change on ocean productivity and fisheries. Among others, in chronological order: Large Marine Ecosystems: Patterns, processes and yields (Sherman et al., 1990[219]), Climate variability, climate change and fisheries (Glanz, 1992), Scaling Fisheries (Smith, 1994), Population production and regulation in the sea. A fisheries perspective (Cushing, 1995), and Patterns in the Ocean: Ocean Processes and Marine Population Dynamics (Bakun, 1996[220]). For fisheries scientist, the aforementioned books by Glantz and Cushing are particularly interesting, in which the history of the major climatic changes and ecological shifts that are well recorded in the global fisheries literature is described. Cyclical times of expansion and abundance vs collapse of these populations are identified, coinciding with periods of marked marine environmental changes and/or by anthropogenic causes on a global scale since the 1920s. Varied human activities and subtle changes in key environmental variables (such as temperature, salinity, wind speed and direction, ocean currents, strength of upwelling, as well as predators), can sharply alter their abundance, distribution and availability of fish populations. And also influenced by many elements of their natural environments during all phases of their life cycle, including the location of spawning grounds and monitoring the survival of their eggs and larvae in the planktonic ecosystem. Our bibliographic review of interest to fisheries ends by citing two monographs. The entire 1990 issue of Fisheries magazine (of the American Fisheries Society) dedicated to the effects of global climate change on fishery resources, and the titled Climate Change and Northern Fish Populations, by National Research Council Canada (Beamish, 1995[221]), which summarizes contemporary studies (of the relationships among climate, the dynamics of fish populations and the aquatic environment) from the North Pacific, North Atlantic and from freshwater.
With respect to the ocean surface circulation system it has been demonstrated that it has a much more important role than previously appreciated, and the paleoclimatic changes on the decadal-to-millennial time scale are primarily driven by the same complex mechanism: the “connection between Earth's rate of rotation, ocean circulation, ocean/atmosphere heating, atmospheric (wind) heat transport and continental paleoclimatic changes” (Mörner, 1995[222]). That author also suggested the importance of the spatiotemporal variability of the branches of the Gulf Stream and the polar front in the North Atlantic, from the Last Glacial Maximum (peaked at around 20 kyr ago) to the 1982/83 ENSO-events. In addition, the first secular reconstructions of solar irradiance since 1610 and their implications for climate change were also analyzed (Lean et al., 1995[223]); and the solar variability is also defended as a decisive forcing function that affects the Earth's rotation, ocean circulation and climate change (Mörner, 1996[224], 2013[225]), as well as the cyclical pattern of about 60 years that relates changes in the length of the day, temperature and sea level in the North-Western Europe region. The correlation of temperature of the northern hemisphere with solar cycle length was initially demonstrated over the period 1850-1970 (Friis-Christense et al. 1991[226]), and then confirmed and extended to further 65 years, back to the end of the 18th century (Butler & Johnston, 1996[227]). Also for pelagic fisheries research, the study of the solar variability is very interesting. In oceanic sedimentary records (fish-scale abundances of Northern anchovy Engraulis mordax and Pacific herring Clupea pallasii, at decadal to centennial scales) an especially well-defined variability at one stationary solar cycle (75-90 years) has been demonstrated (Patterson et al., 2004[228]). On the other hand, solar variability negatively affected bluefin tuna catches in the North Atlantic and Mediterranean for at least 165 years, differently during two phases. In the first one during the intense cooling of the period 1645–1715 (with minimal solar activity, included in the so-called Little Ice Age) (Ganzedo et al. 2009[229]); and in the second one (mainly during 1700–1810) with a strong correlation founded between the total solar irradiance and bluefin catches (Ganzedo et al., 2016[230]). The variability of the Atlantic Iberian sardine (annual landings from 1906 to 2002) has been found to be governed by the alternation of short and longer solar cycles, which will also potentially affect different biological processes (Guisande et al., 2004[231]).
We finished this historical review with the workshop convened at the National Oceanic and Atmospheric Administration (NOAA) in 1996 to examine the uses of environmental data for fisheries research and management, with one of its main objectives: “to assess the current and future needs for environmental data bases (oceanographic, atmospheric, remote sensing, model output, and geological) in fisheries research and management” (Boehlert & Schumacher, 1997[232]).
All of this historical information remains useful in the 21st century to provide appropriate reference models to better understand a century of complex interactions between climate, the marine environment, fish stocks and human activities (e.g. overfishing and the application of certain management schemes or new technologies).
3.2. Anthropogenic Global Warming vs Natural Global Warming
The described different origins of international initiatives to increase global research on climate change (CC) since the 1980s have become evident in the 21st century, even with a decisive challenge to overcome the limitations recognized in 1999: “it is not possible to make a very confident statement about the relative contributions of specific natural and anthropogenic forcings to observed climate change” (Barnett et al., 1999[233]).
When comparing the different criteria for the definitions of CC and it usages in the framework of the IPCC vs UNFCCC, substantial differences have been found (IPCC, 2007[234]). CC in IPCC usage refers to any change in climate over time, whether due to natural variability or as a result of human activity. This usage differs from that in UNFCCC, where CC refers to a change of climate that is attributed directly or indirectly to human activity that alters the composition of the global atmosphere and that is in addition to natural climate variability observed over comparable time periods. That greater or unique anthropocentric responsibility is based on the hypothesis that the change in the concentration of greenhouse gases (mainly CO2) is the most probable cause of the intense and lasting increase in the planet's global temperature recorded since 1850 (IPCC, 2001[235]), and that there is a linear relationship between the increase in atmospheric CO2 and global temperature. Although other scientists believe that sometimes only natural causes can be decisive in CC, and they think that the UNFCCC's overly restrictive definition “has often been adopted by the media, in policy documents and in some scientific literature and has also been used interchangeably with global warming" (Drinkwater, 2010). According to the IGBP synthesis book Global Change and the Earth System: A Planet Under Pressure (Steffen et al., 2004[236]) the humanity was now the main driver of global change and the planet has moved well outside the range of natural variability in the last half million years at least. The concept of the Anthropocene was introduced in 2000 at a meeting of the IGBP, defending the opinion that the cumulative negative impact of human activities over the past two centuries, mostly from 1945 to c. 2015 (called the “Great Acceleration”), having a decisive influence on the continuing to accelerate loss of the planet’s biodiversity that has led the Earth to enter a new geological epoch, towards its sixth great extinction event (eg. Steffen, 2010[237]). However after 15 years of reports prepared by a multidisciplinary group of Quaternary specialists, the responsible international organizations refused in 2024 to consider the Anthropocene as a new geological epoch and include it in the International Chronostratigraphic Scale.
The general opinion that CC is mainly of caused by greenhouse emissions from human activities to the atmosphere resulting in an Anthropogenic Global Warming or AGW is maintained (IPCC, 2022[238]). Thus for instance, for the European Environment Agency in 2024: “Our climate is changing because of greenhouse gases released into the atmosphere”[]. On the other hand, although it is also claimed that global mean sea-level (GMSL) is rising and accelerating due to global warming, however, that claim does not seem to take into account that the most numerous and reliable historical records of GMSL, from tide gauge reconstructions, are only from the 20th century, although structural uncertainty is high before 1950 (Palmer et al., 2021[239]).
The current reality is a lack of full scientific certainty, because there is no international unanimity on identifying the main drivers of global temperature changes and their possible relationship with GMSL. On the other hand, on the contrary to the main current of opinion, part of the international scientific community dissent on the AGW as the main cause of CC. This part of the scientific community support the hypothesis that CC is mainly caused by Natural Global Warming (NGW) due to solar variability. Although internationally this is a minority line of research, it is supported by publications in peer-review journals and must also be taken into account in an impartial and rigorous bibliographic review. In the papers consulted we find experiments to address different scenarios with the relative importance of anthropogenic and solar forcing (Rind et al., 2004[240]). The aforementioned temporal limitation of sea-level records (less than one century) contrasts with studies based on records for the last 500 years in different global locations, which indicate the present sea level stability (“with some oscillations in the 19th, 20th and early 21st centuries”), whose level changes seem to be correlated with the opposite forcing ocean water masses during Grand Solar Minima/Maxima, and “the recorded sea level changes are anti-correlated with the major changes in climate during the last 600 years” (Mörner, 2017[241]).
Since the last decade there is a growing interest in research on climate change vs solar activity variations and the need for the decisive solar influence to be revalued in the 21st century and incorporated prominently into the international concept of Global Warming has been highlighted (Mörner, 2020[242]). From 1963 to 2010, for the Northern Hemisphere in winter, in the region of the North Atlantic Ocean, a high correlation between sea surface temperature and the solar wind speed has been found (Zhou et al., 2016[243]). With extensive long-term reconstructions of the solar irradiance (Velasco et al., 2015[244]), and its relationship with solar activity and ozone thickness (Jardine et al., 2010[245]). Furthermore, it has been suggested that cyclical periodic variations in different direct and indirect climate records are synchronized to solar variations in timescales of a decade to millennia (Miyara et al., 2023[246]), the solar forcing synchronizes the North Atlantic climate decadal variability (Thiéblemont, 2015[247]), and the importance of solar activity (together with atmospheric CO2) on the wind regime and the water column stability in upwelling areas has been described (Patti et al., 2010[248]). With the analysis of the sun's varying activity (cyclic solar forces) in the last two millennia, the surprising prediction of a new potential future Grand Solar Minimum has been reached, that would theoretically cause severe global cooling in coming decades over the Arctic and NW Europe, similar to cold periods detected around 1450, 1690 and 1815 (Landscheidt, 2003[249]; Mörner, 2010[250], 2015[251], 2018[252]). Although some authors believe that “a future Grand Solar Minimum could slow down but not stop global warming” (Meehl et al. 2013[253]). The impact and implications of this hypothetical future solar minimum on climate change on Earth have been assessed (Feulner et al., 2010[254]), and could have important negative consequences on the stratosphere and on the climate in Northern Hemisphere at regional surface (Arsenovic et al., 2013[255]; Maycock et al. 2015[256]; Ineson et al. 2015[257]; Chiodo et al., 2016[258]). Evaluating many hundred scientific arguments, recently several authors have recognized the need to increase of research the relationship between climate and solar activity and incorporate solar radiation in the numerical simulation of weather (daily reports and seasonal forecasts) and for climate projections; taking into account that different variations (eg. solar 11-year cycle, 27-day cycle, and Grand Solar Minimum) can cause short and long-term changes in the atmosphere and oceans (Yoden & Yoshida, 2023[259]). Although the importance of ultraviolet solar irradiance as a climate forcing mechanism is clear, there is still high uncertainty about the temporal evolution of this irradiance during the last millennium and for future research it is advisable to incorporate fully interactive atmospheric chemistry (Tartaglione et al., 2024[260]). In brief, the IPCC suggests that almost 100% of the observed surface warming from 1850 is due to anthropogenic emissions. However, these conclusions are based solely on computer simulations performed with global climate models (useless at regional scale) that do not take into account all the main solar processes proposed in the peer-reviewed literature (Scafetta, 2023[261]). That author has proposed three “balanced” multiproxy models of total solar activity that can generate more realistic global and sea surface temperature warming projections. Various scientists have also demonstrated the decisive influence of solar variability on pelagic fisheries (see 3.1.).
A third group of scientists are equidistant or eclectic specialists (eg. Polyakov et al., 2010; Kopp & Lean, 2011[262]; Hughes et al., 2012) and believe that the detected global trend of temperature increase is the result of very complex processes developed at different time scales. For the particular case of the Iberian Peninsula, it has been confirmed by CLIVAR-Spain that its climate over the last 2000 years is controlled by fluctuations (and interactions) with climate modes of variability (e.g. NAO, EA, SCAND), solar irradiance and volcanic activity (Giralt et al., 2017[263]). It is highlighted that although as well as the long-term trend attributable to anthropogenic climate change, there are other complementary complex regional patterns of decadal and multidecadal scale natural variability. For the period 1980-2010, the combined effect of El Niño-Southern Oscillation (ENSO), solar activity, volcanic aerosols and anthropogenic effects explain 85 % of observed global temperature variance (Kopp & Lean, 2011). Particularly interesting is the use of the AMO climatic index, which aims to represent the variability in the North Atlantic caused by mechanisms other than anthropogenic climate change (Hughes et al., 2012). Therefore, disentangling natural climate variability from anthropogenic effects on CC requires the development of different methods and approaches (historical marine ecology, spatial comparisons, designed experiments, statistical modeling and genetics methodologies) (MacKenzie & Christensen, 2010[264]).
3.3. The Current Rapid Warming and Tropicalization in South European Atlantic Seas and Connecting African Regions
Over recent decades, rapid ocean warming has expanded across Large Marine Ecosystems (Belkin, 2009[265]), with notable impacts on numerous European and African fisheries, as we will describe in this section. Long lasting changes in water temperature and salinity drive fish species with particular thermal preferences to migrate to different latitudes. While this can be a cyclical process that we have observed since the 19th century, the current accelerated rate of ocean warming is alarming (see 3.3.1). It is crucial to analise this process using the available 20th century data to provide a basis for comparison with the present situation.
Historically, shifts of many marine species migrating north and south across different decades have frequently occurred in various biogeographic transition zones. East-to-West movements, away from the coast, often result in certain cold-water species moving into deeper areas with lower temperatures.
In the Atlantic waters of Europe, Boreal, Lusitanian, and Mediterranean species often come into contact, and during prolonged warming periods, tropical African species also appear. In the Eastern Atlantic (from southern and western Iceland all the way down to southern Africa), there exists a continuous set of regions with sharply contrasting oceanographic and ecological characteristics. These differences were manifested through the local presence of multiple species typical of temperate, tropical, and equatorial waters. During decades of water cooling or warming, the physical-chemical and biological differences in their distinct ecoregions are comparable, and it is necessary to analyze these using the data available from at least the 1950s.
Unfortunately, there are numerous regional demarcations proposed by various international bodies and independent researchers that complicate large-scale integrative scientific research. The research and management of fish stocks are conducted separately in the European ICES areas and the African CECAF areas (which extends from the south of the Strait of Gibraltar to the mouth of the Congo River) and the former ICSEAF (Angola-Namibia-South Africa), that has now replaced by the South East Atlantic Fisheries Organization (SEAFO). From an environmental perspective, the MEON (Marine Ecoregions of the World) biogeographic system in the Eastern Atlantic, identifies a total of 7 provinces and 23 ecoregions that are grouped into several realms (Spalding et al., 2007[266]). The Tropical Atlantic realm is centrally located (from Cape Verde-Senegal to Angola), and to the north, the Temperate Northern Atlantic (TNA) realm includes two Atlantic provinces: Northern European Seas (with 7 ecoregions) and Lusitanian (with 3 ecoregions: South European Atlantic Shelf, Saharan Upwelling, and Azores-Canaries-Madeira). Additionally, in the TNA, there is the Canary Current Upwelling System (CCUPS) or Canary/Iberian Current System (CanCS), extending from the northwest of the Iberian Peninsula to Senegal, with both northern and southern environmental limits fluctuating seasonally (Kämpf, 2016[267]). Moreover, based on the scientific rationale of the Large Marine Ecosystem (LME) approach (Sherman & Hempel, 2008[268]), four ecosystems have been delineated: the African Canary Current LME (CCLME) off the NW African coast, and the European ones: North Sea LME, Celtic-Biscay Shelf LME, and Iberian Coastal LME (the latter with three subregions: Galician, Portuguese, Gulf of Cadiz). In addition to these subdivisions in fisheries and biogeographic/oceanographic regions, geopolitical barriers also exist. For example, the European Union recognizes two distinct regions in the Atlantic waters of Spain and Portugal[269]: first is the area from the Bay of Biscay to the Gulf of Cadiz (Bay of Biscay and Iberian Coast) to the north, and the second in southern region, encompassing the Canary, Madeira, and Azores archipelagos (Macaronesia). For all these reasons, comprehensive study and international management of the CCUPS, and its adjacent areas is highly complex. Fortunately, since 2018, a programme funded by the European Union called Programme for Improved Regional Fisheries Governance in Western Africa, has been implemented by the fifteen countries of the Economic Community of West African States (ECOWAS). To evaluate the impact of fishing on the African Large Marine Ecosystems, the long efforts of the global ‘Sea Around Us' initiative have been very valuable (Zeller et al., 2020[270]).
The objectives of this chapter are to consider the historical background across the European spatial extent from the North Sea to the Gulf of Cadiz and to analyze the consequences of the current rapid global warming on fish and fisheries, considering that stocks fluctuate based on varying environmental and biological processes as well as fishing activity. To better understand the situation in the 21st century in southern European waters, a broader retrospective investigation is required, including various regions of West Africa within the CCLME, especially from the decades of relative cooling in the 1960s–1980s and the subsequent relative warming from the 1990s onward. Given its vast geographical expanse, the CCLME includes numerous areas that are particularly sensitive to climatic and oceanographic fluctuations, making them of special interest for detecting changes in pelagic and demersal ecosystems.
