
Submitted:
06 December 2024
Posted:
06 December 2024
You are already at the latest version
Abstract
The human microbiome, a complex ecosystem of microorganisms residing in and on the body, plays a pivotal role in regulating a wide range of physiological processes, including digestion, immune responses, and metabolic functions. In recent years, the rapidly growing field of microbiome-driven therapeutics has garnered significant attention for its potential to revolutionize healthcare. This review explores the evolving landscape of microbiome-based therapies, with a particular focus on the gut microbiome and its implications for both gut health and precision medicine. We highlight recent advances in understanding how microbial communities influence disease pathogenesis and treatment outcomes, spanning conditions such as inflammatory bowel disease (IBD), metabolic disorders, neurological diseases, and even cancer. This article also discusses emerging therapeutic strategies, including probiotics, prebiotics, fecal microbiota transplantation (FMT), and microbial-based drugs, as well as the challenges associated with their clinical implementation. Additionally, we examined how the integration of microbiome profiling and metagenomic data is advancing the field of precision medicine, paving the way for more personalized and effective treatments. This review serves as a comprehensive resource that synthesizes current knowledge, identifies key gaps in microbiome research, and offers insights into the future direction of microbiome-driven therapeutics, thus providing a valuable framework for clinicians, researchers, and policymakers seeking to harness the potential of the microbiome to advance personalized healthcare solutions.
Keywords:
Microbiome-driven therapeutics
; gut microbiome
; precision medicine
; probiotics
; prebiotics
; fecal microbiota transplantation (FMT)
; microbial-based therapies
; personalized healthcare
; microbi-ome profiling
; disease modulation
1. Introduction
Microbiome-driven therapeutics have emerged as a transformative field in medicine, utilizing complex interactions between microbial communities and host biological functions to address various health problems [1]. The human microbiome, often called the “second genome,” is involved in crucial processes such as regulating the immune system, metabolic activities, and maintaining gut homeostasis. Disruption of the microbiome, known as dysbiosis, has been associated with numerous health disorders, including inflammatory bowel disease (IBD), neurological diseases such as autism and Parkinson’s, metabolic conditions such as obesity and diabetes, and certain cancers [2]. Despite significant advances, some notable gaps remain in our understanding of the mechanisms underlying host-microbiome interactions. Understanding these interactions is essential to develop more effective microbiome-based therapies.
Current therapeutic strategies, such as fecal microbiota transplantation (FMT), probiotics, and treatments based on microbial metabolites, have shown varying degrees of success. However, their clinical application is often hindered by factors such as the high interindividual variability of microbiomes, complexity of microbial interactions, and significant gaps in our mechanistic understanding of how these therapies work at the molecular level. Moreover, regulatory hurdles and lack of standardized protocols further complicate their widespread clinical adoption. These challenges underscore the urgent need for advanced research to uncover the underlying mechanisms of host-microbiome interactions and refine microbiome-based interventions for more consistent and predictable clinical outcomes [3,4,5].
One promising direction is the development of personalized microbiome therapies tailored to the specific microbial profiles of individual patients. Precision-based approaches, considering the genetic, environmental, and lifestyle factors that shape the microbiome, could enhance therapeutic efficacy and reduce variability. Although interventions such as FMT and probiotics have demonstrated potential in clinical settings, their inconsistent results highlight the need for more targeted and individualized treatments [6,7]. Another key area of exploration is the engineering of synthetic microbiomes, such as customized microbial consortia, designed to modulate host physiology in a controlled manner. Recent advancements, particularly in CRISPR-based microbiome editing technologies, offer exciting possibilities for precise microbial intervention. However, these technologies are still in their infancy and face challenges related to stability, delivery, and safety that need to be addressed before they can be translated into widespread clinical use [8,9].
The therapeutic potential of the microbiome extends beyond the gut, with emerging research highlighting its role in diverse areas such as cancer immunotherapy, neuroimmune modulation, and the gut-brain axis [10]. These findings underscore the need for interdisciplinary research that integrates microbiology, genomics, immunology, and personalized medicine. However, translating microbiome-based therapies into clinical practice requires the establishment of robust regulatory frameworks and ethical guidelines to ensure patient safety and treatment efficacy [11,12]. Simultaneously, a comprehensive exploration of under-studied microbiome niches, such as those in the skin, oral cavity, and respiratory tract, is essential and holds promise for novel therapeutic applications [12,13]. This review discusses recent developments in the field of microbiome-driven therapeutics, highlights challenges that must be overcome, and proposes future directions for research and clinical implementation. By addressing these gaps and fostering innovative approaches, microbiome-based therapies have the potential to become the cornerstone of precision medicine, offering personalized, effective, and sustainable solutions to a broad spectrum of diseases.
2. Microbiome and Disease Connections
Human microbiome alteration or dysbiosis, which is increasingly associated with various health disorders, ranges from gastrointestinal diseases such as inflammatory bowel disease (IBD) and irritable bowel syndrome (IBS) to systemic diseases such as metabolic syndrome, cardiovascular diseases, neurological diseases, and immune-related diseases. Evidence now points to its role in cancer, especially colorectal cancer, in which some microbial species contribute to inflammation and DNA damage (Figure 1).
2.1. Gastrointestinal Disorders
2.1.1. Irritable Bowel Syndrome (IBS)
IBS is a common functional gastrointestinal disorder with a variety of symptoms, including abdominal pain, bloating, and altered bowel habits such as diarrhea, constipation, or a combination of both. Although the exact cause of IBS remains unclear, several studies have consistently demonstrated that individuals with IBS have distinct changes in their gut microbiota compared with healthy individuals [13,14,15]. A comprehensive meta-analysis revealed a significant reduction in bacterial diversity among IBS patients, particularly a noticeable decrease in beneficial bacteria such as Bifidobacterium and Lactobacillus, which are known to support gut health and maintain microbial balance [16]. This loss of microbial diversity can disrupt the overall harmony of the gut ecosystem, potentially leading to the overgrowth of pathogenic bacteria such as those belonging to the Enterobacteriaceae family [17,18]. These shifts in bacterial composition may worsen IBS symptoms through mechanisms such as increased intestinal permeability (also known as “leaky gut”) and altered immune responses [19]. Elevated levels of pro-inflammatory cytokines in IBS patients trigger inflammation in the gut, leading to pain perception and more severe abdominal discomfort [20,21]. IBS symptomatology is also linked to gut bacterial fermentation of undigested carbohydrates. When these carbohydrates are not properly absorbed, they become substrates for bacterial fermentation, leading to increased gas production and symptoms such as bloating and abdominal distension [22]. In addition to gut dysbiosis and immune dysregulation, IBS is also related to abnormal gut-brain communication [23], and changes in the gut microbiome can affect neurotransmitter levels, potentially leading to emotional disorders in patients with IBS [23].
2.1.2. Inflammatory Bowel Disease (IBD)
IBD is a chronic inflammatory condition affecting the gastrointestinal tract (GIT), primarily in Crohn’s disease (CD) and ulcerative colitis (UC), leading to damage to the intestinal lining [24]. Recent studies have highlighted significant alterations in the gut microbiome of IBD patients, shedding light on its potential role in disease development and progression [25]. One of the most consistent findings is the depletion and imbalance of the beneficial bacterial populations. A study by Sokol et al. (2008) demonstrated that one of the beneficial bacteria, Faecalibacterium prausnitzii is significantly depleted in patients with IBD, particularly those with CD [26]. F. prausnitzii is known for producing short-chain fatty acids (SCFAs), particularly butyrate, which is crucial in maintaining intestinal barrier function and regulating immune responses [27,28]. Its depletion has been linked to increased intestinal permeability, which allows harmful substances and bacteria from the gut lumen to cross the epithelial barrier, leading to immune activation and chronic inflammation [26]. Another protective role of F. prausnitzii is that it can inhibit the secretion of pro-inflammatory cytokines such as TNF-α and IL-6, which are major contributors to the inflammatory cascade in IBD patients [29]. This ability to dampen inflammation underscores the importance of maintaining a balanced gut microbiome to prevent disease exacerbation.
In addition to the loss of beneficial bacteria, the microbiome of patients with IBD is often characterized by an overrepresentation of pathogenic bacteria, such as Escherichia coli. Adherent-invasive E. coli (AIEC) strains have been implicated in mucosal inflammation [30]. These pathogenic strains adhere to and invade epithelial cells of the gut, producing virulence factors that trigger and sustain inflammation. A meta-analysis found that AIEC strains are significantly more prevalent in individuals with CD, supporting the idea that microbial dysbiosis, in the form of pathogenic overgrowth, contributes to disease pathogenesis [31]. Given the strong association between gut microbiome alterations and IBD, therapeutic strategies to restore the microbial balance are gaining significant attention.
2.2. Metabolic Disorders
Emerging evidence suggests that the gut microbiome significantly influences metabolic health and is crucial in developing metabolic disorders such as obesity and type 2 diabetes [32]. The microbiome composition in obese individuals tends to differ from that in lean individuals, with a higher proportion of Firmicutes and a lower proportion of Bacteroidetes. This altered microbial composition may affect energy extraction from dietary sources, leading to increased fat storage and weight gain [33]. The gut microbiome regulates host metabolism by producing metabolites, particularly SCFAs [34]. For instance, certain bacterial taxa, such as F. prausnitzii, are associated with increased SCFA production, which has been linked to improved insulin sensitivity and anti-inflammatory effects [35]. Conversely, dysbiosis can lead to reduced SCFA levels and promote insulin resistance, creating a vicious cycle that exacerbates metabolic disorders [27]. Moreover, the gut microbiome affects lipid metabolism and cholesterol homeostasis by influencing energy balance and insulin sensitivity. Some intestinal bacteria can metabolize bile acids, which play a vital role in the digestion and absorption of fat. Alterations in bile acid metabolism, influenced by gut microbial composition, can affect lipid profiles and potentially lead to disorders such as hyperlipidemia and fatty liver disease [36]. Furthermore, the microbiome influences the host response to dietary interventions [37]. For example, individuals with a diverse and balanced microbiome tend to respond better to dietary changes aimed at weight loss than those with dysbiosis [38]. This finding underscores the importance of considering microbiome composition in the development of personalized dietary recommendations for weight management and metabolic health.
2.3. Mental Health
Emerging research has focused on the complex relationship between the gut and brain, known as the gut-brain axis [39]. The gut-brain axis represents a bidirectional communication pathway between the gastrointestinal tract and central nervous system, mediated by various signaling mechanisms, including hormonal, neural, and immune pathways [40]. Research has uncovered significant connections between the gut microbiome and mental health conditions such as depression, anxiety, and neurodegenerative diseases [41]. Gut microbiota can influence brain function by producing neurotransmitters and neuroactive compounds that directly affect central nervous system (CNS) function, influencing gastrointestinal motility and pain perception [39,42]. For instance, certain bacteria can synthesize gamma-aminobutyric acid (GABA) and serotonin, both of which are critical for mood regulation [43]. Dysbiosis may disrupt this process, potentially contributing to the onset of mood disorders. Moreover, the gut-brain axis is thought to play a role in neurodegenerative diseases such as Alzheimer’s and Parkinson’s [44]. Research has indicated that changes in gut microbiota composition may precede the onset of neurodegenerative symptoms, suggesting that microbial intervention may be a novel approach to preventing or mitigating these conditions [45]. The role of microbial metabolites, such as SCFAs, which are produced by the fermentation of dietary fibers, has also been shown to affect gut-brain signaling pathways by interacting with the nervous system, potentially affecting gut motility, inflammation, and visceral sensitivity-key factors in the manifestation of IBS symptoms [46].
