
Submitted:
27 November 2024
Posted:
29 November 2024
You are already at the latest version
Abstract
Soil salinisation is threatening crop sustainability worldwide mainly due to the anthropogenic climate change. To avoid the excessive ion concentration in soils, plants have developed mechanisms, which have been deeply studied in many model plants and widely reviewed in the scientific literature. Nevertheless, the economically relevant olive tree (Olea europaea subsp. europaea L.), being very exposed to soil salinisation, deserves a specific review, where morphological and biochemical adaptations, signalling pathways involved, and the interplay of transcriptomics and metagenomics adaptations, are described. Variations in the olive tree microbiota due to soil salinisation and usual agricultural practices, and how might biostimulants and chemical amendments may allow the cultivation in salinised soils. Although not concluding, it seems that tolerance in olive cultivars is associated with effective mechanisms of ion exclusion and retention in the root, limiting the accumulation of these ions into actively growing shoots. The emerging importance of post-translational modifications as a source of salt tolerance is highlighted. Also, a comprehensive list of 97 olive cultivars classified by salt tolerance is given. The future advantages of engineering specific metacaspases involved in the programmed cell death and autophagy pathways to rapidly rise salt-tolerant cultivars or rootstocks is considered. An overview of bioinformatic tools focused on olive tree, as well as the machine learning approaches to study plant stress from the multi-omics perspective invites to hypothesise that they can foster the development of salt-tolerant cultivars or rootstocks adapted to soil salinisation and pave the way for a future 'smart oliviculture' in the seeking of more productive and sustainable oliviculture under salt stress.
Keywords:
olive
; cultivar
; salt stress
; salinisation
; salt tolerance
; bioinformatics
; multi-omics
; machine learning
; climate change
; post-translational modification
; programmed cell death
; autophagy
1. Soil Salinisation Is a Major Problem Under Climate Change
1.1. Origins and Consequences of Soil Salinisation
High salts in soil and irrigation water is the more detrimental abiotic factor for agriculture [1,2]. The sixth report of the United Nations Intergovernmental Panel on Climate Change (IPCC) [3] states that deforestation, land degradation, irrigation with brackish or saline water, fertiliser malpractice, and over-exploitation of fresh underground waters, together with the decrease in precipitation, temperature increase and high evapotranspiration due to climate change, are among the main causes of soil salinisation worldwide [2,4,5,6,7,8,9]. Temperature increase, drought waves, and expansion of drylands are especially concerning in the Mediterranean basin, whose coastal areas suffer seawater intrusion in by over-exploitation of groundwater, which is accelerating the problem [10]. Moreover, the alternative use of wastewater, although provides nutrients that may increase soil fertility, is causing unanticipated, serious salt stress [11]. Predictions point out that soil salinisation will affect 50% of total agricultural lands by 2050 [2], with important economic losses [12]. Adding that 33% of irrigated agricultural lands and 20% of the cultivated lands are highly saline (>40 mM), with an expected increase of 10% annually, drives to the urgent need of fighting against salt accumulation in the root zone [13] or the development of salt-tolerant crops [9]. There are some agronomical practices that may mitigate the salt stress, such as restoration of vegetation cover, changes in irrigation pattern and water quality, appropriate use of fertilisers, application of organic amendments to soil (including microbiota changes, compost, charcoal, chemical compounds...), as well as the cultivation of salt-tolerant crop plants [9]. In conclusion, urgent solutions are required to mitigate the effects of soil salinisation.
1.2. Why the Olive Tree?
The effect of soil salinisation in many crop plants (rice, wheat, maize, sorghum, chickpea, soybean, cotton) as well as in the model plant Arabidopsis has been widely studied [14]. But the evergreen sclerophyll olive tree (Olea europaea L subsp. europaea), typically cultivated in dry lands of Europe and Africa [15,16], has not received the same interest. Olive groves have sculpted the Mediterranean landscape and entails important economic, agronomic and agroecological repercussions due to their olive oil and table olives, appreciated worldwide thanks to the benefits for human health [17]. In fact, these benefits have been proposed to be the main driver of its enormous geographical expansion [18]. Additionally, other parts of the plant are of utility for human use, such as its wood for firewood and its seeds for feed supplements (edible oil, nutraceuticals, and proteins) [19]. Different aspects of olive tree biology have been addressed, including fruit maturation [20,21,22,23], oil synthesis [24], alternate bearing [25], juvenility-to-maturity transition [26,27], sexual reproduction and self-incompatibility [28,29,30,31,32,33], pollen allergy [34,35,36,37], pest tolerance [38], etc. Since soil salinisation is also affecting olive fruit yield [39], there are studies about response to salt stress in olive tree (previously reviewed in [15,16,40]). Salt stress is exacerbated in olive groves due to the change from a traditional, extensive, rain-fed crop to intensive or super-intensive groves with high water demands [41]. Hence, it can be hypothesised that soil salinisation might explain why being Europe and Africa the main olive oil producers in the world, are Oceania and America the lands with better yields [15].
In olive tree groves, the soil amendments and salt-tolerant crops are considered the only realistic approaches to mitigate salt stress to date. This is why this review is focused on the biochemical, physiological, genomic and metagenomic studies (in their broadest sense) developed for olive trees to gain molecular insights into its adaptive response to salt stress, the putative roles of microbiota and post-translational modifications in the salt tolerance, as well as the future of approaches based on artificial intelligence, to maintain olive yields in stressed lands.
2. Plant Management of Soil Salinisation
Although there are many reviews considering the plant responses to salt stress, such as the one by Hualpa-Ramírez and coworkers [42] focused on possible biotechnological strategies to cope with climate change harsh conditions, a general overview of the main physiological and molecular mechanisms of salt tolerance will be illustrative (Figure 1).
2.1. Overview of Plant Responses to Salt Exposure
Natural salt stress is usually due to soil salinisation, characterised by the accumulation of water-soluble salts (mainly Na+ and Cl− accompanied by other ions) in the root zone, which provokes a diminution of water uptake [43,44]. This increase induces a short-term osmotic response in plant followed by an long-term response [45,46], as well as other secondary stresses such as nutritional imbalance and oxidative stress (Figure 1). All these factors will limit net photosynthesis rate and consequently plant growth, development, and yield [46]. After long-term salt stress, plants present chlorosis, membrane electrolyte leakage, photosynthesis impairment, reduction of leaf surface and stem diameter, increase of leaf thickness, compartmentalization of salts in old leaves, and stunted growth [47,48,49]. More severe symptoms include leaf burn, scorching, necrosis and premature defoliation [49]. In spite of the usual robustness of roots, they are very vulnerable to salts [50] and try to cope with salinisation by increasing the root/shoot ratio to diminish the demand for element supply and to improve water absorption and osmotic balance [51].
2.2. Biochemical Indicators of Salt Adaptation
The biochemical and molecular adaptation is triggered to evade the adverse effects of salt stress. Most crop plants synthesise and accumulate osmolytes or osmoprotectants (sugars, proline, arginine, glycine betaine, organic acids, etc.) [52] to maintain cellular osmotic potential and turgor, stabilise proteins, and scavenge reactive oxygen species (ROS) that appear at the short-term response (Figure 1). An excess of ROS can produce the oxidative damage of macromolecules [45,53], including cell membrane disorganisation that leads to ion leakage, covalent modifications of chromatin via histone modifications, DNA methylation and alternative splicing [54,55], and alterations in membrane and protein surface charges [54]. Hence, a complex antioxidant defence system is expressed during salt stress, comprising enzymes [56] (e.g., superoxide dismutase [SOD], catalase, peroxidases, glutathione reductase, and glutathione-S-transferase) and non-enzymatic components (e.g., ascorbic acid, tocopherol, carotenoids, phenolic compounds, alkaloids, and glutathione) [45,57]. It has been observed that transgenic Arabidopsis and rice over-expressing SOD or other antioxidant enzymes switch to salt tolerance (up to 250 mM NaCl) and produce seeds in the presence of 100 mM NaCl [45]. Unfortunately, those genetically modified organisms (GMOs) are difficult to bring to orchards, particularly in Europe, due to the restrictive regulations against GMOs [58].
2.3. Adaptive Signalling to Salt Stress
2.3.1. Hormonal Signalling
It is well known that abscisic acid (ABA) and ethylene play a key role in regulating the response to salt stress [59,60]. ABA is a growth and stress hormone that increases its biosynthesis under salt stress to regulate stomatal closure in order to reduce water loss and prevent further salt uptake, resulting in plant survival [61]. Ethylene is involved in signalling pathways that control cytoskeleton dynamics via microtubule repolymerisation, root development, ion transport, and senescence in response to salt stress. Zhao and coworkers [62] and Zhou and coworkers [63] have recently published exhaustive reviews about hormonal signalling.
2.3.2. Post-Translational Modifications
Another adaptive signalling that is gaining interest is the post-translational modification (PTM) of proteins. Even though the role of PTMs in plant salt tolerance is at a very early stage, it has been demonstrated the ubiquitination and SUMOylation of proteins in pathways regulating salt stress, and the importance of phosphorylation in the ion homeostasis under salt stress [64]. PTMs are described to modulate or even inhibit the activity of numerous enzymes and proteins (including transporters), modify their cell localization or binding capacity, as well as minimise the ROS production to provide some tolerance to salts [64]. Additionally, ROS, reactive nitrogen species (RNS) and other free radicals like reactive sulphur species (RSS) are triggered by salt stress and exert their salt-tolerant signalling functions throughout other PTMs such as carbonylation, S- and Tyr-nitration, sulfhydration, persulphidation, and others [65,66]. Lipidation, N-acetylation and N-glycosylation are also promising PTMs for salt tolerance [64].
Many chemical agents, most of them known to produce PTMs, such as hydrogen sulphide (H2S), nitric oxide (NO), hydrogen peroxide (H2O2) or melatonin, have been shown to enhance the salt tolerance of crop plants when used for priming seeds and seedlings [67], probably by means of the PTMs they cause. In any case, it is important to bear in mind that salt tolerance induced by PTMs would make reusable the salinised lands using these 'primed' crops without the need of GMOs or the time-consuming breeding programs or replanting with salt-tolerant crops.
2.3.3. Signalling Crosstalk
It is well established that cytosolic Ca2+ concentration increases rapidly in plants with the exposure to high salt concentrations, suggesting that elevated cytosolic Ca2+ may function as second messenger in salt stress [61,68] (Figure 1). However, Ca2+ signalling enables information integration and physiological coordination in response to a variety of extracellular cues, such as nutrient deprivation and abiotic stresses, as well as intracellular signals (such as plant cell death [69,70]), which is vital for downstream responses. Interplay between Ca2+, ROS, PTMs, hormones and MAPKs seems to shape the acclimation responses in most plants [71]. An inadequate adaptation to a high ion strength may drive however to premature senescence and programmed cell death [72] (Figure 1, red lines and boxes), as will be discussed below.
2.4. Cell Wall Thickening and Leaf Changes Under Salt Stress
Epidermis is the first line of defence against environmental stresses, but high salt concentrations can severely damage the cell wall to disrupt the normal growth and development of plants by affecting the cortical microtubule polymerisation (governed by Ca2+ levels) and the cellulose synthase (CesA) complex assembly. This is why plants respond by altering the synthesis and deposition of epicuticular wax and the main cell wall components to prevent water loss and decrease the transport of ion surplus into the plant. This mainly affects biosynthesis and deposition of cell wall components (cellulose, pectins, hemicelluloses, lignin, suberin and epicuticular waxes) resulting in an increase of both cell wall and leaf thickness when salts are abundant [73,74]. Cell wall suberinisation and lignification becomes a physical root barrier to salts that is regulated by several MYB transcription factors as well as members of the WRKY, ERF and NAC gene families [75]. Other morphological changes described in salt stress are a decreased stomatal density and an increased trichome density [43].
2.5. Avoiding Ion Toxicity
Salt tolerance is a polygenic trait influenced by both genetic and environmental factors [76]. Two main phenotypes can be defined: halophytes (species adapted to perpetually saline conditions of 70-200 mM NaCl [77]) and glycophytes (plants requiring soils with low salt content to maintain low Na+ and Cl- levels in their above-ground tissues [78]). The toxicity of different ions also varies according to the plant species and among cultivars, although accumulation of Na+ in the cytosol is detrimental in both glycophytes and halophytes [46,55,57]. Na+ is toxic for most terrestrial plants, including many crops [46,48,79,80], and its chemical similarity with K+ makes that high levels of external Na+ competitively inhibit K+ uptake systems and stimulate K+ efflux, leading to insufficient intracellular K+ concentrations [81,82,83]. Therefore, the maintenance of balanced cytosolic Na+/K+ ratios has become a key tolerance mechanism in plants, such as Arabidopsis, cereals and other crop plants. Transporters probably avert the excessive accumulation of Na+ and/or prevent the loss of K+ in the cytosol to keep a high K+/Na+ ratio [55,60] by means of (1) NSCC, AKT1, HAK5 and SKOR/GORK, P-H+-ATPases, involved in selective K+ vs Na+ uptake in roots; (2) Na+/H+ antiporter, SOS1, P-ATPases, mediators in the Na+ removal to the external medium or to the apoplast; (3) Na+/H+ antiporter, NHXs, V-H+-ATPase, V-H+-PPase, participating in vacuolar and endosomal compartmentalisation; and (4) HKT1 and SOS1, associated with xylem and phloem loading and unloading. In consequence, ion exclusion, including accumulation in root cells, is used by some plants to prevent the intracellular accumulation in the aerial part.
In other crops, such as citrus, grapevine, avocado and persimmon, excessive accumulation of Cl– (but not Na+) in leaves correlates with a decrease in crop transpiration, photosynthesis, yield, and quality, eventually leading to plant death [43,84,85,86,87]. However, prevention of Cl– toxicity has been poorly studied in spite of the characterisation of several genes exhibiting Cl– transport activity (NPF, NRT2, SLAH, SLAC, CCC, ICln, ALMT, CLC, or NAXT [86,88,89]). Very recently, a new mechanism of Cl– exclusion has been described in which the chloride channel protein f (CLCf) in Arabidopsis thaliana moves from Golgi's apparatus to the plasma membrane to remove the excess of chloride ions when plants are treated with NaCl, which increased plant tolerance to salt [90]. This change of location remembers the behaviour of VPEs (vacuolar processing enzymes [91]) involved in the achievement of programmed cell death (PCD, see below).
There are other mechanisms involved in ion detoxification. For example, aquaporins of the PIP (plasma membrane intrinsic proteins) family accumulate in the root cortex to lower the water conductance, and dehydrins increase their expression to prevent cellular damage caused by dehydration [92]. The tetraspanins (integral membrane proteins, characterized by four transmembrane domains) are involved in development, reproduction and immune responses, but also contribute to salt and drought stress tolerance in rice by (1) maintaining an elevated K+/Na+ ratio to gain the ionic equilibrium, (2) regulating antioxidant pathway enzymes and (2) promoting the proline accumulation [93]. Biotechnological manipulation of model plants with some of the mentioned ion transporters are providing encouraging results [94], but these plants are difficult to bring to orchards, particularly in Europe, due to the restrictive regulations against GMOs [58] (more hopes can be placed on CRISPR-modified plants after recent resolutions [95,96]).
2.6. Programmed Cell Death and Autophagy Under Salt Stress Conditions
Cell wall structure, cytoskeleton dynamics, phytohormone and ROS signalling, metabolism activation, foliar gas exchange control, morphological changes of leaf anatomy, and growth control have been mentioned above as adaptive processes to alleviate salt stress (Figure 1). PCD, which is usually overlooked in salt stress studies, is also among the evolved plant strategies to tolerate extreme environments by an organised removal of superfluous and damaged cells. In fact, Na+ toxicity (and not osmotic stress) leads to PCD activation [97] (Figure 1), the Na+/K+ imbalance due to salt stress being crucial for triggering the effector enzymes of PCD [98,99]. Salt treatments lead to a rapidly rise in cytosolic Ca2+ levels that trigger the Ca2+/SOS cascade, allowing the discrimination of salt stress from mere osmotic stress [70] and resulting in the onset of PCD and death-specific enzymes [69]. PCD depends on caspases as well as on VPEs (located in the apoplast in normal conditions but relocated to the cytoplasm under stress). VEPs have also been recently established as executors of NaCl stress-induced vacuole-mediated PCD [91]. Overexpression of anti-apoptotic proteins such as Bcl-2 drives to the inhibition of salt-induced PCD because it suppresses Ca2+ rise and decreases VPE expressions, which eventually improves salt stress tolerance [100]. Since VPEs became key moderators of salt stress-induced PCD in plants, new transporter-independent strategies can be envisaged to enhance the efficiency of salt tolerance.
Salt stress also triggers autophagy to avoid high salt concentrations in cytosol (Figure 1). Thus, NaCl treatments induce transcription of several autophagy-related genes (ATGs) in plants for Na+ sequestration in the central vacuole, being some of these ATGs and metacaspases required for the autophagosome-vacuole fusion [101]. Hence, plant metacaspases, in addition to be regulators and executors of PCD in a Ca2+-dependent way [102,103], are also involved in a variety of stress responses [104]; for example, metacaspases rapidly enhance their expression in response to salt stress [105]. There are increasing evidences that point out metacaspases as regulators of the autophagy-PCD crosstalk [105,106] since autophagy, together with PCD, is a conserved strategy in plants to resist various environmental stresses [107]. Metacaspases that activate PCD and autophagy, as critical regulators of the balance between cell death and survival, emerge as new biotechnological targets to improve plant salt tolerance. Moreover, manipulation of innate vacuolar PCD pathway emerges as an approach for 'broad stress tolerance'.
2.7. Computer Learning as an Integrative Approach to Study Salt Stress
Generation of new knowledge based on public data becomes the next challenge where the emerging field of artificial intelligence (AI) and its machine learning (ML) and deep learning (DL) subfields are expected to play a key role. A comprehensive understanding of plant response to salt stress would require the combination of image data (source of morphology and phenomics) with genomics datasets in the widest sense [108]. In recent times, the application of ML has gained prominence in the field of plant phenotyping [109,110] as well as in the analysis of plant leaf images to predict the occurrence of biotic and abiotic damage [111]. DL has been successfully applied to identify specific image patterns that can differentiate canopy and non-canopy regions [112] or leaves under stresses [109,110], as well as for the analysis of time-course images to predict plant leaf damages [59,109,110,113,114]. AI techniques, particularly in computer vision, are rapidly progressing in agriculture, encompassing tasks such as counting plants, fruits or leaves, crop classification, phenotyping, vein pattern detection, plant disease diagnosis, weed control, and biotic stress prediction [59,109,110,111]. Combination of AI models was exploited to diagnose plant stress in greenhouses [115]. Other promising studies reveal that γ-aminobutyric acid (GABA) concentration, type of crop, days of treatment, and level or stress (drought or salt), were able to predict the time for optimal physiological properties in pomegranate [116]. It is clear that integration of physiological, biochemical, and molecular markers with AI techniques for predicting plant stress remains an underexplored area that merits more efforts [108,117,118].