3.3.1. North Sea/Bay of Biscay/Iberian Coast
We previously looked at (see 3.1) several decades of ocean cooling/warming variations primarily from 1900–2005, noting significant warming of sea surface temperatures (SST) in different parts of the Northeast Atlantic since the late 20th century. Particularly alarming is the ongoing surface water warming in the Northeast Atlantic, recorded from the mid-1980s to the present. Along the eastern Bay of Biscay (Basque coast shelf), this warming since the 1980s has led to changes in the biology of pelagic fish, gradually altering biodiversity and affecting benthic communities (Chust et al., 2022[271]). In more recent years, concern has grown over non-native fish species in Spanish waters (Bay of Biscay-Iberian shelf and Macaronesia; Zenetos-Carbonell, 2022[272]; Png-Gonzalez et al, 2023[273]). In other southern European coastal waters, the warming-induced biological changes from the 1990s onward have impacted the entire water column, changing both the surface waters (affecting pelagic fish populations) and seabeds. Macroalgae communities on Iberian Peninsula coastal beds have shown a significant increase in non-native species from other latitudes in recent decades. Notable changes include the abundance and distribution of several macroalgae species in Galicia (Carreira-Flores et al., 2023[274]), along Spain's southern coasts (Andalusia) and the Strait of Gibraltar (Pérez-Lloréns et al., 2023[275]). In Portuguese waters, macroalgae have shown a rapid trend toward tropicalization in recent years (Azevedo et al., 2023), while local upwellings have create a transition zone with the coexistence of algae communities from two distinct origins (Boreal and Lusitanian-Mediterranean).
A comprehensive analysis of spatial-temporal evolution in the NE Atlantic has shown that fish species predominantly expanded southward during the relative cooling period of the 1970s and 1980s, and northward during the subsequent decades of relative warming (Heath, 2007[276]). Since the mid-1980s, in ecosystems across the eastern Atlantic, from northern regions (North Sea, Celtic Sea) to southern regions (Canary and Guinea Currents), specific routes and years of fish species migrations have been documented (Kaimuddin et al., 2016[277]). These authors reveal that non-native/exotic species have primarily migrated northward, some southward, or expanded their depth ranges in response to environmental changes depending on the season. In mid-regions (South European Atlantic Shelf, Western Mediterranean Sea), there is an increasing presence of species from both high and low latitudes. The Canary Current exports species to these mid-northern regions and southward toward the Guinea Current (Kaimuddin et al., 2016).
Oceanic warming has accelerated in recent years. According to NASA[278], the last 10 years have been the hottest and “2023 was the warmest year on record since record-keeping began in 1880.” Europe’s situation is particularly concerning as it is considered “the fastest-warming continent on Earth” since the 1980s, warming “twice as fast as the global average” (ESOTC, 2023[279]), with an escalation in temperatures beginning in 2007 and intensifying since 2020. The most recent records for the WMO Regional Association VI (Europe domain) confirm that 2020 and 2023 were the warmest years on record (ECMWF, 2024[280]). The dire situation in European seas is shown in an image produced by the European Environment Agency (EEA), illustrating sea surface temperatures from 1980-2023 and ranking the annual average SSTs for 2023 (Figure 1). The extensive marine warming, from the Canary Islands to the Bay of Biscay and extending south of Iceland and into the Mediterranean, has been monitored by the European Marine Climate Change Index (EMCCI).
Figure 1.
Ranking of the annual average sea surface temperatures in 2023 (EMCCI index), relative to the period 1980-2023 (44 years). Darkest red: the areas where 2023 was the warmest year on record. Lighter shades: areas that were closer to average. [Southwest sector of the original image: MAP1-259111 (14 Dec. 2023]. [Data source: ESA SST CCI Analysis v3.0. Reference data: ©EuroGeographics, ©FAO (UN), ©TurkStat. Source: European Commission-Eurostat/GISCO. Credit: ESACCI/EOCIS/UKMCAS and C3S/ECMWF (2024[281])]. https://www.eea.europa.eu/data-and-maps/figures/european-marine-climate-change-index-emcci [Re-use of content on the EEA website for commercial or non-commercial purposes is permitted free of charge, provided that the source is acknowledged (https://www.eea.europa.eu/legal/copyright). Copyright holder: European Environment Agency (EEA)].
Figure 1.
Ranking of the annual average sea surface temperatures in 2023 (EMCCI index), relative to the period 1980-2023 (44 years). Darkest red: the areas where 2023 was the warmest year on record. Lighter shades: areas that were closer to average. [Southwest sector of the original image: MAP1-259111 (14 Dec. 2023]. [Data source: ESA SST CCI Analysis v3.0. Reference data: ©EuroGeographics, ©FAO (UN), ©TurkStat. Source: European Commission-Eurostat/GISCO. Credit: ESACCI/EOCIS/UKMCAS and C3S/ECMWF (2024[281])]. https://www.eea.europa.eu/data-and-maps/figures/european-marine-climate-change-index-emcci [Re-use of content on the EEA website for commercial or non-commercial purposes is permitted free of charge, provided that the source is acknowledged (https://www.eea.europa.eu/legal/copyright). Copyright holder: European Environment Agency (EEA)].
To monitor and visualize the spatio-temporal evolution of thermophilic fish distributions in Atlantic waters as represented in Figure 1, a portal like NOAA's DisMAP[], for the U.S. marine ecosystems, is needed to understand specific species distributions in European waters. The FAO's recent Aquatic Species Distribution Map Viewer provides historical maximum geographic distribution data for numerous marine species (924), including some European and African species (FAO, 2020[282]). To better understand the tropicalization process, examining selected thermophilic fish species and how warm-water species (native and alien) expand geographically while cold-water species contract, is crucial, as well as examining how biogeographic boundaries have shifted. In European waters, comparing Boreal vs Lusitanian species is a useful indicator. The English Channel serves as a permeable boundary connecting the Bay of Biscay with the southern North Sea, allowing Boreal fish species (characteristic of northern Channel waters and extending northward toward Iceland and Norway) and Lusitanian species (prevalent in the Atlantic and Mediterranean waters of the Iberian Peninsula) to interact.
In the region from NW Iberia to the Strait of Gibraltar, Lusitanian fauna interact with Mediterranean species, while colder-water species are confined to upwelling areas. In the case of the Atlantic chub mackerel (Scomber colias), the Gulf of Cadiz has been suggested as a hinge or mixing zone for the Atlantic Iberian population with elements from the Mediterranean and Atlantic Africa (Domínguez-Petit et al., 2022[283]). Biodiversity increases in the Spanish and Portuguese Atlantic waters south of the Iberian Peninsula, influenced by deep Mediterranean currents exiting through the Strait of Gibraltar. These saltier Mediterranean waters influence several demersal fish species in the Gulf of Cadiz and are adapted to climate variations (Delgado et al., 2018[284]).
For European Atlantic continental regions, the European Environment Agency (EEA) has created maps showing changes in fish distribution over various annual intervals from 1982 to 2022 (EEA, 2024[285]). The maps depict the temporal development of the ratio between the number of warm-favouring (Lusitanian) fish species and the number of cool-favouring (Boreal) fish species by the ICES statistical area[286]. Figure 2 shows the results for three specific years (1990/2006/2022).
Figure 2.
Temporal development of the ratio of the number of Lusitanian species to the number of Boreal species (years 1990/2006/2022) in European regions north of the Strait of Gibraltar. Reference data: ©EuroGeographics, ©FAO (UN), ©TurkStat. Source: European Commission-Eurostat/GISCO.
Figure 2.
Temporal development of the ratio of the number of Lusitanian species to the number of Boreal species (years 1990/2006/2022) in European regions north of the Strait of Gibraltar. Reference data: ©EuroGeographics, ©FAO (UN), ©TurkStat. Source: European Commission-Eurostat/GISCO.
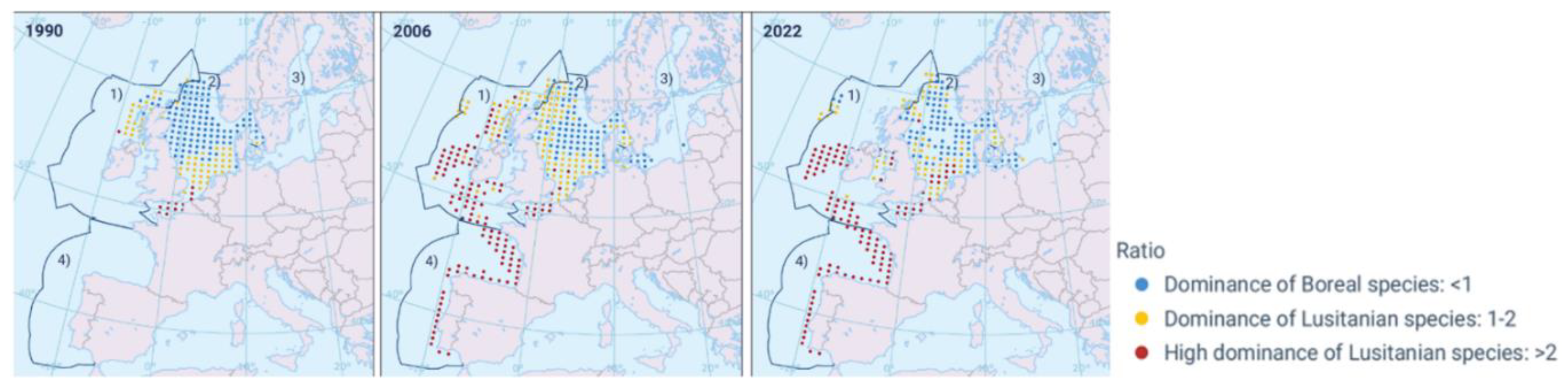
We now have data over the last few decades for nine pelagic and demersal species from Lusitanian fauna (which have increased, especially at the northern limit of their particular distribution areas) and Boreal fauna (which have decreased at the southern edge of their distribution and expanded into deeper waters; Rijnsdorp et al., 2010[287]). Other authors (Hofstede et al. 2010) have tracked different interpretations of the varied geographic origins of species (Boreal, Lusitanian, and Atlantic) and the variability across three key regions of the NE Atlantic (North Sea, Celtic Sea, and the area off the west coast of Scotland), primarily covering the years 1997–2008 and averaging around 100 species per region. An analysis of 50 abundant species in waters around the UK and Ireland showed that 70% of the species responded to warming in the region by changing their distribution and abundance (Simpson et al., 2011[288]). Specifically, small-bodied warm-water species have increased in abundance, while large-bodied cold-water species have decreased. These changes are significant and are linked to water temperature shifts (Simpson et al., 2011; Cheung et al., 2012[289]).
However, to better interpret these maps in Figure 2, a broader review of background data is needed to document the historical information available. By the late 1970s, a total of 224 fish species had been reported in the North Sea, with Lusitanian species being dominant (representing 49% of all species; Yang, 1982[A][290]). Despite this, Boreal fish fauna was considered dominant in terms of biomass (53.8% of the total), with “Atlantic” fauna being the least represented (Yang, 1982[A]). Currently, “Atlantic” species are those widely distributed in the North Atlantic, including "deeper-water species that may be widely distributed along the continental slope" (Hofstede et al. 2010[291], Cheung, 2012). Between 1966–1973, at least thirteen papers were published on the presence of non-native fish in British waters (references in Yang, 1982[B][292]).
The migratory cycles of specific species have been reconstructed for the North and Baltic Seas. Cold-water species that dominated from the 1960s to the 1980s (such as herring —Clupea harengus— and sprat —Sprattus sprattus—) were mainly displaced from the 1990s onward by associations of warmer-water species (e.g., sardine —Sardina pilchardus—, anchovy —Engraulis encrasicolus—, Atlantic mackerel —Scomber scombrus— and horse mackerel —Trachurus trachurus—) (Montero-Serra et al., 2014[293]). The general increase in sea temperatures across the NE Atlantic from the mid-1980s brought about a northward expansion, boosting southern Lusitanian fish species and introducing subtropical species (Pinnegar et al., 2010[294]). Some of these southern pelagic species reached high latitudes, such as Atlantic mackerel in southern Icelandic waters and Atlantic bluefin tuna (Thunnus thynnus) off Eastern Greenland (MacKenzie et al., 2014[295]). Sardines and anchovies returned to the North and Baltic Seas after approximately 40 years of absence (Alheit et al., 2012[296]). In British waters, warm-water species such as the demersal red mullet (Mullus barbatus) and the pelagic sea bass (Dicentrarchus labrax) also increased (Cheung et al., 2012). Particularly in the North Sea, the abundance of southern fish species increased almost exponentially from the mid-1990s, with a peak abundance observed for a highly diverse set of species that was comparable to other peaks in the late 1950s and mid-1970s (Beare et al., 2004[297]). Further south, in Spanish waters off the northern Iberian Peninsula, the presence of the first “uncommon or rare” warm-water fish species from southern waters was detected in the mid-20th century (see 3.1). First noted in Galicia (NW Spain) with four species, around 30 species were recorded a decade later in the Gulf of Biscay (eastern sector, Basque coast). In the same region of northern Iberia and in Spanish waters of the Strait of Gibraltar (Gulf of Cadiz, Strait of Gibraltar, and Alboran Sea), interdecadal fluctuations of various pelagic marine fish species are documented. We have an incomplete “puzzle” of data from the 1920s to the mid-1990s, comparing historical periods of warming versus cooling with fisheries crises affecting various species (Pérez-Rubín, 2008[298], 2023[299]).
3.3.2. Fish Species Useful as Indicators of Environmental Changes
Promonting the integrated study of small and medium-sized, short-lived pelagic fish is essential, as they are valuable indicators of environmental changes and play a key role as forage fish for larger predator species (e.g., Raybaud, 2017[300]; Checkley et al. 2009[301]; Engelhard et al. 2014[302]). For our intercontinental study area, it’s necessary to identify a selection of specific pelagic fish species that hold economic importance and serve as ecological indicators of environmental variability and the process of tropicalization. With a comprehensive spatial-temporal historical review, some of these species can also help detect past variability in ecological transition zones. Within certain ecosystem boundaries, early changes in the species composition of different origins may start to appear. For example, in the English Channel, Boreal and Lusitanian ichthyofauna meet: to the north are cold-water species (e.g., herring, sprat) and to the south, cold-temperate (sardine) and warm-temperate (anchovy) species. In the Strait of Gibraltar region, Atlantic species encounter Mediterranean ones. In Spanish waters off the southern Iberian Peninsula, four small pelagic species have been identified as useful indicators of water temperature: sardine, anchovy, round sardinella or gilt sardine (Sardinella aurita), and boarfish (Capros aper) (see 3.1).
In the Bay of Biscay, that subtropical boarfish, wich were rare in the 1970s, grew exponentially in abundance, eventually becoming a dominant species, largely due to rising water temperatures and the absence of its predators (Blanchard et al., 2005[303]). Locally, boarfish was sporadically caught in large quantities in the second half of the 19th century and in the mid-1950s, with catches increasing exponentially after 2006, reaching 207,882 tons in 2012 (Coad et al., 2014[304]). Boarfish have a broad vertical distribution (from 40–600 m depths), and in the NE Atlantic several European countries (primarily Ireland, Denmark, and Scotland) have developed commercial fisheries for it (Egerton, 2017[305]). In southern Spain, during a 1974 trawling survey in the Gulf of Cádiz, boarfish and snipefish (Macrorhamphosus scolopax) were the two most abundant fish species, comprising 36.80% of catches, with snipefish most abundant at depths of 100–200 m and boarfish at 200–600 m (coinciding with peak cephalopod catches) (Oliver & Fdez., 1974[306]). Since then, boarfish presence has increased in Spain’s Mediterranean waters, particularly in the Alboran Sea (see 3.1).
Sardine and anchovy are of particular interest in different coastal regions of Europe and Africa under the Canary Current Upwelling System (CCUPS), both playing a significant economic and ecological role, especially in coastal ecosystems. While sardines remain primarily associated with cold upwelling waters, significant long-term variations have been observed across their regional populations. These variations are linked to different environmental fluctuations and fisheries trends from Morocco’s coast (Chlaida et al., 2021[307]) to Gambia–Guinea Bissau (Sambe et al., 2016[308]) and the Iberian Peninsula. In the Iberian Peninsula, sardine catches have declined sharply since the early 21st century, while other pelagic species with more subtropical affinities, such as Atlantic chub mackerel (Scomber colias), have increased in number (Malta et al., 2016[309]), a species that continues to expand northward in the eastern Atlantic. The warm-temperate “European” anchovy is also expanding in various Atlantic regions from the southern North Sea to South Africa (previously called Engraulis capensis). Conversely, its stocks have declined since the 1990s in warmer regions, such as the Mediterranean (Raybaud, 2017).
