
Submitted:
11 December 2024
Posted:
12 December 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Coronavirus (CoV) causes intestinal and respiratory diseases in birds, mammals, and humans. In dogs, CoV occurs in two main serotypes, namely, type I and type II, based on mutations and recombination in the spike protein. The pantropical strains are characterized as hypervirulent and multisystemic infections in dogs. Recent outbreaks of coronavirus infection among humans have attracted interest in animal CoV as a source of direct and indirect zoonoses. A literature search was conducted in research databases and open internet sources to analyze coronaviruses in dogs. A total of 189 sources were analyzed, with 142 published in the last decade. Canine coronavirus (CCoV) has undergone mutations and recombination over the past few decades, resulting in changes in tropism, virulence, and the formation of new divergent genetic lines. CCoV is a common component of the canine infectious respiratory disease complex and enteritis of all breeds and ages. CoV infection in pets raises concerns about spreading and transmitting the virus to people and other animals, posing risks to animal and public health. Understanding canine coronaviruses and genetic information sharing could improve prevention and control strategies for future zoonotic coronaviruses.
Keywords:
coronavirus
; canine coronavirus
; coronavirus in dogs
; coronavirus in animals
; SARS-CoV-2
1. Introduction
Coronaviruses (CoVs) cause intestinal and respiratory diseases in birds and mammals, including humans. In dogs, coronavirus (canine coronavirus, CCoV) occurs in two main serotypes, type I and type II, divided based on random point mutations and recombination associated with the spike (S) protein and different serological properties [1,2,3].
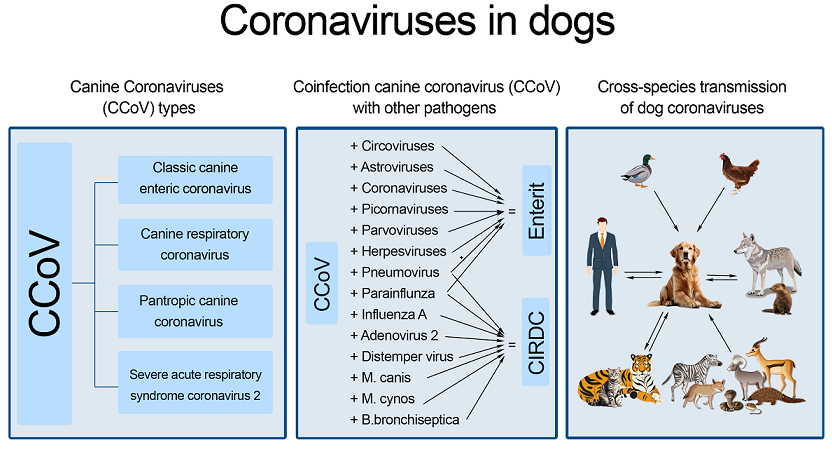
The evolution of CCoV is a clear example of the development of the viral genome through the accumulation of point mutations, insertions, or deletions, manifested by the formation of new genotypes, biotypes, and host variants [3]. Among the various types of CCoV, the latest to be discovered are pantropic strains, characterized by hypervirulent and multisystem infections with a severe course up to a fatal outcome, in contrast to classic enteric coronavirus infections [4,5]. CoVs have been recorded in dogs as an enteric infection (canine enteric coronavirus), a respiratory infection (canine respiratory coronavirus), a multisystem infection (pantropic canine coronavirus), and SARS-CoV-2. CCoV has also been recorded in foxes, wolves, raccoon dogs, and other wild animals [6,7].
The outbreaks of CoVs in humans in recent years (severe acute respiratory syndrome in 2002 (SARS), Middle East respiratory syndrome in 2012 (MERS), and the SARS-CoV-2 pandemic in 2019 (COVID-19) have renewed scientists' interest in animal CoVs as a source of direct and indirect zoonoses [8]. The zoonotic potential of CoVs was confirmed by the zoonotic origin of known outbreaks, as bats and civets acted as sources of pathogens of human coronavirus infections for SARS-CoV and camels for MERS-CoV [9]. The ability of CoVs to overcome interspecies barriers and the zoonotic nature of SARS-CoV-2 allow for the possibility of its emergence in populations of various animals, including pets, which requires further study [10,11,12].
2. Materials and Methods
A literature review was conducted to collect, analyze, and summarize the available scientific data on coronaviruses in dogs.
The data search was conducted in the PubMed, Scopus, MDPI, Elsevier, and CyberLeninka databases and open internet sources without publication date restrictions. The keywords used for the search were "coronavirus", "canine coronavirus", "canine infectious respiratory disease complex", "canine viral enteritis", "canine respiratory coronavirus", "canine pantropic coronavirus", "SARS-CoV-2", and "SARS-CoV-2 in dogs". The received links were checked for compliance with the topic of the article and links containing the full text of the articles or an informative abstract in English or Russian were selected for further work. As a result, 189 sources were analyzed in this review. The number of articles published up to and including 2000 were five, with the earliest article dating back to 1974, and 142 articles included in the work were published in the last 10 years.
Data were obtained from studies conducted in Asia, Europe, America, Africa, New Zealand, and worldwide.
3. Coronaviruses
Coronaviruses are enveloped viruses of the order Nidovirales, family Coronaviridae, and subfamily Orthocoronavirinae. The subfamily Orthocoronavirinae in the family Coronaviridae consists of four genera: Alphacoronavirus, Betacoronavirus, Gammacoronavirus, and Deltacoronavirus [13,14].
Coronaviruses were first identified in dogs in 1971, and, since then, a large number of different strains have been described. CCoV strains are characterized by the presence of significant antigenic differences, on the basis of which they are usually divided into low-virulence strains (mainly type I strains: CCoV-I, causing enteritis) and highly virulent strains (type II: CCoV-II, which are typical strains that cause multisystem diseases in a small percentage of animals) [15]. Low-virulence CCoV strains and feline coronavirus strains are genetically more closely related to each other than to typical CCoV strains. CCoV strains of type I and II are often detected simultaneously in the same dog with diarrhea (36.9–76.8%), which leads to genetic recombination and the emergence of new CCoV variants, contributing to the increased severity of the infection [15,16]. In turn, CCoV-II is divided into two subtypes: CCoV-IIa (highly virulent pantropic CCoV (for example, strain CB/05)) and CCoV-IIb (pathogenic viruses capable of spreading to other organs, more closely related to the porcine transmissible gastroenteritis virus (TGEV)) [17]. In addition, a subtype of CCoV-IIc was distinguished, which is a product of recombination between sequences of CCoV types I and II (prototype strain A76) [15,17].
CoVs are spherical in shape (125 nm in diameter) and have club-shaped spikes on their surface, creating the appearance of the solar corona [18]. The coronavirus genome contains five major open reading frames (ORFs) flanked by 5′ and 3′ untranslated regions. The CoV genome is the largest known RNA virus and is represented by non-segmented RNA with positive polarity. It contains five ORFs flanked by 5′ and 3′ untranslated regions, and replicates in the cytoplasm without integration into the cell genome. The genes encode four major structural proteins: spike (S), membrane (M), envelope (E), and nucleocapsid (N). Some betacoronaviruses have a hemagglutinin esterase protein closely related to the protein of the same name in the influenza C virus. In addition to the main ones, the virus genome encodes a set of additional proteins that can vary significantly among different coronaviruses. The functions of the additional proteins are still poorly understood. These proteins are not essential for virus replication, but they play an important role in virus–host interactions, since they are usually preserved during natural infection, and mutations lead to a decrease in virulence [19]. Genotypic variability due to point mutations and gene recombination results in the emergence of new viral genotypes with different properties or host tropisms [20,21].
The introduction of the virus into the cell is initiated by the interaction between the S protein and its specific receptor on the surface of the cell membrane. Accordingly, the S protein largely determines the tropism of the virus to tissues and the range of hosts, and it also initiates the immune response of the host. After the fusion of the viral and cellular membranes, through the cleavage of the S protein by the protease enzyme, the contents of the virion enter the cytoplasm of the host cell. Then, the viral replicase is synthesized on the RNA matrix of the virion. Newly replicated viral RNA forms a complex with the N protein, and it is then coated with a shell to form a mature virion. After assembly, the virus particles are released from the cell via exocytosis [22].
4. Enteric Coronavirus in Dogs
Enteric CCoV belongs to the Alphacoronavirus genus. CCoV (enteritis-causing strains, or CCoV type I) is related to feline coronaviruses type I and CCoV type II [3]. The CCoV strains studied are branched strains of the CCoV genotype at different distances. Moreover, the degree of similarity of genomes is often not determined by geography, which suggests the genetic recombination and interspecies transmission of CCoV [25].
CCoV was first described during an outbreak in Germany in 1971 [2]; subsequently, it spread rapidly and is now enzootic with prevalence ranging from 6.25% to 72.5% [26,27]. CCoV can be detected in fecal samples of clinically healthy dogs and its serological prevalence is high [28,29]. CCoV was initially identified as a mild enteritis and was long ignored, and vaccination is still not recommended due to the lack of an effective infection model. This virus is a common component of the canine infectious respiratory disease complex (CIRDC) and enteritis in dogs of all breeds and ages, especially in kennels and shelters. Severe and fatal CCoV monoinfections are rare [3,28,30].
5. Canine Respiratory Coronavirus
Canine respiratory coronavirus (CRCoV) is a lineage A betacoronavirus more closely related to bovine coronavirus (BCoV) than to enteric CCoV [31,32,33,34]. This type of CoV was first detected in the UK in 2003 in shelter dogs with symptoms of CIRDC, and it was later identified in dogs worldwide [34,35].
Dogs infected with CRCoV may exhibit varying degrees of clinical signs of respiratory disease, ranging from severe to none [32]. Dogs experimentally infected with CRCoV exhibited signs of mild respiratory disease, with virus dissemination to most respiratory tissues and lymphoid tissues associated with respiratory disease, such as tonsils and local lymphatic tissue. Histopathological changes in lymphoid tissue included damage to or loss of respiratory cilia [36]. Following experimental infection, the virus was detectable for up to 11 days [37]. Shelter dogs have been shown to be at higher risk of acquiring CRCoV [38].
Although CRCoV is an important global respiratory pathogen, its complete genome sequences have eluded retrieval for a long time, with the first complete genome of the South Korean strain K37 being described, showing high similarity to human coronavirus OC43 (HCoV-OC43) and BCoV [39]. The high similarity of CRCoV to BCoV likely indicates descent from BCoV or, alternatively, a common ancestor [20,35].
