
Submitted:
10 December 2024
Posted:
11 December 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Despite the widespread use of Covid-19 vaccines, there is still a global need to find effective therapeutics to deal with the variants of SARS-CoV-2. Moslae Herba (MH) is a herbal medicine crediting with antiviral effect. The study aims to investigate the antiviral effects and the underlying mechanism of aqueous extract from MH (AEMH) for treating SARS-CoV-2. The in vitro anti-SARS-CoV-2 activity of AEMH was evaluated using cell viability and viral load. Component analysis was performed by HPLC-ESI-Q-TOF/MS. The connection between Covid-19 and AEMH was constructed by integrating network pharmacology and transcriptome profile to seek the core targets. The components with antiviral activities were analyze by molecular docking and in vitro pharmacological verification. AEMH exerted anti-SARS-CoV-2 effects through inhibiting viral replication and reducing cell death caused by infection (IC50 is 170 μg/mL for omicron strain). A total of 26 components were identified from AEMH. Through matching 119 intersection targets of ‘disease and drug’ with 1082 differentially expressed genes Covid-19 patients, nine genes were screened. Of the nine, PNP and TPI1 genes were identified as core targets as AEMH treatment significantly regulated the mRNA expression level of the two genes on infected cells. Three components, caffeic acid, luteolin, and rosmarinic acid displayed antiviral activities in verification. Molecular docking also demonstrated they could form stable bonds with the core targets. This study explored the antiviral activity and possible mechanism of AEMH for treating SARS-CoV-2, which could provide basic data and reference for clinical application of MH.
Keywords:
SARS-CoV-2
; Moslae Herba
; HPLC-ESI-Q-TOF/MS
; network pharmacology
; molecular docking
1. Introduction
The coronavirus disease 2019 (Covid-19) pandemic, caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), resulted in devastation on public health and global economy. According to World Health Organization, as of November 17, 2024, in total of 776,897,200 people were confirmed to infected with SARS-CoV-2 worldwide, and above seven million patients died [1]. The swift development and application of vaccines made a great contribution in controlling the spread of Covid-19. However, along with the evolution of SARS-CoV-2, many variants appeared [2]. The variants may escape the humoral immune response and weaken the effectiveness of current vaccines, thereby resulting in new transmission. In terms of treatment, a variety of approaches, including repurposing of previously adopted drugs, development of novel virus-targeting antivirals, as well as search for the immune-modulators and drug-capable host targets, have been used in the research of Covid-19 drugs [2,3]. Of which, Paxlovid, developed on the basis of SARS-CoV-2 Mpro inhibitor nirmatrelvir (PF-07321332), had been approved and widely used for the treatment of Covid-19 [4]. Similar to the vaccines, the antiviral drugs are also facing great challenges in dealing with new variants of SARS-CoV-2, especially the resistance mutants. For instance, most omicron variants were found to produce immune escape to therapeutic antibodies approved previously [5,6]. Hence, the constant search for efficient and safe compounds for treatment of SARS-CoV-2 is still essential.
Chinese herbal medicine (CHM) is a unique resource in medical system of China, and it has been used for prevention and treatment of infectious diseases for thousands of years. After long-time practice, CHM was proved to be efficient and safe for treatment of viral infection, such as influenza [7,8]. In terms of Covid-19, many researches provided evidence that CHM can not only help recovery of mild and ordinary cases, but also be an adjuvant treatment for severe cases [9,10,11]. Integration of CHM with standard care or western medicine can significantly reduce the mortality, improve the clinical symptoms and treatment outcomes of the patients [9,10]. As it is a whole of multi-components, CHM exerted anti-SARS-CoV-2 effects through multiple mechanisms, including down-regulation of the expression of angiotensin converting enzyme 2 (ACE2), blocking of bond between ACE2 and the viral spike protein, and regulation of host immune function as well [12,13,14]. Hence, CHM, with the advantages of low toxicity and diverse activities, is an important source which could provide many potential compounds for further control of SARS-CoV-2 variants.
Moslae Herba (MH) is the aboveground dry part of the Mosla chinensis Maxim (M. chinensis Maxim) or M. chinensis.cv. ‘Jiangxiangru’, a tomentose and aromatic plant belonging to Lamiaceae family, is a traditional medicinal and edible herb mainly produced in southern China. Chinese pharmacopoeia records that the herb has the effects of sweating and relieving surface, as well as removing dampness for regulating stomach. Clinically, it is widely used for treating summer-dampness cold, aversion to cold with fever, headache without sweating, abdominal pain, vomiting and diarrhea. In modern Chinese medicine, MH is generally used as an ingredient of many prescriptions, such as Xinjia Xiangru Yin, Chaihu-Xiangru decoction, and Huanglian Xiangru decoction, which could be used for treatment of common cold due to summer heat and dampness [15], acute upper respiratory infection in summer [16], as well as influenza [17]. Pharmacological researches showed that MH has many biological activities, including antibacterial, antiviral, antioxidant, anti-inflammatory. With the wide application of chromatography and mass spectrometry technologies, the active components of MH were identified, which were roughly divided into volatile components and non-volatile components [18]. So far, approximately 123 compounds were identified from volatile components of MH. Most of them belonged to flavonoids, terpenoids, phenolic acids, and phenylpropanoids [18]. Of which, flavonoids, a group of natural compounds derived from plants and with phenolic structures, was considered to have antiviral activities. The total flavonoids of MH inhibit Influenza A virus through suppressing NOX4/NF-κB/MLCK pathway [19]. And its main components, luteolin and apigenin, have antiviral effects on many viruses, including Japanese encephalitis virus, enterovirus 71 (EV71), respiratory syncytial virus (RSV), and herpes simplex virus 1 (HSV-1) [20,21,22,23]. For SARS-CoV-2, luteolin was considered as an inhibitor of RNA-dependent RNA polymerase (RdRp) and the protease of 3CL [24,25], and a potent blocker of cell entry [26].
Previous researches paid many attentions to the volatile components of MH, however, the herb is traditionally processed as a decoction used for clinical. Whether the aqueous extract of MH (AEMH) has the anti-SARS-CoV-2 activities, that is, whether MH could be a potential herb medicine used for treatment of Covid-19 is the first question that we studied. Next, which components are present in AEMH, and whether flavonoids exist and play an important role are still unknown. Therefore, component analysis of AEMH and explore the active component is essential to understand the antiviral mechanism of AEMH. In this study, we firstly evaluated the anti-SARS-CoV-2 activities of AEMH, and found that AEMH suppressed viral replication intracellularly. Then, the material basis of AEMH was analyzed and a total of 26 components were identified. Integrating network pharmacology and transcriptional profiling, two core regulators was screened and identified. Finally, the active components were screened and verified by molecular docking and in vitro pharmacological experiment. Our results could provide not only a reference for the clinical applications of MH, but also a valuable insight for developing antiviral drugs with herbal medicines.
2. Results
2.1. In Vitro Antiviral Effects of AEMH on SARS-CoV-2
For avoiding the cytotoxicity in antiviral experiments, the maximum non-toxic concentration (MNTC) of AEMH was firstly determined on Vero-E6 cells. When 10 mg/mL of AEMH was inoculated, the cell survival rates were 101.94 ± 4.83%. When AEMH increased to 20 mg/mL, the cell survival rate sharply dropped to 1.44 ± 1.27% (Figure 1a). Hence, the MNTC of AEMH was identified as 10 mg/mL and the CC50 was calculated as 14.31 mg/mL. Also, the MNTC and CC50 of positive control Nirmatrelvir (PF-07321332) was tested as 1 μM and 23.50 μM (Figure 1b).
To evaluate the in vitro antiviral activities of AEMH on SARS-CoV-2, we compared the viral loads of infected cells with or without AEMH treatment, and also investigated the inhibition of AEMH on cell death caused by infection. When the cells were treated with three concentrations of AEMH for 48h after infection, the viral loads of treatment groups were all markedly dropped compared to VC group (P < 0.01), with 67~782 fold reductions (Figure 1C). Cell survival rates further demonstrated that AEMH treatment significantly inhibited the cell death caused by viral infection (Figure 1D), and the inhibition rates have dose-effect relationship with the concentration of AEMH. The median inhibitory concentration (IC50) of AEMH and positive control Nirmatrelvirw was calculated as 170 µg/mL and 80 nM, respectively (Figure 1D and 1E).
2.2. Components Identified from AEMH
For investigating the antiviral mechanism, the material basis of AEMH was analyzed by HPLC-ESI-Q-TOF/MS. The positive ion and negative ion chromatogram are displayed in Figure 2. Trough comparison with database, a total of 27 components were determined from AEMH (Table 1). As one out of the 27, that is, the compound with molecular formula C18H28O9 can’t be exactly distinguished, only 26 components were identified and used for research of network pharmacology.
2.3. Screening of Targets Related to Covid-19 and AEMH
Covid-19 related targets were searched through six databases. There are 61, 214, 40, 540, 1843, and 109 targets obtained from DrugBank, GeneCards, OMIM, NCBI, DisGeNET, and TTD databases, respectively. For screening the AEMH related targets, 26 components identified by HPLC-ESI-Q-TOF/MS were imported to three databases (PharmMapper, SEA and TCMSP), and then 558 related targets were retrieved. Then, by matching Covid-19 related targets with components related targets, a total of 126 targets were found in the intersection, namely the common targets of ‘disease and drug’ (Figure 3A).
Further, we performed Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis based on the 126 intersections targets. From DAVID database, GO enrichment analysis screened 177 GO items (FDR < 0.05). Of which, 119 items were involved in Biological Process (BP), while only 20 were involved in Cellular Component (CC) and 38 were involved in Molecular Function (MF). The bubble charts showed the top 15 pathways that the 126 genes were enriched in BP, CC and MF (Figure 3B-D). KEGG analysis showed that the 126 genes were involved in 124 pathways (FDR<0.05). The top 15 pathways enriched were shown in Figure 3E. Of the 124 pathways, IL-17 signaling pathway is most significantly involved. In addition, the ‘drug-components-targets-disease’ network was constructed using one medicine (AEMH), 26 components, 126 targets, and the disease COVID-19. The network was composed of 154 nodes and 629 edges (Figure 3F).
