
Submitted:
09 December 2024
Posted:
10 December 2024
You are already at the latest version
Abstract
Food is a crucial component affecting the health of individuals, which may have the potential to expand lifespan. It has been shown that the long lifespan may be related to the fine-tuned autophagy. In general, the suitable autophagy could play a significant role in an anti-aging biological exertion of the host. A member of serine and threonine kinases, AMPK, could play vital roles within the autophagy signaling pathway in various cells. In addition, alterations in the kinase activity of AMPK have been shown to be connected to several pathologies of aging related diseases. Therefore, autophagy could control the lifespan-related homeostasis within the host from cells to a body via the modification of AMPK. The design of the diet and/or nutrition targeting the AMPK would be a possibility to expand the lifespan. Some analyses of the molecular biology underlying the autophagy suggest that supplementation of accurate nutraceuticals as well as dietary restriction, mild fasting, and/or appropriate physical exercise could modulate the AMPK signaling which may be advantageous for the life-extension with the alteration of autophagy. Remarkably, it has been revealed that several non-coding RNAs (ncRNAs) might also play significant roles in the regulation of autophagy. In addition, the production of some ncRNAs may be associated with the alteration of gut microbiota with certain dieting. Therefore, the modulation of AMPK action with ncRNAs via choosing the relevant diets could be a therapeutic tactic for grasping the longevity, also accompanying with the reduced risk of several aging-related diseases.
Keywords:
autophagy
; AMPK
; ncRNA
; miRNA
; aging
; aging-related diseases
; longevity
1. Introduction
Aging may refer to gradual weakening processes of physiological integrity and/or functionality, which might consequently increase susceptibility to various aging-related diseases. Aging is a flexible process that can be determined by genetic, environmental, and/or lifestyle factors [1]. Among them, a food nutrition in lifestyle factors may be an important component affecting the health, which could also enlarge the lifespan of human individuals [2]. Interestingly, it has been shown that caloric restriction can significantly extend the lifespan of various species including humans [3,4]. Additionally, appropriate physical exercises may also improve one’s health markers, which has been believed to prolong the lifespan [5]. In their biological mechanisms, it has been discovered that the lifespan may be related with an adjusted autophagy in cells [6]. Autophagy can actually play a vital role in the anti-aging contributions by degrading broken proteins and/or organelles [7]. Therefore, the autophagy has been commonly thought to enhance the health levels and/or the lifespan expansion [8]. In addition, an age-related decrease of autophagy-power might be an important initiator of the cellular senescence that may lead to the development of various aging related diseases such as diabetes and/or cancers [9]. The autophagy is also considered as a cell survival mechanism that an appropriate activity of autophagy could contribute to the high duration of various cells/tissues in organisms [10].
Current studies have revealed the role of non-coding RNAs (ncRNAs) such as microRNAs (miRNAs) in regulating cellular integrity by targeting various components in the autophagy pathway. In addition, the role of the ncRNAs in controlling aging processes has also been demonstrated with the discovery of the specific miRNA that regulate lifespan in the nematode Caenorhabditis elegans (C. elegans) [11]. In addition, the degradation of the miRNA has been shown to contribute to the aging related neurodegeneration, affecting the mechanisms responsible for the development of various neurodegenerative diseases [12]. Hence, the neurodegeneration can be considered as an RNA disorder where the miRNA plays a critical role in the pathology [13]. In the brain, dopaminergic neurons may be dependent on the functionality in the miRNA relevant network observed in vitro and in vivo [14]. The miRNA could also provide therapeutic targets for the neurodegenerative disease. In general, miRNAs are single-stranded RNA molecules that regulate gene expression by complementary binding sequences in the three prime untranslated region (3′UTR) of the target genes, potentially modifying a disease phenotype [15]. Actually, miRNAs could regulate various biological processes, such as cell proliferation, differentiation, apoptosis and cell migration by binding to specific sites of target mRNAs [16]. Interestingly, the silent mating type information regulation 2 homolog-1 (SIRT1) may also be a target of miR-34a [17], which could affect the cellular physiological senescence and/or aging [18,19]. Up to the present time, several miRNAs have been found to be regulated within the process of aging, which have appeared as key regulators of aging at the cellular and/or body levels [20]. A better comprehension of the interaction with the relevant ncRNAs during the autophagy and/or the process of aging related pathologies may be imperative for providing the prevention of aging related diseases as well as for promoting the long-term lifespan. (Figure 1) This review would summarize and discuss the most important nutritional components for the improved autophagy, which might be also associated with longevity. In particular, it should be reasonable to seek safe and practical nutraceuticals which could induce applicable autophagy for achieving the longevity. This review might also be beneficial for designing a daily diet in order to decrease aging related tissue/organ damages.
2. Relationship Between Autophagy and Aging Related Diseases by the Modulation of AMPK Signaling Pathway
Aging may be characterized with the functional weakening of an individual’s cells/tissues/organs, in which aging related diseases such as cardiovascular disease, diabetes, neurodegenerative diseases, and/or cancers might appear. Several characteristics of these diseases and/or aging are often associated with the alteration of autophagy. In general, autophagy is an evolutionarily conserved host defense mechanism, which may also play important roles for some biological key processes. For example, autophagy may involve several cell components being sequestered in auto-phagosomes for molecular recycling [21]. Consequently, suitable induction of the autophagy can extend the lifespan, whereas its deficiency may reduce the lifespan [22]. Therefore, continued and diminished autophagy may initiate an aging-related alterations of cells/tissues, following various disorders in tissues/organs [23]. In addition, altered autophagy may lead to the spread of abnormal proteins to several organelles, which can eventually exacerbate further aging-related diseases including neurodegenerative diseases, metabolic diseases, cardiovascular diseases, and/or immune related diseases. [24,25]. For example, it has been described that a coronary artery disease may be predisposed by levels of autophagy [26]. Accordingly, appropriate induction of autophagy can contribute to the protection against heart failures [27,28]. In addition, autophagy could also protect against neuronal cell-damages in the brain of seniors [29]. For example, age-associated memory waning can be decreased by the autophagic effect of spermidine [30]. Consistently, autophagy has been reported to improve the age-related dementia in the seniors [31]. Furthermore, autophagy may be involved in the regulation of metabolic diseases such as obesity and/or metabolic syndrome [32]. Autophagy could even protect against the ageing-related skeletal muscle atrophy by the modification of autophagy [33]. Levels of autophagy may be gradually decreased in mature lymphocytes of seniors. Remarkably, it has been shown that improved autophagy could be connected to reverse the immune senescence in humans [34]. Consequently, autophagy may be essential for the maintenance of health homeostasis with several advantages to the longevity of the host [35].
Adenosine monophosphate-activated protein kinase (AMPK) might be dominant in a coordinated intracellular signaling network sustaining the homeostasis of cells. [36]. In particular, the AMPK can adjust an energy condition by approving the ATP production and by regulating the ATP consumption in cells [36,37]. The AMPK can be involved in directing an inhibition signaling pathway with the mechanistic/mammalian target of rapamycin (mTOR), which may successfully decrease the apoptosis by regulating the autophagy [38]. Also, AMPK can start the autophagy via the autophagy activating kinase 1 (ULK1) [38,39]. Activation of the serine/threonine kinase ULK1 is an initiation for the formation of phagophores, which are then extended to autophagosomes [40]. The AMPK and mTOR complex 1 (mTORC1) can work as the key mTOR antagonists controlling the ULK1 activity [41]. By external growth factors stimulation and the following activation of the PI3K/AKT and AMPK signaling, the mTORC1 could control the phosphorylation and/or the activation of the ULK1 [42]. (Figure 2) Therefore, the AMPK may be a key regulator of cellular metabolism during several oxidative stresses keeping energy homeostasis, which might be recognized as an energy sensor due to its critical roles [43,44]. Remarkably, it has been well-known that the AMPK can provide several protective effects in cells by controlling the energy homeostasis. In addition, the AMPK can organize several cellular key processes including protein synthesis, DNA repair, and cell differentiation/proliferation [45,46]. Therefore, the AMPK could achieve the endogenous cell protection, which can respond to various cell damages including oxidative stresses [47]. These vital roles of the AMPK in cells/tissues/organs may be indispensable for maintaining the physiological/pathological function [48,49]. Unquestionably, the AMPK should be expected to play a substantial role for the longevity. Physical aging may be also renovated by modulating the AMPK signaling with the alteration of autophagy [50].
