
Submitted:
01 December 2024
Posted:
02 December 2024
You are already at the latest version
Abstract
Integrating metagenomics and synthetic biology has enormous potential to improve biotechnological applications and deepen our understanding of microbial ecosystems. Synthetic biology offers strong tools for creating new microorganisms or altering existing ones to carry out particular tasks because of its capacity to design and build biological systems. Instead of requiring cultivation, metagenomics allows the investigation of microbial diversity and function directly from ambient samples. These disciplines' merger enables scientists to modify genetic material, uncover secret biochemical processes, and understand intricate microbial communities. Synthetic biologists can find new genes, enzymes, and metabolic pathways by using metagenomic data. These can then be synthesized and turned into customised microbes for a range of uses, such as environmental cleanup, agriculture, and medicine. The development of microorganisms that work symbiotically with human, plant, or animal hosts is made easier by this interaction, which also offers up new avenues for microbiome study. Despite its promise, there are still obstacles in closing the gaps between applications of synthetic biology, microbial function, and metagenomic data analysis. The practical application of this convergence will be accelerated by future developments in computational tools, DNA synthesis, and system biology, which will solve microbial mysteries and spur breakthroughs in a variety of scientific and industrial fields.
Keywords:
Metagenomics
; Microbiome
; Microbial Ecosystems
; Synthetic Biology
1. Introduction
Microbes are some of the oldest and most diverse life forms on Earth. They include bacteria, viruses, fungi, protists, and archaea, found in every possible habitat, from the highest mountain peaks to the deepest oceans, playing a crucial role in maintaining balance in ecosystems. Microbial Biodiversity in simple words is defined as several species that include the genetic composition of microorganisms found in soil or other habitats performing major functions in the ecosystem. Studying microbial biodiversity can help researchers understand and manipulate the working of ecosystems, ecosystems’ tolerance to major disturbances, shaping ecosystems, and providing nutrients [1]. These microbes play multifunctional roles which include soil microbes decomposing litter, detoxification, enhancing nutrient cycling, and establishing critical relationships with higher organisms and among themselves [2]. Aquatic microbes found in lakes and ponds are the main habitat for their growth playing an important role in remineralization and restoring nutrients, they produce industrially important enzymes such as proteases that are used in food industries, and waste treatment [3]. Additionally, microbes in the human gut and skin play important roles in digestion, immune system, vitamin production and overall health [4]. Despite its ecological and biological importance, most of the microbial world remains unexplored due to its enormous complexity and the difficulty of cultivating many microbes in laboratory conditions.
1.1. Overview of Synthetic Biology (SynBio) and Metagenomics
The human microbiota plays various physiological functions such as drug metabolism, digestion, and nutrient degradation. It is important to maintain their balance as any alteration in gut microbes can lead to inflammatory diseases, and metabolic diseases [5]. In order to understand and control microbial communities various system biology approaches were implicated. SynBio is an interdisciplinary field which includes principles of engineering, molecular biology, and genetics to create tools and computational modelling that has the potential to construct new biological parts, that permit modify organisms to perform desired tasks [5,6,7] conducted an experiment using genetic circuits that reprogrammed rat cell’s identity to treat diabetes converting normal cells to insulin secreting cells. Such biological system helps to develop novel therapeutic and diagnostic ideas and improve disease modelling studies. Metagenomic, a technique used for collecting genomes of microbial communities, that can reveal the functional and compositional characteristics of microbial populations [5,8]. This technique is also called 16s rRNA sequencing where at a specific region of the 16s rRNA gene, DNA is extracted further amplified and sequenced and generated data is compared to the reference database [9]. Bioinformatic tools such as MicrobiomeAnalyst are used for studying tasks such as profiling, and visualisation of metagenomic networks [10]. Metagenomics has played a huge role in identifying food microbiomes. A recent study was conducted in which 1,950 food microbiomes were metagenomically sequenced [11]. The sequencing data obtained from metagenomic is difficult to analyse due to its complex structure and various software tools such as MicrobiomeAnalyst, Newbler (Roche), MIRA 4,37 or AMOS, as well as the recent MetaAMOS [12]. SynBio and metagenomics when combined, overcome the barrier of metabolic burden, that is, recombinant microorganisms are engineered for producing desired products (e.g., insulin) but their productivity is limited and decreases as commercialization demand grows [13]. Some of the major landmarks in SynBio are depicted in the Figure 1.
The purpose and scope are to explore the transformative potential of SynBio and metagenomics in unravelling microbial mysteries. A primary objective is to show how SynBio, with its ability to engineer and redesign biological systems, and metagenomics, which offers insights into microbial genetic material, can be combined to unlock new insights into microbial diversity and its ecological significance. The integration of these issues will allow us to address pressing global issues such as environmental sustainability, health care, and industrial innovation. The approach also focuses on modularity and innovation, connecting microbial communities to ecosystems to manage antimicrobial resistance [14,15,16,17]. The ultimate purpose is to decode the microbial ecosystems by enabling the vast microbial diversity from various sources, it also enhances the drug discovery and production of various products by exploring the untapped microbial pathways and then uncovering the functional potential of microbes and their interactions in ecosystems [18,19,20,21,22]. SynBio involves redesigning and creating microorganisms with desired functions based on metagenomic data [23]. The study of metagenomics reveals untapped genetic material from a wide range of microbial communities, which facilitates the discovery of new enzymes, metabolic pathways, and genes of interest for industrial and medical applications. By developing antimicrobials and probiotics, these fields contribute to improving healthcare and the environment: bioremediation and carbon capture can be achieved by engineering microorganisms [24,25,26].
2. Tools and Techniques in Synthetic Biology
SynBio holds unique genetic tools and components that can be used to design metabolic pathways or develop an artificial process. They rely upon the most expressed genetic components at higher concentrations. Any variation in the synthetic and native components can lead to reduced performance in the engineered system [27]. Based on the complexity of the system, the chances of failure are bigger [28]. Precise tool kits and a regulatory system of the cells were to be utilized to reach their desired regulation. Traditional approaches, like chemical agents to induce mutation or transposons, can be time-consuming and need increased screening time. DNA viral vectors (Double-stranded) and RNA interference in gene knockdown approaches pose pitfalls [29]. Cellular machinery involved in gene silencing can be disrupted by dsRNA (RNAi technology) when it is added exogenously. The most common form of genetic engineering technique involves DNA-cutting using enzymes to give double strands [30]. Endonucleases-based tools were helpful in target gene inactivation studies [31].
Next Generation Sequencing (NGS) can generate 15 petabases of sequencing data in a year [32]. Advancements like CRISPR/Cas system make deletion, insertion of genes, and transcriptional control much easier [33]. Modular components were interconnected to the assembled fragments to develop biological circuits and control cellular behaviour for metabolic pathways [28]. When a repair template is introduced into the system, they utilize the host double-stranded DNA break homologous repair system as an advantage [34]. Genome editing tools have greater application in controlling pest populations of invasive species (cane toads, malaria, carp, etc.) [35]. Gene drive is introduced into the genetic element (meganucleases), which can distort the sex ratio, fertility, and pathogenicity among the interbreeding population [36]. Well-known genome editing tools are meganucleases (MNs) [37], zinc finger nucleases (ZFNs) [38], transcription activator-like effector nucleases (TALENs) [39], clustered regularly interspaced short palindromic repeat (CRISPR)-associated nuclease Cas9 [40], targetrons [41]. The nonhomologous end-joining repair system (NHEJ) causes frameshift mutation, resulting in gene disruption or knockout [42]. Lesions are generated when a homologous sequence containing a double-stranded DNA template corresponds to a flanking sequence at the break site in a cell and is repaired with the help of HDR machinery [43]. The cleavage at the non-targeted sites creates undesirable mutations, requiring extensive identification screening (Figure 2).
2.1. Gene Editing Mechanism
Breaks in the double-stranded DNA are repaired by two major pathways: Homology-directed repair (HDR) or Non-homology end joining (NHEJ). HDR works template-dependent at the broken end by making homologous sequences [44]. Gene targeting efficiency by homology recombination magnitude can be increased by DSB introduction at the target site [45]. The NHEJ repair mechanism doesn’t use a DSBs repair template at the target cleave ends [46]. The reading frame of a gene can be shifted by using site-specific DSBs with the assistance of NHEJ stimulation [47].
2.2. Gene Knockouts and Gene Deletion
Gene knockout is the most straightforward form of gene editing, which focuses on creating insertion and deletion (indel) at the target site created by NHEJ error. There are two types of NHEJ. Classical NHEJ acts directly on unprocessed DNA, whereas the alternative NHEJ, also called microhomology-mediated end joining (MMEJ), requires end-resection and annealing of single-stranded regions that are shorter in size. Both of these pathways repair DNA with a higher rate of mutagenesis, forming indels at the break site; the indels formed cause frameshift [48]. In Duchenne muscular dystrophy (DMD), disease frameshift is seen due to gene deletion resulting in loss of protein function; indels targeted by NHEJ-induction assist in correcting the reading frame of the gene by restoring the correct one [49]. Knocking out endogenous genes helps in new treatments where the particular gene function can be stopped permanently. They can be used in treating diseases like Huntington’s disease, where an allele gene huntingtin (HTT) tends to expand repeatedly, producing mutant HTT protein (toxic). Gene editing by NHEJ could provide clinical benefits in other diseases like HIV (CCR5 knockouts) [50]. More significant segments of DNA can be deleted by flanking the sequence with the help of two double-stranded base pairs (DSBs). When two targeted breaks are introduced simultaneously, megabases of genomic deletions are created [51]. This can be beneficial in treating hemoglobinopathies where the BCL11A (erythroid-specific) region is deleted [52].
2.3. Gene Correction and Insertion
Exogenous donor template stimulates HDR gene editing with the help of targeted double strands, which tend to be active at the S and G2 phases. During the process, the sister chromatid is used as a template for DNA repair [48]. Delivering targeted nucleases and DNA homologous containing vectors at the break site induces HDR-based gene editing with higher editing efficiency [53]. The mutation responsible for a specific disease can be repaired using donor template sequences on the endogenous locus [54]. The most commonly used donor templates are plasmids; single-stranded oligonucleotides (ssODNs) with 80 base pairs can also serve as a template for HDR [55]. Where transfection is impossible, viral vectors like integrase-deficient lentivirus or adeno-associated virus (AAV) were used as donors [56].
Viral vectors have long been used in traditional gene therapy to introduce the gene of choice (exogenous genes) into the desired genome. Controlling the site of integration of these viruses poses a challenge, so murine retroviral vectors were preferred [57]. Flanking the desired insert present in the donor with the help of the homology arm and the nuclease enzyme site-specific sequence induces the DSB-induced HDR DNA insertion at a particular site [58]. The insertion of therapeutic genes at the target site, a “safe harbor,” increases the risk of insertion mutagenesis, which expresses ubiquitous genes. To control this, a wild type of the same disease-causing gene is inserted at the endogenous locus, where the promoter takes control of the process [59].
2.4. Genome Editing Tools
The Genome editing tools can be subdivided into two groups: (I) MNs (meganucleases), ZFNs (zinc finger nucleases), and TALE (Transcription activator-like effector nucleases). They bind to their specific DNA sequence with the help of protein-DNA interaction [37]. The second group comprises two subgroups (I) RNA-guided systems CRISPR/Cas9 and targetrons [40]. The second subgroup contains DNA-based guided systems peptide nucleic acids (PNAs), triplex-forming oligonucleotides (TFOs), and structure-guided endonucleases (SGNs) [60]. Gene editing initiated by Double-stranded breaks was repaired endogenously. Precise gene editing was implemented by introducing targeted DSBs.
2.4.1. Zinc Finger Nucleases
Zinc finger nucleases (ZFNs) are the most abundant transcription factors, and it is artificially generated by combining DNA binding and recognition domain (Cys2His2), a common DNA-binding domain encoded in the human genome to non-specific cleavage domain (IIS) from FoKI restriction enzyme [61]. The zinc finger domain forms the ββα structure in a zinc atom’s presence. Where the α helical portion of each finger makes contact with 4bp in the DNA [62]. ZFNs bind to an extended target sequence where the three-finger protein binds to the nine-bp site. Structure-based design was used to engineer two ZFNs that efficiently cleave DNA when paired as heterodimer, which can be used as a potential gene-modifying reagent [63]. The dimerization by ZFN was studied using 3D protein modeling and energy calculations with computer assistance, showing a potential dimer within the FoKI interface [64]. Newly designed ZFNs were less toxic (genotoxic) in cellular-based studies. When dimerization energy is lowered, dimeric FoKI formation is prevented [64]. Zif268 has an attractive framework that can be used to design new DNA-binding motifs [65]. ZFNs have been used to inactivate specific core receptors (HIV CCR5 coreceptors) to avoid viral entry into cells [66]. Due to FoKI’s non-specific nature and ZFN modular architecture, there is a high chance of breakage at the ectopic site [67]. Artificial zinc finger protein (AZP) and Staphylococcal nuclease (SNase) give a hybrid AZP-SNase that can be used for anti-viral therapies. This artificial nuclease can target a particular site in human papillomavirus type 18 (HPV-18), which stops viral replication [68]. One drawback of this hybrid is that it cleaves both single and double-stranded host DNA and RNA by switching the moiety in the AZP-SNase to single-chain FoKI dimer (scFoKI) successfully cleaves the viral DNA. It can inactivate human DNA viruses with fewer side effects [69]. ZFN technology was made possible due to FoKI restriction endonuclease, which functions independently (DNA-binding and cleavage domains) [70]. Chimeric nuclease with the novel binding site can be generated by replacing the FokI DNA-binding domain with a zinc finger domain. DSBs formed by ZFN induction can be modified by either NHEJ or HDR [71], where they find their application in somatic cells of humans and stem cells [72].
2.4.2. Transcription Activator-like Effector Protein (TALENs)
These are artificial endonucleases made by the fusion of DNA-binding domain obtained from transcription activator-like effector protein (TALE) with FoKI endonuclease-like cleavage domain [73], primarily isolated from the plant pathogens Xanthomonas sp. [39].TALE repeats are highly conserved, binding to a single base pair guided by the hypervariable residues. The binding of TALEs to DNA forms a helical structure for each repeat connected by a loop that presents the hypervariable region to the major groove where the protein forms a superhelical structure around the DNA [74]. Each corresponding nucleotide is recognized by a TALE repeat with its two variable residues, where the repeats represent the sequence in the binding site. TALENs have higher genome editing than ZFNs [75]. By joining proper repeated units, user-defined TALENs can be redesigned. TALENs are dimeric, where the specificity of the enzymes is higher when compared to monomeric enzymes [39]. Tev-TALE nucleases (Tev-mTALENs) were an alternative to FokI-based architecture, where the sequence-specific I-TevI HEase is joined with TALEs. Thus, the TeVI domain has predetermined recognition sites with limitations over the genomic targets [76].
TALE assembly can recognize 12-20bps of DNA and higher genome editing can be achieved by increasing the base pairs [77]. Each building domain in TALE is more than 34 residues in length. The amino acid residue at the 12 and 13 in each repeat gets in contact with the DNA and is called repeat variable residues (RVDs) [39]. RVDs used in synthetic tale arrays are NI for adenine, HD for cytosine, NG for thymine, and guanine, NN, or HN [39]. One method used to construct TALE DNA-binding domains is the Golden Gate Way assembly approach [78]—other methods include high throughput TALE assembly, Flash Assembly, iterative capped assembly, and Ligation independent cloning. TALENs have two unique advantages in genome editing when compared with ZFNs. No selection is needed to engineer the TALEN array. Also, assembling functional nuclease requires less time and no prior experience. TALENs have increased affinity towards targeted DNA with less toxicity. Due to their larger size and repetitive structure, transportation into cells is done with the help of lentivirus or single adeno-associated virus (AAV) [79].To make the delivery easy, TALENs can be delivered as mRNA (Mock et al., 2015) and protein [80].
2.4.3. CRISPR-Cas9
CRISPR-Cas 9 protein has a significant application ranging from genetics to bioimaging systems. They function as an adaptive immunity role in bacteria, which finds its way inside genome editing [81]. The type II CRISPR system in bacteria helps protect against invading viruses and plasmids with the help of Cas proteins through an RNA-guided DNA cleavage mechanism [82]. CRISPR RNA was created by transcribing the locus containing foreign DNA and annealing it to transactivate crRNA (tracrRNA). Sequence-specific degradation of pathogenic DNA was carried out by the Cas9 protein [83]. Target recognition by Cas9 protein is made easy by integrating a seed sequence that must be in the crRNA and a protospacer-adjacent motif (PAM) upstream of the crRNA binding site [83]. This finds its way inside the world of genome engineering, which contains Cas9 nuclease guide RNA gRNA (crRNA (CRISPR RNA) and tracrRNA (trans-activating CRISPR RNA) elements). Both are expressed on a single construct called single-guided RNA (sgRNA) [83]. CRISPR-Cas9 emerged as a user-friendly and more flexible genome editing platform. One of the significant challenges is that the PAM motif is to be located in the gRNA target during the downstream process. The PAM sequence for the Streptococcus pyogenes Cas9 is 5’ -NGG-3’, where 5’ -NAG-3’ can be tolerated [40]. The gRNA and the complementary strand form a heteroduplex structure between two nuclease domains, RuvC and HNH, inside the Cas9 protein, where the PAM recognition is by the arginine-rich motif in Cas9 [84]. Cas9 and gRNA can be delivered using plasmids, and ECRISP is used to select the target site and construct gRNA [85]. Reports suggest Cas9 can cause off-target mutations [86]. Using paired Cas9 nickases, the specificity can be improved considerably by inducing nick in DSB formation or increasing the sensitivity for mismatches on genomic target sites by truncated gRNA [87]. They use configured Cas9 variants such as Cas9 architecture split induced by rapamycin or Cas9 variant with responsive intein domain [88]. Methods like nucleofection and transient transfection over Cas9 complex reduces the off-target effects in gene editing in human T-cell [89], Stem cells form embryo [90] gonads of Caenorhabditis elegans [91], mouse [92]. The secondary structure of RNA can be protected and made stable by using specific chemical modifications, which further enhance the Cas9 ribonucleoprotein activity [93]. Protein engineering can increase Cas9 specificity and change PAM requirements.