2.4. Screening and Verification of Core Targets
For seeking the core targets, the 126 intersection genes were input into the STRING database to construct a Protein-Protein Interaction (PPI) network (Figure. 4A). The PPI network contained 119 nodes (119 out of 126 genes) and 1341 edges. Figure 4B showed the global change on the transcriptome between patients infected with Omicron or ancestral strains and health donors, and 1082 genes were found to be differentially expressed in omicron infected patients [27]. Then, 119 genes obtained from PPI were matched with the 1082 differential expression genes. Nine genes, as shown in Venn plot, including Heme oxygenase 1 (HMOX1), Lipocalin 2 (LCN2), Monoamine oxidase B (MAOB), Myeloid Cell Leukemia 1 (MCL1), Matrix metallopeptidase 1 (MMP1), Purine nucleoside phosphorylase (PNP), selectin P (SELP), Triosephosphate isomerase 1 (TPI1) and MAPK activated protein kinase 2 (MAPKAPK2) were found in the intersection region (Figure 4C).
The nine genes were validated by qRT-PCR and the mRNA level was presented with fold-change based on Ct value. Of the nine, the mRNA level of two genes, PNP and TPI1, was markedly downregulated in VC group, and then significantly higher transcribed in AEMH treated group (Figure 5). Significant difference was also observed on relative mRNA expression level of MMP1 gene between VC and AEMH treated group, but there was no difference between VC and CC group. Hence, the PNP and TPI1 were selected as the key regulators.
2.5. Screening and Identification of Active Components
For seeking the single components with anti-SARS-CoV-2 activity, the connection between 26 components and proteins of two core targets were analyzed by molecular docking. The binding affinity energies, as shown in Figure. 6, demonstrated that a total of 20 components with binding energies lower than -6 Kcal/mol with PNP, and the lowest one is Luteolin-7-O-glucuronide (-8.7 Kcal/mol). The overall binding energies between components and TPI1 were higher than that with PNP, and the lowest one was Luteolin-7-O-rutinoside, with binding energy of -6.9 Kcal/mol.
Since not all the components are available, only eight with binding energy lower than -6 Kcal/mol were selected to perform in vitro pharmacological verification (Figure 7). The MNTC was tested as 25 µg/mL for Luteolin 7-O-glucuronide, Luteolin, and Rosmarinic acid, and 50 µg/mL for Apigenin-7-O-glucuronide, 5,6-Dihydroxy-7-methoxyflavone, Luteolin 7-O-rutinoside, and Caffeic acid, while only 12.5 µg/mL for Luteolin-7-O-glucoside. After 48h post infection, viral loads of luteolin, rosmarinic acid, and caffeic acid treated groups remarkably dropped compared to VC, indicating the three components have in vitro antiviral effects on SARS-CoV-2 at MNTC. Additionally, molecular docking between three verified components with the proteins of PNP or TPI1 were conducted, and the results were shown in Figure 8.
3. Discussion
In this study, aqueous extract of MH displayed significant in vitro antiviral effects against SARS-CoV-2 through inhibiting viral replication intracellularly and cell death caused by viral infection, and moreover, the inhibitory effects were dose–dependent. MH or compounds containing MH have been employed to treating fever, influenza, pneumonia in China for a long time [15,16,17]. Previous studies demonstrated that MH and its active components, for example total flavonoids, can inhibit influenza A virus induced CPE in MDCK cells, and also reduce viral loads in lung of infected mice [28,29], which is similar to the results of our study.
As herbal medicine is a multi-component substance, it is necessary to clarify its material basis in order to study the mechanism of antiviral action. The active components of MH has been extensively studied [18,30]. Duan et al. summarized 123 components identified from M. chinensis Maxim and M. chinensis.cv. ‘Jiangxiangru’, and most of them belong to flavonoids, terpenoids, phenolic acids, and phenylpropanoids [18]. Also, Wang et al. identified 69 chemical constituents of M. chinensis.cv. ‘Jiangxiangru’ by UPLC-LTQ-Orbitrap-MS [30]. Compared with previous studies which prepared samples with organic solvent, such as ethanol and methyl alcohol, we analyzed the material basis of MH aqueous extract since traditional CHM is usually used as decoction. In total, 26 components were identified in our study. Of which, nine components including citric acid, succinic acid, caffeic acid, 3,4-Dihydroxybenzoic acid, 4-Hydroxybenzoic acid, Syringic acid, rosmarinic acid, luteolin, and 5,6-Dihydroxy-7-methoxyflavone were reported, while other 17 were firstly identified as far as we know. Flavonoids, Luteolin and Apigenin, were considered as representative existed in MH [28], but we identified only luteolin from AEMH. In addition, eight organic acids were identified; of which, citric acid, succinic acid, caffeic acid, and rosmarinic acid were determined from M. chinensis.cv. ‘Jiangxiangru’ [30].
Due to the complex composition, the action of CHM is considered to be multi-target and multi-pathway. Therefore, it is a challenge to explore the possible mode of action of CHM. In recent years, the development of bioinformatics provides a novel and effective strategy for investigation of antiviral mechanism of CHM. During the epidemic of Covid-19, many researchers investigated the potential mechanism and active components of herbal medicine for treating SARS-CoV-2 through network pharmacology combined with molecular docking, such as Andrographis paniculata and Fuzheng Yugan Mixture [31,32] . In this study, we first retrieved Covid-19 related and AEMH related targets, and then obtained the common targets of ‘drug and disease’. Secondly, the ‘drug-disease’ targets were re-intersected with differential expressed genes of Covid-19 patients [27]. As the transcriptome data was generated from real patients and health donor, the re-intersection could narrow the range of core targets and further increase the accuracy of target screening. As a results, nine genes were found based on twice screening. Subsequently, PNP and TPI1 were identified as core regulators through comparing the relative mRNA expression levels amongst VC, CC, and AEMH treated groups.
In order to explore the single component with anti-SARS-COV-2 activity in AEMH, the 26 components were molecularly docked with PNP and TPI1, respectively. The binding energies indicated that 20 components could form stable bonds with the protein of PNP or TPI1, thereby exerting antiviral activity. Of the 20 components, only eight were selected to do in vitro pharmacological verification. Three components, including two organic acids (caffeic acid and rosmarinic acid) and a flavonoid (luteolin) displayed antiviral effects at their MNTC.
Caffeic acid, a phenolic compound found in most plants, was reported to have significantly inhibitory effects against HCoV-NL63, Influenza virus, Hepatitis C virus, HSV, and ILHV [33,34,35,36,37]. Previous studies put forward that polyphenol have potential to combat with COVID-19, and analyze the binding ability between caffeic acid and its derivatives with SARS-CoV-2 by molecular docking [38]. Of the three components verified in our study, caffeic acid displayed the highest anti-SARS-CoV-2 activity, which proved the above point of view. Rosmarinic acid, an ester of caffeic acid and 3,4-dihydroxyphenyllactic acid, has a wide-range of pharmacological properties, such as antiviral, anti-inflammatory and antioxidant. Evidence from in vitro or in vivo studies demonstrated that rosmarinic acid possesses antiviral activities on Chikungunya virus, Monkeypox, influenza virus, and EV71 viruses [39,40,41,42]. For SARS-CoV-2, crystal structure elucidated that rosmarinic acid can form complex with SARS-CoV-2 Mpro [43]. In the in vitro antiviral experiment, rosmarinic acid exerted anti-SARS-CoV-2 activities with a IC50 of 25.47 ng [44]. Rosmarinic acid significantly inhibited the replication of SARS-CoV-2 in our study, which is similar to the previous reports. Luteolin, a nontoxic and non-mutagenic dietary flavonoid existing in a variety of fruits and vegetables, was considered as a promising antiviral agent for many viruses. For instance, Luteolin inhibited RSV replication though regulating the miR-155/SOCS1/STAT1 signaling pathway [45], inhibited HSV-1 through enhancing type I interferon production [23], and conferred survival protection of 91.67% from the lethal EV71 challenge in new born mice [21]. For SARS-CoV-2, luteolin was reported to display antiviral activities through binding with RdRp, 3CLpro, and spike proteins [24,25,26]. Most of the publications were performed by molecular docking or enzymatic inhibition assay, but few research proved the antiviral effects of luteolin using live virus strain. Our results filled the gap by confirming the anti-SARS-CoV-2 activity of luteolin with real omicron strain. However, Apigenin-7-O-glucoside and Luteolin-7-O-glucoside, which has lower binding energies with PNP in our study, and was reported to have activities against RdRp [46], didn’t show any antiviral activities at MNTC against live virus of SARS-CoV-2.
The limitation of the study is the in vivo anti-SARS-CoV-2 activities of AEMH was not assessed, and also, the mechanism of caffeic acid, rosmarinic acid and luteolin needs to be further studied.
4. Materials and Methods
4.1. Medicines
Moslae Herba was collected and manufactured by Zhejiang Chinese Medical University Medical Pieces., LTD. Crude slice of the herb was provided and authenticated by Prof. Qiaofeng Wu in Zhejiang Chinese Medical University (ZCMU). The AEMH was prepared by the Pharmaceutical Preparation Department of ZCMU. Briefly, the herb was boiled and refluxed in 30 volumes of water (~ 3.33% w/v) for 1h. The extraction process was repeated twice followed by filtration. And then, the filtered liquid was concentrated under reduced pressure to 50 mL and was determined to contain 1 g/mL of crude herb. Nirmatrelvir powder (PF-07321332) (MedChemExpress, Monmouth Junction, NJ, USA) was dissolved in dimethyl sulfoxide (DMSO), making the initial concentration 1 mM. CP-100356 hydrochloride (MedChemExpress, Monmouth Junction, NJ, USA), dissolved to 1 mM with DMSO, was used as efflux inhibitor.
Rosmarinic acid (CAS:20283-92-5) standards was purchased from BBI Life Science Corporation, China. Caffeic acid (CAS: 331-39-5) standard was purchased from National Institutes for Food and Drug Control. Luteolin 7-O-glucuronide (CAS: 29741-10-4) standard was purchased from Baoji Herbest Bio-Tech Co., Ltd., China. 5,6-Dihydroxy-7-methoxyflavone (CAS: 29550-13-8), Apigenin-7-O-glucuronide (CAS: 29741-09-1), Luteolin-7-O-glucoside (CAS: 5373-11-5), and Luteolin 7-O-rutinoside (CAS: 20633-84-5) standards were purchased from Chengdu Desite Biotech Co., Ltd., China. Luteolin (CAS: 491-70-3) standard was purchased from Shanghai Macklin Biochemical Co., Ltd., China. All the standards were dissolved with DMSO, making the initial concentration 10 mg/mL and stored at -20℃.
4.3 Viruses and Cells
SARS-CoV-2 strain SARS-CoV-2/VeroE6/DSh/2021ZJ25 (Omicron/B.1.1/EPI_ISL_ 12040149) was isolated from throat swab of a suspected COVID-19 patient and kept at -80°C in Zhejiang CDC [47]. All experiments involving in live virus were performed in a Biosafety Level 3 (BSL-3) laboratory. The initial titre of the virus stock was 105.6 TCID50/0.1mL. Vero-E6 cells were culture in Minimum Essential Medium supplemented with 10% of fetal bovine serum, 1% L-glutathione and 1% penicillin-streptomycin solution.