In West Africa, in addition to sardine and anchovy, areas characterized by the abundance of two species of warm-water sardinellas —round sardinella and flat sardinella (Sardinella maderensis)— are observed (Ba et al., 2017[310]). Both sardinellas compete with each other and are concentrated around the three upwelling zones in West Africa, located south of Cape Blanco in the Eastern Tropical Atlantic (Cury & Fontana, 1988[311]). However, both sardinellas also undertake migrations and occupy distinct areas, with flat sardinella preferring more coastal waters and round sardinella inhabiting the continental shelf in warmer (> 24.0 °C) and more saline waters (Sambe et al., 2016). In fish landings as far north as Gambia–Guinea Bissau, these two sardinella species dominate, along with horse mackerels (Trachurus trachurus and T. trecae) in coastal waters (Sambe et al., 2016). Further south along the West Gulf of Guinea coast, both sardinella species remain important for fisheries, along with anchovy and chub mackerel (Scomber japonicus) (Asiedu, 2021).
A species indicative of the tropicalization of several European marine ecosystems is the grey triggerfish (Balistes capriscus). This tropical reef fish has a broad bathymetric range and is now well-known in the Mediterranean and much of the Atlantic Ocean, extending along both the eastern and western coasts, in the Macaronesian archipelagos, and along West Africa up to Angola (Aggrey-Fynn, 2009[312]), at the southern boundary between the tropical waters of the Eastern Tropical Atlantic and the temperate waters of Temperate Southern Africa. In the Iberian Peninsula, the grey triggerfish was present at least by the mid-20th century, in Galicia and the Bay of Biscay (see 3.1). By the mid-1970s, it was common in southern Galicia during the summer, extending northward in the following decades, and is now considered “the emblematic species with respect to tropicalization of native fauna,” with its fishery catches rising to 9,304 kg in 2022 (Bañón et al., 2024[313]).
3.3.3. North West African Seas (Canary Current LME)
The extensive CCLME ecosystem is one of the most productive and richest in the world, containing around 12,500 species of crustaceans, mollusks, and fish (Sambe, 2016). Since the 1930s, we have data on the great variability of several pelagic fish populations, with multiple episodes of collapse and boom in at least a dozen commercially significant species. For the multi-species fisheries of West Africa –targeting both temperate and tropical water species–, variability, instability, and changes were analyzed together (Cury & Roy, 1991[314]). It was shown that the highest diversity tends to occur in the biogeographic transition zone between temperate-warm and tropical waters, with intermittent processes of local demographic booms in certain species of regional economic interest. The historical cycles of warming/cooling phases from the 1930s to 2012 have been identified in the CCLME (Sambe, 2016): warming (1930s to 1960), cooling (1960s to 1980), and subsequent new warming, which continued into the 2000s. During several periods of extreme temperatures in the CCLME, changes in abundance and distribution boundaries of several species were detected, beginning with the intense cooling from the late 1960s to early 1970s (e.g., sardine, sardinellas, triggerfish, and snipefish) (Sambe, 2016). Starting in the mid-1970s, there were dramatic shifts in the composition and spatial distribution of various fish and cephalopod species. In Western Sahara, from 1970-72, there was a replacement of Sparids by Cephalopods, along with the remarkable expansion of the temperate-cold Sardine, replacing more tropical species like Scomber and Sardinella in the fish catches (Gulland & García, 1984[315]).
Fishery variability in African waters also extended into the tropical waters from southern Senegal to the Gulf of Guinea (Central Eastern Atlantic), and during the 1970s-early 1980s, the onset of drastic changes in catches of several key species were detected in different regional ecosystems. The most notable episodes in the NW Gulf of Guinea (Ivory Coast-Ghana-Togo) began with the collapse of its primary round sardinella fishery in 1971-1972, coinciding with the simultaneous invasion of the grey triggerfish. This species then expanded northward, reaching first Guinea-Guinea Bissau (1974-1975) and later, from 1978 onwards, Senegal-southern Mauritania (Gulland & García, 1984). Another author (Aggrey-Fynn, 2009) reported that peak catches of grey triggerfish in the early 1980s were recorded in the western stock in the Guinea Bissau−Guinea area, with its decline beginning in 1987 in the eastern stock in Ghana, reaching minimum catches in 2004. Conversely, by the late 1980s, the round sardinella stock expanded from Ghana westward (Ivory Coast), with its fishery extending throughout the year (Binet & Servain, 1993[316]). Further changes in Ghana's coastal pelagic fisheries occurred in the first two decades of the 21st century. In 2000, following peak catches of round sardinella, there was a drastic decline in landings of this species. In contrast, catches of other species, like Atlantic horse mackerel (Trachurus trachurus) and anchovy, began increasing from 2014-2016, coinciding with the latest decline phase of chub mackerel (Scomber japonicus), which reached its lowest level in 2018 (Asiedu, 2021[317]).
In the 1980s, explosions of various cephalopod species were also recorded, some benthic like common octopus (Octopus vulgaris) and others not. The first extraordinary abundance ("plague") of octopus was detected in the English Channel in the 1950s (See 3.1), and in the following decades, this phenomenon recurred along the coasts of various African countries. In the 1960s, the first spectacular proliferation of cephalopod populations (mainly octopus) began in the Sahara Bank, extending until 1980, attracting a massive presence of trawlers from different countries to exploit this new fishery (Balguerías 1993[318], 2000[319]). The same phenomenon was repeated in 1986 with a major proliferation of octopus on the coastal grounds of Gambia-Senegal, which became a new fishery resource in the area (Caverivière, 1990[320], 1994[321]). Further north, in the EEZ of Mauritania, octopus displayed significant interannual fluctuations in its spatial distribution during the 1980s-1990s, with an expansion into deeper waters in the cold seasons and into northern areas in the warm seasons (Inejih, 2002[322]).
During the net warming period from 1997-2012 (accompanied by a decrease in upwelling) along the West African coast, sardine biomass (in decline), round sardinella (declines from 1999-2007), and flat sardinella (decline in 2004-2010) fluctuated markedly. In contrast, catches of Atlantic horse mackerel and Cunene horse mackerel (Trachurus trecae) increased, with catch peaks in 1996-1997, 2000, 2005, and 2009 (Sambe, 2016).
In summary, the pronounced variability of fishing catches was intermittently documented from the 1950s to the 1990s in European and African coastal waters. In European Atlantic waters, peaks of comparable abundances have been noted in the North Sea (since the mid-1950s, in the 1970s and 1990s), and in several southern European countries extraordinary abundances of grey triggerfish and boarfish were observed in the 1950s and 1970s. Along the African coasts, up to the Senegal-Gulf of Guinea region, the first detected changes were in Western Sahara from the 1960s, culminating in two species group substitutions in the early 1970s: in pelagic catches, the massive expansion of sardine displacing Scomber and Sardinella; and in demersal catches with cephalopods eventually replacing sparids. Further south, in the mid-1980s, there is also a record of octopus expansion on the coastal grounds of Gambia-Senegal. Several critical episodes of species alternation were described in the Gulf of Guinea over three key periods. In the early 1970s versus late 1980s (alternation between round sardinella versus grey triggerfish) and the first decade of the 2010s (decline of round sardinella and chub mackerel versus an increase of Atlantic horse mackerel and anchovy).
Comparing these fishery fluctuations with the situation in southern Africa (Angola-Namibia-South Africa) –at the Eastern Tropical Atlantic/Temperate Southern Africa boundary– we find a similar change in migratory behavior of African Atlantic ichthyofauna during the 1950s-1980s in fisheries. With environmental modulation affecting several key fish populations, showing long-term changes in their distribution and abundance: southward migrations (between the late 1950s and early 1970s) and northward migrations (from the mid-1970s) (Shelton et al., 1985[323]; Shannon et al., 1988[324]; Shannon & Tauton-Clark, 1989[325]).
4. Some Examples and Proposals for the Future in a Global Warming Context
Some opinions by the American Oscar E. Sette (1900-1972) from 1961 are still relevant:
“...it is my belief that in fishery oceanography the challenge and the opportunity lies in studying the changing sea rather than the equilibrium ocean, and in studying the biological consequences of the changes at various trophic levels. […] In the aggregate this implies the necessity of observation of physical and chemical properties of sea water, its motions and mixings, and the numbers, kinds, and perhaps stages of the biota inhabiting the waters, all with space and time continuity sufficient to describe the events that take place and to investigate their inter-relationships.” (In: Boehlert & Schumacher, 1997).
Although there is abundant scientific evidence that climate and human activities altered marine ecosystems for thousands of years before the pre-industrial world (with low levels of CO2 emissions), differentiating between natural climate variability and human-induced changes remains a challenge in the 21st century. For that reason, there are European projects/programs such as 4-OCEANS–A Human History of Marine Life (“aims to assess the importance of marine life for human societies during the last two millennia, up to c.1860 CE”)[326], and Q-MARE-Disentangling climate and pre-industrial human impacts on marine ecosystems, which also explore “the timing and scale of human impacts on Quaternary marine ecosystems by combining paleontological, paleoclimatic, archaeological, and historical data”[327].
It is still necessary to promote the field of fishery oceanography internationally (Lehodey et al., 2006[328]; Bograd et al., 2023[329]), at least during the last hundred years, with mutual collaboration between fisheries scientists, oceanographers, meteorologists and researchers who are experts in the marine populations history (the most sensitive indicators of climatic change in the ocean), the marine environmental history, together with specialists in paleo-life reconstructions and paleoclimatic fluctuations. With these last two scientific specialties, great advances have been made for the Iberian Peninsula (IP) and adjacent seas, e.g. a review of paleoclimate reconstructions in the IP shows the last glaciation and deglaciation had a great impact on the regional climate (Cacho et al., 2010[330]). In the Mediterranean waters it has been possible to distinguish several episodes of colder and warmer climate of the last 700 kyr (Real et al, 2021[331]), with vestiges of its corresponding fauna and the confirmation that a good number of the Mediterranean marine species traditionally considered “endemic” are truly paleoendemic and currently shared with the coasts of America, Asia and Australia. Also, the current Mediterranean planktonic community contains 40 dinoflagellate taxa considered to be potential Indo-Pacific origin (Gómez, 2006[332]).
During the whole Quaternary period (Pleistocene and Holocene periodes), in the Mediterranean the alternation of cooling/heating cycles (ice ages/warm interglacial) resulted in different immigration occurrences of Atlantic fauna, respectively of boreal or subtropical origin (Würtz, 2010[333]). Other authors (Zazo et al, 2013[334], Zazo, 2015[335]) have described the great sea-level changes that also occurred throughout the Quaternary period in the Mediterranean and Atlantic coasts of Spain, and confirmed that during the warmest part of the last MIS-5 interglacial, faunal migrations from the tropics to the poles occurred globally. In the coastal deposits of the Spanish Mediterranean belonging to the Middle Pleistocene, species of mollusks from tropical Africa ("Senegalese" fauna) appear, useful for estimating the average surface values of sea water at that geological time, with high temperature and low salinity. Originating from previous geological epochs of extreme ocean warming (the Early Carboniferous) in the current southern region of Spain assemblages of tropical rough corals remain emerged in different coastal sectors. Those corals were established because of colonization that occurred during that oldest documented warming of the early ocean (Herbig and Mamet, 1985[336]), and diversified notably in the Paleotethys due to the extension of the tropical regions and sea invasion of large continental shelves, providing many new ecological niches (Somerville and Rodríguez, 2007[337]).
A totally opposite environmental scenario occurred in this Spanish geographical region, transition between the Atlantic and the Mediterranean, during the geological periods of maximum cooling at the poles with the expansion of their respective frozen surfaces, which forced numerous species of cold-water marine fauna to migrate towards the ice-free temperate latitudes. Particularly in the cave of Nerja (Málaga), archaeological evidence of marine fauna has been found that demonstrates the dramatic environmental and faunal changes in two different geologic time scales of extreme cooling. During the Pliocene-Early Pleistocene-Holocene human inhabitants captured species that are currently typical of the North Atlantic, eg. the extinct great auk Pinguinus impennis, Phoca vitulina, and fish of the Salmonidae and Gadidae families (Morales et al., 1994[]; Morales-Muñiz et al. 2008[338]; Jordá J. et al. 2011[339]). On the other hand, during the Last Glacial Period (Upper Magdalenian), in the same cave it is also documented (indirectly by two whale barnacle species) the consumption of whale meat and fat of the today endemic Antarctic species Eubalena australis (Álvarez-Fernández et al. 2014[340]).
Over the Alboran Sea/Western Mediterranean there have also been advances on broad geological millennial time scale. The variability of the western Mediterranean Sea surface temperature during the last 25 kyr have been related to the Northern Hemisphere climatic changes (Cacho et al, 2001[341]). The heating/cooling cycles of the surface temperatures of the Alboran Sea have been reconstructed over the last 50 kyr (Moreno et al., 2005[342]) and between 250 and 150 kyr (González-Mora et al., 2006[343]).
On a global scale, for the past 5 kyr, the Holocene climatic change was also characterized by drastic warming and cooling cycles, which had a very negative impact on different world civilizations. During the last two millennia, two long extreme periods lasting several centuries have been distinguished: the Medieval Warm Period (MWP) and the Little Ice Age (LIA). The LIA had its most severe stage roughly from 1560-1850 and from its end slowly began the current age of global warming. Retrospective studies of those intense and long-lasting warming/cooling in Europe are also recommended. The LIA had three periods of exceptionally low solar activity, with largely destructive consequences for societies in the 15th to 18th centuries (Degroot, 2018[344]) and affected fisheries (eg. Holm et al., 2019[345]; Edvardsson et al. 2022[346]).
Besides, it is necessary to continue investigating and providing multidisciplinary observational facts (confirmed historical data in the past centuries or millennia), preferably using one of the current advanced ecosystem mathematical models (e.g. Moullec et al., 2019[347]), which can explore and predict a variety of research questions related to specific human activities, climate variability, biodiversity and invasive alien species. On the other hand, the currently minority hypothesis of natural global warming (NGW, driven by solar variability) should not be underestimated, and it is recommended to develop research in the field of solar-terrestrial physics, as a necessary complement to oceanographic, meteorological and biological research.
Research on a global scale must be complemented by research focused on regional and local scales. On a regional scale, it is also necessary to advance in the detection of ocean warming hotspots (Popova et al., 2016[348]). Regional studies on the thermal variability of seawater at centennial resolution are also necessary, particularly in areas such as NW Africa-Canaries whose ecosystems present great natural variability depending on upwelling (eg. Arístegui et al., 2009[349]). The upwelling intensity since the 19th century has been calculated for the Canaries latitude (Gallego et al., 2021[350], 2022[351]). At a local scale, more climate change indicators should be developed and detect both marine heat waves (Roselló et al., 2023[352]) and the less studied persisting extremely cold events (Kodra et al., 2011[353]), which tend to go unnoticed in the average values of regional atmospheric temperatures and in global satellite images.
Paradoxically, in southern Europe marine paleontological research is increasing and there is still very little research on fish and fishing vs climate changes during the 16th to 19th centuries. We remember the same pending tasks that the two ICES scientists recognized 75 years ago, the need to investigate environmental changes in the marine environment also in the 19th century: “Apparently there have been certain periods of years in northern areas with higher temperatures and simultaneously increasing occurrence of southern species, for instance in the years about 1820-30, 1840-50, 1870-80 and 1920-?. All data relating to such periodicities will certainly prove to be of great importance” (Rollefsen & Tåning, 1948[354]). Evidence of this warming/cooling of the sea in the last centuries was recorded in both ichthyological and seaweed texts of naturalists from several European countries and in the academic publications of different ancient institutions, as well as in the historical natural collections (fauna and macroalgae) of numerous national museums.
Taking into account our historical review in marine ecosystems, at interdecadal and regional scales, it is foreseeable that when the current slow tropicalization processes (increases of warm-water species) and deborealization (decreases of cold-water species) finish, the opposite “normalization” processes (detropicalizacion/borealization) may occur at medium or long term. Methodological advances related to global change have been recently proposed to unravel these cyclical processes (McLean et al. 2021[355]), as well as the use of new biological indices (eg. Cheung et al., 2013[356]; Chust et al., 2024[357]) and various modeling techniques (eg. Sánchez-Garrido et al., 2019[358]; Derhy et al., 2022[359]; Townhill et al. 2023[360]).
A final recommendation. In the ocean warming presente frame, it is very necessary to increase the short term research on the future socioeconomic implications derived from overfishing and climate changes. In the southern European-African studied area, valuable prospectives projections have been published (eg. Allison et al., 2009[361]; Cheung, 2012[362]; Raybaud, 2017[363]; Erauskin-Extramiana et al., 2019[364]; Townhill, 2023[365]). Several of these authors agree in the idea that sub-Saharan countries will be the most affected area in case of ocean warming and overfishing continuity for more decades. Because of most of these countries, characterized by their poverty and the unstoppable increase in their population, are among the most vulnerable in the world, mainly due to their great dependence on fish in their diet and the uncontrolled development of the fishmeal industry and also the so-called migrant fishery. For all these reasons, it has been estimated that around the year 2050, the current high exports of fish from these African countries abroad (thousands of tonnes annually) will be drastically reduced, including the European markets (Failler, 2020[366]).