2.4.4. Homing Endonucleases
Homing Endonucleases (HEs) cut DNA encoded by the Homing Endonuclease gene (HEGs). These mobile genetic elements also induce the movement of genetic elements where they are seen. They are sequence-specific endonucleases that recognize base pairs that are >14 bp compared to restriction enzymes from bacteria (REs). HEs tolerate sequence degeneracy. It may not interfere with the cleavage process but can reduce efficiency. It is also the final member of the targeted nuclease family. There is an endonuclease family named after the conserved motif region present within these enzymes LAGLIDADG (LHEs). It has the most comprehensive host range from plants, protists, and branching metazoans. The domains (binding and cleavage) are not modular, as seen in ZFNs and TALENs. HEGs perform double-strand break (DSB) or single-stranded nick inside the alleles (cognate), which do not have coding sequences for endonuclease enzyme; such process is called homing.
The most frequently encountered group I intron-containing Open reading frames (ORFs) is LHEs. At certain times, they can be seen in group II introns [94]. LHEs make staggered cuts at the site of cleavage, leaving 3’ overhangs over four nucleotides. Dodecapeptide motifs (one or two) LAGLIDADG helps in protein encoding, where one of the motifs is of ancestral version, which after duplication resulted in a double motif [95].When single motif LHEs act as a homodimer, whereas double motif LHEs are monomers. I-SceI comes under the family of HEs and was the first encoded intron. The intron in the mtDNA of Sacharomyces cerevisiae in the rnl gene is mobile and referred to as the omega (u) locus. The nuclear genome of Saccharomyces species contains two non-intron encoded LHE family members: homothallism (HO) and VMA1-derived endonuclease (VDE). This endonuclease, also known as PI-SceI, was the first structure elucidated intein [96]. Switching between mating types can be achieved through HO, which acts as a catalyst in budding yeast [97]. GIY-YIG endonuclease introns have been reported within the mitochondria, chloroplast, and bacteriophage genome. The GIY-YIG motif is associated with DNA-binding enzymes that require endonuclease activity, like UvrC in archaea and UvrABC in bacteria (excision repair complex) [98]. The group one intron-containing ORF encodes specific domains from proteins such as H-N-H proteins or H-N-H domains, where the H-N-H motif bearing endonuclease domain can be seen associated with inteins, Res of bacteria, transposases, DNA acting factors [99]. Group II introns have varied functional domains with various activities that encode complex proteins. The Intron insertion site is cleaved by Endonucleases [100].
2.5. Synthetic DNA
DNA holds the basic information of life. Recently, many efforts have been made to generate viable microbes using synthetic genome technology [101]. Synthetic DNA can be used as a probe in living organisms, which can be used to amplify inaccessible sequences. Using xeno nucleic acids (non-natural nucleotides) (XNAs), new DNA architectures with functionalities can be created [102]. Single-stranded DNA acts as a donor in genome engineering in synthesizing homo-polynucleotides [103]. Next-generation sequencing (NGS) has dramatically improved the editing of DNA, which has revolutionized population genomics analysis. Technologies like DNA de novo synthesis and gene editing by CRISPR have the most significant advantage over editing and DNA writing [104]. This can be applied in fields like vaccines, data storage, drug delivery, and designing microorganisms with resistant capabilities. DNA synthesis by chemical method has produced DNA chains of <200-nucleotide (oligonucleotides). Technologies that plasmid template approaches are rolling circular amplification [105], Polymerase chain assembly or Gibson assembly [106], template-independent enzymatic oligonucleotide synthesis (TiEOS) in which terminal deoxynucleotidyl transferase (TdT) as a DNA synthesis tool [107]. One of the significant challenges in DNA synthesis is generating >300 nucleotide DNA sequence that entirely depends on the efficiency of the elongation cycle. The elongation cycle (99%) yield of oligonucleotide has 120 nucleotides. During the commercial preparation of the>1 kb sequence,>300 bp polynucleotides were used as the building blocks (Crosby et al., 2019).
2.5.1. Synthetic DNA Using Phosphoramidites
This method synthesizes the DNA from 5ʹ-protected dimethoxy trityl (DMT) nucleotide phosphoramidites [108]. In 1980, applied biosystems designed the first automatic DNA synthesizer (synthetic oligonucleotides) [109]. Individual sequences of oligonucleotides were chemically assembled onto solid support (silicon). By applying varied microarray formats, parallel in situ synthesis of oligonucleotides has been achieved. The sequence assembled in one site can be controlled independently, thus giving an insight into the synthesis of various sequences in a site-specific manner [110]. Compared to the single sequence synthesis approach, the multisequence approach on silicon chip reduces the reaction volume and increases the DNA output [111]. Phosphoramidite synthesis has poor bench stability and requires more significant organic solvents. Depurination occurs when the 5ʹ-DMT protecting group (PG) is removed during acid treatment, leading to the loss of purine bases, which makes the synthesized DNA susceptible to hydrolysis (Ravikumar et al., 1996). Asymmetric PCR can amplify DNA, which is thousands of nucleotides in length [112].
2.5.2. Enzymatic Oligonucleotide Synthesis
They work on the principles of solid phase synthesis, where the DNA of choice is synthesized on solid phase support with the help of DNA polymerase nucleoside 5ʹ-triphosphates (NTPs) [113]. DNA polymerase works on a base-pairing mechanism through the template strand and assists in selecting incoming nucleotides. DNA polymerase is unable to go de novo. The enzyme that can aid in the elongation of the polynucleotide sequence in the absence of a template strand is required, which has been identified as TdT and has been commercially incorporated into the TiEOS Methods [107].
2.5.3. Template Independent Synthesis
Elongation of oligonucleotides occurs promiscuously from 5’-3’ by taking in the nucleotides and raising the concomitant formation of various sequences [114]. One of the best ways to control nucleotide incorporation is through reversible termination. Where modified NTPs are used as the synthesis terminator at the 3’ position. Chemically synthesized single-stranded initiator DNA 19 was preloaded onto resin beads, followed by ligation between the 3ʹ-protected NTPs 20 to the desired sequence 23. Before each elongation cycle, deblocking of 3ʹ-PG was performed along with the washing step to remove any side products and other reagents [115]. A particular deoxyuridine cleavage site is found in initiator DNA 19 at the 3’ end, which is liable enzymatically. Once the synthesis is over, Uracil DNA glycosylase cleaves the site to release the assembled sequence of the resin [116].
There are some limitations reported on TdT. It prefers some nucleotides over others, which could increase the chances of sequence-specific errors [115]. TdT works on single-stranded DNA rather than double-stranded DNA. This is due to the steric shield attributed to the lariat loop in the enzyme that prohibits double-stranded DNA access [117] . The efficiency of the synthesized strand can be reduced if the strand under construction forms a secondary structure (Hoose et al., 2023). TdT-catalysed phosphoryl transfer requires divalent cations to synthesize new strands from NTPs. Unlike other DNA polymerases that require Mg2+ to initiate the synthesis of DNA molecules, TdT can utilize numerous divalent metal cations Co 2+, Mn 2+, Zn 2+, and Mg 2+. Using Mg2+ incorporates deoxyguanosine triphosphate and deoxyadenosine triphosphate, Co2+ deoxycytidine triphosphate, and deoxythymidine triphosphate (Hoose et al., 2023). Placing appropriate 3ʹ-PGs onto NTPs avoids the random incorporation of NTPs onto the extending DNA chain; this technology can be seen in DNA Script and Nuclera Nucleics.
3. Advancement in Metagenomics
3.1. Metagenomics- Exploring the Microbial World
Years back, our awareness of the microbial world was limited to strains that were capable of culturing under laboratory conditions. However, with the advancement in sequencing techniques and with the discovery of metagenomics, genetic materials from vast environmental and clinical niches could be analysed without being isolated or grown under laboratory conditions. This technique has transformed our understanding of microbial communities by providing unparalleled insights into their complexity and functionality[24].Metagenomics enables scientists to visualise microbial communities by separating genetic material (RNA, DNA) straight from an environmental/clinical sample, eliminating the effort of growing them. The isolated genetic materials are a collection of genomic samples that illustrate the microbial diversity of an ecosystem. For instance, the metagenomic data processed from human samples, which claim to include billions of microorganisms, was a significant achievement for the metagenomic method serving the scientific world. The basic steps to metagenomics have been listed below (Figure 3).
3.1.1. Extraction of Metagenomic DNA
DNA from a sample can be extracted directly from a location of interest, providing 1-50 kb of low-quality DNA, or indirectly, by separating microbes and isolating the DNA, yielding greater DNA quality and more precise outcomes. The indirect approach can recover DNA ranging from 20 to 500 kb, although it is time-consuming and elaborate. The indirect method usually results in the loss of some DNA of microbial species[118]
3.1.2. Metagenome Libraries
The method for creating a microorganism DNA library varies depending on the goal of the study. Specific work goals are needed to know what vector to use and what portion of the gene or gene cluster needs to be cloned to make a library. Plasmid, cosmid, fosmid, bacterial artificial chromosome (BAC), yeast artificial chromosome (YAC), and phage are examples of frequent vectors used for making metagenomic libraries[119].
3.1.3. Screening of Metagenome Library
Function-driven screening, sequence-driven screening, and substrate-induced gene expression screening (SIGEX) are the three methods used today for screening environmental metagenomic libraries. Fluorescence in situ hybridisation and DNA stable-isotope probing (DNA-SIP) are further methods. Further, some new microarray, stable isotope labelling, and fluorescence in situ hybridisation-based approaches have also been applied to sequence screening[24]
3.1.4. Metagenomic Sequencing and Analysis
Metagenomic DNA can be sequenced from DNA libraries using next-generation sequencing or third-generation sequencing and obtained high-quality reads are compiled into a complete genome sequence by software. The data is next analysed for quality, followed by functional and taxonomic profiling. The amplified sequences are library-prepared, and the relevant statistical and bioinformatic analysis is conducted [120].
3.2. Advances in DNA Sequencing Technologies
Over the years, sequencing 16S amplicon has been a legacy in identifying and classifying bacterial species. Now, with the recent advances in the field of sequencing techniques, it has become much more feasible to study the mysteries behind different microbial communities. The availability of public depositories of novel metagenome-assembled genomes and protein sequences has accelerated the journey of achieving significant milestones in the field of medicine. The advancement in the field of sequencing techniques is marked by the discoveries of third-generation sequencing and the latest NGS-based sequencing
3.2.1. Illumina Sequencing
The majority of metagenomic research uses second-generation sequencing, which produces billions of short sequences in a single run. The most popular platforms for this type of sequencing include Illumina. Illumina sequencing is the most commonly used sequencing technique that can run up to a read length of 150-30 bp with errors ranging from 0.1% to a maximum of 1%. For instance, when combined with metagenomic WGS the system can generate up to 1.5Tb of read information through Illumina NovaSeq 6000. The read separations obtained from the Illumina sequencing are of the top quality [121].
3.2.2. Applied Biosystems Sequencer
The Applied Biosystems (AB) sequencer is a popular tool in metagenomics research because of its special “sequencing by ligation” technique, which enables double-base encoding. Because each base is questioned twice during sequencing, the platform has a far lower chance of error compared to other similar ones, which contributes to its high accuracy. Despite having higher accuracy they have a read length of 85bp which is prone to cause errors in the assembly and generate increased sequencing time [122].
3.2.3. Ion Torrent Sequencing
The Ion torrent sequencer senses the changes in hydrogen ion concentration that happen when a nucleotide is inserted into the sequence. The output of the sequencing quality is great and reliable and is cost-effective when compared to Illumina or pyrosequencing[122]. Further, the “Ion 16S Metagenomics Solution” offers a specialised workflow for analysing bacterial diversity within a sample by targeting and sequencing a specific gene like the 16S rRNA[123].
3.2.4. Roche 454 Genome Sequencer
The Roche 454 genome sequencer is a high-throughput sequencing tool that is frequently used in metagenomics studies. The 454 genome sequencer is useful for building and analysing complex microbial communities from environmental samples. Finding the pyrophosphate (PPi) chemical that is generated when a nucleotide is integrated into the polymerase is the foundation of a real-time sequencing-by-synthesis (SBS) pyrosequencing technique. However, the efficacy of this sequencer is not as great as Illumina sequencing [124].
3.2.5. PacBio Technology
PacBio eliminates the need for DNA amplification, allowing for faster sequence reading. It also reads the complete DNA molecule in real-time as it traverses through a specialised sequencing device. The typical sequencing read length ranges between 8 and 15 kb, with the most complex sequences spanning 40 to 70 kb. Can identify small variations at a rate of 0.1%. Because there is no PCR amplification stage, base changes may be identified promptly during sequencing having a precision of as high as 99% [125,126,127].
3.2.6. Oxford Nanopore Technology (ONT)
ONT can overcome the limitations of short reads, such as identifying structural variations and highly repetitive areas, thanks to its ultra-long read capabilities. In ONT, DNA and RNA bases are characterized by detecting different ionic current signals according to their characteristics when they pass through nanopores[128]. In addition to sequencing nucleic acid molecules with lengths well above 100,000 bp, ONT produces reads with N50 values up to 40,000 bp, which are longer than the average read length due to the unique sequencing technology. In addition to real-time base detection the MinION (ONT commercialised product) is a low-cost technology that collects samples and sequences them in a lab. Consequently, nanopore sequencing will eventually replace all other sequencing devices [129]. Other currently available versions of ONT include, benchtop GridION, VolTRAX, and high-throughput PromethION [130].
4. Synergistic Benefits of Synthetic Biology and Metagenomics
To investigate, comprehend, and create biological systems, the areas of SynBio and metagenomics have come together to form a potent synergy. Scientists can find novel genes and antibiotics from a variety of habitats by merging SynBio and metagenomics. As resistance mechanisms continue to evolve and outstrip our strategies for fighting these diseases, antimicrobial-resistant (AMR) bacterial pathogens pose an ever-increasing threat to human health. Many antibiotics are classified as natural products (NPs) because they are naturally occurring substances that are released by bacteria and fungus. Numerous small molecule NPs called secondary metabolites in bacteria are encoded by biosynthetic gene clusters (BGC), which may be found by genomic sequencing and automated genome mining technologies [131]. Advanced computer bioinformatics methods are used by programs like Secondary Metabolite Unique Regions Finder (SMURF) and Antibiotics and Secondary Metabolite Analysis Shell (antiSMASH) to predict and locate BGCs in genomic data [132,133].
BGCs that are found, but not connected with known secondary metabolites, are called “cryptic” since they have been previously undetected or transcriptionally quiet. Following the discovery of these cryptic BGCs in the genome, bioengineering techniques may be used to edit the genome and either stimulate the production of the encoded NP or possibly change it to produce novel antibiotics and more chemical diversity [134]. With the right methods, bioengineering and molecular cloning concepts may be applied to biological systems that generate protein or small molecule products with specific, controlled characteristics. To alter, reimagine, or create new biological systems, these methods often seek to change gene circuits, genes, and metabolic pathways [135]. Through the activation and manipulation of cryptic BGCs encoding potentially new antibiotic secondary metabolites, SynBio and metagenomics can leverage these strategies to help combat AMR bacterial infections by increasing the chemical diversity of antibiotics. To extract and investigate potentially new antibiotics from understudied and environmental microorganisms, methods centred on genomic mutagenesis, endogenous gene expression, and heterologous gene expression can be employed [136].
The convergence of metagenomics and SynBio is increasingly being used as an approach to anticipate and induce new, natural antibiotics. By using SynBio perspective, advances in cloning and mutagenesis techniques may be used to study the natural control of antibiotic biosynthetic gene clusters (BGCs) and find possible changes that could result in stronger antibiotic action. Furthermore, a large number of cryptic antibiotic BGCs originate from the biosynthesis pathways of polyketide synthase (PKS) and non-ribosomal peptide synthase (NRPS); these routes are complicated, clustered genetic sequences that result in natural compounds produced from amino acids [134].
To regulate and manipulate gene expression, gene circuits, or metabolic pathways in non-native species, SynBio may be applied to current genome editing methods through the use of designed and constructed components. Numerous methods exist that may be adjusted and customized for the activation of cryptic BGC. To activate BGCs and create their encoded natural product as antibiotics, SynBio uses genomic mutagenesis techniques to induce genetic changes inside a bacterium’s genome. Cloning and genetic engineering methods are being employed in SynBio to activate cryptic BGCs utilizing endogenous and heterologous expression platforms. Activating these mysterious BGCs aims to find, describe, and employ new natural compounds as antibiotics. Researchers and scientists might be able to start a new generation of antibiotic production that can go back in time and stop the impending threat of the growing list of MDR and XDR bacterial pathogens if these techniques can be successfully modified and used to concentrate on antibiotic BGC discovery and characterization [134].