ML models for the identification of genes responsible for six types of abiotic stress (salinisation, oxidation, light, heat, cold and drought) were investigated [119], concluding that SVMs (support vector machines) outperformed other ML approaches including DL. Another successful case was the prediction of miRNAs associated with four specific abiotic stress factors (salt, drought, cold and heat) using SVM models that also surpassed the results obtained by these same authors using more computationally consuming DL algorithms [120]. Yield prediction under salt stress for breeding programmes in alfalfa and many other polyploid crops was another successful use of ML to integrate polymorphisms and genomic context [121,122]. Other ML approach that integrates time-series transcriptome data from various stress types (heat, cold, salt, and drought) drove to more accurate stress classification, with high specificity in the discovery of known stress-related genes [123]. Recently, salt stress was tackled with a multi-omics approach in Fabaceae plants (that enabled the development of 'smart farming' with genome-wide marker-assisted breeding and removal of deleterious genes [124]) and rice (with the application of AI to obtain accurate predictions [125]). As a result, more and more pipelines and web tools are emerging designed for multi-omics datasets, such as 'mixOmics' [126], 'OmicsIntegrator' [127] or 'MiBiOmics' [128], as well as AI-driven tools that reduce the need for human intervention such as 'AutoBA' [129] (AI tools and databases to feed algorithms have been recently compiled [108]). Hence multi-omics AI strategies have the potential to revolutionise not only plant stress research but also crop breeding programs [118,130,131,132].
3. Olive Tree Management of Soil Salinisation
Even though the olive tree has been described as an intermediate tolerant species to salinity compared to other fruit trees (but less resistant than barley, cotton, or sugar beet), a sustained excessive salt exposure may result in a cultivar-dependent, non-adaptive response presenting with reduction of stalk length, leaf surface, dry weight and root length. Easily seen phenotypes of a non-adaptive response are leaf tip burn, leaf chlorosis, leaf rolling, wilting of flowers, and even root necrosis, shoot dieback and defoliation [40]. Identifying and understanding the adaptive response to salts play a pivotal role in enhancing orchard management practices, ultimately contributing to increased yield, cost reduction, and sustainability [133].
3.1. Ion Exclusion and Retention in Olive Tree
The level of tolerance to salts in olive tree is associated with effective mechanisms of ion exclusion and retention of Na+ and Cl− in the root to limit the accumulation of these ions into actively growing shoots [134,135,136]. This suggests that ion concentrations in leaves (mainly Na+ and Cl−) is a good indicator of salt stress [136,137], since salt stress damage occurred when the leaf content of Na+ and Cl– exceed 0.5% and 0.2% dry weight, respectively [40]. In fact, the tolerance limit for olive trees was established on 137 mM NaCl [138]. As occurs in other plants, the early response of the olive tree is also interlaced with osmotic shock, including a decrease in water potential and plant turgor, leading to ABA-dependent stomata closure and photosynthesis decay. Later, salt-specific effects are deployed depending on the concentration of Na+ and Cl– ions, and the stress prolongation, presenting typical phenomics effects, more correlated to Na+ than to Cl– accumulation in leaves [139]. Stem elongation and gas exchange in olive leaves may be misleading since they were not highly correlated with Na+ and Cl– contents or salt treatments [139,140]. In spite of the decrease in net photosynthesis, leaf chlorophyll content and leaves dry weight [141], there was no relationship between salt accumulation and photosynthesis decay in either young or old leaves, confirming that olive photosynthetic sensitivity to salt did not depend on salt exclusion or compartmentalisation in old leaves [142]. But in agreement with the observed findings in other plants, leaf lignin content is increased in olive tree as response to salt stress [143,144]. Additionally, growing parameters such as shoot elongation, trunk diameter, dry weight of total plant, number of leaves, leaf thickness, stem and roots, internodes length, total leaf area, shoot/root ratio, are good surrogates to determine the salt effects on olive tree [15].
A comparative study [145] revealed a lower osmotic potential in salinised olive trees (100 mM NaCl) than in citrus (50 mM NaCl); additionally, the detected increase in Cl– and Na+ concentration in leaves and roots of both species caused a decrease in leaf chlorophyll a in citrus but not in olive trees. As a result, young olive plantlets do not seem to suffer any apparent damage when treated with 100 mM NaCl for 57 days or even 18 months in field-grown trees irrigated with saline water (6.6 dS/m) [40], while adults growing with moderate salinity (40-60 mM NaCl) reduce viability and germinability of pollen, mean number of perfect flowers per inflorescence, and fruit set [40]. Leaf drop phenomenon was observed from 60 days after high salt treatments [136]. High-salt irrigation water (>6 dS/m) has a detrimental effect by accelerating olive fruit ripening and inhibiting the linoleic desaturase activity, driving to an increase in phenol content and a decrease in the unsaturated/saturated fatty acid content [146].
Olive trees protect their sensitive shoot organs from the detrimental effects of excess toxic-ion concentration by means of a moderate growth and a reduced transport of water that would prevent salt translocation to the aerial part of the plant [140]. Only after a maintained high salt stress (>6-8 dS/m), ions are translocated to and accumulated in the stem and old leaves [139,147]. This exclusion driven by a drastic decline in the water mass flow seems an untenable strategy for most annual crops, but since olive cultivars have evolved under adverse environments, they are able to assimilate CO2 and produce new growth at considerable rate, allowing a rapid recovery of plant performance when good-quality water is available to the roots [148]. High salt also reduces the number and length of roots, increases the root turnover, and rises the concentration of Na+ and Cl− 5 to 10 times greater compared to leaf tissues, indicating that the protection of aerial tissues from salt ends up having a high cost to roots [137].
3.2. Biochemical Adaptations to Salt in Olive Cultivars
Although is well established that, as occurs in 'Frantoio', long-term salt tolerance is mediated by Na+ exclusion in leaves and accumulation in roots, there are other possibilities. For example, in one study, 'Frantoio' presents a higher mobilisation of Ca2+, with expression of transporter activities in cytoplasmic membrane instead of tonoplast [149]. This setting may remember the recently described tolerance mechanism involving CLCf in A. thaliana [90].
A Croatian wild olive tree genotype was considered salt-resilient due to several factors: its ability to accumulate a huge amount of Na+ and Cl− in leaves, avoid the loss of K+, and deploy adaptable antioxidative mechanisms [150]. It is known that ROS are increased in olive trees due to salt stress, with a concomitant increase in the activity of antioxidant enzymes that differs between cultivars [141]. In fact, some degree of salt tolerance in olive trees is depending on the time required to increase SOD levels in plant tissues, since more sensitive olive tree genotypes accumulate higher concentrations of SOD over a shorter time [151].
Another way of defence against oxidation due to salt stress is the accumulation of phenolic compounds, including the highly abundant oleuropein, especially in leaves, due to the highly significant correlation observed between total phenol content and antioxidant activity in both leaves and roots in different olive cultivars [152]. It has also been observed a strong up-regulation of key genes of the phenylpropanoid pathway in salt-sensitive 'Leccino' under salt stress while salt-tolerant 'Frantoio' showed a less intense up-regulation of the phenylpropanoid genes but overall higher content of phenolic compounds, suggesting that 'Frantoio' faces NaCl toxicity not only with mechanisms of Na+ exclusion, but also through activating antioxidant defences [153,154].
Regarding osmoprotection, in some cultivars such as 'Picual', 'Gemlik' or 'Chemlali', proline accumulates in roots and leaves in response to salts due to its high hydration capacity so that it can bind proteins to prevent their denaturation [144,155,156,157]. However, a decrease in proline levels under salt stress has been described for cultivars 'Chétoui', 'Arbequina', 'Koroneiki', 'Royal de Cazorla' and 'Fadak 86' [141,143]. Other osmoprotectant adjustments in olive trees to counteract oxidative damage under salt stress involved mannitol, violaxanthin-cycle pigment, and flavonoid accumulation [148].
Exploring the Italian olive germplasm also allowed to compare 'Canino' (salt-tolerant) and 'Sirole' (salt-sensitive) transgenic olive trees overexpressing a tobacco osmotin gene with control non-transgenic plants [158]. After four weeks of treatment with 200 mM NaCl, stunted growth and ultimate leaf drop was observed in both cultivars, but not in the transgenic lines. The authors provide evidence that the S assimilation pathway plays a key role in adaptive response of olive trees under salt stress conditions. Tobacco transgenic plants overexpressing OeSCR1 gene, which encodes a Ks-type dehydrin, also showed an increased tolerance to salt and drought stresses [159].
3.3. Salt Tolerance of Olive Tree is Cultivar-Dependent
Since the natural habitat of olive trees are calcareous soils, the available Ca2+ limits the toxic effects of Na+ on plasma membrane integrity and avoids the transport of Na+ and Cl− to sensitive shoots and leaves [147], explaining at least partially why olive trees appear more tolerant to salts than other fruit trees. Additionally, olive grooves based on salt tolerant cultivars may have positive consequences for soil fertilisation, since salt-sensitive trees have difficulties in capturing the available nitrogen compounds in salinised soils while salt-tolerant trees can capture them [160], resulting in a clear improvement of plant growth in high salt. As occurs in other fruit trees, the use of salt-tolerant rootstocks grafted with salt-sensitive cultivars can mitigate salt stress issues. To avoid the time-consuming efforts for improving salt tolerance in cultivars or rootstocks, the use of salt tolerant cultivars can be encouraged on salinised soils. The knowledge of the salt-tolerance levels of olive cultivars is then helpful since this trait is cultivar-dependent [138]. Hence, we have collected this information from literature and presented it in Table 1 and Table 2. As a result, 89 cultivars have been unambiguously described as salt-tolerant (30), intermediate tolerance (33) and salt-sensitive (26) nowadays (Table 1), even if they have been analysed in different reports (such as 'Chemlali', 'Frantoio', 'Kalamon', 'Megaritiki', 'Royal de Cazorla', 'Manzanillo', ' Chondrolia Chalkidikis' and 'Leccino' in Table 1). 'Frantoio' is commonly considered a salt-tolerant cultivar despite the fact that in one study [157] the authors found that ‘Ocal’ (Table 1) and ‘Picudo’ (Table 2) were more salt-tolerant cultivars than 'Frantoio'. 'Changlot Real' was included in the intermediate tolerance group in Table 1, but nurseries (https://gardencenterejea.com/olivos/382-olivo-changlot-realacebuche-c1.html) and the experience of the authors indicate that it can tolerate high salts. Treatment time has been very variable in the different studies, as well as the strength of the salt treatment. More surprisingly, the salt concentration is not described in some cases (ND in Table 1).
Further 8 cultivars (including the very common 'Arbequina', 'Koronieki' and 'Picual') have been assigned to different levels of salt tolerance depending on the published studies (Table 2). Surprisingly inconsistent cases are 'Arbequina' and 'Picual (qualified as tolerant, intermediate and sensitive), or 'Chétoui' and 'Gemlik'' (qualified as sensitive or tolerant). Divergences in salt concentration and treatment time, in addition to the research laboratory, might partially explain the inconsistencies in the tolerance qualification.
4. Studies About Salt Stress Genomics in Olive Tree
4.1. More and More Genome Sequences of Olive Cultivars Are Available
In recent years, olive tree genome sequences started to populate databases. The first one corresponded to the genome of a very old tree of cultivar 'Farga' [183], whose assembly and annotation have been recently improved [184]. Next was the wild olive or oleaster (O. europaea L. subsp. europaea var. sylvestris) from a Turkey-located individual [18], although it was recently suggested that it would be more consistent with a feral genome [184]. The 'Picual' cultivar, widely used in Spain but without a clear salt-tolerance phenotype (Table 2), was published on the scaffold stage [185], chromosomes being pending. Another widely used cultivar in Spain and throughout the world called 'Arbequina', whose salt tolerance also seems inconsistent (Table 2), was recently sequenced at the level of chromosomes combining short- and long-reads [186]. Finally, using the most recent advances in sequencing platforms, the first gap-free, telomere-to-telomere, olive tree genome has been published for the salt-sensitive cultivar 'Leccino' (Table 1) [187], representing the highest contiguous and complete olive genome to date. There are two other Olea europaea sequenced genomes corresponding to Olea europaea subsp. cuspidata, a wild olive tree present from northeast Africa and southwest Asia to drier parts of Yunnan and Sichuan in China. This subspecies, considered the ancestor of the European olive, was also originally introduced into Australia and New Zealand as ornamental plant, but now it is considered relevant as rootstock because of its fungal resistance, growth strength and the survival rate of olive cultivar grafts [188,189,190].
4.2. Transcriptional Response to Salt Stress in Olive Tree
Over the last decade, many key genes for salt tolerance were reported in plants, many of them relating to oxidation-reduction processes, ion transport and chloride channels, cell wall synthesis, as well as hormone-related genes, supporting the above introduced idea that salt tolerance is a multigenic response [60]. Genes related to transcription factor families such as MYB, WRKY, basic leucine zipper (bZIPs), basic helix–loop–helix (bHLH), NAC, homeobox (HB) and GATA are key in the early response to salt stress [193,194]. This can explains why transgenic plants overexpressing transcription factors such as DREB6 [52] or SNAC1 [195], among others, gain salt tolerance under controlled conditions. Unfortunately, a limited success was obtained when these strategies were translated from laboratory to field [196].
Olive breeding was only initiated in the second half of the twentieth century and currently represents the most promising strategy to provide farmers with new, well adapted cultivars or rootstocks [197]. Since (1) genomics tools can shorten the protracted crossbreeding process [118], and (2) high-throughput analyses of differentially expressed genes in horticultural plants support the already known physiological findings [198], genomic information from olive tree regarding salt stress is required to foster new salt-tolerant varieties or rootstocks. A pioneer transcriptomic study was carried out by Bazakos and coworkers [172], using 'Kalamon' (salt tolerant) and 'Chondrolia Chalkidikis' (salt sensitive) cultivars treated with 120 mM NaCl for 90 days. In the salt tolerant cultivar 'Kalamon', up to 159 genes were up-regulated in response salt treatment and 50 transcripts were synthesized de novo after 45 days of treatment, all of them decreasing their levels after plants recovered from stress. However, only 20 genes responded to salt stress in the salt sensitive cultivar. This lack of transcriptional activation might be partly responsible for the sensitivity of 'Chondrolia Chalkidikis' compared to 'Kalamon'. The same authors analysed leaves and roots of the salt-tolerant olive cultivar 'Kalamon' using 454-pyrosequencing [173]. Only 24 genes in roots (15 up- and 8 down-regulated) and 70 in leaves (56 up- and 14 down-regulated) were differentially expressed in salt stress. Among them, the typical responsive transcription factors were reported, but also other salt-responsive genes including AP2/ERF, NF-Y, JERF, HMG and GRAS, ABA-related transcripts, a vacuole-type H+-ATPase, a Na+/H+ antiporter to accumulate Na+ in the vacuole, and the plasma membrane antiporter SOS1. The presence of solute transporters and enzymes involved in the synthesis of osmoprotectants corroborates that salt stress in olive trees triggers an osmotic response at early stages.
In another study involving only the salt-sensitive cultivar 'Chondrolia Chalkidikis', concentration of metabolites was contrasted with expression levels of genes involved in tyrosine (OePPO, OehisC), flavonoid (OeAtF3H, OeFNSII, OeCA4H), lignan (OePLRTp2) and secoiridoid (OeGTF) metabolism during 45 days at 75 mM NaCl to reveal salt-responsive tissue-specific metabolic pathways [180]. Tyrosine and flavonoid metabolism-related genes were clearly modulated by salt in old leaves, while OeGTF gene up-regulation in new leaves suggests that oleuropein metabolism was modified by salt stress. The same authors studied the molecular basis of the priming response in roots and leaves to moderate salt (75 mM NaCl for 45 days) in the same cultivar following a proteogenomic approach [181]. This study revealed major differences between primed and non-primed tissues mainly associated with hormone signalling and defence-related interactions, as well as the key role of protein PTMs, particularly phosphorylation, carbonylation and S-nitrosylation, in promoting salt priming. Salt stress also produces a significant reduction of OeFAD6 transcript levels together with a synergic up-regulation of OeSAD1 gene, increasing the oleic/linoleic ratio in the maturing fruit mesocarp of salt-sensitive cultivar 'Leccino' [146].
Up to ten salt-responsive candidate genes were tested in shoots of high- and medium-tolerant Spanish cultivars ('Royal de Cazorla' and 'Picual', respectively), and compared with low-tolerant Greek ('Koroneiki') and Iranian ('Fadak86') varieties [177]. Only OeNHX7, OeP5CS, OeRD19A and OePetD genes were up-regulated in salt tolerant cultivars. These data suggest that the maintenance of ionic homeostasis, the increase of turgor pressure, the modulation of electron transport and generation of ATP, and the induction of stress-related proteins (SRPs) are key processes to cope with salt stress. Moreover, expression of OePIP1.1 (an aquaporin), OePetD (a cytochrome b6), OePI4Kg4 (a phosphatidylinositol 4-kinase) and OeXylA (a xylose isomerase) genes was differentially regulated by DNA methylation among cultivars, suggesting that this mechanism could be also involved in the adaptation to high salt of the salt-tolerant cultivar 'Royal de Cazorla' [159]. In another study, the differential expression of novel salt-responsive genes (OeMO1, OeSTO, OePMP3, OeUSP2, OeAP-4, OeWRKY1, OeCCX1 and OeKT2) was detected when comparing ‘Picual’ (tolerant) and ‘Nabali’ (moderately tolerant) cultivars [199], suggesting that different sets of salt-responsive genes are triggered depending both on the stress level (i.e. salt concentration) and the cultivar genotype.
These very early high-throughput studies on the response to salt-stress in olive tree seem to corroborate the general findings described for most plants. However, after the early onset of an osmotic response secondary to salt stress, the number of genes that significantly change their expression in sensitive and tolerant cultivars is very low, suggesting that seed priming, epigenetics and PTMs may be playing an important role in salt response that would merit the effort to analyse. In addition, more comparative studies with more cultivars should be undertaken to have the complete range of olive tree responses to salt stress.
5. Microbiota Changes in Salinised Soils of Olive Groves
5.1. Distribution of Plant-Associated Microorganisms
Plant-associated microorganisms consist of large numbers of bacteria, fungi and archaea taxa potentially interacting among them. They live either inside the plant tissue (endophytes) or on the surface of plant organs (epiphytes) in both the aerial part (phyllosphere or 'above-ground' microbiota) and root region (the 'below-ground' microbiota). The below-ground microbiota, considered as the key hot-spot for plant–microbe interactions, is usually divided in four compartments: the bulk soil, the rhizosphere (outside the root), the rhizoplane (the root surface) and the endosphere (inside the root), each compartment sustaining distinctive microbial communities. The compartments are not isolated since some endophytes are vertically-transmitted to reach the endosphere of the phyllosphere [200]. This has also been described in olive tree, where the phyllosphere seems to come from the soil and reaches aerial plant parts through xylem sap [201] and also protects the plant from diseases and stresses [202]. Plant root-associated microbiota hosts a reservoir of beneficial microorganisms that improve growth, nutritional status, development, and fitness of the host plants, including tolerance to drought and salt stresses; those enhancements are achieved directly by improving nutrient assimilation and protection against the stressing factor [203,204,205] and indirectly by shaping agronomically relevant traits [206,207].