2.4. Immune-Mediated Diseases
The gut microbiome plays a pivotal role in regulating the immune system, and its interactions with immune cells are critical for maintaining immune homeostasis. [47]. The gut houses a large proportion of the body’s immune cells, particularly within gut-associated lymphoid tissue (GALT). It is a major interface between the microbiome and the immune system [48]. The microbiome can influence innate and adaptive immune responses, particularly by shaping the functions of T-cells, dendritic cells, and macrophages. Studies have shown that specific microbial populations are involved in the differentiation of T helper (Th) cells and regulatory T cells (Tregs), which are key to maintaining immune tolerance and preventing autoimmunity and chronic inflammation [49]. For example, Lactobacillus and Bifidobacterium strains have been shown to promote Treg differentiation, which can help prevent inflammatory diseases such as IBD and reduce excessive immune responses [50]. Dysbiosis, or an imbalance in microbial composition, has been implicated in a range of autoimmune and inflammatory diseases, including rheumatoid arthritis (RA) and IBD [51]. In IBD, the microbial diversity is often reduced, causing an increased abundance of pro-inflammatory bacteria such as E. coli and Fusobacterium [52]. This dysbiosis contributes to the inappropriate activation of the immune system, leading to chronic inflammation and tissue damage in the gut [53]. Similarly, altered gut microbiota composition in RA has been associated with increased intestinal permeability and systemic inflammation, highlighting the role of the gut in modulating joint health [54].
Dysbiosis, or microbial imbalance, is increasingly recognized as a driver of various immune-mediated diseases, including IBD, rheumatoid arthritis, and autoimmune disorders such as type 1 diabetes and multiple sclerosis. Disruptions in the gut microbial diversity or overgrowth of pathogenic bacteria can lead to a dysregulated immune response, resulting in chronic inflammation and tissue damage. For instance, reduced microbial diversity in the gut has been observed in IBD patients, with an overabundance of pro-inflammatory bacteria and depletion of anti-inflammatory species [54]. The gut microbiome influences systemic immune responses by producing metabolites, such as SCFAs, which have potent anti-inflammatory effects. SCFAs produced by fiber-fermenting, particularly butyrate bacteria, can modulate immune function by promoting Treg activity and inhibiting the activation of pro-inflammatory cytokines [52]
2.5. Cardiovascular Diseases
The relationship between the microbiome and cardiovascular diseases (CVD) has emerged as a prominent area of research, highlighting the critical role of the microbiome in systemic health. However, significant gaps remain in our understanding of the precise mechanisms and the multifactorial nature of these interactions. A foundational study showed that gut bacteria can metabolize dietary phosphatidylcholine to produce trimethylamine-N-oxide (TMAO), a metabolite associated with atherosclerosis. These findings indicate a correlation between elevated TMAO level and cardiovascular events [55]. F Tang et al. (2013) suggested that a more diverse gut microbiome might counteract the harmful effects of TMAO, further highlighting the importance of overall microbial diversity in maintaining cardiovascular health [56]. Chronic inflammation is another critical mechanism linking the microbiome to cardiovascular diseases (CVD). Cani et al. (2009) demonstrated that dysbiosis increases intestinal permeability, allowing bacterial endotoxins to enter the bloodstream and provoking systemic inflammation. This pathway, involving glucagon-like peptide-2 (GLP-2), has been implicated in developing atherosclerosis and other cardiovascular conditions [57].
The impact of the microbiome on key cardiovascular risk factors such as obesity and diabetes has also been extensively studied. A recent study found that specific microbial communities are associated with obesity and insulin resistance, suggesting that the microbiome may influence metabolic pathways contributing to cardiovascular risk [58]. However, the translational potential of these results remains unclear. For instance, a systematic review by Dixon et al. (2020) reported mixed outcomes in clinical trials assessing the efficacy of probiotics in reducing cardiovascular risk factors, highlighting the need for standardized protocols and clearer definitions of success in these studies [59]. Moreover, literature often overlooks the individual variability inherent to microbiome composition. Genetics, age, sex, and ethnicity can significantly influence microbiome diversity and its interactions with cardiovascular health [60].
2.6. Cancer
Colorectal Cancer (CRC) is one of the most prevalent cancers worldwide, and growing evidence suggests a critical link between the composition of gut microbiota and the development of this malignancy [61]. Research has shown that certain bacterial profiles, particularly those enriched in pathogenic species, are associated with an increased likelihood of CRC development [62,63]. Fusobacterium nucleatum is one of the most notable bacteria associated with colorectal cancer [64]. It is known to activate oncogenic signaling pathways, thereby facilitating tumor growth and progression. Specifically, the bacterium produces virulence factors such as adhesins and inflammatory mediators, which can alter the tumor microenvironment by promoting chronic inflammation [65]. Rubinstein et al. (2013) demonstrated that F. nucleatum plays a key role in colorectal tumorigenesis by interacting with colorectal cells and inducing an inflammatory response through the modulation of E-cadherin/catenin signaling [64]. In turn, this inflammation contributes to the initiation and progression of cancer. Clinical studies have shown that patients with elevated F. nucleatum levels have poorer prognoses and increased tumor metastasis, suggesting that this pathogen may contribute not only to tumor development but also to disease progression [66]. This highlights F. nucleatum as a potential biomarker for CRC severity and a target for therapeutic interventions to reduce bacterial load and inflammation in the tumor environment.
Dysbiosis in colorectal cancer is an overrepresentation of pathogenic bacteria and the depletion of beneficial anti-inflammatory bacteria [67]. In particular, there was a reduction in butyrate-producing bacteria, such as those belonging to the phylum Firmicutes. Butyrate, an SCFA, plays a crucial role in maintaining the integrity of the intestinal epithelium, protecting it against DNA damage, and regulating immune responses [68,69]. Studies have shown that butyrate has anticancer properties, including the ability to induce apoptosis (programmed cell death) in cancer cells [70]. Sanchez-Alcoholado et al. (2020) also reported that CRC patients exhibited significantly reduced levels of butyrate-producing bacteria, which may contribute to compromised gut barrier function and increased susceptibility to colorectal cancer [71].
In addition to colorectal cancer, the human microbiome appears to be involved in the development and progression of several other types of cancers. For example, Helicobacter pylori infection is a well-known risk factor for gastric cancer, and chronic inflammation of the stomach lining increases the risk of malignancy [72]. Similarly, some oral microbiota have been associated with an increased pancreatic cancer risk, suggesting that oral pathogens may influence pancreatic carcinogenesis [73,74]. Moreover, lung microbiomes have been found to differ between individuals with lung cancer and healthy controls, suggesting a potential role in lung tumorigenesis [75,76]. The complex connections between the microbiome and various disease states underscore the potential of microbiome-driven therapeutics in clinical practice [77]. Understanding the mechanisms underlying these associations may lead to novel treatment strategies aimed at restoring the microbial balance and improving health outcomes across a spectrum of conditions. Table 1 summarizes the bacterial strains implicated in gastrointestinal and other diseases, including their alterations and clinical significance.
3. Current Microbiome-Driven Therapies
3.1. Fecal Microbiome Transplantation
One of the most promising interventions in microbiome-driven therapy is fecal microbiota transplantation (FMT), a procedure in which fecal matter from a healthy donor enters the recipient’s gastrointestinal tract to reestablish a healthy microbial composition (Figure 2) [90]. This is most often accomplished using colonoscopy, enemas, or capsules. The aim of transplanting healthy fecal matter is to restore gut balance by repopulating it with bacteria that are beneficial to the body and eliminating harmful viruses [91]. The procedure begins by selecting a donor with no family history of autoimmune, metabolic, or malignant disease, followed by infection testing of the donor. The fecal matter from the donor is suspended in a solution containing either water or saline and is strained to eliminate particulate matter. The resulting concoction may be delivered to the recipient by numerous means, such as a nasogastric or nasojejunal tube, esophagogastroduodenoscopy, colonoscopy, or retention enema [92,93].
Fecal microbiota transplantation (FMT) has a rich history in ancient Chinese medicine, predating modern scientific microbiome understanding. The earliest records of fecal therapy in China, dating back to the 4th century AD, were attributed to physician Ge Hong. He described the use of a fecal slurry known as “yellow soup” to treat severe gastrointestinal issues, including diarrhea and food poisoning. This practice has been noted for its effectiveness in protecting patients from critical conditions [94,95]. Currently, FMT is being studied in various patients across North America. The participating investigators entered de-identified data into an online platform, including the FMT protocol, baseline patient characteristics, Clostridioides difficile infection (CDI) cure and recurrence, and short- and long-term safety outcomes. FMT has demonstrated a 90-92% success rate in resolving recurrent CDI after a single treatment [4,96]. A systematic review and meta-analysis showed that fecal microbiota transplantation had a clinical cure rate of 76.1 % %after the first administration. It also revealed that open-label studies had high cure rates of 82.7% compared to 67.7% in randomized trials. [97].
Several studies have reported that FMT can lead to significant clinical improvement in patients with IBD. IBD is closely linked to disruptions in the gut microbiome, characterized by the depletion of beneficial bacteria and the overgrowth of pathogenic strains. The primary goal of FMT in IBD is to restore a balanced gut microbiota and improve the interaction between the microbiota and host immune system [98]. A randomized controlled trial by Moayyedi et al. (2015) found that FMT successfully induced remission in a subset of patients with UC, providing strong evidence that restoring microbiome balance can positively affect disease outcomes [99]. According to a comprehensive study, FMT can alleviate symptoms in approximately 76% of IBD patients, with 63% exhibiting disease improvement. However, recent large-scale meta-analyses have reported decreased efficacy rates, with approximately 36% of patients with UC and 50.5% of patients with CD receiving symptomatic relief after FMT [90,100].
3.2. Probiotics, Prebiotics and Synbiotics
Probiotics, prebiotics, and synbiotics have become key interventions for restoring and maintaining a balanced gut microbiota [101]. Probiotics are live microorganisms that provide health benefits to hosts when consumed in sufficient quantities. Commonly used probiotic strains include Lactobacillus, Bifidobacterium, and Saccharomyces boulardii. These beneficial bacteria enhance intestinal health by inhibiting pathogenic microorganisms, modulating immune responses, and strengthening the intestinal barrier. Clinical applications of probiotics have successfully managed conditions such as IBS, IBD, and antibiotic-associated diarrhea. For instance, certain strains, such as Bifidobacterium infantis, have been found to reduce IBS symptoms, particularly bloating and abdominal pain [102,103]. Although probiotics have been associated with symptom relief in IBD, evidence supporting their role in remission induction is limited. According to some studies, they may be advantageous in sustaining remission in UC but less effective in Crohn’s disease [104,105]. Probiotics have been explored for their ability to modulate immune responses against allergic diseases. Research has indicated that early life administration of probiotics reduces the incidence of eczema during infancy, suggesting its preventative role in allergic diseases [106]. The gut-brain axis was another focus of this probiotic research, where results from a randomized controlled trial showed that a multispecies probiotic supplement reduced symptoms of depression in patients with a major depressive disorder, indicating a potential adjunctive treatment for the management of mood disorders [107]. Animal studies have demonstrated that altering the gut microbiome through diet or probiotics can affect behavior and cognitive function [108]. For example, germ-free mice, which lack gut microbiota, exhibit increased anxiety-like behaviors. Clinical studies have shown that probiotics can improve anxiety and depression symptoms, highlighting the therapeutic potential of targeting the gut microbiome for mental health treatment [109].
Prebiotics are non-digestible food components that selectively stimulate the growth and activity of beneficial intestinal bacteria. Common prebiotics include dietary fibers, such as inulin, fructooligosaccharides (FOS), and galactooligosaccharides (GOS). Prebiotics act as substrates for beneficial microorganisms, promoting a beneficial intestinal environment and increasing microbial diversity and metabolic activity. This may lead to enhanced digestion, an improved immune response, and reduced inflammation. Research has shown that prebiotic consumption promotes the growth of Bifidobacterium and Lactobacillus species, thereby enhancing overall gut health. Additionally, supplementation with prebiotic inulin-type fructans improves insulin sensitivity and reduces body weight in overweight adults, emphasizing the role of prebiotics in metabolic health [110,111].
Synbiotics are combinations of probiotics and prebiotics with synergistic advantages. The rationale behind synbiotics is that the prebiotic component provides a specific substrate that supports the growth and activity of the probiotic strain. This would eventually lead to an increase in the therapeutic outcome because the prebiotics guarantee that the beneficial bacteria not only survive the gastrointestinal transit but also become active and start exerting their health-promoting effects [101]. Clinical research on synbiotics has investigated their potential applications in various health areas including gastrointestinal diseases, metabolic disorders, and immune system modulation. Certain synbiotic formulations are being investigated for their potential to reduce the incidence of necrotizing enterocolitis in preterm infants, most of which show positive results [85]. Synbiotics have been evaluated as adjunctive therapies for IBD management. Randomized controlled trials have shown that the synbiotic combination of Bifidobacterium breve and prebiotic GOS improves the clinical outcome of patients with UC, suggesting a possible synergistic effect in modulating gut inflammation [111]. The effects of synbiotics on cardiovascular health have also been explored. A meta-analysis showed that synbiotic supplementation significantly lowered systolic and diastolic blood pressure levels in patients with hypertension, suggesting a positive impact on blood pressure regulation [112].