3. Several ncRNAs Involved in the Longevity via the Modulation of Autophagy
ncRNAs can induce the degradation of mRNAs for the repression of their translation, which can direct to the specific mRNA transcript through the sequence-specific binding to the 3’ untranslated region (3′UTR) of the mRNA [51]. In general, the biogenesis of ncRNAs including miRNAs may start with their transcription in the nucleus [52], which is further processed by an RNase enzyme to generate a precursor of the ncRNA [53]. The precursors of ncRNAs are exported to the cytoplasm to generate a mature ncRNA with the help of another RNase enzyme DICER [54]. Some of ncRNAs can bind its specific target mRNA to form the RNA-induced silencing complex (RISC) [55]. In these ways, some of ncRNAs could regulate the autophagy related mRNAs [56]. In addition, certain ncRNAs have been shown to be upregulated during the age-associated diseases such as Parkinson’s disease [57]. In particular, an increased expression of miR-301b, miR-26b, and miR-106a may regulate the autophagy, which might aggravate the alpha-synuclein pathology of Pardinson’s disease [57]. These ncRNAs could also regulate protein folding, which may serve as potential therapeutic targets to treat the protein misfolding of several age-associated diseases. Autophagy may also play an imperative role in the pathogenesis of age-related macular degeneration, which has to do with the harmful aggregation of damaged proteins. In C. elegans, secreted ncRNAs such as miR-29 and/or miR-83 could improve the age-related decrease of autophagy across different tissues for the longevity [58]. Additionally, loss-of-function mutations in miR-34 can significantly extend the lifespan, which has been shown to bind with the autophagy related gene [59]. Some ncRNAs are also known to play role in ultraviolet ray A (UVA) and ultraviolet ray B (UVB)-induced photo-related aging in the skin fibroblasts by regulating the autophagy [60]. In this case, the level of miR-23 might be upregulated during the skin exposure to UVA/B irradiation, which is a positive regulator of the autophagy [60]. Inhibition of miR-23 could stimulate the activation of autophagy, which might protect the skin fibroblasts from UVA/B-induced cellular senescence [60]. The knockdown of miR-23 is also known to increase autophagy in lens epithelial cells, which might alleviate oxidative stresses [61]. Therefore, the miR-23 may play a role in age-associated increase in oxidative stresses in lens epithelial cells, where one of the prime causes of eye cataracts with aging might occur. In addition, the inhibition of miR-331-30 and miR-9-5p have been shown to prevent the progression of Alzheimer’s disease by activating the autophagy essential for the clearance of amyloid beta proteins [62]. It has been shown that miR-331-3p and miR-9-5p can affect the autophagy receptors, which may act as potential markers of the Alzheimer’s disease [62]. Another miRNA, miR-101, could also inhibit the autophagy. The reduction of miR-101 has been reported in the hippocampus of the brain with Alzheimer's disease [63]. Mimicking age-associated loss of the miR-101 in hippocampal neurons may bring cognitive decline in the model mice of Alzheimer's disease [63]. (Figure 1)
The featuring role of miRNAs for the regulation of autophagy in other neurodegenerative diseases has been reviewed elsewhere [64]. Several miRNAs are dysregulated in these aging-related diseases with the alteration of autophagy [65]. Some miRNAs may also play a vital role in cardiovascular diseases via the alteration of autophagy for the longevity [66]. The induction of autophagy by some drugs such as rapamycin may be controlled by several post-transcriptional mechanisms including the modulation of miRNAs for the longevity [66,67]. Interestingly, the alliance of miR-506 may be related to the development of atherosclerosis and autophagy [68,69]. In addition, it has been shown that a football training might down-regulate the expression of miR-1303, indicating the molecular mechanisms of the physical training linking to autophagy and/or the longevity [70]. Furthermore, it has also been shown that a secreted miR-29 or miR-83 could manipulate the age-related decrease with the modulation of the autophagy in C. elegans [58,71]. Up to the present time, various miRNAs have been detected to be regulated with aging, suggesting that some of these may become beneficial indicators and/or regulators of aging [72].
4. Possible Tactics with Certain Dieting for the Longevity
Agreed with the noteworthy role of autophagy in the pathogenesis of aging and aging related diseases, the autophagy could work as a promising therapeutic target. In fact, several inhibitors such as metformin can target AMPK pathway, thereby mediating the modulation of autophagy for the treatment of several aging related diseases. Hence, the promotion or inhibition of autophagy could be actually used to get the longevity. Remarkably, it has been shown that modulation of autophagy might be useful both in improving cardiac function and in treating cardiovascular disease [73]. For example, a study in animal model of an aging heart has shown that mild fasting with the induction of autophagy may advance the cardiac function and/or the durability of heart, which may be linked to the clearance of damaged cellular components by the autophagy [73].
A popular form of repeated mild fasting has been confirmed to suggest multiple health profits, including an extension of healthy lifespan in preclinical models, in which several miRNAs may be expressed to work for [74]. Remarkably, fasting are also effective in the treatment of various solid tumors in mice models [75]. Several cancers could be treated for the prevention of cell proliferation by increasing cellular apoptosis via the autophagy modulatory mechanism of several miRNAs [76]. Mild fasting could activate the AMPK signaling for the modulation of autophagy [74,76]. The ghrelin release can control the mTOR signaling pathway, which could subsequently control the autophagy [77]. In addition, the ghrelin could also stimulate glucose metabolism for energy production in the brain, thereby probably preventing the deterioration of memory function in dementia [78]. Interestingly, it has been suggested that the ghrelin can attenuate the TGFβ-induced fibrosis via the miR-125a-5p action by antagonizing the TGF receptor signaling [79].
With regard to the AMPK signaling for the modulation of autophagy, the nutraceutical berberine, a compound observed in a diverse range of herbs used in Chinese traditional medicine, has shown beneficial activity for the regulation of hyperlipidemia and type 2 diabetes via the activation of the AMPK signaling [80]. Actually, berberine could stimulate the AMPK in a manner equivalent to the metformin [81]. Therefore, the capability of berberine to promote the autophagy has been recognized in rodent animal models in vivo [82]. The berberine could protect against several tissues injury by inducing the autophagy. In fact, it has been shown that berberine may possess a therapeutic effect on a cardiovascular disease [83]. Berberine may also serve the cardioprotective effect for the myocardial damage from an ischemia/reperfusion of heart by inducing the autophagy [84,85]. The miR-29b expression can be increased by the berberine usage, which may be abolished by the treatment with AMPK inhibitors [86]. Flavonoids are broadly present in grains, vegetables, fruits, and medicinal plants, which may have the potential to work as protective mediators against several aging related diseases. For example, specific flavonoids such as quercetin, puerarin, luteolin, and hesperidin have verified a cardioprotective activity in animal models [87]. In addition, quercetin can also induce neurogenesis enhancing the longevity of neuronal cells probably by modulating the AMPK signaling pathway [88]. The antioxidative nutraceutical components in Humulus japonicus, such as luteolin have been shown to be able to scavenge reactive oxygen species (ROS) within various cellular systems including mitochondria [89]. In neurodegenerative diseases such as Alzheimer’s disease, it has been shown that these flavonoids have a potential to modify the regulation of several miRNAs [90]. For example, there are considerable alterations in the number of miRNAs during antioxidant responses triggered by flavonoids [14,91]. In addition, neuronal cells treated with the flavonoid have exhibited that 14 miRNAs linked to the antioxidant system have altered the expression brought by oxidative stresses [92]. It has been suggested that miRNA-dependent anti-inflammatory mechanisms can validate the neuroprotective effects of flavonoids including quercetin [93]. Therefore, these modifications of miRNAs may indicate that certain flavonoids treatment can induce the differential expression of miRNAs in specific brain regions that can contribute to the development/decrease of several neurodegenerative diseases [94]. Similarly, luteolin can improve the lipid accumulation in nonalcoholic fatty liver disease (NAFLD) by increasing the oxidation of fatty acids via the increased mitochondrial biogenesis by upregulating the AMPK signaling [95]. Furthermore, puerarin has been shown to be a promising flavonoid for increasing the longevity of Drosophila melanogaster by stimulating the autophagy [96]. Interestingly, it has been shown that the puerarin can decrease the level of miR-7, which may activate the AMPK signaling [97]. It has been shown that hesperidin could also slow down aging speed with the prolonged longevity via the modulation of the AMPK signaling [98,99]. Damaged lung tissue may show a substantial increase of several miRNAs level within the lung injury place in mice, suggesting that the hesperidin has a protective effect with the modulation of miRNAs for the improved autophagy [100]. Likewise, it has been described that resveratrol has also a protective effect on the cardiac dysfunction by modifying the autophagy [101]. In line with this, resveratrol could protect mitochondria by modulating autophagy through the alteration of the AMPK signaling in myocardial cells [102]. Additionally, resveratrol can inhibit hyperglycemia-induced cardiomyocyte hypertrophy by reducing oxidative stresses via the stimulation of autophagy to sustain the mitochondrial homeostasis [103]. Resveratrol could also improve retinal arterioles, which can eventually protect against aging related retinal neurodegeneration via the alteration of the AMPK dependent signaling pathway [104]. In these ways, certain nutraceutical treatments may indeed have potential for the regulation of aging related diseases as well as for the longevity. (Figure 3)
5. Future Perspectives
Accumulating data suggest that regulatory ncRNAs including various miRNAs, circRNAs, and lncRNAs can affect the host-microbiota as well as the microbiota-associated diseases including cancers and/or diabetes [105]. Interestingly, the expression of several ncRNAs in the host could be repressed by certain gut microbiota [106]. It has also been shown that there are intricate connections between the gut microbiota and the expression of ncRNAs in the host, suggesting that gut microbiota may have a potential role for the protection against aging related diseases [107]. The intricate collaboration could even regulate the growth of cancer cells [108]. (Figure 3) Some studies have demonstrated that the miR-30a-3p expression exerts tumor-suppressive functions in some forms of cancers [109]. In addition, the ectopic expression of miR-30a-3p could also attenuate the aggressiveness of cancer cells [109]. Interestingly, the miR-30a-3p could enhance the chemosensitivity of several cancer cells [110]. Actually, the miR-30a-3p expression can repress cell growth, migration, and/or inflammatory responses in fibroblastic cells via the alteration of autophagy [110]. Interestingly, the nutrient starvation may induce a considerable intensification of the autophagy by Ulk1 dephosphorylation with the decreased expression of the miR-30a-3p [111]. In contrast, the upregulation of the miR-30a-3p could reverse the effects of autophagy with the use of rapamycin [112]. Rapamycin can promote the autophagy, which has therefore been used as a potent inducer of the autophagy [112]. Thus, some ncRNAs could regulate the autophagy. It has been shown that the downregulated miR-30a-3p expression could also promote the autophagy in mice mammary glands [113]. On the contrary, the autophagy may be inhibited by the overexpression of miR-30a-3p in mammary glands [113]. Interestingly, the increased expression of miR-34a or the inhibited autophagy may be associated with aging and/or metabolic syndrome [114]. The miR34a could repress the expression of the nicotinamide phosphoribosyltransferase gene, which may lead to reduced levels of the SIRT1 activity [115]. It has been shown that inflammatory transcription factors such as nuclear factor-kappa B (NF-κB) may be involved in the expression of miR-34a [116]. Interestingly, the upregulation of miR-34a could increase the activity of NF-κB in the same way [117]. Therefore, the overexpression of the miR-34a can promote the apoptosis in various cancer cells [118]. Instead, the inhibition of the miR-34a expression can not only increase the expression of SIRT1 to induce the proper activity of autophagy but also strengthen the ability of cells to oppose the apoptosis. Consequently, the downregulation of miR-34a could further improve the outcomes of several therapies against the progression of aging related diseases including cancers. Cancers prevention might also contribute to the longevity.