5.2. The Olive Grove Microbiota
Soil microbial composition of olive groves varies depending on the genotype and the studied plant compartment [208,209,210,211], and seems to be modulated by other factors like edaphic condition and soil management [201,212,213], as well as domestication [214]. Microbiota in olive tree roots across different seasonal patterns is highly stable in taxa composition except in cold-susceptible genotypes [206], although a vast amount of novel diversity remains to be discovered in olive grove soils [210].
Proteobacteria, Actinobacteria, Firmicutes and Bacteriodetes dominated the olive tree bacterial microbiota, and in many cases Planctomycetes, Acidobacteria, Myxococcota, Verrucomicrobiota, Gemmatimonadetes, Betaproteobacteria can also be relevant [206,208,214]. It seems that the presence of Proteobacteria in the soil is an indicator of high soil nutrient-content [201]. Proteobacteria and its class Grammaproteobacteria, Firmicutes and Actinobacteria were also highly represented in the olive tree phyllosphere [208,214] while Bacteroidetes were more variable [208,215]. Going deeper in the taxonomy level, the most common genus in soil, leaves and xylem sap [201] were Alphaproteobacteria (more abundant in soil and leaves), Actinobacteria, Sphingobacteria, Bacilli (more abundant in xylem), Gammaproteobacteria and Betaproteobacteria. The most abundant classes in the root endosphere were Alphaproteobacteria, Gammaproteobacteria and Deltaproteobacteria, belonging to Proteobacteria, with (i) Actinophytocola spp. especially relevant for olive tree fitness and health, (ii) Streptomyces, Pseudonocardia, all belonging to Actinobacteria, the most abundant genus in rhyzosphere, and (iii) α-Proteobacteria, Rhizobium, Sphingomonas and Sphingomonas (all Proteobacteria) and Acinetobacter the most abundant in soil [209,210,216].
Concerning the mycobiota whose symbiotic relationships is beneficial under salt stress [217], minor differences were found between the olive tree root endosphere and the soil rhizosphere while there were significant differences between above- and below-ground communities [209,211,218], with predominance of Basidiomycota and inconsistent data about Ascomycota, with four genera (Canalisporium, Macrophomina, Aspergillus and Malassezia) constituting the belowground fungal core [206,208,214]. Sordariomycetes was also the most abundant class found in olive tree fruits and roots [209,218].
5.3. Decrease of Microbiota Biodiversity due to Agricultural Practices
Recent studies support the idea that plant microbiota is a consequence of millions of years of coevolution, where plants likely seek cooperation with microorganisms by means of chemical stimuli as a kind of 'cry for help' strategy to fight stresses [203]. In other level of co-evolution, the beneficial effects of the natural above- and below-ground microbiota are threatened by human agricultural practices related to pesticides, fertilisers or tillage, since they tend to decrease microbiota biodiversity [219]. This prompted to propose a defensebiome [203] to guide the design and construction of beneficial microbial synthetic communities. Low-quality soils of olive groves make endosphere and rhizosphere highly affected by the agronomic practices in a cultivar-dependent way [201,208,220,221]. For example, it was reported in three olive groves with differences in soil and climate (Jaén, Córdoba and Málaga) that Proteobacteria, Actinobacteria and Bacteroidetes were the most abundant bacteria, and that Ascomycota and Basidiomycota were the most abundant fungi. Moreover, Actinophytocola, Streptomyces and Pseudonocardia are the most abundant bacterial genera in the olive tree root endosphere in the Spanish germplasm bank, Actinophytocola being the most prevalent genus by far [209].
5.4. The Olive Tree Microbiota Changes Under Salt Stress
It is known that salt stress affects microbial diversity, community structure, and their functions in many crops [222]: bacterial diversity decreases in high salt, whereas the response of fungi to this stress is more complex. In general, soil salinisation leads to the replacement of salt-sensitive taxons by salt-tolerant ones [223], resulting in a salt-resistant microbial community that may sustain and even promote plant growth [72,224]. The microbiota contributes to plant growth in salinised soils by means of biofilm formation, extracellular polymeric substance production, nitrogen fixation, phytohormone production, nutrient uptake promotion and homeostasis [72,223] in addition to the above mentioned response of phytohormone biosynthesis, signalling pathways, accumulation of osmoprotectants, and regulation of ion transporters. Other microorganisms induce the expression of salt-responsive genes via the action of transcription factors, as well as by post-transcriptional modifications and PTMs [223].
Moderate levels of salt stress can favour bacterial diversity of soil microbial communities instead of their rapid replacement by salt-resistant fungus at high salt levels, suggesting that the interaction of the epigenetic adaptation process in the plant and the microbiota occurs under high salt stress but not at mild salt stress. In olive tree there is a shift of the phyllosphere bacterial communities, with a significant increase of Burkholderia and Ralstonia taxa (both belonging to Burkholderiales) and a decrease of their antagonistic counterpart Pseudomonas (a genus that improves salt tolerance in plants [222,225,226] and is the main taxon of native olive leaf microbiota [215]). In summary, there are many evidences that salt tolerant bacteria can protect plants (including olive tree) from salinisation and would improve productivity and yield under salt stress regimes.
5.5. Soil Amendments and Salt Stress
Restoring the quality of degraded soils by salinisation is a challenging task, especially in arid and semi-arid regions, even though soil health restoration has recently gained increased attention to recover the global biodiversity and habitat (loss driven by human activities and climate change) and rebuild ecosystem resilience and sustainability [227]. Biostimulants (materials of biological origin that enhance plant growth or development when applied to the soil) as well as chemical compounds (such as biochar [228], melatonin [229], silicon or ascorbic acid [67]) have been used in crop (including olive tree) groves as cost-effective soil amendments to increase plant yield, stress tolerance and even soil health and fertility [230]. In some cases, the amendments induce microbial cells to counteract the osmotic stress with the production of osmolytes that mitigate the effects of salinisation [231], demonstrating a close crosstalk between plants and the inhabitant microbiota favoured by the amendment [216]. Chemical amendments have also been used with olive tree cultivars Koroneiki and Mastoidis showing that salt-toxicity symptoms are partially reverted in the presence of 100 mM K+ [232]. Other chemical compounds with CaCl2, MgCl2, H2SO4 (the three added to the saline irrigation water), and gypsiferous material (added to the soil) were able to maintain the tree productivity, although high Mg2+ concentration led to some nutrient imbalances [233]. Adverse effects of salinisation were partially ameliorated by proline in 'Arbequina', 'Arbosana', and 'Koroneiki' [175]. The use of compost, olive mill wastewater and legume cover crops was also beneficial for 'Chemlali' irrigated with saline water under semi-arid conditions [234]. Since Zn2+ is required for ROS detoxification, its supplement improves the antioxidative defence and increases the salt tolerance in many plants, including the olive cultivars Frontoio and Conservolea, by alleviating oxidative injuries [235]. Hence, chemical amendments can be beneficial to olive fruit production when other salt-toxicity mitigation strategies are not available.
When the described conventional strategies cannot alleviate the ion toxicity, are difficult to implement, or are harmful for soil health [236], enhancing biodiversity to promote pedosphere processes appears as an alternative. This is why the manipulation of plant microbiota is an emerging practice to mitigate salt stress, even in olive tree [237]. Several studies have demonstrated that, when inoculated to soils, growth-promoting rhizobacteria (PGPR) from genera Klebsiella, Streptomyces, Pseudomonas, Agrobacterium, Bacillus, Enterobacter, Stenotrophomonas, Rhizobium, and Ochromobacter avoided the over-accumulation of Na+ and maintained the ion homeostasis under salt stress, resulting in an improved crop yield [238]. Several successful studies have identified soil and rhizosphere bacteria (and in less extent fungal taxa) as potential plant growth-promoting microorganism (PGPM) able to reduce the impact of salt stress in crops [222,226,239]. Regarding olive tree, it was described that Glomus mosseae was the most efficient fungus in reducing the detrimental effects of salts in olive plantlets, concomitant with an enhanced K+ concentration in the plant [240], its inoculation in olive tree pot soils being a current practice in nurseries.
6. Bioinformatics Studies in Olive Tree Concerning Salt Stress
6.1. From Olive Tree Transcriptome to OliveAtlas
The first sequencing studies concerning olive tree were performed in the 21th century to obtain information, among other aspects, about the response to biotic and abiotic stresses [241]. The high-throughput sequencing data were later collected to produce the first olive tree transcriptome [242] and then a database for the transcriptome of olive tree reproductive tissues [28]. Microarrays and 454/Roche pyrosequencing were also used, including studies regarding the salt–tolerant and salt–sensitive olive tree cultivars [172,173] discussed above. The authors of the present review have also developed olive-focused bioinformatic tools that were also useful to study salt stress in citrus [243]: (1) RSeqFlow [33] to study differential gene expression that is currently in use to study salt and drought stresses, (2) a comprehensive list of Arabidopsis orthologues to 'Picual' [33] adapted for functional analyses based on the clusterProfiler package (https://github.com/bullones/FunRichR), (3) an R markdown pipeline for new olive allergen detection (https://github.com/bullones/AllergenDetectR), and (4) an R markdown pipeline to detect gene co-expression based on WGCNA package (https://github.com/Javiersdr/Co-expression_analysis).
The continuous growth of high-throughput sequencing data, the availability of five olive cultivar genome sequences and the continuous advances of artificial intelligence algorithms required new bioinformatics approaches. It is time for expression atlas, that (1) integrates expression data from different sources such as RNA-seq, proteomics, metabolomics, expression microarrays, etc., (2) precomputes many analyses using standardised methods, and (3) incorporates multiple interactive visualization methods to explore and analyse expression data. Successful cases of atlas in plants are the Tomato Expression Atlas (TEA) [244], the atlas of the Norway spruce needle seasonal transcriptome [245] and many plants in ePlants (http://bar.utoronto.ca) at BAR [246], PEATmoss for Physcomitrium patens [247], and MangoBase for Mangifera indica [248]. The OliveAtlas (https://www.oliveatlas.uma.es/) for Olea europaea subsp. europaea cultivar Picual was recently published [249] integrating information of 70 RNA-seq experiments, organised by datasets that include, among others, the response to biotic and abiotic stresses. Our ongoing multi-omics data about responses to salt stress of four olive cultivars will be soon integrated in the OliveAtlas. Expression data from other research groups, as well as other olive tree genome references, will be integrated and subjected to ML approaches to provide clues (i.e. genes, biomarkers) for selection of the most suitable cultivars tolerating salt stress. Researchers can find the gene expression levels under different environmental conditions without performing expensive, time-consuming experiments. This would help to select biomarkers and biosensors (genes, polymorphism/alleles, phenotypes, and even microbiota) that can guide both research and agricultural practices to include by introgression the desired tolerance traits in new olive rootstocks or cultivars. Hence, OliveAtlas can be considered the first step towards a future smart (or digital) breeding under the current climate change scenario.
6.2. Machine Learning to Study the Olive Tree
AI in general and ML in particular are virtually unexplored fields in olive tree. A pioneer study developed an algorithm to detect symptoms of olive quick decline syndrome on leaves infected by Xylella fastidiosa from leaf images [250]. The method was able to discover veins and colours that lead to symptomatic leaves. Authors also demonstrate that transfer learning (the use of other model plants where more data can be found to obtain models that will then be tested in another organism) of the same disease in other plants can be leveraged when it is not possible to collect thousands of new leaf images in olive trees, with a promising true positive rate of 98.60 ± 1.47%. A more recent approach was able to detect symptoms of this infection using image processing techniques applied to high-resolution visible and multispectral images [251]. Detection was not only fast but high sensitive (98%) and precise (93%). Another study described the accurate (89.57%) identification of olive cultivars using a combination of morphological characteristics and ISSR (Inter Simple Sequence Repeats) markers analysed through DL [252]. However, studies about salt stress in olive trees remains to be done, in spite of its high potential for elucidating the salt tolerance mechanisms that would foster the selection of salt tolerant rootstock and cultivars [132], as well as optimal microbiota, for salinised soils.
7. Conclusions: Stepping Towards a 'Smart Oliviculture'
There is an urgent need of combat soil salinisation to mitigate the future economic losses and the difficulties of world food supply under the current climate change. The usual agronomical practices to fight against soil salinisation are many times difficult to apply to orchards, particularly olive groves, leaving the use of soil amendments (including microbiota) and salt-tolerant crops as the only realistic approaches for salt stress mitigation. The exploitation of salt tolerant bacteria, both natural and adapted populations, as well as other biostimulants or amendments can protect sensitive plants from salt stress, may improve productivity and yield under salt stress regimes, and would enable the use of salinised lands as olive groves. The use of salt tolerant cultivars or the restoration of the microbial community in salinised soils also protects biodiversity, that is another way to mitigate soil salinisation under climate change. It must be considered that non adaptive responses to salt trigger the mechanisms of PCD and the plant dies. However, metacaspases involved in PCD and autophagy can be the target of new, CRISPR-based biotechnological strategies that can enhance salt tolerance (and likely other stresses) in crop cultivars.
The economical and nutritional interest of olive oil, the abundance of olive groves in lands affected by soil salinisation, and its resilience to ion toxicity makes olive tree a good model to study salt tolerance from the genomics, and preferably multi-omics, point of view. An agricultural advantage of natural or induced salt tolerant olive cultivars is that they exhibited an improved growth and probably required less fertilisers because their roots can better take up nitrogen, while salt-sensitive cultivars did not and required more fertilisers, which exacerbates soil salinisation. The use of salt-tolerant olive rootstocks or cultivars is a good way to mitigate salt stress issues without the need of develop new salt-tolerant cultivars, since most olive cultivars are clones grafted on wild olive trees (any of Olea europaea subsp. europaea var. sylvestris or Olea europaea subsp. cuspidata). When such rootstocks are not available, treatment of olive seed, seedling or even soils with organic amendments that induce PTMs is a promising alternative.
There are many successful cases of transgenic plants becoming salt tolerant by the insertion of one ion transporter in the genome, but this approach is not currently suitable due to the laboratory difficulties to produce transgenic olive plants and to the restrictive regulations about GMOs, even though other biotechnological approaches such as CRISPR-Cas have more opportunities in future field cultures. In any case, it is compulsory that breeding programs aiming to produce new salt-tolerant cultivars or rootstocks know what genome loci will affect the phenotype, so that the availability of genome sequences would facilitate the genetic dissection of complex traits. The olive tree genomics studies are congruent with the morphological and phenomical findings reported for this species in the literature. However, it is a little bit surprising the limited number of genes that were differentially expressed after long-term salt treatments in olive trees and other fruit trees, suggesting that studies about epigenetics and PTMs may be accomplished in olive tree to have the complete picture of salt stress. If the involvement of these processes is clearly demonstrated, new biotechnological approaches may be designed in place to increase the salt tolerance of olive groves.
Future research in the field of salinisation should explore more remote sensors of soil and plant properties besides the analytical and imaging methods already available. The use of AI, particularly ML and SVMs, should guide future research lines and oliviculture practices to cope with climate change in a kind of 'smart oliviculture'. Unfortunately, AI requires an arsenal of data compiled in a compatible way that is unavailable nowadays for olive tree. Transfer learning would fill the gap, but future research should include the collection of large amounts of olive tree-specific data and their integration in an open science resource (for example, by means of the OliveAtlas) that would give scientists and growers a powerful way to find information about basic and applied knowledge related to the soil salinisation and the consequent olive damages. Application of AI to salt stress in olive tree remains an unexplored field yet. Recent advances in sensors, imaging and automation technologies combined with AI have promoted the development of platforms that will facilitate the multi-omics integrations and predictions. The proposed central role of AI in olive tree research, rootstock genetic improvement, microbiota selection and PTM targets should also integrate data from agrisensors, physical mapping, and any other smart agriculture technology to increase the productivity of farmers' operations, minimising risks through more accurate predictions, resulting in improvement of biodiversity, productivity, sustainability and resilience of oliviculture practices and post-harvest processing. This is not science fiction speculation, since Arable (https://www.arable.com) and Semios (https://semios.com/es/) already offer in-field remote sensors to provide farmers real-time data on factors affecting crop health and overall yield, such as soil quality, leaf water potential, and precipitation to help them to manage supply to meet crop demands, identify disease and infection in their crops, and earlier treatment interventions. As a result, the combination of multi-omics (and beyond the realm of omics) data and in-field data with the aid of AI is expected to stimulate sustainable and resilient 'smart oliviculture' by (1) providing clues to manage olive cultivars and groves to cope with salinisation to maintain or increase productivity; and (2) providing information about the possibility to maintain current cultivars irrigated with poor quality water while waiting for new rootstocks that support salt stress, which is known to require less nitric fertilisers (diminishing their leaching into underground water).
Author Contributions
All authors contributed to the conceptualisation and writing—review and editing. MGC was also involved in writing—original draft preparation. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by (i) MCIN/AEI/ERDF-UE grants TED2021-130015B-C21, TED2021-130015B-C22, PID2020-113324GB-I00, PID2020-115853RR-C33, PID2021-125805OA-I00, RED2022-134072-T, RYC2020-030219-I, and Conexión Biología Computacional y Bioinformática 2304210057, and (ii) Consejería de Economía, Conocimiento, Empresas y Universidad, Junta de Andalucía/ERDF-UE grants EMERGIA20_00286, UMA20-FEDERJA-029, PYC20 RE 009 CSIC EEZ and BIO-267.
Institutional Review Board Statement
The study did not require ethical approval.
Acknowledgments
The authors thank Javier Santos del Río (iBIP at IHSM) for sharing its bioinformatics resources and Diana F. Nicolás (BReMAP at EEZ) for her technical support.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
ABA, abscisic acid; AI, artificial intelligence; ATG, autophagy-related gene; DL, deep learning; GMO, genetically modified organism; ISSR, inter simple sequence repeats; ML, machine learning; PCD, programmed cell death; PTM, post-translational modification; RNS, reactive nitrogen species; ROS, reactive oxygen species; RSS, reactive sulphur species; SOD, superoxide dismutase; SOS, salt overly sensitive; SUMO, small ubiquitin-like modifier; SVM, support vector machine; VPEs, vacuolar processing enzymes.