5. Diverse Applications of Synthetic Biology and Metagenomics
Although microbes have their immense significance, only a small percentage of microbes have been successfully cultivated in the lab. It is now evident that molecular methods have been crucial in identifying and revealing the vast diversity of microorganisms. Characterization of a large and unexplored portion of the microbial diversity was made possible by “omic” technologies such as single-cell genomics and metagenomics etc. [137]
SynBio is a scientific field that is initiated at the start of the current millennium aa visionary extension of what was previously known as genetic engineering or recombinant DNA technology in the 1970s. Enhancing the genetic engineering process through a multidisciplinary approach that combines the fundamental sciences of chemistry, biology, mathematics, and biophysics with biological, chemical, and electrical engineering techniques is one of the main objectives of SynBio. Its broadest definition includes cell-state analysis, systems biology and bioinformatics, DNA synthesis and assembly, genetic programming, protein engineering, pathway engineering, metabolic engineering, computer-aided design, in vitro biochemistry, and artificial life [138]. Many of these SynBio fields have obvious uses in the biotechnology and biomanufacturing sectors microbial fermentations can be optimized to produce chemicals, industrial enzymes, and biofuels; microbial fermentations can be used to produce natural products for plant-crop protection, animal health, and human medicine; and bacterial and mammalian cells can be engineered to produce human therapeutic proteins. This viewpoint presents reviews and original [139,140]
Metagenomics facilitates the examination of all microorganisms, irrespective of their culturing capabilities, by analyzing genomic data sourced directly from environmental samples. This approach yields insights into the species present and enables the extraction of information concerning the functional roles of microbial communities within their natural environments. Function-based screenings, which involve the cloning and expression of metagenomic DNA in a heterologous host, can be utilized to identify novel proteins of industrial significance that are encoded by genes from microorganisms that were previously difficult to access. [141]
The integration of SynBio and metagenomics is anticipated to represent a major advancement in the engineering of intricate biological systems. The complete biosphere, encompassing its myriad life forms, can be viewed as a reservoir of evolutionary innovations and a valuable source of modules for SynBio. Additionally, metagenomics, which represents a significant aspect of biodiversity, ought to be recognized as a crucial source of these modules.
Standardized integration of components, devices, circuits, and modules into functional chassis is part of the abstraction hierarchy of combining SynBio and metagenomics (also known as “synthetic metagenomics”). The engineering of biodiversity sustainability in multicellular entities, their interconnections, and their dynamics in communities and entire ecosystems are all covered by these principles, which go beyond the confines of single cell design. [14]
In order to design basic functional units, which are typically bottom-up and could be referred to as modules, SynBio has essentially “borrowed” concepts from electrical engineering or software. Therefore, SynBio is a technoscience founded on the core notion that modules can be integrated, combined, or arranged as interchangeable systems to create complexity. [142]
In order to produce ever-more intricate designs, scientists have taken components from natural gene circuits and altered them since the development of recombinant DNA technology. These gene circuits use separate and interacting components to produce particular responses, and they depend on regulatory factors for cellular functions. This has led to the development of flexible components and tools for gene circuit engineering. In SynBio, a “device” is a group of biological components that cooperate to produce a specific beneficial function or behavior. More intricate structures can be created by scaling up or further integrating these devices. For instance, a bacterium that has been genetically modified to break down plastic polymers into their monomers will be able to both catabolically “burn” the hydrolysis products of the broken polymer and use anabolic processes to turn the monomers into biomass. [143,144]
By utilizing the genetic and biochemical diversity found in the genomes of microorganisms from environmental samples or through metagenomics, SynBio’s integration of metagenomics encourages the development of sophisticated modular design that uses host cells for customized bioactivities. Since its fundamental ideas—modular integration, standardization, abstraction, and orthogonality—apply to the engineering of biodiversity sustainability in single cells as well as in multicellular entities, their interconnections, and their dynamics in communities and entire ecosystems, we expect that this integration will greatly advance the entire SynBio field. The biosphere as a whole, with all of its varied life forms, can also be viewed as a source of SynBio modules and a repository for evolutionary innovations. Unquestionably, a significant portion of biodiversity is derived from metagenomics, which should be regarded as a valuable source of modules. Standardized processes can be used to take advantage of the bioengineering of these modules in particular situations to develop new biobased economies and technologies, as well as artificial cells and communities.[14]
Analyzing and sequencing the DNA or RNA found in a sample from a specific ecological niche is known as metagenomics. Genetic information is retrieved using sophisticated sequencing technologies because this material is typically fragmented and piecemeal. Metagenomics, in theory, enables us to investigate the genetic makeup of entire microbial communities in their native environments, uncovering the ecosystems’ hidden biodiversity and functional potential. In this regard, metagenomics is developing into a technology that uses SynBio’s system design concepts and enhances gene and protein engineering techniques. Utilizing the abundance of genetic and biochemical diversity found in the genomes of microorganisms found in environmental samples metagenomics provides a variety of new technologies for screening environmental samples for novel catalytic activities that may have biotechnological or even diagnostic uses. For instance, the extent of novel compounds encoded in the enormous bacterial diversity and richness of the marine ecosystem is starting to be revealed by metagenomic analyses of marine ecosystems.[14] [145] [146]
Our knowledge of ecosystems and biodiversity has increased as a result of metagenomic analyses in a number of areas, including plastic biodegradation, oil spill bioremediation, and antimicrobial resistance in waste management systems. Consequently, metagenomes are functionally studied using the main tenets of SynBio in the developing field of synthetic metagenomics. The list of recognized biological activities and building blocks is significantly increased by this method. [147]
The field of SynBio makes it possible to create controllable metabolic and regulatory networks, circuits, and pathways. Additionally, it makes it possible to create whole cell systems, pathways, and new enzymes. As a result, SynBio makes it possible to create new organisms that can be engineered to generate useful products like medications, as well as chemicals, fuels, and materials that avoid using non-renewable fossil fuels. Human, bacterial, viral, and plant cells have all been manipulated using SynBio to create compounds or change processes that may improve human health. SynBio has had a big impact on human health and disease treatment, from fortifying crops to modifying T cells. A fresh avenue for disease treatment research is provided by SynBio. This entails the therapeutic and diagnostic engineering of bacterial communities or cells. Salmonella, for instance, can regulate tumour growth by releasing drugs in a time-dependent manner in response to the hypoxic microenvironment of the tumour. Salmonella carries genes for synthetic antechamber drugs. 2010 saw the development of viruses that specifically target brain tumour cells; oncolytic viral therapy was used to stop the growth of blood vessels in brain tumour cells by focusing on a protein known as a vascular statin, which prevents blood vessel growth. Additionally, SynBio can be applied to biosensing and cellular homeostasis regulation, which can change how a disease progresses. The use of engineered T cells, such as chimeric antigen receptor T cell immunotherapy (CAR-T), which is used to treat cancers, is an example of SynBio in this context. Tisagenlecleucel was the first CAR-T treatment to receive FDA approval [148].
5.1. Impact on Biotechnology-Based Research and Development
SynBio and omics technologies (Metagenomics) have had a significant impact on biotechnology through a deeper understanding of biological systems and the ability to design and engineer organisms with specialized functions. This has led to advancements in fields such as biomanufacturing, medicine, and agriculture. SynBio and metagenomics have an important role in biotechnology like the development of organisms with specialized characteristics for particular uses, such as the production of useful macromolecules or the invention of novel treatments. New techniques in biotechnology can optimize an organism’s metabolic processes to produce more of the desired chemicals and to alter cells to produce new protein-based medications, cell treatments, and customized medicine techniques. Below are the modern techniques in which SynBio and metagenomics help revolutionise biotechnology by making it possible to create new biological systems with tailored functions.
5.1.1. Synthetic Biology and CRISPR-Cas9
CRISPR-Cas9 has been profoundly impacted by SynBio and this is a powerful genome editing tool that allows scientists to make precise changes to DNA sequences in living organisms. It has revolutionized the field of genetic engineering and has enormous potential for developing new treatments for genetic diseases. Through system optimization and fine-tuning, more accurate gene editing with fewer off-target effects, better delivery systems, and the creation of intricate genetic circuits that can regulate CRISPR activity depending on particular conditions, SynBio has had a substantial impact on CRISPR-Cas9 and increased its potential uses in industrial biotechnology, medicine, and agriculture. [149] Metagenomics has had a significant impact on CRISPR-Cas9 technology by offering a large reservoir of novel CRISPR systems from various microbial communities. This has enabled researchers to find new Cas enzymes with distinct properties, which could result in increased efficiency, specificity, and applications for genome editing across a variety of organisms, including ones that were previously challenging to target with current CRISPR systems. Scientists can discover novel CRISPR-Cas systems that might differ from the widely used CRISPR-Cas9 system in terms of target recognition sequences or cutting capabilities by using metagenomics to analyze DNA sequences from environmental samples. Natural anti-CRISPR proteins can also be found through metagenomic research, which is useful for comprehending CRISPR mechanisms and creating instruments to regulate CRISPR activity. [150]
5.1.2. DNA Synthesis
The creation of unique DNA sequences is now simpler and less expensive thanks to developments in DNA synthesis technology. This has made it possible for scientists to start from scratch and create completely new genes and genetic circuits. The design, build, test, learn, and repeat cycle that underpins advancements in SynBio is made possible by the chemical synthesis of DNA oligonucleotides and their assembly into synthons, genes, circuits, and even entire genomes using gene synthesis techniques. [151] Metagenomics fundamentally expands the potential for DNA synthesis beyond known organisms by enabling researchers to access and sequence genetic material directly from environmental samples. This opens up a vast pool of novel DNA sequences from previously uncultured microorganisms, allowing the discovery of new genes and potential applications in areas like drug discovery, bioremediation, and enzyme development by eliminating the need to first isolate and cultivate individual species in a lab.[152]
5.1.3. Directed Evolution
Through the use of genetic engineering, scientists can produce vast collections of mutant genes and subsequently select for desired characteristics. New proteins, enzymes, and other molecules with a variety of uses have been created using this method. Even though directed evolution’s therapeutic uses are still in their infancy, this method has potential in several therapeutic domains that are significant for cardiovascular disease patients, including the following: Enzyme replacement therapies, drug development, enhancing tissue repair, and antibody development. [153] By giving researchers access to a large and hitherto untapped reservoir of genetic diversity from uncultured microorganisms, metagenomics has had a significant impact on directed evolution. This has sped up the process of discovering new enzymes, antibodies, and other functional molecules with unique properties that are difficult to find in well-studied organisms. All things considered, metagenomics has greatly increased the potential of directed evolution by providing access to a large genetic diversity reservoir from unidentified microorganisms, which has made it possible to find new proteins with useful roles for a range of applications. [154]
5.1.4. Synthetic Cells
Researchers have succeeded in producing artificial cells that can carry out fundamental tasks like reproduction and stimulus-response. These cells may find use in environmental remediation, biotechnology, and medicine. SynBio and its evolution have made its application in many genetic tools for reprogramming and engineering cells for enhanced performance, new functions, and a variety of applications. Research and development of new treatments may benefit greatly from the use of such cell engineering resources. With the help of SynBio, synthetic cells have the potential to develop engineered cell-based therapeutics for cancer, and engineered cell-based therapeutics for infectious diseases. engineered cell-based therapeutics for imbalanced flora-related diseases. [135] Through metagenomics, scientists can find genes with distinct roles not present in well-studied model organisms. These genes can then be integrated into artificial cells to produce novel cellular behaviours or metabolic pathways. All things considered, metagenomics gives synthetic biologists access to a large genetic reservoir, facilitating the production of artificial cells with distinct functions and potential uses in a range of industries, including environmental remediation, bioenergy, and medicine [155].
5.1.5. Metabolic Engineering
The development of metabolic engineering and SynBio has been greatly impacted by the new tools and techniques introduced by the “-omics” era. Metabolic engineering entails optimizing metabolic pathways in organisms, like yeast and bacteria, through genetic engineering to generate useful chemicals and fuels. While SynBio is primarily concerned with basic biological research made possible by the use of synthetic DNA and genetic circuits, metabolic engineering is about designing cell factories for the biological production of chemical and pharmaceutical products. These two exciting fields have made significant biotechnology advancements and helped to solve issues with the production of medications, vaccines, chemical compounds, etc. are metabolic engineering and SynBio. [156] Metagenomics help researchers to discover new proteins, enzymes, and biochemical pathways, which can also identify viruses and microorganisms. This provides biological information that is useful for metabolic engineering goals. One of the best ways to inform strategies to engineer a cell to produce a target compound is to learn how the cell is currently using its biochemical resources. It is fairly common practice to drive metabolic engineering by analyzing the target compound’s (or a few closely related molecules’) extracellular or intracellular levels. Additionally, metagenomics has the potential to have a major biotechnological impact on metabolic engineering because it naturally supports the goal of manipulating metabolite production, which is the aim of metabolic engineering. Furthermore, using metagenomics to study different strains originating from both directed evolution strategies and rational design can be interpreted and understood in the rich biological context provided by the extensive study of organisms like S. cerevisiae and E. coli. [156].
5.1.6. Microbial Biosensing
SynBio has facilitated the development of microbial biosensors, organisms designed to detect and respond to specific environmental stimuli. These biosensors are important tools for monitoring environmental pollutants [157], detecting pathogens [158], and assessing soil health in agriculture [159]. Synthetic pathways enable microbes to produce detectable signals like fluorescence upon exposure to target molecules, providing real-time monitoring capabilities [160]. An example includes the engineering of Pseudomonas putida to sense and degrade environmental pollutants such as toluene and other hydrocarbons [158,161]. These microbial sensors offer cost-effective, scalable, and in situ detection solutions.
5.1.7. Therapeutic Applications
SynBio has enabled advances in microbial research that are transforming therapeutic applications, particularly in designing probiotics and engineered bacteria for disease treatment. Engineered strains of Lactobacillus and Escherichia coli have been developed to deliver therapeutic molecules directly to the gut, modulate the gut microbiome, and even target tumours [162]. For instance, synthetic gene circuits have allowed E. coli strains to sense and respond to inflammation markers, releasing anti-inflammatory compounds in conditions like inflammatory bowel disease [163]; [164]. Furthermore, genetically modified microbes are being explored as live biotherapeutics for metabolic disorders, cancer, and infection control [165].
5.2. Using Synthetic Microbes to Improve the Environment
Synthetic biology has a significant impact on the development of synthetic microbes enabling scientists to create custom microorganisms with improved capacities to degrade particular pollutants in the bioremediation. These microorganisms can target complex pollutants, withstand harsh conditions, and even detect contaminants through biosensing mechanisms. In other words, synthetic biology creates “tailor-made” microbes for specific bioremediation efforts.
SynBio helps the use of extremophilic microorganisms and is one of the essential SynBio resources for the development of bioremediation technologies in particular. These organisms offer a multitude of functionally adapted enzymes. In harsh conditions that would cause other proteins to denature. For instance, thermotolerant microalgae Galdieria sulphyraria, when combined with heterotrophic bacteria, can remediate ammonium and phosphates from wastewater systems, producing biofuel without requiring the energy-intensive cooling of the photobioreactor system. Halophilic bacteria can also break down petroleum and polycyclic aromatic hydrocarbons from highly saline wastewater [166].
According to purists, prokaryotic organisms are currently modified or artificially created using SynBio techniques. Although SynBio’s use in more intricate, multicellular organisms is still in its early stages. However, with incredible potential, ambitious projects like the C4 Rice project (c4rice.com/the-project-2), engineering nitrogen-fixing cereals are currently in progress. However, the development of microbial-based systems remains the primary focus of the application of SynBio techniques in the field of bioremediation. Although plants play a significant part in bioremediation, current technologies only express one or a small number of transgenes, and fully SynBio techniques have not yet been developed [166,167].
5.2.1. Engineered Biofilms for Bioremediation
A new area of study in bioremediation is bioengineered biofilms, which are designed to overcome the inherent limitations of biofilms and lessen the effects of hydrocarbons, pesticides, and hazardous metals in the environment. Biofilms’ capacity for regeneration makes them suitable for sustainable use, while the composition of extracellular polymeric substance (EPS) can be tuned to perform particular tasks for a range of bioremediation applications. Among the conventional bioremediation uses that biofilm engineering can enhance are bioleaching (heavy metal dissolution from the solid matrix), metal corrosion prevention, and wastewater treatment using microbial fuel cells (MFCs). For example, a mercury detector (MerR promoter) was used in a selective mercury detoxification system that was. For example, a mercury detector (MerR promoter) was constitutively expressed in E. coli as part of a selective mercury detoxification system [16].