4.4. Cytotoxicity of Medicines
The cytotoxicity of AEMH, Nirmatrelvir, and seven chemical components were determined on Vero-E6 cells by the cell counting Kit-8 (CCK-8) (Beyotime Biotechnology, Shanghai). After cell monolayer formed in 96-well plate, the growth medium was decanted and displaced by 100 μL of different concentrations of medicine. Each concentration was performed in five replicates. 100 μL of maintenance medium (MM) instead of medicine was added to cell control. After incubation for 72 hrs, medicine solution was removed and 100 μL of MM containing 10 μL of CCK-8 was added to the cells. The plates were re-incubated for 1.5 h at 37°C and the optical density value was measured at 450 nm after incubation. The highest concentration with cell viability rate higher than 90% was determined as MNTC of the medicine.
4.5. In vitro Antiviral Verification Experiment
The in vitro antiviral activity was validated using both gene amplification reduction assay and cell viability rate assay. In gene amplification reduction experiments, Vero-E6 seeded in 24-well plates were infected with 100 TCID50 of SARS-CoV-2 strain for 1h. The cells were then washed twice with PBS to remove the unbound viral particles. Then, the cells were treated with 1mL of AEMH (4, 2 and 1 mg/mL), Nirmatrelvir (1 μM), or seven chemical compounds with MNTC, respectively. Each concentration of AEMH and Nirmatrelvir were performed in five replicates, while the chemical compounds were performed in triplicates. For virus control (VC) group, the infected cells were treated with 1 mL of MM. After 48h of incubation at 37°C in a 5% CO2 incubator, viral RNA was extracted from each group and viral load was tested by qRT-PCR. The antiviral effects were confirmed by comparing the viral loads between VC and medicine treated groups.
In regard to cell viability rate assay, vero-E6 cells were infected with 100 TCID50 SARS-CoV-2 in 96-well plates for 1 h at 37°C. After infection, the virus fluid was decanted and 100 μL of different concentrations of medicine was added to the cells. For VC, the cells were covered with 100 µL MM without medicine after infection. Each concentration was performed in five replicates. After incubation for 72hrs, cell viability rate was measured by the same assay as for the cytotoxicity testing. The IC50 of medicine was calculated using the Graphpad software (version 6.01). Results are shown as the mean ± standard deviation.
4.6. HPLC-ESI-Q-TOF/MS Conditions
Component analysis of AEMH was performed on an Agilent1260-6530 QTOF system (Agilent, CA, USA). Separation was carried out on an Agilent ZORBAX Eclipse XDB-C18 column (5µm, 4.6*250mm) (Agilent, CA, USA) with the injection volume of 10 μL (The aliquot of 1mL AEMH (1g/mL) was diluted with 25 mL water and then processed with 0.22 µm filter). Methanol was used as mobile phase A and 0.05% acetic acid solution as mobile phase B. The gradient elution program was shown in Supplementary Table S1. The flow rate was1 mL/min. The column temperature was 30°C and the detection wavelengths are 254 nm and 360 nm (full wavelength enabled). The mass spectrometer was operated in the positive and negative ion mode. Other parameters were set as follows: capillary voltage (Vcap), 3.5 KV; nebulizer pressure, 35 psig; capillary outlet voltage (Fragmentor), 65 V; skimmer voltage, 65 V; dring Gas flow rate, 10 L/min; collision energy, 20 V; scanning range, 100~1700 m/z. MS data was acquired by Waters MassLynx 4.1 and processed using UNIFI 1.7 software (Waters).
4.7. Network Pharmacology
4.7.1. Screening of the SARS-CoV-2 Related Targets and the AEMH Related Targets
SARS-CoV-2 related targets were screened from six databases, including DrugBank (https://www.drugbank.ca/), GeneCards (https://www.genecards.org/), OMIM (https://www.omim.org/), NCBI (https://www.ncbi.nlm.nih.gov/), DisGeNET (https://www.disgenet.org/home/) and TTD (http://db.idrblab.net/ttd/) with keywords ‘COVID-19’, ‘Coronavirus 2019’, ‘Novel coronavirus’ or ‘Coronavirus disease 2019’. Of the targets searched from GeneCards, those with scores ≥10.0 were selected as SARS-CoV-2 related targets.
In total of 26 components identified by the HPLC-ESI-Q-TOF/MS were used for screening of AEMH related targets. Briefly, the components which was downloaded from PUbChem (https://pubchem.ncbi.nlm.nih.gov/) in SDF format were imported into PharmMapper server database (http://www.lilab-ecust.cn/pharmmapper/) for target searching. Genes with Norm Fit ≥ 0.9 were selected as targets. Another two databases SEA (https://sea.bkslab.org/) and TCMSP (https://old.tcmsp-e.com/tcmsp.php) were used to retrieve the component related targets as well. All the targets were converted into the uniform gene names using the UniProt protein database (http://www.uniprot.org/).
The target intersections were analyzed by matching the SARS-CoV-2 related targets and the AEMH related targets. Venn diagram drawn by package VennDiagram (https://cran.r-project.org/web/packages/VennDiagram/index.html). Network of medicine-components-targets-disease was drawn using Cytoscape (version 3.9.1) software. Genes in the intersection were imported to David online database (https://david.ncifcrf.gov/tools.jsp), and the Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment and Gene Ontology (GO) analysis was conducted.
4.7.2. Screening of Core Targets
For screening core targets, the PPI network was constructed based on the genes in the intersection using String database (http://string.embl.de/) and visualized by Cytoscape (version 3.9.1) software. Then, the differential expressed genes between healthy donor and patients infected with omicron or ancestral strain were analyzed based on the the transcriptome data of platelets published by Hong Wang et al. [27]. Finally, the genes obtained from PPI were intersected with the differential expressed genes, and the genes in the intersection were defined as core targets and used for verification.
4.7. RNA Extraction and Quantitative real-Time PCR (qRT-PCR)
Genomic RNA was extracted from 200 µL of culture fluid and eluted into 30 µL RNase-free water using RNeasy Mini Kit (Qiagen, Hilden, Germany) according to manufacturer’s instructions. In antiviral experiments, the extracted vRNA was quantitatively detected by the Nucleic Acid Detection Kit for 2019-nCoV (EDIAGNOSIS, Wuhan, China). The pseudovirus standard (High value H1) used for quantify the viral loads of N gene of SARS-CoV-2 was purchased from FANTASIABIO. Co. Ltd. The initial concentration of the standard is 107 copies/mL. qRT-PCR was also used to validate the changes in the transcripts of core targets selected. The extracted RNA was detected by One Step TB Green™ PrimeScript PLUS RT-PCR Kit (Perfect Real-time) (TaKaRa, Kyoto, Japan). Primers used for detection of the nine host genes were shown in Supplementary Table S1. The relative RNA expression was calculated using a classical 2−ΔΔCt method after normalizing against the expression level of β-actin.
4.8. Molecular Docking
The 3D structures of two core target proteins, PNP (PDB ID: 1V3Q) and TPI1 (PDB ID: 6UP5), were downloaded from the RSCB PDB (https://www. rcsb.org/). The proteins were preprocessed using PyMOL 2.3 software, including water removal and active site definition. The processed proteins were then saved as PDB format. After that, the 26 components of AEMH were imported into Chemdraw3D (version 20.0) for energy minimization and saved as PDB files. The proteins and the components were subsequently added hydrogens and converted to PDBQT format with the AutoDock Tools 1.5.7. Molecular docking simulations were performed with AutoDock vina (version 1.2.2). The binding mode, affinity, and key interactions were analyzed using PyMOL 2.5.4 software.
4.9. Statistic Analysis
Results of viral loads and mRNA expression level are shown as the mean ± standard deviation. Ordinary one-way ANOVA (multiple comparison) was used to compare the significance of differences between the means of the VC and each medicine treated groups. All statistical tests were considered statistically significant at P < 0.05.
5. Conclusions
In summary, aqueous abstract of herbal medicine MH has in vitro antiviral effects on SARS-CoV-2. It may exert antiviral effects through regulating PNP and TPI1 genes on cells. Caffeic acid, rosmarinic acid, and luteolin were identified as single components with anti-SARS-CoV-2 activities from AEMH. Our results could provide detailed date and reference for the clinical application of MH, and also a valuable insight for developing antiviral drugs with herbal medicines.
Author Contributions
Conceptualization, Y.F. and C.X.; methodology, J.G. and Q. G; software, J.G.; validation, Q.G. and Y.Z.; formal analysis, Y.F.; investigation, Y.F., Y.Z., Q.G., J.G, and Z.W.; resources, H.M.; data curation, Z.W, and Y.Z.; writing—original draft preparation, Y.F; writing—review and editing, C.X.; visualization, Y.F.; supervision, H.M and B.W.; project administration, H.M. and C.X.; funding acquisition, B.W. All authors have read and agreed to the published version of the manuscript.”
Funding
This research was funded by the Major Program of Natural Science Foundation of Zhejiang, grant number LDT23H19013H19, and the National Health Commission Scientific Research Projects, grant number WKJ-ZJ-2220.
Institutional Review Board Statement
The study project was conducted in accordance with the Declaration of Helsinki, and approved by the Ethics Committee of Zhejiang provincial Center for Disease Control and Prevention (Zhejiang CDC), China (approve no. 2020-015). SARS-CoV-2 strain used is an isolate stored in laboratory of Zhejiang CDC; hence there was no operation of sampling in this study and pose no extra burden for patient. In addition, the study does not involve other clinical samples, human or animals.
Data Availability Statement
We encourage all authors of articles published in MDPI journals to share their research data. In this section, please provide details regarding where data supporting reported results can be found, including links to publicly archived datasets analyzed or generated during the study. Where no new data were created, or where data is unavailable due to privacy or ethical restrictions, a statement is still required. Suggested Data Availability Statements are available in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Acknowledgments
We are grateful to Zhejiang Conba Pharmaceutical Co., Ltd. for assisting in the HPLC-Q-TOF-MS/MS analysis.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Word Health Organization (WH). WHO COVID-19 dashboard. Available online: https://data.who.int/dashboards/covid19/cases?n=c (accessed on November 17, 2024).