References
- Ocean Past Initiative. Tracing human interactions with marine ecosystems through deep time: implications for policy and management. In: Oceans Past VII. International Conference in Bremerhaven, Germany (22-26 October 2018). 2018. https://www.awi.de/forschung/besondere-gruppen/wissensplattform-erde-und-umwelt/opp7.html. https://www.tcd.ie/history/opi/.
- Pitcher, T.J. Fisheries managed to rebuild ecosystems? Reconstructing the past to salvage the future. Ecological Applications 2001, 11, 2, 601-617.
- Gordon, T.A.C.; Harding, H.R.; Clever, F.K.; Davidson, I.K.; Davison, W.; Montgomery, D.W.; Weatherhead, R.C.; Windsor, F.M.; Armstrong, J.D.; Bardonnet, A.; et al. Fishes in a changing world: learning from the past to promote sustainability of fish populations. J. Fish Biol. 2018, 92, 804–827. [CrossRef]
- Cooke, S.J.; Fulton, E.A.; Sauer, W.H.H.; Lynch, A.J.; Link, J.S.; Koning, A.A.; Jena, J.; Silva, L.G.M.; King, A.J.; Kelly, R.; et al. Towards vibrant fish populations and sustainable fisheries that benefit all: learning from the last 30 years to inform the next 30 years. Rev. Fish Biol. Fish. 2023, 33, 317–347. [CrossRef]
- Holm, P.; Smith, T.D.; Starkey, D.J. The exploited seas: new directions for marine environmental history. International Maritime Economic History Association. St. John’s (Canada). 2001 .
- Holm, P.; Marboe, A.H; Poulsen, B.; MacKenzie, B. Marine animal populations: a new look back in time. In Life in the World's Oceans: Diversity, Distribution and Abundance. Wiley-Blackwell. Wiley-Blackwell, Oxford, 2010, pp. 3-23.
- Starkey, D.J.; Holm, P; Barnard, M. (eds.) 2008. Ocean Past: management insights from the history of marine animal populations. Earthscan. New York.
- Lotze, H.K.; Mcclenachan, L. Marine historical ecology: informing the future by learning from the past. Marine community ecology and conservation, 2013, 165-201.
- ICES. 2018. Report of the Working Group on the History of Fish and Fisheries (WGHIST), 4–7 September 2017, Lysekil, Sweden. ICES CM 2017/SSGEPI, 19. 56 p.
- Southward A.J.; Langmead O.; Hardman-Mountford, N.J.; Aiken J.; Boalch, G.T.; Dando, P.R.; Genner, M,J.; Joint, I.; Kendall, M.A.; Halliday, N.C.; Harris, R.P.; Leaper, R.; Mieszkowska, N.; Pingree, R.D.; Richardson, A.J.; Sims, D.W.; Smith, T.; Walne, A.W.; Hawkins, S.J. Long-term oceanographic and ecological research in the Western English Channel. Adv Mar Biol. 2005, 47, 1-105. doi: 10.1016/S0065-2881(04)47001-1.
- Poulsen, B.; Holm, P.; MacKenzie, B.R. A long-term (1667–1860) perspective on impacts of fishing and environmental variability on fisheries for herring, eel, and whitefish in the Limfjord, Denmark. Fish. Res. 2007, 87, 181–195. [CrossRef]
- Philippart, C.; Anadón, R.; Danovaro, R.; Dippner, J.; Drinkwater, K.; Hawkins, S.; Oguz, T.; O'Sullivan, G.; Reid, P.C. Impacts of climate change on European marine ecosystems: Observations, expectations and indicators. J. Exp. Mar. Biol. Ecol. 2011, 400, 52–69. [CrossRef]
- Malta, T.; Santos, P.; Santos, A.; Rufino, M.; Silva, A. Long-term variations in Ibero-Atlantic sardine ( Sardina pilchardus ) population dynamics: Relation to environmental conditions and exploitation history. Fish. Res. 2016, 179, 47–56. [CrossRef]
- Van Beveren, E.; Fromentin, J.-M.; Rouyer, T.; Bonhommeau, S.; Brosset, P.; Saraux, C. The fisheries history of small pelagics in the Northern Mediterranean. ICES J. Mar. Sci. 2016, 73, 1474–1484. [CrossRef]
- Fortibuoni, T.; Libralato, S.; Arneri, E.; Giovanardi, O.; Solidoro, C.; Raicevich, S. Fish and fishery historical data since the 19th century in the Adriatic Sea, Mediterranean. Sci. Data 2017, 4, 170104–170104. [CrossRef]
- Hernvann, P.-Y.; Gascuel, D. Exploring the impacts of fishing and environment on the Celtic Sea ecosystem since 1950. Fish. Res. 2020, 225, 105472. [CrossRef]
- Sánchez-Garrido, J.; Werner, F.; Fiechter, J.; Rose, K.; Curchitser, E.; Ramos, A.; Lafuente, J.G.; Arístegui, J.; Hernández-León, S.; Santana, A.R.; et al. Decadal-scale variability of sardine and anchovy simulated with an end-to-end coupled model of the Canary Current ecosystem. Prog. Oceanogr. 2018, 171, 212–230. [CrossRef]
- Pérez-Rubín, J.; A Historical Approach to Living Resources on the Spanish Coasts from the Alboran Sea Between the Sixteenth and Twentieth Centuries. In: Alboran Sea Ecosystems and Marine Resources. Báez, J.C; Vázquez, J-T.; Camiñas, J.A.; Malouli, M., Eds. Springer Nature, Switzerland. 2021, pp. 775-795.
- Thurstan, R.H. The potential of historical ecology to aid understanding of human-ocean interactions throughout the Anthropocene. J. Fish Biol. 2022. [CrossRef]
- Finney, B.P.; Alheit, J.; Emeis, K.-C.; Field, D.B.; Gutiérrez, D.; Struck, U. Paleoecological studies on variability in marine fish populations: A long-term perspective on the impacts of climatic change on marine ecosystems. J. Mar. Syst. 2010, 79, 316–326. [CrossRef]
- Barrett, J.H. An environmental (pre)history of European fishing: past and future archaeological contributions to sustainable fisheries. J. Fish Biol. 2019, 94, 1033–1044. [CrossRef]
- Pauly, D. Anecdotes and the shifting baseline syndrome of fisheries. Trends Ecol. Evol. 1995, 10, 430. [CrossRef]
- Pinnegar, J.K.; Engelhard, G.H. The ‘shifting baseline’ phenomenon: a global perspective. Rev. Fish Biol. Fish. 2007, 18, 1–16. [CrossRef]
- Bonebrake, T.C.; Christensen, J.; Boggs, C.L.; Ehrlich, P.R. Population decline assessment, historical baselines, and conservation. Conserv. Lett. 2010, 3, 371–378. [CrossRef]
- Soga, M.; Soga, M.; Gaston, K.J.; Gaston, K.J. Shifting baseline syndrome: causes, consequences, and implications. Front. Ecol. Environ. 2018, 16, 222–230. [CrossRef]
- Gilliland, R. L. Solar, volcanic and CO2 forcing of recent climatic changes. Climatic Change, 1982, 4, 111-131.
- Carbon Dioxide Review: 1982, W. C. Clark (ed.), Oxford University Press, 427 pp. Changing Climate: Report of the Carbon Dioxide Assessment Committee: 1983, National Research Council, National Academy Press, Washington D.C., 496 pp.
- Sherman K. and G. Hempel. 2008. Perspectives on Regional Seas and the Large Marine Ecosystem Approach. In: Sherman, K. and Hempel, G. (Editors) 2008. The UNEP Large Marine Ecosystem Report: A perspective on changing conditions in LMEs of the world’s Regional Seas. UNEP Regional Seas. Report and Studies, 182: 3-21.
- https://www.clivar.org/.
- http://www.ipcc.ch/.
- http://www.igbp.net/.
- Glantz, M.H.; Feingold, L.E. Climate variability, climate change, and fisheries: a summary. In: Climate Variability, Climate Change and Fisheries. Cambridge: Cambridge University Press. 1992, pp. 417-438.
- Everett, J.T.; A, Krovnin; D, Lluch-Belda.; E, Okemwa.; H.A, Regier.; and J.-P Troadec. Climate Change 1995: Impacts, Adaptations, and Mitigation of Climate Change: Scientific-Technical Analyses. In: Contribution of Working Group II to the Second Assessment Report of the IPCC. Watson, R.T.; Zinyowera M.C.; Moss, R.H. Eds. Cambridge University Press, New York, USA. 1996, 511-537.
- https://www.unep.org/unepmap/.
- https://eurogoos.eu/.
- http://www.esf.org/marineboard.
- https://argo.ucsd.edu/.
- IPCC. 2023. Climate Change 2021. The Physical Science Basis. Working Group I Contribution to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press.
- Resumen Ejecutivo de CLIVAR-España 2019. El clima en la Península Ibérica.
- National Research Council (NRC). The Effects of Solar Variability on Earth's Climate: A Workshop Report; The National Academies Press: Washington, DC, USA, 2012. [CrossRef]
- Kusano, K. (ed.). Solar-Terrestrial Environmental Prediction. Springer Nature Singapore. 2023, XXIV, pp. 462. [CrossRef]
- Barange, M.; Bahri, T.; Beveridge, M. C. M.; Cochrane, K. L.; Funge-Smith, S.; Poulain, F. Impacts of climate change on fisheries and aquaculture. Synthesis of current knowledge, adaptation and mitigation options. FAO Fisheries and Aquaculture Technical Paper, 2018, 627,1-628.
- IPCC, 2019. IPCC Special Report on the Ocean and Cryosphere in a Changing Climate [H.-O. Pörtner, D.C. Roberts, V. Masson-Delmotte, P. Zhai, M. Tignor, E. Poloczanska, K. Mintenbeck, A. Alegría, M. Nicolai, A. Okem, J. Petzold, B. Rama, N.M. Weyer (eds.)].
- Bindoff, N. L.; Cheung, W. W. L.; Arístegui, J. G. K. J.; Guinder, V. A.; Hallberg, R.; Hilmi, N.; Williamson, P. Chapter 5: Changing Ocean, Marine Ecosystems, and Dependent Communities. In: IPCC Special Report on the Ocean and Cryosphere in a Changing Climate. Pörtner, D.C.; Roberts, V.; Masson-Delmotte, P.; Zhai, M.; Tignor, E.; Poloczanska, K.; Mintenbeck, A.; Alegría, M.; Nicolai, A.; Okem, J.; Petzold, B.; Rama, N.M. Weyer.; Eds. Cambridge University Press, Cambridge, UK and New York, NY, USA, 2019; vol 1155, 447-587. [CrossRef]
- Cooley, S.; Schoeman, D.; Bopp, L.; Boyd P.; Donner, S.; Ghebrehiwet, D.Y.; Ito, S.I.; Kiessling, W.; Martinetto, P.; Ojea, E.; Racault, M.-F.; Rost, B.; Skern-Mauritzen, M. 2022: Oceans and Coastal Ecosystems and Their Services. In: Climate Change 2022: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change [H.-O. Pörtner, D.C. Roberts, M. Tignor, E.S. Poloczanska, K. Mintenbeck, A. Alegría, M. Craig, S. Langsdorf, S. Löschke, V. Möller, A. Okem, B. Rama (eds.)]. Cambridge University Press, Cambridge, UK and New York, NY, USA, 2023, 379–550. [CrossRef]
- Global Environment Monitoring System & World Water Quality Alliance: Freshwater, Air and Ocean. 2022. https://wedocs.unep.org/handle/20.500.11822/35879.
- https://marine.copernicus.eu/about#copernicus-marine-service.
- www.oceandecade.org.
- https://climate.copernicus.eu/.
- https://climate.copernicus.eu/sites/default/files/custom-uploads/ESOTC%202023/Summary_ESOTC2023.pdf.
- https://climate.copernicus.eu/esotc/2023/european-ocean.
- Ocean Decade Vision 2030. Draft Outcomes Report and White Papers (January 2024). https://oceanexpert.org/document/33599.
- UNESCO-IOC. The United Nations Decade of Ocean Science for Sustainable Development (2021-2030): Implementation Plan. Ocean Decade Science-Policy Interface Guidelines. Paris: United Nations Educational, Scientific and Cultural Organization. 2021. url: https://unesdoc.unesco.org/ark:/48223/pf0000377082.
- UNESCO-IOC. Ocean Decade Progress Report 2022–2023. UNESCO, Paris. The Ocean Decade Series, 2023, 46.
- UNESCO, 2017. Global Ocean Science Report—The current status of ocean science around the world.
- IOC-UNESCO. 2020. Global Ocean Science Report 2020–Charting Capacity for Ocean Sustainability.
- Potter, R.W.K.; Pearson, B.C. Assessing the global ocean science community: understanding international collaboration, concerns and the current state of ocean basin research. npj Ocean Sustain. 2023, 2, 1–12. [CrossRef]
- https://oceanexpert.org/event/3430.
- Schroeder, K.; Cardin, V.; Font, J.; Fuda, J.; Garcia Lafuente J.; Puig, P.; Taupier-Letage, I; Bengara, L.; Ismail, S. B; Bensi, M.; et al. Hydrochanges network: latest observations and links between time series. Rapp. Comm. Int. Mer Medit. 2010, 39, 180.
- Schroeder, K.; Millot, C.; Bengara, L.; Ben Ismail, S.; Bensi, M.; Borghini, M.; Budillon, G.; Cardin, V.; Coppola, L.; Curtil, C.; et al. Long-term monitoring programme of the hydrological variability in the Mediterranean Sea: a first overview of the HYDROCHANGES network. Ocean Sci. 2013, 9, 301–324. [CrossRef]
- Schroeder, K.; Chiggiato, J.; Bryden, H.L.; Borghini, M.; Ben Ismail, S. Abrupt climate shift in the Western Mediterranean Sea. Sci. Rep. 2016, 6, 23009. [CrossRef]
- Schroeder, K.; Chiggiato, J.; Josey, S.A.; Borghini, M.; Aracri, S.; Sparnocchia, S. Rapid response to climate change in a marginal sea. Sci. Rep. 2017, 7, 4065. [CrossRef]
- CIESM 2008. Climate warming and related changes in Mediterranean marine biota. CIESM Workshop Monographs, 2008, 35: 1-152.
- https://www.ciesm.org/marine/programs/tropicalization.htm.
- https://ciesm.org/online/atlas/.
- Golani, D.; Azzurro, E.; Dulčić, J.; Massutí, E.; Orsi-Relini, L. Atlas of Exotic Fishes in the Mediterranean Sea. F. Briand.; Ed. CIESM Publishers, Monaco. 2021, pp. 365.
- Galil, B.; Froglia, C.; Noël, P. Vol. 2: Crustaceans: decapods and stomatopods. In: CIESM Atlas of Exotic Species in the Mediterranean. Briand, F.; Ed. CIESM Publishers, Monaco. 2002, pp. 192.
- Zenetos, A.; Gofas, S.; Russo, G.; Templado, J. Vol. 3: Molluscs. In: CIESM Atlas of Exotic Species in the Mediterranean. Briand, F. Ed. CIESM Publishers, Monaco. 2003, pp. 376.
- Verlaque, M.; Ruitton, S.; Mineur, F.; Boudouresque, Ch-F. Vol. 4: Macrophytes. In: CIESM Atlas of Exotic Species. Briand, F.; Ed, CIESM Publisher, Monaco. 2015, pp. 362.
- Rijnsdorp, A. D., Peck, M. A., Engelhard, G. H., Möllmann, C., and Pinnegar, J. K. (Eds). 2010. Resolving climate impacts on fish stocks. ICES Cooperative Research Report, 301: 1-371.
- Drinkwater, K.F. Marine European climate: past, present, and future. Resolving climate impacts on fish stocks. In: ICES Cooperative Research Report. Rijnsdorp, A. D.; Peck, M. A.; Engelhard, G. H.; Möllmann, C.; and Pinnegar, J. K.; Eds. 2010, 301, 49-65.
- Pinnegar, J.K.; Engelhard, G. H.; Daskalov, G.M. Changes in the distribution of fish. Resolving climate impacts on fish stocks. In: ICES Cooperative Research Report. Rijnsdorp, A. D.; Peck, M. A.; Engelhard, G. H.; Möllmann, C.; and Pinnegar, J. K.; Eds. 2010, 301, 94-110.
- Predragovic, M.; Cvitanovic, C.; Karcher, D.B.; Tietbohl, M.D.; Sumaila, U.R.; e Costa, B.H. A systematic literature review of climate change research on Europe's threatened commercial fish species. Ocean Coast. Manag. 2023, 242. [CrossRef]
- Meckler, A.N.; Trofimova, T.; Risebrobakken, B. & S. De Schepper. 14th International Conference on Paleoceanography. Past Global Changes Magazine, 2023, 31, 2, 116-117. [CrossRef]
- https://www.historicalclimatology.com/cliwoc.html.
- https://pastglobalchanges.org/about/general-overview.
- https://water.europa.eu/marine.
- https://www.eea.europa.eu/data-and-maps/figures/european-marine-climate-change-index-emcci.