3.3. Microbial Metabolite-Based Therapies
Microbial metabolite-based treatments offer a way to capitalize on the positive effects of the gut microbiota on host health, marking a new chapter in the treatment of illness. Their use includes the treatment of neurological illnesses, metabolic diseases, and inflammatory bowel disease. Further research is needed to improve and incorporate these treatments into routine clinical practice. Microbial metabolites are small molecules of gut bacteria produced during the fermentation of dietary fibers and other substrates. These metabolites, including SCFAs, bile acids, and other signaling molecules, play crucial roles in host physiology and health [113]. Metabolites can influence various biological pathways, including immune response, inflammation regulation, and metabolic processes. For instance, SCFAs, such as butyrate, are known to enhance gut barrier integrity, modulate immune function, and exert anti-inflammatory effects [114,115].
Microbial metabolites play an important role in the gut-brain axis. SCFAs, such as butyrate, produced by the intestinal microbiota modulate cognitive functions and behavior. Butyrate exhibits neuroprotective effects and can modulate immune responses, suggesting great therapeutic applications for neurodegenerative diseases, including Alzheimer’s and Parkinson’s diseases [116]. SCFAs also have potent effects on host metabolism. They provide energy to colonocytes, regulate glucose and lipid metabolism, and modulate appetite through hormonal actions. Restoring the healthy levels of these metabolites improves insulin sensitivity and reduces inflammation. This opens up new prospects for the treatment of metabolic disorders such as obesity and type 2 diabetes [117]. In IBD, intestinal dysbiosis leads to a change in the microbial metabolite profile, which contributes to chronic intestinal inflammation. Treatments to re-establish beneficial metabolites, such as butyrate, have been promising for reducing inflammation and facilitating mucosal healing. For instance, butyrate enemas are used to reduce the symptoms in patients with UC [118].
Microbial metabolite-based and probiotic treatments are two distinct approaches for modulating the gut microbiome for health benefits. Table 2 shows the aspects of microbial metabolite-based and probiotic treatments.
3.4. Emerging Microbiome Editing Therapeutics
Recent breakthroughs in microbiome editing therapeutics have paved the way for innovative treatments that precisely target microbial genes in the human gut. These emerging techniques, mostly CRISPR and base editing, offer promising approaches for manipulating microbial communities in situ with high specificity and efficacy, thus showing great promise for treating various diseases. This in-situ technique manipulates the microbiome in its native setting. Although microbiome transplants can create large-scale changes but lack specificity, in-situ engineered microbiomes are designed to target specific bacteria, limiting their impact on the rest of the microbial community [121]. Base editing is a genome-editing technique that enables precise modification of specific DNA bases without causing double-strand breaks or relying on homologous recombination, without introducing double-strand breaks or homologous recombination. This fusion technology combines a catalytically weakened CRISPR-associated nuclease (Cas) protein and nucleobase deaminase enzyme. The guide RNA guides these complexes to the target DNA sequence, where deaminase catalyzes chemical alterations and produces a point mutation. This method allows accurate genetic modifications with a lower risk of unintended genomic changes [122]. Together, these approaches represent a new paradigm in microbiome manipulation and have tremendous potential to address the challenges associated with antibiotic resistance and complex populations of microbes.
Several research groups have developed new gene editing tools specifically for individual gut bacteria or entire communities. Techniques such as CRISPR-Cas9 and inducible CRISPR interference (CRISPRi) have been used to effectively modify bacterial genetic materials [123]. For example, researchers have engineered bacteriophages and viruses that infect bacteria using CRISPR-Cas systems that selectively target antibiotic-resistant bacteria, thereby providing a new approach to combat antibiotic resistance. Recently, clinical trials have tested whether microbiome modification may be a therapeutic agent for infection. Locus Biosciences performed a phase 2 trial of CRISPR-enhanced bacteriophages against E. coli in patients with tract infections. The results showed significant reductions in both bacterial numbers and symptoms of urinary tract infections, suggesting that CRISPR may prove useful in treating bacterial infections [124]. In addition, researchers at the University of California have used CRISPR gene-cutting tools to modify the gut microbiota to prevent childhood asthma by changing the structure of the microbiome. This shows a promising avenue in which diseases related to microbial dysbiosis can be prevented through microbiome modification [125].
Despite these promising improvements, several challenges remain in the field of microbiome-edited therapeutics. Specificity and safety must be guaranteed for genome editing techniques to avoid off-target effects that may disrupt the sensitive balance of the microbiome. Moreover, knowledge of the complex interactions between microbial communities and their consequences on human health is essential for the successful application of such therapies. Ongoing research on developing more precise editing tools and delivery methods aims to overcome these challenges. For instance, the Innovative Genomics Institute (IGI) is developing new CRISPR-based strategies to increase both the safety and accuracy of microbiome editing, which could improve the therapeutic potential of such interventions. Moreover, IGI’s Berkeley Initiative for Optimized Microbiome Editing (BIOME) is dedicated to advancing CRISPR technologies to edit microbiomes within their natural environments precisely. The goal is to develop safe and effective solutions to complex problems by understanding and manipulating the microbial communities [126].
4. Microbiome-Based Drug Development
4.1. Microbiome as a Drug Target
Recent microbiome research has opened up avenues for novel strategies to develop drugs that precisely target specific microbial metabolic pathways for treating infections and as a weapon against antibiotic resistance. Understanding and manipulating these pathways will help to devise therapeutic interventions that inhibit or modulate selective microbial function without damaging the host. One strategy involves targeting vital bacterial processes, including the synthesis of cell walls, proteins, nucleic acids, and other metabolic processes. For example, antibiotics in the β-lactam family inhibit cell wall biosynthesis by attaching penicillin-binding proteins, which leads to bacterial cell lysis. Similarly, drugs such as tetracyclines and macrolides disrupt bacterial protein synthesis by binding to ribosomal subunits, thereby hindering bacterial proliferation [127]. Another promising strategy is the inhibition of bacterial fatty acid biosynthesis. This pathway provides membrane production and energy storage for the bacteria. Compounds that target the essential enzymes in this pathway, such as enoyl-acyl carrier protein reductase (FabI), have demonstrated potent activity against Staphylococcus aureus, including strains resistant to methicillin [128]. The application of genomics has enabled the discovery of new drug targets in microbial metabolic pathways. Studies on bacterial genomes have enabled researchers to identify unique enzymes or mechanisms that are not present in human cells as drug targets, which facilitates the design of highly specific drugs with low toxicity. For example, targets include two-component signal transduction systems (TCSTS) and histidine kinases, which play crucial roles in bacterial adaptation and virulence [129]. Furthermore, the integration of synthetic biology and metabolic engineering has enabled the production of antimicrobial drugs through engineered microbial pathways [130].
Gut microbiota, referred to as the second human genome, is critically important in drug metabolism and significantly influences therapeutic efficacy and adverse effects. Recent research has shown that the microbiota itself is responsible for several drug modifications, such as activation, inactivation, and even toxification. Variations in the gut microbiota of different individuals may result in divergent microbe-drug interactions, underscoring the importance of personalized approaches in pharmacotherapy [131,132,133].
4.2. Engineered Microbiomes
Engineered microbiomes, or synthetic microbial communities, are designed to perform specific functions in a host or environment for precise interventions in health and disease management. Researchers have assembled defined microbial consortia with tailored functionalities to modulate the host physiology, enhance therapeutic outcomes, and mitigate disease processes. The development of synthetic microbiomes involves the careful selection and integration of microbial species with specific traits to achieve targeted effects. This process can be achieved through two approaches: bottom-up and top-down. The bottom-up approach isolates and assembles specific microbial strains into an organized community with defined functions. This method allows for complete and precise control of the community, composition, and interactions. The top-down approach involves altering existing microbial communities by adding or deleting members of the community to change community behavior and function [134]. Advances in genetic engineering and synthetic biology have enabled the programming of microbial consortia to execute complex tasks including biosensing, metabolic production, and immune modulation [135].
Engineered microbiomes have significant potential in precision medicine by facilitating treatments that are customized to each patient’s unique characteristics. In disease therapy, engineered microbiomes can modulate metabolic pathways, generate therapeutic compounds, or outcompete pathogenic microbes to treat conditions such as IBD, infections, and metabolic disorders [135]. Engineered gut microbiota composition also controls drug metabolism, thereby improving efficacy and decreasing adverse effects [7]. In addition, engineered microbial communities modulate immune responses, thus offering potential treatments for autoimmune diseases and allergies [135]. Despite their promise, engineered microbiomes face several challenges in terms of stability in the host, avoidance of off-target interactions, and ethical and regulatory concerns. Ongoing studies are focused on areas such as developing strategies to control engineered microbes and prevent horizontal gene transfer to native microbiota [136], altering synthetic microbiomes to individual genetic and microbiome profiles to maximize therapeutic benefits [7], and establishing regulations for the clinical use of engineered microbiomes to confirm safety and efficacy [137].
4.3. Microbiome as a Drug Target
Recent microbiome research has opened up avenues for novel strategies to develop drugs that precisely target specific microbial metabolic pathways for treating infections and as a weapon against antibiotic resistance. Understanding and manipulating these pathways will help to devise therapeutic interventions that inhibit or modulate selective microbial function without damaging the host. One strategy involves targeting vital bacterial processes, including the synthesis of cell walls, proteins, nucleic acids, and other metabolic processes. For example, antibiotics in the β-lactam family inhibit cell wall biosynthesis by attaching penicillin-binding proteins, which leads to bacterial cell lysis. Similarly, drugs such as tetracyclines and macrolides disrupt bacterial protein synthesis by binding to ribosomal subunits, thereby hindering bacterial proliferation [127]. Another promising strategy is the inhibition of bacterial fatty acid biosynthesis. This pathway provides membrane production and energy storage for the bacteria. Compounds that target the essential enzymes in this pathway, such as enoyl-acyl carrier protein reductase (FabI), have demonstrated potent activity against Staphylococcus aureus, including strains resistant to methicillin [128]. The application of genomics has enabled the discovery of new drug targets in microbial metabolic pathways. Studies on bacterial genomes have enabled researchers to identify unique enzymes or mechanisms that are not present in human cells as drug targets, which facilitates the design of highly specific drugs with low toxicity. For example, targets include two-component signal transduction systems (TCSTS) and histidine kinases, which play crucial roles in bacterial adaptation and virulence [129]. Furthermore, the integration of synthetic biology and metabolic engineering has enabled the production of antimicrobial drugs through engineered microbial pathways [130].
Gut microbiota, referred to as the second human genome, is critically important in drug metabolism and significantly influences therapeutic efficacy and adverse effects. Recent research has shown that the microbiota itself is responsible for several drug modifications, such as activation, inactivation, and even toxification. Variations in the gut microbiota of different individuals may result in divergent microbe-drug interactions, underscoring the importance of personalized approaches in pharmacotherapy [131,132,133].
4.4. Engineered Microbiomes
Engineered microbiomes, or synthetic microbial communities, are designed to perform specific functions in a host or environment for precise interventions in health and disease management. Researchers have assembled defined microbial consortia with tailored functionalities to modulate the host physiology, enhance therapeutic outcomes, and mitigate disease processes. The development of synthetic microbiomes involves the careful selection and integration of microbial species with specific traits to achieve targeted effects. This process can be achieved through two approaches: bottom-up and top-down. The bottom-up approach isolates and assembles specific microbial strains into an organized community with defined functions. This method allows for complete and precise control of the community, composition, and interactions. The top-down approach involves altering existing microbial communities by adding or deleting members of the community to change community behavior and function [134]. Advances in genetic engineering and synthetic biology have enabled the programming of microbial consortia to execute complex tasks including biosensing, metabolic production, and immune modulation [135].