The epidemiology of CRCoV is currently poorly understood, and knowledge on its global prevalence, pathology, and genetic characteristics is incomplete. The prevalence of CRCoV has been studied using serological methods since the 2000s. It was found that the prevalence of serum antibodies to CRCoV in dog populations in different countries of Europe and North America ranges from 30.1% to 54.7%, with cross-reactivity between CRCoV antibodies from different countries and the virus isolated from the UK, suggesting immunological similarity [31,34,40,41]. ELISA studies of dog sera in Italy in 2006-2007 confirmed its antigenic relationship with BCoV [34]. More than half of the serologically tested dogs in New Zealand showed a positive reaction to CRCoV [42]. In Canada in 2016, CRCoV was detected in 9.4% of dogs with CIRDC using PCR [43]. In New Zealand in 2014-2016, CRCoV was detected in 1 out of 116 dogs tested with CIRDC symptoms (1/56, 2%) [44].
In Sweden, the sequencing of 13 CRCoV isolates in 2013-2015 showed a low level of diversity of CRCoV endemic to Sweden [45]. In contrast, in Thailand, the molecular biological analysis of 21 CRCoV gene sequences from dogs collected in 2013-2015 and 2021-2022 showed variability in the genomic characteristics of Thai CRCoV. The phylogenetic analysis of the S genes allowed us to divide the studied CRCoV strains into five clades. The 2021-2022 strains exhibit characteristic non-synonymous mutations in the S gene that are absent in the 2013-2015 strains, indicating an ongoing evolutionary process of Thai CRCoV. Evolutionary analysis showed that CRCoV in Thailand shares a common ancestor with CRCoV strains from Korea [46].
In the United States in 2024, the serologic prevalence of CRCoV was retrospectively estimated at 23.7%, with the PCR screening of nasal swabs finding 14% of dogs positive for CRCoV. The phylogenetic analysis of the genomes allowed them to be grouped into two distinct groups, with most isolates more closely related to CRCoV from Sweden and only one more closely related to CRCoV from Asia [47].
Although CRCoV is one of the main causative agents of CIRDC and its prevalence in different countries demonstrates its international importance, its epidemiology is still poorly understood. A more complete study of the genomic characteristics and evolutionary patterns of this virus is also needed.
6. Pantropic Canine Coronavirus
Canine pantropic coronavirus (pCCoV), a variant of enteric CCoV, was first isolated in Italy in 2005 from dogs with systemic fatal disease [4]. In contrast to cases with purely enteric tropism, the virus with systemic manifestations is called canine pantropic coronavirus. The disease that it causes can manifest with severe clinical signs and lesions in various organs, including the intestine, liver and spleen, lymph nodes, kidneys, brain, and even the cornea of the eye [48,49]. Histopathological examination has shown multiple severe lung and bronchial lesions that can be observed postmortem in puppies, which may indicate a role for this virus in the pathogenesis of CIRDC [50]. In experimental infections in dogs, the severity of the disease has been found to vary depending on the age and immune status of the animals and may lead to lymphopenia over time [51,52,53]. Later, new outbreaks were reported worldwide, and coinfection with canine parvovirus was also reported [26,54,55,56,57,58,59,60,61]. In addition, other members of the canine family (Canidae) are also known to be susceptible to pCCoV infection [60].
The prototype isolate pCCoV CB/05 belongs to CCoV-IIa and has a high degree of similarity to FCoV type II 79-1683 in the S gene [4]. The complete genome sequence of the prototype strain CB/05 showed that this virus is genetically similar to CCoV-IIa viruses but differs in the presence of an ORF3b deletion [62]. Despite the fact that the CB/05 isolate is recognized as the prototype, a large number of pCCoV outbreaks are caused by viruses that differ significantly, both genetically and antigenically. Thus, it was found that out of 24 pCCoV strains collected throughout Europe, none contained an ORF3b deletion, although they were also CCoV-IIa strains. Moreover, no common genetic marker or specific pathogenicity marker was found for any of the obtained sequences [63]. In Brazil in 2014, pCCoV type IIa was also detected in dogs. The phylogenetic analysis of Brazilian isolates showed their genetic similarity to strains found in other countries [59].
In Italy in 2014-2017, a phylogenetic analysis of S gene sequences of 15 pCCoV strains showed that 3 strains were more closely related to alphacoronaviruses circulating in Asia than to European strains [64].
A large European surveillance study of 354 fatal cases identified pCCoV in 33 cases. Of these, 24 were CCoV type IIa and 9 were CCoV IIb, indicating that systemic infection is not limited to CCoV type IIa [63]. CCoV type IIb viruses, which are more closely related to TGEV, may also be highly pathogenic and pantropic [3,16,55,65,66,67,68].
In addition to the outbreaks of pCCoV subtypes IIa and IIb, the first case of fatal pCCoV infection caused by CCoV-I was reported in France in 2008 [55].
Thus, pCCoV is a polyphyletic group of CoVs, united by tropism and clinical manifestations. In this regard, to date, no consistent genetic marker common to pCCoV has been identified [69]. The accurate test for pCCoV infection is the detection of CCoV in extraintestinal tissues [70].
The discovery of TGEV-like pCCoV in 2005 and again in 2008 in Italy confirmed the observation that CCoV can cause more severe infections than the classical enteric forms [65]. This should be an alarming signal about the pathogenetic evolutionary potential of CCoV. The discovery of virulent CCoV strains associated with the ability to disseminate in different internal organs strongly supports the ability of CoV to change its virulence and organ tropism, which indicates the importance of investigating the molecular basis of these mechanisms [4]. The genetic differences observed in the S protein between classical enteric strains and recombinant CCoV may have special implications for vaccine efficacy. Currently available inactivated vaccines based on CCoV type IIa reference strains were ineffective against challenge with pantropic CCoV IIa [52,65], but the clinical signs were milder in vaccinated dogs. Close monitoring of these viruses is necessary to better understand the clinical, pathogenetic, and epidemiological implications of different CCoV genotypes. Due to the relatively high mutation rate of RNA viruses, CCoV is able to rapidly adapt to the host immune response. Recombination events affecting CCoV could shed light on the evolutionary processes leading to the emergence of new strains, as observed for SARS-CoV-1 and SARS-CoV-II, as well as for CCoV-IIa (CB/05) and CCoV-IIb (TGEV-like CCoV). Many aspects of the pathobiology and evolutionary features of CoVs remain to be elucidated, including the significance of co-infection with CCoV-I and CCoV-II, their pathogenetic role, immune response, and the efficacy of currently used vaccines in the development of CCoV prophylaxis and therapy [5,24,71].
Veterinarians should be informed about the prevalence of pantropic CCoV infection. Moreover, increased data collection will help improve surveillance systems in breeders, veterinary clinics, and animal shelters.
7. Severe Acute Respiratory Syndrome Coronavirus 2
Coronavirus disease 2019 (COVID-19), which was discovered in China, has spread very quickly around the world. On February 29, 2020, the World Health Organization (WHO) classified it as a high-risk disease, and on March 11, 2020, the virus was declared as pandemic [72,73].
The human membrane protein angiotensin-converting enzyme 2 (ACE2) is a receptor for the SARS-CoV-2 S-binding glycoprotein [74]. Two research groups in China and the United States experimentally confirmed this independently in 2020 [75,76,77]. SARS-CoV-2 has incorporated a furin proteolytic site into its S protein that facilitates virus activation in any tissue, making this CoV strain highly polytropic and pathogenic [78]. ACE2 is expressed in many tissues and organs, on the membranes of different cells: type II pneumocytes, enterocytes in the small intestine, endothelial cells of arteries and veins, and smooth muscle cells in most organs. In addition, ACE2 has been found in the membranes of neurons in different parts of the brain [79], which makes these cells susceptible to infection by SARS-CoV-2. This explains the loss of smell and the development of neurological deficits observed in COVID-19 [80].
The unprecedented spread of SARS-CoV-2 worldwide has resulted in the emergence of multiple SARS-CoV-2 variants that have changed transmissibility, infectivity, or immune evasion, causing disease in a wide range of animals in addition to humans [81]. More than 675 SARS-CoV-2 outbreaks in animals have been reported worldwide, including more than 2,000 natural infections [82]. Constant human–animal interaction can lead to new variants [83,84]. A comparative analysis of proteins homologous to ACE2 showed high homology and complete conservation of five amino acid residues: 353-KGDFR-357 of humans, dogs, and other animals, with the exception of snakes. It was shown that SARS-CoV-2 could not be specifically adapted to any of the putative intermediate hosts [85]. Potential risks of reciprocal infections associated with the transmission of SARS-CoV-2 from animals to humans remain [86].
The limited susceptibility of dogs to SARS-CoV-2 is likely due to relatively low levels of ACE2 in their airways and differences in dog and human ACE2 [87], resulting in dogs rarely spreading the disease [88,89]. However, ACE2 undergoes many amino acid changes, which may result in a lower species barrier. Since approximately 470 million pet dogs worldwide live with their human owners, the detection of natural SARS-CoV-2 infection in these animals has important implications for potential zoonotic transmission. Worldwide surveillance efforts to assess the prevalence of SARS-CoV-2 in animals and the intra- and interspecies transmission of CoVs are needed to identify potential reservoir sources of the virus, as well as its evolution and transmission dynamics [90,91,92]. A number of studies conducted in Europe, Asia, America, and Africa have shown the prevalence of SARS-CoV-2 among dogs.
For example, in Germany and Italy, seven veterinary clinics in areas affected by COVID-19 were tested for SARS-CoV-2 early in the pandemic (March–July 2020). Of 877 dog samples tested using real-time PCR, only one dog tested positive. Even in areas where outbreaks of the pandemic were reported, the prevalence of infection in animals admitted to veterinary clinics was low. However, subsequent studies of household animals found higher levels of SARS-CoV-2 infection [93]. According to other data, in Germany, the seroprevalence of SARS-CoV-2 in dogs was 56.8% [94]. In Italy, 2.3-2.7% of dog blood sera showed positivity for SARS-CoV-2 in 2021 and 2022. The seroprevalence was higher among animals living in close contact with SARS-CoV-2-positive owners [95,96].
Switzerland and the Netherlands also demonstrated a higher prevalence of SARS-CoV-2 infection in animals that were in close contact with SARS-CoV-2-positive humans [97,98].
When assessing SARS-CoV-2 seropositivity in dogs during the COVID-19 pandemic in Serbia, the relative incidence of infection was 1.45%. The relative incidence of infection among pet owners was 11%. Seropositivity among pets in contact with COVID-19-infected owners was 9%. High seropositivity was found among stray animals. Of the five tested, two animals were positive [99].