- Chan, J.F.-W.; Yuan, S.; Chu, H.; Sridhar, S.; Yuen, K.-Y. COVID-19 drug discovery and treatment options. Nat. Rev. Microbiol. 2024, 22, 391–407. [CrossRef]
- Choi, H.S.; Choi, A.Y.; Kopp, J.B.; Winkler, C.A.; Cho, S.K. Review of COVID-19 Therapeutics by Mechanism: From Discovery to Approval. J. Korean Med Sci. 2024, 39, e134. [CrossRef]
- Owen, D.R.; Allerton, C.M.N.; Anderson, A.S.; Aschenbrenner, L.; Avery, M.; Berritt, S.; Boras, B.; Cardin, R.D.; Carlo, A.; Coffman, K.J.; et al. An oral SARS-CoV-2 M pro inhibitor clinical candidate for the treatment of COVID-19. Science 2021, 374, 1586–1593. [CrossRef]
- Iketani, S.; Iketani, S.; Liu, L.; Liu, L.; Guo, Y.; Guo, Y.; Liu, L.; Liu, L.; Chan, J.F.-W.; Chan, J.F.-W.; et al. Antibody evasion properties of SARS-CoV-2 Omicron sublineages. Nature 2022, 604, 553–556. [CrossRef]
- Cao, Y.; Wang, J.; Jian, F.; Xiao, T.; Song, W.; Yisimayi, A.; Huang, W.; Li, Q.; Wang, P.; An, R.; et al. Omicron escapes the majority of existing SARS-CoV-2 neutralizing antibodies. Nature 2021, 602, 657–663. [CrossRef]
- Wang, C.; Wang, H.; Liu, X.; Xu, D.; Tang, Y.; Luo, P. Traditional Chinese Medicine for the treatment of influenza: a systematic review and Meta-analysis of randomized controlled trials. J. Tradit. Chin. Med. 2014, 34, 527–531. [CrossRef]
- Duan, Z.-P.; Jia, Z.-H.; Zhang, J.; Liu, S.; Chen, Y.; Liang, L.-C.; Zhang, C.-Q.; Zhang, Z.; Sun, Y.; Zhang, S.-Q.; et al. Natural herbal medicine Lianhuaqingwen capsule anti-influenza A (H1N1) trial: a randomized, double blind, positive controlled clinical trial.. 2011, 124, 2925–33.
- Hung, S.-W.; Liao, Y.-C.; Chi, I.-C.; Lin, T.-Y.; Lin, Y.-C.; Lin, H.-J.; Huang, S.-T. Integrated Chinese herbal medicine and Western medicine successfully resolves spontaneous subcutaneous emphysema and pneumomediastinum in a patient with severe COVID-19 in Taiwan: A case report. EXPLORE 2021, 19, 147–152. [CrossRef]
- Fan, A.Y.; Gu, S.; Alemi, S.F. Chinese herbal medicine for COVID-19: Current evidence with systematic review and meta-analysis. J. Integr. Med. 2020, 18, 385–394. [CrossRef]
- Han, R.; Xie, Y.; Zhao, H.; Li, B.; Yu, X.; Wang, M.; Li, S.; Li, J. The efficacy of traditional Chinese medicine in the treatment of the COVID-19 pandemic in Henan Province: a retrospective study. Eur. J. Med Res. 2023, 28, 1–10. [CrossRef]
- Brevini, T.; Maes, M.; Webb, G.J.; John, B.V.; Fuchs, C.D.; Buescher, G.; Wang, L.; Griffiths, C.; Brown, M.L.; Scott, W.E.; et al. FXR inhibition may protect from SARS-CoV-2 infection by reducing ACE2. Nature 2022, 615, 134–142. [CrossRef]
- He, M.-F.; Liang, J.-H.; Shen, Y.-N.; Zhang, J.-W.; Liu, Y.; Yang, K.-Y.; Liu, L.-C.; Wang, J.; Xie, Q.; Hu, C.; et al. Glycyrrhizin Inhibits SARS-CoV-2 Entry into Cells by Targeting ACE2. Life 2022, 12, 1706. [CrossRef]
- Qian, L.; Zeng, Z. Network-based pharmacology and UHPLC-Q-Exactive-Orbitrap-MS reveal Jinhua Qinggan granule's mechanism in reducing cellular inflammation in COVID-19. Front. Immunol. 2024, 15, 1382524. [CrossRef]
- Yuan, H.; Sun, Y.X. Treating 200 cases of cold of the Shushi type with Xinjia Xiangru Yin. Clin J Chin Med 2018, 26, 122-123. [CrossRef]
- Yao, J.C.; Ying, Y.F.; Zheng, H.W.; Hu, G.H.; Lin, Y.P. Study of Chaihu-Xiangru decoction on acute upper respiratory infection in summer. Mod J of Integr Tradit Chin West Med 2005, 4, 441-442.
- Wu, Q.-F.; Zhu, W.-R.; Yan, Y.-L.; Zhang, X.-X.; Jiang, Y.-Q.; Zhang, F.-L. Anti-H1N1 influenza effects and its possible mechanism of Huanglian Xiangru Decoction. J. Ethnopharmacol. 2016, 185, 282–288. [CrossRef]
- Duan, Z.-Y.; Sun, Y.-P.; Wang, Z.-B.; Kuang, H.-X. Moslae Herba: Botany, Traditional Uses, Phytochemistry, and Pharmacology. Molecules 2024, 29, 1716. [CrossRef]
- Yu, W.-Y.; Li, L.; Wu, F.; Zhang, H.-H.; Fang, J.; Zhong, Y.-S.; Yu, C.-H. Moslea Herba flavonoids alleviated influenza A virus-induced pulmonary endothelial barrier disruption via suppressing NOX4/NF-κB/MLCK pathway. J. Ethnopharmacol. 2020, 253, 112641. [CrossRef]
- Fan, W.; Qian, S.; Qian, P.; Li, X. Antiviral activity of luteolin against Japanese encephalitis virus. Virus Res. 2016, 220, 112–116. [CrossRef]
- Dai, W.; Bi, J.; Li, F.; Wang, S.; Huang, X.; Meng, X.; Sun, B.; Wang, D.; Kong, W.; Jiang, C.; et al. Antiviral Efficacy of Flavonoids against Enterovirus 71 Infection in Vitro and in Newborn Mice. Viruses 2019, 11, 625. [CrossRef]
- Wang, S.; Ling, Y.; Yao, Y.; Zheng, G.; Chen, W. Luteolin inhibits respiratory syncytial virus replication by regulating the MiR-155/SOCS1/STAT1 signaling pathway. Virol. J. 2020, 17, 1–14. [CrossRef]
- Wang, Y.; Li, F.; Wang, Z.; Song, X.; Ren, Z.; Wang, X.; Wang, Y.; Zheng, K. Luteolin inhibits herpes simplex virus 1 infection by activating cyclic guanosine monophosphate-adenosine monophosphate synthase-mediated antiviral innate immunity. Phytomedicine 2023, 120, 155020. [CrossRef]
- Munafò, F.; Donati, E.; Brindani, N.; Ottonello, G.; Armirotti, A.; De Vivo, M. Quercetin and luteolin are single-digit micromolar inhibitors of the SARS-CoV-2 RNA-dependent RNA polymerase. Sci. Rep. 2022, 12, 1–9. [CrossRef]
- Wang, W.; Yang, C.; Xia, J.; Li, N.; Xiong, W. Luteolin is a potential inhibitor of COVID-19: An in silico analysis. Medicine 2023, 102, e35029. [CrossRef]
- Shadrack, D.M.; Deogratias, G.; Kiruri, L.W.; Onoka, I.; Vianney, J.-M.; Swai, H.; Nyandoro, S.S. Luteolin: a blocker of SARS-CoV-2 cell entry based on relaxed complex scheme, molecular dynamics simulation, and metadynamics. J. Mol. Model. 2021, 27, 1–15. [CrossRef]
- Wang, H.; Liu, C.; Xie, X.; Niu, M.; Wang, Y.; Cheng, X.; Zhang, B.; Zhang, D.; Liu, M.; Sun, R.; et al. Multi-omics blood atlas reveals unique features of immune and platelet responses to SARS-CoV-2 Omicron breakthrough infection. Immunity 2023, 56, 1410–1428.e8. [CrossRef]
- Liu, A.-L.; Liu, B.; Qin, H.-L.; Lee, S.; Wang, Y.-T.; Du, G.-H. Anti-Influenza Virus Activities of Flavonoids from the Medicinal PlantElsholtzia rugulosa. Planta Medica 2008, 74, 847–851. [CrossRef]
- Wu, Q.; Yu, C.; Yan, Y.; Chen, J.; Zhang, C.; Wen, X. Antiviral flavonoids from Mosla scabra. Fitoterapia 2009, 81, 429–433. [CrossRef]