- Heip, C.; Barange, M.; Danovaro, R.; Gehlen, M.; Grehan, A.; Meysman, F.; Oguz, T.; Papathanassiou, V.; Philippart, C.; She, J.; et al. Climate Change and Marine Ecosystem Research. Synthesis of European Research on the Effects of Climate Change on Marine Environments. Marine Board Special Report. 2011. Ostend, Belgium, 1-151.
- https://www.ecmwf.int/en/about.
- http://ciesm.org/online/GISBiblio.php.
- https://ices-library.figshare.com/.
- Grainger, R.J.R.; Garcia, S.M. Chronicles of marine fishery landings (1950-1994): Trend analysis and fisheries potential. FAO Fisheries Technical Paper, 1996, 359, 1-51.
- https://www.cprsurvey.org/1409.
- Richardson, A.; Walne, A.; John, A.; Jonas, T.; Lindley, J.; Sims, D.; Stevens, D.; Witt, M. Using continuous plankton recorder data. Prog. Oceanogr. 2006, 68, 27–74. [CrossRef]
- http://biblioteca.ieo.es/biblio_ext.html.
- https://digital.csic.es/handle/10261/152009 ; https://digital.csic.es/handle/10261/158325.
- Pérez-Rubín, J. 2020. Investigaciones ictiológicas y pesqueras de la familia Lozano en el MNCN y su faceta divulgadora (1912-1970). In: Martín Albaladejo C. (ed.). Del elefante a los dinosaurios. 45 años de historia del Museo Nacional de Ciencias Naturales 1940-1985: 105-171.
- Tel E. Los repositorios marinos del IEO: la larga trayectoria de la Red de Mareógrafos y el Centro Español de Datos Oceanográficos durante más de 50 años. In: El estrecho de Gibraltar: llave natural entre dos mares y dos continentes. Pérez-Rubín J. & Teodoro Ramírez T. Eds. Bol. R. Soc. Esp. Hist. Nat. 2023, 16, 385-394.
- Valdés, L.; Lavín, A.; Fernández de Puelles, M.L.; Varela, M.; Anadón, R.; Miranda, A.; Camiñas, J.; Mas, J. Spanish Ocean Observation System. IEO Core Project: Studies on time series of oceanographic data. In: Operational Oceanography. Implementation at the European and Regional Scales. Flemming N.; Vallerga S.; Pinardi N.; et al.; Eds. 2002, pp. 99-105.
- IGN. 1991. El Medio Marino. In: Atlas Nacional de España, Sección III (13). Instituto Geográfico Nacional, MOPT, Madrid.
- https://repositorio.aemet.es/.
- https://www.aemet.es/en/serviciosclimaticos/datosclimatologicos/atlas_climatico.
- https://www.ipcc.ch/reports/.
- http://clivar.es/?page_id=1253.
- https://obis.org/about/.
- Smith T. D. Scaling Fisheries. The Science of Measuring the Effects of Fishing, 1855-1955. Cambridge University Press. 1994.
- Bigelow, H. Oceanography. Its Scope, Problems, and Economic Importance. The Riverside Press Cambridge, Boston. 1931.
- Garstang, W. The Impoverishment of the Sea. A Critical Summary of the Experimental and Statistical Evidence bearing upon the Alleged Depletion of the Trawling Grounds. J. Mar. Biol. Assoc. United Kingd. 1900, 6, 1–69. [CrossRef]
- Hjort, J. Fluctuations in the great fisheries of northern Europe viewed in the light of biological research. ICES. 1914, 20, 1–228.
- Petterson, O.; Pettersson, O. Climatic variations in historic and prehistoric time. Springer Berlin Heidelberg. 1914, 1-27. [CrossRef]
- Jones, P.D.; Wigley, T.M.L.; Wright, P.B. Global temperature variations between 1861 and 1984. Nature 1986, 322, 430–434. [CrossRef]
- Johannessen, O.M.; Bengtsson, L.; Miles, M.W.; Kuzmina, S.I.; Semenov, V.A.; Alekseev, G.V.; Nagurnyi, A.P.; Zakharov, V.F.; Bobylev, L.P.; Pettersson, L.H.; Hasselmann, K.; Cattle, H.P. Arctic climate change: observed and modelled temperature and sea-ice variability. Tellus, 2004, 56, 4, 328–341.
- Hughes, S.L.; Holliday, N.P.; Gaillard, F. Variability in the ICES/NAFO region between 1950 and 2009: observations from the ICES Report on Ocean Climate. ICES J. Mar. Sci. 2012, 69, 706–719. [CrossRef]
- Hodson, D.L.R.; Robson, J.I.; Sutton, R.T. An Anatomy of the Cooling of the North Atlantic Ocean in the 1960s and 1970s. J. Clim. 2014, 27, 8229–8243. [CrossRef]
- Lluch-Belda, D.; Lluch-Cota, S.; LLuch-Cota, D.; Hernández-Vázquez, S. La variabilidad oceánica interanual y su impacto sobre las pesquerías. Rev. Soc. Mex. Hist. Natural, 1999, 49, 219-227.
- Cort, J.L.; Abaunza, P. The fall of the tuna traps and the collapse of the Atlantic Bluefin Tuna, Thunnus thynnus L.; fisheries of Northern Europe from the 1960s. Rev. Fish. Sci. Aquacult. 2015, 23, 4, 346-373. [CrossRef]
- Cort, J. L.; Abaunza, P. The bluefin tuna fishery in the Bay of Biscay. Its relationship with the crisis of catches of large specimens in the East Atlantic Fisheries from the 1960s. Springer Briefs in Biology. 2019. [CrossRef]
- Philippart, C.; Anadón, R.; Danovaro, R.; Dippner, J.; Drinkwater, K.; Hawkins, S.; Oguz, T.; O'Sullivan, G.; Reid, P.C. Impacts of climate change on European marine ecosystems: Observations, expectations and indicators. J. Exp. Mar. Biol. Ecol. 2011, 400, 52–69. [CrossRef]
- Belkin, I.M. Rapid warming of Large Marine Ecosystems. Prog Oceanogr. 2009, 81, 207–213. [CrossRef]
- Nykjaer, L. Mediterranean Sea surface warming 1985–2006. Clim. Res. 2009, 39, 11–17. [CrossRef]
- Mann, M.E.; Jones, P.D. Global surface temperatures over the past two millennia. Geophys. Res. Lett. 2003, 30. [CrossRef]
- Moberg, A.; Sonechkin, D.M.; Holmgren, K.; Datsenko, N.M.; Karlén, W. Highly variable Northern Hemisphere temperatures reconstructed from low- and high-resolution proxy data. Nature 2005, 433, 613–617. [CrossRef]
- Bakun, A. Global Climate Change and Intensification of Coastal Ocean Upwelling. Science 1990, 247, 198–201. [CrossRef]
- Russell, F.S.; Southward, A.J.; Boalch, G.T.; Butler, E.I. Changes in Biological Conditions in the English Channel off Plymouth during the Last Half Century. Nature 1971, 234, 468–470. [CrossRef]
- Southward, A. Fluctuations in the “indicator” chaetognaths Sagitta elegans and Sagitta setosa in the Western Channel. Oceanol. Acta, 1984, 7 (2), 220-239.
- Hansen, P. M. The stock of cod in Greenland waters during the years 1924-1952. Rapp. Reun. Cons. int. Explor. Mer. 1954, 136, 65–71.
- Binet, D.; Leroy, C. La pêcherie du ton rouge (Thynnus thynnus) dans l’Atlantique nord-est était-elle liée au réchauffement séculaire. ICCAT/SCRS, 1986, 86 (52), pp. 16.
- MacKenzie, B.R.; Myers, R.A. The development of the northern European fishery for north Atlantic bluefin tuna Thunnus thynnus during 1900–1950. Vol. Sci. Pap. ICCAT, 2009, 63, 233-234. [CrossRef]
- Crowley, T.J.; Lowery, T.S. How Warm Was the Medieval Warm Period?. AMBIO 2000, 29, 51–54. [CrossRef]
- Drinkwater, K. F. Comparing the response of Atlantic cod stocks during the warming period of the 1920s-1930s to that in the 1990s-2000s. Deep Sea Res, 2009, 56, 2087-2096.
- Polyakov, I.V.; Alexeev, V.A.; Bhatt, U.S.; Polyakova, E.I.; Zhang, X. North Atlantic warming: patterns of long-term trend and multidecadal variability. Clim. Dyn. 2010, 34, 439–457. [CrossRef]
- Yamanouchi, T. Early 20th century warming in the Arctic: A review. Polar Sci. 2011, 5, 53–71. [CrossRef]
- Fridiksson, A. Boreo-tended changes in the marine vertebrata fauna of Iceland during the last twenty five years. Rapports et Procès-verbaux des Réunions. ICES, 1949, 125, 30-32.
- Johansen, A.C. On the Remarkable Quantities of Haddock in the Belt Sea during the Winter of 1925-26, and Causes leading to the same. ICES J. Mar. Sci. 1926, 1, 140–156. [CrossRef]
- Walford, L. A. Northward occurrence of southern fish off San Pedro in 1931. Calif. Fish. Game. 1931, 17, 401-405.
- McKenzie, R. A.; Homans, R. Rare and interesting fishes and salps in the Bay of Fundy and off Nova Scotia. Proc. N. S. Inst. Sci. 1938, 19 (3), 277.
- Lumby, J.R.; Atkinson, G.T. On the unusual Mortality amongst Fish during March and April 1929, in the North Sea. ICES J. Mar. Sci. 1929, 4, 309–332. [CrossRef]
- Jensen, A. S. Concerning a change of climate during recent decades in the artic and subarctic regions, from Greenland in the West to Eurasia in the East and contemporary biological and geophysical changes. Det Konglike Videnskabernes Selska Biologiske Meddelelser, 1939, 14 (8), 1-75.
- Cushing, D. H.; Dickson, R. R. Biological and hydrographic changes in British waters during the last thirty years. Biol. Rev. 1976a, 41 (2), 221-258.
- Stephen, A.C. Temperature and the Incidence of Certain Species in Western European Waters in 1932-1934. J. Anim. Ecol. 1938, 7, 125–129. [CrossRef]
- Desbrosses, P. Capture d’une Tortue (Dermatochelys coriacea) dans la baie d’Etel. Bull Soc Zool Fr. 1932, 57, 274-277.
- Desbrosses, P. Echouage d’un Tetrodon T. lagocephalus pres de Quiberon et remarques sur la présense de cette éspèce et de Balistes capriscus au nord du 44º L. N. Bull Soc Zool Fr. 1935, 60, 43-48.
- Desbrosses, P. Présence à l’entrée occidentale de la Manche de Callanthias ruber. Bull Soc Zool Fr. 1936, 61, 406-407.
- Buen, F. Fluctuaciones en la sardina Sardina pilchardus. Pesca y medidas. Notas Resum. 1929, 35, 1-80.
- Anadón, E. Sobre la sustitución alternativa en el litoral gallego de los llamados peces emigrantes sardina, espadín, anchoa y jurel. Bol. Inst. Esp. Oceanogr. 1950, 24, 1-20.
- ICES. Comparative Studies of the Fluctuations in the Stocks of Fish in the seas of North and West Europe. Conseil Internat. pour l’Explor. de la Mer. Rapp. Et Proc.-Verb. Des Réunions, 1936, CI, part 3.
- Kemp, S. Oceanography and the fluctuations in the abundance of marine animals. Report of the British Association for the Advancement of Science, 1938, 85-101.
- Carruthers, J, N. Fluctuations n the herrins of the East Anglian Autumm fishery, the yield of the Ostend spent herring fishery and the haddock of the North Sea in the light of the relevant wind conditions. Rapports et Procès-verbaux des Réunions. ICES, 1938, 107, 1-15.
- Simpson, A.C. Some Observations on the Mortality of Fish and the Distribution of Plankton in the Southern North Sea during the Cold Winter, 1946-1947. ICES J. Mar. Sci. 1953, 19, 150–177. [CrossRef]
- Woodhead, P. M. Changes in the behavior of the sole Solea vulgaris during cold winters and the relation between the winter catch and sea temperatures. Helgoländer wissenschaftliche Meeresuntersuchungen, 1964, 10, 328-342.
- ICES. Contributions to Special Scientific Meetings 1948. Climatic changes in the Arctic in relation to plants and animals. Rapp. Cons. Explor. Mer, 1949, 125, 5-51.
- Brooks, C. Climate through the ages. A study of the climatic factors and their variations. E. Benn limited, 1926. 395 pp.
- Hubbs, C. Changes in the fish fauna of Western North America correlated with changes in ocean temperature. J. Mar. Res. 1948, 459-482.
- Fitch, J. E. The decline of the pacific mackerel fishery. Calif. Fish Game 1952, 38, 381-403.
- Ketchen, K. Climate trends and fluctuations in yield of marine fisheries in the North Pacific. J. Fish. Res. Board. Can. 1956, 13, 357-374.
- Uda, M. On the relation between the variation of the important fisheries conditions and the oceanographical conditions in the adjacent waters of Japan. J. Tokyo. Univ. Fish. 1952, 38 (3), 364-389.
- Bell, F.H.; Pruter, A.T. Climatic temperature changes and commercial yields of some marine fishes. J. Fish. Res. Board. Can 1958, 15, 625-83.
- Schweigger E. Die Westküste Südamerikas im Bereich des Peru-Stroms. Keysersche Verlagsbuchhandlung GmbH, Heidelberg-München. 1959.
- Rollefsen, G. Fluctuations in two of the most important stocks of fish in northern waters, the cod and the herring. Rapports et Procès-verbaux des Réunions. ICES, 1949, 125, 33-35.
- Hansen, P. Studies on the biology of the cod in Greenland waters. Rapports et Procès-verbaux des Réunions. ICES, 1949, 5-77.
- Rasmussen, B. On the migration pattern of the West Greenland stock of cod. Ann. Biol. 1959, 14, 123.
- Cushing, D.H.; Burd, A.C. On the herring of the Southern North Sea. Fishery Invest. 1957, 20 (11), 1-31.
- Cushing, D.H. The number of pilchards in the Channel. Fishery Invest. 1957, 21 (5), 27.
- Rees, W.J. The Distribution of Octopus Vulgaris Lamarck in British Waters. J. Mar. Biol. Assoc. United Kingd. 1950, 29, 361–378. [CrossRef]
- Rees, W.J.; Lumby, J. R. The abundance of Octopus in the English Channel J. Mar. Biol. Ass. UK. 1954, 33, 515-136.
- Holme, N.A. The Bottom Fauna of the English Channel. J. Mar. Biol. Assoc. United Kingd. 1961, 41, 397–461. [CrossRef]
- Shurin, T.A. Characteristic features of the bottom fauna in the eastern Baltic in 1959. Ann. Biol. 1961, 16 (1959), 86.
- Meyer, P.F.; Kalle, K. Die biologische Umstimmung in der Ostsee in den letzen Jahrezenten, eine Folge hydrographischer Wasserumschictungen. Archiv für Fischereiwissenschaft, 1950, 2, 1-9.
- Wyrtki, K. Der grosse Salzeinbruch in die Ostsee im november un december 1951. Kieler Meeresforschungen, 1954, 10 (1), 19-25.
- Fischer-Piette, E. Sur les déplacements des frontièrs biogeographiques intercotidales observables en Espagne: situation en 1954-55. CR. Acad. Sci. Paris. 1955, 241, 447-449.
- Fisher-Piette E. Sur les déplacements des frontières biogéographiques intercotidales, actuellement en cours en Espagne: situation en 1956. CR. Acad. Sci. Paris. 1956, 242, 2782-2784.
- Fisher-Piette E. Sur les progrés des espèces septentrionales dans le bios inter-cotidale ibérique: situation en 1956-1957. CR. Acad. Sci. Paris. 1957a, 245, 373–375.
- Fisher-Piette E. Sur les déplacements de frontières biogéographiques, observables au long des cotes ibériques dans le domaine intercotidal. Inst. Biol. Aplic. 1957b, 26, 35–40.
- Fisher-Piette E. Sur l’écologie intercotidale ouest-ibérique. Comptes Rendus. Academie des Sciences, Paris, 1958, 245, 373–375.
- Fisher-Piette E. La distribution des principaux organismes nord iberiques en 1954-1955. Ann. Inst. Ocean. Monaco, 1963, 40 (3), 165-311.
- Navaz, J. Mª. Nuevos datos sobre la sustitución alternativa en la pesca de peces emigrantes en el litoral de Galicia. Notas Resum. 1946a, 132, 1-9.
- Navaz J. Mª. Sobre algunos peces poco frecuentes o desconocidos en las costas de Galicia. Notas Resum. 1946b, 133, 1-9.
- Navaz, J. Mª. Sobre algunos peces poco frecuentes o desconocidos en la costa vasca. Bol. Inst. Esp. Oceanogr. 1961, 106, 1-48.
- Carson. The Sea Around Us, Oxford University Press, USA, 1952, pp. 182.
- Conti A. Râcleurs d’Océans. Paris, Payot & Rivages. 1953.