References
- Daliakopoulos, I.N.; Tsanis, I.K.; Koutroulis, A.; Kourgialas, N.N.; Varouchakis, A.E.; Karatzas, G.P.; Ritsema, C.J. The Threat of Soil Salinity: A European Scale Review. Sci. Total Environ. 2016, 573, 727–739. [Google Scholar] [CrossRef] [PubMed]
- Hassani, A.; Azapagic, A.; Shokri, N. Global Predictions of Primary Soil Salinization under Changing Climate in the 21st Century. Nat. Commun. 2021, 12, 6663. [Google Scholar] [CrossRef] [PubMed]
- Calvin, K.; Dasgupta, D.; Krinner, G.; Mukherji, A.; Thorne, P.W.; Trisos, C.; Romero, J.; Aldunce, P.; Barrett, K.; Blanco, G.; et al. IPCC, 2023: Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change [Core Writing Team, H. Lee and J. Romero (Eds.)]. IPCC, Geneva, Switzerland.; First.; Intergovernmental Panel on Climate Change (IPCC), 2023;
- Shahid, S.A.; Zaman, M.; Heng, L. Introduction to Soil Salinity, Sodicity and Diagnostics Techniques. In Guideline for Salinity Assessment, Mitigation and Adaptation Using Nuclear and Related Techniques; Springer International Publishing: Cham, 2018; ISBN 978-3-319-96189-7. [Google Scholar]
- Whittle, A.; Barnett, R.L.; Charman, D.J.; Gallego-Sala, A.V. Low-Salinity Transitions Drive Abrupt Microbial Response to Sea-Level Change. Ecol. Lett. 2022, 25, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, P.; Kumar, R. Soil Salinity: A Serious Environmental Issue and Plant Growth Promoting Bacteria as One of the Tools for Its Alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef]
- Singh, A. Soil Salinity: A Global Threat to Sustainable Development. Soil Use Manag. 2022, 38, 39–67. [Google Scholar] [CrossRef]
- Stavi, I.; Thevs, N.; Priori, S. Soil Salinity and Sodicity in Drylands: A Review of Causes, Effects, Monitoring, and Restoration Measures. Front. Environ. Sci. 2021, 9, 712831. [Google Scholar] [CrossRef]
- Majeed, A.; Siyyar, S. Salinity Stress Management in Field Crops: An Overview of the Agronomic Approaches. In Plant Ecophysiology and Adaptation under Climate Change: Mechanisms and Perspectives II.; Hasanuzzaman, M., Ed.; Springer Singapore: Singapore, 2020; ISBN 9789811521713. [Google Scholar]
- Romero-Trigueros, C.; Vivaldi, G.A.; Nicolás, E.N.; Paduano, A.; Salcedo, F.P.; Camposeo, S. Ripening Indices, Olive Yield and Oil Quality in Response to Irrigation With Saline Reclaimed Water and Deficit Strategies. Front. Plant Sci. 2019, 10, 1243. [Google Scholar] [CrossRef]
- Meena, M.D.; Yadav, R.K.; Narjary, B.; Yadav, G.; Jat, H.S.; Sheoran, P.; Meena, M.K.; Antil, R.S.; Meena, B.L.; Singh, H.V.; et al. Municipal Solid Waste (MSW): Strategies to Improve Salt Affected Soil Sustainability: A Review. Waste Manag. 2019, 84, 38–53. [Google Scholar] [CrossRef]
- Munns, R.; Gilliham, M. Salinity Tolerance of Crops – What Is the Cost? New Phytol. 2015, 208, 668–673. [Google Scholar] [CrossRef]
- Chele, K.H.; Tinte, M.M.; Piater, L.A.; Dubery, I.A.; Tugizimana, F. Soil Salinity, a Serious Environmental Issue and Plant Responses: A Metabolomics Perspective. Metabolites 2021, 11, 724. [Google Scholar] [CrossRef]
- Atta, K.; Mondal, S.; Gorai, S.; Singh, A.P.; Kumari, A.; Ghosh, T.; Roy, A.; Hembram, S.; Gaikwad, D.J.; Mondal, S.; et al. Impacts of Salinity Stress on Crop Plants: Improving Salt Tolerance through Genetic and Molecular Dissection. Front. Plant Sci. 2023, 14, 1241736. [Google Scholar] [CrossRef]
- El Yamani, M.; Cordovilla, M.D.P. Tolerance Mechanisms of Olive Tree (Olea europaea) Under Saline Conditions. Plants 2024, 13, 2094. [Google Scholar] [CrossRef] [PubMed]
- Fraga, H.; Moriondo, M.; Leolini, L.; Santos, J.A. Mediterranean Olive Orchards under Climate Change: A Review of Future Impacts and Adaptation Strategies. Agronomy 2020, 11, 56. [Google Scholar] [CrossRef]
- Dini, I.; Laneri, S. Spices, Condiments, Extra Virgin Olive Oil and Aromas as Not Only Flavorings, but Precious Allies for Our Wellbeing. Antioxidants 2021, 10, 868. [Google Scholar] [CrossRef]
- Unver, T.; Wu, Z.; Sterck, L.; Turktas, M.; Lohaus, R.; Li, Z.; Yang, M.; He, L.; Deng, T.; Escalante, F.J.; et al. Genome of Wild Olive and the Evolution of Oil Biosynthesis. Proc. Natl. Acad. Sci. 2017, 114, E9413–E9422. [Google Scholar] [CrossRef]
- Maestri, D.; Barrionuevo, D.; Bodoira, R.; Zafra, A.; Jiménez-López, J.; Alché, J. de D. Nutritional Profile and Nutraceutical Components of Olive (Olea europaea L.) Seeds. J. Food Sci. Technol. 2019, 56, 4359–4370. [Google Scholar] [CrossRef]
- Georgiadou, E.C.; Koubouris, G.; Goulas, V.; Sergentani, C.; Nikoloudakis, N.; Manganaris, G.A.; Kalaitzis, P.; Fotopoulos, V. Genotype-dependent Regulation of Vitamin E Biosynthesis in Olive Fruits as Revealed through Metabolic and Transcriptional Profiles. Plant Biol. 2019, 21, 604–614. [Google Scholar] [CrossRef]
- Bruno, L.; Picardi, E.; Pacenza, M.; Chiappetta, A.; Muto, A.; Gagliardi, O.; Muzzalupo, I.; Pesole, G.; Bitonti, M.B. Changes in Gene Expression and Metabolic Profile of Drupes of Olea europaea L. Cv Carolea in Relation to Maturation Stage and Cultivation Area. BMC Plant Biol. 2019, 19, 428. [Google Scholar] [CrossRef]
- Mougiou, N.; Trikka, F.; Trantas, E.; Ververidis, F.; Makris, A.; Argiriou, A.; Vlachonasios, K.E. Expression of Hydroxytyrosol and Oleuropein Biosynthetic Genes Are Correlated with Metabolite Accumulation during Fruit Development in Olive, Olea Europaea, Cv. Koroneiki. Plant Physiol. Biochem. 2018, 128, 41–49. [Google Scholar] [CrossRef]
- Iaria, D.L.; Chiappetta, A.; Muzzalupo, I. A De Novo Transcriptomic Approach to Identify Flavonoids and Anthocyanins “Switch-Off” in Olive (Olea europaea L.) Drupes at Different Stages of Maturation. Front. Plant Sci. 2016, 6, 1246. [Google Scholar] [CrossRef]
- Parvini, F.; Sicardo, M.D.; Hosseini-Mazinani, M.; Martínez-Rivas, J.M.; Hernández, M.L. Transcriptional Analysis of Stearoyl-Acyl Carrier Protein Desaturase Genes from Olive (Olea europaea ) in Relation to the Oleic Acid Content of the Virgin Olive Oil. J. Agric. Food Chem. 2016, 64, 7770–7781. [Google Scholar] [CrossRef] [PubMed]
- Yanik, H.; Turktas, M.; Dundar, E.; Hernandez, P.; Dorado, G.; Unver, T. Genome-Wide Identification of Alternate Bearing-Associated microRNAs (miRNAs) in Olive (Olea europaea L.). BMC Plant Biol. 2013, 13, 10. [Google Scholar] [CrossRef] [PubMed]
- García-López, M.C.; Vidoy, I.; Jiménez-Ruiz, J.; Muñoz-Mérida, A.; Fernández-Ocaña, A.; de la Rosa, R.; Barroso, J.B.; Navarro, F.; Trelles, O.; Beuzón, C.R.; et al. Genetic Changes Involved in the Juvenile-to-Adult Transition in the Shoot Apex of Olea europaea L. Occur Years before the First Flowering. Tree Genet. Genomes 2014, 10, 585–603. [Google Scholar] [CrossRef]
- Jiménez-Ruiz, J.; García-López, M.C.; Vidoy, I.; de la O Leyva-Pérez, M.; Fernández-Ocaña, A.; Barroso, J.B.; Barceló, A.; Beuzón, C.R.; de la Rosa, R.; Luque, F. Transcriptional Analysis of Adult Cutting and Juvenile Seedling Olive Roots. Tree Genet. Genomes 2015, 11, 77. [Google Scholar] [CrossRef]
- Carmona, R.; Zafra, A.; Seoane, P.; Castro, A.J.; Guerrero-Fernández, D.; Castillo-Castillo, T.; Medina-García, A.; Cánovas, F.M.; Aldana-Montes, J.F.; Navas-Delgado, I.; et al. ReprOlive: A Database with Linked Data for the Olive Tree (Olea europaea L.) Reproductive Transcriptome. Front. Plant Sci. 2015, 6, 625. [Google Scholar] [CrossRef]
- Rejón, J.; Delalande, F.; Schaeffer-Reiss, C.; Alché, J.; Rodríguez-García, M.; Van Dorsselaer, A.; Castro, A. The Pollen Coat Proteome: At the Cutting Edge of Plant Reproduction. Proteomes 2016, 4, 5. [Google Scholar] [CrossRef]
- Zafra, A.; Carmona, R.; Traverso, J.A.; Hancock, J.T.; Goldman, M.H.S.; Claros, M.G.; Hiscock, S.J.; Alche, J.D. Identification and Functional Annotation of Genes Differentially Expressed in the Reproductive Tissues of the Olive Tree (Olea europaea L.) through the Generation of Subtractive Libraries. Front. Plant Sci. 2017, 8, 1576. [Google Scholar] [CrossRef]
- Farinelli, D.; Breton, C.; Koubouris, G.; Famiani, F.; Villemur, P.; Bervillé, A. Reply to Saumitou-Laprade et al. (2017) “Controlling for Genetic Identity of Varieties, Pollen Contamination and Stigma Receptivity Is Essential to Characterize the Self-Incompatibility System of Olea europaea L.”. Evol. Appl. 2018, 11, 1465–1470. [Google Scholar] [CrossRef]
- Alagna, F.; Caceres, M.E.; Pandolfi, S.; Collani, S.; Mousavi, S.; Mariotti, R.; Cultrera, N.G.M.; Baldoni, L.; Barcaccia, G. The Paradox of Self-Fertile Varieties in the Context of Self-Incompatible Genotypes in Olive. Front. Plant Sci. 2019, 10, 725. [Google Scholar] [CrossRef]
- Bullones, A.; Castro, A.J.; Lima-Cabello, E.; Fernandez-Pozo, N.; Bautista, R.; Alché, J.D.D.; Claros, M.G. Transcriptomic Insight into the Pollen Tube Growth of Olea europaea L. subsp. europaea Reveals Reprogramming and Pollen-Specific Genes Including New Transcription Factors. Plants 2023, 12, 2894. [Google Scholar] [CrossRef]
- Gros-Balthazard, M.; Besnard, G.; Sarah, G.; Holtz, Y.; Leclercq, J.; Santoni, S.; Wegmann, D.; Glémin, S.; Khadari, B. Evolutionary Transcriptomics Reveals the Origins of Olives and the Genomic Changes Associated with Their Domestication. Plant J. 2019, 100, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Castro, A.J.; de Dios Alché, J.; Cuevas, J.; Romero, P.J.; Alché, V.; Rodríguez-García, M.I. Pollen from Different Olive Tree Cultivars Contains Varying Amounts of the Major Allergen Ole e 1. Int. Arch. Allergy Immunol. 2003, 131, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Alché, J.D.; Castro, A.J.; Jiménez-López, J.C.; Morales, S.; Zafra, A.; Hamman-Khalifa, A.M.; Rodríguez-García, M.I. Differential Characteristics of Olive Pollen from Different Cultivars: Biological and Clinical Implications. J. Investig. Allergol. Clin. Immunol. 2007, 17 Suppl 1, 17–23. [Google Scholar]
- Hamman-Khalifa, A.; Castro, A.; Jiménez-López, J.; Rodríguez-García, M.; Alché, J. Olive Cultivar Origin Is a Major Cause of Polymorphism for Ole e 1 Pollen Allergen. BMC Plant Biol. 2008, 8, 10. [Google Scholar] [CrossRef] [PubMed]
- Grasso, F.; Coppola, M.; Carbone, F.; Baldoni, L.; Alagna, F.; Perrotta, G.; Pérez-Pulido, A.J.; Garonna, A.; Facella, P.; Daddiego, L.; et al. The Transcriptional Response to the Olive Fruit Fly (Bactrocera oleae) Reveals Extended Differences between Tolerant and Susceptible Olive (Olea europaea L.) Varieties. PLOS ONE 2017, 12, e0183050. [Google Scholar] [CrossRef]
- León, L.; De La Rosa, R.; Arriaza, M. Prioritization of Olive Breeding Objectives in Spain: Analysis of a Producers and Researchers Survey. Span. J. Agric. Res. 2021, 19, e0701. [Google Scholar] [CrossRef]
- Gucci, R.; Tattini, M. Salinity Tolerance in Olive. In Horticultural Reviews; Janick, J., Ed.; John Wiley & Sons, Inc.: Oxford, UK, 2010; ISBN 978-0-470-65066-0. [Google Scholar]
- Lavee, S. Evaluation of the Need and Present Potential of Olive Breeding Indicating the Nature of the Available Genetic Resources Involved. Sci. Hortic. 2013, 161, 333–339. [Google Scholar] [CrossRef]
- Hualpa-Ramirez, E.; Carrasco-Lozano, E.C.; Madrid-Espinoza, J.; Tejos, R.; Ruiz-Lara, S.; Stange, C.; Norambuena, L. Stress Salinity in Plants: New Strategies to Cope with in the Foreseeable Scenario. Plant Physiol. Biochem. 2024, 208, 108507. [Google Scholar] [CrossRef]
- Romero-Aranda, R.; Moya, J.L.; Tadeo, F.R.; Legaz, F.; Primo-Millo, E.; Talon, M. Physiological and Anatomical Disturbances Induced by Chloride Salts in Sensitive and Tolerant Citrus: Beneficial and Detrimental Effects of Cations. Plant Cell Environ. 1998, 21, 1243–1253. [Google Scholar] [CrossRef]
- Romero-Aranda, R.; Soria, T.; Cuartero, J. Tomato Plant-Water Uptake and Plant-Water Relationships under Saline Growth Conditions. Plant Sci. 2001, 160, 265–272. [Google Scholar] [CrossRef]
- Muchate, N.S.; Nikalje, G.C.; Rajurkar, N.S.; Suprasanna, P.; Nikam, T.D. Plant Salt Stress: Adaptive Responses, Tolerance Mechanism and Bioengineering for Salt Tolerance. Bot. Rev. 2016, 82, 371–406. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Zhu, J.-K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef]
- Romero-Aranda, M.R.; González-Fernández, P.; Pérez-Tienda, J.R.; López-Diaz, M.R.; Espinosa, J.; Granum, E.; Traverso, J.Á.; Pineda, B.; Garcia-Sogo, B.; Moreno, V.; et al. Na+ Transporter HKT1;2 Reduces Flower Na+ Content and Considerably Mitigates the Decline in Tomato Fruit Yields under Saline Conditions. Plant Physiol. Biochem. 2020, 154, 341–352. [Google Scholar] [CrossRef]
- Niu, G.; Rodriguez, D.S.; Starman, T. Response of Bedding Plants to Saline Water Irrigation. HortScience 2010, 45, 628–636. [Google Scholar] [CrossRef]
- Sánchez-Blanco, M.J.; Álvarez, S.; Ortuño, M.F.; Ruiz-Sánchez, M.C. Root System Response to Drought and Salinity: Root Distribution and Water Transport. In Root Engineering; Morte, A., Varma, A., Eds.; Soil Biology; Springer Berlin Heidelberg: Berlin, Heidelberg, 2014; ISBN 978-3-642-54275-6. [Google Scholar]
- Plant Roots. The Hidden Half; Waisel, Y., Eshel, A., Beeckman, T., Kafkafi, U., Eds.; 3rd ed.; CRC Press: Boca Ratón, 2002; ISBN 978-0-203-90942-3. [Google Scholar]