Engineered microbiomes have significant potential in precision medicine by facilitating treatments that are customized to each patient’s unique characteristics. In disease therapy, engineered microbiomes can modulate metabolic pathways, generate therapeutic compounds, or outcompete pathogenic microbes to treat conditions such as IBD, infections, and metabolic disorders [135]. Engineered gut microbiota composition also controls drug metabolism, thereby improving efficacy and decreasing adverse effects [7]. In addition, engineered microbial communities modulate immune responses, thus offering potential treatments for autoimmune diseases and allergies [135]. Despite their promise, engineered microbiomes face several challenges in terms of stability in the host, avoidance of off-target interactions, and ethical and regulatory concerns. Ongoing studies are focused on areas such as developing strategies to control engineered microbes and prevent horizontal gene transfer to native microbiota [136] altering synthetic microbiomes to individual genetic and microbiome profiles to maximize therapeutic benefits [7], and establishing regulations for the clinical use of engineered microbiomes to confirm safety and efficacy [137].
5. Challenges and Limitations
5.1. Individual Microbiome Variation
Research has demonstrated a significant correlation between gut microbiome diversity and factors such as food, demographics, health problems, and hygiene [138]. While ethnicity has an impact on the microbiome, other factors such as geographic location, culture, tradition, dietary pattern, lifestyle, and exposure to toxins and diseases largely contribute to differences in microbiomes among different populations of humans, which poses challenges in microbiome-driven therapy [139]. Recent research indicates that a “one size fits all” approach to dietary interventions for treating metabolic disorders could be insufficient because the gut microbiota plays a critical role in inter-individual variability in the metabolism of key nutrients [140]. Despite considerable heterogeneity, identifying reliable microbiological biomarkers that predict therapeutic responses is challenging because each individual’s microbiome is unique; individuals with similar symptoms may show divergent reactions to identical treatments [141].
5.2. Mechanistic Gaps in Microbiome Research
Although microbiome data have enormous therapeutic potential, they are still difficult to analyze and interpret owing to several factors, including compositional structure, which introduces a negative correlation bias, sparsity, and collinearity [142]. The complexity of microbial ecosystems has led to a lack of understanding of specific microbial-host interactions. Consequently, the effects of probiotics and synbiotics are not yet fully understood [143]. The human microbiome comprises a complex array of microorganisms, and our knowledge of their individual roles in human health remains limited. There is a lack of comprehensive understanding of how successfully these microbial species perform their roles, their interactions with the host and other microbial species, and their specific locations within the host where they exert their influence. This knowledge gap highlights the need for further research to understand these complex relationships.
5.3. Regulatory and Ethical Considerations
According to research ethics, the potential societal benefits of conducting a study must be weighed against the potential risks. Studies on the human microbiome must explain a favorable risk-benefit ratio. However, this can be extremely challenging owing to several unresolved microbiological, clinical, and social problems. Balancing potential risks and benefits is often difficult and nearly impossible in some cases [144]. To ensure safety and effectiveness, therapies involving live organisms such as FMT, probiotics, and prebiotics are under strict regulatory oversight. Selecting the right regulatory category, such as drug, biological, or dietary supplement, can be challenging, as it profoundly impacts the requirements and approval process. Depending on their function and characteristics, developers must classify their products under regulatory frameworks as biological goods, medications, or medical devices. Preclinical studies, clinical evaluations, and assessments of adverse effects on the microbiome and host physiology must be performed to determine both safety and efficacy. The manufacturing, distribution, and storage of live biotherapeutic products (LBPs) follows standard good manufacturing practices (GMP) guidelines [11]. Ethical considerations in FMT include the major elements necessary to protect patients and ensure informed participation. Rigorous screening and precise selection can reduce the risk of transferring novel diseases to recipients through FMT. Informed consent is another crucial component. It is a voluntary agreement to participate in clinical trials based on knowledge of research goals, risks, potential advantages, and safety issues. Also, a proper assessment of safety issues and benefit-to-risk ratio should be carried out to avoid adverse effects and ensure the therapy is safe and effective overall [145].
6. Future Directions
Microbiome-driven therapeutics are rapidly advancing, with future research focusing on addressing these challenges and broadening therapeutic applications. Precision medicine will play a key role by utilizing individual microbial profiles to develop personalized treatments that enhance efficacy and reduce side effects [146]. Progress in synthetic biology and computational modeling is expected to improve the design of engineered microbiomes and predictive tools, thereby increasing their stability and functionality. Understanding the gut-brain axis and immune modulation promises new therapies for neurological diseases and autoimmune disorders [147]. Future research has also highlighted the potential of microbiome modulation in cancer therapy to improve responses to immunotherapy and reduce side effects through targeted interventions such as probiotics or engineered microbiomes [10,148]. Therefore, one of the critical avenues that will ensure the safe and effective translation of these therapies into the clinic is the creation of strong regulatory frameworks and ethical guidelines [11]. This will be accelerated by public education, access equality, and expansion into understudied areas, including research on the skin and oral microbiomes [149,150,151]. Together, these efforts will establish microbiome therapeutics as a cornerstone of precision medicine and improve health outcomes worldwide.
7. Conclusions
Microbiome-driven therapeutics are a next-generation approach for treating complex diseases, highlighting host-microbial community interplay. This review covers the current developments in FMT, probiotics, and engineered microbiomes, each of which holds promise to meet unmet medical needs. However, the remaining challenges are significant, such as individual variability, knowledge gaps, and regulatory hurdles. However, with interdisciplinary approaches, innovation, and adherence to ethical standards, microbiome-based treatments could revolutionize healthcare by providing targeted, effective, and sustainable solutions for numerous conditions.
Author Contributions
Conceptualization, M.O.Y.; writing—original draft preparation, M.O.Y., A.J., C.E.J., and L.K.E.; writing—review and editing, L.K.E; supervision, L.K.E. All authors have read and agreed to the published version of the manuscript.
Acknowledgments
The authors are grateful to the American Society of Microbiology (ASM) Future Leader Mentorship Fellowship (FLMF) Program.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Young, V.B. The Role of the Microbiome in Human Health and Disease: An Introduction for Clinicians. BMJ (Online) 2017, 356. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Wang, X.; Li, L. Human Gut Microbiome: The Second Genome of Human Body. Protein Cell 2010, 1, 718–725. [Google Scholar] [CrossRef] [PubMed]
- Khanna, S.; Pardi, D.S. Clostridium Difficile Infection: New Insights into Management. Mayo Clin Proc 2012, 87, 1106–1117. [Google Scholar] [CrossRef] [PubMed]
- Kelly, C.R.; Yen, E.F.; Grinspan, A.M.; Kahn, S.A.; Atreja, A.; Lewis, J.D.; Moore, T.A.; Rubin, D.T.; Kim, A.M.; Serra, S.; et al. Fecal Microbiota Transplantation Is Highly Effective in Real-World Practice: Initial Results From the FMT National Registry. Gastroenterology 2021, 160, 183–192.e3. [Google Scholar] [CrossRef]
- Al-Fakhrany, O.M.; Elekhnawy, E. Next-Generation Probiotics: The Upcoming Biotherapeutics. Mol Biol Rep 2024, 51, 505. [Google Scholar] [CrossRef]
- Petrosino, J.F. The Microbiome in Precision Medicine: The Way Forward. Genome Med 2018, 10, 12. [Google Scholar] [CrossRef]
- Ryu, G.; Kim, H.; Koh, A. Approaching Precision Medicine by Tailoring the Microbiota. Mammalian Genome 2021, 32, 206–222. [Google Scholar] [CrossRef]
- Rasul, M.F.; Hussen, B.M.; Salihi, A.; Ismael, B.S.; Jalal, P.J.; Zanichelli, A.; Jamali, E.; Baniahmad, A.; Ghafouri-Fard, S.; Basiri, A.; et al. Strategies to Overcome the Main Challenges of the Use of CRISPR/Cas9 as a Replacement for Cancer Therapy. Mol Cancer 2022, 21, 64. [Google Scholar] [CrossRef]
- Sioson, V.A.; Kim, M.; Joo, J. Challenges in Delivery Systems for CRISPR-Based Genome Editing and Opportunities of Nanomedicine. Biomed Eng Lett 2021, 11, 217–233. [Google Scholar] [CrossRef]
- Lu, Y.; Yuan, X.; Wang, M.; He, Z.; Li, H.; Wang, J.; Li, Q. Gut Microbiota Influence Immunotherapy Responses: Mechanisms and Therapeutic Strategies. J Hematol Oncol 2022, 15. [Google Scholar] [CrossRef]
- Waheed Rashem and Farooq, A.Z. and Farooq, A.Z. and A.L. Regulatory Considerations for Microbiome-Based Therapeutics. In Human Microbiome: Techniques, Strategies, and Therapeutic Potential; Khurshid Mohsin and Akash, M.S.H., Ed.; Springer Nature Singapore: Singapore, 2024; ISBN 978-981-97-3790-1. [Google Scholar]
- Rhodes, R. Ethical Issues in Microbiome Research and Medicine. BMC Med 2016, 14, 156. [Google Scholar] [CrossRef] [PubMed]
- Carroll, I.M.; Ringel-Kulka, T.; Siddle, J.P.; Ringel, Y. Alterations in Composition and Diversity of the Intestinal Microbiota in Patients with Diarrhea-Predominant Irritable Bowel Syndrome. Neurogastroenterology and Motility 2012, 24. [Google Scholar] [CrossRef] [PubMed]
- Carroll, I.M.; Chang, Y.H.; Park, J.; Sartor, R.B.; Ringel, Y. Luminal and Mucosal-Associated Intestinal Microbiota in Patients with Diarrhea-Predominant Irritable Bowel Syndrome. Gut Pathog 2010, 2. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, S.D.; Sun, N.; Canakis, A.; Park, W.Y.; Weber, H.C. Irritable Bowel Syndrome and the Gut Microbiome: A Comprehensive Review. J Clin Med 2023, 12. [Google Scholar] [CrossRef]
- Wang, L.; Alammar, N.; Singh, R.; Nanavati, J.; Song, Y.; Chaudhary, R.; Mullin, G.E. Gut Microbial Dysbiosis in the Irritable Bowel Syndrome: A Systematic Review and Meta-Analysis of Case-Control Studies. J Acad Nutr Diet 2020, 120, 565–586. [Google Scholar] [CrossRef]
- Degruttola, A.K.; Low, D.; Mizoguchi, A.; Mizoguchi, E. Current Understanding of Dysbiosis in Disease in Human and Animal Models. Inflamm Bowel Dis 2016, 22, 1137–1150. [Google Scholar] [CrossRef]
- Baldelli, V.; Scaldaferri, F.; Putignani, L.; Del Chierico, F. The Role of Enterobacteriaceae in Gut Microbiota Dysbiosis in Inflammatory Bowel Diseases. Microorganisms 2021, 9. [Google Scholar] [CrossRef]
- Di Vincenzo, F.; Del Gaudio, A.; Petito, V.; Lopetuso, L.R.; Scaldaferri, F. Gut Microbiota, Intestinal Permeability, and Systemic Inflammation: A Narrative Review. Intern Emerg Med 2024, 19, 275–293. [Google Scholar] [CrossRef]
- Jang, J.H.; Jang, S.Y.; Ahn, S.; Oh, J.Y.; Yeom, M.; Ko, S.J.; Park, J.W.; Kwon, S.K.; Kim, K.; Lee, I.S.; et al. Chronic Gut Inflammation and Dysbiosis in IBS: Unraveling Their Contribution to Atopic Dermatitis Progression. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef]
- Coates, M.D.; Clarke, K.; Williams, E.; Jeganathan, N.; Yadav, S.; Giampetro, D.; Gordin, V.; Smith, S.; Vrana, K.; Bobb, A.; et al. Abdominal Pain in Inflammatory Bowel Disease: An Evidence-Based, Multidisciplinary Review. Crohns Colitis 360 2023, 5. [Google Scholar] [CrossRef]
- Pasta, A.; Formisano, E.; Calabrese, F.; Plaz Torres, M.C.; Bodini, G.; Marabotto, E.; Pisciotta, L.; Giannini, E.G.; Furnari, M. Food Intolerances, Food Allergies and IBS: Lights and Shadows. Nutrients 2024, 16. [Google Scholar] [CrossRef] [PubMed]
- Gros, M.; Gros, B.; Mesonero, J.E.; Latorre, E. Neurotransmitter Dysfunction in Irritable Bowel Syndrome: Emerging Approaches for Management. J Clin Med 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Fakhoury, M.; Negrulj, R.; Mooranian, A.; Al-Salami, H. Inflammatory Bowel Disease: Clinical Aspects and Treatments. J Inflamm Res 2014, 7, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Pittayanon, R.; Lau, J.T.; Leontiadis, G.I.; Tse, F.; Yuan, Y.; Surette, M.; Moayyedi, P. Differences in Gut Microbiota in Patients With vs Without Inflammatory Bowel Diseases: A Systematic Review. Gastroenterology 2020, 158, 930–946.e1. [Google Scholar] [CrossRef]
- Sokol, H.; né dicte Pigneur, B.; Watterlot, L.; Lakhdari, O.; Bermú dez-Humará, L.G.; Gratadoux, J.-J.; bastien Blugeon, S.; Bridonneau, C.; Furet, J.-P.; rard Corthier, G.; et al. 2008.