Finding desirable characteristics in artificially selected microbial communities synthesized by SynBio can greatly aid bioremediation. A consortium of synthetic microorganisms entails the co-cultivation of specific strains with favorable characteristics. Every microbe can take part in the degradation pathway in this configuration. Therefore, it is possible to prevent over-engineering of a single strain and the associated metabolic load. With the help of SynBio it is easy to develop bioremediation as an inexpensive, labor-, energy-, and chemical-saving technique for managing pollution. Heavy metals pose environmental and longer times and inadequate metal removal are frequent drawbacks of using these procedures on a large scale. Consequently, synthetic biologists began addressing the issues by altering the systems of living organisms. Inserting resistance genes is one method of increasing tolerance. Recently, several engineering techniques have been proposed to improve microorganisms’ resistance to Cu. The use of SynBio in hydrocarbon bioremediation is becoming more widespread. Hexachlorocyclohexane (HCH), petroleum and its derivatives, polyaromatic hydrocarbons (PAHs), Polychlorinated biphenyls (PCBs), chloroalkanes, organic halides, and aromatic dyes are removed applying new degradation pathways through SynBio. [168] [169] Multiple microorganisms, such as actinomycetes, algae, bacteria, and fungi, use enzymes to drive the microbial decontamination of plastics. With the help of SynBio and metagenomics the ability of Pseudomonas, Escherichia, and Bacillus species to enzymatically depolymerize plastics has been improved. The majority of plastic biodegradation engineering research has concentrated on altering the genes that code for efficient degradative enzymes. Particularly useful were Pseudomonas species that provided effective enzymes for cloning in other bacterial hosts. Similarly, SynBio and metagenomics have been used to enhance the breakdown of organophosphorus compounds (OPCs), a common class of pesticides, the enzyme organophosphate hydrolase (OPH) was developed. Key players in the breakdown of xenobiotics, such as cytochrome P450 systems and dioxygenases, have been discovered and characterized thanks to functional metagenomics techniques [16,170].
5.2.2. Ecosystem Restoration
SynBio and metagenomics can address problems like invasive species, pollution removal, and the recovery of endangered native populations within an ecosystem. SynBio can be used to actively restore a degraded environment by employing genetic engineering techniques to create and introduce modified organisms that are specifically suited to address environmental degradation. This could improve biodiversity, ecosystem services, and resilience. Targeted interventions can be designed with the help of SynBio. Researchers can create organisms with particular characteristics to address specific environmental issues. For example, bacteria can be created to break down particular pollutants in contaminated soil, or algae can be created to absorb heavy metals from water. The basic research and application of SynBio as a tool for eco-engineering or working directly with biodiversity, the field of ecological restoration is now beginning. Landscape designers or council employees may use databases of socially engaged SynBio to guide their designs for restoring biodiversity habitats.[171]
Crude oil spills contain thousands of compounds and polycyclic aromatic compounds (PACs) are particularly dangerous because they can cause cancer [172], mutagenesis, and acute and chronic toxicity to living things. By incorporating synthetic biology and metagenomics techniques designed to understand the microbiome linked to hydrocarbon plastics, bioremediation procedures for oil spills can be improved, which in turn helps in ecological restoration. A thorough grasp of the degradation processes of bacteria and fungi, their complex interactions within microbiomes, and their interactions with the soil matrix are essential to achieving this goal. Although petroleum-based synthetic plastics like polyethylene (PE), polypropylene (PP), polyvinyl chloride (PVC), polyethylene terephthalate (PET), and polystyrene (PS) are essential to modern industry and daily life, their extreme resistance to environmental biodegradation has resulted in serious environmental issues worldwide. According to several studies, Arthrobacter and Mortierella were either able to break down plastics or were enriched on plastics in soils. According to metagenomic analyses, and SynBio certain and prevalent microbial groups in both terrestrial and marine environments have adapted to degrade the plastics and helps in ecological restoration.[171,173]. In addition to potentially alleviating the strain on ecosystems by providing sustainable food for a future world of nine billion people, SynBio may offer novel answers to well-known “wicked” problems like animal disease. [174]
5.2.3. Microbial Consortia Engineering
SynBio has also driven the design of microbial consortia, groups of different microorganisms engineered to work collaboratively. These consortia can be structured to perform complex functions that single organisms cannot efficiently achieve. In microbial research, consortia can be engineered for applications like nutrient cycling, pollutant degradation, and bioproduction [175]. For example, synthetic consortia are used in wastewater treatment, where various microbes sequentially degrade organic pollutants, heavy metals, and other contaminants [176]. This multi-organism approach optimizes resource utilization and enhances resilience in fluctuating environments.
5.3. Potential for Novel Applications in Medicine and Agriculture
The integration of SynBio and metagenomics offers transformative potential for both medicine and agriculture by harnessing microbial diversity and engineering capabilities. The combination of these fields allows for precise manipulation of microbial genomes and exploration of previously inaccessible microbial communities, opening doors for innovation across several domains.
5.3.1. Medicine
5.3.1.1. Microbiome-Based Therapies
The human microbiome, consisting of trillions of microbes inhabiting various parts of the body, plays a critical role in maintaining health and preventing disease. Recent advances in SynBio and metagenomics are enabling unprecedented exploration and manipulation of these microbial communities, particularly within the gut, where microbial imbalances are linked to a wide range of diseases, including gastrointestinal disorders, metabolic syndromes, and even cancers [177]. By mapping out the vast genetic diversity of the gut microbiota through metagenomic approaches, researchers can identify specific bacterial strains that are either beneficial or pathogenic.
SynBio offers the toolkit to engineer these microbes, turning them into “living medicines” that can address health conditions with greater precision than traditional pharmaceuticals. For example, engineered bacteria can be designed to release anti-inflammatory compounds directly in the gut, which can help treat inflammatory bowel diseases like Crohn’s disease or ulcerative colitis [178]. Similarly, synthetic microbes can be programmed to degrade toxins, regulate metabolic processes, or produce essential metabolites that are deficient in patients with certain genetic disorders [179].
Another promising area is cancer treatment. Certain engineered microbial strains have been developed to either stimulate the immune system to recognize and attack cancer cells or to deliver anti-cancer drugs in a targeted manner. For instance, synthetic bacteria have been designed to home in on tumors and produce tumor-killing agents locally, reducing side effects and improving efficacy over systemic treatments [177]. This approach not only opens new therapeutic avenues but also underscores the versatility of SynBio in utilizing the human microbiome to address complex diseases.
Moreover, microbiome-based therapies provide an alternative to broad-spectrum antibiotics. Traditional antibiotics often disrupt healthy microbial communities, leading to issues such as antibiotic-associated diarrhea or secondary infections like Clostridium difficile [180]. Engineered probiotics, on the other hand, can selectively target pathogens while preserving the balance of beneficial microbes, offering a more nuanced and sustainable approach to treating infections.
5.3.1.2. Antibiotic Resistance Management
Antibiotic resistance is one of the most critical public health threats of the 21st century, with resistant bacteria causing millions of infections and thousands of deaths annually [181]. The overuse and misuse of antibiotics in both medicine and agriculture have accelerated the emergence of resistant strains, often referred to as “superbugs.” Metagenomics has become a powerful tool in this fight, allowing scientists to scan microbial communities—whether from clinical, environmental, or agricultural sources—for genes that confer resistance to antibiotics [182]. By understanding how these genes spread within and between microbial populations, researchers can devise more effective strategies to counteract them.
SynBio can build on this knowledge to combat resistance in novel ways. One promising approach involves the use of CRISPR-Cas systems, originally discovered as a bacterial defense mechanism against viruses, to selectively target and disable antibiotic resistance genes in pathogenic bacteria [183]. By engineering CRISPR-based tools, scientists can design systems that recognize and cleave resistance genes, rendering resistant bacteria susceptible to antibiotics once again [183]. This targeted strategy avoids the collateral damage to healthy microbiota that is often seen with broad-spectrum antibiotics.
Additionally, SynBio can be used to create new classes of antibiotics. Using metagenomic data, researchers have identified previously unknown biosynthetic gene clusters in environmental samples that are responsible for the production of natural antibiotics. These gene clusters can be transferred into laboratory strains of bacteria using SynBio techniques, enabling the mass production of novel antibiotics that have not yet faced resistance [184]. These “mined” antibiotics from metagenomic databases represent a new frontier in drug discovery, offering hope in the fight against resistant pathogens.
Engineered microbes can also play a role in preventing the spread of resistance. For example, synthetic bacteria could be designed to outcompete resistant strains in the human body or in agricultural environments by producing compounds that inhibit resistance mechanisms or disrupt biofilms where resistant bacteria tend to thrive [185]. These engineered organisms could be deployed as part of a preventative strategy to reduce the prevalence of resistant bacteria in hospitals, food production, or natural ecosystems.
Figure 4.
Diverse applications of SynBio and Metagenomics.
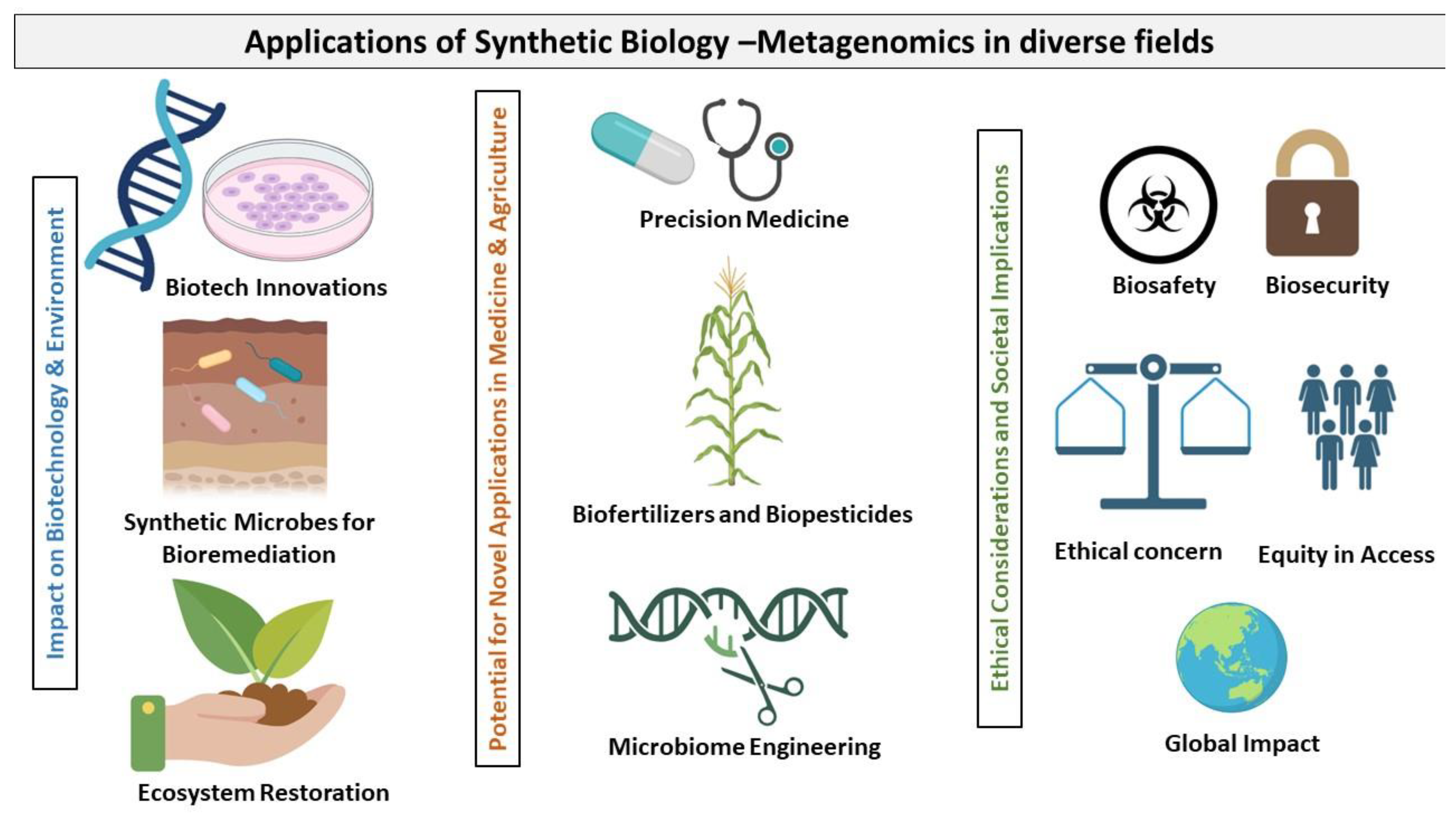
5.3.2. Agriculture
5.3.2.1. Engineered Biofertilizers and Biopesticides
Agriculture has long relied on chemical fertilizers and pesticides to meet the growing demand for food production. However, the environmental costs of these practices—such as soil degradation, water pollution, and loss of biodiversity—have prompted the search for more sustainable alternatives. The convergence of SynBio and metagenomics offers a powerful solution by tapping into the vast diversity of soil microorganisms and enhancing their natural abilities to promote plant growth and protect crops.
Metagenomics allows for the comprehensive exploration of soil microbial communities, uncovering microbes that play pivotal roles in nutrient cycling, such as nitrogen-fixing bacteria (Rhizobium, Azotobacter) and phosphate-solubilizing fungi (Penicillium, Aspergillus) [186]. By identifying and cataloging the genes involved in these processes, researchers can gain insights into how microbial consortia contribute to soil fertility and plant health. These metagenomic datasets serve as the foundation for SynBio, which enables the precise manipulation of microbial genomes to enhance or modify their functions.
Through SynBio techniques, beneficial microbes can be engineered to optimize nutrient cycling, providing a next-generation biofertilizer that reduces dependence on chemical inputs. For instance, engineered nitrogen-fixing bacteria can be designed to thrive in diverse soil conditions and enhance nitrogen availability to plants, promoting higher yields while minimizing the environmental impact of synthetic nitrogen fertilizers [187]. Similarly, phosphate-solubilizing microbes can be genetically engineered to increase their efficiency in making phosphorus—an essential but often limiting nutrient—available to crops.
In addition to biofertilizers, the integration of metagenomics and SynBio is revolutionizing biopesticides. Beneficial soil microbes that produce natural antimicrobial or insecticidal compounds can be identified through metagenomic screening and subsequently enhanced using SynBio to create robust, environmentally friendly biopesticides. These engineered organisms can target specific pests or pathogens without the negative ecological consequences associated with chemical pesticides. For example, microbes engineered to produce specific insecticidal proteins or antifungal compounds can protect crops from harmful pathogens while preserving beneficial organisms, such as pollinators [188].
Moreover, engineered microbial consortia can be tailored to local soil ecosystems, ensuring that biofertilizers and biopesticides are both effective and environmentally compatible. This shift from synthetic chemicals to biologically engineered solutions hold great promise for advancing sustainable farming practices, improving soil health, and reducing the environmental footprint of agriculture.
5.3.2.2. Crop Microbiome Engineering
Plants, like humans, have their own microbiomes—complex communities of bacteria, fungi, and other microorganisms that live on and within them. These microbial communities play a critical role in promoting plant health, resilience, and productivity. By studying plant-associated microbiomes through metagenomic analysis, scientists are gaining new insights into how these microbes influence crop performance, opening up the possibility of engineering plant microbiomes to enhance agricultural output in the face of challenges such as climate change, disease outbreaks, and soil degradation [189].
SynBio is enabling researchers to design microbes that can confer specific benefits to crops, such as increased tolerance to drought, heat, or salinity. For instance, microbes that produce plant growth-promoting hormones like auxins or gibberellins can be engineered to enhance root development, enabling crops to access water and nutrients more efficiently, even in poor soil conditions [190]. These engineered microbes can also help plants cope with abiotic stress, such as extreme temperatures or water scarcity, by enhancing their resilience mechanisms, such as antioxidant production or osmotic adjustment.
Moreover, crop microbiome engineering has the potential to boost plant immunity against pathogens. Using metagenomic data, scientists can identify beneficial microbes that naturally suppress plant diseases, such as bacterial or fungal infections. By applying SynBio, these microbes can be enhanced to provide stronger or more targeted protection. For example, bacteria that produce antifungal compounds can be engineered to colonize plant roots and protect against soil-borne pathogens like Fusarium or Phytophthora [191]. This approach reduces the need for chemical fungicides, making crop protection both safer and more sustainable.
The ability to engineer the crop microbiome also has significant implications for improving agricultural sustainability. By designing microbial consortia that promote nutrient uptake, protect against pests, and improve resilience to environmental stressors, SynBio could help shift agriculture toward a more sustainable model that relies less on synthetic inputs and more on natural processes. This is particularly critical in the context of climate change, which is expected to increase the frequency of droughts, floods, and other extreme weather events that threaten global food security [186]. Engineered plant-associated microbes could help crops withstand these challenges, ensuring stable food production even under changing environmental conditions.
5.3.3. Ethical Considerations and Societal Implications
The convergence of SynBio and metagenomics offers transformative opportunities but also presents significant ethical and societal concerns. These technologies could dramatically alter ecosystems, human health, and agricultural systems, raising critical questions regarding safety, equity, and environmental impact.
5.3.3.1. Biosafety and Biosecurity
5.3.3.1.1 Unintended Consequences:
One of the primary concerns with SynBio and metagenomics is the unpredictability of engineered microbes once they are released into the environment or the human body. Synthetic organisms could disrupt natural microbial communities, potentially leading to ecological imbalances. Horizontal gene transfer between engineered microbes and native organisms could result in the spread of synthetic traits in unintended ways, which may lead to the emergence of new pathogenic strains [192]. For example, if engineered microbes designed for agricultural purposes transfer traits to environmental microbes, they could inadvertently disrupt soil health or nutrient cycles.
5.3.3.1.2 Containment and Control
Developing reliable containment strategies for engineered organisms is essential to minimize risks. Features like “kill switches,” which enable synthetic organisms to self-destruct in case of uncontrolled spread, or other biocontainment mechanisms are crucial in mitigating biosafety hazards [193]. The global release of these organisms, however, necessitates the creation of international regulations and monitoring frameworks to prevent their misuse and manage biosafety risks effectively [194].