- Tu, T.Q.; Vaciaxa, P.; Lo, T.T.M.; Nguyen, N.H.; Pham, N.T.T.; Nguyen, Q.H.; Do, P.T.; Nguyen, L.T.N.; Nguyen, Y.T.H.; Chu, M.H. GmDREB6, a Soybean Transcription Factor, Notably Affects the Transcription of the NtP5CS and NtCLC Genes in Transgenic Tobacco under Salt Stress Conditions. Saudi J. Biol. Sci. 2021, 28, 7175–7181. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive Oxygen Species (ROS) and Response of Antioxidants as ROS-Scavengers during Environmental Stress in Plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- Singh, A.; Roychoudhury, A. Gene Regulation at Transcriptional and Post-transcriptional Levels to Combat Salt Stress in Plants. Physiol. Plant. 2021, 173, 1556–1572. [Google Scholar] [CrossRef]
- Balasubramaniam, T.; Shen, G.; Esmaeili, N.; Zhang, H. Plants’ Response Mechanisms to Salinity Stress. Plants 2023, 12, 2253. [Google Scholar] [CrossRef]
- Mansour, M.M.F.; Hassan, F.A.S. How Salt Stress-Responsive Proteins Regulate Plant Adaptation to Saline Conditions. Plant Mol. Biol. 2022, 108, 175–224. [Google Scholar] [CrossRef]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.-K.; Bohnert, H.J. Plant Celular and Molecular Responses to High Salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef]
- Bruetschy, C. The EU Regulatory Framework on Genetically Modified Organisms (GMOs). Transgenic Res. 2019, 28, 169–174. [Google Scholar] [CrossRef]
- Soltabayeva, A.; Ongaltay, A.; Omondi, J.O.; Srivastava, S. Morphological, Physiological and Molecular Markers for Salt-Stressed Plants. Plants 2021, 10, 243. [Google Scholar] [CrossRef]
- van Zelm, E.; Zhang, Y.; Testerink, C. Salt Tolerance Mechanisms of Plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef]
- Afzal, M.Z.; Jia, Q.; Ibrahim, A.K.; Niyitanga, S.; Zhang, L. Mechanisms and Signaling Pathways of Salt Tolerance in Crops: Understanding from the Transgenic Plants. Trop. Plant Biol. 2020, 13, 297–320. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of Plant Responses to Salt Stress. Int. J. Mol. Sci. 2021, 22, 4609. [Google Scholar] [CrossRef]
- Zhou, H.; Shi, H.; Yang, Y.; Feng, X.; Chen, X.; Xiao, F.; Lin, H.; Guo, Y. Insights into Plant Salt Stress Signaling and Tolerance. J. Genet. Genomics Yi Chuan Xue Bao 2024, 51, 16–34. [Google Scholar] [CrossRef]
- Najar, M.A. Updates on Protein Post-Translational Modifications for Modulating Response to Salinity. In Genetics of Salt Tolerance in Plants; Ganie, S.A., Wani, S.H., Eds.; CABI: GB, 2024; ISBN 978-1-80062-301-9. [Google Scholar]
- Mata-Pérez, C.; Sánchez-Vicente, I.; Arteaga, N.; Gómez-Jiménez, S.; Fuentes-Terrón, A.; Oulebsir, C.S.; Calvo-Polanco, M.; Oliver, C.; Lorenzo, Ó. Functions of Nitric Oxide-Mediated Post-Translational Modifications under Abiotic Stress. Front. Plant Sci. 2023, 14, 1158184. [Google Scholar] [CrossRef]
- Benjamin, J.J.; Miras-Moreno, B.; Araniti, F.; Salehi, H.; Bernardo, L.; Parida, A.; Lucini, L. Proteomics Revealed Distinct Responses to Salinity between the Halophytes Suaeda maritima (L.) Dumort and Salicornia brachiata (Roxb). Plants 2020, 9, 227. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Nafees, M.; Chen, J.; Darras, A.; Ferrante, A.; Hancock, J.T.; Ashraf, M.; Zaid, A.; Latif, N.; Corpas, F.J.; et al. Chemical Priming Enhances Plant Tolerance to Salt Stress. Front. Plant Sci. 2022, 13, 946922. [Google Scholar] [CrossRef]
- Tang, R.-J.; Wang, C.; Li, K.; Luan, S. The CBL-CIPK Calcium Signaling Network: Unified Paradigm from 20 Years of Discoveries. Trends Plant Sci. 2020, 25, 604–617. [Google Scholar] [CrossRef]
- Ismail, A.; Takeda, S.; Nick, P. Life and Death under Salt Stress: Same Players, Different Timing? J. Exp. Bot. 2014, 65, 2963–2979. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of Plant Responses to Salt Stress. Int. J. Mol. Sci. 2021, 22, 4609. [Google Scholar] [CrossRef]
- Raja, V.; Majeed, U.; Kang, H.; Andrabi, K.I.; John, R. Abiotic Stress: Interplay between ROS, Hormones and MAPKs. Environ. Exp. Bot. 2017, 137, 142–157. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, S.; Gaurav, A.K.; Srivastava, S.; Verma, J.P. Plant Growth-Promoting Bacteria: Biological Tools for the Mitigation of Salinity Stress in Plants. Front. Microbiol. 2020, 11, 1216. [Google Scholar] [CrossRef]
- Dabravolski, S.A.; Isayenkov, S.V. The Regulation of Plant Cell Wall Organisation under Salt Stress. Front. Plant Sci. 2023, 14, 1118313. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, W.; Long, S.; Zhao, C. Maintenance of Cell Wall Integrity under High Salinity. Int. J. Mol. Sci. 2021, 22, 3260. [Google Scholar] [CrossRef]
- Lei, X.; Liu, Z.; Xie, Q.; Fang, J.; Wang, C.; Li, J.; Wang, C.; Gao, C. Construction of Two Regulatory Networks Related to Salt Stress and Lignocellulosic Synthesis under Salt Stress Based on a Populus davidiana × P. bolleana Transcriptome Analysis. Plant Mol. Biol. 2022, 109, 689–702. [Google Scholar] [CrossRef]
- Venkataraman, G.; Shabala, S.; Véry, A.-A.; Hariharan, G.N.; Somasundaram, S.; Pulipati, S.; Sellamuthu, G.; Harikrishnan, M.; Kumari, K.; Shabala, L.; et al. To Exclude or to Accumulate? Revealing the Role of the Sodium HKT1;5 Transporter in Plant Adaptive Responses to Varying Soil Salinity. Plant Physiol. Biochem. 2021, 169, 333–342. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Salinity Tolerance in Halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef]
- Cheeseman, J.M. The Evolution of Halophytes, Glycophytes and Crops, and Its Implications for Food Security under Saline Conditions. New Phytol. 2015, 206, 557–570. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, P.M. Sodium (Na+) Homeostasis and Salt Tolerance of Plants. Environ. Exp. Bot. 2013, 92, 19–31. [Google Scholar] [CrossRef]
- Hussain, S.; Zhang, J.; Zhong, C.; Zhu, L.; Cao, X.; Yu, S.; Allen Bohr, J.; Hu, J.; Jin, Q. Effects of Salt Stress on Rice Growth, Development Characteristics, and the Regulating Ways: A Review. J. Integr. Agric. 2017, 16, 2357–2374. [Google Scholar] [CrossRef]
- Kronzucker, H.J.; Coskun, D.; Schulze, L.M.; Wong, J.R.; Britto, D.T. Sodium as Nutrient and Toxicant. Plant Soil 2013, 369, 1–23. [Google Scholar] [CrossRef]
- Munns, R.; James, R.A.; Gilliham, M.; Flowers, T.J.; Colmer, T.D. Tissue Tolerance: An Essential but Elusive Trait for Salt-Tolerant Crops. Funct. Plant Biol. 2016, 43, 1103. [Google Scholar] [CrossRef]
- Tester, M. Na+ Tolerance and Na+ Transport in Higher Plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef]
- Brumós, J.; Colmenero-Flores, J.M.; Conesa, A.; Izquierdo, P.; Sánchez, G.; Iglesias, D.J.; López-Climent, M.F.; Gómez-Cadenas, A.; Talón, M. Membrane Transporters and Carbon Metabolism Implicated in Chloride Homeostasis Differentiate Salt Stress Responses in Tolerant and Sensitive Citrus Rootstocks. Funct. Integr. Genomics 2009, 9, 293–309. [Google Scholar] [CrossRef]
- Brumós, J.; Talón, M.; Bouhlal, R.; Colmenero-Flores, J.M. Cl- Homeostasis in Includer and Excluder Citrus Rootstocks: Transport Mechanisms and Identification of Candidate Genes. Plant Cell Environ. 2010, 33, 2012–2027. [Google Scholar] [CrossRef]
- Li, B.; Tester, M.; Gilliham, M. Chloride on the Move. Trends Plant Sci. 2017, 22, 236–248. [Google Scholar] [CrossRef]
- Celis, N.; Suarez, D.L.; Wu, L.; Li, R.; Arpaia, M.L.; Mauk, P. Salt Tolerance and Growth of 13 Avocado Rootstocks Related Best to Chloride Uptake. HortScience 2018, 53, 1737–1745. [Google Scholar] [CrossRef]
- Colmenero-Flores, J.M.; Franco-Navarro, J.D.; Cubero-Font, P.; Peinado-Torrubia, P.; Rosales, M.A. Chloride as a Beneficial Macronutrient in Higher Plants: New Roles and Regulation. Int. J. Mol. Sci. 2019, 20, 4686. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Sandhu, N.; Kumar, P.; Pruthi, G.; Singh, J.; Kaur, S.; Chhuneja, P. Genome-Wide Identification and in Silico Analysis of NPF, NRT2, CLC and SLAC1/SLAH Nitrate Transporters in Hexaploid Wheat (Triticum aestivum). Sci. Rep. 2022, 12, 11227. [Google Scholar] [CrossRef] [PubMed]
- Rajappa, S.; Krishnamurthy, P.; Huang, H.; Yu, D.; Friml, J.; Xu, J.; Kumar, P.P. The Translocation of a Chloride Channel from the Golgi to the Plasma Membrane Helps Plants Adapt to Salt Stress. Nat. Commun. 2024, 15, 3978. [Google Scholar] [CrossRef] [PubMed]
- Wleklik, K.; Borek, S. Vacuolar Processing Enzymes in Plant Programmed Cell Death and Autophagy. Int. J. Mol. Sci. 2023, 24, 1198. [Google Scholar] [CrossRef] [PubMed]
- Aina, O.; Bakare, O.O.; Fadaka, A.O.; Keyster, M.; Klein, A. Plant Biomarkers as Early Detection Tools in Stress Management in Food Crops: A Review. Planta 2024, 259, 60. [Google Scholar] [CrossRef]
- Mani, B.; Kaur, I.; Dhingra, Y.; Saxena, V.; Krishna, G.K.; Kumar, R.; Chinnusamy, V.; Agarwal, M.; Katiyar-Agarwal, S. Tetraspanin 5 Orchestrates Resilience to Salt Stress through the Regulation of Ion and Reactive Oxygen Species Homeostasis in Rice. Plant Biotechnol. J. 2024, pbi.14476. [CrossRef]
- Kotula, L.; Garcia Caparros, P.; Zörb, C.; Colmer, T.D.; Flowers, T.J. Improving Crop Salt Tolerance Using Transgenic Approaches: An Update and Physiological Analysis. Plant Cell Environ. 2020, 43, 2932–2956. [Google Scholar] [CrossRef]
- Stokstad, E. European Parliament Votes to Ease Regulation of Gene-Edited Crops. Science 2024. [Google Scholar] [CrossRef]
- Puchta, H. Regulation of Gene-Edited Plants in Europe: From the Valley of Tears into the Shining Sun? aBIOTECH 2023, 5, 231–238. [Google Scholar] [CrossRef]
- Affenzeller, M.J.; Darehshouri, A.; Andosch, A.; Lutz, C.; Lutz-Meindl, U. Salt Stress-Induced Cell Death in the Unicellular Green Alga Micrasterias denticulata. J. Exp. Bot. 2009, 60, 939–954. [Google Scholar] [CrossRef]
- Joseph, B.; Jini, D. Salinity Induced Programmed Cell Death in Plants: Challenges and Opportunities for Salt-Tolerant Plants. J. Plant Sci. 2010, 5, 376–390. [Google Scholar] [CrossRef]
- Shabala, S. Salinity and Programmed Cell Death: Unravelling Mechanisms for Ion Specific Signalling. J. Exp. Bot. 2009, 60, 709–712. [Google Scholar] [CrossRef]
- Kim, Y.; Wang, M.; Bai, Y.; Zeng, Z.; Guo, F.; Han, N.; Bian, H.; Wang, J.; Pan, J.; Zhu, M. Bcl-2 Suppresses Activation of VPEs by Inhibiting Cytosolic Ca2+ Level with Elevated K+ Efflux in NaCl-Induced PCD in Rice. Plant Physiol. Biochem. 2014, 80, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Minina, E.A.; Filonova, L.H.; Fukada, K.; Savenkov, E.I.; Gogvadze, V.; Clapham, D.; Sanchez-Vera, V.; Suarez, M.F.; Zhivotovsky, B.; Daniel, G.; et al. Autophagy and Metacaspase Determine the Mode of Cell Death in Plants. J. Cell Biol. 2013, 203, 917–927. [Google Scholar] [CrossRef]
- Bozhkov, P.V.; Suarez, M.F.; Filonova, L.H.; Daniel, G.; Zamyatnin, A.A.; Rodriguez-Nieto, S.; Zhivotovsky, B.; Smertenko, A. Cysteine Protease mcII-Pa Executes Programmed Cell Death during Plant Embryogenesis. Proc. Natl. Acad. Sci. U. S. A. 2005, 102, 14463–14468. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.; Yu, X.-H.; Wang, C.; Zhang, Q.; Liu, W.; McSweeney, S.; Shanklin, J.; Lam, E.; Liu, Q. Structural Basis for Ca2+-Dependent Activation of a Plant Metacaspase. Nat. Commun. 2020, 11, 2249. [Google Scholar] [CrossRef] [PubMed]
- Garcia, N.; Kalicharan, R.E.; Kinch, L.; Fernandez, J. Regulating Death and Disease: Exploring the Roles of Metacaspases in Plants and Fungi. Int. J. Mol. Sci. 2022, 24, 312. [Google Scholar] [CrossRef] [PubMed]
- Yue, J.-Y.; Wang, Y.-J.; Jiao, J.-L.; Wang, W.-W.; Wang, H.-Z. The Metacaspase TaMCA-Id Negatively Regulates Salt-Induced Programmed Cell Death and Functionally Links With Autophagy in Wheat. Front. Plant Sci. 2022, 13, 904933. [Google Scholar] [CrossRef]
- Minina, E.A.; Smertenko, A.P.; Bozhkov, P.V. Vacuolar Cell Death in Plants: Metacaspase Releases the Brakes on Autophagy. Autophagy 2014, 10, 928–929. [Google Scholar] [CrossRef]
- Chen, H.; Dong, J.; Wang, T. Autophagy in Plant Abiotic Stress Management. Int. J. Mol. Sci. 2021, 22, 4075. [Google Scholar] [CrossRef]
- Koh, E.; Sunil, R.S.; Lam, H.Y.I.; Mutwil, M. Confronting the Data Deluge: How Artificial Intelligence Can Be Used in the Study of Plant Stress. Comput. Struct. Biotechnol. J. 2024, 23, 3454–3466. [Google Scholar] [CrossRef]
- Kamilaris, A.; Prenafeta-Boldú, F.X. Deep Learning in Agriculture: A Survey. Comput Electron Agric 2018, 147, 70–90. [Google Scholar] [CrossRef]
- Khalifani, S.; Darvishzadeh, R.; Azad, N.; Seyed Rahmani, R. Prediction of Sunflower Grain Yield under Normal and Salinity Stress by RBF, MLP and, CNN Models. Ind Crops Prod 2022, 189, 115762. [Google Scholar] [CrossRef]
- Navarro, A.; Nicastro, N.; Costa, C.; Pentangelo, A.; Cardarelli, M.; Ortenzi, L.; Pallottino, F.; Cardi, T.; Pane, C. Sorting Biotic and Abiotic Stresses on Wild Rocket by Leaf-Image Hyperspectral Data Mining with an Artificial Intelligence Model. Plant Methods 2022, 18, 45. [Google Scholar] [CrossRef]
- Zhou, C.; Ye, H.; Sun, D.; Yue, J.; Yang, G.; Hu, J. An Automated, High-Performance Approach for Detecting and Characterizing Broccoli Based on UAV Remote-Sensing and Transformers: A Case Study from Haining, China. Int J Appl Earth Obs Geoinf 2022, 114, 103055. [Google Scholar] [CrossRef]
- Ho Tong Minh, D.; Ienco, D.; Gaetano, R.; Lalande, N.; Ndikumana, E.; Osman, F.; Maurel, P. Deep Recurrent Neural Networks for Winter Vegetation Quality Mapping via Multitemporal SAR Sentinel-1. IEEE Geosci. Remote Sens. Lett. 2018, 15, 464–468. [Google Scholar] [CrossRef]