Low seropositivity for SARS-CoV-2 has been established among domestic animals in Croatia. Only 0.31% of dogs were confirmed to have neutralizing antibodies. In an ELISA, 7.56% of the dog sera tested were reactive. Low titers of neutralizing antibodies and a small number of seropositive animals indicate that infections are rare and occur following infections in the human population. In addition, there appears to be no occupational risk for practicing veterinarians when in contact with animals [100].
In Poland, 343 dogs were tested for SARS-CoV-2-specific antibodies and/or sera. The seroprevalence was 1.17% (95% CI 0.45–2.96), respectively (p ≥ 0.05). No significant gender or age differences in seroprevalence were found in dogs (p ≥ 0.05). None of the animals with SARS-CoV-2 antibodies showed respiratory or gastrointestinal symptoms at the time of sampling. The results confirmed previous findings that SARS-CoV-2 infections in pets are uncommon [11].
In Omsk, Russia, nasal swabs were collected from companion animals kept by owners infected with COVID-19. In a study of 90 blood serum samples, 40 were positive. It was found that cats were more likely to become sick (62.96%) than dogs (35.94%). Mongrel animals were less susceptible to infection (42.5%) than purebred animals (57.5%), and the number of females seropositive to coronavirus exceeded the number of males. Animals over 10 years old were most susceptible, and those aged 3 to 5 years were least susceptible [101].
During the COVID-19 pandemic in Japan, 8 (15.1%) of the 53 dogs studied tested positive for SARS-CoV-2. A virus neutralization test also showed antibodies in six dogs. The virus was often transmitted from infected owners to their pets, which then shed the virus despite mild or no clinical signs [102].
Blood samples from 180 dogs were collected in Lebanon during the COVID-19 pandemic from 12 veterinary clinics. The seroprevalence was 19.44% (35/180) in dogs [103].
The low prevalence of SARS-CoV-2 in pets has been confirmed in two of Asia's most populous cities (Hong Kong and Seoul), where contact between people and their pets is very close. The ELISA positivity rate in dog samples was 4.94% for Seoul and 6.46% for Hong Kong [104]. In 2020, 2 of 15 dogs in Hong Kong that had contact with confirmed human cases of COVID-19 were infected with SARS-CoV-2. The genetic sequences of the viruses from two dogs were identical to the virus found in corresponding human cases, indicating human-to-animal transmission. However, it is not yet clear whether dogs can transmit the virus to other animals and back to humans [105]. Final test report from the University of Hong Kong on seroconversion confirms antibody production in one dog following mild infection and real disease due to human-animal transmission [106,107].
Fatal cases of natural SARS-CoV-2 infection in dogs were reported in Thailand [108].
In Korea in 2022, 16 dogs (2.38%) tested positive in ELISA and 14 dogs (2.08%) tested positive in plaque reduction neutralization tests. SARS-CoV-2 antigens and/or antibodies were detected in animals regardless of whether their household was infected; this occurred even in animal shelters that were considered relatively safe from transmission. This indicates continuous circulation of the virus between humans and animals [109].
In household samples from dogs and people with COVID-19 in Ontario, Canada, 41% of serum samples were seropositive, and 2% (1/49) of swabs were PCR-positive. In shelter samples, the seropositivity was 9.3% (7/75) and in veterinary clinics, 16% (35/221). The data suggest that there is a significant risk of infection in pets living with COVID-19 carriers [110].
In Idaho, USA, 119 dogs in households where humans had >1 confirmed SARS-CoV-2 infection were examined. Clinical signs consistent with SARS-CoV-2 were reported in 20 dogs (21%). Of the 81 dogs sampled, 40% were seropositive and 5% were PCR-positive. This discrepancy may be due to delays in sampling [111].
Similar results were obtained during the second wave of COVID-19 in Mexico. In several veterinary clinics in the city of Pueblo, RNA was detected in swabs of four dogs from households infected with COVID-19, and 31.82% were seropositive in ELISA [112]. In another study in Mexico, between February and November 2022, positive results for SARS-CoV-2 infection were detected. Although SARS-CoV-2 infection in dogs is uncommon, these results suggest that transmission from owners to dogs occurs [113].
In Rio de Janeiro, Brazil, in a study of 29 dogs, 31% were found to be infected or seropositive for SARS-CoV-2 and 1 dog developed virus-neutralizing antibodies. Moreover, 6 of 13 dogs developed mild signs of the disease. This indicates the presence and persistence of SARS-CoV-2 infection in dogs from households where human cases of COVID-19 have been reported [114].
Of the 86 dogs tested in Minas Gerais, Brazil, only 1 tested positive for SARS-CoV-2. Among dogs tested in Almenara, SARS-CoV-2 had a low prevalence (1.16%) compared to 15.62% for CCoV. However, younger animals were 2.375 times more likely to be infected. Similarly, purebred animals were more likely to become infected [115].
In Brazil, 11 out of 100 dogs (11.00%) were positive for SARS-CoV-2. A genome comparison showed similarities between animal and human variants that were circulating at the time of sampling, thus confirming the existence of zoonotic transmission [116].
In domestic dogs in Guayaquil, Ecuador's most populous city, 26.2% of dogs tested positive for SARS-CoV-2 in whole-genome sequencing [117].
The overall prevalence of IgG to SARS-CoV-2 in domestic dogs in Villavicencio, Colombia, in November 2020 was 3.67% [118,119].
From March to September 2022, a large molecular biology study of Namibian dogs living in urban and rural areas showed a low but non-negligible prevalence of SARS-CoV-2 (1%). In only one case was a link between the owner and the animal confirmed. In the other cases, no evidence of human infection could be obtained [120].
In contrast to humans, descriptions of animal infection with different SARS-CoV-2 lineages are still rare. Three dogs in COVID-19-positive households in Switzerland were RT-qPCR and/or serologically positive for the SARS-CoV-2 Delta variant. NGS data identified SNPs that were found at higher frequencies in pet viral sequences than in human viral sequences, as well as SNPs that were found exclusively in animals and not in their owners [121]. None of the dogs tested were positive for SARS-CoV-2 during the circulation of new SARS-CoV-2 variants (Delta and Omicron) in Iran [122]. In Chile, the first domestic animal infection with the Omicron BA.4.1 strain was reported in a comprehensive household COVID-19 study. The positive result for SARS-CoV-2 was found in 6.06% (three dogs). In a phylogenetic analysis, the sequences of two SARS-CoV-2 genomes from animals clustered with human sequences collected in Chile during the same period when the BA.4.1 variant was circulating in the country indicated that the virus was transmitted from humans to domestic animals [123].
In a study of the prevalence of SARS-CoV-2 in 2021-2022 among domestic and zoo animals in the south of France, covering submicron and omicron waves, antibodies to the virus were detected not only in domestic dogs and cats, but also in some zoo mammals [124].
There is currently no consensus that infected pets pose a risk of infecting people or other pets. A number of studies of pets for SARS-CoV-2 have given completely negative results [125,126,127,128]. However, suspected cases of human SARS-CoV-2 infection from infected animals that had previous contact with infected people have been identified in Thailand [129].
A study of the susceptibility to SARS-CoV-2 was conducted on animals that interacted closely with humans. Ferrets and cats were found to be susceptible to infection, but SARS-CoV-2 did not replicate well in dogs, pigs, chickens, and ducks [130,131]. The origin, reservoir, and intermediate host of SARS-CoV-2 remain unknown, although related viruses have been found in bats [132].
In summary, the risk to public health and the rate of transmission from infected dogs back to humans are considered low; however, it is critical to investigate dogs as potential hosts for the spread of SARS-CoV-2, with the possibility of establishing natural infection or adaptation in animals prior to back-spread to humans, associated with the genetic evolution of the virus [133,134]. For example, probable transmission of SARS-CoV-2 variants from hamsters to humans, AY.127 and B.1.258, has been demonstrated in pet-store-associated COVID-19 outbreaks [135,136]. Similarly, the latest variant of concern, SARS-CoV-2 Omicron (B.1.1.529), may have evolved in mice before spreading to humans [137]. Thus, monitoring SARS-CoV-2 transmission in animals is important to understand how viruses evolve in animals, whether new variants arise in animals, and whether new variants pose any risk to public health and prevention and control strategies based on One Health approaches [81,84,138,139].
Epidemiological studies and case reports indicate that dogs become infected with SARS-CoV-2 naturally or experimentally; however, the global number of animals infected naturally is much lower than the number of people with COVID-19. These studies show that pets become infected through direct contact with owners infected with COVID-19. The emergence of SARS-CoV-2 infection in pets worldwide raises concerns that pets are at higher risk of spreading and transmitting SARS-CoV-2 to people and other animals, posing risks to animal health and public health. Thus, studying the role of dogs in the transmission and epidemiology of SARS-CoV-2 will help to develop and implement appropriate preventive measures against further transmission of SARS-CoV-2 [140].
8. Coinfection of CCoV with Other Pathogens
Canine infectious respiratory disease (CIRDC), commonly known as kennel cough, is a syndrome in which multiple viral and bacterial pathogens sequentially or synergistically contribute to the disease. One of the key emerging pathogens of CIRDC is CCoV [141]. Numerous studies have been conducted in kennels and veterinary clinics for CIRDC pathogens in dogs seeking veterinary care, both symptomatic and asymptomatic.
In a large multicenter European study, CIRDC affected 66% (381/572) of the dogs studied, including both pet and kennel-based dogs. The incidence and severity of disease were significantly reduced in dogs vaccinated against the classic CIRDC pathogens, canine distemper virus (CDV), canine adenovirus 2, and canine parainfluenza virus (CPIV), but a significant proportion (65.7%; 201/306) of vaccinated dogs remained affected. CRCoV and canine pneumovirus (CnPnV) were widely distributed in various dog populations, with overall seropositivity and detection rates of 47% and 7.7% for CRCoV and 41.7% and 23.4% for CnPnV, respectively, and their presence was associated with an increased incidence and severity of clinical disease. Dogs were more likely to be seropositive for CnPnV, CRCoV, and Mycoplasma cynos simultaneously, which could indicate their frequent coinfection or cocirculation [142].
A molecular study of CIRDC-associated pathogens conducted in Italy between 2011 and 2013 showed that CPIV was the most frequently identified pathogen in CIRDC, followed by CRCoV and CnPnV. The most severe forms of respiratory infections were observed with CPIV, CRCoV, and M. cynos, either alone or in combination with other pathogens. The study, along with CPIV, highlighted the increasing role of viruses such as CRCoV and CnPnV, for which there are no effective vaccines [143].