Aging may be a promoter in the development of aging-related diseases, whereas aging-related diseases could exacerbate the aging process. Increase in the incidence of aging related diseases may lead to the decrease in the quality of life (QOL) of individuals. Better health care can ensure a significant elevation of the QOL. In addition, aging molecular mechanisms including the control of autophagy at cellular, tissue, and/or body levels should be accurately characterized in order to achieve novel strategies for the longevity. Although it should be noted that some beneficial effects might also be related to the modification of the cellular antioxidant system, particular nutraceutical regimens within physiological doses may have potentials for the treatment of aging-related diseases. Some natural bioactive molecules could induce autophagy, which may also improve several aging-related diseases via the alteration of gut microbiota. Again, a longevity-promoting effect with appropriate autophagy could be provided, in which ncRNAs might play imperative roles. Nonetheless, the relationship among autophagy, ncRNAs and gut microbiota should intensely be investigated in the future studies.
6. Conclusions
The conclusion of this review is that supplementation of certain nutraceuticals and/or mild caloric restriction could modify the autophagy for the longevity. Several ncRNAs might also play imperative roles in the process of the autophagy regulation. Dieting may be associated with the alteration of gut microbiota, which could provide beneficial ncRNAs for the reduced risk of several aging-related diseases via the alteration of autophagy.
Author Contributions
Conceptualization, AF, MN, and SM; original draft preparation and editing, AF, and SM; visualization, AF and SM; supervision, SM. Each author (AF, MN, and SM) has participated sufficiently in this work of drafting the article and/or revising the article for the important rational content. Then, all authors gave final approval of the version to be submitted. Finally, all authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
A special thanks to all the members in Matsuda’s lab for sincerely assisting with the search of this meta-analysis and consultation in the drafting of this manuscript. The project was partially supported by Nara Women’s University of Japan.
Conflicts of Interest
The authors declare that they have no competing financial interests.
Abbreviations
| AMP | adenosine monophosphate |
| ATP | adenosine triphosphate |
| AMPK | adenosine monophosphate-activated protein kinase |
| C. elegans | Caenorhabditis elegans |
| FMT | fecal microbiota transplantation |
| miRNA | microRNA |
| mTOR | mechanistic/mammalian target of rapamycin |
| mTORC1 | mTOR complex 1 |
| NAFLD | nonalcoholic fatty liver disease |
| QOL | quality of life |
| ROS | reactive oxygen species |
| 3′UTR | three prime untranslated region |
| ULK1 | autophagy activating kinase 1 |
| UVA | ultraviolet ray A |
| UVB | ultraviolet ray B |
References
- Longo, V.D.; Antebi, A.; Bartke, A.; Barzilai, N.; Brown-Borg, H.M.; Caruso, C.; Curiel, T.J.; de Cabo, R.; Franceschi, C.; Gems, D.; et al. Interventions to Slow Aging in Humans: Are We Ready? Aging Cell 2015, 14, 497–510. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Jo, D.G.; Park, D.; Chung, H.Y.; Mattson, M.P. ; Jo, D.G.; Park, D.; Chung, H.Y.; Mattson, M.P. Adaptive cellular stress pathways as therapeutic targets of dietary phytochemicals: focus on the nervous system. Pharmacological Reviews 2014, 66, 815–868. [Google Scholar] [CrossRef]
- Mattison, J.A.; Colman, R.J.; Beasley, T.M.; Allison, D.B.; Kemnitz, J.W.; Roth, G.S.; Ingram, D.K.; Weindruch, R.; de Cabo, R.; Anderson, R.M. Caloric restriction improves health and survival of rhesus monkeys. Nature Communications 2017, 8, 14063. [Google Scholar] [CrossRef] [PubMed]
- Fontana, L.; Partridge, L.; Longo, V.D. Extending healthy life span--from yeast to humans. Science 2010, 328, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Gremeaux, V.; Gayda, M.; Lepers, R.; Sosner, P.; Juneau, M.; Nigam, A. Exercise and longevity. Maturitas 2012, 73, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Ekmekcioglu, C. Nutrition and longevity - From mechanisms to uncertainties. Critical Reviews in Food Science and Nutrition 2020, 60, 3063–3082. [Google Scholar] [CrossRef]
- Rubinsztein, D.C.; Mariño, G.; Kroemer, G. Autophagy and aging. Cell 2011, 146, 682–695. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, S.; Tasset, I.; Arias, E.; Pampliega, O.; Wong, E.; Martinez-Vicente, M.; Cuervo, A.M. Autophagy and the hallmarks of aging. Aging Research Reviews 2021, 72, 101468. [Google Scholar] [CrossRef]
- Zhang, H.; Puleston, D.J.; Simon, A.K. Autophagy and Immune Senescence. Trends in Molecular Medicine 2016, 22, 671–686. [Google Scholar] [CrossRef]
- Miceli, C.; Leri, M.; Stefani, M.; Bucciantini, M. Autophagy-related proteins: Potential diagnostic and prognostic biomarkers of aging-related diseases. Aging Research Reviews 2023, 89, 101967. [Google Scholar] [CrossRef] [PubMed]
- Thalyana, S.V.; Slack, F.J. MicroRNAs and their roles in aging. J Cell Sci 2012, 125, 7–17. [Google Scholar]
- Maciotta, S.; Meregalli, M.; Torrente, Y. The involvement of microRNAs in neurodegenerative diseases. Front Cell Neurosci 2013, 7, 265. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.; Noble, W.; Tartaglia, G.G.; Buckley, N.J. Neurodegeneration as an RNA disorder. Prog Neurobiol 2012, 99, 293–315. [Google Scholar] [CrossRef]
- Kim, J.; Inoue, K.; Ishii, J.; Vanti, W.B.; Voronov, S.V.; Murchison, E.; Hannon,G. ; Abeliovich, A. A MicroRNA Feedback Circuit in Midbrain Dopamine Neurons. Science 2007, 317, 1220–1224. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Beermann, J.; Piccoli, M.T.; Viereck, J.; Thum, T. Non-coding RNAs in development and disease: Background, mechanisms, and therapeutic approaches. Physiol Rev 2016, 96, 1297–1325. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; He, P.; Tian, Q.; Luo, Y.; He, Y.; Liu, C.; Gong, P.; Guo, Y.; Ye, Q.; Li, M. Genetic modification of miR-34a enhances efficacy of transplanted human dental pulp stem cells after ischemic stroke. Neural Regen. Res 2023, 18, 2029–2036. [Google Scholar]
- Guo, Y.; Li, P.; Gao, L.; Zhang, J.; Yang, Z.; Bledsoe, G.; Chang, E.; Chao, L.; Chao, J. Kallistatin reduces vascular senescence and aging by regulating microRNA-34a-SIRT1 pathway. Aging Cell 2017, 16, 837–846. [Google Scholar] [CrossRef]
- Yamakuchi, M.; Ferlito, M.; Lowenstein, C.J. miR-34a repression of SIRT1 regulates apoptosis. Proc Natl Acad Sci USA 2008, 105, 13421–13426. [Google Scholar] [CrossRef]