- Wang, X.; Cheng, K.; Liu, Z.; Sun, Y.; Zhou, L.; Xu, M.; Dai, X.; Xiong, Y.; Zhang, H. Bioactive constituents of Mosla chinensis-cv. Jiangxiangru ameliorate inflammation through MAPK signaling pathways and modify intestinal microbiota in DSS-induced colitis mice. Phytomedicine 2021, 93, 153804. [CrossRef]
- Nguyen, H.T.; Do, V.M.; Phan, T.T.; Huynh, D.T.N. The Potential of Ameliorating COVID-19 and Sequelae From Andrographis paniculata via Bioinformatics. Bioinform. Biol. Insights 2023, 17. [CrossRef]
- Jiang, X.M.; Zhou, J.; Yu, Z.; Gu, X.M.; Lu, Y.M.; Ruan, Y.M.; Wang, T.M. Exploration of Fuzheng Yugan Mixture on COVID-19 based on network pharmacology and molecular docking. Medicine 2023, 102, e32693. [CrossRef]
- Weng, J.-R.; Lin, C.-S.; Lai, H.-C.; Lin, Y.-P.; Wang, C.-Y.; Tsai, Y.-C.; Wu, K.-C.; Huang, S.-H. Antiviral activity of Sambucus FormosanaNakai ethanol extract and related phenolic acid constituents against human coronavirus NL63. Virus Res. 2019, 273, 197767–197767. [CrossRef]
- Kanazawa, R.; Morimoto, R.; Horio, Y.; Sumitani, H.; Isegawa, Y. Inhibition of influenza virus replication by Apiaceae plants, with special reference to Peucedanum japonicum (Sacna) constituents. J. Ethnopharmacol. 2022, 292, 115243. [CrossRef]
- Shen, J.; Wang, G.; Zuo, J. Caffeic acid inhibits HCV replication via induction of IFNα antiviral response through p62-mediated Keap1/Nrf2 signaling pathway. Antivir. Res. 2018, 154, 166–173. [CrossRef]
- Yamasaki, H.; Ikeda, K.; Tsujimoto, K.; Uozaki, M.; Nishide, M.; Suzuki, Y.; Koyama, A.H. Inhibition of multiplication of herpes simplex virus by caffeic acid. Int. J. Mol. Med. 2011, 28, 595–598. [CrossRef]
- Saivish, M.V.; Pacca, C.C.; da Costa, V.G.; Menezes, G.d.L.; da Silva, R.A.; Nebo, L.; da Silva, G.C.D.; Milhim, B.H.G.d.A.; Teixeira, I.d.S.; Henrique, T.; et al. Caffeic Acid Has Antiviral Activity against Ilhéus Virus In Vitro. Viruses 2023, 15, 494. [CrossRef]
- Adem, S.; Eyupoglu, V.; Sarfraz, I.; Rasul, A.; Zahoor, A.F.; Ali, M.; Abdalla, M.; Ibrahim, I.M.; A Elfiky, A. Caffeic acid derivatives (CAFDs) as inhibitors of SARS-CoV-2: CAFDs-based functional foods as a potential alternative approach to combat COVID-19. Phytomedicine 2020, 85, 153310–153310. [CrossRef]
- Liao, X.; Xin, J.; Yu, Z.; Yan, W.; Li, C.; Cao, L.; Zhang, H.; Wang, W. Unlocking the antiviral potential of rosmarinic acid against chikungunya virus via IL-17 signaling pathway. Front. Cell. Infect. Microbiol. 2024, 14, 1396279. [CrossRef]
- Hu, X.; An, S.; Chu, J.; Liang, B.; Liao, Y.; Jiang, J.; Lin, Y.; Ye, L.; Liang, H. Potential Inhibitors of Monkeypox Virus Revealed by Molecular Modeling Approach to Viral DNA Topoisomerase I. Molecules 2023, 28, 1444. [CrossRef]
- Jheng, J.-R.; Hsieh, C.-F.; Chang, Y.-H.; Ho, J.-Y.; Tang, W.-F.; Chen, Z.-Y.; Liu, C.-J.; Lin, T.-J.; Huang, L.-Y.; Chern, J.-H.; et al. Rosmarinic acid interferes with influenza virus A entry and replication by decreasing GSK3β and phosphorylated AKT expression levels. J. Microbiol. Immunol. Infect. 2022, 55, 598–610. [CrossRef]
- Lin, W.-Y.; Yu, Y.-J.; Jinn, T.-R. Evaluation of the virucidal effects of rosmarinic acid against enterovirus 71 infection via in vitro and in vivo study. Virol. J. 2019, 16, 1–9. [CrossRef]
- Li, Q.; Zhou, X.; Wang, W.; Xu, Q.; Wang, Q.; Li, J. Structural basis of rosmarinic acid inhibitory mechanism on SARS-CoV-2 main protease. Biochem. Biophys. Res. Commun. 2024, 724, 150230. [CrossRef]
- Elebeedy, D.; Elkhatib, W.F.; Kandeil, A.; Ghanem, A.; Kutkat, O.; Alnajjar, R.; Saleh, M.A.; El Maksoud, A.I.A.; Badawy, I.; Al-Karmalawy, A.A. Anti-SARS-CoV-2 activities of tanshinone IIA, carnosic acid, rosmarinic acid, salvianolic acid, baicalein, and glycyrrhetinic acid between computational and in vitro insights. RSC Adv. 2021, 11, 29267–29286. [CrossRef]
- Wang, S.; Ling, Y.; Yao, Y.; Zheng, G.; Chen, W. Luteolin inhibits respiratory syncytial virus replication by regulating the MiR-155/SOCS1/STAT1 signaling pathway. Virol. J. 2020, 17, 1–14. [CrossRef]
- Majrashi, T.A.; El Hassab, M.A.; Mahmoud, S.H.; Mostafa, A.; Wahsh, E.A.; Elkaeed, E.B.; Hassan, F.E.; Eldehna, W.M.; Abdelgawad, S.M. In vitro biological evaluation and in silico insights into the antiviral activity of standardized olive leaves extract against SARS-CoV-2. PLOS ONE 2024, 19, e0301086. [CrossRef]
- Li, J.; Li, X.; Wang, E.; Yang, J.; Li, J.; Huang, C.; Zhang, Y.; Chen, K. Neutralizing Antibodies against the SARS-CoV-2 Delta and Omicron BA.1 following Homologous CoronaVac Booster Vaccination. Vaccines 2022, 10, 2111. [CrossRef]
Figure 1.
Evaluation of in vitro anti-SARS-CoV-2 activities of AEMH. (A) Cytotoxicity induced by AEMH on Vero-E6 cells. (B) Cytotoxicity induced by Nirmatrelvir on Vero-E6 cells. (C) SARS-CoV-2 viral loads on cells with or without AEMH treatment. Vero-E6 cells were infected with 100 TCID50 of SARS-CoV-2 and treated with different concentrations of AEMH or Nirmatrelvir (1 μM) for 48 h. Viral loads were tested by quantitative real-time PCR (qRT-PCR) targeting N gene of SARS-CoV-2. **, significant with P < 0.01 compared to VC. (D) AEMH inhibited cell deaths induced by SARS-CoV-2 infection. (E) Nirmatrelvir inhibited cell deaths induced by SARS-CoV-2 infection.
Figure 1.
Evaluation of in vitro anti-SARS-CoV-2 activities of AEMH. (A) Cytotoxicity induced by AEMH on Vero-E6 cells. (B) Cytotoxicity induced by Nirmatrelvir on Vero-E6 cells. (C) SARS-CoV-2 viral loads on cells with or without AEMH treatment. Vero-E6 cells were infected with 100 TCID50 of SARS-CoV-2 and treated with different concentrations of AEMH or Nirmatrelvir (1 μM) for 48 h. Viral loads were tested by quantitative real-time PCR (qRT-PCR) targeting N gene of SARS-CoV-2. **, significant with P < 0.01 compared to VC. (D) AEMH inhibited cell deaths induced by SARS-CoV-2 infection. (E) Nirmatrelvir inhibited cell deaths induced by SARS-CoV-2 infection.
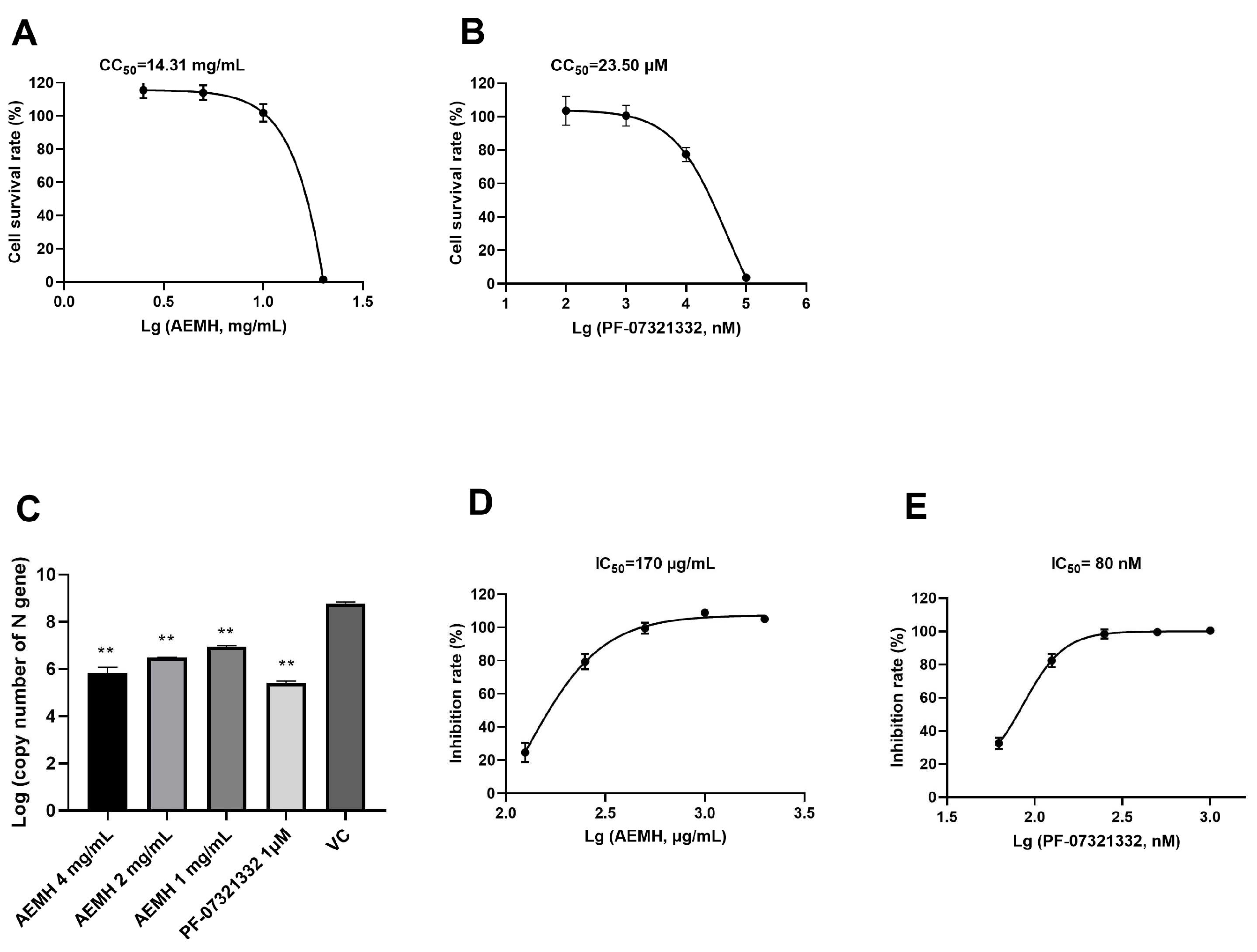
Figure 2.
Components analysis performed by HPLC-ESI-Q-TOF/MS. The top is negative ion chromatogram and the bottom is positive ion chromatogram of AEMH.
Figure 2.
Components analysis performed by HPLC-ESI-Q-TOF/MS. The top is negative ion chromatogram and the bottom is positive ion chromatogram of AEMH.