In France and Belgium, dogs with systemic CCoV infection were commonly coinfected with parvovirus [63].
As previously reported, CRCoV has been detected in the UK dog population [31]. CRCoV infections occurred predominantly during the first week of a dog's stay in a kennel. In addition, other pathogens, such as canine herpes virus (CHV), may play a role in the pathogenesis of canine respiratory diseases [144].
A study of swabs from kennels in southern and central England found the presence of antibodies to CRCoV, either during seasonal respiratory outbreaks or in dogs on admission, consistent with the presence of antibodies to CHV throughout the year [145].
In Germany, of 37.7% of dogs with acute respiratory symptoms, coinfection with CPIV and CRCoV was observed in 82.6% [146].
In New Zealand, CRCoV, canine adenovirus 2, CPIV, CHV, Bordetella bronchiseptica, and M. cynos were circulating among sampled dogs. In total, 47 of 93 (50%) dogs were seropositive for CRCoV [147].
In Thailand, a study of 209 dogs with respiratory signs using multiplex PCR detected six common viruses associated with CIRDC, with CRCoV and CIV coinfection predominantly found along with other CIRDC viruses. It was also noted that the level of clinical severity was significantly associated with the age of infected dogs rather than vaccination status, sex, and route of transmission [148].
In samples from clinically sick and asymptomatic dogs obtained from the Athens Veterinary Diagnostic Laboratory, CCoV was not among the most frequently detected pathogens, but still constituted a significant proportion (4.6%, 26/559), occurring in different combinations with CPIV (29%, 33/114), M. canis (23.6%, 27/144), M. cynos (24.5%, 28/144), influenza A (11.2%, 63/559), B. bronchiseptica (9%, 51/559), canine adenovirus 2 (2.5%, 14/559), and CDV (2%, 11/559). A statistical analysis of CIRDC pathogens highlighted the impact of coinfections on the severity of clinical manifestations [149].
In Canada in 2007, a study of wolf diseases revealed virus exchange between dogs and wolves. Dogs were found to be exposed to parvovirus, CDV, and CRCoV, indicating the need for vaccination to prevent morbidity and mortality in dogs, as well as the potential transmission of viruses to sympatric wild animals, leading to their extinction or population decline [150].
In China, a metagenomics approach was used to investigate the enteric virome status of 45 healthy domestic dogs with extensive human contact. CCoV was uniquely identified in all samples with high abundance. The top 30 contigs with the highest read counts were mapped to the DNA virus families Circoviridae, Parvoviridae, and Herpesviridae, and the RNA virus families Astroviridae, Coronaviridae, and Picornaviridae [151].
In Georgia, USA, from 2018 to 2022, of 459 cases, viral agents were detected in 34% of cases, of which two or more pathogens were detected in 24% of cases. The percentages of viral agents detected were canine adenovirus 2 (4%), CDV (3%), CPIV (16%), CRCoV (7%), and CIV (2%) [152].
Studies of 20 dogs aged 3-5 years during an outbreak of intestinal infection in the K-9 kennel in Irkutsk showed that, in 18 (90%) animals, the pathology was caused by an association of microorganisms involving two or three species. At the same time, only in 1 (5%) dog were the associations represented by two types of viruses (CoV and parvovirus), and in 17 (85%) by viruses and bacteria. CoV was observed in association with the following agents: parvovirus, Clostridium freundii, E. coli, and Proteus mirabilis [153].
Thus, CCoV is one of the most widespread and important viral components of CIRDC and, when acting together with other pathogens, determines the severity of the disease to a large extent. Therefore, one of the important issues remains the development and distribution of an up-to-date vaccine against CCoV and a set of preventive measures. Current data on the prevalence and role of coinfections with individual viruses and bacteria in the etiology of CIRDC emphasize the importance of molecular diagnostics and vaccination.
9. Cross-Species Transmission of Dog Coronaviruses
Almost 60% of human pathogens and approximately 60% of emerging infectious diseases are zoonoses, and zoonoses are an increasingly serious public health problem as their incidence has increased in recent years [154,155]. In line with the One Health concept, the genetic plasticity of CoVs should raise alarm and prompt the scientific community to conduct continuous genomic surveillance in both wildlife and domestic animals to avoid possible future major health emergencies [21,23,156,157].
In recent years, advances in metagenomics have enabled the discovery and characterization of many native viral particles. Next-generation sequencing and bioinformatics analysis have revealed the enormous phylogenetic diversity of viromes in domestic, farm, and wild animals. For example, the repository of publicly available animal-associated SarbeCoV (subgenus Betacoronavirus) strains is quite extensive, including 1,535 strains identified in 63 animal species in 43 countries (as of February 14, 2023) [158]. The importance of new (SARS/MERS) and re-emerging (COVID-19) infections is currently increasing [159,160,161]. Therefore, it is necessary to constantly study their genetic characteristics to identify the emergence of new types, changes in pathogenesis, and mechanisms of immune escape [5,162,163].
Although the exact public health implications are not yet clear, RNA viruses, including CCoV, may be the most problematic because their genomes are characterized by high mutation rates and the ability to evade the immune system [164]. Coinfection with multiple viruses accelerates viral variation in the host organism [17,27,65,165]. Following the global shock of the COVID-19 pandemic and its zoonotic nature, it has become even more evident that monitoring animal viruses is critical. Timely research and vigilance can potentially save many lives, both animal and human [166,167,168,169].
Known human pandemics have been caused by members of two families of viruses: Orthomyxoviridae (influenza A viruses: subtypes H1N1, H2N2, and H3N2) and Coronaviridae (SARS-CoV-2) [161]. In the genome of these viruses, mutations occurred in the body of the intermediate host animal, which acquired an adaptive nature and contributed to overcoming species barriers and zoonosis. These adaptive mutations contributed to the horizontal transmission of the variant strain in addition to humans to other contact species of animals, including domestic and wild animals [170]. Monitoring viral zoonosis and viral reverse zoonosis of pandemic strains within the framework of the One Health, One World approach helps reduce unexpected risks, such as the emergence and adaptation of new strains to the human host, the drug resistance of pathogens, and unknown preventive and therapeutic measures [171].
Domestic dogs share their habitat with humans, making them a potential source of zoonotic viruses. Cohabitation and daily contact between pets and their owners create favorable conditions for the interspecies transmission of viruses.
In 2015, a recombinant CCoV strain, HLJ-073, was isolated from a dog in China, which is more closely related to FCoV II than to CCoV I or CCoV II strains. Experiments showed that HLJ-073 can replicate efficiently in canine macrophages/monocytes and human THP-1 cells. This strain has, thus, acquired the ability to change tissue tropism from the intestinal tract to systemic infection and to change host tropism from dogs to humans [67].
In 2017–2018, a virus most similar to CCoV was isolated from children with pneumonia in Malaysia. The HuPn-2018 strain, classified as a novel canine–feline recombinant virus, had a unique deletion in the nucleoprotein similar to the deletion found in SARS-CoV-1 and SARS-CoV-2, which occurred very soon after their introduction into humans [172], supposedly moving from dogs to humans. A new canine-origin CoV, HuCCoV_Z19Haiti, related to the Malaysian strain, was detected in a man after a trip to Haiti, suggesting that Malaysian CCoV strains may infect humans [173]. According to phylogenetic studies, the S gene was similar to CCoV-HuPn-2018, while the M gene was closer to CCoV B639_ZI_2019 [26], and the N gene was more similar to TGEV [173]. This strain confirms the high potential for the genetic recombination of these viruses, defining the emergence of new variants that overcome the species barrier and pose a potential threat to humans. These data and the emergence of highly pathogenic CoVs in humans (SARS-CoV-1, MERS-CoV, and SARS-CoV-2) highlight the significant threat that the spread of CCoV poses to humans [24,174].
A study of fecal samples from urban domestic dogs in China revealed the presence of Coronaviridae in the intestine, along with other viruses. A phylogenetic analysis of CCOV samples showed that they were grouped into the Alphacoronavirus genus and were closely related to canine coronaviruses circulating in China, the United States, and Italy, as well as to human coronavirus strains 229E and NL63, indicating a potential risk of human infection by canine coronaviruses through the acquisition of cross-species transmission. The long preservation of CCOV in dog feces (156 days) [3,175] and the high frequency of virus detection in healthy exposed animals raises the risk of infection of other animals and humans [151].
In 2020, Hong Kong reported its first positive result for SAR-CoV-2, where 2 out of 15 dogs from COVID-19-positive households showed decreased neutralizing antibodies, suggesting the human-to-dog transmission of SARS-CoV-2 [105]. Several more countries then confirmed cases of positive SARS-CoV-2 infection in pet dogs living in COVID-19-positive households [176,177]. By May 2023, the United States had recorded over a hundred confirmed cases of various SARS-CoV-2 variants (B.1.1.7, AY.3, AY.43) in dogs [178,179,180,181].
Experimentally infected dogs with SARS-CoV-2 developed low viral loads compared to experimentally infected cats and ferrets, suggesting that dogs are susceptible to SARS-CoV-2 infection but to a lesser extent than cats and ferrets [130].
The first ever case of SARS-CoV-2 transmission from a human host to a domestic cat was confirmed on March 27, 2020. A tiger at a zoo in New York, USA, was also reported to be infected with SARS-CoV-2. In the Netherlands, two employees were infected with SARS-CoV-2 from minks [86].
SARS-CoV-2 was recently transmitted from an infected cat to a veterinarian in Thailand [182], confirming the potential for animal-to-human transmission of the coronavirus.
The challenge associated with emerging zoonoses such as SARS-CoV-2 is to identify their sources and mechanism(s) of origin. Such possible mechanisms of viral adaptation include mutations, genomic recombination, viral host physiology and immune response, ecogeography, and human factors including ACE2 receptor structure and immune function [183,184,185]. SARS-CoV-2 is widespread among animals and can infect a wide range of domestic and wild species. To effectively control the spread of viruses, it is essential to have a clear understanding of the mechanisms of interspecies transmission and the risk factors for zoonotic infections [186,187].
10. Conclusions
Emerging infectious diseases in animals have two major biological consequences: first, many wildlife species serve as reservoirs of pathogens that threaten the health of domestic animals and humans; second, the infections pose a significant threat to the conservation of global biodiversity [150,188].