5.3.3.2. Equity and Access
5.3.3.2.1 Unequal Access to Technologies
The benefits of SynBio and metagenomics may not be evenly distributed across societies. Developing advanced microbial therapies or bioengineered agricultural products can be expensive, limiting access to wealthier nations or large corporations. As a result, low-income communities and small-scale farmers may be excluded from the benefits of microbial biofertilizers or therapeutics [195]. This raises concerns about exacerbating global inequalities, particularly in agriculture, where smallholder farmers in developing regions might not have access to high-tech biofertilizers or biopesticides [196].
5.3.3.2.2 Patents and Ownership
Intellectual property issues further complicate the use of SynBio and metagenomics. Innovations in SynBio are often subject to patent protections, which may limit access to genetic resources derived from metagenomic studies. Questions about who holds the rights to engineered organisms, particularly those based on microbial communities from nature, are increasingly important. There are also concerns about “biopiracy,” where genetic resources from developing countries may be exploited without fair compensation [197].
5.3.3.3. Environmental Impact and Sustainability
5.3.3.3.1 Long-Term Ecological Impact
The release of engineered organisms into ecosystems poses ecological risks, and their long-term effects are not fully understood. Synthetic organisms could outcompete natural species, potentially leading to biodiversity loss or disrupting food webs [198]. This requires comprehensive ecological risk assessments to evaluate potential long-term environmental effects. SynBio also holds the promise of solving environmental challenges, such as developing organisms that help restore degraded ecosystems, but their impacts need to be carefully considered.
5.3.3.3.2 Sustainability and Environmental Justice
While SynBio and metagenomics provide promising solutions for sustainability, such as reducing chemical use in agriculture or developing new methods for environmental remediation, their equitable implementation is crucial. For instance, communities most affected by environmental degradation should have a role in deciding how these technologies are applied locally [199]. Ensuring that the benefits of these innovations contribute to environmental justice and that marginalized communities are not further disadvantaged is a key ethical challenge.
6. Challenges and Limitations
Metagenome sequencing offers more promise than just new understandings of microbiome function since it opens up new avenues for microbial separation and culture. Specifically, the standard of genomic data from single organisms and species via MAGs may be enhanced for more precise metabolic interpretation. Metagenomic samples frequently offer intriguing novel biological activities and contain distinct genetically encoded information acquired from various geographical habitats, such as severe conditions (hot, cold, acidic, basic, and salty). Finding a suitable host or chassis that can express metagenomic DNA effectively is a major obstacle. This task is made more difficult by species-specific variations in metabolic profiles, regulatory networks, transcription, translation, and post-translation processes [200].
Therefore, in order to fully use the biological novelties of metagenomics, standardized hosts (chassis) and SynBio principles must be introduced. Thermophilic, psychrophilic, halophilic, mesophilic, and other species should all be included in these flexible chassis, which should be standardized platforms with integrated metagenomic libraries. For instance, microbial hosts (such as a chassis based on Escherichia coli) might be used in live bacterial therapies to guarantee robust colonization and drug delivery in difficult luminal conditions [201]. Other strategies include synthetic metagenomics in yeast, which has the potential to replicate the metabolic processes of a whole ecosystem in a single Saccharomyces cerevisiae cell, or the discovery of standardized chassis for genomic clusters encoding different natural products [14,202,203,204].
The creation of novel culture media is supported by data from SynBio and metagenomics. It is reported that targeted culture procedures are intricate and now necessitate a thorough understanding of genetic data to forecast cultivation requirements. After the target organisms have been separated, the right growth medium and physicochemical conditions must be determined to sustain their growth over time. However, because the chemical makeup of natural settings is sometimes unknown, genome sequences by themselves frequently do not give enough information to precisely determine all needs to grow a specific bacterium effectively. For instance, Lavy et al. created a medium for Candidatus Poribacteria sp. WGA-4E is based on the available genetic information [205]. Poribacteria were not captured in culture using this particular medium under the conditions used, even though metabolic characteristics of this phylum, such as the potential use of urea as the source of nitrogen and assimilatory reduced sulfate metabolism, were inferred from genomic data [206]. It is possible that the medium’s component concentrations weren’t ideal for Poribacteria, or that signal molecules needed to be added [205]. Researchers may be able to accomplish cultivation by combining metagenomic methodologies with other empirical techniques including culturomics, in situ culture, single-cell isolation, and factorial trial and error [204].
Author Contributions
Conceptualization, S.H. and P.S.; Validation, S.H. and P.S.; Data curation, S.H., M.M., E.H., A.T. and R.M; Writing—original draft preparation, M.M., E.H., A.T., A.Y.A., S.A.F., K.K.G., T.P., R.D., N.K., P.S. and S.H.; Writing—review and editing, S.H., A.T and P.S.; Visualization, E.H., R.M. and P.S.; Supervision, S.H. All authors have read and agreed to the published version of the manuscript.
Funding
None
Institutional Review Board Statement
Not Applicable.
Informed Consent Statement
Not Applicable.
Data Availability Statement
No new data has been generated in this work.
Acknowledgements
None
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Nannipieri, P.; Ascher, J.; Ceccherini, M.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. European journal of soil science 2017, 68, 12–26. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D.; Berdugo, M.; Campbell, C.D.; Singh, B.K. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nature communications 2016, 7, 10541. [Google Scholar] [CrossRef]
- Joshi, P.; Pande, V.; Joshi, P. Microbial diversity of aquatic ecosystem and its industrial potential. J. Bacteriol. Mycol. Open Access 2016, 3, 177–179. [Google Scholar]
- Structure, function and diversity of the healthy human microbiome. nature 2012, 486, 207–214. [CrossRef]
- Ezzamouri, B.; Shoaie, S.; Ledesma-Amaro, R. Synergies of systems biology and synthetic biology in human microbiome studies. Frontiers in microbiology 2021, 12, 681982. [Google Scholar] [CrossRef]
- Padilla-Vaca, F.; Anaya-Velázquez, F.; Franco, B. Synthetic biology: Novel approaches for microbiology. Int. Microbiol 2015, 18, 71–84. [Google Scholar] [PubMed]
- Duan, F.F.; Liu, J.H.; March, J.C. Engineered commensal bacteria reprogram intestinal cells into glucose-responsive insulin-secreting cells for the treatment of diabetes. Diabetes 2015, 64, 1794–1803. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.; Cao, L.; Ning, K. Microbiome big-data mining and applications using single-cell technologies and metagenomics approaches toward precision medicine. Frontiers in Genetics 2019, 10, 972. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Martins, L.; Page, A.J.; Mather, A.E.; Charles, I.G. Taxonomic resolution of the ribosomal RNA operon in bacteria: implications for its use with long-read sequencing. NAR Genomics and Bioinformatics 2020, 2, lqz016. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Li, Y.; Zhang, X. An overview of major metagenomic studies on human microbiomes in health and disease. Quantitative Biology 2016, 4, 192–206. [Google Scholar] [CrossRef]
- Carlino, N.; Blanco-Míguez, A.; Punčochář, M.; Mengoni, C.; Pinto, F.; Tatti, A.; Manghi, P.; Armanini, F.; Avagliano, M.; Barcenilla, C. Unexplored microbial diversity from 2,500 food metagenomes and links with the human microbiome. Cell 2024, 187, 5775–5795. e5715. [Google Scholar] [CrossRef]
- Oulas, A.; Pavloudi, C.; Polymenakou, P.; Pavlopoulos, G.A.; Papanikolaou, N.; Kotoulas, G.; Arvanitidis, C.; Iliopoulos, l. Metagenomics: tools and insights for analyzing next-generation sequencing data derived from biodiversity studies. Bioinformatics and biology insights 2015, 9, BBI. S12462. [Google Scholar] [CrossRef] [PubMed]
- Roell, G.W.; Zha, J.; Carr, R.R.; Koffas, M.A.; Fong, S.S.; Tang, Y.J. Engineering microbial consortia by division of labor. Microbial cell factories 2019, 18, 1–11. [Google Scholar] [CrossRef]
- Levin, D.B.; Budisa, N. Synthetic biology encompasses metagenomics, ecosystems, and biodiversity sustainability within its scope. Frontiers in Synthetic Biology 2023, 1, 1255472. [Google Scholar] [CrossRef]
- Ke, J.; Wang, B.; Yoshikuni, Y. Microbiome engineering: synthetic biology of plant-associated microbiomes in sustainable agriculture. Trends in Biotechnology 2021, 39, 244–261. [Google Scholar] [CrossRef]
- Aminian-Dehkordi, J.; Rahimi, S.; Golzar-Ahmadi, M.; Singh, A.; Lopez, J.; Ledesma-Amaro, R.; Mijakovic, I. Synthetic biology tools for environmental protection. Biotechnology Advances 2023, 108239. [Google Scholar] [CrossRef]
- Lee, S.J.; Lee, S.-J.; Lee, D.-W. Design and development of synthetic microbial platform cells for bioenergy. Frontiers in microbiology 2013, 4, 92. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Z.; Zhu, Y.; Zhang, Q.; Qiao, X.; Mu, R.; Xu, Z.; Yan, Y.; Wang, F.; Zhang, T.; Zhuang, W.-Q. Unravelling biosynthesis and biodegradation potentials of microbial dark matters in hypersaline lakes. Environmental Science and Ecotechnology 2024, 20, 100359. [Google Scholar] [CrossRef]
- Rahgu, K.; Choudhary, S.; Kushwaha, T.N.; Shekhar, S.; Tiwari, S.; Sheikh, I.A.; Srivastava, P. Microbes as a promising frontier in drug discovery: A comprehensive exploration of nature's microbial marvels. Acta Botanica Plantae. V02i02 2023, 24, 30. [Google Scholar]
- Weber, W.; Fussenegger, M. The impact of synthetic biology on drug discovery. Drug Discovery Today 2009, 14, 956–963. [Google Scholar] [CrossRef]
- Jain, A.; Bhatia, P.; Chugh, A. Microbial synthetic biology for human therapeutics. Systems and synthetic biology 2012, 6, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Seghal Kiran, G.; Ramasamy, P.; Sekar, S.; Ramu, M.; Hassan, S.; Ninawe, A.S.; Selvin, J. Synthetic biology approaches: Towards sustainable exploitation of marine bioactive molecules. International Journal of Biological Macromolecules 2018, 112, 1278–1288. [Google Scholar] [CrossRef]
- Leggieri, P.A.; Liu, Y.; Hayes, M.; Connors, B.; Seppälä, S.; O'Malley, M.A.; Venturelli, O.S. Integrating systems and synthetic biology to understand and engineer microbiomes. Annual Review of Biomedical Engineering 2021, 23, 169–201. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chen, F.; Zeng, Z.; Xu, M.; Sun, F.; Yang, L.; Bi, X.; Lin, Y.; Gao, Y.; Hao, H. Advances in metagenomics and its application in environmental microorganisms. Frontiers in microbiology 2021, 12, 766364. [Google Scholar] [CrossRef]
- Dhanjal, D.S.; Sharma, D. Microbial metagenomics for industrial and environmental bioprospecting: the unknown envoy. Microbial bioprospecting for sustainable development 2018, 327–352. [Google Scholar]
- Culligan, E.P.; Sleator, R.D.; Marchesi, J.R.; Hill, C. Metagenomics and novel gene discovery: promise and potential for novel therapeutics. Virulence 2014, 5, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R. Metabolic load and heterologous gene expression. Biotechnology advances 1995, 13, 247–261. [Google Scholar] [CrossRef]
- Brophy, J.A.; Voigt, C.A. Principles of genetic circuit design. Nature methods 2014, 11, 508–520. [Google Scholar] [CrossRef] [PubMed]
- Frantz, S. Studies reveal potential pitfalls of RNAi. Nature Reviews Drug Discovery 2003, 2, 763–765. [Google Scholar] [CrossRef]
- Kim, H.; Kim, J.-S. A guide to genome engineering with programmable nucleases. Nature Reviews Genetics 2014, 15, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Gaj, T.; Gersbach, C.A.; Barbas, C.F. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends in biotechnology 2013, 31, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Schatz, M.C.; Phillippy, A.M. The rise of a digital immune system. Gigascience 2012, 1, 2047-2217X-2041-2044. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; La Russa, M.; Qi, L.S. CRISPR/Cas9 in genome editing and beyond. Annual review of biochemistry 2016, 85, 227–264. [Google Scholar] [CrossRef] [PubMed]
- Stark, J.M.; Pierce, A.J.; Oh, J.; Pastink, A.; Jasin, M. Genetic steps of mammalian homologous repair with distinct mutagenic consequences. Molecular and cellular biology 2004. [Google Scholar] [CrossRef] [PubMed]
- Saunders, G.; Cooke, B.; McColl, K.; Shine, R.; Peacock, T. Modern approaches for the biological control of vertebrate pests: an Australian perspective. Biological control 2010, 52, 288–295. [Google Scholar] [CrossRef]
- Scharenberg, A.M.; Stoddard, B.L.; Monnat, R.J.; Nolan, A. Retargeting: an unrecognized consideration in endonuclease-based gene drive biology. bioRxiv 2016, 089946. [Google Scholar]
- Stoddard, B.L. Homing endonuclease structure and function. Quarterly reviews of biophysics 2005, 38, 49–95. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-G.; Cha, J.; Chandrasegaran, S. Hybrid restriction enzymes: zinc finger fusions to Fok I cleavage domain. Proceedings of the National Academy of Sciences 1996, 93, 1156–1160. [Google Scholar] [CrossRef]
- Boch, J.; Scholze, H.; Schornack, S.; Landgraf, A.; Hahn, S.; Kay, S.; Lahaye, T.; Nickstadt, A.; Bonas, U. Breaking the code of DNA binding specificity of TAL-type III effectors. Science 2009, 326, 1509–1512. [Google Scholar] [CrossRef]
- Jiang, W.; Bikard, D.; Cox, D.; Zhang, F.; Marraffini, L.A. RNA-guided editing of bacterial genomes using CRISPR-Cas systems. Nature biotechnology 2013, 31, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Davis, G. Mobile group II intron targeting: applications in prokaryotes and perspectives in eukaryotes. Front Biosci 2007, 12, 4972–4985. [Google Scholar] [CrossRef]
- You, K.T.; Li, L.S.; Kim, N.-G.; Kang, H.J.; Koh, K.H.; Chwae, Y.-J.; Kim, K.M.; Kim, Y.K.; Park, S.M.; Jang, S.K. Selective translational repression of truncated proteins from frameshift mutation-derived mRNAs in tumors. PLoS biology 2007, 5, e109. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Staahl, B.T.; Alla, R.K.; Doudna, J.A. Enhanced homology-directed human genome engineering by controlled timing of CRISPR/Cas9 delivery. elife 2014, 3, e04766. [Google Scholar] [CrossRef]
- Szostak, J.W.; Orr-Weaver, T.L.; Rothstein, R.J.; Stahl, F.W. The double-strand-break repair model for recombination. Cell 1983, 33, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Rouet, P.; Smih, F.; Jasin, M. Expression of a site-specific endonuclease stimulates homologous recombination in mammalian cells. Proceedings of the National Academy of Sciences 1994, 91, 6064–6068. [Google Scholar] [CrossRef] [PubMed]
- Lieber, M.R.; Ma, Y.; Pannicke, U.; Schwarz, K. Mechanism and regulation of human non-homologous DNA end-joining. Nature reviews Molecular cell biology 2003, 4, 712–720. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Noyes, M.B.; Zhu, L.J.; Lawson, N.D.; Wolfe, S.A. Targeted gene inactivation in zebrafish using engineered zinc-finger nucleases. Nature biotechnology 2008, 26, 695–701. [Google Scholar] [CrossRef]