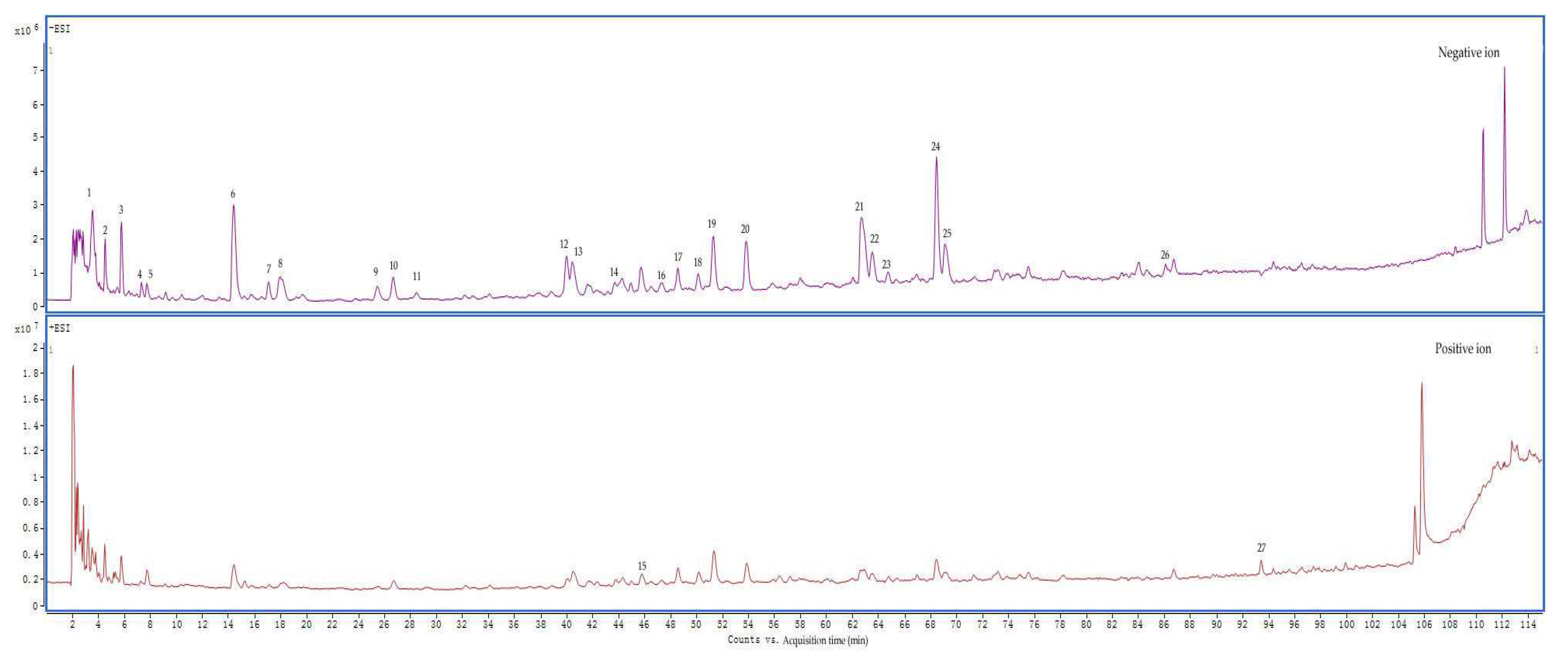
Figure 3.
Screening and analysis of targets related to Covid-19 and AEMH. (A) Venn plot constructed based on Covid-19 related targets retrieved from six databases and the related targets of 26 components identified from AEMH. (B-D) GO enrichment analysis of 126 intersection genes. The top 15 items involved in BP (B), CC (C), and MF (D). (E) The top 15 pathways enriched in KEGG analysis (FDR<0.05). (F) AEMH-components-targets-COVID-19 network diagram.
Figure 3.
Screening and analysis of targets related to Covid-19 and AEMH. (A) Venn plot constructed based on Covid-19 related targets retrieved from six databases and the related targets of 26 components identified from AEMH. (B-D) GO enrichment analysis of 126 intersection genes. The top 15 items involved in BP (B), CC (C), and MF (D). (E) The top 15 pathways enriched in KEGG analysis (FDR<0.05). (F) AEMH-components-targets-COVID-19 network diagram.
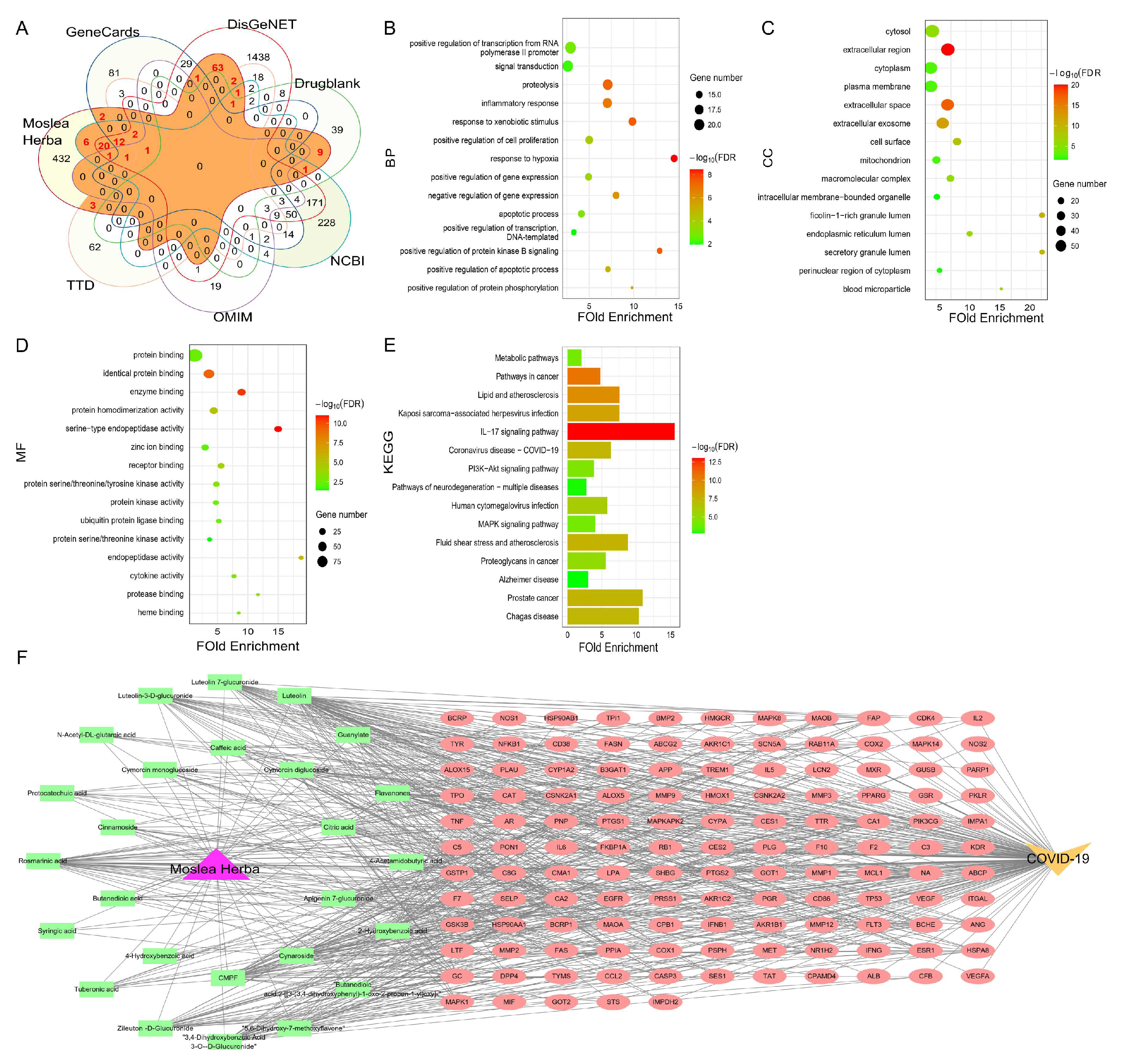
Figure 4.
Screening of core targets. (A) PPI network diagram. (B) Heat map plotted with differential expressed genes of patients in comparison with healthy donors. (C) Venn plot constructed based on 119 genes obtained in PPI with 1082 differential expression genes of patients infected with omicron.
Figure 4.
Screening of core targets. (A) PPI network diagram. (B) Heat map plotted with differential expressed genes of patients in comparison with healthy donors. (C) Venn plot constructed based on 119 genes obtained in PPI with 1082 differential expression genes of patients infected with omicron.
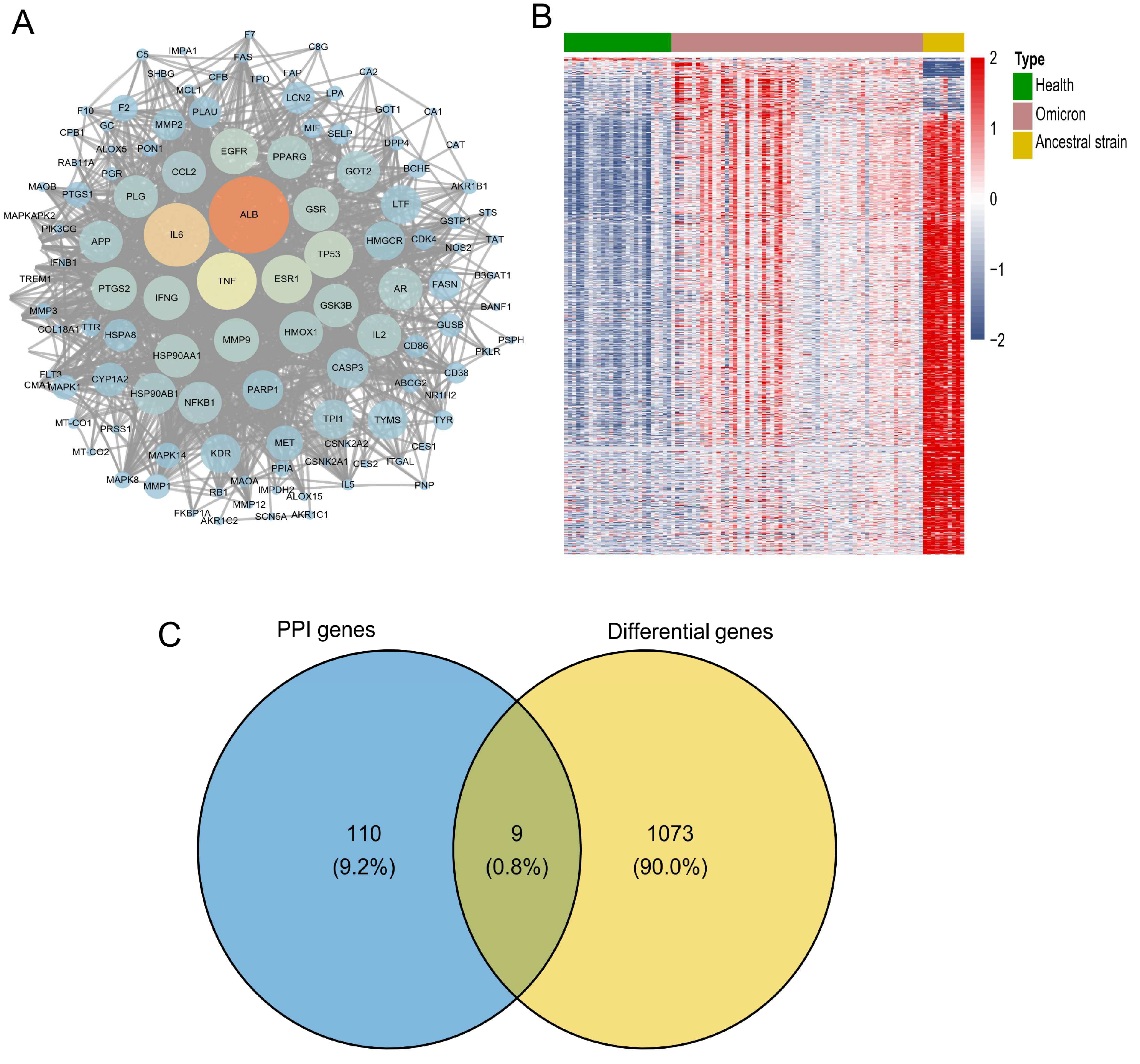
Figure 5.