- Atef, M.; Khattab, A.; Agamy, E.A.; Khairy, M.M. Deep Learning Based Time-Series Forecasting Framework for Olive Precision Farming. In Proceedings of the 2021 IEEE International Midwest Symposium on Circuits and Systems (MWSCAS); August 2021; pp. 1062–1065. [Google Scholar]
- Alhnaity, B.; Pearson, S.; Leontidis, G.; Kollias, S. Using Deep Learning to Predict Plant Growth and Yield in Greenhouse Environments. Acta Hortic. 2019, 1296, 55. [Google Scholar] [CrossRef]
- Zarbakhsh, S.; Shahsavar, A.R. Artificial Neural Network-Based Model to Predict the Effect of γ-Aminobutyric Acid on Salinity and Drought Responsive Morphological Traits in Pomegranate. Sci Rep 2022, 12, 16662. [Google Scholar] [CrossRef]
- Gao, G.; Tester, M.A.; Julkowska, M.M. The Use of High-Throughput Phenotyping for Assessment of Heat Stress-Induced Changes in Arabidopsis. Plant Phenomics 2020, 2020, 3723916. [Google Scholar] [CrossRef]
- Raza, A.; Gangurde, S.S.; Singh Sandhu, K.; Lv, Y. Omics-Assisted Crop Improvement under Abiotic Stress Conditions. Plant Stress 2024, 100626. [Google Scholar] [CrossRef]
- Meher, P.K.; Sahu, T.K.; Gupta, A.; Kumar, A.; Rustgi, S. ASRpro: A Machine-Learning Computational Model for Identifying Proteins Associated with Multiple Abiotic Stress in Plants. Plant Genome 2022, 17, e20259. [Google Scholar] [CrossRef]
- Pradhan, U.K.; Meher, P.K.; Naha, S.; Rao, A.R.; Kumar, U.; Pal, S.; Gupta, A. ASmiR: A Machine Learning Framework for Prediction of Abiotic Stress–Specific miRNAs in Plants. Funct Integr Genomics 2023, 23, 92. [Google Scholar] [CrossRef] [PubMed]
- Medina, C.A.; Hawkins, C.; Liu, X.-P.; Peel, M.; Yu, L.-X. Genome-Wide Association and Prediction of Traits Related to Salt Tolerance in Autotetraploid Alfalfa (Medicago sativa L.). Int J Mol Sci 2020, 21, 3361. [Google Scholar] [CrossRef] [PubMed]
- Medina, C.A.; Kaur, H.; Ray, I.; Yu, L.-X. Strategies to Increase Prediction Accuracy in Genomic Selection of Complex Traits in Alfalfa (Medicago sativa L.). Cells 2021, 10, 3372. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.; Ahn, H.; Lee, S.; Lee, C.-J.; Hur, J.; Jung, W.; Kim, S. StressGenePred: A Twin Prediction Model Architecture for Classifying the Stress Types of Samples and Discovering Stress-Related Genes in Arabidopsis. BMC Genomics 2019, 20, 949. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Chaudhary, P.; Taunk, J.; Singh, C.K.; Singh, D.; Tomar, R.S.S.; Aski, M.; Konjengbam, N.S.; Raje, R.S.; Singh, S.; et al. Fab Advances in Fabaceae for Abiotic Stress Resilience: From “Omics” to Artificial Intelligence. Int J Mol Sci 2021, 22, 10535. [Google Scholar] [CrossRef]
- Ullah, M.A.; Abdullah-Zawawi, M.-R.; Zainal-Abidin, R.-A.; Sukiran, N.L.; Uddin, M.I.; Zainal, Z. A Review of Integrative Omic Approaches for Understanding Rice Salt Response Mechanisms. Plants 2022, 11, 1430. [Google Scholar] [CrossRef]
- Rohart, F.; Gautier, B.; Singh, A.; Lê Cao, K.-A. mixOmics: An R Package for ‘omics Feature Selection and Multiple Data Integration. PLOS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef]
- Tuncbag, N.; Gosline, S.J.C.; Kedaigle, A.; Soltis, A.R.; Gitter, A.; Fraenkel, E. Network-Based Interpretation of Diverse High-Throughput Datasets through the Omics Integrator Software Package. PLOS Comput. Biol. 2016, 12, e1004879. [Google Scholar] [CrossRef]
- Zoppi, J.; Guillaume, J.-F.; Neunlist, M.; Chaffron, S. MiBiOmics: An Interactive Web Application for Multi-Omics Data Exploration and Integration. BMC Bioinformatics 2021, 22, 6. [Google Scholar] [CrossRef]
- Zhou, J.; Zhang, B.; Li, G.; Chen, X.; Li, H.; Xu, X.; Chen, S.; He, W.; Xu, C.; Liu, L.; et al. An AI Agent for Fully Automated Multi-Omic Analyses. Adv. Sci. 2024, 2407094. [Google Scholar] [CrossRef]
- Wang, H.; Cimen, E.; Singh, N.; Buckler, E. Deep Learning for Plant Genomics and Crop Improvement. Curr. Opin. Plant Biol. 2020, 54, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Zaied, Y.B.; Zouabi, O. Impacts of Climate Change on Tunisian Olive Oil Output. Clim. Change 2016, 139, 535–549. [Google Scholar] [CrossRef]
- Harfouche, A.L.; Jacobson, D.A.; Kainer, D.; Romero, J.C.; Harfouche, A.H.; Scarascia Mugnozza, G.; Moshelion, M.; Tuskan, G.A.; Keurentjes, J.J.B.; Altman, A. Accelerating Climate Resilient Plant Breeding by Applying Next-Generation Artificial Intelligence. Trends Biotechnol. 2019, 37, 1217–1235. [Google Scholar] [CrossRef] [PubMed]
- Bey, N.E.; Maazoun, A.M.; Nahdi, O.; Krima, N.B.; Aounallah, M.K. Water Stress Indicators in Citrus, Olive and Apple Trees: A Review. J. Appl. Hortic. 2024, 26, 3–9. [Google Scholar] [CrossRef]
- Chartzoulakis, K.; Loupassaki, M.; Androulakis, I. Comparative Study on NaCl Salinity Tolerance of Six Olive Cultivars. Acta Hortic. 2002, 497–501. [Google Scholar] [CrossRef]
- Chartzoulakis, K.; Loupassaki, M.; Bertaki, M.; Androulakis, I. Effects of NaCl Salinity on Growth, Ion Content and CO2 Assimilation Rate of Six Olive Cultivars. Sci. Hortic. 2002, 96, 235–247. [Google Scholar] [CrossRef]
- Kchaou, H.; Larbi, A.; Gargouri, K.; Chaieb, M.; Morales, F.; Msallem, M. Assessment of Tolerance to NaCl Salinity of Five Olive Cultivars, Based on Growth Characteristics and Na+ and Cl− Exclusion Mechanisms. Sci. Hortic. 2010, 124, 306–315. [Google Scholar] [CrossRef]
- Soda, N.; Ephrath, J.E.; Dag, A.; Beiersdorf, I.; Presnov, E.; Yermiyahu, U.; Ben-Gal, A. Root Growth Dynamics of Olive (Olea europaea L.) Affected by Irrigation Induced Salinity. Plant Soil 2017, 411, 305–318. [Google Scholar] [CrossRef]
- Rugini, E.; Fedeli, E. Olive (Olea Europaea L.) as an Oilseed Crop. In Legumes and Oilseed Crops I.; Bajaj, Y.P.S., Ed.; Biotechnology in Agriculture and Forestry; Springer Berlin Heidelberg: Berlin, Heidelberg, 1990; ISBN 978-3-642-74450-1. [Google Scholar]
- Chartzoulakis, K.S. Salinity and Olive: Growth, Salt Tolerance, Photosynthesis and Yield. Agric. Water Manag. 2005, 78, 108–121. [Google Scholar] [CrossRef]
- Tattini, M.; Mergar, J.C.; Traversi, M.L. Responses of Olea Europaea to High Salinity : A Brief-Ecophysiological-Review. Adv. Hortic. Sci. 2008. [Google Scholar] [CrossRef]
- Regni, L.; Del Pino, A.M.; Mousavi, S.; Palmerini, C.A.; Baldoni, L.; Mariotti, R.; Mairech, H.; Gardi, T.; D’Amato, R.; Proietti, P. Behavior of Four Olive Cultivars During Salt Stress. Front. Plant Sci. 2019, 10, 867. [Google Scholar] [CrossRef]
- Loreto, F.; Centritto, M.; Chartzoulakis, K. Photosynthetic Limitations in Olive Cultivars with Different Sensitivity to Salt Stress: Photosynthetic Limitations and Salt Stress in Olive Cultivars. Plant Cell Environ. 2003, 26, 595–601. [Google Scholar] [CrossRef]
- Ben Abdallah, M.; Trupiano, D.; Polzella, A.; De Zio, E.; Sassi, M.; Scaloni, A.; Zarrouk, M.; Ben Youssef, N.; Scippa, G.S. Unraveling Physiological, Biochemical and Molecular Mechanisms Involved in Olive (Olea europaea L. Cv. Chétoui ) Tolerance to Drought and Salt Stresses. J. Plant Physiol. 2018, 220, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Sofo, A.; Dichio, B.; Xiloyannis, C.; Masia, A. Lipoxygenase Activity and Proline Accumulation in Leaves and Roots of Olive Trees in Response to Drought Stress. Physiol. Plant. 2004, 121, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Melgar, J.C.; Syvertsen, J.P.; Martínez, V.; García-Sánchez, F. The Relative Salt Tolerance of ‘Rangpur’ Seedlings and ‘Arbequina’ Olive Cuttings. Proc Fla State Hort Soc 2007, 120, 79–84. [Google Scholar]
- Moretti, S.; Francini, A.; Hernández, M.L.; Martínez-Rivas, J.M.; Sebastiani, L. Effect of Saline Irrigation on Physiological Traits, Fatty Acid Composition and Desaturase Genes Expression in Olive Fruit Mesocarp. Plant Physiol. Biochem. 2019, 141, 423–430. [Google Scholar] [CrossRef]
- Tattini, M.; Traversi, M. On the Mechanism of Salt Tolerance in Olive (Olea europaea L.) under Low- or High-Ca2+ Supply. Environ. Exp. Bot. 2009, 65, 72–81. [Google Scholar] [CrossRef]
- Cimato, A.; Castelli, S.; Tattini, M.; Traversi, M.L. An Ecophysiological Analysis of Salinity Tolerance in Olive. Environ. Exp. Bot. 2010, 68, 214–221. [Google Scholar] [CrossRef]
- Sodini, M.; Astolfi, S.; Francini, A.; Sebastiani, L. Multiple Linear Regression and Linear Mixed Models Identify Novel Traits of Salinity Tolerance in Olea europaea L. Tree Physiol. 2022, 42, 1029–1042. [Google Scholar] [CrossRef]
- Tadić, J.; Dumičić, G.; Veršić Bratinčević, M.; Vitko, S.; Radić Brkanac, S. Physiological and Biochemical Response of Wild Olive (Olea europaea subsp. europaea var. sylvestris) to Salinity. Front. Plant Sci. 2021, 12, 712005. [Google Scholar] [CrossRef]
- Goreta, S.; Bučević-Popović, V.; Pavela-Vrančić, M.; Perica, S. Salinity-induced Changes in Growth, Superoxide Dismutase Activity, and Ion Content of Two Olive Cultivars. J. Plant Nutr. Soil Sci. 2007, 170, 398–403. [Google Scholar] [CrossRef]
- Petridis, A.; Therios, I.; Samouris, G.; Tananaki, C. Salinity-Induced Changes in Phenolic Compounds in Leaves and Roots of Four Olive Cultivars (Olea europaea L.) and Their Relationship to Antioxidant Activity. Environ. Exp. Bot. 2012, 79, 37–43. [Google Scholar] [CrossRef]
- Rossi, L.; Borghi, M.; Francini, A.; Lin, X.; Xie, D.-Y.; Sebastiani, L. Salt Stress Induces Differential Regulation of the Phenylpropanoid Pathway in Olea europaea Cultivars Frantoio (Salt-Tolerant) and Leccino (Salt-Sensitive). J. Plant Physiol. 2016, 204, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Palm, E.R.; Salzano, A.M.; Vergine, M.; Negro, C.; Guidi Nissim, W.; Sabbatini, L.; Balestrini, R.; De Pinto, M.C.; Fortunato, S.; Gohari, G.; et al. Response to Salinity Stress in Four Olea europaea L. Genotypes: A Multidisciplinary Approach. Environ. Exp. Bot. 2024, 218, 105586. [Google Scholar] [CrossRef]
- Ahmed, Ch.B.; Rouina, B.B.; Sensoy, S.; Boukhris, M.; Abdallah, F.B. Changes in Gas Exchange, Proline Accumulation and Antioxidative Enzyme Activities in Three Olive Cultivars under Contrasting Water Availability Regimes. Environ. Exp. Bot. 2009, 67, 345–352. [Google Scholar] [CrossRef]
- Cordovilla, M. del P.; Aparicio, C.; Melendo, M.; Bueno, M. Exogenous Application of Indol-3-Acetic Acid and Salicylic Acid Improves Tolerance to Salt Stress in Olive Plantlets (Olea europaea L. cultivar Picual) in Growth Chamber Environments. Agronomy 2023, 13, 647. [Google Scholar] [CrossRef]
- Aparicio, C.; Urrestarazu, M.; Cordovilla, M.D.P. Comparative Physiological Analysis of Salinity Effects in Six Olive Genotypes. HortScience 2014, 49, 901–904. [Google Scholar] [CrossRef]
- Bashir, M.A.; Silvestri, C.; Coppa, E.; Brunori, E.; Cristofori, V.; Rugini, E.; Ahmad, T.; Hafiz, I.A.; Abbasi, N.A.; Nawaz Shah, M.K.; et al. Response of Olive Shoots to Salinity Stress Suggests the Involvement of Sulfur Metabolism. Plants 2021, 10, 350. [Google Scholar] [CrossRef]
- Mousavi, S.; Regni, L.; Bocchini, M.; Mariotti, R.; Cultrera, N.G.M.; Mancuso, S.; Googlani, J.; Chakerolhosseini, M.R.; Guerrero, C.; Albertini, E.; et al. Physiological, Epigenetic and Genetic Regulation in Some Olive Cultivars under Salt Stress. Sci. Rep. 2019, 9, 1093. [Google Scholar] [CrossRef]
- Tabatabaei, S.J. Salinity Stress and Olive: An Overview. Plant Stress 2007, 1, 105–112. [Google Scholar]
- Azimi, M.; Khoshzaman, T.; Taheri, M.; Dadras, A. Evaluation of Salinity Tolerance of Three Olive (Olea europaea L.) Cultivars. J. Cent. Eur. Agric. 2021, 22, 571–581. [Google Scholar] [CrossRef]
- Rahemi, M.; Karimi, S.; Sedaghat, S.; Ali Rostami, A. Physiological Responses of Olive Cultivars to Salinity Stress. Adv. Hortic. Sci. 2017, 31, 53–59. [Google Scholar] [CrossRef]
- Ayaz, M.; Varol, N.; Yolcu, S.; Pelvan, A.; Kaya, Ü.; Aydoğdu, E.; Bor, M.; Özdemir, F.; Türkan, İ. Three (Turkish) Olive Cultivars Display Contrasting Salt Stress-Coping Mechanisms under High Salinity. Trees 2021, 35, 1283–1298. [Google Scholar] [CrossRef]
- Chartzoulakis, K.S. The Potential of Saline and Residual Water Use in Olive Growing. Acta Hortic. 2014, 257–273. [Google Scholar] [CrossRef]
- Marin, L.; Benlloch, M.; Fernández-Escobar, R. Screening of Olive Cultivars for Salt Tolerance. Sci. Hortic. 1995, 64, 113–116. [Google Scholar] [CrossRef]
- Kchaou, H.; Larbi, A.; Gargouri, K.; Chaieb, M.; Morales, F.; Msallem, M. Assessment of Tolerance to NaCl Salinity of Five Olive Cultivars, Based on Growth Characteristics and Na+ and Cl− Exclusion Mechanisms. Sci. Hortic. 2010, 124, 306–315. [Google Scholar] [CrossRef]
- Aragüés, R.; Guillén, M.; Royo, A. Five-Year Growth and Yield Response of Two Young Olive Cultivars (Olea europaea L., Cvs. Arbequina and Empeltre) to Soil Salinity. Plant Soil 2010, 334, 423–432. [Google Scholar] [CrossRef]
- Bakshi, P. ; Amit; Wali, V.K. Mitigation Strategies of Abiotic Stress in Fruit Crops. In Abiotic & Biotic Stress Management in Plants, Ed.; CRC Press: London, 2022; ISBN 978-1-00-328198-6. [Google Scholar]
- Al-Absi, K.; Qrunfleh, M.; Abu-Sharar, T. Mechanism of Salt Tolerance of Two Olive Olea europaea L. Cultivars as Related to Electrolyte Concentration and Toxicity. Acta Hortic. 2003, 618, 281–290. [Google Scholar] [CrossRef]
- Perica, S.; Brkljača, M.; Goreta, S.; Romić, M. Vegetative Growth and Salt Accumulation of Six Olive Cultivars under Salt Stress. Acta Hortic. 2004, 664, 555–560. [Google Scholar] [CrossRef]
- AL-Ouda, A. Evaluation of the Response of Some Syrian Olive (Olea Europea L.) Cultivars to Salinity Stress.