- Ciccia, A.; Elledge, S.J. The DNA damage response: making it safe to play with knives. Molecular cell 2010, 40, 179–204. [Google Scholar] [CrossRef] [PubMed]
- Ousterout, D.G.; Perez-Pinera, P.; Thakore, P.I.; Kabadi, A.M.; Brown, M.T.; Qin, X.; Fedrigo, O.; Mouly, V.; Tremblay, J.P.; Gersbach, C.A. Reading frame correction by targeted genome editing restores dystrophin expression in cells from Duchenne muscular dystrophy patients. Molecular Therapy 2013, 21, 1718–1726. [Google Scholar] [CrossRef]
- Tebas, P.; Stein, D.; Tang, W.W.; Frank, I.; Wang, S.Q.; Lee, G.; Spratt, S.K.; Surosky, R.T.; Giedlin, M.A.; Nichol, G. Gene editing of CCR5 in autologous CD4 T cells of persons infected with HIV. New England Journal of Medicine 2014, 370, 901–910. [Google Scholar] [CrossRef]
- Lee, H.J.; Kim, E.; Kim, J.-S. Targeted chromosomal deletions in human cells using zinc finger nucleases. Genome research 2010, 20, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Bauer, D.E.; Kamran, S.C.; Lessard, S.; Xu, J.; Fujiwara, Y.; Lin, C.; Shao, Z.; Canver, M.C.; Smith, E.C.; Pinello, L. An erythroid enhancer of BCL11A subject to genetic variation determines fetal hemoglobin level. Science 2013, 342, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Choulika, A.; Perrin, A.; Dujon, B.; Nicolas, J.-F. Induction of homologous recombination in mammalian chromosomes by using the I-SceI system of Saccharomyces cerevisiae. Molecular and cellular biology 1995. [Google Scholar] [CrossRef] [PubMed]
- Urnov, F.D.; Miller, J.C.; Lee, Y.-L.; Beausejour, C.M.; Rock, J.M.; Augustus, S.; Jamieson, A.C.; Porteus, M.H.; Gregory, P.D.; Holmes, M.C. Highly efficient endogenous human gene correction using designed zinc-finger nucleases. Nature 2005, 435, 646–651. [Google Scholar] [CrossRef]
- Chen, F.; Pruett-Miller, S.M.; Huang, Y.; Gjoka, M.; Duda, K.; Taunton, J.; Collingwood, T.N.; Frodin, M.; Davis, G.D. High-frequency genome editing using ssDNA oligonucleotides with zinc-finger nucleases. Nature methods 2011, 8, 753–755. [Google Scholar] [CrossRef] [PubMed]
- Händel, E.-M.; Gellhaus, K.; Khan, K.; Bednarski, C.; Cornu, T.I.; Müller-Lerch, F.; Kotin, R.M.; Heilbronn, R.; Cathomen, T. Versatile and efficient genome editing in human cells by combining zinc-finger nucleases with adeno-associated viral vectors. Human gene therapy 2012, 23, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Hacein-Bey-Abina, S.; Von Kalle, C.; Schmidt, M.; McCormack, M.; Wulffraat, N.; Leboulch, P.a.; Lim, A.; Osborne, C.; Pawliuk, R.; Morillon, E. LMO2-associated clonal T cell proliferation in two patients after gene therapy for SCID-X1. science 2003, 302, 415–419. [Google Scholar] [CrossRef]
- Moehle, E.A.; Rock, J.M.; Lee, Y.-L.; Jouvenot, Y.; DeKelver, R.C.; Gregory, P.D.; Urnov, F.D.; Holmes, M.C. Targeted gene addition into a specified location in the human genome using designed zinc finger nucleases. Proceedings of the National Academy of Sciences 2007, 104, 3055–3060. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Haurigot, V.; Doyon, Y.; Li, T.; Wong, S.Y.; Bhagwat, A.S.; Malani, N.; Anguela, X.M.; Sharma, R.; Ivanciu, L. In vivo genome editing restores haemostasis in a mouse model of haemophilia. Nature 2011, 475, 217–221. [Google Scholar] [CrossRef]
- Algasaier, S.I.; Exell, J.C.; Bennet, I.A.; Thompson, M.J.; Gotham, V.J.; Shaw, S.J.; Craggs, T.D.; Finger, L.D.; Grasby, J.A. DNA and protein requirements for substrate conformational changes necessary for human flap endonuclease-1-catalyzed reaction. Journal of Biological Chemistry 2016, 291, 8258–8268. [Google Scholar] [CrossRef] [PubMed]
- Tupler, R.; Perini, G.; Green, M.R. Expressing the human genome. Nature 2001, 409, 832–833. [Google Scholar] [CrossRef]
- Pavletich, N.P.; Pabo, C.O. Zinc finger-DNA recognition: crystal structure of a Zif268-DNA complex at 2.1 Å. Science 1991, 252, 809–817. [Google Scholar] [CrossRef]
- Miller, J.C.; Holmes, M.C.; Wang, J.; Guschin, D.Y.; Lee, Y.-L.; Rupniewski, I.; Beausejour, C.M.; Waite, A.J.; Wang, N.S.; Kim, K.A. An improved zinc-finger nuclease architecture for highly specific genome editing. Nature biotechnology 2007, 25, 778–785. [Google Scholar] [CrossRef] [PubMed]
- Szczepek, M.; Brondani, V.; Büchel, J.; Serrano, L.; Segal, D.J.; Cathomen, T. Structure-based redesign of the dimerization interface reduces the toxicity of zinc-finger nucleases. Nature biotechnology 2007, 25, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Gersbach, C.A.; Gaj, T.; Barbas III, C.F. Synthetic zinc finger proteins: the advent of targeted gene regulation and genome modification technologies. Accounts of chemical research 2014, 47, 2309–2318. [Google Scholar] [CrossRef]
- Didigu, C.A.; Wilen, C.B.; Wang, J.; Duong, J.; Secreto, A.J.; Danet-Desnoyers, G.A.; Riley, J.L.; Gregory, P.D.; June, C.H.; Holmes, M.C. Simultaneous zinc-finger nuclease editing of the HIV coreceptors ccr5 and cxcr4 protects CD4+ T cells from HIV-1 infection. Blood, The Journal of the American Society of Hematology 2014, 123, 61–69. [Google Scholar] [CrossRef]
- Pattanayak, V.; Ramirez, C.L.; Joung, J.K.; Liu, D.R. Revealing off-target cleavage specificities of zinc-finger nucleases by in vitro selection. Nature methods 2011, 8, 765–770. [Google Scholar] [CrossRef]
- Mino, T.; Mori, T.; Aoyama, Y.; Sera, T. Gene-and protein-delivered zinc finger–staphylococcal nuclease hybrid for inhibition of DNA replication of human papillomavirus. PloS one 2013, 8, e56633. [Google Scholar] [CrossRef]
- Mino, T.; Mori, T.; Aoyama, Y.; Sera, T. Inhibition of DNA replication of human papillomavirus by using zinc finger–single-chain foki dimer hybrid. Molecular biotechnology 2014, 56, 731–737. [Google Scholar] [CrossRef]
- Li, L.; Wu, L.P.; Chandrasegaran, S. Functional domains in Fok I restriction endonuclease. Proceedings of the National Academy of Sciences 1992, 89, 4275–4279. [Google Scholar] [CrossRef] [PubMed]
- Porteus, M.H.; Baltimore, D. Chimeric nucleases stimulate gene targeting in human cells. Science 2003, 300, 763–763. [Google Scholar] [CrossRef] [PubMed]
- Hockemeyer, D.; Soldner, F.; Beard, C.; Gao, Q.; Mitalipova, M.; DeKelver, R.C.; Katibah, G.E.; Amora, R.; Boydston, E.A.; Zeitler, B. Efficient targeting of expressed and silent genes in human ESCs and iPSCs using zinc-finger nucleases. Nature biotechnology 2009, 27, 851–857. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.C.; Tan, S.; Qiao, G.; Barlow, K.A.; Wang, J.; Xia, D.F.; Meng, X.; Paschon, D.E.; Leung, E.; Hinkley, S.J. A TALE nuclease architecture for efficient genome editing. Nature biotechnology 2011, 29, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Deng, D.; Yan, C.; Pan, X.; Mahfouz, M.; Wang, J.; Zhu, J.-K.; Shi, Y.; Yan, N. Structural basis for sequence-specific recognition of DNA by TAL effectors. Science 2012, 335, 720–723. [Google Scholar] [CrossRef]
- Kim, Y.; Kweon, J.; Kim, J.-S. TALENs and ZFNs are associated with different mutation signatures. Nature methods 2013, 10, 185–185. [Google Scholar] [CrossRef]
- Kleinstiver, B.P.; Wang, L.; Wolfs, J.M.; Kolaczyk, T.; McDowell, B.; Wang, X.; Schild-Poulter, C.; Bogdanove, A.J.; Edgell, D.R. The I-TevI nuclease and linker domains contribute to the specificity of monomeric TALENs. G3: Genes, Genomes, Genetics 2014, 4, 1155–1165. [Google Scholar] [CrossRef] [PubMed]
- Guilinger, J.P.; Thompson, D.B.; Liu, D.R. Fusion of catalytically inactive Cas9 to FokI nuclease improves the specificity of genome modification. Nature biotechnology 2014, 32, 577–582. [Google Scholar] [CrossRef] [PubMed]
- Cermak, T.; Doyle, E.L.; Christian, M.; Wang, L.; Zhang, Y.; Schmidt, C.; Baller, J.A.; Somia, N.V.; Bogdanove, A.J.; Voytas, D.F. Efficient design and assembly of custom TALEN and other TAL effector-based constructs for DNA targeting. Nucleic acids research 2011, 39, e82–e82. [Google Scholar] [CrossRef] [PubMed]
- Holkers, M.; Maggio, I.; Liu, J.; Janssen, J.M.; Miselli, F.; Mussolino, C.; Recchia, A.; Cathomen, T.; Goncalves, M.A. Differential integrity of TALE nuclease genes following adenoviral and lentiviral vector gene transfer into human cells. Nucleic acids research 2013, 41, e63–e63. [Google Scholar] [CrossRef]
- Cai, Y.; Bak, R.O.; Mikkelsen, J.G. Targeted genome editing by lentiviral protein transduction of zinc-finger and TAL-effector nucleases. Elife 2014, 3, e01911. [Google Scholar] [CrossRef] [PubMed]
- Horvath, P.; Barrangou, R. CRISPR/Cas, the immune system of bacteria and archaea. Science 2010, 327, 167–170. [Google Scholar] [CrossRef]
- Sorek, R.; Lawrence, C.M.; Wiedenheft, B. CRISPR-mediated adaptive immune systems in bacteria and archaea. Annual review of biochemistry 2013, 82, 237–266. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA–guided DNA endonuclease in adaptive bacterial immunity. science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Anders, C.; Niewoehner, O.; Duerst, A.; Jinek, M. Structural basis of PAM-dependent target DNA recognition by the Cas9 endonuclease. Nature 2014, 513, 569–573. [Google Scholar] [CrossRef]
- Heigwer, F.; Kerr, G.; Boutros, M. E-CRISP: fast CRISPR target site identification. Nature methods 2014, 11, 122–123. [Google Scholar] [CrossRef]
- Fu, Y.; Foden, J.A.; Khayter, C.; Maeder, M.L.; Reyon, D.; Joung, J.K.; Sander, J.D. High-frequency off-target mutagenesis induced by CRISPR-Cas nucleases in human cells. Nature biotechnology 2013, 31, 822–826. [Google Scholar] [CrossRef]
- Fu, Y.; Sander, J.D.; Reyon, D.; Cascio, V.M.; Joung, J.K. Improving CRISPR-Cas nuclease specificity using truncated guide RNAs. Nature biotechnology 2014, 32, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Davis, K.M.; Pattanayak, V.; Thompson, D.B.; Zuris, J.A.; Liu, D.R. Small molecule–triggered Cas9 protein with improved genome-editing specificity. Nature chemical biology 2015, 11, 316–318. [Google Scholar] [CrossRef] [PubMed]
- Schumann, K.; Lin, S.; Boyer, E.; Simeonov, D.R.; Subramaniam, M.; Gate, R.E.; Haliburton, G.E.; Ye, C.J.; Bluestone, J.A.; Doudna, J.A. Generation of knock-in primary human T cells using Cas9 ribonucleoproteins. Proceedings of the National Academy of Sciences 2015, 112, 10437–10442. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Gaj, T.; Yang, Y.; Wang, N.; Shui, S.; Kim, S.; Kanchiswamy, C.N.; Kim, J.-S.; Barbas III, C.F. Efficient delivery of nuclease proteins for genome editing in human stem cells and primary cells. Nature protocols 2015, 10, 1842–1859. [Google Scholar] [CrossRef]
- Paix, A.; Folkmann, A.; Rasoloson, D.; Seydoux, G. High efficiency, homology-directed genome editing in Caenorhabditis elegans using CRISPR-Cas9 ribonucleoprotein complexes. Genetics 2015, 201, 47–54. [Google Scholar] [CrossRef]
- Ménoret, S.; De Cian, A.; Tesson, L.; Remy, S.; Usal, C.; Boulé, J.-B.; Boix, C.; Fontanière, S.; Crénéguy, A.; Nguyen, T.H. Homology-directed repair in rodent zygotes using Cas9 and TALEN engineered proteins. Scientific reports 2015, 5, 14410. [Google Scholar] [CrossRef] [PubMed]
- Rahdar, M.; McMahon, M.A.; Prakash, T.P.; Swayze, E.E.; Bennett, C.F.; Cleveland, D.W. Synthetic CRISPR RNA-Cas9–guided genome editing in human cells. Proceedings of the National Academy of Sciences 2015, 112, E7110–E7117. [Google Scholar] [CrossRef]
- Mullineux, S.-T.; Costa, M.; Bassi, G.S.; Michel, F.; Hausner, G. A group II intron encodes a functional LAGLIDADG homing endonuclease and self-splices under moderate temperature and ionic conditions. Rna 2010, 16, 1818–1831. [Google Scholar] [CrossRef]
- Haugen, P.; Bhattacharya, D. The spread of LAGLIDADG homing endonuclease genes in rDNA. Nucleic acids research 2004, 32, 2049–2057. [Google Scholar] [CrossRef]
- Moure, C.M.; Gimble, F.S.; Quiocho, F.A. Crystal structure of the intein homing endonuclease PI-Sce I bound to its recognition sequence. Nature structural biology 2002, 9, 764–770. [Google Scholar] [CrossRef] [PubMed]
- Butler, G.; Kenny, C.; Fagan, A.; Kurischko, C.; Gaillardin, C.; Wolfe, K.H. Evolution of the MAT locus and its Ho endonuclease in yeast species. Proceedings of the National Academy of Sciences 2004, 101, 1632–1637. [Google Scholar] [CrossRef] [PubMed]
- Dunin-Horkawicz, S.; Feder, M.; Bujnicki, J.M. Phylogenomic analysis of the GIY-YIG nuclease superfamily. BMC genomics 2006, 7, 1–19. [Google Scholar] [CrossRef]
- Cascales, E.; Buchanan, S.K.; Duché, D.; Kleanthous, C.; Lloubes, R.; Postle, K.; Riley, M.; Slatin, S.; Cavard, D. Colicin biology. Microbiology and molecular biology reviews 2007, 71, 158–229. [Google Scholar] [CrossRef] [PubMed]
- Lambowitz, A.M.; Zimmerly, S. Mobile group II introns. Annu. Rev. Genet. 2004, 38, 1–35. [Google Scholar] [CrossRef]
- Kosuri, S.; Church, G.M. Large-scale de novo DNA synthesis: technologies and applications. Nature methods 2014, 11, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.; Berdis, A.J. Non-natural nucleotides as probes for the mechanism and fidelity of DNA polymerases. Biochimica et Biophysica Acta (BBA)-Proteins and Proteomics 2010, 1804, 1064–1080. [Google Scholar] [CrossRef] [PubMed]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A. Multiplex genome engineering using CRISPR/Cas systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Ran, F.; Hsu, P.D.; Wright, J.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome engineering using the CRISPR-Cas9 system. Nature protocols 2013, 8, 2281–2308. [Google Scholar] [CrossRef] [PubMed]
- Mohsen, M.G.; Kool, E.T. The discovery of rolling circle amplification and rolling circle transcription. Accounts of chemical research 2016, 49, 2540–2550. [Google Scholar] [CrossRef]
- Stemmer, W.P.; Crameri, A.; Ha, K.D.; Brennan, T.M.; Heyneker, H.L. Single-step assembly of a gene and entire plasmid from large numbers of oligodeoxyribonucleotides. Gene 1995, 164, 49–53. [Google Scholar] [CrossRef]
- Sarac, I.; Hollenstein, M. Terminal deoxynucleotidyl transferase in the synthesis and modification of nucleic acids. ChemBioChem 2019, 20, 860–871. [Google Scholar] [CrossRef]
- Caruthers, M.; Barone, A.; Beaucage, S.; Dodds, D.; Fisher, E.; McBride, L.; Matteucci, M.; Stabinsky, Z.; Tang, J.-Y. [15] Chemical synthesis of deoxyoligonucleotides by the phosphoramidite method. In Methods in enzymology; Elsevier, 1987; Volume 154, pp. 287–313. [Google Scholar]
- Caruthers, M.H. The chemical synthesis of DNA/RNA: our gift to science. Journal of biological chemistry 2013, 288, 1420–1427. [Google Scholar] [CrossRef]
- LeProust, E.M.; Peck, B.J.; Spirin, K.; McCuen, H.B.; Moore, B.; Namsaraev, E.; Caruthers, M.H. Synthesis of high-quality libraries of long (150mer) oligonucleotides by a novel depurination controlled process. Nucleic acids research 2010, 38, 2522–2540. [Google Scholar] [CrossRef]
- Cleary, M.A.; Kilian, K.; Wang, Y.; Bradshaw, J.; Cavet, G.; Ge, W.; Kulkarni, A.; Paddison, P.J.; Chang, K.; Sheth, N. Production of complex nucleic acid libraries using highly parallel in situ oligonucleotide synthesis. Nature methods 2004, 1, 241–248. [Google Scholar] [CrossRef]
- Veneziano, R.; Shepherd, T.R.; Ratanalert, S.; Bellou, L.; Tao, C.; Bathe, M. In vitro synthesis of gene-length single-stranded DNA. Scientific reports 2018, 8, 6548. [Google Scholar] [CrossRef]
- Jensen, M.A.; Davis, R.W. Template-independent enzymatic oligonucleotide synthesis (TiEOS): its history, prospects, and challenges. Biochemistry 2018, 57, 1821–1832. [Google Scholar] [CrossRef]
- Boulé, J.-B.; Rougeon, F.; Papanicolaou, C. Terminal deoxynucleotidyl transferase indiscriminately incorporates ribonucleotides and deoxyribonucleotides. Journal of Biological Chemistry 2001, 276, 31388–31393. [Google Scholar] [CrossRef] [PubMed]
- Hoose, A.; Vellacott, R.; Storch, M.; Freemont, P.S.; Ryadnov, M.G. DNA synthesis technologies to close the gene writing gap. Nature Reviews Chemistry 2023, 7, 144–161. [Google Scholar] [CrossRef]
- Loftie-Eaton, W.; Heinisch, T.; Soskine, M.; Champion, E.; Godron, X.; Ybert, T. Novel Variants of Endonuclease V and Uses Thereof. 2023.