- Sette, 0. E. The long term historical record of meteorological, oceanographic and biological data. Calif. Coop. Oceanic Fisheries Invest. Rep. 1960, 7, 181-194.
- Sette, 0. E. Problems in fish population fluctuations. Calif. Coop. Oceanic Fisheries Invest. Rep. 1961, 8, 21-24.
- Crisp, D.J. The Effects of the Severe Winter of 1962-63 on Marine Life in Britain. J. Anim. Ecol. 1964, 33, 165. [CrossRef]
- Rodewald, M. Sea-surface temperatures of the North Atlantic Ocean during the decade 1951–60. In: Changes of Climate, Proceedings of the Rome Symposium, Arid Zone Research, 1963, 20, 97–107. UNESCO, Paris.
- Stearns, F. Sea-surface temperature anomaly study of records from Atlantic coast stations. J. Geophys. Res. 1965, 70, 283–296. [CrossRef]
- Fairbridge, R.W. Mean sea level related to solar radiation during the last 20,000 years. In: Changes of Climate. Proceedings, Rome Symposium, 1960 UNESCO, Paris, 1963, 229–242.
- Beverton, R. J. H.; Lee, A. J. Hydrodynamic fluctuations in the North Atlantic Ocean and some biological consequences. In: Johnson C. G. & Smith L. P. eds. The Biological significance of Climatic Changes in Britain. Institute of Biology Symposia, 1965, 14, 79-107.
- Zupanovitch, S. Causes of fluctuations in sardine catches along the Eastern coast of the Adriatic Sea. Anali Jadranskog Instituta, 1968, 4, 401-489.
- Wyatt, T.; Porteiro, C. The Iberian Sardine Fisheries: Trends and Crises. In: Large Marine Ecosystems. Sherman, K.; Skjoldal, H.; Eds. Elsevier, 2002, 321-338.
- Cendrero, O. Sardine and anchovy crises in northern Spain: natural variations or an effect of human activities?. ICES Marine Science Symposia, 2002, 215, 279-285. [CrossRef]
- Hernvann, P.-Y.; Gascuel, D. Exploring the impacts of fishing and environment on the Celtic Sea ecosystem since 1950. Fish. Res. 2020, 225, 105472. [CrossRef]
- Lamb, H.H. Climate: Past, Present and Future. I. Fundamentals and Climate Now. 1972, pp. 1-624.
- Cushing, D.H. Marine Ecology and Fisheries. CUP Archive, 1975.
- Cushing, D.H.; Dickson R.R. The biological response in the sea to climatic changes. Adv. Mar. Biol. 1976b, 14, 1-122. [CrossRef]
- Mörner, N-A.; Karlén, W. 1984 Climatic changes on a yearly to millennial basis: geological, historical, and instrumental records. Proceedings of the II Nordic Symposium on Climatic Changes and Related Problems (Stockholm, Sweden, May 16-20, 1983). Reidel Pub., Dordrecht. 1984.
- Mörner, N.A. Oceanic circulation changes and redistribution of energy and mass on a yearly to centenary time-scale. Symposium Long Term Changes in Marine Fish Populations. Vigo, Spain. Contribution 3. 1986.
- Cushing, D.H. Climate and fisheries. Academic Press, London (UK). 1982. XIV, pp. 373.
- Caddy, J.; Gulland, J. Historical patterns of fish stocks. Mar. Policy 1983, 7, 267–278. [CrossRef]
- Kawasaki, T.; Omori, M. Fluctuations in the three major sardine stocks in the Pacific and the global trend in temperature. In: Proc. Long Term Changes in Marine Fish Populations. Vigo, Spain: Instituto de Investigaciones Marinas de Vigo. 1988, 37–53.
- Dickson, R.R.; Meinecke, J.; Malmberg, S.A.; Lee, A.J. The "great salinity anomaly" in the northern North Atlantic 1968-1982. Prog Oceanogr. 1988, 20, 103-151.
- Cushing, D. Population, production and regulation in the sea: A fisheries perspective. Cambridge University Press. 1995.
- Bakun, A.; Parrish, R.H. Environmental inputs to fishery population models for eastern boundary current regions. In: Workshop on the effects of environmental variation on the survival of larval pelagic fishes. Sharp, G.D. Ed. UNESCO, Paris. IOC Workshop Rep. 1980, 28, 67-104.
- Southward, A.J.; Boalch, G.T.; Maddock, L. Fluctuations in the herring and pilchard fisheries of devon and cornwall linked to change in climate since the 16th century. J. Mar. Biol. Assoc. United Kingd. 1988, 68, 423–445. [CrossRef]
- Backhaus, J.O. A three-dimensional model for the simulation of shelf sea dynamics. Dtsch. Hydrogr. Z. 1985, 38, 165–187. [CrossRef]
- Bartsch, J.; Brander, K.; Heath, M.; Munk, P.; Richardson, K.; Svendsen, E. Modelling the advection of herring larvae in the North Sea. Nature 1989, 340, 632–636. [CrossRef]
- Bardach, J.E.; Santerre, R.M. Climate and Aquatic Food Production. In: Food Climate Interactions. Bach, W.; Pankrath, I.; Schneider. S. H.; Eds. 1981, Kluwer Academic Publishers, Dordrecht, The Netherlands.
- Frye, R. Climatic change and fisheries management. Nat. Resour. J. 1983, 23, 77-96.
- Sibley, T.H.; Strickland, R.M. Fisheries: Some Relationships to Climate Change and Marine Environmental Factors. In: Characterization of Information Requirements for Studies of CO2 Effects: Water, Resources, Agriculture, Fisheries, Forests and Human Health. White, M. R.; Ed. United States Department of Energy, Carbon Dioxide Research Division, Washington, D.C.; 1985, 95-143.
- Shepherd, J.G.; Pope, J.G.; Cousen, R.D. Variations in fish stocks and hypotheses concerning their links with climate. Rapp. Procès-Verb. Réun. Cons. Int. Explor. Mer, 1984, 185, 255-267.
- Pérez-Rubín J. El Ictioplancton del mar de Alborán: Relación de su distribución espacio-temporal y composición con diferentes parámetros ambientales y con la distribución de los peces adultos. PhD Thesis. 1996. [CrossRef]
- Pérez-Rubín, J.; Mafalda, P. Abnormal domination of gilt sardine (Sardinella aurita) in the middle shelf ichthyoplankton community of Gulf of Cádiz (SW Iberian Peninsula) in summer: related changes in the hydrologic structure and implications in the larval fish and mesozooplankton assemblages finded. ICES Annual Science Conference Vigo, Spain 22–25 September 2004. ICES CM 2004/Q: 23, 1-9.
- Pérez-Rubín, J.; Abad, R. La presencia masiva ocasional de larvas y adultos de Capros aper (Pisces) en el Golfo de Cádiz y Mar de Alborán. Gaia Rev Geociencias, 1994, 9, 23-26.
- Sharp, G.D.; Csirke, J. eds. Proceedings of the Expert Consultation to examine changes in abundance and species composition of neritic fish resources. San José, Costa Rica, 18-29 April 1983. FAO Fisheries Report, 1983, 291 (2-3), 1-1224.
- Csirke, J.; Sharp, G.D. eds. Reports of the Expert Consultation to examine changes in abundance and species composition of neritic fish resources. San José, Costa Rica, 18-29 April 1983. FAO Fisheries Report, 1984, 291 (1), 1-102.
- Wyatt, T.; Larrañeta, M.G. eds. Long Term Changes in Marine Fish Populations. International Symposium Vigo, Spain, 18-21 Nov, 1986. 1988.
- López-Areta, J.M.; Villegas, M.L. Annual and seasonal changes in the captures of Sardina pilchardus, Trachurus trachurus, Engraulis encrasicolus and Scomber scombrus on the coast of Asturias (1952-1985). International Symposium on Long Term Changes in Marine Fish Populations 18-21 November 1986, Vigo, Spain. 1986. Abstract nº 35.
- Villegas, M.L.; López-Areta, J.M. Annual and seasonal changes in captures of Brama raii, Pagellus bogaraveo, Thunnus alalunga, Micromesistius poutassou, Merluccius merluccius and Lophius piscatorius on the coast of Asturias (1952-1985). International Symposium on Long Term Changes in Marine Fish Populations 1986. 18-21 November 1986, Vigo, Spain. Abstract nº 39.
- Junquera, S. Changes in the anchovy fishery of the Bay of Biscay in relation to climatic and oceanographic variations in the Nort Atlantic. In: Long Term Changes in Marine Fish Populations. Wyatt, T.; Larrañeta, M.G.; Eds. International Symposium 1988. Vigo, Spain, 18-21 Nov. 1986, 543-554.
- Porteiro, C.; Álvarez, F.; Pérez, N. Variations in the sardine Sardina pilchardus stock of the Atlantic coast of the Iberian Peninsula (1976-1985). International Symposium on Long Term Changes in Marine Fish Populations, 1986, 18-21 November 1986, Vigo, Spain. Abstract nº 36.
- Wyatt, T.; Pérez-Gándaras, C. Ekman transport and sardine yields in western Iberia. International Symposium on Long Term Changes in Marine Fish Populations, 1986, 18-21 November 1986, Vigo, Spain. Abstract nº 37.
- Binet, D.; French sardine and herring fisheries: a tentative description of their fluctuations since the XVIII th century. In: Long Term Changes in Marine Fish Populations. Wyatt, T.; Larraneta, M.G. Eds. Symposium held 18-20 November 1986, Vigo, Spain, 1988, 253-272.
- Regier, H.A.; Magnuson, J.J.; Coutant, C.C. Introduction to Proceedings: Symposium on Effects of Climate Change on Fish. Trans. Am. Fish. Soc. 1990, 119, 173–175. [CrossRef]
- Kawasaki, T.; Tanaka, S.; Toba, Y.; Taniguchi, A. eds. Long-term variability of pelagic fish populations and their environment. Proceedings of the international symposium, Sendai, Japan, 14-18 November 1989. Pergamon Press, Oxford UK, 1991, 402.
- Wyatt, T.; Cushing D.H.; Junquera, S. Stock distinctions and evolution of European sardine. In: T. Kawasaki, S. Tanaka, Y. Toba, and A. Taniguchi eds. Long-term Variability of Pelagic Fish Populations and their Environment. Pergamon Press, 1991, 229-238.
- Souter. A.; Isaacs, J. D. Abundance of pelagic fish during the 19th and 20th centuries as recorded in anaerobic sediments off California. Fish. Bull. 1974, 72, 257-275.
- De Vries, Th. J.; Pearcy, W. G. Fish debris in sediments of the upwelling zone off central Peru: a late Quaternary record. Deep. Sea. Res. Part A. Oceanographic Research Papers, 1982, 29 (1), 87-109.
- Shackleton, L.Y. Fossil pilchard and anchovy scales: indicators of past fish populations off Namibia. In: Proceedings of the International Symposium on Long Term Changes in Fish Populations. Vigo, Spain: Instituto de Investigaciones Marinas de Vigo, 1988, 55-68.
- Shackleton, L.Y. A comparative study of fossil fish scales from three upwelling regions. South Afr. J. Mar. Sci. 1987, 5, 79–84. [CrossRef]
- Sherman, K.; Alexander, L.M. Large marine ecosystems: Patterns, processes and yields. American Association for the Advancement of Science: Washington D.C. XIII, 1990, pp. 242.
- Bakun, A. Patterns in the Ocean: Ocean Processes and Marine Population Dynamics. La Jolla, CA: California Sea Grant. 1996, pp. 323.
- Beamish, R.J. ed. Climate change and northern fish populations. NRC Research Press, 1995, 121, pp. 739.
- Mörner, N.-.-A. Earth rotation, ocean circulation and paleoclimate. GeoJournal 1995, 37, 419–430. [CrossRef]
- Lean, J.; Beer, J.; Bradley, R. Reconstruction of solar irradiance since 1610: Implications for climate change. Geophys. Res. Lett. 1995, 22, 3195–3198. [CrossRef]
- Mörner, N.A. Global Change and Interaction of Earth Rotation, Ocean Circulation and Paleoclimate. An. Acad. Bras. Ci.; 1996, 68 (1), 77-94.
- Mörner N.A. Solar wind, earth’s rotation and changes in terrestrial climate. Physical Review & Research International. 2013, 3 (2), 117-136.
- Friis-Christensen, E.; Lassen, K. Length of the Solar Cycle: An Indicator of Solar Activity Closely Associated with Climate. Science 1991, 254, 698–700. [CrossRef]
- Butler, C.; Johnston, D. A provisional long mean air temperature series for Armagh observatory. J. Atmospheric Terr. Phys. 1996, 58, 1657–1672. [CrossRef]
- Patterson, R.T.; Prokoph, A.; Wright, C.; Chang, A.S.; Thomson, R.E.; Ware, D.M. Holocene Solar Variability and Pelagic Fish Productivity in the NE Pacfic. Palaeontologia Electronica, 2004, 7 (4), 17 p. http://palaeo-electronica.org/paleo/2004_3/fish2/issue1_04.htm.
- Ganzedo, U.; Zorita, E.; Solari, A.P.; Chust, G.; del Pino, A.S.; Polanco, J.; Castro, J.J. What drove tuna catches between 1525 and 1756 in southern Europe?. ICES J. Mar. Sci. 2009, 66, 1595–1604. [CrossRef]
- Ganzedo, U.; Polanco-Martínez, J.M.; Caballero-Alfonso, .M.; Faria, S.H.; Li, J.; Castro-Hernández, J.J. Climate effects on historic bluefin tuna captures in the Gibraltar Strait and Western Mediterranean. J. Mar. Syst. 2016, 158, 84–92. [CrossRef]
- Guisande, C.; Ulla, A.; Thejll, P. Solar activity governs abundance of Atlantic Iberian sardine Sardina pilchardus. Mar. Ecol. Prog. Ser. 2004, 269, 297–301. [CrossRef]
- Boehlert G.W.; Schumacher J.D. Changing Oceans and Changing Fisheries: Environmental Data for Fisheries Research and Management. NOAA-TM-NMFS-SWFSC-239. U.S. Department of Commerce. 1997, pp. 147.
- Barnett, T.P.; Hasselmann, K.; Chelliah, M.; Delworth, T.; Hegerl, G.; Jones, P.; Rasmusson, E.; Roeckner, E.; Ropelewski, C.; Santer, B.; et al. Detection and Attribution of Recent Climate Change: A Status Report. Bull. Am. Meteorol. Soc. 1999, 80, 2631–2659. [CrossRef]
- IPCC. Climate Change 2007. Synthesis Report. Contribution of Working Groups I, II and III to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. IPCC, 2007, Geneva, Switzerland, 104 pp.
- IPCC. IPCC Third Assessment Report: Climate Change 2001. The Scientific Basis, Ginebra. 2001.
- Steffen W.; Sanderson A.; Tyson P.; Jäger J.; Matson P.; Moore B.; Oldfield F.; Richardson K.; Schellnhuber H.J.; Turner B.L.; Wasson R.J. Global Change and the Earth System: A Planet Under Pressure. Springer Berlin, Heidelberg. 2004, 336 pp. [CrossRef]
- Stef, W. Observed trends in Earth System behavior. WIREs Clim. Chang. 2010, 1, 428–449. [CrossRef]
- Pörtner H.-O.; Roberts, D.C; Eds. Climate Change 2022: Impacts, Adaptation and Vulnerability. IPCC 2022. Cambridge University Press, Cambridge, UK. 2022, pp. 3056. [CrossRef]
- European Environment Agency. Climate change mitigation: reducing emissions (Modified 25 Mar 2024). URL: https://www.eea.europa.eu/en/topics/in-depth/climate-change-mitigation-reducing-emissions.
- Palmer, M.D.; Domingues, C.M.; A Slangen, A.B.; Dias, F.B. An ensemble approach to quantify global mean sea-level rise over the 20th century from tide gauge reconstructions. Environ. Res. Lett. 2021, 16, 044043. [CrossRef]
- Rind, D.; Shindell, D.; Perlwitz, J.; Lerner, J.; Lonergan, P.; Lean, J.; McLinden, C. The Relative Importance of Solar and Anthropogenic Forcing of Climate Change between the Maunder Minimum and the Present. J. Clim. 2004, 17, 906–929. [CrossRef]
- Mörner, N.-A. Our Oceans-Our Future: New Evidence-based Sea Level Records from the Fiji Islands for the Last 500 years Indicating Rotational Eustasy and Absence of a Present Rise in Sea Level. Int. J. Earth Environ. Sci. 2017, 2. [CrossRef]
- Mörner, N.A. Chapter 2: 2500 Years of Observations, Deductions, Models and Geoethics: Global Perspective. Boll. Soc. Geol. It, 2006, vol. 125, pp. 259-264. [CrossRef]
- Zhou, L.; Tinsley, B.; Chu, H.; Xiao, Z. Correlations of global sea surface temperatures with the solar wind speed. J. Atmospheric Solar-Terrestrial Phys. 2016, 149, 232–239. [CrossRef]
- Herrera, V.V.; Mendoza, B.; Herrera, G.V. Reconstruction and prediction of the total solar irradiance: From the Medieval Warm Period to the 21st century. New Astron. 2015, 34, 221–233. [CrossRef]
- Jardine, P.E.; Fraser, W.T.; Gosling, W.D.; Roberts, C.N.; Eastwood, W.J.; Lomax, B.H. Proxy reconstruction of ultraviolet-B irradiance at the Earth’s surface, and its relationship with solar activity and ozone thickness. Holocene, 2010, 30, 155–161.