- Bazakos, C.; Manioudaki, M.E.; Therios, I.; Voyiatzis, D.; Kafetzopoulos, D.; Awada, T.; Kalaitzis, P. Comparative Transcriptome Analysis of Two Olive Cultivars in Response to NaCl-Stress. PLoS ONE 2012, 7, e42931. [Google Scholar] [CrossRef]
- Bazakos, C.; Manioudaki, M.E.; Sarropoulou, E.; Spano, T.; Kalaitzis, P. 454 Pyrosequencing of Olive (Olea europaea L.) Transcriptome in Response to Salinity. PLOS ONE 2015, 10, e0143000. [Google Scholar] [CrossRef] [PubMed]
- Demir, S.; Cetinkaya, H. Effects of Saline Conditions on Polyphenol and Protein Content and Photosynthetic Response of Different Olive (Olea europaea L.). Appl. Ecol. Environ. Res. 2020, 18, 2599–2610. [Google Scholar] [CrossRef]
- Poury, N.; Seifi, E.; Alizadeh, M. Effects of Salinity and Proline on Growth and Physiological Characteristics of Three Olive Cultivars. Gesunde Pflanz. 2023, 75, 1169–1180. [Google Scholar] [CrossRef]
- El-Sayed Emtithal, H.; El-Said, M.E.; El-Sherif, A.H.; Sari El-Deen, S.A. Estudios químicos sobre la tolerancia a la sanidad de algunos cultivares. Olivae 1996, 64, 52–57. [Google Scholar]
- Mousavi, S.; Mariotti, R.; Valeri, M.C.; Regni, L.; Lilli, E.; Albertini, E.; Proietti, P.; Businelli, D.; Baldoni, L. Characterization of Differentially Expressed Genes under Salt Stress in Olive. Int. J. Mol. Sci. 2021, 23, 154. [Google Scholar] [CrossRef] [PubMed]
- Hassan, I.; Gaballah, M.; Bedour, A.; Proietti, P.; Regni, L. Salinity Stress Effects on Three Different Olive Cultivars and the Possibility of Their Cultivation in Reclaimed Lands. Plant Arch. 2020, 20, 2378–2382. [Google Scholar]
- Therios, I.N.; Misopolinos, N.D. Genotypic Response to Sodium Chloride Salinity of Four Major Olive Cultivars (Olea europea L.). Plant Soil 1988, 106, 105–111. [Google Scholar] [CrossRef]
- Skodra, C.; Michailidis, M.; Dasenaki, M.; Ganopoulos, I.; Thomaidis, N.S.; Tanou, G.; Molassiotis, A. Unraveling Salt-responsive Tissue-specific Metabolic Pathways in Olive Tree. Physiol. Plant. 2021, 173, 1643–1656. [Google Scholar] [CrossRef]
- Skodra, C.; Michailidis, M.; Moysiadis, T.; Stamatakis, G.; Ganopoulou, M.; Adamakis, I.-D.S.; Angelis, L.; Ganopoulos, I.; Tanou, G.; Samiotaki, M.; et al. Disclosing the Molecular Basis of Salinity Priming in Olive Trees Using Proteogenomic Model Discovery. Plant Physiol. 2023, 191, 1913–1933. [Google Scholar] [CrossRef]
- Briccoli-Bati, C.; Basta, P.; Tocci, C.; Turco, D. Influencia del riego con agua salobre en jóvenes plantas de oliva. Olivae 1994, 53, 35–38. [Google Scholar]
- Cruz, F.; Julca, I.; Gómez-Garrido, J.; Loska, D.; Marcet-Houben, M.; Cano, E.; Galán, B.; Frias, L.; Ribeca, P.; Derdak, S.; et al. Genome Sequence of the Olive Tree, Olea europaea. GigaScience 2016, 5, 29. [Google Scholar] [CrossRef] [PubMed]
- Julca, I.; Marcet-Houben, M.; Cruz, F.; Gómez-Garrido, J.; Gaut, B.S.; Díez, C.M.; Gut, I.G.; Alioto, T.S.; Vargas, P.; Gabaldón, T. Genomic Evidence for Recurrent Genetic Admixture during the Domestication of Mediterranean Olive Trees (Olea europaea L.). BMC Biol. 2020, 18, 148. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Ruiz, J.; Ramírez-Tejero, J.A.; Fernández-Pozo, N.; Leyva-Pérez, M. de la O.; Yan, H.; Rosa, R. de la; Belaj, A.; Montes, E.; Rodríguez-Ariza, M.O.; Navarro, F.; et al. Transposon Activation Is a Major Driver in the Genome Evolution of Cultivated Olive Trees (Olea europaea L.). Plant Genome 2020, 13, e20010. [Google Scholar] [CrossRef] [PubMed]
- Rao, G.; Zhang, J.; Liu, X.; Lin, C.; Xin, H.; Xue, L.; Wang, C. De Novo Assembly of a New Olea europaea Genome Accession Using Nanopore Sequencing. Hortic. Res. 2021, 8, 64. [Google Scholar] [CrossRef]
- Lv, J.; Jiang, C.; Wu, W.; Mao, K.; Wei, Q.; Zheng, Y.; Gao, C.; Niu, Z.; Jin, G.; Zhang, R.; et al. The Gapless Genome Assembly and Multi-Omics Analyses Unveil a Pivotal Regulatory Mechanism of Oil Biosynthesis in the Olive Tree. Hortic. Res. 2024, 11, uhae168. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, J.; Peng, D.; Tian, Y.; Zhao, D.; Ni, W.; Long, J.; Li, J.; Zeng, Y.; Wu, Z.; et al. High-Quality Genome Assembly of Olea europaea subsp. cuspidata Provides Insights Into Its Resistance to Fungal Diseases in the Summer Rain Belt in East Asia. Front. Plant Sci. 2022, 13, 879822. [Google Scholar] [CrossRef]
- Wu, T.; Ma, T.; Xu, T.; Pan, L.; Zhang, Y.; Li, Y.; Ning, D. The De Novo Genome Assembly of Olea europaea subsp. cuspidata, a Widely Distributed Olive Close Relative. Front. Genet. 2022, 13, 868540. [Google Scholar] [CrossRef]
- Khadivi, A.; Mirheidari, F.; Saeidifar, A.; Moradi, Y. Morphological Characterizations of Olea europaea subsp. cuspidata. Genet. Resour. Crop Evol. 2024, 71, 1837–1853. [Google Scholar] [CrossRef]
- Yolcu, S.; Alavilli, H.; Ganesh, P.; Panigrahy, M.; Song, K. Salt and Drought Stress Responses in Cultivated Beets (Beta vulgaris L.) and Wild Beet (Beta maritima L.). Plants 2021, 10, 1843. [Google Scholar] [CrossRef]
- Zhang, J.; Jiang, D.; Liu, B.; Luo, W.; Lu, J.; Ma, T.; Wan, D. Transcriptome Dynamics of a Desert Poplar (Populus pruinosa) in Response to Continuous Salinity Stress. Plant Cell Rep. 2014, 33, 1565–1579. [Google Scholar] [CrossRef]
- Jamil, A.; Riaz, S.; Ashraf, M.; Foolad, M.R. Gene Expression Profiling of Plants under Salt Stress. Crit. Rev. Plant Sci. 2011, 30, 435–458. [Google Scholar] [CrossRef]
- Hoang, X.L.T.; Nhi, D.N.H.; Thu, N.B.A.; Thao, N.P.; Tran, L.-S.P. Transcription Factors and Their Roles in Signal Transduction in Plants under Abiotic Stresses. Curr. Genomics 2017, 18, 483–497. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Dai, M.; Yao, J.; Xiao, B.; Li, X.; Zhang, Q.; Xiong, L. Overexpressing a NAM, ATAF, and CUC (NAC) Transcription Factor Enhances Drought Resistance and Salt Tolerance in Rice. Proc. Natl. Acad. Sci. 2006, 103, 12987–12992. [Google Scholar] [CrossRef] [PubMed]
- Cabello, J.V.; Lodeyro, A.F.; Zurbriggen, M.D. Novel Perspectives for the Engineering of Abiotic Stress Tolerance in Plants. Curr. Opin. Biotechnol. 2014, 26, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Rallo, L.; Barranco, D.; Díez, C.M.; Rallo, P.; Suárez, M.P.; Trapero, C.; Pliego-Alfaro, F. Strategies for Olive (Olea europaea L.) Breeding: Cultivated Genetic Resources and Crossbreeding. In Advances in Plant Breeding Strategies: Fruits; Al-Khayri, J.M., Jain, S.M., Johnson, D.V., Eds.; Springer International Publishing: Cham, 2018; ISBN 978-3-319-91943-0. [Google Scholar]
- Jiang, X.; Zhang, W.; Fernie, A.R.; Wen, W. Combining Novel Technologies with Interdisciplinary Basic Research to Enhance Horticultural Crops. Plant J. 2022, 109, 35–46. [Google Scholar] [CrossRef]
- Sadder, M.T.; Ateyyeh, A.F.; Alswalmah, H.; Zakri, A.M.; Alsadon, A.A.; Al-Doss, A.A. Characterization of Putative Salinity-Responsive Biomarkers in Olive (Olea europaea L.). Plant Genet. Resour. Charact. Util. 2021, 19, 133–143. [Google Scholar] [CrossRef]
- Qin, Y.; Druzhinina, I.S.; Pan, X.; Yuan, Z. Microbially Mediated Plant Salt Tolerance and Microbiome-Based Solutions for Saline Agriculture. Biotechnol. Adv. 2016, 34, 1245–1259. [Google Scholar] [CrossRef]
- Fausto, C.; Mininni, A.N.; Sofo, A.; Crecchio, C.; Scagliola, M.; Dichio, B.; Xiloyannis, C. Olive Orchard Microbiome: Characterisation of Bacterial Communities in Soil-Plant Compartments and Their Comparison between Sustainable and Conventional Soil Management Systems. Plant Ecol. Divers. 2018, 11, 597–610. [Google Scholar] [CrossRef]
- Costa, D.; Fernandes, T.; Martins, F.; Pereira, J.A.; Tavares, R.M.; Santos, P.M.; Baptista, P.; Lino-Neto, T. Illuminating Olea europaea L. Endophyte Fungal Community. Microbiol. Res. 2021, 245, 126693. [Google Scholar] [CrossRef]
- Liu, H.; Brettell, L.E.; Qiu, Z.; Singh, B.K. Microbiome-Mediated Stress Resistance in Plants. Trends Plant Sci. 2020, 25, 733–743. [Google Scholar] [CrossRef]
- Pascale, A.; Proietti, S.; Pantelides, I.S.; Stringlis, I.A. Modulation of the Root Microbiome by Plant Molecules: The Basis for Targeted Disease Suppression and Plant Growth Promotion. Front. Plant Sci. 2020, 10, 1741. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Dias, M.C.; Freitas, H. Drought and Salinity Stress Responses and Microbe-Induced Tolerance in Plants. Front. Plant Sci. 2020, 11, 591911. [Google Scholar] [CrossRef] [PubMed]
- Chialva, M.; De Rose, S.; Novero, M.; Lanfranco, L.; Bonfante, P. Plant Genotype and Seasonality Drive Fine Changes in Olive Root Microbiota. Curr. Plant Biol. 2021, 28, 100219. [Google Scholar] [CrossRef]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant-Microbiome Interactions: From Community Assembly to Plant Health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef] [PubMed]
- Mina, D.; Pereira, J.A.; Lino-Neto, T.; Baptista, P. Epiphytic and Endophytic Bacteria on Olive Tree Phyllosphere: Exploring Tissue and Cultivar Effect. Microb. Ecol. 2020, 80, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Fernández-González, A.J.; Villadas, P.J.; Gómez-Lama Cabanás, C.; Valverde-Corredor, A.; Belaj, A.; Mercado-Blanco, J.; Fernández-López, M. Defining the Root Endosphere and Rhizosphere Microbiomes from the World Olive Germplasm Collection. Sci. Rep. 2019, 9, 20423. [Google Scholar] [CrossRef]
- Malacrinò, A.; Mosca, S.; Li Destri Nicosia, M.G.; Agosteo, G.E.; Schena, L. Plant Genotype Shapes the Bacterial Microbiome of Fruits, Leaves, and Soil in Olive Plants. Plants 2022, 11, 613. [Google Scholar] [CrossRef]
- Abdelfattah, A.; Li Destri Nicosia, M.G.; Cacciola, S.O.; Droby, S.; Schena, L. Metabarcoding Analysis of Fungal Diversity in the Phyllosphere and Carposphere of Olive (Olea europaea). PLOS ONE 2015, 10, e0131069. [Google Scholar] [CrossRef]
- Caliz, J.; Montes-Borrego, M.; Triadó-Margarit, X.; Metsis, M.; Landa, B.B.; Casamayor, E.O. Influence of Edaphic, Climatic, and Agronomic Factors on the Composition and Abundance of Nitrifying Microorganisms in the Rhizosphere of Commercial Olive Crops. PLOS ONE 2015, 10, e0125787. [Google Scholar] [CrossRef]
- Sofo, A.; Ricciuti, P.; Fausto, C.; Mininni, A.N.; Crecchio, C.; Scagliola, M.; Malerba, A.D.; Xiloyannis, C.; Dichio, B. The Metabolic and Genetic Diversity of Soil Bacterial Communities Depends on the Soil Management System and C/N Dynamics: The Case of Sustainable and Conventional Olive Groves. Appl. Soil Ecol. 2019, 137, 21–28. [Google Scholar] [CrossRef]
- Müller, H.; Berg, C.; Landa, B.B.; Auerbach, A.; Moissl-Eichinger, C.; Berg, G. Plant Genotype-Specific Archaeal and Bacterial Endophytes but Similar Bacillus Antagonists Colonize Mediterranean Olive Trees. Front. Microbiol. 2015, 6, 138. [Google Scholar] [CrossRef] [PubMed]
- Vita, F.; Sabbatini, L.; Sillo, F.; Ghignone, S.; Vergine, M.; Guidi Nissim, W.; Fortunato, S.; Salzano, A.M.; Scaloni, A.; Luvisi, A.; et al. Salt Stress in Olive Tree Shapes Resident Endophytic Microbiota. Front. Plant Sci. 2022, 13, 992395. [Google Scholar] [CrossRef] [PubMed]
- Fernández-González, A.J.; Ramírez-Tejero, J.A.; Nevado-Berzosa, M.P.; Luque, F.; Fernández-López, M.; Mercado-Blanco, J. Coupling the Endophytic Microbiome with the Host Transcriptome in Olive Roots. Comput. Struct. Biotechnol. J. 2021, 19, 4777–4789. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Meng, Y.; Zhao, M.; Wang, M.; Wang, C.; Dong, J.; Fan, W.; Xu, F.; Wang, D.; Xie, Z. Advances and Mechanisms of Fungal Symbionts in Improving the Salt Tolerance of Crops. Plant Sci. 2024, 349, 112261. [Google Scholar] [CrossRef]
- Martins, F.; Pereira, J.A.; Bota, P.; Bento, A.; Baptista, P. Fungal Endophyte Communities in Above- and Belowground Olive Tree Organs and the Effect of Season and Geographic Location on Their Structures. Fungal Ecol. 2016, 20, 193–201. [Google Scholar] [CrossRef]
- Fernández-González, A.J.; Wentzien, N.M.; Villadas, P.J.; Valverde-Corredor, A.; Lasa, A.V.; Gómez-Lama Cabanás, C.; Mercado-Blanco, J.; Fernández-López, M. Comparative Study of Neighboring Holm Oak and Olive Trees-Belowground Microbial Communities Subjected to Different Soil Management. PLOS ONE 2020, 15, e0236796. [Google Scholar] [CrossRef]
- Llimós, M.; Segarra, G.; Sancho-Adamson, M.; Trillas, M.I.; Romanyà, J. Impact of Olive Saplings and Organic Amendments on Soil Microbial Communities and Effects of Mineral Fertilization. Front. Microbiol. 2021, 12, 653027. [Google Scholar] [CrossRef]
- Sofo, A.; Palese, A.M.; Casacchia, T.; Xiloyannis, C. Sustainable Soil Management in Olive Orchards. In Emerging Technologies and Management of Crop Stress Tolerance; Elsevier, 2014; pp. 471–483 ISBN 978-0-12-800875-1.
- Kumar, V.; Kumar, P.; Khan, A. Optimization of PGPR and Silicon Fertilization Using Response Surface Methodology for Enhanced Growth, Yield and Biochemical Parameters of French Bean (Phaseolus vulgaris L.) under Saline Stress. Biocatal. Agric. Biotechnol. 2020, 23, 101463. [Google Scholar] [CrossRef]
- Roy, S.; Chakraborty, A.P.; Chakraborty, R. Understanding the Potential of Root Microbiome Influencing Salt-tolerance in Plants and Mechanisms Involved at the Transcriptional and Translational Level. Physiol. Plant. 2021, 173, 1657–1681. [Google Scholar] [CrossRef]
- Mercado-Blanco, J.; Abrantes, I.; Barra Caracciolo, A.; Bevivino, A.; Ciancio, A.; Grenni, P.; Hrynkiewicz, K.; Kredics, L.; Proença, D.N. Belowground Microbiota and the Health of Tree Crops. Front. Microbiol. 2018, 9, 1006. [Google Scholar] [CrossRef]
- Costa-Gutierrez, S.B.; Lami, M.J.; Santo, M.C.C.-D.; Zenoff, A.M.; Vincent, P.A.; Molina-Henares, M.A.; Espinosa-Urgel, M.; de Cristóbal, R.E. Plant Growth Promotion by Pseudomonas putida KT2440 under Saline Stress: Role of eptA. Appl. Microbiol. Biotechnol. 2020, 104, 4577–4592. [Google Scholar] [CrossRef] [PubMed]
- Vives-Peris, V.; Gómez-Cadenas, A.; Pérez-Clemente, R.M. Salt Stress Alleviation in Citrus Plants by Plant Growth-Promoting Rhizobacteria Pseudomonas putida and Novosphingobium Sp. Plant Cell Rep. 2018, 37, 1557–1569. [Google Scholar] [CrossRef] [PubMed]
- Raupp, P.; Carrillo, Y.; Nielsen, U.N. Soil Health to Enhance Ecological Restoration and Conservation. J. Sustain. Agric. Environ. 2024, 3, e70022. [Google Scholar] [CrossRef]
- Kapoor, A.; Sharma, R.; Kumar, A.; Sepehya, S. Biochar as a Means to Improve Soil Fertility and Crop Productivity: A Review. J. Plant Nutr. 2022, 45, 2380–2388. [Google Scholar] [CrossRef]
- Khan, Z.; Jan, R.; Asif, S.; Farooq, M.; Jang, Y.-H.; Kim, E.-G.; Kim, N.; Kim, K.-M. Exogenous Melatonin Induces Salt and Drought Stress Tolerance in Rice by Promoting Plant Growth and Defense System. Sci. Rep. 2024, 14, 1214. [Google Scholar] [CrossRef]
- Ding, Z.; Kheir, A.M.S.; Ali, M.G.M.; Ali, O.A.M.; Abdelaal, A.I.N.; Lin, X.; Zhou, Z.; Wang, B.; Liu, B.; He, Z. The Integrated Effect of Salinity, Organic Amendments, Phosphorus Fertilizers, and Deficit Irrigation on Soil Properties, Phosphorus Fractionation and Wheat Productivity. Sci. Rep. 2020, 10, 2736. [Google Scholar] [CrossRef]
- Khan, I.; Mahmood, S.; Chattha, M.U.; Bilal Chattha, M.; Ahmad, S.; Awan, M.I.; Alqahtani, F.M.; Hashem, M.; Alhaithloul, H.A.S.; Qari, S.H.; et al. Organic Amendments Improved the Productivity and Bio-Fortification of Fine Rice by Improving Physiological Responses and Nutrient Homeostasis under Salinity Stress. Plants 2023, 12, 1644. [Google Scholar] [CrossRef]
- Chartzoulakis, K.; Psarras, G.; Vemmos, S.; Loupassaki, M.; Bertaki, M. Response of Two Olive Cultivars to Salt Stress and Potassium Supplement. J. Plant Nutr. 2006, 29, 2063–2078. [Google Scholar] [CrossRef]
- Ziskin, R.; Dag, A.; Yermiyahu, U.; Levy, G.J. Different Amendments for Combating Soil Sodicity in an Olive Orchard. Agric. Water Manag. 2024, 299, 108837. [Google Scholar] [CrossRef]
- Chehab, H.; Tekaya, M.; Hajlaoui, H.; Abdelhamid, S.; Gouiaa, M.; Sfina, H.; Chihaoui, B.; Boujnah, D.; Mechri, B. Complementary Irrigation with Saline Water and Soil Organic Amendments Modified Soil Salinity, Leaf Na+, Productivity and Oil Phenols of Olive Trees (Cv. Chemlali) Grown under Semiarid Conditions. Agric. Water Manag. 2020, 237, 106183. [Google Scholar] [CrossRef]
- Naeini, N.R.; Esna-Ashari, M.; Khoshgoftarmanesh, A.; Cheraghi, H.; Hatami, A.; Mirzapour, M. The Effect of Zinc Nutrition on Two Olive (Olea europaea L.) Cultivars Components and Alleviate Oxidative Damage in Salinity Conditions. J. Med. Plants Byprod. 2016, 1, 23–31. [Google Scholar]
- Rodriguez, R.J.; Henson, J.; Van Volkenburgh, E.; Hoy, M.; Wright, L.; Beckwith, F.; Kim, Y.-O.; Redman, R.S. Stress Tolerance in Plants via Habitat-Adapted Symbiosis. ISME J. 2008, 2, 404–416. [Google Scholar] [CrossRef] [PubMed]
- Kumawat, K.C.; Razdan, N.; Saharan, K. Rhizospheric Microbiome: Bio-Based Emerging Strategies for Sustainable Agriculture Development and Future Perspectives. Microbiol. Res. 2022, 254, 126901. [Google Scholar] [CrossRef] [PubMed]
- Kumawat, K.C.; Nagpal, S.; Sharma, P. Potential of Plant Growth-Promoting Rhizobacteria-Plant Interactions in Mitigating Salt Stress for Sustainable Agriculture: A Review. Pedosphere 2022, 32, 223–245. [Google Scholar] [CrossRef]
- Barnawal, D.; Bharti, N.; Pandey, S.S.; Pandey, A.; Chanotiya, C.S.; Kalra, A. Plant Growth-Promoting Rhizobacteria Enhance Wheat Salt and Drought Stress Tolerance by Altering Endogenous Phytohormone Levels and TaCTR1/TaDREB2 Expression. Physiol. Plant. 2017, 161, 502–514. [Google Scholar] [CrossRef]
- Porras-Soriano, A.; Soriano-Martín, M.L.; Porras-Piedra, A.; Azcón, R. Arbuscular Mycorrhizal Fungi Increased Growth, Nutrient Uptake and Tolerance to Salinity in Olive Trees under Nursery Conditions. J. Plant Physiol. 2009, 166, 1350–1359. [Google Scholar] [CrossRef]
- Galla, G.; Barcaccia, G.; Ramina, A.; Collani, S.; Alagna, F.; Baldoni, L.; Cultrera, N.G.; Martinelli, F.; Sebastiani, L.; Tonutti, P. Computational Annotation of Genes Differentially Expressed along Olive Fruit Development. BMC Plant Biol. 2009, 9, 128. [Google Scholar] [CrossRef]
- Munoz-Merida, A.; Gonzalez-Plaza, J.J.; Canada, A.; Blanco, A.M.; Garcia-Lopez, M. d. C.; Rodriguez, J.M.; Pedrola, L.; Sicardo, M.D.; Hernandez, M.L.; De la Rosa, R.; et al. De Novo Assembly and Functional Annotation of the Olive (Olea europaea) Transcriptome. DNA Res. 2013, 20, 93–108. [Google Scholar] [CrossRef]
- Asins, M.J.; Bullones, A.; Raga, V.; Romero-Aranda, M.R.; Espinosa, J.; Triviño, J.C.; Bernet, G.P.; Traverso, J.A.; Carbonell, E.A.; Claros, M.G.; et al. Combining Genetic and Transcriptomic Approaches to Identify Transporter-Coding Genes as Likely Responsible for a Repeatable Salt Tolerance QTL in Citrus. Int. J. Mol. Sci. 2023, 24, 15759. [Google Scholar] [CrossRef]
- Fernandez-Pozo, N.; Zheng, Y.; Snyder, S.I.; Nicolas, P.; Shinozaki, Y.; Fei, Z.; Catala, C.; Giovannoni, J.J.; Rose, J.K.C.; Mueller, L.A. The Tomato Expression Atlas. Bioinforma. Oxf. Engl. 2017, 33, 2397–2398. [Google Scholar] [CrossRef]
- Bag, P.; Lihavainen, J.; Delhomme, N.; Riquelme, T.; Robinson, K.M.; Jansson, S. An Atlas of the Norway Spruce Needle Seasonal Transcriptome. Plant J. 2021, 108, 1815–1829. [Google Scholar] [CrossRef] [PubMed]
- Toufighi, K.; Brady, S.M.; Austin, R.; Ly, E.; Provart, N.J. The Botany Array Resource: E-Northerns, Expression Angling, and Promoter Analyses: The Botany Array Resource. Plant J. 2005, 43, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Pozo, N.; Haas, F.B.; Meyberg, R.; Ullrich, K.K.; Hiss, M.; Perroud, P.-F.; Hanke, S.; Kratz, V.; Powell, A.F.; Vesty, E.F.; et al. PEATmoss (Physcomitrella Expression Atlas Tool): A Unified Gene Expression Atlas for the Model Plant Physcomitrella patens. Plant J. Cell Mol. Biol. 2020, 102, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Ollé, A.; Bullones, A.; Hormaza, J.I.; Mueller, L.A.; Fernandez-Pozo, N. MangoBase: A Genomics Portal and Gene Expression Atlas for Mangifera indica. Plants 2023, 12, 1273. [Google Scholar] [CrossRef]
- Bullones, A.; Castro, A.J.; Lima-Cabello, E.; Alché, J.D.D.; Luque, F.; Claros, M.G.; Fernandez-Pozo, N. OliveAtlas: A Gene Expression Atlas Tool for Olea europaea. Plants 2023, 12, 1274. [Google Scholar] [CrossRef]
- Cruz, A.C.; Luvisi, A.; De Bellis, L.; Ampatzidis, Y. X-FIDO: An Effective Application for Detecting Olive Quick Decline Syndrome with Deep Learning and Data Fusion. Front. Plant Sci. 2017, 8, 1741. [Google Scholar] [CrossRef]
- Di Nisio, A.; Adamo, F.; Acciani, G.; Attivissimo, F. Fast Detection of Olive Trees Affected by Xylella fastidiosa from UAVs Using Multispectral Imaging. Sensors 2020, 20. [Google Scholar] [CrossRef]
- Sesli, M.; Yegenoğlu, E.D.; Altıntaa, V. Determination of Olive Cultivars by Deep Learning and ISSR Markers. J Env. Biol 2020, 41, 426–431. [Google Scholar] [CrossRef]
Figure 1.