Coronaviruses are generally zoonotic pathogens with significant interspecies transmission. They are widespread in nature and have recently become a significant component of infectious diseases in many wild and domestic animals, including dogs. The prevalence of coronavirus diseases is a result of the high mutation rate of the virus, the abundance of circulated strains, and the slow pace of vaccine development. To effectively combat such infections, the development of new antiviral strategies is necessary [189].
A better understanding of canine coronaviruses and their ability to transmit across species, as well as the sharing of genetic information, could improve prevention and control strategies for future emerging zoonotic coronaviruses.
Author Contributions
Conceptualization, G.L. and T.G.; formal analysis, N.K., N.S. and R.W.; investigation, G.L.; resources, G.L.; data curation, N.K.; writing—original draft preparation, T.G. and R.W.; writing—review and editing, G.L. and T.G.; supervision, N.K. and R.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Science and Higher Education of the Republic of Kazakhstan, grant number AP19677712.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Parkhe, P.; Verma, S. Evolution, Interspecies Transmission, and Zoonotic Significance of Animal Coronaviruses. Front Vet Sci 2021, Oct 18, 8, 719834. [CrossRef]
- Binn, L.N.; Lazar, E.C.; Keenan, K.P.; Huxsoll, D.L.; Marchwicki, R.H.; Strano, A.J. Recovery and characterization of a coronavirus from military dogs with diarrhea. Proc Annu Meet U S Anim Health Assoc 1974, 78, 359–66. http://surl.li/gfpprn.
- Decaro, N.; Buonavoglia, C. An update on canine coronaviruses: viral evolution and pathobiology. Vet Microbiol 2008, Dec 10, 132(3-4), 221-34. [CrossRef]
- Buonavoglia, C.; Decaro, N.; Martella, V.; Elia, G.; Campolo, M.; Desario, C.; Castagnaro, M.; Tempesta, M. Canine coronavirus highly pathogenic for dogs. Emerg Infect Dis 2006, 12, 492–494. [CrossRef]
- Timurkan, M.O.; Aydin, H.; Dincer, E.; Coskun, N.; Molecular characterization of canine coronaviruses: an enteric and pantropic approach. Arch Virol 2021, Jan,166(1), 35-42. [CrossRef]
- Ma, G.G.; Lu, C.P. Two genotypes of Canine coronavirus simultaneously detected in the fecal samples of healthy foxes and raccoon dogs. Wei Sheng Wu Xue Bao 2005, Apr,45,2,305-8. [Chinese] https://europepmc.org/article/med/15989282.
- Zarnke, R.L.; Evermann, J.; Ver Hoef, J.M.; McNay, M.E.; Boertje. R.D.; Gardner, C.L.; Adams, L.G.; Dale, B.W.; Burch, J. Serologic survey for canine coronavirus in wolves from Alaska. J Wildl Dis 2001, Oct, 37(4), 740-5. [CrossRef]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A Pneumonia Outbreak Associated with a New Coronavirus of Probable Bat Origin. Nature 2020, 579, 270–273. [CrossRef]
- Malik, Y.A. Properties of Coronavirus and SARS-CoV-2. Malays J Pathol 2020, Apr, 42(1), 3-11. https://pubmed.ncbi.nlm.nih.gov/32342926/.
- WHO (2003) Consensus document on the epidemiology of severe acute respiratory syndrome (SARS) available at https://apps.who.int/iris/bitstream/handle/10665/70863/WHO_CDS_CSR_GAR_2... accessed on 11 November 2024.
- Pomorska-Mól, M.; Turlewicz-Podbielska, H.; Gogulski, M.; Ruszkowski, J.J.; Kubiak, M.; Kuriga, A.; Barket, P.; Postrzech M. A cross-sectional retrospective study of SARS-CoV-2 seroprevalence in domestic cats, dogs and rabbits in Poland. BMC Vet Res 2021 Oct 7, 17(1), 322. [CrossRef]
- Subbotina, A.; Semenov, V.M.; Kupriyanov, I.I. Biological and molecular genetic features of SARS-CoV-2. Bulletin of Vitebsk State Medical University 2023, 22(6), 76-82. [in Russ]. [CrossRef]
- Adams, M.J.; Carstens, E.B. Ratification vote on taxonomic proposals to the International Committee on Taxonomy of Viruses. Arch Virol 2012, 157, 1411–1422. [CrossRef]
- Adams, M.J.; Lefkowitz, E.J.; King, A.M.Q.; Bamford, D.H.; Breitbart, M.; Davison, A.J.; Ghabrial, S.A.; Gorbalenya, A.E.; Knowles, N.J.; Krell, P.; et al. Ratification Vote on Taxonomic Proposals to the International Committee on Taxonomy of Viruses. Arch. Virol. 2015, 160, 1837–1850. [CrossRef]
- Nguyen, T.L.; Pham Thi, H.H. Genome-wide comparison of coronaviruses derived from veterinary animals: A canine and feline perspective. Comp Immunol Microbiol Infect Dis 2021, Jun, 76, 101654. [CrossRef]
- Decaro, N.; Mari, V.; Elia, G.; Addie, D.D.; Camero, M.; Lucente, M.S.; Martella, V.; Buonavoglia, C. Recombinant canine coronaviruses in dogs, Europe. Emerg Infect Dis 2010, Jan, 16(1), 41-7. [CrossRef]
- Regan, A.D.; Millet, J.K.; Tse, L.P.V.; Chillag, Z.; Rinaldi, V.D.; Licitra, B.N.; Dubovi, E.J.; Town, C.D.; Whittaker, G.R. Characterization of a recombinant canine coronavirus with a distinct receptor-binding (S1) domain. Virology 2012, 430(2), 90-99. (https://www.sciencedirect.com/science/article/pii/S0042682212002085). [CrossRef]
- Fehr, A.R.; Perlman, S. Coronaviruses: an overview of their replication and pathogenesis. Methods Mol Biol. 2015, 1282, 1–23. [CrossRef]
- Enjuanes, L.; Brian, D.; Cavanagh, D. et al. Family Coronaviridae. In Virus taxonomy, classification and nomenclature of viruses van Regenmortel, M.H.V.; Fauquet, C.M.; Bishop, D.H.L. et al. Eds., Academic Press, New York. 2000, pp. 835–849. http://surl.li/qrkxau.
- Lorusso, A.; Desario, C.; Mari, V.; Campolo, M.; Lorusso, E.; Elia, G.; Martella, V.; Buonavoglia, C.; Decaro, N. Molecular characterization of a canine respiratory coronavirus strain detected in Italy. Virus Res 2009, Apr, 141(1), 96-100. [CrossRef]
- Pratelli, A.; Buonavoglia, A.; Lanave, G.; Tempesta, M.; Camero, M.; Martella, V.; Decaro, N. One world, one health, one virology of the mysterious labyrinth of coronaviruses: the canine coronavirus affair. Lancet Microbe 2021, Dec, 2, 12, e646-e647. [CrossRef]
- Haake, C.; Cook, S.; Pusterla, N.; Murphy, B. Coronavirus Infections in Companion Animals: Virology, Epidemiology, Clinical and Pathologic Features. Viruses 2020, Sep, 13, 12(9), 1023. [CrossRef]
- Pratelli, A.; Tempesta, M.; Elia, G.; Martella, V.; Decaro, N.; Buonavoglia, C. The knotty biology of canine coronavirus: A worrying model of coronaviruses' danger. Res Vet Sci 2022, May, 144, 190-195. [CrossRef]
- Buonavoglia, A.; Pellegrini, F.; Decaro, N.; Galgano, M.; Pratelli, A. A One Health Perspective on Canine Coronavirus: A Wolf in Sheep's Clothing? Microorganisms 2023, Apr,2, 11(4), 921. [CrossRef]
- Hossain, M.E.; Islam, A.; Islam, S.; Rahman, M.K.; Miah, M.; Alam, M.S.; Rahman, M.Z. Detection and Molecular Characterization of Canine Alphacoronavirus in Free-Roaming Dogs, Bangladesh. Viruses 2021, Dec,30, 14(1), 67. [CrossRef]
- He, H.J.; Zhang, W.; Liang, J.; Lu, M.; Wang, R.; Li, G.; He, J.W.; Chen, J.; Chen, J.; Xing, G.; Chen, Y. Etiology and genetic evolution of canine coronavirus circulating in five provinces of China, during 2018-2019. Microb Pathog 2020, Aug, 145, 104209. [CrossRef]
- Wang, X.; Li, C.; Guo, D.; Wang, X.; Wei, S.; Geng, Y.; Wang, E.; Wang, Z.; Zhao, X.; Su, M.; Liu, Q.; Zhang, S.; Feng, L.; Sun, D. Co-Circulation of Canine Coronavirus I and IIa/b with High Prevalence and Genetic Diversity in Heilongjiang Province, Northeast China. PLoS One 2016, Jan, 15, 11(1), e0146975. [CrossRef]
- Tennant, B.J.; Gaskell, R.M.; Jones, R.C.; et al. Studies on the epizootiology of canine coronavirus. Vet Rec 1993, 132, 7–11. [CrossRef]
- Tupler, T.; Levy, J.K.; Sabshin, S.J.; et al. Enteropathogens identified in dogs entering a Florida animal shelter with normal feces or diarrhea. J Am Vet Med Assoc 2012, 241, 338–343. [CrossRef]
- Pratelli, A. The evolutionary processes of canine coronaviruses. Adv Virol 2011, 562831. [CrossRef]
- Erles, K.; Toomey, C.; Brooks, H.W.; Brownlie, J. Detection of a group 2 coronavirus in dogs with canine infectious respiratory disease. Virology 2003, Jun,5, 310(2), 216-23. [CrossRef]
- Erles, K.; Brownlie, J. Canine respiratory coronavirus: an emerging pathogen in the canine infectious respiratory disease complex. Vet Clin North Am Small Anim Pract 2008, Jul, 38(4), 815-25. [CrossRef]
- An, D.J.; Jeong, W.; Yoon, S.H.; Jeoung, H.Y.; Kim, H.J.; Park, B.K. Genetic analysis of canine group 2 coronavirus in Korean dogs. Vet Microbiol 2010, Feb, 24, 141(1-2), 46-52. [CrossRef]
- Decaro, N.; Desario, C.; Elia, G.; Mari, V.; Lucente, M.S.; Cordioli, P.; Colaianni, M.L.; Martella, V.; Buonavoglia, C. Serological and molecular evidence that canine respiratory coronavirus is circulating in Italy. Vet Microbiol 2007, Apr,15, 121(3-4), 225-30. [CrossRef]
- Erles, K.; Shiu, K.B.; Brownlie, J. Isolation and sequence analysis of canine respiratory coronavirus. Virus Res 2007, Mar, 124(1-2), 78-87. [CrossRef]
- Yachi, A.; Mochizuki, M. Survey of dogs in Japan for group 2 canine coronavirus infection. J Clin Microbiol 2006, Jul, 44(7), 2615-8. [CrossRef]
- Kaneshima, T.; Hohdatsu, T.; Hagino, R.; Hosoya, S.; Nojiri, Y.; Murata, M.; Takano, T.; Tanabe, M.; Tsunemitsu, H.; Koyama, H. The infectivity and pathogenicity of a group 2 bovine coronavirus in pups. J Vet Med Sci 2007, Mar, 69(3), 301-3. [CrossRef]
- Okonkowski, L.K.; Szlosek, D.; Ottney, J.; Coyne, M.; Carey, S.A. Asymptomatic carriage of canine infectious respiratory disease complex pathogens among healthy dogs. J Small Anim Pract. 2021, Aug, 62(8), 662-668. [CrossRef]
- Lu, S.; Wang, Y.; Chen, Y.; Wu, B.; Qin, K.; Zhao, J.; Lou, Y.; Tan, W. Discovery of a novel canine respiratory coronavirus support genetic recombination among betacoronavirus1. Virus Res 2017, Jun, 2, 237, 7–13. [CrossRef]
- Priestnall, S.L.; Brownlie, J.; Dubovi, E.J.; Erles, K. Serological prevalence of canine respiratory coronavirus. Vet Microbiol 2006, Jun, 15, 115(1-3), 43-53. [CrossRef]
- Ellis, J.; Anseeuw, E.; Gow, S.; Bryan, H.; Salb, A.; Goji, N.; Rhodes, C.; La Coste S.; Smits, J.; Kutz, S. Seroepidemiology of respiratory (group 2) canine coronavirus, canine parainfluenza virus, and Bordetella bronchiseptica infections in urban dogs in a humane shelter and in rural dogs in small communities. Can Vet J. 2011, Aug, 52, 8, 861–8. https://pubmed.ncbi.nlm.nih.gov/22294792/.