- Miyahara H.; Ayumi Asai A.; Ueno S. Solar Activity in the Past and Its Impacts on Climate. In: Solar-Terrestrial Environmental Prediction. Kusano, K. Eds. Springer, Singapore, 2023, 403-419.
- Thiéblemont, R.; Matthes, K.; Omrani, N.-E.; Kodera, K.; Hansen, F. Solar forcing synchronizes decadal North Atlantic climate variability. Nat. Commun. 2015, 6, 8268. [CrossRef]
- Patti, B.; Guisande, C.; Riveiro, I.; Thejll, P.; Cuttitta, A.; Bonanno, A.; Basilone, G.; Buscaino, G.; Mazzola, S. Effect of atmospheric CO2 and solar activity on wind regime and water column stability in the major global upwelling areas. Estuarine, Coast. Shelf Sci. 2010, 88, 45–52. [CrossRef]
- Landscheidt, T. New Little ICE Age Instead of Global Warming?. Energy Environ. 2003, 14, 327–350. [CrossRef]
- Mörner NA. Solar minima, earth’s rotation and little ice ages in the past and in the future. The North Atlantic – European case. Global Planet. Change. 2010, 72, 282-293.
- Mörner, N.-A. The Approaching New Grand Solar Minimum and Little Ice Age Climate Conditions. Nat. Sci. 2015, 07, 510–518. [CrossRef]
- Mörner, N.-A. Anthropogenic Global Warming (AGW) or Natural Global Warming (NGM). Voice Publ. 2018, 04, 51–59. [CrossRef]
- Meehl, G.A.; Arblaster, J.M.; Marsh, D.R. Could a future “Grand Solar Minimum” like the Maunder Minimum stop global warming? Geophys. Res. Lett. 2013, 40, 1789–1793. [CrossRef]
- Feulner, G.; Rahmstorf, S. On the effect of a new grand minimum of solar activity on the future climate on Earth. Geophys. Res. Lett. 2010, 37, L05707. [CrossRef]
- Arsenovic, P.; Rozanov, E.; Anet, J.; Stenke, A.; Schmutz, W.; Peter, T. Implications of potential future grand solar minimum for ozone layer and climate. Atmos. Chem. Phys. 2013, 18, 3469–3483. [CrossRef]
- Maycock, A.C.; Ineson, S.; Gray, L.J.; Scaife, A.A.; Anstey, J.A.; Lockwood, M.; Butchart, N.; Hardiman, S.C.; Mitchell, D.M.; Osprey, S.M. Possible impacts of a future grand solar minimum on climate: Stratospheric and global circulation changes. J. Geophys. Res. Atmos. 2015, 120, 9043–9058. [CrossRef]
- Ineson, S.; Maycock, A.C.; Gray, L.J.; Scaife, A.A.; Dunstone, N.J.; Harder, J.W.; Knight, J.R.; Lockwood, M.; Manners, J.C.; Wood, R.A. Regional climate impacts of a possible future grand solar minimum. Nat. Commun. 2015, 6, 7535. [CrossRef]
- Chiodo, G.; García-Herrera, R.; Calvo, N.; Vaquero, J.M.; A Añel, J.; Barriopedro, D.; Matthes, K. The impact of a future solar minimum on climate change projections in the Northern Hemisphere. Environ. Res. Lett. 2016, 11, 034015. [CrossRef]
- Yoden, S.; Yoshida, K. Impacts of Solar Activity Variations on Climate. In: Kusano, K. eds Solar-Terrestrial Environmental Prediction. Springer, Singapore. 2023. [CrossRef]
- Tartaglione, N.; Toniazzo, T.; Otterå, O.H.; Orsolini, Y. Equilibrium Climate after Spectral and Bolometric Irradiance Reduction in Grand Solar Minimum Simulations. Climate 2023, 12, 1. [CrossRef]
- Scafetta, N. Empirical assessment of the role of the Sun in climate change using balanced multi-proxy solar records. Geosci. Front. 2023, 14. [CrossRef]
- Kopp, G.; Lean, J.L. A new, lower value of total solar irradiance: Evidence and climate significance. Geophys. Res. Lett. 2011, 38. [CrossRef]
- Giralt, S.; Moreno, A.; Cacho, I.; Valero-Garcés, B. A comprehensive overview of the last 2,000 years Iberian Peninsula climate history. In: Special Issue on climate over the Iberian Peninsula: an overview of CLIVAR-Spain coordinated science. Clivar Exchanges, 2017, 73, 5-10.
- MacKenzie, B.; Christensen. Disentangling climate from antropogenic effects. In: Resolving climate impacts on fish stocks. Rijnsdorp A. D.; Peck M. A.; Engelhard G.H.; Möllmann C.; Pinnegar J. K. eds. ICES Cooperative Research Report, 2010, 301, 44-48.
- Belkin, I.M. Rapid warming of Large Marine Ecosystems. 2009, 81, 207–213. [CrossRef]
- Spalding, M.D.; Fox, H.E.; Allen, G.R.; Davidson, N.; Ferdaña, Z.A.; Finlayson, M.; Halpern, B.S.; Jorge, M.A.; Lombana, A.; Lourie, S.A.; et al. Marine Ecoregions of the World: A Bioregionalization of Coastal and Shelf Areas. Bioscience 2007, 57, 573–583. [CrossRef]
- Kämpf, J.; Chapman, P. 2016. The Canary/Iberia Current Upwelling System. In: Upwelling Systems of the World. Springer, Cham. [CrossRef]
- Sherman K.; Hempel, G. 2008. Perspectives on Regional Seas and the Large Marine Ecosystem Approach: pp. 3-21. In: Sherman, K. and Hempel, G. (Editors) 2008. The UNEP Large Marine Ecosystem Report: A perspective on changing conditions in LMEs of the world’s Regional Seas. UNEP Regional Seas. Report and Studies, 182. United Nations Environment Programme. Nairobi, Kenya.
- https://www.eea.europa.eu/data-and-maps/figures/marine-regions-and-subregions.
- Zeller, D.; Hood, L.; Palomares, M.; Sumaila, U.; Khalfallah, M.; Belhabib, D.; Woroniak, J.; Pauly, D. Comparative fishery yields of African Large Marine Ecosystems. Environ. Dev. 2020, 36, 100543. [CrossRef]
- Chust, G.; González, M.; Fontán, A.; Revilla, M.; Alvarez, P.; Santos, M.; Cotano, U.; Chifflet, M.; Borja, A.; Muxika, I.; et al. Climate regime shifts and biodiversity redistribution in the Bay of Biscay. Sci. Total. Environ. 2022, 803, 149622. [CrossRef]
- Zenetos, A.; Tsiamis, K.; Galanidi, M.; Carvalho, N.; Bartilotti, C.; Canning-Clode, J.; Castriota, L.; Chainho, P.; Comas-González, R.; Costa, A.C.; et al. Status and Trends in the Rate of Introduction of Marine Non-Indigenous Species in European Seas. Diversity 2022, 14, 1077. [CrossRef]
- Png-Gonzalez, L.; Comas-González, R.; Calvo-Manazza, M.; Follana-Berná, G.; Ballesteros, E.; Díaz-Tapia, P.; Falcón, J.M.; Raso, J.E.G.; Gofas, S.; González-Porto, M.; et al. Updating the National Baseline of Non-Indigenous Species in Spanish Marine Waters. Diversity 2023, 15, 630. [CrossRef]
- Carreira-Flores, D.; Rubal, M.; Moreira, J.; Guerrero-Meseguer, L.; Gomes, P.T.; Veiga, P. Recent changes on the abundance and distribution of non-indigenous macroalgae along the southwest coast of the Bay of Biscay. Aquat. Bot. 2023, 189. [CrossRef]
- Pérez-Lloréns, J.L.; Brun F.G.; Hernández, I.; Bermejo R.; Vergara J.J. 2023. Macrofitos marinos (algas y angiospermas) de las costas de Cádiz. In: Pérez-Rubín J. & Ramírez T. (ed). El estrecho de Gibraltar: llave natural entre dos mares y dos continentes. R. Soc. Esp. Hist. Nat. Memorias, 2023, 16, 133-151.
- Heath, M. R. 2007. Responses of fish to climate fluctuations in the Northeast Atlantic. In: The Practicalities of Climate Change: Adaptation and Mitigation: 102-116. Ed. by L. E. Emery. Proceedings of the 24th Conference of the Institute of Ecology and Environmental Management, Cardiff, 14-16 November 2006.
- Kaimuddin, A.H.; Laë, R.; De Morais, L.T. Fish Species in a Changing World: The Route and Timing of Species Migration between Tropical and Temperate Ecosystems in Eastern Atlantic. Front. Mar. Sci. 2016, 3, 162. [CrossRef]
- NASA's Goddard Institute for Space Studies. https://science.nasa.gov/climate-change/scientific-consensus/.
- https://climate.copernicus.eu/esotc/2023https://climate.copernicus.eu/sites/default/files/custom-uploads/ESOTC%202023/Summary_ESOTC2023.pdf.
- https://www.ecmwf.int/en/about/media-centre/news/2024/europe-saw-widespread-flooding-and-severe-heatwaves-2023-report.
- ECMWF, 2024. Europe saw widespread flooding and severe heatwaves in 2023-Report [https://www.ecmwf.int/en/about/media-centre/news/2024/europe-saw-widespread-flooding-and-severe-heatwaves-2023-report]. https://www.ecmwf.int/sites/default/files/styles/wide/public/2024-04/Ranking-annual-sea-surface-temperatures-2023_690px.png?itok=02E1PCTC.
- The NOAA Fisheries Distribution Mapping and Analysis Portal (DisMAP). https://apps-st.fisheries.noaa.gov/dismap/.
- FAO. 2020. FAO Fisheries and Aquaculture - Aquatic Species Distribution Map Viewer. In: FAO Fisheries and Aquaculture Division [online]. Rome. https://www.fao.org/figis/geoserver/factsheets/species.html.
- Domínguez-Petit, R.; Navarro, M.R.; Cousido-Rocha, M.; Tornero, J.; Ramos, F.; Jurado-Ruzafa, A.; Nunes, C.; Hernández, C.; Silva, A.V.; Landa, J. Spatial variability of life-history parameters of the Atlantic chub mackerel (Scomber colias), an expanding species in the northeast Atlantic. Sci. Mar. 2022, 86, e048–e048. [CrossRef]
- Delgado, M.; Hidalgo, M.; Puerta, P.; Sánchez-Leal, R.; Rueda, L.; Sobrino, I. Concurrent changes in spatial distribution of the demersal community in response to climate variations in the southern Iberian coastal Large Marine Ecosystem. Mar. Ecol. Prog. Ser. 2018, 607, 19–36. [CrossRef]
- European Environment Agency (EEA). 2024. “Changes in fish distribution in Europe’s seas” (published 16 Feb 2024): https://www.eea.europa.eu/en/analysis/indicators/changes-in-fish-distribution-in.
- https://www.eea.europa.eu/data-and-maps/figures/temporal-development-of-the-ratio-1/.
- Rijnsdorp, A.D.; Peck, M.A.; Engelhard, G.H.; Möllmann, C.; Pinnegar J.K. (eds.). 2010. Resolving climate impacts on fish stocks. ICES Cooperative Research Report, 301. 371 p.
- Simpson, S.D.; Jennings, S.; Johnson, M.P.; Blanchard, J.L.; Schön, P.-J.; Sims, D.W.; Genner, M.J. Continental Shelf-Wide Response of a Fish Assemblage to Rapid Warming of the Sea. 2011, 21, 1565–1570. [CrossRef]
- Cheung, W.W.L.; Pinnegar, J.; Merino, G.; Jones, M.C.; Barange, M. Review of climate change impacts on marine fisheries in the UK and Ireland. Aquat. Conserv. Mar. Freshw. Ecosyst. 2012, 22, 368–388. [CrossRef]
- Jiming, Y. The dominant fish fauna in the North Sea and its determination. J. Fish Biol. 1982, 20, 635–643. [CrossRef]
- ter Hofstede, R.; Hiddink, J.; Rijnsdorp, A. Regional warming changes fish species richness in the eastern North Atlantic Ocean. Mar. Ecol. Prog. Ser. 2010, 414, 1–9. [CrossRef]
- Yang, J. An estimate of the fish biomass in the North Sea. ICES J. Mar. Sci. 1982, 40, 161–172. [CrossRef]
- Montero-Serra, I.; Edwards, M.; Genner, M.J. Warming shelf seas drive the subtropicalization of European pelagic fish communities. Glob. Chang. Biol. 2014, 21, 144–153. [CrossRef]
- Pinnegar, J.K.; Engelhard G.H.; Daskalov, G.M. Changes in the distribution of fish. In: Rijnsdorp A. D., Peck M. A., Engelhard G.H., Möllmann C., Pinnegar J. K. (eds.). Resolving climate impacts on fish stocks. ICES Cooperative Research Report, 2010, 301: 94-120.
- MacKenzie, B.R.; Payne, M.R.; Boje, J.; Høyer, J.L.; Siegstad, H. A cascade of warming impacts brings bluefin tuna to Greenland waters. Glob. Chang. Biol. 2014, 20, 2484–2491. [CrossRef]
- Alheit, J.; Pohlmann, T.; Casini, M.; Greve, W.; Hinrichs, R.; Mathis, M.; O’driscoll, K.; Vorberg, R.; Wagner, C. Climate variability drives anchovies and sardines into the North and Baltic Seas. Prog. Oceanogr. 2012, 96, 128–139. [CrossRef]
- Beare, D.; Burns, F.; Greig, A.; Jones, E.; Peach, K.; Kienzle, M.; McKenzie, E.; Reid, D. Long-term increases in prevalence of North Sea fishes having southern biogeographic affinities. Mar. Ecol. Prog. Ser. 2004, 284, 269–278. [CrossRef]
- Pérez-Rubín J., 2008. Un siglo de historia oceanográfica del golfo de Vizcaya (1850-1950). Ciencia, técnica y vida en sus aguas y costas. Aquarium Donostia-San Sebastián, Spain.
- Pérez-Rubín J., 2023. 80 años de series de investigaciones periódicas del IEO en el ecosistema pelágico del estrecho de Gibraltar y mares adyacentes (1914-1995): zoología, biología, ecología y medio ambiente marino. Una revisión bibliográfica anotada. In: El estrecho de Gibraltar: llave natural entre dos mares y dos continentes (Pérez-Rubín J. & Ramírez T. (Eds.). Memorias de la Real Sociedad Española de Historia Natural, XVI: 337-383.
- Raybaud, V.; Bacha, M.; Amara, R.; Beaugrand, G. Forecasting climate-driven changes in the geographical range of the European anchovy (Engraulis encrasicolus). ICES J. Mar. Sci. 2017, 74, 1288–1299. [CrossRef]
- Checkley, D.; Alheit, J.; Oozeki, Y.; Roy, C.; Csirke, J.; Glantz, M.H.; Hurrell, J.; Bakun, A.; MacCall, A.D.; Checkley, D.M.; et al. Climate Change and Small Pelagic Fish; Cambridge University Press (CUP): Cambridge, United Kingdom, 2001.
- Engelhard, G.H.; Peck, M.A.; Rindorf, A.; Smout, S.C.; van Deurs, M.; Raab, K.; Andersen, K.H.; et al.. Forage fish, their fisheries, and their predators: who drives whom?. ICES Journal of Marine Science: Journal Du Conseil, 2014, 71, 90–104.
- Blanchard, F.; Vandermeirsch, F. Warming and exponential abundance increase of the subtropical fish Capros aper in the Bay of Biscay (1973–2002). Comptes Rendus Biol. 2005, 328, 505–509. [CrossRef]
- Coad, J.O.; Hüssy, K.; Farrell, E.D.; Clarke, M.W. The recent population expansion of boarfish, Capros aper (Linnaeus, 1758): interactions of climate, growth and recruitment. J. Appl. Ichthyol. 2014, 30, 463–471. [CrossRef]
- Egerton, S.; Culloty, S.; Whooley, J.; Stanton, C.; Ross, R.P. Boarfish (Capros aper): review of a new capture fishery and its valorization potential. ICES J. Mar. Sci. 2017, 74, 2059–2068. [CrossRef]
- Oliver, P., Fernández, A.. Prospecciones pesqueras en la región suratlántica española. Biocenosis de la plataforma y del talud continental. Bol. Inst. Esp. Oceanor., 1974, 180: 1-31.