- Kinser, H.E.; Pincus, Z. MicroRNAs as modulators of longevity and the aging process. Hum Genet 2020, 139, 291. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Baehrecke, E.H.; Ballabio, A.; Boya, P.; Bravo-San Pedro, J.M.; Cecconi, F.; Choi, A.M.; Chu, C.T.; Codogno, P.; Colombo, M.I.; et al. Molecular definitions of autophagy and related processes. The EMBO Journal 2017, 36, 1811–1836. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, O.; Otsu, K. Role of autophagy in aging. Journal of Cardiovasc Pharmacol 2012, 60, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Han, Z.; Ding, L.; Wang, P.; He, X.; Lin, L. The molecular mechanism of aging and the role in neurodegenerative diseases. Heliyon 2024, 10, e24751. [Google Scholar] [CrossRef]
- He, L.Q.; Lu, J.H.; Yue, Z.Y. Autophagy in aging and aging-associated diseases. Acta Pharmacologica Sinica 2013, 34, 605–611. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, J.; Ouyang, C.; Meng, N. The association between ferroptosis and autophagy in cardiovascular diseases. Cell Biochemistry and Function 2024, 42, e3985. [Google Scholar] [CrossRef] [PubMed]
- Soda, K.; Kano, Y.; Chiba, F. Food polyamine and cardiovascular disease--an epidemiological study. Global Journal of Health Science 2012, 4, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, O. Autophagy in the Heart. Official Journal of the Japanese Circulation Society 2019, 83, 697–704. [Google Scholar] [CrossRef]
- Eisenberg, T.; Abdellatif, M.; Zimmermann, A.; Schroeder, S.; Pendl, T.; Harger, A.; Stekovic, S.; Schipke, J.; Magnes, C.; Schmidt, A.; et al. Dietary spermidine for lowering high blood pressure. Autophagy 2017, 13, 767–769. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, S.; Zhang, Y.; Lin, X.; Song, Y.; Xue, Z.; Qian, H.; Wang, S.; Wan, G.; Zheng, X.; et al. Induction of autophagy by spermidine is neuroprotective via inhibition of caspase 3-mediated Beclin 1 cleavage. Cell Death & Disease 2017, 8, e2738. [Google Scholar]
- Sigrist, S.J.; Carmona-Gutierrez, D.; Gupta, V.K.; Bhukel, A.; Mertel, S.; Eisenberg, T.; Madeo, F. Spermidine-triggered autophagy ameliorates memory during aging. Autophagy 2014, 10, 178–179. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.T.; Li, H.; Dai, Z.; Lau, G.K.; Li, B.Y.; Zhu, W.L.; Liu, X.Q.; Liu, H.F.; Cai, W.W.; Huang, S.Q.; et al. Spermidine and spermine delay brain aging by inducing autophagy in SAMP8 mice. Aging (Albany NY) 2020, 12, 6401–6414. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Molina, B.; Queipo-Ortuño, M.I.; Lambertos, A.; Tinahones, F.J.; Peñafiel, R. Dietary and Gut Microbiota Polyamines in Obesity- and Age-Related Diseases. Frontiers in Nutrition 2019, 6, 24. [Google Scholar] [CrossRef] [PubMed]
- Cervelli, M.; Leonetti, A.; Duranti, G.; Sabatini, S.; Ceci, R.; Mariottini, P. Skeletal Muscle Pathophysiology: The Emerging Role of Spermine Oxidase and Spermidine. Medical Sciences (Basel) 2018, 6, 14. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Alsaleh, G.; Feltham, J.; Sun, Y.; Napolitano, G.; Riffelmacher, T.; Charles, P.; Frau, L.; Hublitz, P.; Yu, Z.; et al. Polyamines Control eIF5A Hypusination, TFEB Translation, and Autophagy to Reverse B Cell Senescence. Molecular Cell 2019, 76, 110–125.e119. [Google Scholar] [CrossRef]
- Klionsky, D.J.; Petroni, G.; Amaravadi, R.K.; Baehrecke, E.H.; Ballabio, A.; Boya, P.; Bravo-San Pedro, J.M.; Cadwell, K.; Cecconi, F.; Choi, A.M.K.; et al. Autophagy in major human diseases. The EMBO Journal 2021, 40, e108863. [Google Scholar] [CrossRef] [PubMed]
- Cantó, C.; Gerhart-Hines, Z.; Feige, J.N.; Lagouge, M.; Noriega, L.; Milne, J.C.; Elliott, P.J.; Puigserver, P.; Auwerx, J. AMPK regulates energy expenditure by modulating NAD+ metabolism and SIRT1 activity. Nature 2009, 458, 1056–1060. [Google Scholar] [CrossRef]
- Hwang, J.S.; Lai, T.H.; Ahmed, M.; Pham, T.M.; Elashkar, O.; Bahar, E.; Kim, D.R. Regulation of TGF-beta1-Induced EMT by Autophagy-Dependent Energy Metabolism in Cancer Cells. Cancers (Basel) 2022, 14, 4845. [Google Scholar] [CrossRef]
- Li, S.X.; Li, C.; Pang, X.R.; Zhang, J.; Yu, G.C.; Yeo, A.J.; Lavin, M.F.; Shao, H. Jia, Q.; Peng, C. Metformin attenuates silica-induced pulmonary fibrosis by activating autophagy via the AMPK-mTOR signaling pathway. Frontiers in Pharmacology 2021, 12, 719589. [Google Scholar] [CrossRef]
- Trelford, C.B.; Di Guglielmo, G.M. Canonical and Non-canonical TGFbeta Signaling Activate Autophagy in an ULK1-Dependent Manner. Frontiers in Cell and Developmental Biology 2021, 9, 712124. [Google Scholar] [CrossRef] [PubMed]
- Zachari, M.; Ganley, I.G. The mammalian ULK1 complex and autophagy initiation. Essays in Biochemistry 2017, 61, 585–596. [Google Scholar]
- Egan, D.; Kim, J.; Shaw, R.J.; Guan, K.L. The autophagy initiating kinase ULK1 is regulated via opposing phosphorylation by AMPK and mTOR. Autophagy 2011, 7, 643–644. [Google Scholar] [CrossRef]
- Chan, EY. mTORC1 Phosphorylates the ULK1-mAtg13-FIP200 Autophagy Regulatory Complex. Science Signaling 2009, 2, pe51. [Google Scholar] [CrossRef]
- González, A.; Hall, M.N.; Lin, S.C.; Hardie, D.G. AMPK and TOR: the Yin and Yang of cellular nutrient sensing and growth control. Cell Metabolism 2020, 31, 472–492. [Google Scholar] [CrossRef]
- Steinberg, G.R.; Hardie, D.G. New insights into activation and function of the AMPK. Nature Reviews Molecular Cell Biology 2023, 24, 255–272. [Google Scholar] [CrossRef] [PubMed]
- Maharajan, N.; Ganesan, C.D.; Moon, C.; Jang, C.H.; Oh, W.K.; Cho, G.W. Licochalcone D ameliorates oxidative stress-induced senescence via AMPK activation. International Journal of Molecular Sciences 2021, 22, 7324. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Anand, S.K.; Singh, N.; Dwivedi, U.N.; Kakkar, P. AMP-activated protein kinase: an energy sensor and survival mechanism in the reinstatement of metabolic homeostasis. Experimental Cell Research 2023, 428, 113614. [Google Scholar] [CrossRef]
- Hu, Y.; Dong, Y.D.; Wu, Y.C.; Wang, Q.X.; Nan, X.; Wang, D.L. AMPK inhibitor BML-275 induces neuroprotection through decreasing cyt c and AIF expression after transient brain ischemia. Bioorganic & Medicinal Chemistry 2021, 52, 116522. [Google Scholar]
- Spaulding, H.R.; Yan, Z. AMPK and the adaptation to exercise. Annual Review Physiology 2022, 84, 209–227. [Google Scholar] [CrossRef]
- Qiu, Z.; Li, Y.; Fu, Y.; Yang, Y. Research progress of AMP-activated protein kinase and cardiac aging. Open Life Sciences 2023, 18, 20220710. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yang, Z.; Bai, J.; Wang, X.; Yuan, Q.; Mi, Y.; Zhang, C. Bioactive Lignan Honokiol Alleviates Ovarian Oxidative Stress in Aging Laying Chickens by Regulating SIRT3/AMPK Pathway. Antioxidants (Basel) 2024, 13, 377. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of microRNA biogenesis, mechanisms of actions, and circulation. Front. Endocrinol 2018, 9, 402. [Google Scholar] [CrossRef]