- More, G.D.; Dunowska, M.; Acke, E.; Cave, N.J. A serological survey of canine respiratory coronavirus in New Zealand. N Z Vet J 2020, Jan, 68(1), 54-59. [CrossRef]
- Joffe, D.J.; Lelewski, R.; Weese, J.S.; Mcgill-Worsley, J.; Shankel. C.; Mendonca, S.; Sager, T.; Smith, M.; Poljak, Z. Factors associated with development of Canine Infectious Respiratory Disease Complex (CIRDCCCC) in dogs in 5 Canadian small animal clinics. Can Vet J 2016, Jan, 57(1), 46-51. https://typeset.io/papers/factors-associated-with-development-of-canine-infectious-nbfram7rhq.
- More, G.D.; Cave, N.J.; Biggs, P.J.; Acke, E.; Dunowska, M. A molecular survey of canine respiratory viruses in New Zealand. N Z Vet J 2021, Jul, 69(4), 224-233. [CrossRef]
- Wille, M.; Wensman, J.J.; Larsson, S.; van Damme, R.; Theelke, A.K.; Hayer, J.; Malmberg, M. Evolutionary genetics of canine respiratory coronavirus and recent introduction into Swedish dogs. Infect Genet Evol 2020, Aug, 82, 104290. [CrossRef]
- Poonsin, P.; Wiwatvisawakorn, V.; Chansaenroj, J.; Poovorawan, Y.; Piewbang, C.; Techangamsuwan. S. Canine respiratory coronavirus in Thailand undergoes mutation and evidences a potential putative parent for genetic recombination. Microbiol Spectr. 2023, Sep, 14, 11(5), e0226823. [CrossRef]
- De Luca, E.; Álvarez-Narváez, S.; Baptista, R.P.; Maboni, G.; Blas-Machado, U.; Sanchez, S. Epidemiologic investigation and genetic characterization of canine respiratory coronavirus in the Southeastern United States. J Vet Diagn Invest 2024, Jan, 36(1), 46-55. [CrossRef]
- Decaro, N.; Buonavoglia, C. Canine coronavirus: not only an enteric pathogen. Vet Clin North Am Small Anim Pract 2011, Nov, 41(6), 1121-32. [CrossRef]
- Gndoyan, I.A.; Dvoretskaya, Yu.A.; Solodov, E.V.; Chernov, S.A. Corneal lesions in dogs due to coronavirus infection. Veterinarian 2022, 6, 20–24. [in Russ.]. [CrossRef]
- Zappulli, V.; Caliari, D.; Cavicchioli, L.; et al. Systemic fatal type II coronavirus infection in a dog: pathological findings and immunohistochemistry. Res Vet Sci. 2008, 84, 278–282. [CrossRef]
- Decaro, N.; Campolo, M.; Lorusso, A.; Desario, C.; Mari, V.; Colaianni, M.L.; Elia, G.; Martella, V.; Buonavoglia, C. Experimental infection of dogs with a novel strain of canine coronavirus causing systemic disease and lymphopenia. Vet Microbiol 2008, Apr, 30, 128(3-4), 253-60. [CrossRef]
- Decaro, N.; Elia, G.; Martella, V.; Campolo, M.; Mari, V.; Desario, C.; Lucente, M.S.; Lorusso, E.; Kanellos, T.; Gibbons, R.H.; Buonavoglia, C. Immunity after natural exposure to enteric canine coronavirus does not provide complete protection against infection with the new pantropic CB/05 strain. Vaccine 2010, 28, 724–729. [CrossRef]
- Marinaro, M.; Mari, V.; Bellacicco, A.L.; Tarsitano, E.; Elia, G.; Losurdo, M.; Rezza, G.; Buonavoglia, C.; Decaro, N. Prolonged depletion of circulating CD4 T lymphocytes and acute monocytosis after pantropic canine coronavirus infection in dogs. Virus Res 2010, 152, 73–78. [CrossRef]
- Soma, T.; Ohinata, T.; Ishii, H.; Takahashi, T.; Taharaguchi, S.; Hara, M.; Detection and genotyping of canine coronavirus RNA in diarrheic dogs in Japan. Res Vet Sci 2011, Apr, 90(2), 205-7. [CrossRef]
- Zicola, A.; Jolly, S.; Mathijs, E.; et al. Fatal outbreaks in dogs associated with pantropic canine coronavirus in France and Belgium. J Small Anim Pract 2012, 53, 297–300 . [CrossRef]
- Ntafis, V.; Xylouri, E.; Mari, V.; Papanastassopoulou, M.; Papaioannou, N.; Thomas, A.; Buonavoglia, C.; Decaro, N. Molecular characterization of a canine coronavirus NA/09 strain detected in a dog’s organs. Arch Virol 2012, 157, 171–175. [CrossRef]
- Decaro, N.; Mari, V.; von Reitzenstein, M.; Lucente, M.S.; Cirone, F.; Elia, G.; Martella, V.; King, V.L.; Di Bello, A.; Varello, K.; Zhang, S.; Caramelli, M.; Buonavoglia, C. A pantropic canine coronavirus genetically related to the prototype isolate CB/05. Vet Microbiol 2012, 159, 239–244. [CrossRef]
- Ntafis, V.; Mari, V.; Decaro, N.; Papanastassopoulou, M.; Pardali, D.; Rallis, T.S.; Kanellos, T.; Buonavoglia, C.; Xylouri, E. Canine coronavirus, Greece. Molecular analysis and genetic diversity characterization. Infect Genet Evol 2013, Jun, 16, 129-36. [CrossRef]
- Pinto, L.D.; Barros, I.N.; Budaszewski, R.F.; Weber, M.N.; Mata, H.; Antunes, J.R.; Canal, C.W. Characterization of pantropic canine coronavirus from Brazil. Vet J 2014, 202(3), 659–662. [CrossRef]
- Alfano, F.; Dowgier, G.; Valentino, M.P.; Galiero, G.; Tinelli, A.; Decaro, N.; Fusco, G. Identification of pantropic canine coronavirus in a wolf (Canis lupus italicus) in Italy. J Wildl Dis 2019, 55, 504–508. [CrossRef]
- Zobba, R.; Visco, S.; Sotgiu, F.; Pinna Parpaglia, M.L.; Pittau, M.; Alberti, A. Molecular survey of parvovirus, astrovirus, coronavirus, and calicivirus in symptomatic dogs. Vet Res Commun. 2021, Feb, 45(1), 31-40. [CrossRef]
- Decaro, N.; Mari, V.; Dowgier, G.; Elia, G.; Lanave, G.; Colaianni, M.L.; Buonavoglia, C. Full-genome sequence of pantropic canine coronavirus. Genome Announc 2015, May, 7, 3(3), e00401-15. [CrossRef]
- Decaro, N.; Cordonnier, N.; Demeter, Z.; Egberink, H.; Elia, G.; Grellet, A.; Le Poder, S.; Mari, V.; Martella, V.; Ntafis, V.; von Reitzenstein, M.; Rottier, P.J.; Rusvai, M.; Shields, S.; Xylouri, E.; Xu, Z.; Buonavoglia, C. European surveillance for pantropic canine coronavirus. J Clin Microbiol 2013, Jan, 51(1), 83-8. [CrossRef]
- Alfano, F.; Fusco, G.; Mari, V.; Occhiogrosso, L.; Miletti, G.; Brunetti, R.; Galiero, G.; Desario, C.; Cirilli, M.; Decaro, N. Circulation of pantropic canine coronavirus in autochthonous and imported dogs, Italy. Transbound Emerg Dis 2020, Sep., 67(5), 1991-1999. [CrossRef]
- Decaro, N.; Mari, V.; Campolo, M.; et al. Recombinant canine coronaviruses related to transmissible gastroenteritis virus of swine are circulating in dogs. J Virol 2009, 83, 1532–1537. [CrossRef]
- Erles, K.; Brownlie, J. Sequence analysis of divergent canine coronavirus strains present in a UK dog population. Virus Res. 2009, 141, 21–25. [CrossRef]
- Chen, S.; Liu, D.; Tian, J.; Kang, H.; Guo, D.; Jiang, Q.; Liu, J.; Li, Z.; Hu, X.; Qu, L. Molecular characterization of HLJ-073, a recombinant canine coronavirus strain from China with an ORF3abc deletion. Arch Virol. 2019, Aug, 164(8), 2159-2164. [CrossRef]
- Ntafis, V.; Mari, V.; Decaro, N.; Papanastassopoulou, M.; Papaioannou, N.; Mpatziou, R.; Buonavoglia, C.; Xylouri, E. Isolation, tissue distribution and molecular characterization of two recombinant canine coronavirus strains. Vet. Microbiol 2011, 151(3–4), 238-244. [CrossRef]
- Priestnall, S.L.; Mitchell, J.A.; Walker, C.A.; Erles, K.; Brownlie, J. New and Emerging Pathogens in Canine Infectious Respiratory Disease. Veterinary Pathology 2014, 51(2), 492-504. doi:10.1177/0300985813511130 https://journals.sagepub.com/doi/10.1177/0300985813511130.