- Motea, E.A.; Berdis, A.J. Terminal deoxynucleotidyl transferase: the story of a misguided DNA polymerase. Biochimica et Biophysica Acta (BBA)-Proteins and Proteomics 2010, 1804, 1151–1166. [Google Scholar] [CrossRef] [PubMed]
- Breitwieser, F.P.; Lu, J.; Salzberg, S.L. A review of methods and databases for metagenomic classification and assembly. Briefings in bioinformatics 2019, 20, 1125–1136. [Google Scholar] [CrossRef]
- Kakirde, K.S.; Wild, J.; Godiska, R.; Mead, D.A.; Wiggins, A.G.; Goodman, R.M.; Szybalski, W.; Liles, M.R. Gram negative shuttle BAC vector for heterologous expression of metagenomic libraries. Gene 2011, 475, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.; Gilbert, J.; Meyer, F. Metagenomics-a guide from sampling to data analysis. Microbial informatics and experimentation 2012, 2, 1–12. [Google Scholar] [CrossRef]
- Glenn, T.C. Field guide to next-generation DNA sequencers. Molecular ecology resources 2011, 11, 759–769. [Google Scholar] [CrossRef]
- Liu, L.; Li, Y.; Li, S.; Hu, N.; He, Y.; Pong, R.; Lin, D.; Lu, L.; Law, M. Comparison of next-generation sequencing systems. BioMed research international 2012, 2012, 251364. [Google Scholar]
- Fantini, E.; Gianese, G.; Giuliano, G.; Fiore, A. Bacterial metabarcoding by 16S rRNA gene ion torrent amplicon sequencing. Bacterial Pangenomics: Methods and Protocols 2015, 77–90. [Google Scholar]
- Marx, V. Method of the year: long-read sequencing. Nature Methods 2023, 20, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Zhang, S.; Xie, Q.; Wang, W.; Lin, Z.; Wang, H.; Yuan, Y.; Chen, Q. De Novo Transcriptome Analysis by PacBio SMRT-Seq and Illumina RNA-Seq Provides New Insights into Polyphenol Biosynthesis in Chinese Olive Fruit. Horticulturae 2024, 10, 293. [Google Scholar] [CrossRef]
- Kong, N.; Ng, W.; Thao, K.; Agulto, R.; Weis, A.; Kim, K.S.; Korlach, J.; Hickey, L.; Kelly, L.; Lappin, S. Automation of PacBio SMRTbell NGS library preparation for bacterial genome sequencing. Standards in genomic sciences 2017, 12, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Cuber, P.; Chooneea, D.; Geeves, C.; Salatino, S.; Creedy, T.J.; Griffin, C.; Sivess, L.; Barnes, I.; Price, B.; Misra, R. Comparing the accuracy and efficiency of third generation sequencing technologies, Oxford Nanopore Technologies, and Pacific Biosciences, for DNA barcode sequencing applications. Ecological Genetics and Genomics 2023, 28, 100181. [Google Scholar] [CrossRef]
- Xu, Y.; Luo, H.; Wang, Z.; Lam, H.-M.; Huang, C. Oxford Nanopore Technology: revolutionizing genomics research in plants. Trends in Plant Science 2022, 27, 510–511. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Fiddes, I.T.; Miga, K.H.; Olsen, H.E.; Paten, B.; Akeson, M. Improved data analysis for the MinION nanopore sequencer. Nature methods 2015, 12, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Greninger, A.L.; Naccache, S.N.; Federman, S.; Yu, G.; Mbala, P.; Bres, V.; Stryke, D.; Bouquet, J.; Somasekar, S.; Linnen, J.M. Rapid metagenomic identification of viral pathogens in clinical samples by real-time nanopore sequencing analysis. Genome medicine 2015, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhao, Y.; Huang, C.; Luo, Y. Recent advances in silent gene cluster activation in Streptomyces. Frontiers in Bioengineering and Biotechnology 2021, 9, 632230. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Augustijn, H.E.; Reitz, Z.L.; Biermann, F.; Alanjary, M.; Fetter, A.; Terlouw, B.R.; Metcalf, W.W.; Helfrich, E.J. antiSMASH 7.0: new and improved predictions for detection, regulation, chemical structures and visualisation. Nucleic acids research 2023, 51, W46–W50. [Google Scholar] [CrossRef]
- Mózsik, L.; Iacovelli, R.; Bovenberg, R.A.; Driessen, A.J. Transcriptional activation of biosynthetic gene clusters in filamentous fungi. Frontiers in Bioengineering and Biotechnology 2022, 10, 901037. [Google Scholar] [CrossRef]
- Cook, G.D.; Stasulli, N.M. Employing synthetic biology to expand antibiotic discovery. SLAS technology 2024, 29, 100120. [Google Scholar] [CrossRef]
- Zhao, N.; Song, Y.; Xie, X.; Zhu, Z.; Duan, C.; Nong, C.; Wang, H.; Bao, R. Synthetic biology-inspired cell engineering in diagnosis, treatment, and drug development. Signal Transduction and Targeted Therapy 2023, 8, 112. [Google Scholar] [CrossRef]
- Kim, E.; Moore, B.S.; Yoon, Y.J. Reinvigorating natural product combinatorial biosynthesis with synthetic biology. Nature chemical biology 2015, 11, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Tsiamis, G.; Cherif, A.; Karpouzas, D.; Ntougias, S. Microbial diversity for biotechnology 2014. BioMed Research International 2015, 2015. [Google Scholar] [CrossRef]
- Voigt, C.A. Synthetic biology. 2012, 1, 1-2.
- Katz, L.; Chen, Y.Y.; Gonzalez, R.; Peterson, T.C.; Zhao, H.; Baltz, R.H. Synthetic biology advances and applications in the biotechnology industry: a perspective. Journal of Industrial Microbiology and Biotechnology 2018, 45, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-H.P.; Sun, J.; Ma, Y. Biomanufacturing: history and perspective. Journal of Industrial Microbiology and Biotechnology 2017, 44, 773–784. [Google Scholar] [CrossRef]
- Coughlan, L.M.; Cotter, P.D.; Hill, C.; Alvarez-Ordóñez, A. Biotechnological applications of functional metagenomics in the food and pharmaceutical industries. Frontiers in microbiology 2015, 6, 672. [Google Scholar] [CrossRef] [PubMed]
- Endy, D. Foundations for engineering biology. Nature 2005, 438, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Bradley, R.W.; Buck, M.; Wang, B. Tools and principles for microbial gene circuit engineering. Journal of Molecular Biology 2016, 428, 862–888. [Google Scholar] [CrossRef]
- Müller, K.M.; Arndt, K.M. Standardization in synthetic biology. Synthetic gene networks: methods and protocols 2012, 23–43. [Google Scholar]
- Bouhajja, E.; Agathos, S.N.; George, I.F. Metagenomics: probing pollutant fate in natural and engineered ecosystems. Biotechnology advances 2016, 34, 1413–1426. [Google Scholar] [CrossRef]
- Tseng, C.-H.; Tang, S.-L. Marine microbial metagenomics: from individual to the environment. International Journal of Molecular Sciences 2014, 15, 8878–8892. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.L.; Piel, J.; Sunagawa, S. A roadmap for metagenomic enzyme discovery. Natural Product Reports 2021, 38, 1994–2023. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Ellis, T. The second decade of synthetic biology: 2010–2020. Nature Communications 2020, 11, 5174. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Huang, C.; Chen, J. The application of CRISPR/Cas mediated gene editing in synthetic biology: Challenges and optimizations. Frontiers in Bioengineering and Biotechnology 2022, 10, 890155. [Google Scholar] [CrossRef] [PubMed]
- Forsberg, K.J.; Bhatt, I.V.; Schmidtke, D.T.; Javanmardi, K.; Dillard, K.E.; Stoddard, B.L.; Finkelstein, I.J.; Kaiser, B.K.; Malik, H.S. Functional metagenomics-guided discovery of potent Cas9 inhibitors in the human microbiome. elife 2019, 8, e46540. [Google Scholar] [CrossRef]
- Hughes, R.A.; Ellington, A.D. Synthetic DNA synthesis and assembly: putting the synthetic in synthetic biology. Cold Spring Harbor perspectives in biology 2017, 9, a023812. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Yin, Y.; Zhu, L.; Zhang, Y.; Li, Y.-Z. Metagenomic sequencing-driven multidisciplinary approaches to shed light on the untapped microbial natural products. Drug Discovery Today 2022, 27, 730–742. [Google Scholar] [CrossRef]
- Mann, D.L. Synthetic Biology, Directed Evolution, and the Rational Design of New Cardiovascular Therapeutics: Are We There Yet? 2023, 8, 905–906. [Google Scholar] [PubMed]
- Suenaga, H. Targeted metagenomics unveils the molecular basis for adaptive evolution of enzymes to their environment. Frontiers in microbiology 2015, 6, 1018. [Google Scholar] [CrossRef]
- Gunjal, A.; Gupta, S.; Nweze, J.E.; Nweze, J.A. Metagenomics in bioremediation: Recent advances, challenges, and perspectives. Metagenomics to Bioremediation 2023, 81–102. [Google Scholar]
- García-Granados, R.; Lerma-Escalera, J.A.; Morones-Ramírez, J.R. Metabolic engineering and synthetic biology: synergies, future, and challenges. Frontiers in bioengineering and biotechnology 2019, 7, 36. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, S.; Shukla, P. Alternative strategies for microbial remediation of pollutants via synthetic biology. Frontiers in microbiology 2020, 11, 808. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Kumar, V.; Dhanjal, D.S.; Datta, S.; Prasad, R.; Singh, J. Biological biosensors for monitoring and diagnosis. Microbial biotechnology: basic research and applications 2020, 317–335. [Google Scholar]
- Bharti, A.; Jain, U.; Chauhan, N. From Lab to Field: Nano-Biosensors for Real-time Plant Nutrient Tracking. Plant Nano Biology 2024, 100079. [Google Scholar] [CrossRef]
- Tanniche, I.; Behkam, B. Engineered live bacteria as disease detection and diagnosis tools. Journal of Biological Engineering 2023, 17, 65. [Google Scholar] [CrossRef]
- Duque, E.; Udaondo, Z.; Molina, L.; de la Torre, J.; Godoy, P.; Ramos, J.L. Providing octane degradation capability to Pseudomonas putida KT2440 through the horizontal acquisition of oct genes located on an integrative and conjugative element. Environmental Microbiology Reports 2022, 14, 934–946. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Chen, J.; Dharmasiddhi, I.P.W.; Chen, S.; Liu, Y.; Liu, H. Review of the Potential of Probiotics in Disease Treatment: Mechanisms, Engineering, and Applications. Processes 2024, 12, 316. [Google Scholar] [CrossRef]
- Archer, E.J.; Robinson, A.B.; Süel, G.r.M. Engineered E. coli that detect and respond to gut inflammation through nitric oxide sensing. ACS synthetic biology 2012, 1, 451–457. [Google Scholar] [CrossRef]
- Riglar, D.T.; Silver, P.A. Engineering bacteria for diagnostic and therapeutic applications. Nature Reviews Microbiology 2018, 16, 214–225. [Google Scholar] [CrossRef] [PubMed]
- Mahdizade Ari, M.; Dadgar, L.; Elahi, Z.; Ghanavati, R.; Taheri, B. Genetically engineered microorganisms and their impact on human health. International Journal of Clinical Practice 2024, 2024, 6638269. [Google Scholar] [CrossRef] [PubMed]
- Rylott, E.L.; Bruce, N.C. How synthetic biology can help bioremediation. Current Opinion in Chemical Biology 2020, 58, 86–95. [Google Scholar] [CrossRef]
- Ermakova, M.; Danila, F.R.; Furbank, R.T.; von Caemmerer, S. On the road to C4 rice: advances and perspectives. The Plant Journal 2020, 101, 940–950. [Google Scholar] [CrossRef]
- Giachino, A.; Focarelli, F.; Marles-Wright, J.; Waldron, K.J. Synthetic biology approaches to copper remediation: bioleaching, accumulation and recycling. FEMS microbiology ecology 2021, 97, fiaa249. [Google Scholar] [CrossRef] [PubMed]
- Dangi, A.K.; Sharma, B.; Hill, R.T.; Shukla, P. Bioremediation through microbes: systems biology and metabolic engineering approach. Critical reviews in biotechnology 2019, 39, 79–98. [Google Scholar] [CrossRef]
- Skariyachan, S.; Taskeen, N.; Kishore, A.P.; Krishna, B.V.; Naidu, G. Novel consortia of Enterobacter and Pseudomonas formulated from cow dung exhibited enhanced biodegradation of polyethylene and polypropylene. Journal of environmental management 2021, 284, 112030. [Google Scholar] [CrossRef] [PubMed]
- Bizimana Rukundo, T. Ecological Restoration Using Synthetic Biology.
- Haque, E.; Bin Riyaz, M.A.; Shankar, S.; Hassan, S. Compositional Characterization of Biosurfactant Produced from Pseudomonas aeruginosa ENO14-MH271625 and its Application in Crude Oil Bioremediation. International Journal of Pharmaceutical Investigation 2021, 11. [Google Scholar] [CrossRef]
- Gao, B.; Yao, H.; Li, Y.; Zhu, Y. Microplastic addition alters the microbial community structure and stimulates soil carbon dioxide emissions in vegetable-growing soil. Environmental Toxicology and Chemistry 2021, 40, 352–365. [Google Scholar] [CrossRef]
- Fisher, M.C.; Henk, D.A.; Briggs, C.J.; Brownstein, J.S.; Madoff, L.C.; McCraw, S.L.; Gurr, S.J. Emerging fungal threats to animal, plant and ecosystem health. Nature 2012, 484, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Duncker, K.E.; Holmes, Z.A.; You, L. Engineered microbial consortia: strategies and applications. Microbial Cell Factories 2021, 20, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Vo, T.P.; Danaee, S.; Chaiwong, C.; Pham, B.T.; Kim, M.; Kuzhiumparambil, U.; Songsomboon, C.; Pernice, M.; Ngo, H.H.; Ralph, P.J. Microalgae-bacteria consortia for organic pollutants remediation from wastewater: A critical review. Journal of Environmental Chemical Engineering 2024, 114213. [Google Scholar] [CrossRef]
- Zitvogel, L.; Ma, Y.; Raoult, D.; Kroemer, G.; Gajewski, T.F. The microbiome in cancer immunotherapy: Diagnostic tools and therapeutic strategies. Science 2018, 359, 1366–1370. [Google Scholar] [CrossRef] [PubMed]
- Lazar, V.; Ditu, L.-M.; Pircalabioru, G.G.; Picu, A.; Petcu, L.; Cucu, N.; Chifiriuc, M.C. Gut microbiota, host organism, and diet trialogue in diabetes and obesity. Frontiers in nutrition 2019, 6, 21. [Google Scholar] [CrossRef]
- Buffie, C.G.; Pamer, E.G. Microbiota-mediated colonization resistance against intestinal pathogens. Nature Reviews Immunology 2013, 13, 790–801. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D. A metagenome-wide association study of gut microbiota in type 2 diabetes. nature 2012, 490, 55–60. [Google Scholar] [CrossRef]
- Control, C.f.D.; Prevention. Antibiotic resistance threats in the United States, 2019; US Department of Health and Human Services, Centres for Disease Control and …: 2019.