- Rodriguez, A.; Griffiths-Jones, S.; Ashurst, J.L.; Bradley, A. Identification of mammalian microRNA host genes and transcription units. Genome Res 2004, 14, 1902–1910. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Lee, Y.; Yeom, K.H.; Kim, Y.K.; Jin, H.; Kim, V.N. The Drosha-DGCR8 complex in primary microRNA processing. Genes Dev 2004, 18, 3016–3027. [Google Scholar] [CrossRef] [PubMed]
- Cullen, B.R. Transcription and processing of human microRNA precursors. Mol Cell 2004, 16, 861–865. [Google Scholar] [CrossRef]
- van den Berg, A.; Mols, J.; Han, J. RISC-target interaction: Cleavage and translational suppression. Biochim Biophys Acta 2008, 1779, 668–677. [Google Scholar] [CrossRef] [PubMed]
- Majeski, A.E.; Fred Dice, J. Mechanisms of chaperone-mediated autophagy. Int J Biochem Cell Biol 2004, 36, 2435–2444. [Google Scholar] [CrossRef]
- Alvarez-Erviti, L.; Seow, Y.; Schapira, A.H.V.; Rodriguez-Oroz, M.C.; Obeso, J.A.; Cooper, J.M. Influence of microRNA deregulation on chaperone-mediated autophagy and α-synuclein pathology in Parkinson’s disease. Cell Death Dis 2013, 4, e545. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wang, X.; Song, M.; He, Z.; Cui, G.; Peng, G.; Dieterich, C.; Antebi, A.; Jing, N.; Shen, Y. A secreted microRNA disrupts autophagy in distinct tissues of Caenorhabditis elegans upon ageing. Nat Commun 2019, 10, 4827. [Google Scholar] [CrossRef]
- Yang, J.; Chen, D.; He, Y.; Meléndez, A.; Feng, Z.; Hong, Q.; Bai, X.; Li, Q.; Cai, G.; Wang, J.; et al. MiR-34 modulates Caenorhabditis elegans lifespan via repressing the autophagy gene atg9. Age 2013, 35, 11. [Google Scholar] [CrossRef]
- Zhang, J.; Zhou, B.; Xu, Y.; Chen, X.; Liu, J.; Gozali, M.; Wu, D.; Yin, Z.; Luo, D. MiR-23a-depressed autophagy is a participant in PUVA- and UVB-induced premature senescence. Oncotarget 2016, 7, 37420–37435. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Xu, J.; Wang, C.; Shi, D.; Yan, Q. miR-23b-3p regulates apoptosis and autophagy via suppressing SIRT1 in lens epithelial cells. J Cell Biochem 2019, 120, 19635–19646. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.L.; Hong, C.G.; Yue, T.; Li, H.M.; Duan, R.; Hu, W.B.; Cao, J.; Wang, Z.X.; Chen, C.Y.; Hu, X.K.; et al. Inhibition of miR-331-3p and miR-9-5p ameliorates Alzheimer’s disease by enhancing autophagy. Theranostics 2021, 11, 2395–2409. [Google Scholar] [CrossRef] [PubMed]
- Barbato, C.; Giacovazzo, G.; Albiero, F.; Scardigli, R.; Scopa, C.; Ciotti, M.T.; Strimpakos, G.; Coccurello, R.; Ruberti, F. Cognitive Decline and Modulation of Alzheimer's Disease-Related Genes After Inhibition of MicroRNA-101 in Mouse Hippocampal Neurons. Mol Neurobiol 2020, 57, 3183–3194. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.Z.A.; Zhao, D.; Hussain, T.; Sabir, N.; Yang, L. Regulation of MicroRNAs-Mediated Autophagic Flux: A New Regulatory Avenue for Neurodegenerative Diseases with Focus on Prion Diseases. Front Aging Neurosci 2018, 10, 139. [Google Scholar] [CrossRef]
- Hyttinen, J.M.T.; Blasiak, J.; Felszeghy, S.; Kaarniranta, K. MicroRNAs in the regulation of autophagy and their possible use in age-related macular degeneration therapy. Ageing Res Rev 2021, 67, 101260. [Google Scholar] [CrossRef]
- Pulakat, L.; Chen, H.H. Pro-Senescence and Anti-Senescence Mechanisms of Cardiovascular Aging: Cardiac MicroRNA Regulation of Longevity Drug-Induced Autophagy. Front Pharm 2020, 11, 774. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, Q.; Zhang, H.; He, Z.; Song, Y.; Chen, Y.; Tang, N.; Zhou, Y.; Li, Y.; Antebi, A.; et al. Tissue-specific profiling of age-dependent miRNAomic changes in Caenorhabditis elegans. Nat Commun. 2024, 15, 955. [Google Scholar] [CrossRef]
- Song, Z.; Wei, D.; Chen, Y.; Chen, L.; Bian, Y.; Shen, Y.; Chen, J.; Pan, Y. Association of astragaloside IV-inhibited autophagy and mineralization in vascular smooth muscle cells with lncRNA H19 and DUSP5-mediated ERK signaling. Toxicology and Applied Pharmacology 2019, 364, 45–54. [Google Scholar] [CrossRef]
- Chen, Y.; Zeng, A.; He, S.; He, S.; Li, C.; Mei, W.; Lu, Q. Autophagy-Related Genes in Atherosclerosis. J Healthc Eng 2021, 2021, 6402206. [Google Scholar] [CrossRef]
- Mancini, A.; Vitucci, D.; Orlandella, FM.; Terracciano, A.; Mariniello, RM.; Imperlini, E.; Grazioli, E.; Orrù, S.; Krustrup, P.; Salvatore, G.; et al. Regular football training down-regulates miR-1303 muscle expression in veterans. Eur J Appl Physiol 2021, 121, 2903–2912. [Google Scholar] [CrossRef]
- Burke, S.L.; Hammell, M.; Ambros, V. Robust Distal Tip Cell Pathfinding in the Face of Temperature Stress Is Ensured by Two Conserved microRNAS in Caenorhabditis elegans. Genetics. 2015, 200, 1201–1218. [Google Scholar] [CrossRef] [PubMed]
- Kinser, H.E.; Pincus, Z. MicroRNAs as modulators of longevity and the aging process. Hum Genet 2020, 139, 291. [Google Scholar] [CrossRef]
- Parvaresh, H.; Paczek, K.; Al-Bari, MAA. ; Eid, N. Mechanistic insights into fasting-induced autophagy in the aging heart. World Journal of Cardiology 2024, 16, 109–117. [Google Scholar] [CrossRef]
- Saini, S.K.; Singh, A.; Saini, M.; Gonzalez-Freire, M.; Leeuwenburgh, C.; Anton, SD. Time-Restricted Eating Regimen Differentially Affects Circulatory miRNA Expression in Older Overweight Adults. Nutrients. 2022, 14, 1843. [Google Scholar] [CrossRef]
- Buono, R.; Tucci, J.; Cutri, R.; Guidi, N.; Mangul, S.; Raucci, F.; Pellegrini, M.; Mittelman, S.D.; Longo, V.D. Fasting-Mimicking Diet Inhibits Autophagy and Synergizes with Chemotherapy to Promote T-Cell-Dependent Leukemia-Free Survival. Cancers (Basel) 2023, 15, 5870. [Google Scholar] [CrossRef]
- Ulas, M.; Akbas, E.; Akbas, S.; Aktemur, G.; Durcanoglu, N.; Aksak, K.; Atalar, A.A.; Yıldırım, S.; Akalin, I. Physiological aspect of apoptosis-regulating microRNAs expressions during fasting. Eur Rev Med Pharmacol Sci. 2023, 27, 2210–2215. [Google Scholar]
- Mao, Y.; Cheng, J.; Yu, F.; Li, H.; Guo, C.; Fan, X. Ghrelin attenuated lipotoxicity via autophagy induction and nuclear factor-κB inhibition. Cellular Physiology and Biochemistry 2015, 37, 563–576. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Moon, N.R.; Kim, D.S.; Kim, S.H.; Park, S. Central acylated ghrelin improves memory function and hippocampal AMPK activation and partly reverses the impairment of energy and glucose metabolism in rats infused with β-amyloid. Peptides 2015, 71, 84–93. [Google Scholar] [CrossRef]
- Qiu, J.; Ma, C.; Dai, W.; Fang, E.; Li, W.; Yang, F. Ghrelin attenuates transforming growth factor-beta1-induced pulmonary fibrosis via the miR-125a-5p/Kruppel-like factor 13 axis. Arch Biochem Biophys 2022, 715, 109082. [Google Scholar] [CrossRef]