- Pratelli, A.; Tempesta, M.; Greco, G.; et al. Development of a nested PCR assay for the detection of canine coronavirus. J Virol Methods 1999, 80, 11–15. [CrossRef]
- Komarova, A.A., Galkina, T.S. Antigenic activity of the «Rich» strain of canine coronavirus enteritis virus in experiments with rabbits, ferrets and guinea pigs. Veterinary pathology 2023, 22 (4), 12-18. [in Russ.]. [CrossRef]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; Niu, P.; Zhan, F.; Ma, X.; Wang, D.; Xu, W.; Wu, G.; Gao, G.F.; Tan, W. China Novel Coronavirus Investigating and Research Team. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N Engl J Med 2020, Feb, 20, 382(8), 727-733. [CrossRef]
- World Health Organization. Coronavirus disease (COVID-19) Epidemiological Updates and Monthly Operational Updates. https://www.who.int/emergencies/diseases/novel-coronavirus-2019/situation-reports accessed on 29 November 2024.
- Xu, X.; Chen, P.; Wang, J.; Feng, J.; Zhou, H.; Li, X.; Zhong, W.; Hao, P. Evolution of the novel coronavirus from the ongoing Wuhan outbreak and modeling of its spike protein for risk of human transmission. Science China. Life sciences 2020, 63(3), 457–460. [CrossRef]
- Letko, M.; Munster, V. Functional assessment of cell entry and receptor usage for lineage B β-coronaviruses, including 2019-nCoV. BiorXiv : journal 2020, 22 January, 2020.01.22.915660. doi:10.1101/2020.01.22.915660.
- Zhou, P.; Shi, Zh.-L. Discovery of a novel coronavirus associated with the recent pneumonia outbreak in humans and its potential bat origin. BiorXiv : journal 2020, 2020.01.22.914952. doi:10.1101/2020.01.22.914952.
- Gralinski, L.E.; Menachery V.D. Return of the Coronavirus : 2019-nCoV. Viruses 2020, Vol. 12, no. 2 (24 January), 135. [CrossRef]
- Enjuanes, L.; Sola, I.; Zúñiga, S.; Honrubia, J.M.; Bello-Pérez, M.; Sanz-Bravo, A.; González-Miranda, E.; Hurtado-Tamayo, J.; Requena-Platek, R.; Wang, L.; Muñoz-Santos, D.; Sánchez, C.M.; Esteban, A.; Ripoll-Gómez, J. Nature of viruses and pandemics: Coronaviruses. Curr Res Immunol 2022, 3, 151–158. [CrossRef]
- Kabbani, N.; Olds, J.L. Does COVID19 infect the brain? If so, smokers might be at a higher risk. Molecular Pharmacology: journal 2020, 1 April, vol. 97, no. 5, 351-353. [CrossRef]
- Baig, A. M. Neurological manifestations in COVID-19 caused by SARS-CoV-2. CNS Neuroscience & Therapeutics 2020, May, vol. 26, no. 5, 499—501. [CrossRef]
- Pandit, R.; Matthews, Q.L. A SARS-CoV-2: Companion Animal Transmission and Variants Classification. Pathogens 2023, May 29, 12(6), 775. [CrossRef]
- Chan, T.; Klaus, J.; Meli, M.L.; Hofmann-Lehmann, R.; SARS-CoV-2 Infektionen bei Katzen, Hunden und anderen Tieren: Erkenntnisse zur Infektion und Daten aus der Schweiz [SARS-CoV-2 infections in cats, dogs, and other animal species: Findings on infection and data from Switzerland]. Schweiz Arch Tierheilkd 2021, Dec, 163(12), 821-835. [German.] . [CrossRef]
- Valencak, T.G.; Csiszar, A.; Szalai, G.; Podlutsky, A.; Tarantini, S.; Fazekas-Pongor, V.; Papp, M.; Ungvari. Z. Animal reservoirs of SARS-CoV-2: calculable COVID-19 risk for older adults from animal to human transmission. Geroscience 2021, Oct, 43(5), 2305-2320. [CrossRef]
- Miller, M.R.; Braun, E.; Ip, H.S.; Tyson, G.H. Domestic and wild animal samples and diagnostic testing for SARS-CoV-2. Vet Q 2023, Dec, 43(1), 1-11. [CrossRef]
- Zhai, X.; Sun, J.; Yan, Z.; Zhang, J.; Zhao, J.; Zhao, Z.; Gao, Q.; He, W.T.; Veit, M.; Su, S. Comparison of Severe Acute Respiratory Syndrome Coronavirus 2 Spike Protein Binding to ACE2 Receptors from Human, Pets, Farm Animals, and Putative Intermediate Hosts. J Virol 2020, Jul 16, 94(15), e00831-20. [CrossRef]
- Hayashi, T.; Abiko, K.; Mandai, M.; Yaegashi, N.; Konishi, I.; Highly conserved binding region of ACE2 as a receptor for SARS-CoV-2 between humans and mammals. Vet Q 2020, Dec, 40(1), 243-249. [CrossRef]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 2020, 581, 215–220. [CrossRef]
- Kamel, M.S.; El-Sayed, A.A.; Munds, R.A.; Verma, M.S. Interactions between Humans and Dogs during the COVID-19 Pandemic: Recent Updates and Future Perspectives. Animals (Basel) 2023, Feb, 2, 13(3), 524. https://www.mdpi.com/2076-2615/13/3/524. [CrossRef]
- Kulichenko, A.N., Maletskaya, O.V., Sarkisyan, N.S., Volynkina, A.S. COVID-19 as a Zoonotic Infection. Infection and Immunity 2021, 11 (4), 617-623. [in Russ.] https://iimmun.ru/iimm/article/view/1621. [CrossRef]
- Murphy, H.L.; Ly, H. Understanding the prevalence of SARS-CoV-2 (COVID-19) exposure in companion, captive, wild, and farmed animals. Virulence 2021, Dec, 12(1), 2777-2786. [CrossRef]
- Islam, A.; Ferdous, J.; Islam, S.; Sayeed, M.A.; Rahman, M.K.; Saha, O.; Hassan, M.M.; Shirin, T. Transmission dynamics and susceptibility patterns of SARS-CoV-2 in domestic, farmed and wild animals: Sustainable One Health surveillance for conservation and public health to prevent future epidemics and pandemics. Transbound Emerg Dis 2022, Sep, 69(5), 2523-2543. [CrossRef]
- Sharun, K.; Dhama, K.; Pawde, A.M.; Gortázar, C.; Tiwari, R.; Bonilla-Aldana, D.K.; Rodriguez-Morales, A.J.; de la Fuente, J.; Michalak, I.; Attia, Y.A. SARS-CoV-2 in animals: potential for unknown reservoir hosts and public health implications. Vet Q 2021, Dec, 41(1), 181-201. [CrossRef]
- Klaus, J.; Zini, E.; Hartmann, K.; Egberink, H.; Kipar, A.; Bergmann, M.; Palizzotto, C.; Zhao, S.; Rossi, F.; Franco, V.; Porporato, F.; Hofmann-Lehmann, R. Meli ML SARS-CoV-2 Infection in Dogs and Cats from Southern Germany and Northern Italy during the First Wave of the COVID-19 Pandemic. Viruses 2021, Jul 26, 13(8), 1453. [CrossRef]
- Michelitsch, A.; Allendorf ,V.; Conraths, F.J.; Gethmann, J.; Schulz, J.; Wernike, K.; Denzin, N. SARS-CoV-2 Infection and Clinical Signs in Cats and Dogs from Confirmed Positive Households in Germany. Viruses 2023, Mar, 24, 15(4), 837. [CrossRef]
- Colitti, B.; Bertolotti, L.; Mannelli, A.; Ferrara, G.; Vercelli, A.; Grassi, A.; Trentin, C.; Paltrinieri, S.; Nogarol, C.; Decaro, N.; Brocchi, E.; Rosati, S. Cross-Sectional Serosurvey of Companion Animals Housed with SARS-CoV-2-Infected Owners, Italy. Emerg Infect Dis 2021, Jul, 27(7), 1919-1922. [CrossRef]
- Bianco, A.; Bortolami, A.; Miccolupo, A.; Sottili, R.; Ghergo, P.; Castellana, S.; Del Sambro, L.; Capozzi, L.; Pagliari, M.; Bonfante, F.; Ridolfi, D.; Bulzacchelli, C.; Giannico, A.; Parisi, A. SARS-CoV-2 in Animal Companions: A Serosurvey in Three Regions of Southern Italy. Life (Basel) 2023, Dec, 16, 13(12), 2354. [CrossRef]
- Kuhlmeier, E.; Chan, T.; Meli, M.L.; Willi, B.; Wolfensberger, A.; Reitt, K.; Hüttl, J.; Jones, S.; Tyson, G.; Hosie, M.J.; Zablotski, Y.; Hofmann-Lehmann, R. A Risk Factor Analysis of SARS-CoV-2 Infection in Animals in COVID-19-Affected Households. Viruses 2023, Mar, 11, 15(3), 731. [CrossRef]
- Fischer, E.A.J.; Broens, E.M.; Kooistra, H.S.; De Rooij, M.M.T.; Stegeman, J.A.; De Jong, M.C.M. Contribution of cats and dogs to SARS-CoV-2 transmission in households. Front Vet Sci 2023, Jul. 14, 10, 1151772. [CrossRef]
- Stanojevic, S.; Radojicic, S.; Misic, D.; Srejić, D.; Vasiljevic, D.V.; Prokic, K.; Ilić, N. Frequency of SARS-CoV-2 infection in dogs and cats: Results of a retrospective serological survey in Šumadija District, Serbia. Prev Vet Med 2022, Nov, 208, 105755. [CrossRef]
- Stevanovic, V.; Vilibic-Cavlek, T.; Tabain, I.; Benvin, I.; Kovac, S.; Hruskar, Z.; Mauric, M.; Milasincic, L.; Antolasic, L.; Skrinjaric, A.; Staresina, V.; Barbic, L. Seroprevalence of SARS-CoV-2 infection among pet animals in Croatia and potential public health impact. Transbound Emerg Dis 2021, Jul, 68(4), 1767-1773. [CrossRef]
- Paltseva, E.D.; Pleshakova, V.I.; Rudakova, S.A. Detection of infection of companion animals with the SARS-COV-2 virus. Perm Agrarian Bulletin 2022, 3(39), 118-125. URL: https://cyberleninka.ru/article/n/vyyavlenie-infitsirovannosti-zhivotnyh-kompanonov-virusom-sars-cov-2 accessed on 12 November 2024 [in Russ.].