- Nogal, A.; Valdes, A.M.; Menni, C. The Role of Short-Chain Fatty Acids in the Interplay between Gut Microbiota and Diet in Cardio-Metabolic Health. Gut Microbes 2021, 13, 1–24. [Google Scholar] [CrossRef]
- Miquel, S.; Martín, R.; Rossi, O.; Bermúdez-Humarán, L.G.; Chatel, J.M.; Sokol, H.; Thomas, M.; Wells, J.M.; Langella, P. Faecalibacterium Prausnitzii and Human Intestinal Health. Curr Opin Microbiol 2013, 16, 255–261. [Google Scholar] [CrossRef]
- Miquel, S.; Leclerc, M.; Martin, R.; Chain, F.; Lenoir, M.; Raguideau, S.; Hudault, S.; Bridonneau, C.; Northene, T.; Bowene, B.; et al. Identification of Metabolic Signatures Linked to Anti-Inflammatory Effects of Faecalibacterium Prausnitzii. mBio 2015, 6, 1–10. [Google Scholar] [CrossRef]
- Chloé Mirsepasi-Lauridsen, H.; Vallance, B.A.; Krogfelt, K.A.; Petersen, A.M. Escherichia Coli Pathobionts Associated with Inflammatory Bowel Disease. 2019. [Google Scholar] [CrossRef]
- Kamali Dolatabadi, R.; Feizi, A.; Halaji, M.; Fazeli, H.; Adibi, P. The Prevalence of Adherent-Invasive Escherichia Coli and Its Association With Inflammatory Bowel Diseases: A Systematic Review and Meta-Analysis. Front Med (Lausanne) 2021, 8. [Google Scholar] [CrossRef]
- Festi, D.; Schiumerini, R.; Eusebi, L.H.; Marasco, G.; Taddia, M.; Colecchia, A. Gut Microbiota and Metabolic Syndrome. World J Gastroenterol 2014, 20, 16079–16094. [Google Scholar] [CrossRef]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes Ratio: A Relevant Marker of Gut Dysbiosis in Obese Patients? Nutrients 2020, 12. [Google Scholar] [CrossRef]
- Fusco, W.; Lorenzo, M.B.; Cintoni, M.; Porcari, S.; Rinninella, E.; Kaitsas, F.; Lener, E.; Mele, M.C.; Gasbarrini, A.; Collado, M.C.; et al. Short-Chain Fatty-Acid-Producing Bacteria: Key Components of the Human Gut Microbiota. Nutrients 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, T.S.; Shanahan, F.; O’Toole, P.W. The Gut Microbiome as a Modulator of Healthy Ageing. Nat Rev Gastroenterol Hepatol 2022, 19, 565–584. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Pérez, O.; Cruz-Ramón, V.; Chinchilla-López, P.; Méndez-Sánchez, N. The Role of the Gut Microbiota in Bile Acid Metabolism. Ann Hepatol 2017, 16, S21–s26. [Google Scholar] [CrossRef] [PubMed]
- Corbin, K.D.; Carnero, E.A.; Dirks, B.; Igudesman, D.; Yi, F.; Marcus, A.; Davis, T.L.; Pratley, R.E.; Rittmann, B.E.; Krajmalnik-Brown, R.; et al. Host-Diet-Gut Microbiome Interactions Influence Human Energy Balance: A Randomized Clinical Trial. Nat Commun 2023, 14. [Google Scholar] [CrossRef]
- Alcock, J.; Maley, C.C.; Aktipis, C.A. Is Eating Behavior Manipulated by the Gastrointestinal Microbiota? Evolutionary Pressures and Potential Mechanisms. BioEssays 2014, 36, 940–949. [Google Scholar] [CrossRef]
- Yarandi, S.S.; Peterson, D.A.; Treisman, G.J.; Moran, T.H.; Pasricha, P.J. Modulatory Effects of Gut Microbiota on the Central Nervous System: How Gut Could Play a Role in Neuropsychiatric Health and Diseases. J Neurogastroenterol Motil 2016, 22, 201–212. [Google Scholar] [CrossRef]
- Kasarello, K.; Cudnoch-Jedrzejewska, A.; Czarzasta, K. Communication of Gut Microbiota and Brain via Immune and Neuroendocrine Signaling. Front Microbiol 2023, 14. [Google Scholar] [CrossRef]
- Grau-Del Valle, C.; Fernández, J.; Solá, E.; Montoya-Castilla, I.; Morillas, C.; Bañuls, C. Association between Gut Microbiota and Psychiatric Disorders: A Systematic Review. Front Psychol 2023, 14. [Google Scholar] [CrossRef]
- Chen, Y.; Xu, J.; Chen, Y. Regulation of Neurotransmitters by the Gut Microbiota and Effects on Cognition in Neurological Disorders. Nutrients 2021, 13. [Google Scholar] [CrossRef]
- Braga, J.D.; Thongngam, M.; Kumrungsee, T. Gamma-Aminobutyric Acid as a Potential Postbiotic Mediator in the Gut–Brain Axis. NPJ Sci Food 2024, 8. [Google Scholar] [CrossRef]
- Caradonna, E.; Nemni, R.; Bifone, A.; Gandolfo, P.; Costantino, L.; Giordano, L.; Mormone, E.; Macula, A.; Cuomo, M.; Difruscolo, R.; et al. The Brain–Gut Axis, an Important Player in Alzheimer and Parkinson Disease: A Narrative Review. J Clin Med 2024, 13. [Google Scholar] [CrossRef] [PubMed]
- Boehme, M.; Guzzetta, K.E.; Wasén, C.; Cox, L.M. The Gut Microbiota Is an Emerging Target for Improving Brain Health during Ageing. Gut Microbiome 2023, 4. [Google Scholar] [CrossRef] [PubMed]
- Carabotti, M.; Scirocco, A.; Antonietta Maselli, M.; Severi, C. The Gut-Brain Axis: Interactions between Enteric Microbiota, Central and Enteric Nervous Systems; 2015; Vol. 28;
- Wiertsema, S.P.; van Bergenhenegouwen, J.; Garssen, J.; Knippels, L.M.J. The Interplay between the Gut Microbiome and the Immune System in the Context of Infectious Diseases throughout Life and the Role of Nutrition in Optimizing Treatment Strategies. Nutrients 2021, 13, 1–14. [Google Scholar] [CrossRef]
- Wu, H.J.; Wu, E. The Role of Gut Microbiota in Immune Homeostasis and Autoimmunity. Gut Microbes 2012, 3. [Google Scholar] [CrossRef]
- Belkaid, Y.; Hand, T.W. Role of the Microbiota in Immunity and Inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef]
- Lee, N.; Kim, W.U. Microbiota in T-Cell Homeostasis and Inflammatory Diseases. Exp Mol Med 2017, 49. [Google Scholar] [CrossRef]
- Mousa, W.K.; Chehadeh, F.; Husband, S. Microbial Dysbiosis in the Gut Drives Systemic Autoimmune Diseases. Front Immunol 2022, 13. [Google Scholar] [CrossRef]
- Schirmer, M.; Garner, A.; Vlamakis, H.; Xavier, R.J. Microbial Genes and Pathways in Inflammatory Bowel Disease. Nat Rev Microbiol 2019, 17, 497–511. [Google Scholar] [CrossRef]
- Santana, P.T.; Rosas, S.L.B.; Ribeiro, B.E.; Marinho, Y.; de Souza, H.S.P. Dysbiosis in Inflammatory Bowel Disease: Pathogenic Role and Potential Therapeutic Targets. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Bodkhe, R.; Balakrishnan, B.; Taneja, V. The Role of Microbiome in Rheumatoid Arthritis Treatment. Ther Adv Musculoskelet Dis 2019, 11. [Google Scholar] [CrossRef]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; Dugar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.M.; et al. Gut Flora Metabolism of Phosphatidylcholine Promotes Cardiovascular Disease. Nature 2011, 472, 57–65. [Google Scholar] [CrossRef]
- Tang, W.H.W.; Wang, Z.; Levison, B.S.; Koeth, R.A.; Britt, E.B.; Fu, X.; Wu, Y.; Hazen, S.L. Intestinal Microbial Metabolism of Phosphatidylcholine and Cardiovascular Risk. New England Journal of Medicine 2013, 368, 1575–1584. [Google Scholar] [CrossRef]
- Cani, P.D.; Possemiers, S.; Van De Wiele, T.; Guiot, Y.; Everard, A.; Rottier, O.; Geurts, L.; Naslain, D.; Neyrinck, A.; Lambert, D.M.; et al. Changes in Gut Microbiota Control Inflammation in Obese Mice through a Mechanism Involving GLP-2-Driven Improvement of Gut Permeability. Gut 2009, 58, 1091–1103. [Google Scholar] [CrossRef] [PubMed]
- Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Cheng, J.; Duncan, A.E.; Kau, A.L.; Griffin, N.W.; Lombard, V.; Henrissat, B.; Bain, J.R.; et al. Gut Microbiota from Twins Discordant for Obesity Modulate Metabolism in Mice. Science (1979) 2013, 341. [Google Scholar] [CrossRef] [PubMed]
- Dixon, A.; Robertson, K.; Yung, A.; Que, M.; Randall, H.; Wellalagodage, D.; Cox, T.; Robertson, D.; Chi, C.; Sun, J. Efficacy of Probiotics in Patients of Cardiovascular Disease Risk: A Systematic Review and Meta-Analysis. Curr Hypertens Rep 2020, 22. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between Microbiota and Immunity in Health and Disease. Cell Res 2020, 30, 492–506. [Google Scholar] [CrossRef]
- Kim, J.; Lee, H.K. Potential Role of the Gut Microbiome In Colorectal Cancer Progression. Front Immunol 2022, 12. [Google Scholar] [CrossRef]
- White, M.T.; Sears, C.L. The Microbial Landscape of Colorectal Cancer. Nat Rev Microbiol 2024, 22, 240–254. [Google Scholar] [CrossRef]
- Zhao, L.; Cho, W.C.; Nicolls, M.R. Colorectal Cancer-Associated Microbiome Patterns and Signatures. Front Genet 2021, 12. [Google Scholar] [CrossRef]
- Rubinstein, M.R.; Wang, X.; Liu, W.; Hao, Y.; Cai, G.; Han, Y.W. Fusobacterium Nucleatum Promotes Colorectal Carcinogenesis by Modulating E-Cadherin/β-Catenin Signaling via Its FadA Adhesin. Cell Host Microbe 2013, 14, 195–206. [Google Scholar] [CrossRef]
- Pignatelli, P.; Nuccio, F.; Piattelli, A.; Curia, M.C. The Role of Fusobacterium Nucleatum in Oral and Colorectal Carcinogenesis. Microorganisms 2023, 11. [Google Scholar] [CrossRef]
- Udayasuryan, B.; Zhou, Z.; Ahmad, R.N.; Sobol, P.; Deng, C.; Nguyen, T.T.D.; Kodikalla, S.; Morrison, R.; Goswami, I.; Slade, D.J.; et al. Fusobacterium Nucleatum Infection Modulates the Transcriptome and Epigenome of HCT116 Colorectal Cancer Cells in an Oxygen-Dependent Manner. Commun Biol 2024, 7. [Google Scholar] [CrossRef]
- Tabowei, G.; Gaddipati, G.N.; Mukhtar, M.; Alzubaidee, M.J.; Dwarampudi, R.S.; Mathew, S.; Bichenapally, S.; Khachatryan, V.; Muazzam, A.; Hamal, C.; et al. Microbiota Dysbiosis a Cause of Colorectal Cancer or Not? A Systematic Review. Cureus 2022. [Google Scholar] [CrossRef]
- Wang, H.-B.; Wang, P.-Y.; Wang, X.; Wan, Y.-L.; Liu, Y.-C. Butyrate Enhances Intestinal Epithelial Barrier Function via Up-Regulation of Tight Junction Protein Claudin-1 Transcription. Dig Dis Sci 2012, 57, 3126–3135. [Google Scholar] [CrossRef]
- Berni Canani, R.; Di Costanzo, M.; Leone, L. The Epigenetic Effects of Butyrate: Potential Therapeutic Implications for Clinical Practice. Clin Epigenetics 2012, 4, 4. [Google Scholar] [CrossRef]
- Chen, J.; Zhao, K.N.; Vitetta, L. Effects of Intestinal Microbial-Elaborated Butyrate on Oncogenic Signaling Pathways. Nutrients 2019, 11. [Google Scholar] [CrossRef]
- Sánchez-Alcoholado, L.; Ramos-Molina, B.; Otero, A.; Laborda-Illanes, A.; Ordóñez, R.; Medina, J.A.; Gómez-Millán, J.; Queipo-Ortuño, M.I. The Role of the Gut Microbiome in Colorectal Cancer Development and Therapy Response. Cancers (Basel) 2020, 12. [Google Scholar] [CrossRef]
- Helicobacter Pylori (H. Pylori) and Cancer Available online:. Available online: https://www.cancer.gov/about-cancer/causes-prevention/risk/infectious-agents/h-pylori-fact-sheet?utm_source=chatgpt.com (accessed on 23 November 2024).