Integrative scheme from the main causes of natural and anthropogenic soil salinisation (white boxes) to the salt adaptation and growth (green box) or non-adaptive response (red lines and boxes). Stresses are represented by orange boxes; the main signals are in pink boxes; adaptive processes are in blue boxes; and arrows depict known cause-effect relations. ABA: abscisic acid; ROS: reactive oxygen species; SOS: salt overly sensitive; PCD: programmed cell death.
Figure 1.
Integrative scheme from the main causes of natural and anthropogenic soil salinisation (white boxes) to the salt adaptation and growth (green box) or non-adaptive response (red lines and boxes). Stresses are represented by orange boxes; the main signals are in pink boxes; adaptive processes are in blue boxes; and arrows depict known cause-effect relations. ABA: abscisic acid; ROS: reactive oxygen species; SOS: salt overly sensitive; PCD: programmed cell death.
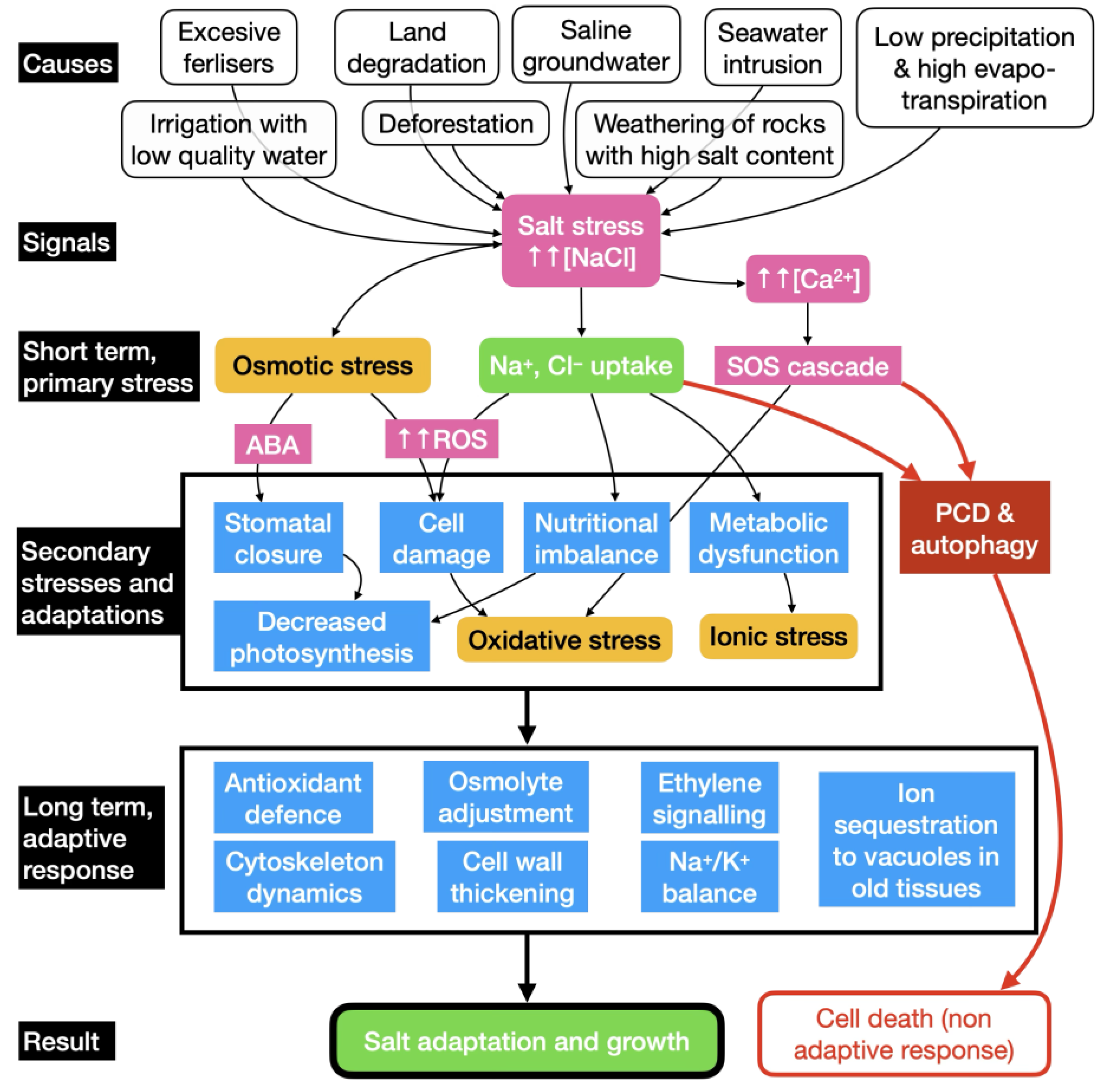

Table 1.
Unambiguous classification of 89 olive cultivars according to their salt tolerance.
| Cultivar | Reference | Country | Salt treatment | Treatment time |
|---|---|---|---|---|
| Salt-tolerant cultivars | ||||
| Abou-Satl | [161] | Syria | 12 dS/m NaCl | 90 days |
| Amigdalilolia | [162] | Iran | 12 dS/m NaCl | 90 days |
| Ayvalık | [163] | Turkey | 300 mM NaCl | 30 days |
| Barnea | [164] | Israel | 7.5 dS/m EC | 9 years |
| Cañivano | [165] | Spain | 100 mM NaCl | 49 days |
| Chemlali | [164] [166] |
Tunisia | brackish water | 8 months |
| 200 mM NaCl | 5 months | |||
| Dakal | [162] | Iran | 12 dS/m NaCl | 90 days |
| Empeltre | [167] | Spain | 10 dS/m Eciw | 4 years |
| Escarabajuelo | [165] | Spain | 100 mM NaCl | 49 days |
| Frantoio | [168] | Italy | 200 mM NaCl | 60 days |
| [169] | 100 molc/m3 SIW | ND | ||
| [157] | 200 mM NaCl | 84 days | ||
| [154] | 200 mM NaCl | 56 days | ||
| Hamed | [164] | Egypt | 6000 ppm NaCl, NaSO4, CaCl2 and MgSO4 | 2 years |
| Istarska bjelica | [170] | Croatia | 100 mM NaCl | 70 days |
| Jabaluna | [165] | Spain | 100 mM NaCl | 49 days |
| Jlot | [171] | Syria | 8000 ppm NaCl | ND |
| Kalamata | [164] | Greece | 200 mM NaCl | 5 months |
| Kalamon | [172] | Greece | 120 mM NaCl | 90 days |
| [173] | 120 mM NaCl | 90 days | ||
| Kerkiras | [164] | Greece | ND | ND |
| Kilis | [163] | Turkey | 300 mM NaCl | 30 days |
| Kilis Yağlık | [174] | Turkey | 150 mM NaCl | 4 months |
| Koroneiki I38 | [175] | Spain | 200 mM NaCl | 5 months |
| Kothreiki | [164] | Greece | 200 mM NaCl | 5 months |
| Lechín de Sevilla | [165] | Spain | 100 mM NaCl | 49 days |
| Lianolia | [164] | Greece | ND | ND |
| Megaritiki | [168] | Greece | 150 meq NaCl | 4 months |
| [135] | 200 mM NaCl | 5 months | ||
| Nevadillo | [165] | Spain | 100 mM NaCl | 49 days |
| Oblica | [170] | Croatia | 100 mM NaCl | 70 days |
| Ocal | [157] | Spain | 200 mM NaCl | 84 days |
| Pocama | [176] | Egypt | 6000 ppm NaCl, NaSO4, CaCl2 and MgSO4 | 2 years |
| Royal de Cazorla | [141] | Spain | 100 mM and 200 mM NaCl | 8 months |
| [159] | 100 mM and 200 mM NaCl | 60 days | ||
| [177] | 200 mM NaCl | 8 months | ||
| Verdale | [176] | France | 6000 ppm NaCl, NaSO4, CaCl2 and MgSO4 | 2 years |
| Intermediate tolerance to salt | ||||
| Adramitini | [164] | Greece | ND | ND |
| Aggezi | [164] | Egypt | 6000 ppm NaCl, NaSO4, CaCl2 and MgSO4 | 2 years |
| Alameño | [165] | Spain | 100 mM NaCl | 49 days |
| Arbosana I43 | [166] | Spain | 200 mM NaCl | 5 months |
| Blanqueta | [165] | Spain | 100 mM NaCl | 49 days |
| Cañivano Negro | [165] | Spain | 100 mM NaCl | 49 days |
| Carolea | [168] | Italy | 14.97 mS/cm | 11 months |
| Casta Cabra | [157] | Spain | 200 mM NaCl | 84 days |
| Changlot Real | [165] | Spain | 100 mM NaCl | 49 days |
| Chorruo | [165] | Spain | 100 mM NaCl | 49 days |
| Coratina | [168] | Italy | 200 mM NaCl | 60 days |
| Cornicabra | [157] | Spain | 200 mM NaCl | 84 days |
| Dezful | [162] | Iran | 12 dS/m NaCl | 90 days |
| Gordal Sevillana | [165] | Spain | 100 mM NaCl | 49 days |
| Hojiblanca | [165] | Spain | 100 mM NaCl | 49 days |
| Khaisi | [171] | Syria | 8000 ppm NaCl | ND |
| Lechín de Granada | [165] | Spain | 100 mM NaCl | 49 days |
| Manzanilla de Sevilla | [165] | Spain | 100 mM NaCl | 49 days |
| Manzanillo | [168] | Spain | 60 mM NaCl | 115 days |
| [178] | 4000 mg/L MgSO4, CaSO4, NaCl, MgCl2 and CaCO3 | 9 months | ||
| [176] | 6000 ppm NaCl, NaSO4, CaCl2 and MgSO4 | 2 years | ||
| Maraiolo | [164] | Italy | ND | ND |
| Maurino | [164] | Italy | 200 mM NaCl | 60 days |
| Moraiolo | [168] | Italy | 200 mM NaCl | 60 days |
| Mostazal | [164] | Peru | 6000 ppm NaCl, NaSO4, CaCl2 and MgSO4 | 2 years |
| Nabali Muhassan | [164] | Jordan | 100 molc/m3 SIW | ND |
| Nizip Yağlik | [174] | Turkey | 150 mM NaCl | 4 months |
| Oblonga | [165] | Italy | 100 mM NaCl | 49 days |
| Redondil | [165] | Spain | 100 mM NaCl | 49 days |
| Toffahi | [164] | Egypt | 6000 ppm NaCl, NaSO4, CaCl2 and MgSO4 | 2 years |
| Tokhm-e-Kabki | [162] | Iran | 12 dS/m NaCl | 90 days |
| Valanolia | [164] | Greece | ND | ND |
| Verdial de Vélez-Málaga | [165] | Spain | 100 mM NaCl | 49 days |
| Wardan | [164] | Egypt | 6000 ppm NaCl, NaSO4, CaCl2 and MgSO4 | 2 years |
| Zorzariega | [165] | Spain | 100 mM NaCl | 49 days |
| Salt-sensitive cultivars | ||||
| Aggezi Shami | [178] | Egypt | 4000 mg/L MgSO4, CaSO4, NaCl, MgCl2 and CaCO3 | 275 days |
| Aguromanaki | [164] | Greece | ND | ND |
| Arbequina I18 | [166] | Spain | 200 mM NaCl | 5 months |
| Arbosana | [175] | Spain | 200 mM NaCl | 5 months |
| Bezeri | [171] | Syria | 8000 ppm NaCl | ND |
| Bouteillan | [164] | France | 6000 ppm NaCl, NaSO4, CaCl2 and MgSO4 | 2 years |
| Buža | [170] | Croatia | 100 mM NaCl | 70 days |
| Chondrolia Chalkidikis | [172] | Greece | 120 mM NaCl | 90 days |
| [179] | 150 meq NaCl | 4 months | ||
| [172] | 120 mM NaCl | 90 days | ||
| [180] | 75 mM NaCl | 45 days | ||
| [181] | 75 mM and 150 mM NaCl | 45 days | ||
| Cobrançosa | [165] | Portugal | 100 mM NaCl | 49 days |
| Conservalia | [162] | Iran | 12 dS/m NaCl | 90 days |
| Drobnica | [170] | Croatia | 100 mM NaCl | 70 days |
| Fadaq86 | [177] | Iran | 200 mM NaCl | 8 months |
| Galego | [165] | Spain | 100 mM NaCl | 49 days |
| Khederi | [171] | Syria | 8000 ppm NaCl | ND |
| Lastovka | [170] | Croatia | 100 mM NaCl | 70 days |
| Leccino | [168] | Italy | 200 mM NaCl | 60 days |
| [170] | 100 mM NaCl | 70 days | ||
| [154] | 200 mM NaCl | 56 days | ||
| Mastoidis | [134] | Greece | 200 mM NaCl | 5 months |
| Meski | [165] | Tunisia | 100 mM NaCl | 49 days |
| Mission | [176] | US | 6000 ppm NaCl, NaSO4, CaCl2 and MgSO4 | 2 years |
| Nabal | [164] | Egypt | 6000 ppm NaCl, NaSO4, CaCl2 and MgSO4 | 2 years |
| Nocellara del Belice | [182] | Italy | 14.97 mS/cm | 11 months |
| Pajarero | [168] | Spain | 100 mM NaCl | 49 days |
| Rosciola | [176] | Italy | 6000 ppm NaCl, NaSO4, CaCl2 and MgSO4 | 2 years |
| Shiraz | [162] | Iran | 12 dS/m NaCl | 90 days |
| Sourani | [171] | Syria | 8000 ppm NaCl | ND |
| Throubolia | [164] | Greece | ND | ND |
ND: no data available.
Table 2.
Inconsistent classification of 8 olive cultivars according to their salt tolerance.
| Cultivar | Tolerant | Intermediate | Sensitive | Country | Salt treatment | Treatment time |
|---|---|---|---|---|---|---|
| Amphissis | [179] | Greece | 150 meq NaCl | 4 months | ||
| [134] | 200 mM NaCl | 5 months | ||||
| Arbequina | [168] | Spain | 100 mM NaCl | 49 days | ||
| [167] | 10 dS/m Eciw | 4 years | ||||
| [175] | 200 mM NaCl | 5 months | ||||
| [159] | 100 mM and 200 mM NaCl | 60 days | ||||
| [161] | 12 dS/m NaCl | 90 days | ||||
| Chétoui | [166] | Tunisia | 200 mM NaCl | 5 months | ||
| [165] | 100 mM NaCl | 49 days | ||||
| Gemlik | [163] | Turkey | 300 mM NaCl | 30 days | ||
| [174] | 150 mM NaCl | 4 months | ||||
| Koroneiki | [176] | Greece | 6000 ppm NaCl, NaSO4, CaCl2 and MgSO4 | 2 years | ||
| [179] | 150 meq NaCl | 4 months | ||||
| [159] | 100 mM and 200 mM NaCl | 60 days | ||||
| [141] | 100 mM and 200 mM NaCl | 8 months | ||||
| [177] | 200 mM NaCl | 8 months | ||||
| [166] | 200 mM NaCl | 5 months | ||||
| [134] | 200 mM NaCl | 5 months | ||||
| Picual | [168] | Spain | 100 mM NaCl | 49 days | ||
| [178] | 4000 mg/L MgSO4, CaSO4, NaCl, MgCl2 and CaCO3 | 9 months | ||||
| [176] | 6000 ppm NaCl, NaSO4, CaCl2 and MgSO4 | 2 years | ||||
| [159] | 100 mM and 200 mM NaCl | 60 days | ||||
| [177] | 200 mM NaCl | 8 months | ||||
| [157] | 200 mM NaCl | 84 days | ||||
| Picudo | [157] | Spain | 200 mM NaCl | 84 days | ||
| [165] | 100 mM NaCl | 49 days | ||||
| Zard | [161] | Iran | 12 dS/m NaCl | 90 days | ||
| [162] | 12 dS/m NaCl | 90 days |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