- Liang, Y.; Xu, X.; Yin, M.; Zhang, Y.; Huang, L.; Chen, R.; Ni, J. Effects of berberine on blood glucose in patients with type 2 diabetes mellitus: A systematic literature review and a meta-analysis. Endocrine Journal 2019, 66, 51–63. [Google Scholar] [CrossRef]
- Hawley, S.A.; Ross, F.A.; Chevtzoff, C.; Green, K.A.; Evans, A.; Fogarty, S.; Towler, M.C.; Brown, L.J.; Ogunbayo, O.A.; Evans, A.M.; et al. Use of cells expressing gamma subunit variants to identify diverse mechanisms of AMPK activation. Cell Metabolism 2010, 11, 554–565. [Google Scholar] [CrossRef] [PubMed]
- Mohammadinejad, R.; Ahmadi, Z.; Tavakol, S.; Ashrafizadeh, M. Berberine as a potential autophagy modulator. Journal of Cellular Physiology 2019, 234, 14914–14926. [Google Scholar] [CrossRef]
- Yang, M.; Yang, T.; Mei, L.; Zhang, Y.; Liang, C.; Bai, X.; Zhang, Z.; Shi, Y.; Chen, Q. The Potential of Berberine to Target Telocytes in Rabbit Heart. Planta Medica 2014, 90, 84–95. [Google Scholar]
- Zhu, N.; Li, J.; Li, Y.; Zhang, Y.; Du, Q.; Hao, P.; Li, J.; Cao, X.; Li, L. Berberine Protects Against Simulated Ischemia/Reperfusion Injury-Induced H9C2 Cardiomyocytes Apoptosis In Vitro and Myocardial Ischemia/Reperfusion-Induced Apoptosis In Vivo by Regulating the Mitophagy-Mediated HIF-1alpha/BNIP3 Pathway. Frontiers in Pharmacology 2020, 11, 367. [Google Scholar] [CrossRef]
- Zhu, N.; Cao, X.; Hao, P.; Zhang, Y.; Chen, Y.; Zhang, J.; Li, J.; Gao, C.; Li, L. Berberine attenuates mitochondrial dysfunction by inducing autophagic flux in myocardial hypoxia/reoxygenation injury. Cell Stress and Chaperones 2020, 25, 417–426. [Google Scholar] [CrossRef]
- Zhu, M.L.; Yin, Y.L.; Ping, S.; Yu, H.Y.; Wan, G.R.; Jian, X. Li, P. Berberine promotes ischemia-induced angiogenesis in mice heart via upregulation of microRNA-29b. Clin Exp Hypertens 2017, 39, 672–679. [Google Scholar] [CrossRef]
- Xu, H.; Yu, S.; Lin, C.; Dong, D.; Xiao, J.; Ye, Y.; Wang, M. Roles of flavonoids in ischemic heart disease: Cardioprotective effects and mechanisms against myocardial ischemia and reperfusion injury. Phytomedicine 2024, 126, 155409. [Google Scholar] [CrossRef] [PubMed]
- Grewal, A.K.; Singh, T.G.; Sharma, D.; Sharma, V.; Singh, M.; Rahman, M.H.; Najda, A.; Walasek-Janusz, M.; Kamel, M.; Albadrani, G.M.; et al. Mechanistic insights and perspectives involved in neuroprotective action of quercetin. Biomedicine & Pharmacotherapy 2021, 140, 111729. [Google Scholar]
- Sung, B.; Chung, J.W.; Bae, H.R.; Choi, J.S.; Kim, C.M.; Kim, N.D. Humulus japonicus extract exhibits antioxidative and anti-aging effects via modulation of the AMPK-SIRT1 pathway. Experimental and Therapeutic Medicine 2015, 9, 1819–1826. [Google Scholar] [CrossRef] [PubMed]
- Benameur, T.; Soleti, R.; Porro, C. The Potential Neuroprotective Role of Free and Encapsulated Quercetin Mediated by miRNA against Neurological Diseases. Nutrients 2021, 13, 1318. [Google Scholar] [CrossRef] [PubMed]
- Khater, S.I.; El-Emam, M.M.A.; Abdellatif, H.; Mostafa, M.; Khamis, T.; Soliman, R.H.M.; Ahmed, H.S.; Ali, S.K.; Selim, H.M.R.M.; Alqahtani, L.S.; et al. Lipid nanoparticles of quercetin (QU-Lip) alleviated pancreatic microenvironment in diabetic male rats: The interplay between oxidative stress - unfolded protein response (UPR) - autophagy, and their regulatory miRNA. Life Sci 2024, 344, 122546. [Google Scholar] [CrossRef]
- Santa-Maria, I.; Alaniz, M.E.; Renwick, N.; Cela, C.; Fulga, T.A.; Van Vactor, D.; Tuschl, T.; Clark, L.N; Shelanski, M.L.; McCabe, B.D.; et al. Dysregulation of microRNA-219 promotes neurodegeneration through post-transcriptional regulation of tau. J Clin Investig 2015, 125, 681–686. [Google Scholar] [CrossRef] [PubMed]
- Konovalova J, Gerasymchuk D, Parkkinen I, Chmielarz P, Domanskyi A. Interplay between MicroRNAs and Oxidative Stress in Neurodegenerative Diseases. Int J Mol Sci 2019, 20, 6055. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yi, P.; Yi, M.; Tong, X.; Cheng, X.; Yang, J.; Hu, Y.; Peng, W. Protective Effect of Quercetin against H2O2-Induced Oxidative Damage in PC-12 Cells: Comprehensive Analysis of a lncRNA-Associated ceRNA Network. Oxid Med Cell Longev 2020, 2020, 6038919. [Google Scholar] [CrossRef]
- Wang, T.; Xu, Q.; Cao, Y.; Zhang, C.; Chen, S.; Zhang, Y.; Liang, T. Luteolin Ameliorates Hepatic Steatosis and Enhances Mitochondrial Biogenesis via AMPK/PGC-1alpha Pathway in Western Diet-Fed Mice. Journal of Nutritional Science and Vitaminology 2023, 69, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Kang, A.W.; Sun, C.; Li, H.T.; Zhong, K.; Zeng, X.H.; Gu, Z.F.; Li, B.Q.; Zhang, X.N.; Gao, J.L.; Chen, T.X. Puerarin is a promising compound for improving the longevity of Drosophila melanogaster by activating autophagy. Food & Function journal 2023, 14, 2149–2161. [Google Scholar]
- Li, C.H.; Gong, D.; Chen, L.Y.; Zhang, M.; Xia, X.D.; Cheng, H.P.; Huang, C.; Zhao, Z.W.; Zheng, X.L.; Tang, X.E.; et al. Puerarin promotes ABCA1-mediated cholesterol efflux and decreases cellular lipid accumulation in THP-1 macrophages. Eur J Pharmacol 2017, 811, 74–86. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.H.; Shen, Z.Q.; Wang, T.W.; Kao, C.H.; Teng, Y.C.; Yeh, T.K.; Lu, C.T.; Yeh, T.K.; Lu, C.K.; Tsai, T.F. Hesperetin promotes longevity and delays aging via activation of Cisd2 in naturally aged mice. Journal of Biomedical Science 2022, 9, 53. [Google Scholar] [CrossRef]
- Park, H.Y.; Yu, J.H. Hesperidin enhances intestinal barrier function in Caco-2 cell monolayers via AMPK-mediated tight junction-related proteins. FEBS Open Bio 2013, 13, 532–544. [Google Scholar] [CrossRef]
- Dong, J.; Zhou, H.; Zhao, H.; Zhao, Y.; Chang, C. Hesperetin ameliorates lipopolysaccharide-induced acute lung injury via the miR-410/SOX18 axis. J Biochem. Mol. Toxicol. 2020, 34, e22588. [Google Scholar] [CrossRef]
- Bal, N.B.; Bostanci, A.; Sadi, G.; Dönmez, M.O.; Uludag, M.O.; Demirel-Yilmaz, E. Resveratrol and regular exercise may attenuate hypertension-induced cardiac dysfunction through modulation of cellular stress responses. Life Sciences 2022, 96, 120424. [Google Scholar] [CrossRef]
- Wang, P.; Yang, Y.; Guo, J.; Ma, T.; Hu, Y.; Huang, L.; He, Y.; Xi, J. Resveratrol Inhibits Zinc Deficiency-Induced Mitophagy and Exerts Cardiac Cytoprotective Effects. Biological Trace Element Research 2024, 202, 1669–1682. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Zhang, Y.; Yun, Q.; Du, R.; Li, L.; Li, Y.; Gao, Q. Resveratrol alleviates hyperglycemia-induced cardiomyocyte hypertrophy by maintaining mitochondrial homeostasis via enhancing SIRT1 expression. Nan Fang Yi Ke Da Xue Xue Bao 2024, 44, 45–51. [Google Scholar]