- Chlaida, M.; Fauvelot, C.; Ettahiri O.; Charouki, N.; Elayoubi, S. et al. Relationship between migratory behavior and environmental features revealed by genetic structure of Sardina pilchardus populations along the Moroccan Atlantic coast. Frontiers in Science and Engineering, 2021, 11 (1), 75 p. [CrossRef]
- Sambe, B.; Tandstad, M.; Caramelo, A.M.; Brown, B.E. Variations in productivity of the Canary Current Large Marine Ecosystem and their effects on small pelagic fish stocks. Environ. Dev. 2016, 17, 105–117. [CrossRef]
- Malta, T.; Santos, P.; Santos, A.; Rufino, M.; Silva, A. Long-term variations in Ibero-Atlantic sardine ( Sardina pilchardus ) population dynamics: Relation to environmental conditions and exploitation history. Fish. Res. 2016, 179, 47–56. [CrossRef]
- Ba, A.; Schmidt, J.; Dème, M.; Lancker, K.; Chaboud, C.; Cury, P.; Thiao, D.; Diouf, M.; Brehmer, P. Profitability and economic drivers of small pelagic fisheries in West Africa: A twenty year perspective. Mar. Policy 2017, 76, 152–158. [CrossRef]
- Cury, P.; Fontana, A. Compétition et strategies démographiques comparées de deux espéces de sardinelles (S. aurita et S. maderensis) de cotes ouest-africaines. Aquat. Living Resour., 1988, 1, 165-180.
- Aggrey-Fynn, J. Distribution and Growth of Grey Triggerfish, Balistes capriscus (Family: Balistidae), in Western Gulf of Guinea. West Afr. J. Appl. Ecol. 2010, 15. [CrossRef]
- Bañón, R.; Pardo, P.C.; Álvarez-Salgado, X.A.; de Carlos, A.; Arronte, J.C.; Piedracoba, S. Tropicalization of fish fauna of Galician coastal waters, in the NW Iberian upwelling system. Reg. Stud. Mar. Sci. 2024, 70. [CrossRef]
- Cury, P.; Roy, C. (eds.). 1991: Pêcheries Ouest-Africaines: Variabilité, Instabilité et Changement. ORSTOM, Paris, France, 525 p.
- Gulland, J.A., Garcia, S., 1984. Observed patterns in multispecies fisheries. In Exploitation of marine communities. In: May, R.M. (Ed.), Dahlem Konferenzen, Springler-Verlag, Berlin, 155–190.
- Binet, D.; Servain, J. Have the recent hydrological changes in the Northern Gulf of Guinea induced the Sardinella aurita outburst?. Oceanologica Acta, 1993, 16(3), 247-260.
- Asiedu, B.; Okpei, P.; Nunoo, F.K.E.; Failler, P. A fishery in distress: An analysis of the small pelagic fishery of Ghana. Mar. Policy 2021, 129, 104500. [CrossRef]
- Balguerías, E. Análisis de los descartes producidos en la pesquería española de cefalópodos del Banco Sahariano. Bol. Inst. Esp. Oceanogr., 1993, 9(1), 75-87.
- Balguerías E. The origin of the Saharan Bank cephalop fishery. ICES Journal of Marine Science, 2000, 57, 15-23.
- Caverivière, A. Étude de la pêche du poulpe (Octopus vulgaris) dans les eaux côtières de la Gambie et du Sénégal. L'explosion démographique de l'été 1986. Centre Rech. Oceanogr. Dakar-Thiaroye, Doc. Scient, 1990, 116, 1-42.
- Caverivière, A. Le poulpe (Octopus vulgaris) au Sénégal: une nouvelle ressource. In: Oiouf T. & Fonteneau A. (eds). L'évaluation des ressources exploitables par la pêche artisanale sénégalaise. Colloques et séminaires Orstom, 1994, 2, 245-256.
- Inejih, C.A.O.; Quiniou, L.; Dochi, T. Variabilite de la distribution spatio-temporelle du poulpe (octopus vulgaris) le long des côtes de Mauritanie. Bulletin Scientifique IMROP, 2002, 29, 19-38.
- Shelton, P.A.; Boyd, A.J.; Armstrong, M.J. The influence of large-scale environmental processes on neritic fish populations in the Benguela current system. Rep. Calif. Coop. Oceanic. Fish. Invest, 1985, 26, 72-92.
- Shannon, L.V.; Crawford, R.J.M.; Brundrit, G.B.; Underhill, L.G. Responses of fish populations in the Benguela ecosystem to environmental change. ICES J. Mar. Sci. 1988, 45, 5–12. [CrossRef]
- Shannon, L.V.; Tauton-Clark, J. Long-term environmental índices for the ICSEAF Area. Draganik B. (ed.). Sel. Pap. Int. Comm SE. Atl. Fish, 1989, 1, 5-15.
- https://www.ntnu.edu/museum/4-oceans.
- https://pastglobalchanges.org/science/wg/q-mare/intro.
- Lehodey, P.; Alheit, J.; Barange, M.; Baumgartner, T.; Beaugrand, G.; Drinkwater, K.; Fromentin, J.-M.; Hare, S.R.; Ottersen, G.; Perry, R.I.; et al. Climate Variability, Fish, and Fisheries. J. Clim. 2006, 19, 5009–5030. [CrossRef]
- Bograd, S.J.; Edwards, M.; Ito, S.; Nye, J.; Chappell, E. Fisheries Oceanography: The first 30 years and new challenges in the 21st century. Fish. Oceanogr. 2022, 32, 3–9. [CrossRef]
- Cacho, I.; Valero-Garcés, B.; González-Sampériz, P. Review of paleoclimate reconstructions in the Iberian Peninsula since the last glacial period. Report: Climate in Spain: Past, present and future.: Pérez, F.F.; Boscolo, R.; Eds.; 2010, 9-24.
- Real, R.; Gofas, S.; Altamirano, Mª.; Salas, C.; Báez, J.C.; Camiñas, J. A.; García-Raso, J. E.; Gil de Sola, L.; Olivero, J.; Reina-Hervás, J. A.; et al. Biogeographical and macroecological context of the Alboran Sea. In: Alboran sea-ecosystems and marine resources. Springer International Publishing, 2021, 431-457.
- Gómez, F. Endemic and Indo-Pacific plankton in the Mediterranean Sea: a study based on dinoflagellate records. J. Biogeogr. 2005, 33, 261–270. [CrossRef]
- Würtz, M. Mediterranean pelagic habitat: oceanographic and biological processes. An overview. 2010. UNCN, Malaga, Spain.
- Zazo, C.; Goy, J.L.; Dabrio, C.J.; Lario, J.; González-Delgado, J.A.; Bardají, T.; Hillaire-Marcel, C.; Cabero, A.; Ghaleb, B.; Borja, F.; et al. Retracing the Quaternary history of sea-level changes in the Spanish Mediterranean-Atlantic coasts: Geomorphological and sedimentological approach. Geomorphology, 2013, 196, 36-49. [CrossRef]
- Zazo, C.; Cendrero, A. Explorando las costas de un pasado reciente: los cambios del nivel del mar. Real Academia de Ciencias (Madrid). 2015, Discursos, 253, 1-112.
- Herbig, H. G.; Mamet, B. L. Stratigraphy of the limestone boulders, Marbella Formation Betic Cordillera, Southern Spain. In: Compte Rendu 10 Congrès International de Stratigraphie du Carbonifère, Madrid, 1985, 1, 199-212.
- Somerville, I.D.; Rodríguez, S. Rugose coral associations from the late Visean Carboniferous of Ireland and SW Spain. In: Fossil Corals and Sponges. Hubmann, B.; Piller, W.E. Eds.; Österreichische Akademie der Wissenschaften, Schriftenreihe der Erdwissenschaftlichen Kommissionen, 2007, 17, 329-351.
- Morales, A.; Rosello, E.; Cañas, J.M. Cueva de Nerja (Málaga), a close look at a twelve thousand year ichthyofaunal sequence from southern Spain. Annales du Musée Royal de l’Afrique centrale. Zool Sci, 1994, 274, 253-264.
- Morales-Muñiz, A. Twenty Thousand Years of Fishing in the Strait. Archaeological Fish and Shellfish assemblages from southern Iberia. In: Human Impacts on Ancient Marine Ecosystems: A Global Perspective. Torben C.; Erlandson, J. Ed. 2008. University of California Press.
- Jordá, J.; Maestro, A.; Aura, E.; Álvarez, E.; Avezuela, B.; Badal, E.; Morales, J.V.; Pérez, M.; Villalba, M.P. Evolución paleogeográfica, paleoclimática y paleoambiental de la costa meridional de la Península Ibérica durante el Pleistoceno superior. El caso de la Cueva de Nerja (Málaga, Andalucía, España). Bol. R. Soc. Esp. Hist. Nat. 2011, 105 (1-4), 137-147.
- Álvarez-Fernández, E.; Carriol, R.-P.; Jordá, J.F.; Aura, J.E.; Avezuela, B.; Badal, E.; Carrión, Y.; García-Guinea, J.; Maestro, A.; Morales, J.V.; et al. Occurrence of whale barnacles in Nerja Cave (Málaga, southern Spain): Indirect evidence of whale consumption by humans in the Upper Magdalenian. Quat. Int. 2014, 337, 163–169. [CrossRef]
- Cacho, I.; Grimalt, J.O.; Canals, M.; Sbaffi, L.; Shackleton, N.J.; Schönfeld, J.; Zahn, R. Variability of the western Mediterranean Sea surface temperature during the last 25,000 years and its connection with the Northern Hemisphere climatic changes. Paleoceanogr. 2001, 16, 40–52. [CrossRef]
- Moreno, A.; Cacho, I.; Canals, M.; Grimalt, J.O.; Sánchez-Goñi, M.F.; Shackleton, N.; Sierro, F.J. Links between marine and atmospheric processes oscillating on a millennial time-scale. A multi-proxy study of the last 50,000yr from the Alboran Sea (Western Mediterranean Sea). Quat. Sci. Rev. 2005, 24, 1623–1636. [CrossRef]
- González-Mora, B.; Sierro, F.J.; Flores, J.A. Study of paleotemperatures in the Alboran Sea between 250 and 150 kyr with the modern analog technique. Geogaceta, 2006, 40, 219-222.
- Degroot, D. Climate change and society in the 15th to 18th centuries. WIREs Clim. Chang. 2018, 9. [CrossRef]
- Holm, P.; Ludlow, F.; Scherer, C.; Travis, C.; Allaire, B.; Brito, C.; Hayes, P.W.; Al Matthews, J.; Rankin, K.J.; Breen, R.J.; et al. The North Atlantic Fish Revolution (ca. AD 1500). Quat. Res. 2019, 108, 92–106. [CrossRef]
- Edvardsson, R.; Patterson, W.P.; Bárðarson, H.; Timsic, S.; Ólafsdóttir, G.. Change in Atlantic cod migrations and adaptability of early land-based fishers to severe climate variation in the North Atlantic. Quat. Res. 2019, 108, 81–91. [CrossRef]
- Moullec, F.; Veleza, L.; Verley, P.; Barrier, N.; Ulses, C.; Carbonara, P.; Esteban, A.; Follesa, C.; Gristina, M.; Jadaud, A.; Ligas A.; López Díaz E.; Maiorano P.; Peristeraki P.; Spedicato Mª. T.; Thasitis I.; Valls, Mª.; Guilhaumon, F.; Shin, Y-J. Catching the big picture of the Mediterranean Sea biodiversity with an end-to-end model of climate and fishing impacts. BioRxiv. 2019. [CrossRef]
- Popova, E.; Yool, A.; Byfield, V.; Cochrane, K.; Coward, A.C.; Salim, S.S.; Gasalla, M.A.; Henson, S.A.; Hobday, A.J.; Pecl, G.T.; et al. From global to regional and back again: common climate stressors of marine ecosystems relevant for adaptation across five ocean warming hotspots. Glob. Chang. Biol. 2016, 22, 2038–2053. [CrossRef]
- Arístegui, J.; Barton, E.D.; Álvarez-Salgado, X.A.; Santos, A.M.P.; Figueiras, F.G.; Kifani, S.; Hernández-León, S.; Mason, E.; Machú, E.; Demarcq, H. Sub-regional ecosystem variability in the Canary Current upwelling. Progress in Oceanography 2009, 83, 33–48. [CrossRef]
- Gallego, D.; García-Herrera, R.; Losada, T.; Mohino, E.; de Fonseca, B.R. A Shift in the Wind Regime of the Southern End of the Canary Upwelling System at the Turn of the 20th Century. J. Geophys. Res. Oceans 2021, 126. [CrossRef]
- Gallego, D.; García-Herrera, R.; Mohino, E.; Losada, T.; Rodríguez-Fonseca, B. Secular Variability of the Upwelling at the Canaries Latitude: An Instrumental Approach. J. Geophys. Res. Oceans 2022, 127. [CrossRef]
- Rosselló, P.; Pascual, A.; Combes, V. Assessing marine heat waves in the Mediterranean Sea: a comparison of fixed and moving baseline methods. Front. Mar. Sci. 2023, 10, 1168368. [CrossRef]
- Kodra, E.; Steinhaeuser, K.; Ganguly, A.R. Persisting cold extremes under 21st-century warming scenarios. Geophys. Res. Lett. 2011, 38. [CrossRef]
- Rollefsen & Tåning, 1948. Inquiry into the Problem of Climatic and Ecological Changes in Northern Waters. (mimeographed).
- McLean, M.; Mouillot, D.; Maureaud, A.A.; Hattab, T.; MacNeil, M.A.; Goberville, E.; Lindegren, M.; Engelhard, G.; Pinsky, M.; Auber, A. Disentangling tropicalization and deborealization in marine ecosystems under climate change. Curr. Biol. 2021, 31, 4817–4823.e5. [CrossRef]
- Cheung, W.W.L.; Watson, R.; Pauly, D. Signature of ocean warming in global fisheries catch. Nature 2013, 497, 365–368. [CrossRef]
- Chust, G.; Villarino, E.; McLean, M.; Mieszkowska, N.; Benedetti-Cecchi, L.; Bulleri, F.; Ravaglioli, C.; Borja, A.; Muxika, I.; Fernandes-Salvador, J.A.; et al. Cross-basin and cross-taxa patterns of marine community tropicalization and deborealization in warming European seas. Nat. Commun. 2024, 15, 1–13. [CrossRef]
- Sánchez-Garrido, J.; Werner, F.; Fiechter, J.; Rose, K.; Curchitser, E.; Ramos, A.; Lafuente, J.G.; Arístegui, J.; Hernández-León, S.; Santana, A.R.; et al. Decadal-scale variability of sardine and anchovy simulated with an end-to-end coupled model of the Canary Current ecosystem. Prog. Oceanogr. 2018, 171, 212–230. [CrossRef]
- Derhy, G.; Macías, D.; Elkalay, K.; Khalil, K.; Rincón, M.M. Stochastic Modelling to Assess External Environmental Drivers of Atlantic Chub Mackerel Population Dynamics. Sustainability 2022, 14, 9211. [CrossRef]
- Townhill, B.L.; Couce, E.; Tinker, J.; Kay, S.; Pinnegar, J.K. Climate change projections of commercial fish distribution and suitable habitat around north western Europe. Fish Fish. 2023, 24, 848–862. [CrossRef]
- Allison, E.H.; Perry, A.L.; Badjeck, M.; Adger, W.N.; Brown, K.; Conway, D.; Halls, A.S.; Pilling, G.M.; Reynolds, J.D.; Andrew, N.L.; et al. Vulnerability of national economies to the impacts of climate change on fisheries. Fish Fish. 2009, 10, 173–196. [CrossRef]
- Cheung, W.W.L.; Pinnegar, J.; Merino, G.; Jones, M.C.; Barange, M. Review of climate change impacts on marine fisheries in the UK and Ireland. Aquat. Conserv. Mar. Freshw. Ecosyst. 2012, 22, 368–388. [CrossRef]
- Raybaud, V.; Bacha, M.; Amara, R.; Beaugrand, G. Forecasting climate-driven changes in the geographical range of the European anchovy (Engraulis encrasicolus). ICES J. Mar. Sci. 2017, 74, 1288–1299. [CrossRef]
- Erauskin-Extramiana, M.; Alvarez, P.; Arrizabalaga, H.; Ibaibarriaga, L.; Uriarte, A.; Cotano, U.; Santos, M.; Ferrer, L.; Cabré, A.; Irigoien, X.; et al. Historical trends and future distribution of anchovy spawning in the Bay of Biscay. Deep. Sea Res. Part II: Top. Stud. Oceanogr. 2018, 159, 169–182. [CrossRef]
- Townhill, B.L.; Couce, E.; Tinker, J.; Kay, S.; Pinnegar, J.K. Climate change projections of commercial fish distribution and suitable habitat around north western Europe. Fish Fish. 2023, 24, 848–862. [CrossRef]
- Failler, P. Fisheries of the Canary Current Large Marine Ecosystem: From capture to trade with a consideration of migratory fisheries. Environ. Dev. 2020, 36, 100573. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