- Dang, C.; Xia, Y.; Zheng, M.; Liu, T.; Liu, W.; Chen, Q.; Ni, J. Metagenomic insights into the profile of antibiotic resistomes in a large drinking water reservoir. Environment International 2020, 136, 105449. [Google Scholar] [CrossRef] [PubMed]
- Javed, M.U.; Hayat, M.T.; Mukhtar, H.; Imre, K. CRISPR-Cas9 system: A prospective pathway toward combatting antibiotic resistance. Antibiotics 2023, 12, 1075. [Google Scholar] [CrossRef]
- Loeffler, C.; Gibson, K.M.; Martin, L.; Chang, L.; Rotman, J.; Toma, I.V.; Mason, C.E.; Eskin, E.; Zackular, J.P.; Crandall, K.A. Metagenomics for clinical diagnostics: technologies and informatics. arXiv 2019, arXiv:1911.11304 2019. [Google Scholar]
- Durão, P.; Balbontín, R.; Gordo, I. Evolutionary mechanisms shaping the maintenance of antibiotic resistance. Trends in microbiology 2018, 26, 677–691. [Google Scholar] [CrossRef]
- Niu, B.; Wang, W.; Yuan, Z.; Sederoff, R.R.; Sederoff, H.; Chiang, V.L.; Borriss, R. Microbial interactions within multiple-strain biological control agents impact soil-borne plant disease. Frontiers in Microbiology 2020, 11, 585404. [Google Scholar] [CrossRef] [PubMed]
- Panke-Buisse, K.; Poole, A.C.; Goodrich, J.K.; Ley, R.E.; Kao-Kniffin, J. Selection on soil microbiomes reveals reproducible impacts on plant function. The ISME journal 2015, 9, 980–989. [Google Scholar] [CrossRef]
- Jin, S.; Clark, B.; Kuznesof, S.; Lin, X.; Frewer, L.J. Synthetic biology applied in the agrifood sector: Public perceptions, attitudes and implications for future studies. Trends in Food Science & Technology 2019, 91, 454–466. [Google Scholar]
- Berg, G.; Raaijmakers, J.M. Saving seed microbiomes. The ISME journal 2018, 12, 1167–1170. [Google Scholar] [CrossRef]
- Ray, P.; Lakshmanan, V.; Labbé, J.L.; Craven, K.D. Microbe to microbiome: a paradigm shift in the application of microorganisms for sustainable agriculture. Frontiers in Microbiology 2020, 11, 622926. [Google Scholar] [CrossRef] [PubMed]
- Naylor, D.; Coleman-Derr, D. Drought stress and root-associated bacterial communities. Frontiers in plant science 2018, 8, 2223. [Google Scholar] [CrossRef]
- Cerqueira, F.; Christou, A.; Fatta-Kassinos, D.; Vila-Costa, M.; Bayona, J.M.; Pina, B. Effects of prescription antibiotics on soil-and root-associated microbiomes and resistomes in an agricultural context. Journal of Hazardous Materials 2020, 400, 123208. [Google Scholar] [CrossRef] [PubMed]
- Wright, O.; Stan, G.-B.; Ellis, T. Building-in biosafety for synthetic biology. Microbiology 2013, 159, 1221–1235. [Google Scholar] [CrossRef]
- Jacquier, E.F.; van de Wouw, M.; Nekrasov, E.; Contractor, N.; Kassis, A.; Marcu, D. Local and Systemic Effects of Bioactive Food Ingredients: Is There a Role for Functional Foods to Prime the Gut for Resilience? Foods 2024, 13, 739. [Google Scholar] [CrossRef]
- Wani, S.H.; Kumar, V.; Khare, T.; Guddimalli, R.; Parveda, M.; Solymosi, K.; Suprasanna, P.; Kavi Kishor, P. Engineering salinity tolerance in plants: progress and prospects. Planta 2020, 251, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Jasanoff, S. Technologies of humility: Citizen participation in governing science; Springer, 2005. [Google Scholar]
- Keiper, F.; Atanassova, A. Regulation of synthetic biology: developments under the convention on biological diversity and its protocols. Frontiers in bioengineering and biotechnology 2020, 8, 310. [Google Scholar] [CrossRef] [PubMed]
- Berg, G.; Grube, M.; Schloter, M.; Smalla, K. The plant microbiome and its importance for plant and human health. 2014, 5, 491. [Google Scholar] [CrossRef]
- Bohua, L.; Yuexin, W.; Yakun, O.; Kunlan, Z.; Huan, L.; Ruipeng, L. Ethical framework on risk governance of synthetic biology. Journal of Biosafety and Biosecurity 2023, 5, 45–56. [Google Scholar] [CrossRef]
- Amarelle, V.; Roldán, D.M.; Fabiano, E.; Guazzaroni, M.-E. Synthetic biology toolbox for antarctic Pseudomonas sp. strains: toward a psychrophilic nonmodel chassis for function-driven metagenomics. ACS Synthetic Biology 2023, 12, 722–734. [Google Scholar] [CrossRef] [PubMed]
- Russell, B.J.; Brown, S.D.; Siguenza, N.; Mai, I.; Saran, A.R.; Lingaraju, A.; Maissy, E.S.; Machado, A.C.D.; Pinto, A.F.; Sanchez, C. Intestinal transgene delivery with native E. coli chassis allows persistent physiological changes. Cell 2022, 185, 3263–3277. e3215. [Google Scholar] [CrossRef]
- Belda, I.; Williams, T.C.; de Celis, M.; Paulsen, I.T.; Pretorius, I.S. Seeding the idea of encapsulating a representative synthetic metagenome in a single yeast cell. Nature Communications 2021, 12, 1599. [Google Scholar] [CrossRef]
- Liu, J.; Wang, X.; Dai, G.; Zhang, Y.; Bian, X. Microbial chassis engineering drives heterologous production of complex secondary metabolites. Biotechnology Advances 2022, 59, 107966. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Moon, C.D.; Zheng, N.; Huws, S.; Zhao, S.; Wang, J. Opportunities and challenges of using metagenomic data to bring uncultured microbes into cultivation. Microbiome 2022, 10, 76. [Google Scholar] [CrossRef] [PubMed]
- Lavy, A.; Keren, R.; Haber, M.; Schwartz, I.; Ilan, M. Implementing sponge physiological and genomic information to enhance the diversity of its culturable associated bacteria. FEMS Microbiology Ecology 2014, 87, 486–502. [Google Scholar] [CrossRef] [PubMed]
- M., *!!! REPLACE !!!*; Nishikawa, Y.; Ide, K.; Sakanashi, C.; Takahashi, K.; Takeyama, H. Single-cell genomics of uncultured bacteria reveals dietary fiber responders in the mouse gut microbiota. Microbiome 2020, 8, 1–14. [Google Scholar]
Figure 1.
Some major SynBio landmarks between 2010-2024.
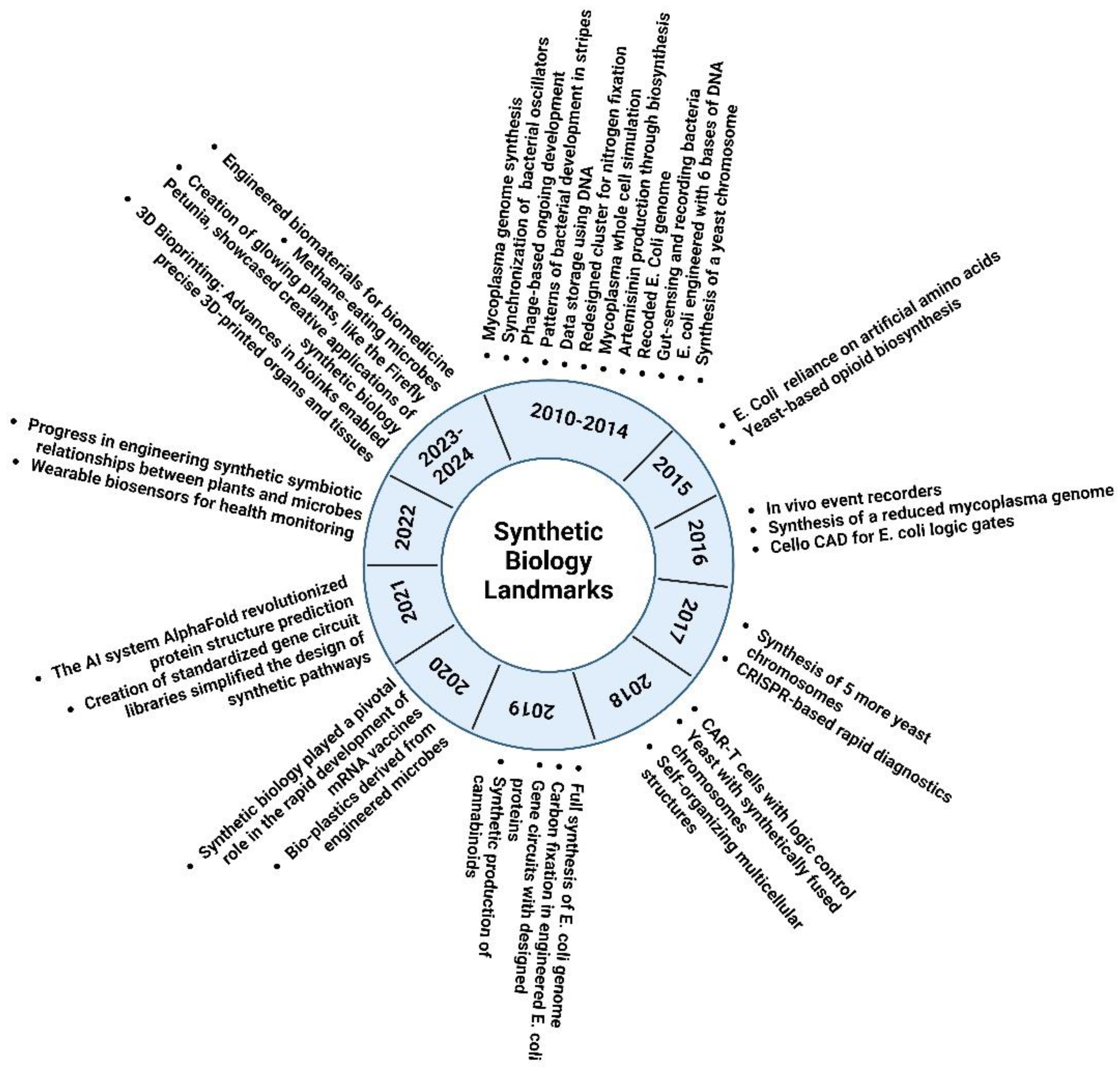
Figure 2.
Mechanism of gene editing via Homology-Directed Repair (HDR) or Non-Homologous End Joining (NHEJ).(a) Zinc Finger Nucleases (ZFNs), (b) Transcription Activator-Like Effector Nucleases (TALENs), (c) CRISPR-Cas9, and (d) Homing Endonucleases create targeted DNA double-strand breaks, repaired by HDR or NHEJ. (e) Synthetic DNA acts as a template for precise HDR-based gene editing.
Figure 2.
Mechanism of gene editing via Homology-Directed Repair (HDR) or Non-Homologous End Joining (NHEJ).(a) Zinc Finger Nucleases (ZFNs), (b) Transcription Activator-Like Effector Nucleases (TALENs), (c) CRISPR-Cas9, and (d) Homing Endonucleases create targeted DNA double-strand breaks, repaired by HDR or NHEJ. (e) Synthetic DNA acts as a template for precise HDR-based gene editing.
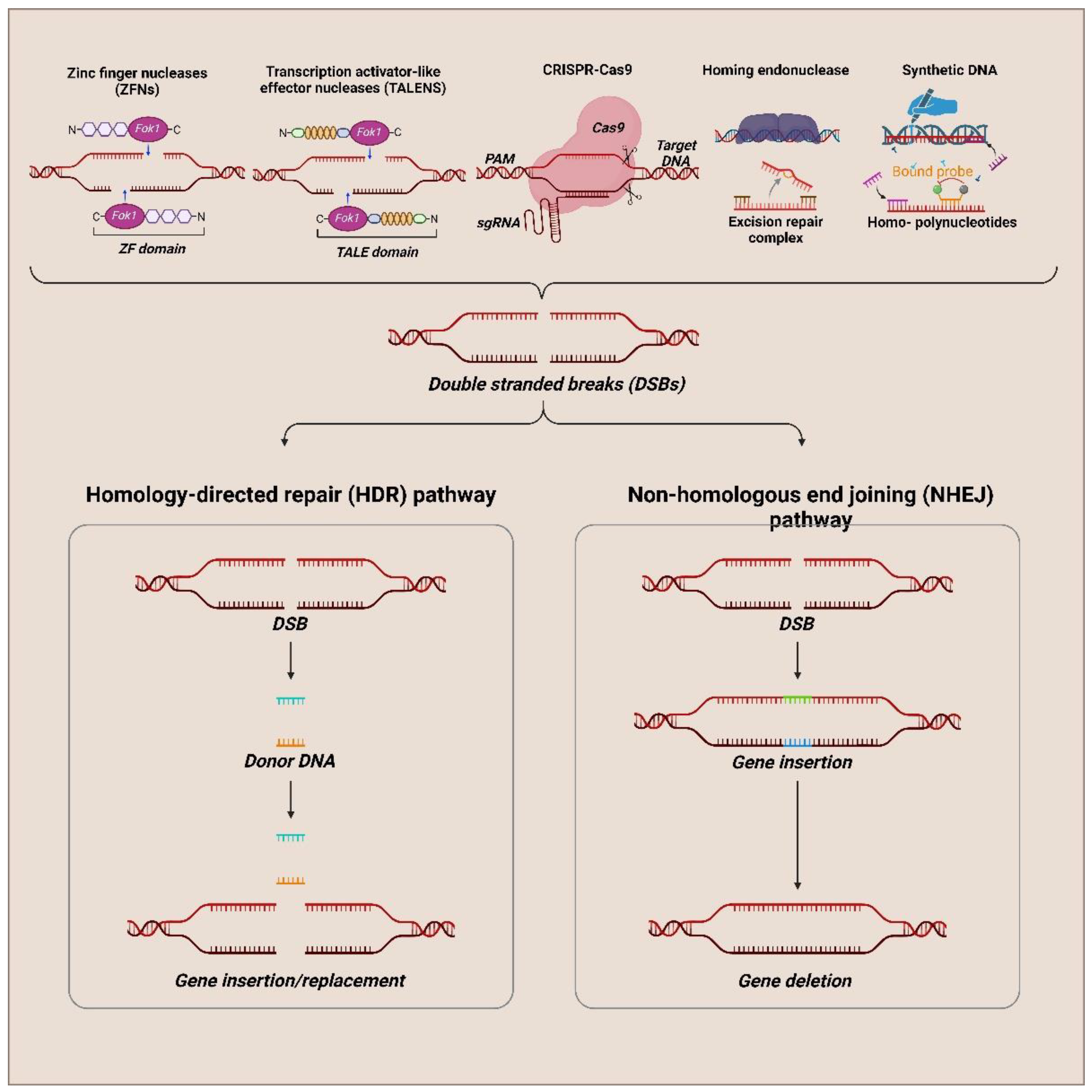
Figure 3.
Workflow of next-generation sequencing (NGS): DNA/RNA extraction, library preparation, sequencing, and bioinformatic analysis for data interpretation.
Figure 3.
Workflow of next-generation sequencing (NGS): DNA/RNA extraction, library preparation, sequencing, and bioinformatic analysis for data interpretation.
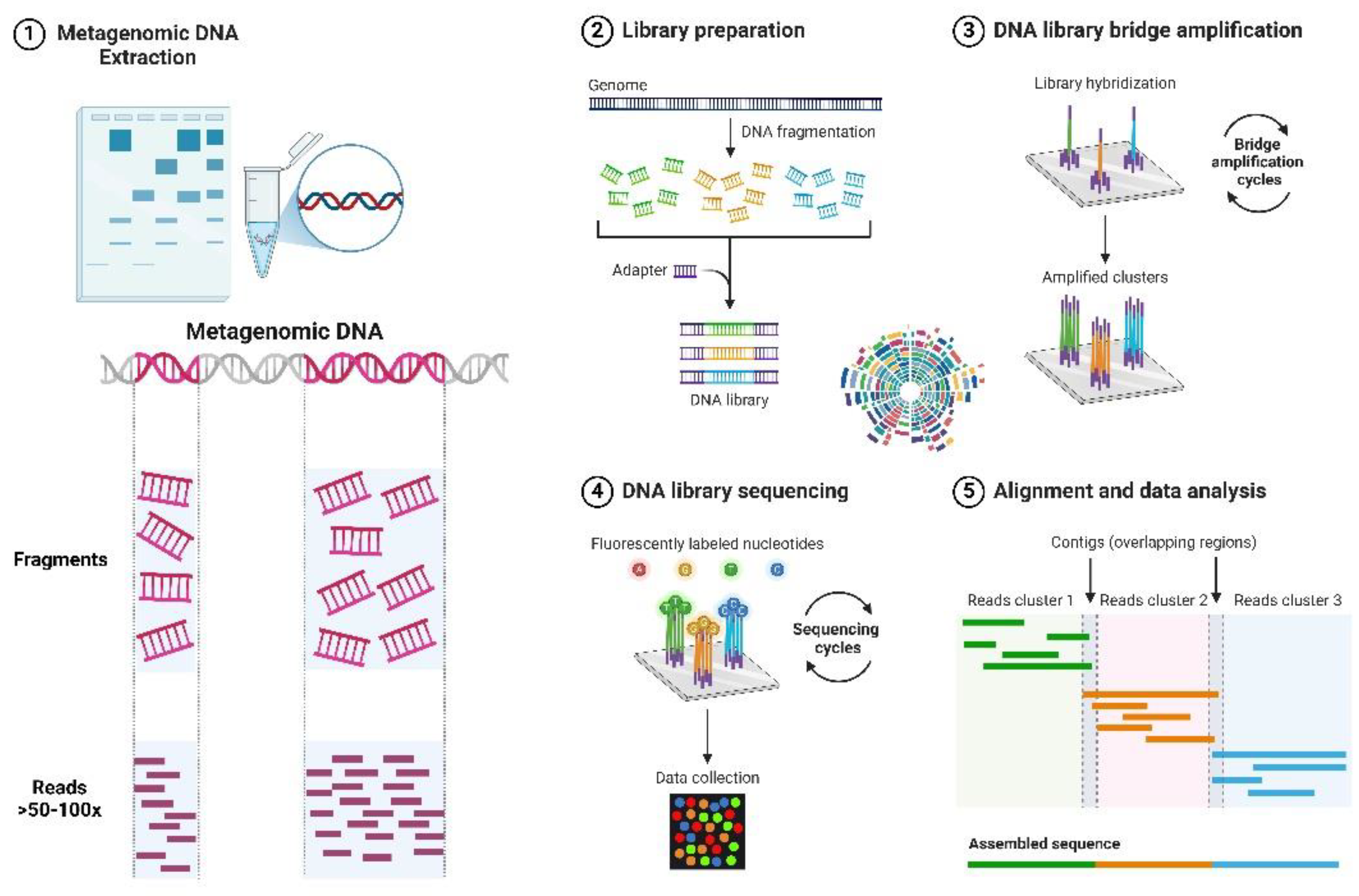
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