- Mori, A.; Ezawa, Y.; Asano, D.; Kanamori, T.; Morita, A.; Kashihara, T. Sakamoto K.; Nakahara, T. Resveratrol dilates arterioles and protects against N-methyl-d-aspartic acid-induced excitotoxicity in the rat retina. Neuroscience Letters 2023, 793, 136999. [Google Scholar] [CrossRef]
- Fardi, F.; Khasraghi, L.B.; Shahbakhti, N.; Salami Naseriyan, A.; Najafi, S.; Sanaaee, S.; Alipourfard, I.; Zamany, M.; Karamipour, S.; Jahani, M.; et al. An interplay between non-coding RNAs and gut microbiota in human health. Diabetes Res. Clin. Pract. 2023, 201, 110739. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, M.; Chen, J.; Li, Y. ; Kuang, Z.; Dende, C.; Raj, P.; Quinn, G.; Hu, Z.; Srinivasan, T.; et al. The gut microbiota reprograms intestinal lipid metabolism through long noncoding RNA Snhg9. Science. 2023, 381, 851–857. [Google Scholar] [CrossRef]
- Ma, H.; Liu, Y.; Yan, R.; Wang, R.; Zhang, P.; Bai, Z.; Liu, Y.; Ren, Y.; Li, Y.; Jiang, X.; et al. Butyrate suppresses atherosclerotic inflammation by regulating macrophages and polarization via GPR43/HDAC-miRNAs axis in ApoE−/− mice. PLoS ONE. 2023, 18, e0282685. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, S.T.; Abdullah, S.R.; Hussen, BM.; Younis, YM.; Rasul, MF.; Taheri, M. Role of circular RNAs and gut microbiome in gastrointestinal cancers and therapeutic targets. Noncoding RNA Res. 2023, 9, 236–252. [Google Scholar] [CrossRef]
- Tanigawa, K.; Misono, S.; Mizuno, K.; Asai, S.; Suetsugu, T.; Uchida, A.; Kawano, M.; Inoue, H.; Seki, N. MicroRNA signature of small-cell lung cancer after treatment failure: Impact on oncogenic targets by miR-30a-3p control. Mol Oncol 2023, 17, 328–343. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Jing, W.; Jin, S.; Wang, B. Circ_0088194 regulates proliferation, migration, apoptosis, and inflammation by miR-30a-3p/ADAM10 axis in rheumatoid arthritis fibroblastic synovial cells. Inflammation 2023, 46, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Shang, L.; Chen, S.; Du, F.; Li, S.; Zhao, L.; Wang, X. Nutrient starvation elicits an acute autophagic response mediated by Ulk1 dephosphorylation and its subsequent dissociation from AMPK. Proc Natl Acad Sci USA 2011, 108, 4788–4793. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, M.A.; Biggers, C.D.; Li, P.A. Rapamycin treatment increases hippocampal cell viability in an mTOR-independent manner during exposure to hypoxia mimetic, cobalt chloride. BMC Neurosci. 2018, 19, 82. [Google Scholar] [CrossRef] [PubMed]
- Hannan, F.M.; Elajnaf, T.; Vandenberg, L.N.; Kennedy, S.H.; Thakker, R.V. Hormonal regulation of mammary gland development and lactation. Nat Rev Endocrinol 2023, 19, 46–61. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, Q. miR-34a and endothelial biology. Life Sci. 2023, 330, 121976. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.E.; Fu, T.; Seok, S.; Kim, D.H.; Yu, E.; Lee, K.W.; Kang, Y.; Li, X.; Kemper, B.; Kemper, J.K. Elevated microRNA-34a in obesity reduces NAD+ levels and SIRT1 activity by directly targeting NAMPT. Aging Cell 2013, 12, 1062–1072. [Google Scholar] [CrossRef]
- Li, J.; Wang, K.; Chen, X.; Meng, H.; Song, M.; Wang, Y.; et al. Transcriptional activation of microRNA-34a by NF-kappa B in human esophageal cancer cells. BMC Mol Biol. 2012, 13, 4. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Yin, B.; Guo, S.; Umar, T.; Liu, J.; Wu, Z.; Zhou, Q.; Zahoor, A.; Deng, G. Enhanced Expression of miR-34a Enhances Escherichia coli Lipopolysaccharide-Mediated Endometritis by Targeting LGR4 to Activate the NF-kappaB Pathway. Oxid Med Cell Longev 2021, 2021, 1744754. [Google Scholar] [CrossRef]
- Li, S.; Wei, X.; He, J.; Cao, Q.; Du, D.; Zhan, X.; Zeng. Y.; Yuan, S.; Sun, L. The comprehensive landscape of miR-34a in cancer research. Cancer Metastasis Rev 2021, 40, 925–948. [Google Scholar] [CrossRef]
Figure 1.
Schematic image of the autophagy involved in both physical aging and the aging-related diseases including neurodegenerative disease, diabetes, hypertension, cardio vascular disease, muscular atrophy, osteoporosis, and cancers. Excess reactive oxygen species (ROS) production may be involved in the development of autophagy. The autophagy could also be affected from various ncRNAs including miR-34a,miR-301b, miR-26b, miR-106a, miR-29, miR-83, miR-23, miR-331, miR-9, and miR-101, etc. Note that some of important factors including inflammation, and/or redox imbalance triggering several aging-related diseases have been omitted for clarity.
Figure 1.
Schematic image of the autophagy involved in both physical aging and the aging-related diseases including neurodegenerative disease, diabetes, hypertension, cardio vascular disease, muscular atrophy, osteoporosis, and cancers. Excess reactive oxygen species (ROS) production may be involved in the development of autophagy. The autophagy could also be affected from various ncRNAs including miR-34a,miR-301b, miR-26b, miR-106a, miR-29, miR-83, miR-23, miR-331, miR-9, and miR-101, etc. Note that some of important factors including inflammation, and/or redox imbalance triggering several aging-related diseases have been omitted for clarity.
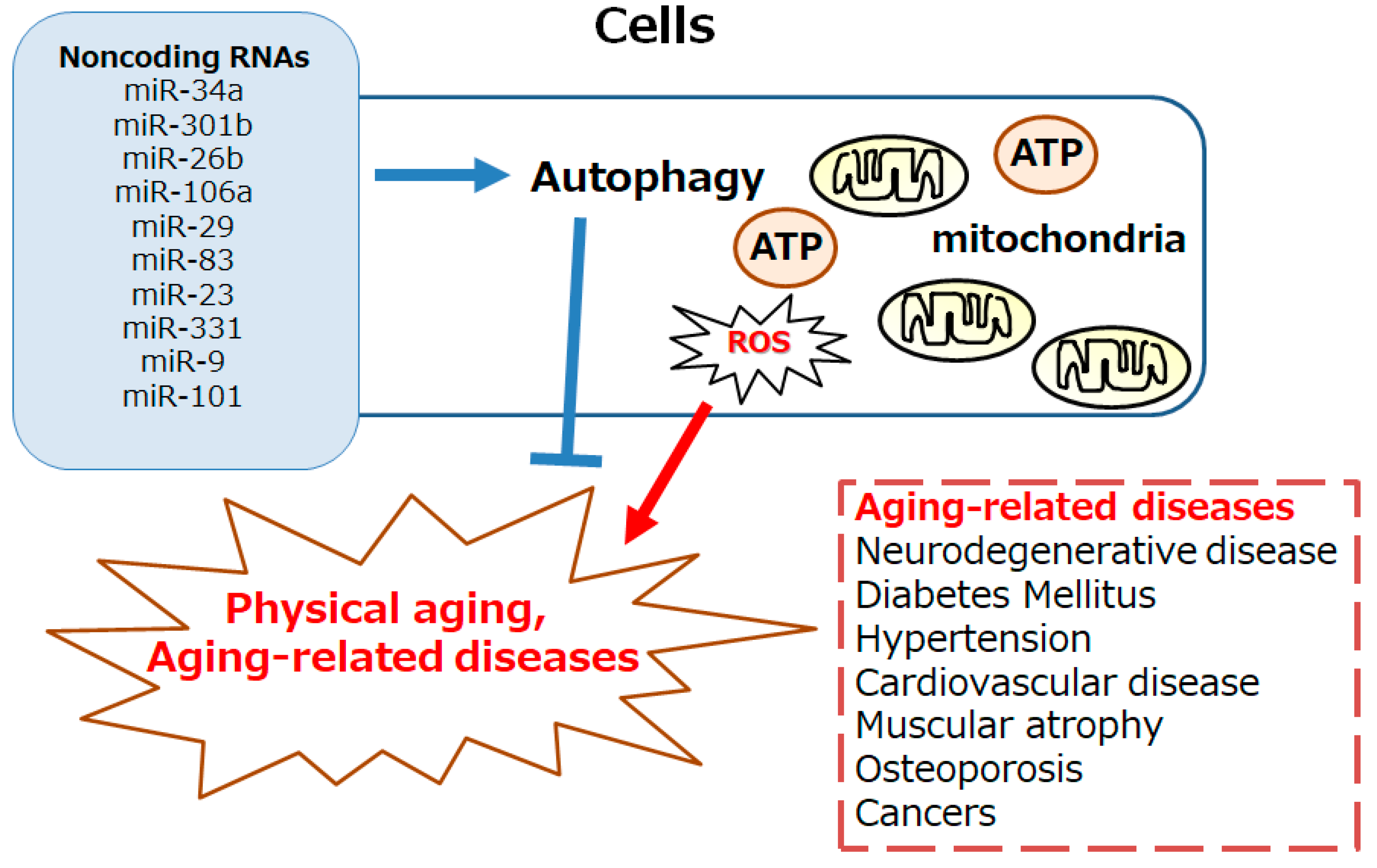
Figure 2.
An overview of the intracellular signaling pathway relevant to the autophagy. With several key molecules such as AMPK, PI3K, AKT, mTOR, and ULK1, the autophagy signaling might be involved in the regulation of physical aging and/or aging-related diseases. Inflammation, starvation, fasting, and/or hypoxia are known to initiate the autophagy signaling pathway. Arrowhead indicates stimulation whereas hammerhead shows inhibition. Note that several signaling pathways such as cytokine-induction and/or inflammatory responses have been omitted for clarity. Abbreviation: mTOR, mammalian/mechanistic target of rapamycin; PI3K, phosphoinositide-3 kinase; AKT, protein kinase B; ROS, reactive oxygen species; AMPK, adenosine monophosphate-activated protein kinase; ULK1, autophagy activating kinase 1; mTORC1, mTOR complex 1.
Figure 2.
An overview of the intracellular signaling pathway relevant to the autophagy. With several key molecules such as AMPK, PI3K, AKT, mTOR, and ULK1, the autophagy signaling might be involved in the regulation of physical aging and/or aging-related diseases. Inflammation, starvation, fasting, and/or hypoxia are known to initiate the autophagy signaling pathway. Arrowhead indicates stimulation whereas hammerhead shows inhibition. Note that several signaling pathways such as cytokine-induction and/or inflammatory responses have been omitted for clarity. Abbreviation: mTOR, mammalian/mechanistic target of rapamycin; PI3K, phosphoinositide-3 kinase; AKT, protein kinase B; ROS, reactive oxygen species; AMPK, adenosine monophosphate-activated protein kinase; ULK1, autophagy activating kinase 1; mTORC1, mTOR complex 1.