Relative mRNA expression level of the selected nine genes. Each gene contains data from CC, VC and AEMH treated groups. Experiments were performed in four replicates.
Figure 5.
Relative mRNA expression level of the selected nine genes. Each gene contains data from CC, VC and AEMH treated groups. Experiments were performed in four replicates.
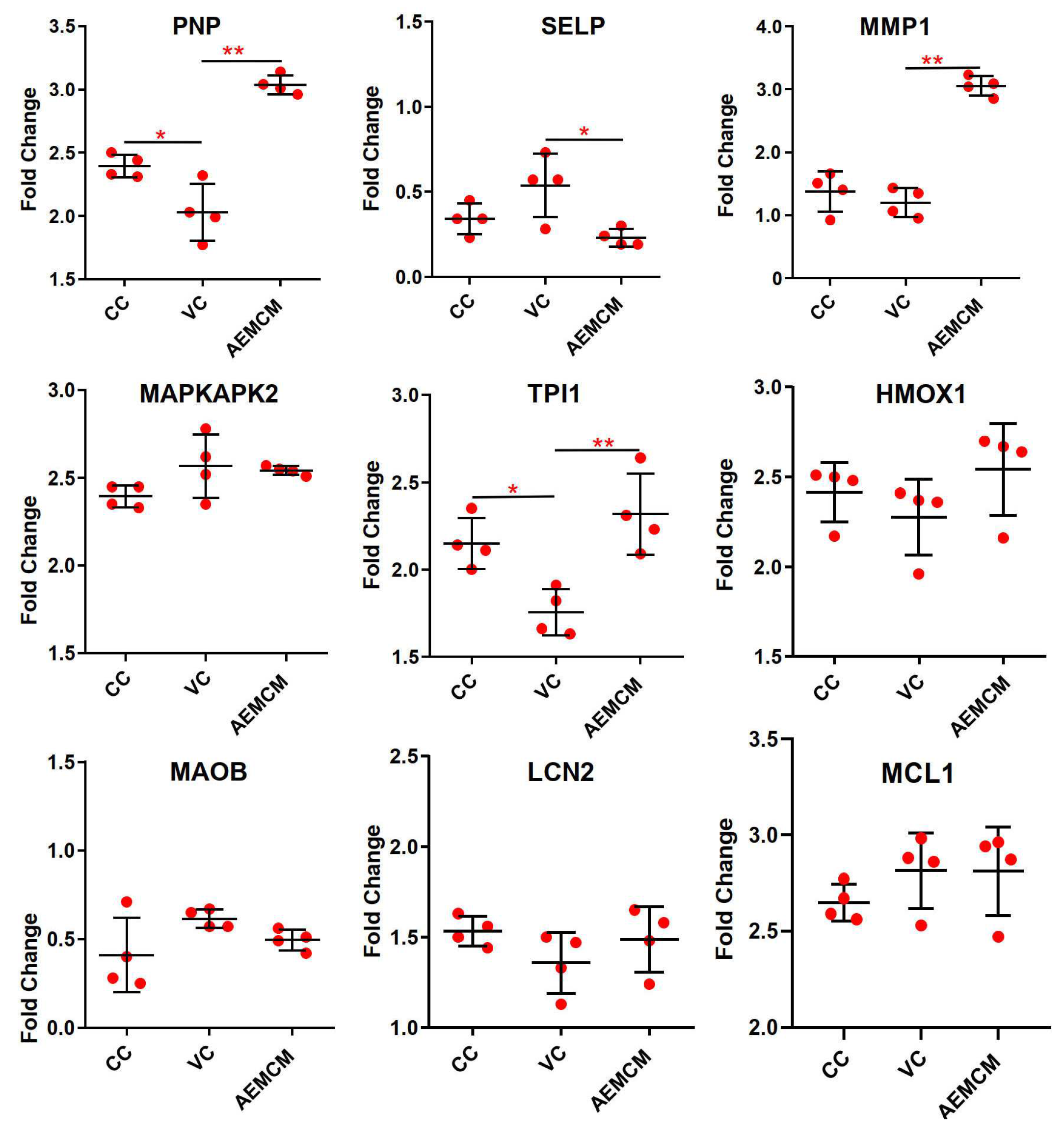
Figure 6.
Binding affinity energies between core targets and 26 components. The binding affinity energy was evaluated by the docking scores of components and core targets.
Figure 6.
Binding affinity energies between core targets and 26 components. The binding affinity energy was evaluated by the docking scores of components and core targets.
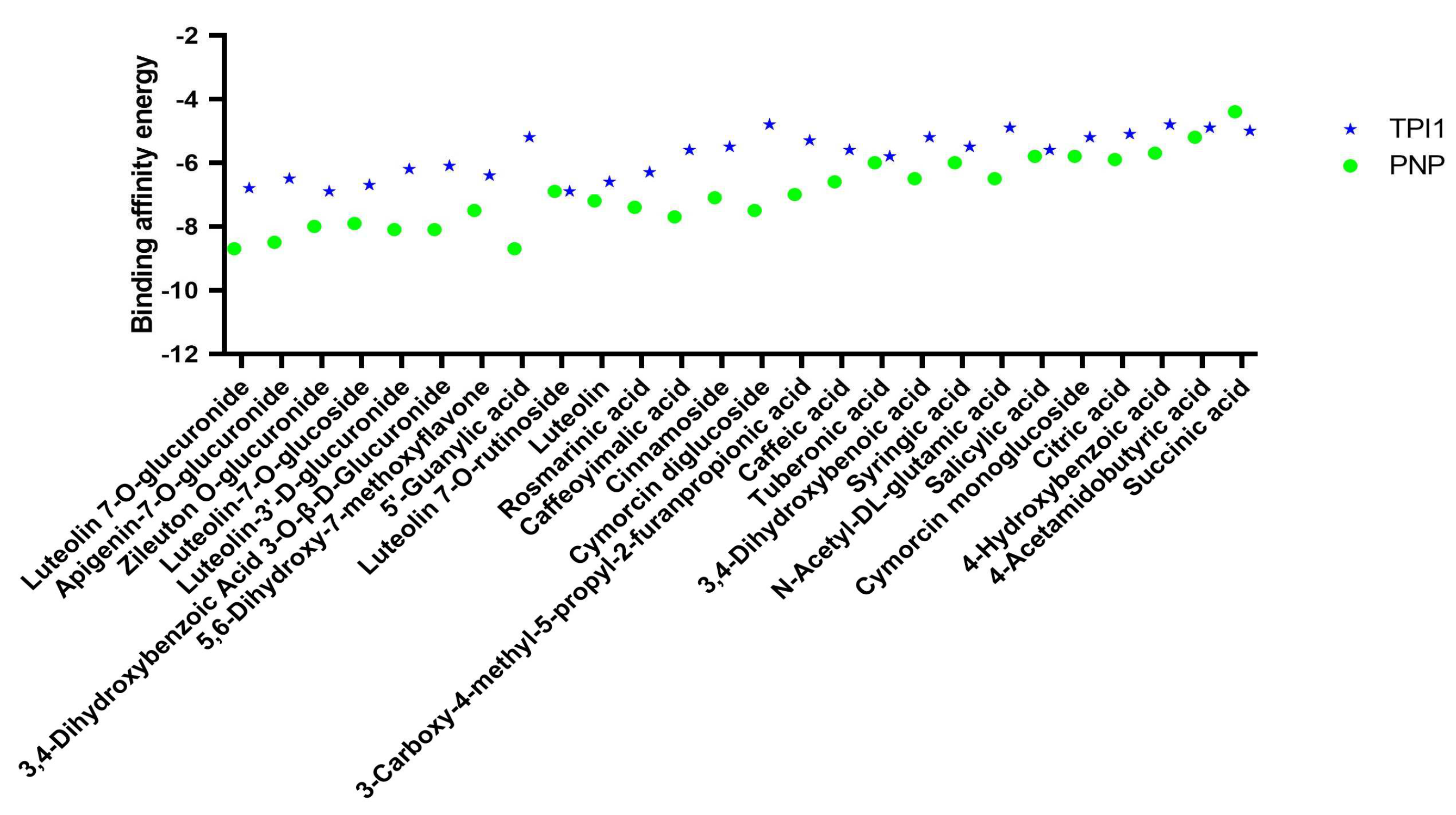
Figure 7.
In vitro pharmacological verification of eight selected components. Vero-E6 cells were infected with 100 TCID50 of SARS-CoV-2 and then treated with each component at MNTC or 4 mg/mL of AEMH for 48 h. Viral loads were tested by qRT-PCR targeting N gene of SARS-CoV-2. *, significant with P < 0.05 compared to VC. **, significant with P < 0.01 compared to VC.
Figure 7.
In vitro pharmacological verification of eight selected components. Vero-E6 cells were infected with 100 TCID50 of SARS-CoV-2 and then treated with each component at MNTC or 4 mg/mL of AEMH for 48 h. Viral loads were tested by qRT-PCR targeting N gene of SARS-CoV-2. *, significant with P < 0.05 compared to VC. **, significant with P < 0.01 compared to VC.