- Fan, X.; Alekseyenko, A. V.; Wu, J.; Peters, B.A.; Jacobs, E.J.; Gapstur, S.M.; Purdue, M.P.; Abnet, C.C.; Stolzenberg-Solomon, R.; Miller, G.; et al. Human Oral Microbiome and Prospective Risk for Pancreatic Cancer: A Population-Based Nested Case-Control Study. Gut 2018, 67, 120–127. [Google Scholar] [CrossRef]
- Doocey, C.M.; Finn, K.; Murphy, C.; Guinane, C.M. The Impact of the Human Microbiome in Tumorigenesis, Cancer Progression, and Biotherapeutic Development. BMC Microbiol 2022, 22. [Google Scholar] [CrossRef]
- Parida, S.; Sharma, D. The Microbiome and Cancer: Creating Friendly Neighborhoods and Removing the Foes with in A C. Cancer Res 2021, 81, 790–800. [Google Scholar] [CrossRef]
- Sexton, R.E.; Uddin, M.H.; Bannoura, S.; Khan, H.Y.; Mzannar, Y.; Li, Y.; Aboukameel, A.; Al-Hallak, M.N.; Al-Share, B.; Mohamed, A.; et al. Connecting the Human Microbiome and Pancreatic Cancer. Cancer and Metastasis Reviews 2022, 41, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Kamel, M.; Aleya, S.; Alsubih, M.; Aleya, L. Microbiome Dynamics: A Paradigm Shift in Combatting Infectious Diseases. J Pers Med 2024, 14. [Google Scholar] [CrossRef]
- Evangelia Legaki and Karanasou, E.A. and Karanasou, E.A. and G.M. Gut Microbiome and Gastrointestinal Disorders. In Gut Microbiome-Related Diseases and Therapies; Gazouli Maria and Theodoropoulos, G., Ed.; Springer International Publishing: Cham, 2021; ISBN 978-3-030-59642-2. [Google Scholar]
- Han, Z.; Min, Y.; Pang, K.; Wu, D. Therapeutic Approach Targeting Gut Microbiome in Gastrointestinal Infectious Diseases. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Bakhti, S.Z.; Latifi-Navid, S. Interplay and Cooperation of Helicobacter Pylori and Gut Microbiota in Gastric Carcinogenesis. BMC Microbiol 2021, 21. [Google Scholar] [CrossRef]
- Shariati, A.; Razavi, S.; Ghaznavi-Rad, E.; Jahanbin, B.; Akbari, A.; Norzaee, S.; Darban-Sarokhalil, D. Association between Colorectal Cancer and Fusobacterium Nucleatum and Bacteroides Fragilis Bacteria in Iranian Patients: A Preliminary Study. Infect Agent Cancer 2021, 16. [Google Scholar] [CrossRef]
- Li, S.; Liu, J.; Zheng, X.; Ren, L.; Yang, Y.; Li, W.; Fu, W.; Wang, J.; Du, G. Tumorigenic Bacteria in Colorectal Cancer: Mechanisms and Treatments. Cancer Biol Med 2022, 19, 147–162. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, N.; Tan, H.Y.; Li, S.; Zhang, C.; Feng, Y. Function of Akkermansia Muciniphila in Obesity: Interactions With Lipid Metabolism, Immune Response and Gut Systems. Front Microbiol 2020, 11. [Google Scholar] [CrossRef]
- Akobeng, A.K.; Singh, P.; Kumar, M.; Al Khodor, S. Role of the Gut Microbiota in the Pathogenesis of Coeliac Disease and Potential Therapeutic Implications. Eur J Nutr 2020, 59, 3369–3390. [Google Scholar] [CrossRef]
- Marasco, G.; Di Biase, A.R.; Schiumerini, R.; Eusebi, L.H.; Iughetti, L.; Ravaioli, F.; Scaioli, E.; Colecchia, A.; Festi, D. Gut Microbiota and Celiac Disease. Dig Dis Sci 2016, 61, 1461–1472. [Google Scholar] [CrossRef]
- Ho, L.K.H.; Tong, V.J.W.; Syn, N.; Nagarajan, N.; Tham, E.H.; Tay, S.K.; Shorey, S.; Tambyah, P.A.; Law, E.C.N. Gut Microbiota Changes in Children with Autism Spectrum Disorder: A Systematic Review. Gut Pathog 2020, 12, 6. [Google Scholar] [CrossRef]
- Ding, H.T.; Taur, Y.; Walkup, J.T. Gut Microbiota and Autism: Key Concepts and Findings. J Autism Dev Disord 2017, 47, 480–489. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.H.W.; Kitai, T.; Hazen, S.L. Gut Microbiota in Cardiovascular Health and Disease. Circ Res 2017, 120, 1183–1196. [Google Scholar] [CrossRef] [PubMed]
- Kazemian, N.; Mahmoudi, M.; Halperin, F.; Wu, J.C.; Pakpour, S. Gut Microbiota and Cardiovascular Disease: Opportunities and Challenges. Microbiome 2020, 8, 36. [Google Scholar] [CrossRef] [PubMed]
- Boicean, A.; Birlutiu, V.; Ichim, C.; Anderco, P.; Birsan, S. Fecal Microbiota Transplantation in Inflammatory Bowel Disease. Biomedicines 2023, 11. [Google Scholar] [CrossRef]
- Khoruts, A. Fecal Microbiota Transplantation–Early Steps on a Long Journey Ahead. Gut Microbes 2017, 8, 199–204. [Google Scholar] [CrossRef]
- Cammarota, G.; Ianiro, G.; Gasbarrini, A. 2014.
- Brandt, L.J.; Aroniadis, O.C.; Mellow, M.; Kanatzar, A.; Kelly, C.; Park, T.; Stollman, N.; Rohlke, F.; Surawicz, C. Long-Term Follow-up of Colonoscopic Fecal Microbiota Transplant for Recurrent Clostridium Difficile Infection. American Journal of Gastroenterology 2012, 107, 1079–1087. [Google Scholar] [CrossRef]
- Zhang, F.; Cui, B.; He, X.; Nie, Y.; Wu, K.; Fan, D.; Feng, B.; Chen, D.; Ren, J.; Deng, M.; et al. Microbiota Transplantation: Concept, Methodology and Strategy for Its Modernization. Protein Cell 2018, 9, 462–473. [Google Scholar] [CrossRef]
- Fecal Microbiota Transplants (FMT): Past, Present and Future Available online:. Available online: https://asm.org/Articles/2024/February/Fecal-Microbiota-Transplants-Past-Present-Future?utm_source=chatgpt.com (accessed on 23 November 2024).
- Khanna, S.; Tande, A.; Rubin, D.T.; Khoruts, A.; Kahn, S.A.; Pardi, D.S. Fecal Microbiota Transplantation for Recurrent C Difficile Infection During the COVID-19 Pandemic: Experience and Recommendations. Mayo Clin Proc 2021, 96, 1418–1425. [Google Scholar] [CrossRef]
- Tariq, R.; Pardi, D.S.; Bartlett, M.G.; Khanna, S. Low Cure Rates in Controlled Trials of Fecal Microbiota Transplantation for Recurrent Clostridium Difficile Infection: A Systematic Review and Meta-Analysis. In Proceedings of the Clinical Infectious Diseases; Oxford University Press, April 8 2019; Vol. 68; pp. 1351–1358. [Google Scholar]
- Pigneur, B.; Sokol, H. Fecal Microbiota Transplantation in Inflammatory Bowel Disease: The Quest for the Holy Grail. Mucosal Immunol 2016, 9, 1360–1365. [Google Scholar] [CrossRef]
- Moayyedi, P.; Surette, M.G.; Kim, P.T.; Libertucci, J.; Wolfe, M.; Onischi, C.; Armstrong, D.; Marshall, J.K.; Kassam, Z.; Reinisch, W.; et al. Fecal Microbiota Transplantation Induces Remission in Patients With Active Ulcerative Colitis in a Randomized Controlled Trial. Gastroenterology 2015, 149, 102–109.e6. [Google Scholar] [CrossRef]
- Yue, B.; Yu, Z.L.; Lv, C.; Geng, X.L.; Wang, Z.T.; Dou, W. Regulation of the Intestinal Microbiota: An Emerging Therapeutic Strategy for Inflammatory Bowel Disease. World J Gastroenterol 2020, 26, 4378–4393. [Google Scholar] [CrossRef] [PubMed]
- Markowiak, P.; Ślizewska, K. Effects of Probiotics, Prebiotics, and Synbiotics on Human Health. Nutrients 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Quigley, E.M.M. Prebiotics and Probiotics in Digestive Health. Clinical Gastroenterology and Hepatology 2019, 17, 333–344. [Google Scholar] [CrossRef]
- The Role of Prebiotics and Probiotics in Human Health: A Systematic Review with a Focus on Gut and Immune Health. Food & Nutrition Journal 2022, 7. [CrossRef]
- Carlson, J.L.; Erickson, J.M.; Lloyd, B.B.; Slavin, J.L.; Slavin, J. Health Effects and Sources of Prebiotic Dietary Fiber. Curr Dev Nutr 2018. [Google Scholar] [CrossRef]
- Prebiotics: Understanding Their Role in Gut Health Available online:. Available online: https://www.health.harvard.edu/nutrition/prebiotics-understanding-their-role-in-gut-health (accessed on 26 November 2024).