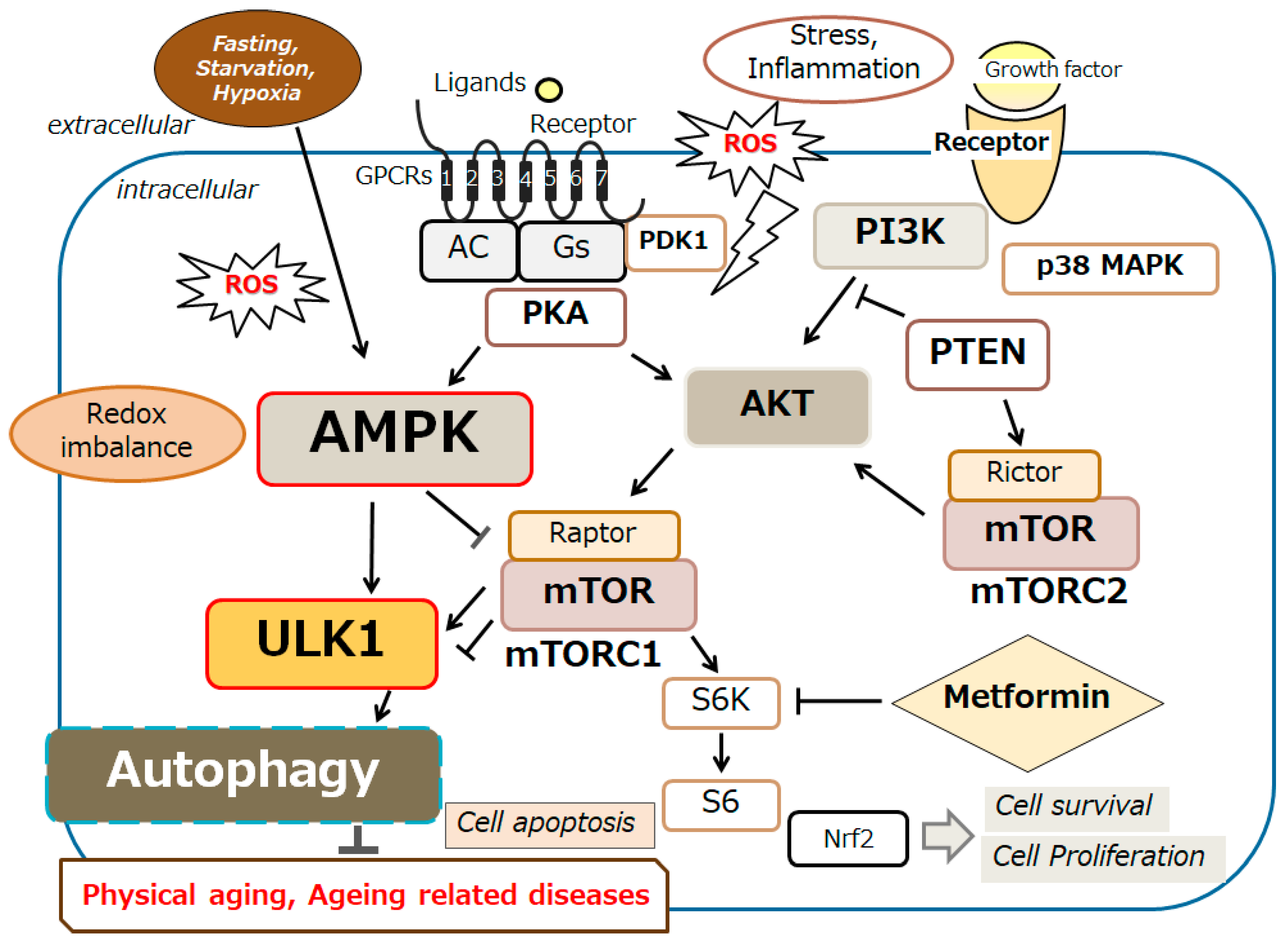
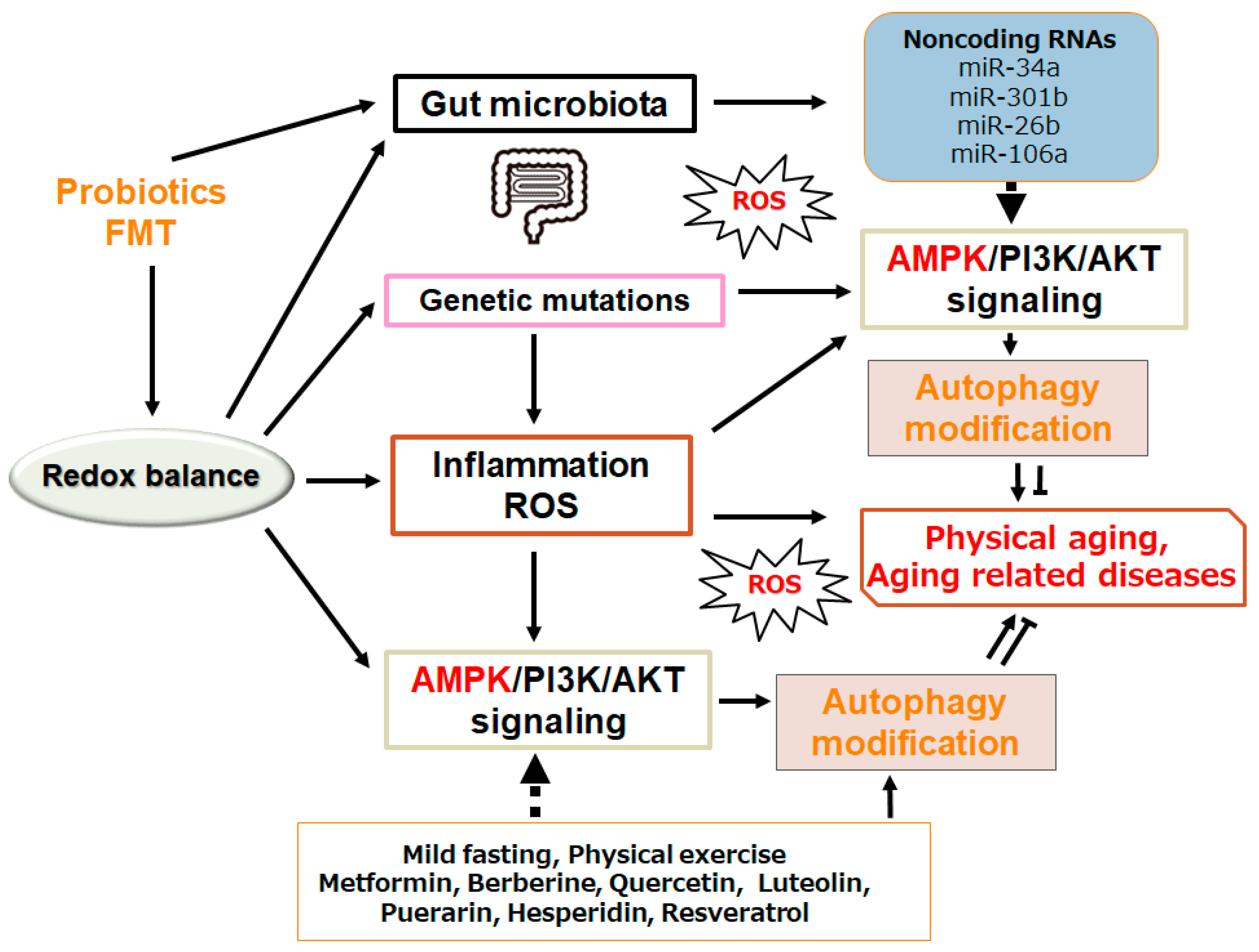
Figure 3.
Schematic demonstration of the potential inhibitory tactics against the development of physical aging and/or aging-related diseases. Example implements including metformin treatment and several nutraceuticals dieting as well as mild fasting/physical exercise known to act on the autophagy modification are also shown. Note that some of significant activities such as inflammatory responses and/or reactive oxygen species (ROS) production from inflammatory conditions have been misplaced for clarity. Some probiotics and/or fecal microbiota transplantation (FMT) could support the positive modification of gut microbiota for the improved autophagy via the alteration of noncoding RNAs production, which might also be advantageous for the treatment of physical aging and/or several aging-related diseases.
Figure 3.
Schematic demonstration of the potential inhibitory tactics against the development of physical aging and/or aging-related diseases. Example implements including metformin treatment and several nutraceuticals dieting as well as mild fasting/physical exercise known to act on the autophagy modification are also shown. Note that some of significant activities such as inflammatory responses and/or reactive oxygen species (ROS) production from inflammatory conditions have been misplaced for clarity. Some probiotics and/or fecal microbiota transplantation (FMT) could support the positive modification of gut microbiota for the improved autophagy via the alteration of noncoding RNAs production, which might also be advantageous for the treatment of physical aging and/or several aging-related diseases.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