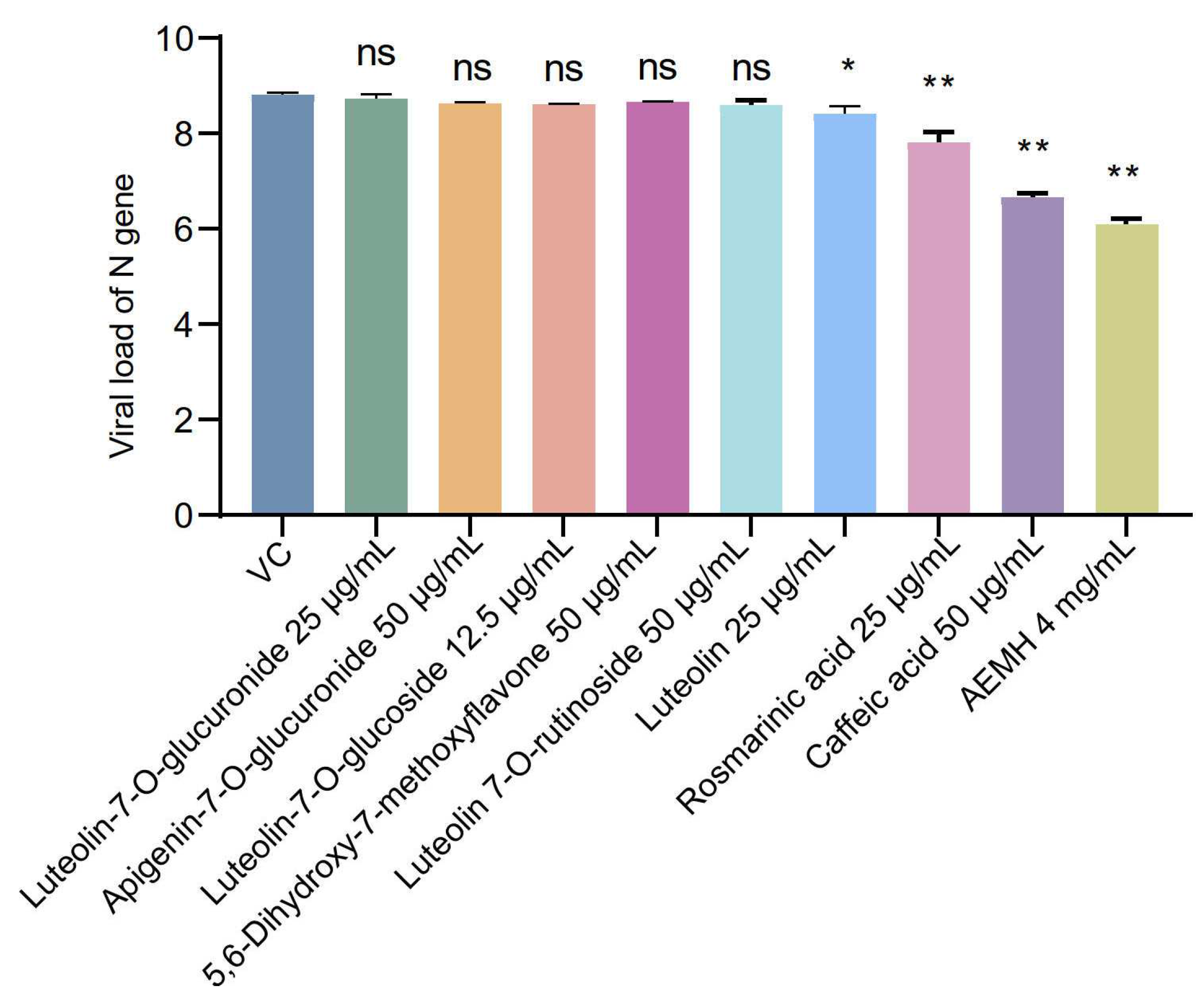
Figure 8.
Binding patterns between three verified components and two core targets. Binding pattern between PNP to caffeic acid (A), rosmarinic acid (B), and luteolin (C). Binding pattern between TPI1 to caffeic acid (D), rosmarinic acid (E), and luteolin (F).
Figure 8.
Binding patterns between three verified components and two core targets. Binding pattern between PNP to caffeic acid (A), rosmarinic acid (B), and luteolin (C). Binding pattern between TPI1 to caffeic acid (D), rosmarinic acid (E), and luteolin (F).
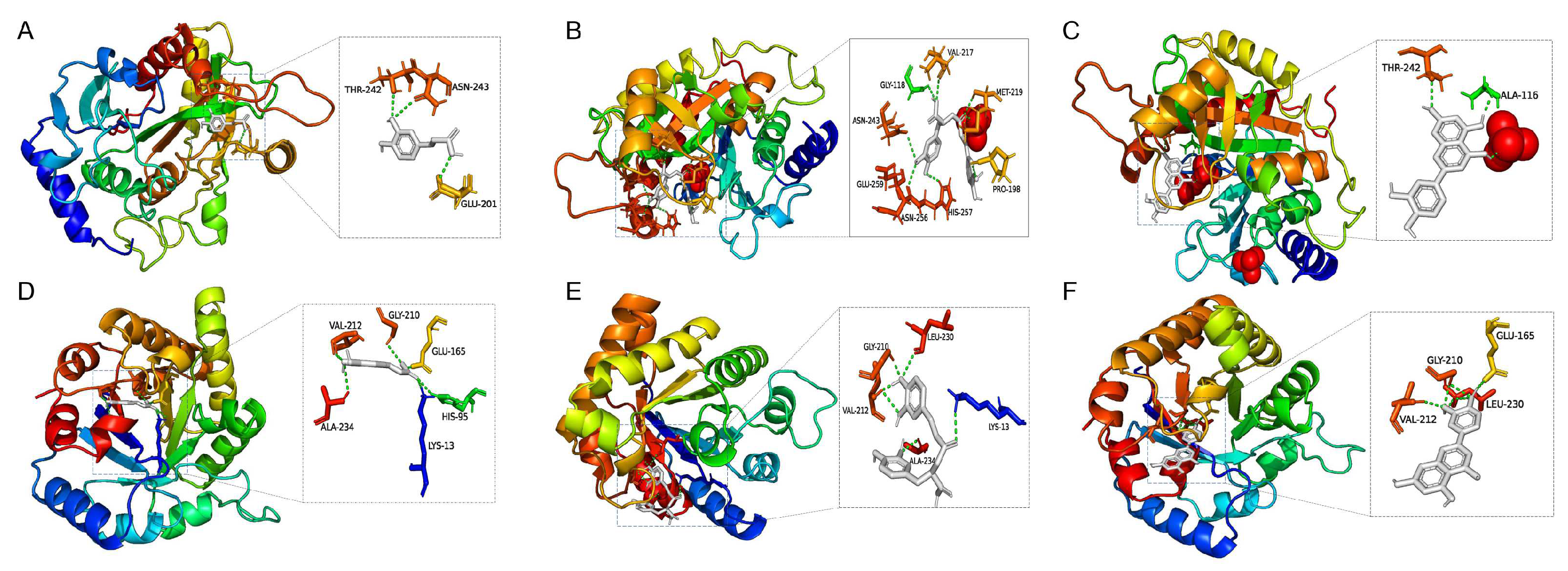

Table 1.
Components identified for AEMH by HPLC-ESI-Q-TOF/MS.
| No. | Retention time (min) | m/z | Molecular formula | ms/ms | Compound name |
| 1 | 3.486 | 191.0199[M-H]- | C6H8O7 | 129 | Citric acid |
| 2 | 4.459 | 188.0563[M-H]- | C7H11NO5 | 128;146 | N-Acetyl-DL-glutamic acid |
| 3 | 5.71 | 117.0191[M-H]- | C4H6O4 | - | Succinic acid |
| 4 | 7.332 | 344.0389[M-H2O-H]- | C10H14N5O8P | 150; 211 | 5'-Guanylic acid |
| 5 | 7.749 | 144.0659[M-H]- | C6H11NO3 | 126 | 4-Acetamidobutyric acid |
| 6 | 14.421 | 329.0517[M-H]- | C13H14O10 | 153 | 3,4-Dihydroxybenzoic acid 3-O-β-D-Glucuronide |
| 7 | 17.063 | 153.0194[M-H]- | C7H6O4 | 109 | 3,4-Dihydroxybenzoic acid |
| 8 | 17.897 | 295.0464[M-H]- | C13H12O8 | 163 | Caffeoyimalic acid |
| 9 | 25.403 | 137.0237[M-H]- | C7H6O3 | - | Salicylic acid /4-Hydroxybenzoic acid |
| 10 | 26.562 | 549.2166[M+Hac-H]- | C22H34O12 | 327 | Cymorcin diglucoside |
| 11 | 28.462 | 137.0237[M-H]- | C7H6O3 | - | 4-Hydroxybenzoic acid /Salicylic acid |
| 12 | 40 | 179.0347[M-H]- | C9H8O4 | 135 | Caffeic acid |
| 13 | 40.463 | 387.1647[M+Hac-H]- | C16H24O7 | 165 | Cymorcin monoglucoside |
| 14 | 43.661 | 197.0447[M-H]- | C9H10O5 | 179;135 | Syringic Acid |
| 15 | 45.838 | 395.0931[M+H-H2O]- | C17H20N2O8S | 219 | Zileuton O-glucuronide |
| 16 | 47.368 | 517.2257[M-H]- | C24H38O12 | 385;161;205 | Cinnamoside |
| 17 | 48.619 | 387.1649[M-H]- | C18H28O9 | - | - |
| 18 | 50.148 | 239.092[M-H]- | C12H16O5 | 151;177;195 | 3-Carboxy-4-methyl-5-propyl-2-furanpropionic acid |
| 19 | 51.306 | 225.1129[M-H]- | C12H18O4 | 147 | Tuberonic acid |
| 20 | 53.809 | 593.1510[M-H]- | C27H30O15 | 353;503 | Luteolin 7-O-rutinoside |
| 21 | 62.659 | 461.0708[M-H]- | C21H18O12 | 285 | Luteolin-3'-D-glucuronide/Luteolin-7-O-glucuronide |
| 22 | 63.54 | 461.0706[M-H]- | C21H18O12 | 285 | Luteolin-3'-D-glucuronide/Luteolin-7-O-glucuronide |
| 23 | 64.744 | 447.0914[M-H]- | C21H20O11 | 285 | Luteolin-7-O-glucoside |
| 24 | 68.451 | 359.0756[M-H]- | C18H16O8 | 197 | Rosmarinic acid |
| 25 | 69.1 | 445.0755[M-H]- | C21H18O11 | 175; 269 | Apigenin-7-O-glucuronide |
| 26 | 86.106 | 285.0393[M-H]- | C15H10O6 | 267 | Luteolin |
| 27 | 93.427 | 285.0748[M+H]+ | C16H12O5 | 270 | 5,6-Dihydroxy-7-methoxyflavone |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