- Martinez, R.C.R.; Bedani, R.; Saad, S.M.I. Scientific Evidence for Health Effects Attributed to the Consumption of Probiotics and Prebiotics: An Update for Current Perspectives and Future Challenges. British Journal of Nutrition 2015, 114, 1993–2015. [Google Scholar] [CrossRef]
- Prajapati, K.; Bisani, K.; Prajapati, H.; Prajapati, S.; Agrawal, D.; Singh, S.; Saraf, M.; Goswami, D. Advances in Probiotics Research: Mechanisms of Action, Health Benefits, and Limitations in Applications. Systems Microbiology and Biomanufacturing 2024, 4, 386–406. [Google Scholar] [CrossRef]
- Mhanna, A.; Martini, N.; Hmaydoosh, G.; Hamwi, G.; Jarjanazi, M.; Zaifah, G.; Kazzazo, R.; Haji Mohamad, A.; Alshehabi, Z. The Correlation between Gut Microbiota and Both Neurotransmitters and Mental Disorders: A Narrative Review. Medicine (United States) 2024, 103, E37114. [Google Scholar] [CrossRef]
- Xiong, R.G.; Li, J.; Cheng, J.; Zhou, D.D.; Wu, S.X.; Huang, S.Y.; Saimaiti, A.; Yang, Z.J.; Gan, R.Y.; Li, H. Bin The Role of Gut Microbiota in Anxiety, Depression, and Other Mental Disorders as Well as the Protective Effects of Dietary Components. Nutrients 2023, 15. [Google Scholar]
- Kumar Yadav, M.; Kumari, I.; Singh, B.; Kant Sharma, K.; Kumar Tiwari, S. MINI-REVIEW Probiotics, Prebiotics and Synbiotics: Safe Options for next-Generation Therapeutics. [CrossRef]
- Yadav, M.; Sehrawat, N.; Sharma, A.K.; Kumar, S.; Singh, R.; Kumar, A.; Kumar, A. Synbiotics as Potent Functional Food: Recent Updates on Therapeutic Potential and Mechanistic Insight. J Food Sci Technol 2024, 61, 1–15. [Google Scholar] [CrossRef]
- Wang, G.; Ding, T.; Ai, L. Editorial: Effects and Mechanisms of Probiotics, Prebiotics, Synbiotics and Postbiotics on Intestinal Health and Disease. Front Cell Infect Microbiol 2024, 14. [Google Scholar] [CrossRef] [PubMed]
- Martin-Gallausiaux, C.; Marinelli, L.; Blottière, H.M.; Larraufie, P.; Lapaque, N. Conference on Diet and Digestive Disease Symposium 2: Sensing and Signalling of the Gut Environment: Scfa: Mechanisms and Functional Importance in the Gut. In Proceedings of the Proceedings of the Nutrition Society; pp. 120218037–49.
- Duncan, S.H.; Holtrop, G.; Lobley, G.E.; Calder, A.G.; Stewart, C.S.; Flint, H.J. Contribution of Acetate to Butyrate Formation by Human Faecal Bacteria. British Journal of Nutrition 2004, 91, 915–923. [Google Scholar] [CrossRef] [PubMed]
- Progress Report.
- Banfi, D.; Moro, E.; Bosi, A.; Bistoletti, M.; Cerantola, S.; Crema, F.; Maggi, F.; Giron, M.C.; Giaroni, C.; Baj, A. Impact of Microbial Metabolites on Microbiota–Gut–Brain Axis in Inflammatory Bowel Disease. Int J Mol Sci 2021, 22, 1–42. [Google Scholar] [CrossRef] [PubMed]
- Martinez, K.B.; Leone, V.; Chang, E.B. Microbial Metabolites in Health and Disease: Navigating the Unknown in Search of Function. Journal of Biological Chemistry 2017, 292, 8553–8559. [Google Scholar] [CrossRef] [PubMed]
- Jamka, M.; Kokot, M.; Kaczmarek, N.; Bermagambetova, S.; Nowak, J.K.; Walkowiak, J. The Effect of Sodium Butyrate Enemas Compared with Placebo on Disease Activity, Endoscopic Scores, and Histological and Inflammatory Parameters in Inflammatory Bowel Diseases: A Systematic Review of Randomised Controlled Trials. Complement Med Res 2021, 28, 344–356. [Google Scholar] [CrossRef]
- Sarada, J. and Anju, S. and A.Y. and A.K. Microbiome Therapeutics: Emerging Concepts and Challenges. In Microbiome in Human Health and Disease; Bramhachari, P.V., Ed.; Springer Singapore: Singapore, 2021; ISBN 978-981-16-3156-6. [Google Scholar]
- Plaza-Diaz, J.; Ruiz-Ojeda, F.J.; Gil-Campos, M.; Gil, A. Mechanisms of Action of Probiotics. In Proceedings of the Advances in Nutrition; Oxford University Press, January 1 2019; Vol. 10; pp. S49–S66. [Google Scholar]
- Sheth, R.U.; Cabral, V.; Chen, S.P.; Wang, H.H. Manipulating Bacterial Communities by in Situ Microbiome Engineering. Trends in Genetics 2016, 32, 189–200. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Y.; Zheng, P.; Sun, J.; Wang, M. Microbial Base Editing: A Powerful Emerging Technology for Microbial Genome Engineering. Trends Biotechnol 2021, 39, 165–180. [Google Scholar] [CrossRef]
- Reshaping the Gut Microbiome Using New Genetic Tools. Available online: https://asm.org/articles/2021/january/reshaping-the-gut-microbiome-using-new-genetic-too (accessed on 27 November 2024).
- Crispr-Enhanced Viruses Are Being Deployed Against UTIs. Available online: https://www.wired.com/story/crispr-enhanced-viruses-are-being-deployed-against-utis/?utm_source=chatgpt.com (accessed on 27 November 2024).
- New Medicine Could Prevent Disease before It Begins — but at a Cost. Available online: https://www.ft.com/content/0df1781d-c224-40d2-9bf1-43bf3d13b2c1?utm_source=chatgpt.com (accessed on 27 November 2024).
- IGI’s ‘Audacious’ New Frontier for CRISPR: Editing Microbiomes for Climate and Health. Available online: https://innovativegenomics.org/news/audacious-project-crispr-microbiome/ (accessed on 27 November 2024).
- Dabhi, M.; Patel, R.; Shah, V.; Soni, R.; Saraf, M.; Rawal, R.; Goswami, D. Penicillin-Binding Proteins: The Master Builders and Breakers of Bacterial Cell Walls and Its Interaction with β-Lactam Antibiotics. J Proteins Proteom 2024, 15, 215–232. [Google Scholar] [CrossRef]
- Serral, F.; Castello, F.A.; Sosa, E.J.; Pardo, A.M.; Palumbo, M.C.; Modenutti, C.; Palomino, M.M.; Lazarowski, A.; Auzmendi, J.; Ramos, P.I.P.; et al. From Genome to Drugs: New Approaches in Antimicrobial Discovery. Front Pharmacol 2021, 12. [Google Scholar] [CrossRef]
- Alkatheri, A.H.; Yap, P.S.X.; Abushelaibi, A.; Lai, K.S.; Cheng, W.H.; Erin Lim, S.H. Microbial Genomics: Innovative Targets and Mechanisms. Antibiotics 2023, 12. [Google Scholar] [CrossRef]
- León-Buitimea, A.; Balderas-Cisneros, F. de J.; Garza-Cárdenas, C.R.; Garza-Cervantes, J.A.; Morones-Ramírez, J.R. Synthetic Biology Tools for Engineering Microbial Cells to Fight Superbugs. Front Bioeng Biotechnol.
- Tan, J.; Fu, B.; Zhao, X.; Ye, L. Novel Techniques and Models for Studying the Role of the Gut Microbiota in Drug Metabolism. Eur J Drug Metab Pharmacokinet 2024, 49, 131–147. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Chen, Y.; Huang, W.; Zhou, H.; Zhang, W. Drug-Microbiota Interactions: An Emerging Priority for Precision Medicine. Signal Transduct Target Ther 2023, 8. [Google Scholar] [CrossRef] [PubMed]
- Gulnaz, A.; Chang, J.E.; Maeng, H.J.; Shin, K.H.; Lee, K.R.; Chae, Y.J. A Mechanism-Based Understanding of Altered Drug Pharmacokinetics by Gut Microbiota. J Pharm Investig 2023, 53, 73–92. [Google Scholar] [CrossRef]
- Orukotan Kesioluwa Eunice and Elughi, G.N. and Elughi, G.N. and A.B.S. and A.A.D. and A.E.F. and O.O. Plant Microbiome Engineering: Principles, Methods, and Current Trends. In Biotechnological Approaches to Sustainable Development Goals; Isibor Patrick Omoregie and Akinduti, P. and O.S.U. and P.J.O., Ed.; Springer Nature Switzerland: Cham, 2023; ISBN 978-3-031-33370-5. [Google Scholar]
- Krishnan Aswathy and Dasgupta, A. Microbiome Engineering towards Disease Therapy. In Industrial Microbiology and Biotechnology: A New Horizon of the Microbial World; Verma, P., Ed.; Springer Nature Singapore: Singapore, 2024; ISBN 978-981-97-6270-5. [Google Scholar]
- InCoGenTEC: Intrinsic Control for Genome and Transcriptome Editing in Communities Available online:. Available online: https://www.genomicscience.energy.gov/snl-incogentec/#:~:text=The%20Intrinsic%20Control%20for%20Genome,controllably%20transform%20bacteria%20with%20enhanced (accessed on 27 November 2024).
- European Medicines Agency: Clinical Efficacy and Safety Guidelines Available online:. Available online: https://www.ema.europa.eu/en/human-regulatory-overview/research-development/scientific-guidelines/clinical-efficacy-safety-guidelines (accessed on 27 November 2024).
- Govender, P.; Ghai, M. Population-Specific Differences in the Human Microbiome: Factors Defining the Diversity. Gene 2025, 933. [Google Scholar] [CrossRef]
- Fang, C.; Fang, W.; Xu, L.; Gao, F.; Hou, Y.; Zou, H.; Ma, Y.; Moll, J.M.; Yang, Y.; Wang, D.; et al. Distinct Functional Metagenomic Markers Predict the Responsiveness to Anti-PD-1 Therapy in Chinese Non-Small Cell Lung Cancer Patients. Front Oncol 2022, 12. [Google Scholar] [CrossRef]
- Sharpton, S.R.; Schnabl, B.; Knight, R.; Loomba, R. Current Concepts, Opportunities, and Challenges of Gut Microbiome-Based Personalized Medicine in Nonalcoholic Fatty Liver Disease. Cell Metab 2021, 33, 21–32. [Google Scholar] [CrossRef]
- Verstockt, B.; Parkes, M.; Lee, J.C. How Do We Predict a Patient’s Disease Course and Whether They Will Respond to Specific Treatments? Gastroenterology 2022, 162, 1383–1395. [Google Scholar] [CrossRef]
- Acharjee, A.; Singh, U.; Choudhury, S.P.; Gkoutos, G. V. The Diagnostic Potential and Barriers of Microbiome Based Therapeutics. Diagnosis 2022, 9, 411–420. [Google Scholar] [CrossRef]
- Liang, D.; Leung, R.K.K.; Guan, W.; Au, W.W. Involvement of Gut Microbiome in Human Health and Disease: Brief Overview, Knowledge Gaps and Research Opportunities. Gut Pathog 2018, 10. [Google Scholar] [CrossRef]
- Ma, Y.; Chen, H.; Lan, C.; Ren, J. Help, Hope and Hype: Ethical Considerations of Human Microbiome Research and Applications. Protein Cell 2018, 9, 404–415. [Google Scholar] [CrossRef]
- Daloiso, V. , Minacori, R. G. ( 19(17), 3173–80.
- Shukla, V.; Singh, S.; Verma, S.; Verma, S.; Rizvi, A.A.; Abbas, M. Targeting the Microbiome to Improve Human Health with the Approach of Personalized Medicine: Latest Aspects and Current Updates. Clin Nutr ESPEN 2024, 63, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Aljeradat, B.; Kumar, D.; Abdulmuizz, S.; Kundu, M.; Almealawy, Y.F.; Batarseh, D.R.; Atallah, O.; Ennabe, M.; Alsarafandi, M.; Alan, A.; et al. Neuromodulation and the Gut–Brain Axis: Therapeutic Mechanisms and Implications for Gastrointestinal and Neurological Disorders. Pathophysiology 2024, 31, 244–268. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.; Singh, S.K. The Microbiome Modulates the Immune System to Influence Cancer Therapy. Cancers (Basel) 2024, 16. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.M.; Redvers, N.; Camargo, A.; Bosch, C.A.; Breed, M.F.; Brenner, L.A.; Carney, M.A.; Chauhan, A.; Dasari, M.; Dietz, L.G.; et al. 2022.
- Chen, H.; Zhao, Q.; Zhong, Q.; Duan, C.; Krutmann, J.; Wang, J.; Xia, J. Skin Microbiome, Metabolome and Skin Phenome, from the Perspectives of Skin as an Ecosystem. Phenomics 2022, 2, 363–382. [Google Scholar] [CrossRef]
- Baker, J.L. Illuminating the Oral Microbiome and Its Host Interactions: Recent Advancements in Omics and Bioinformatics Technologies in the Context of Oral Microbiome Research. FEMS Microbiol Rev 2023, 47. [Google Scholar] [CrossRef]
Figure 1.
Gut microbiota in health and diseases (Created in https://BioRender.com).
Figure 1.
Gut microbiota in health and diseases (Created in https://BioRender.com).
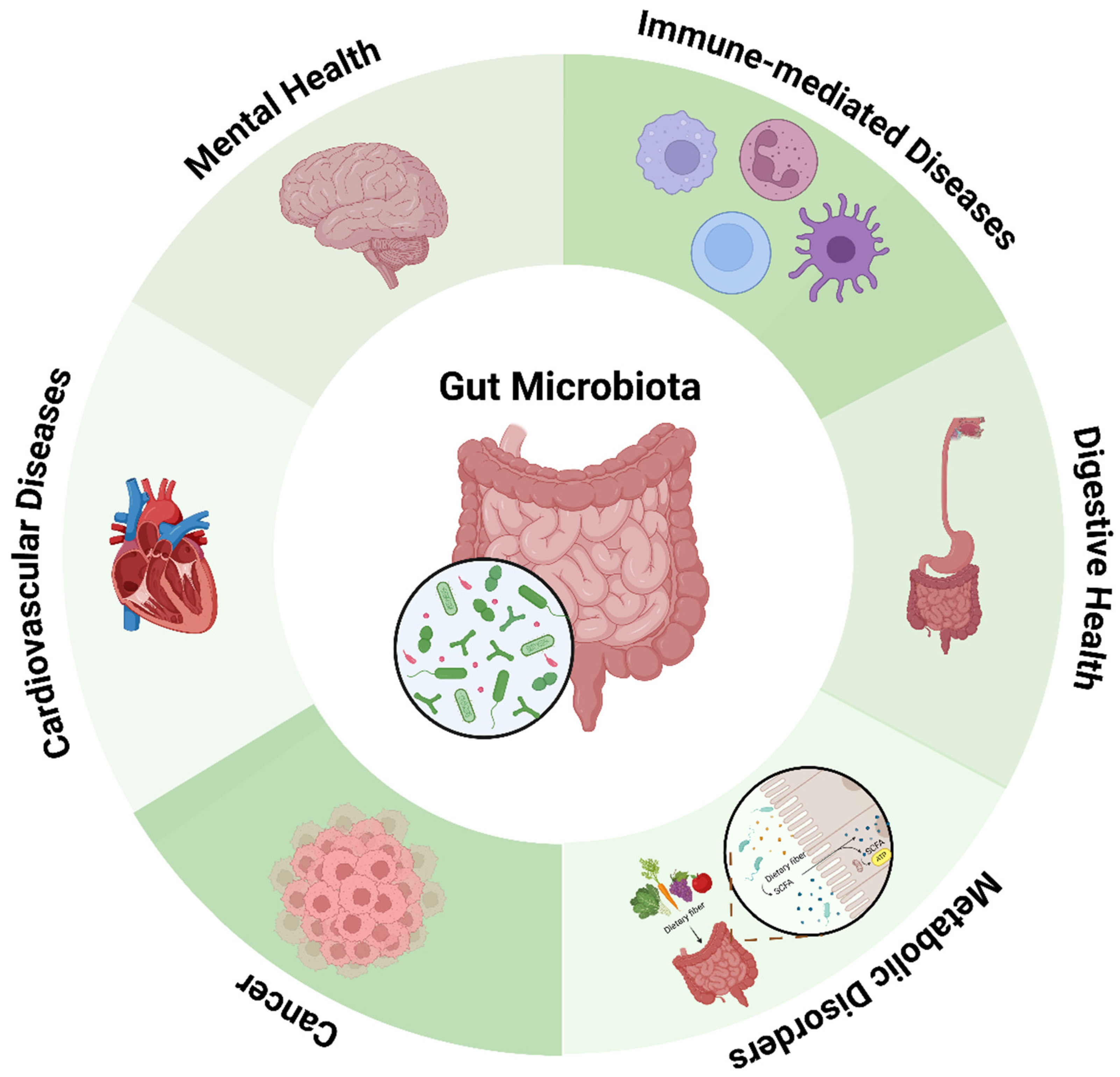
Figure 2.
Gut Microbiota Restoration through Fecal Microbiota Transplantation (FMT) in patients with Clostridioides difficile infection (Created in https://BioRender.com).
Figure 2.
Gut Microbiota Restoration through Fecal Microbiota Transplantation (FMT) in patients with Clostridioides difficile infection (Created in https://BioRender.com).
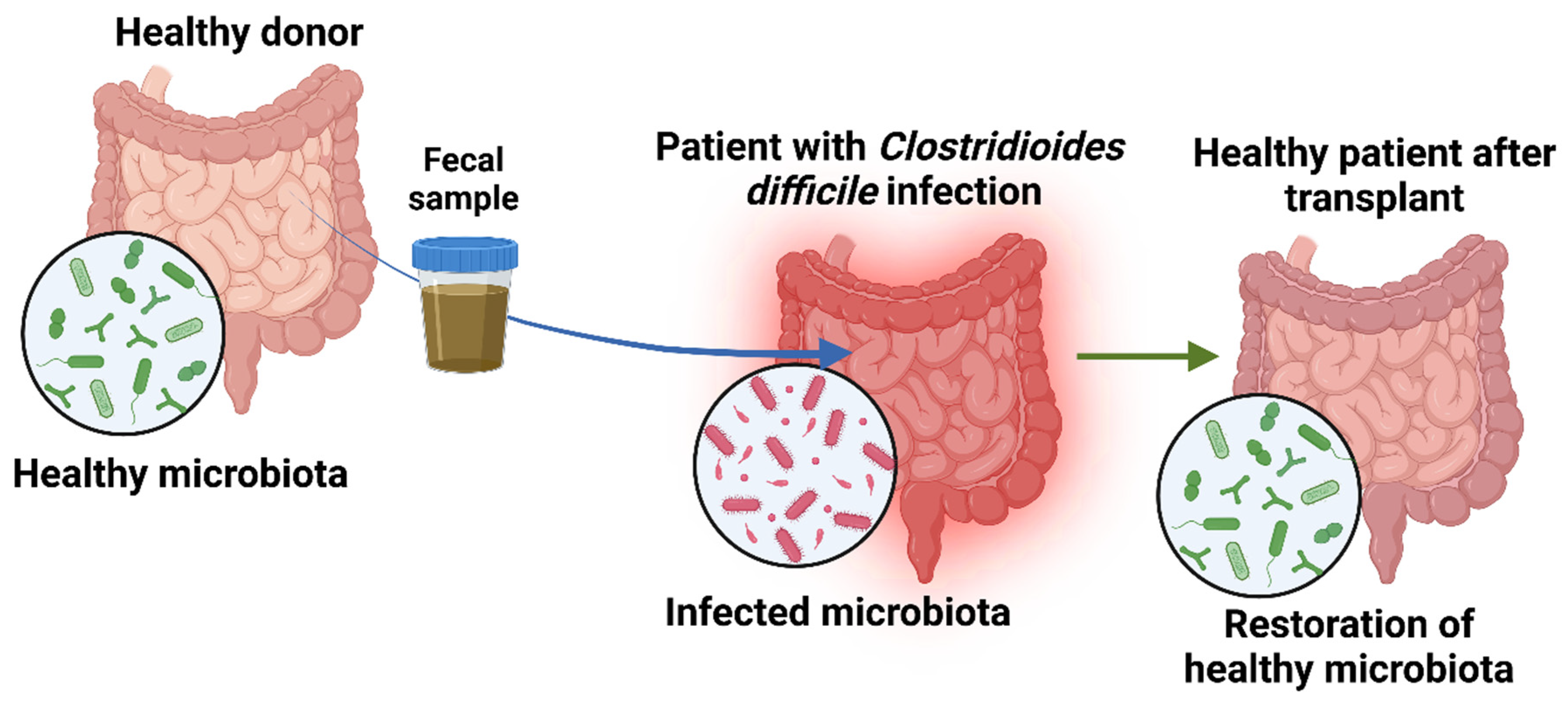

Table 1.
Key bacterial strains and clinical implications associated with microbiome-related disorders.
Table 1.
Key bacterial strains and clinical implications associated with microbiome-related disorders.
| Disorder | Associated Bacterial Strains | Alterations | Clinical Implications | References |
|---|---|---|---|---|
| Irritable Bowel Syndrome (IBS) | Bifidobacterium spp., Lactobacillus spp., Methanobrevibacter smithii, Escherichia coli | Decreased Bifidobacterium and Lactobacillus; increased Methanobrevibacter and E. coli. | Increased gas production, altered motility, and inflammation. | [78] |
| Inflammatory Bowel Disease (IBD) | Faecalibacterium prausnitzii, Roseburia spp., Eubacterium rectale, Akkermansia muciniphila, Escherichia coli | Depletion of anti-inflammatory species (F. prausnitzii, Roseburia); overgrowth of pathogenic E. coli. | Loss of gut barrier integrity, chronic inflammation. | [79] |
| Clostridioides difficile Infection | Clostridioides difficile, reduced Bacteroidetes and Firmicutes diversity | Overgrowth due to disrupted microbiota (e.g., post-antibiotics). | Severe diarrhea and colitis. | [79] |
| Helicobacter pylori Infection | Helicobacter pylori | Colonizes the stomach lining, reduces protective microbial diversity. | Gastritis, ulcers, increased gastric cancer risk. | [80] |
| Colorectal Cancer (CRC) | Fusobacterium nucleatum, Bacteroides fragilis, Escherichia coli | Enrichment of F. nucleatum and B. fragilis. | Promotes tumorigenesis via inflammation and DNA damage. | [81] |
| Diverticulitis | Bacteroides fragilis, Escherichia coli, Enterococcus spp. | Altered microbial diversity, increased inflammation. | Pain, fever, abscess formation. | [82] |
| Metabolic Disorders | Akkermansia muciniphila, Bacteroidetes spp., Firmicutes spp. | Reduced Akkermansia; altered Firmicutes/Bacteroidetes ratio. | Obesity, insulin resistance, increased inflammation. | [83] |
| Celiac Disease | Bifidobacterium spp., Lactobacillus spp., increased Enterobacteriaceae | Reduced beneficial bacteria; increased pathogenic strains. | Triggers inflammatory responses in the gut. | [84,85] |
| Autism Spectrum Disorders (ASD) | Bacteroides spp., Clostridium spp., Prevotella spp. | Decreased Prevotella; increased Clostridium. | Altered gut-brain axis signaling, behavioral symptoms. | [86,87] |
| Cardiovascular Diseases | Lactobacillus spp., Bifidobacterium spp., Firmicutes | Increased trimethylamine-N-oxide (TMAO)-producing species. | Links to atherosclerosis and hypertension. | [88,89] |
Table 2.
Comparative Overview of Microbial Metabolite-Based Treatments and Probiotic Treatments.
| Aspect | Microbial Metabolite Based Therapies | Probiotic Treatment |
|---|---|---|
| Definition | Utilizes metabolites produced by gut microbiota (e.g., SCFAs) to exert therapeutic effects [119] | Involves the administration of live microorganisms that confer health benefits when consumed in adequate amounts [120] |
| Mechanism | Directly modulates host signaling via metabolites, influencing immune responses and gut health [119,120] | Restores or maintains a healthy gut microbiota, inhibiting pathogenic bacteria and enhancing mucosal barrier function [120] |
| Efficacy in Disease Treatment | Effective for conditions like IBD, metabolic disorders, and neurological conditions through targeted action [119] | Variable efficacy; beneficial for gastrointestinal disorders like diarrhea and IBD, but effectiveness can be strain-specific [120] |
| Advantages | Provides targeted therapy; may overcome limitations of probiotics related to colonization and individual microbiomes [119]. | Generally safe; can restore gut flora balance effectively in many individuals [120]. |
| Limitation | Less research on long-term effects; potential need for personalized approaches based on specific metabolites. | Variable efficacy based on strain and individual microbiome composition; may pose risks for immunocompromised individuals |
| Safety Profile | Generally low toxicity; fewer risks associated with live organisms. | Safe for most; caution advised in immunocompromised individuals or those with severe underlying conditions |
| Administration | Can be administered as stable compounds (e.g., supplements). | Requires viable organisms; may have storage and viability issues. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