- Kuroda, Y.; Watanabe, K.; Yamamoto, T.; Suzuki, H.; Park, E.S.; Ishijima, K.; Tatemoto, K.; Virhuez-Mendoza, M.; Inoue, Y.; Harada, M.; Nishino, A.; Sekizuka, T.; Kuroda, M.; Fujimoto, T.; Ishihara, G.; Horie, R.; Kawamoto, K.; Maeda, K. Pet Animals Were Infected with SARS-CoV-2 from Their Owners Who Developed COVID-19: Case Series Study. Viruses 2023, Sep, 29, 15(10), 2028. [CrossRef]
- Khalife, S.; Abdallah, M. High seroprevalence of SARS-CoV-2 antibodies in household cats and dogs of Lebanon. Res Vet Sci 2023, Apr, 157, 13-16. [CrossRef]
- Go, Y.Y.; Carrai, M.; Choi, Y.R.; Brackman, C.J.; Tam, K.W.S.; Law, P.Y,T.; Woodhouse, F.; Gray, J.; Kim, J.H.; Park, J.; Jeon, C.W.; Jang, H.; Magouras, I.; Decaro, N.; Cheng, S.M.S.; Peiris, M.; Beatty, J.A.; Barrs, V.R. Low Prevalence of SARS-CoV-2 Antibodies in Canine and Feline Serum Samples Collected during the COVID-19 Pandemic in Hong Kong and Korea. Viruses 2023, Feb, 20, 15(2), 582. [CrossRef]
- Sit, T.H.C.; Brackman, C.J.; Ip, S.M.; Tam, K.W.S.; Law, P.Y.T.; To, E.M.W.; Yu, V.Y.T.; Sims, L.D.; Tsang, D.N.C.; Chu, D.K.W.; Perera, R.A.P.M.; Poon, L.L.M.; Peiris, M. Infection of dogs with SARS-CoV-2. Nature 2020, Oct, 586(7831), 776-778. [CrossRef]
- AFCD. Blood test result of pet dog with low-level infection of COVID-19 released. www.afcd.gov.hk/english/publications/publications_press/pr2343.html accessed 31 March 2020.
- GHKSAR. Pet dog further tests positive for antibodies for COVID-19 virus. www.info.gov.hk/gia/general/202003/26/P2020032600756.htm accessed 31 March 2020.
- Piewbang, C.; Poonsin, P.; Lohavicharn, P.; Punyathi, P.; Kesdangsakonwut, S.; Kasantikul, T.; Techangamsuwan, S. Natural SARS-CoV-2 infection in dogs: Determination of viral loads, distributions, localizations, and pathology. Acta Trop. 2024, Jan, 249, 107070. [CrossRef]
- Bae, D.Y.; Tark, D.; Moon, SH.; Oem, J.K.; Kim, W.I.; Park, C.; Na, K.J.; Park, C.K.; Oh, Y.; Cho, H.S. Evidence of Exposure to SARS-CoV-2 in Dogs and Cats from Households and Animal Shelters in Korea. Animals (Basel) 2022, Oct, 15, 12(20), 2786. [CrossRef]
- Bienzle, D.; Rousseau, J.; Marom, D.; MacNicol, J.; Jacobson, L.; Sparling, S.; Prystajecky, N.; Fraser, E.; Weese, J.S. Risk Factors for SARS-CoV-2 Infection and Illness in Cats and Dogs1. Emerg Infect Dis 2022, Jun, 28(6), 1154-1162. [CrossRef]
- Meisner, J.; Baszler, T.V.; Kuehl, K.E.; Ramirez, V.; Baines, A.; Frisbie, L.A.; Lofgren, E.T.; de Avila, D.M; Wolking, R.M.; Bradway, D.S.; Wilson, H.R.; Lipton, B.; Kawakami, V.; Rabinowitz, P.M. Household Transmission of SARS-CoV-2 from Humans to Pets, Washington and Idaho, USA. Emerg Infect Dis 2022, Dec, 28(12), 2425-2434. [CrossRef]
- Espinosa-Gómez, F.C.; Bautista, E.; Palacios-Cruz, O.E.; Téllez-Ramírez, A.; Vázquez-Briones, D.B.; Flores de Los Ángeles, C.; Abella-Medrano, C.A.; Escobedo-Straffón, J.L.; Aguirre-Alarcón, H.; Pérez-Silva, N.B.; Solís-Hernández, M.; Navarro-López, R.; Aguirre, A.A. Host traits, ownership behaviour and risk factors of SARS-CoV-2 infection in domestic pets in Mexico. Zoonoses Public Health 2023, Jun, 70(4), 327-340. [CrossRef]
- Fernández-Figueroa, E.A.; Espinosa-Martínez, D.V.; Miranda-Ortiz, H.; Ruiz-García, E.; Figueroa-Esquivel, J.M.; Becerril-Moctezuma, M.L.; Muñoz-Rivas, A.; Ríos-Muñoz, C.A. Evidence of SARS-CoV-2 infection in companion animals from owners who tested positive for COVID-19 in the Valley of Mexico. Mol Biol Rep 2024, Jan, 25, 51(1), 186. [CrossRef]
- Calvet, G.A.; Pereira, S.A.; Ogrzewalska, M.; Pauvolid-Corrêa, A.; Resende, P.C.; Tassinari, W.S.; Costa, A.P.; Keidel, L.O.; da Rocha, A.S.B.; da Silva, M.F.B.; Dos Santos, S.A.; Lima, A.B.M.; de Moraes, I.C.V.; Mendes Junior, A.A.V.; Souza, T.D.C.; Martins, E.B.; Ornellas, R.O.; Corrêa, M.L.; Antonio, I.M.D.S.; Guaraldo, L.; Motta, F.D.C.; Brasil, P.; Siqueira, M.M.; Gremião, I.D.F.; Menezes, R.C. Investigation of SARS-CoV-2 infection in dogs and cats of humans diagnosed with COVID-19 in Rio de Janeiro, Brazil. PLoS One 2021, Apr, 28, 16(4), e0250853. [CrossRef]
- Santos, H.O.; Santos, E.M.S.; de Oliveira, H.D.S.; Dos Santos, W.S.; Tupy, A.A.; Souza, E.G.; Ramires, R.; Luiz, A.C.O.; de Almeida, A.C. Screening for canine coronavirus, canine influenza virus, and severe acute respiratory syndrome coronavirus 2 in dogs during the coronavirus disease-2019 pandemic. Vet World 2023, Sep, 16(9), 1772-1780. [CrossRef]
- Galhardo, J.A.; Barbosa, D.S.; Kmetiuk, L.B.; de Carvalho, O.V.; Teixeira, A.I.P.; Fonseca, P.L.C.; de Araújo, E.; Santos, L.C.G.; Queiroz, D.C.; Miranda, J.V.O.; da Silva Filho, A.P.; Castillo, A.P.; Araujo, R.N.; da Silveira, J.A.G.; Ristow, L.E.; Brandespim, D.F.; Pettan-Brewer, C.; de Sá Guimarães, A.M.; Dutra, V.; de Morais, H.A.; Dos Santos, A.P.; Agopian, R.G.; de Aguiar, R.S.; Biondo, A.W. Molecular detection and characterization of SARS-CoV-2 in cats and dogs of positive owners during the first COVID-19 wave in Brazil. Sci Rep 2023, Sep, 2, 13(1), 14418. [CrossRef]
- Alberto-Orlando, S.; Calderon, J.L.; Leon-Sosa, A.; Patiño, L.; Zambrano-Alvarado, M.N.; Pasquel-Villa, L.D.; Rugel-Gonzalez, D.O.; Flores, D.; Mera, M.D.; Valencia, P.; Zuñiga-Velarde, J.J.; Tello-Cabrera, C.; Garcia-Bereguiain, M.A. SARS-CoV-2 transmission from infected owner to household dogs and cats is associated with food sharing. Int J Infect Dis 2022, Sep, 122, 295-299. [CrossRef]
- Gautam, A.; Kaphle, K.; Shrestha, B.; Phuyal, S. Susceptibility to SARS, MERS, and COVID-19 from animal health perspective. Open Vet J 2020, Aug, 10(2), 164-177. [CrossRef]
- Jaramillo Hernández, D.A.; Chacón, M.C.; Velásquez, M.A.; Vásquez-Trujillo, A.; Sánchez, A.P.; Salazar Garces, L.F.; García, G.L.; Velasco-Santamaría, Y.M.; Pedraza, L.N.; Lesmes-Rodríguez, L.C. Seroprevalence of exposure to SARS-CoV-2 in domestic dogs and cats and its relationship with COVID-19 cases in the city of Villavicencio, Colombia. F1000Res. 2023, Aug 10, 11, 1184. [CrossRef]
- Molini, U.; Coetzee, L.M.; Engelbrecht, T.; de Villiers, L.; de Villiers, M.; Mangone, I.; Curini, V.; Khaiseb, S.; Ancora, M.; Cammà, C.; Lorusso, A.; Franzo, G.; SARS-CoV-2 in Namibian Dogs. Vaccines (Basel) 2022, Dec 13, 10(12), 2134. [CrossRef]
- Kuhlmeier, E.; Chan, T.; Agüí, C.V.; Willi, B.; Wolfensberger, A.; Beisel, C.; Topolsky, I.; Beerenwinkel, N.; Stadler, T. Detection and Molecular Characterization of the SARS-CoV-2 Delta Variant and the Specific Immune Response in Companion Animals in Switzerland. Viruses. 2023, 15(1), 245. [CrossRef]
- Akhtardanesh, B.; Jajarmi, M.; Shojaee, M.; Salajegheh Tazerji, S.; Khalili Mahani, M.; Hajipour, P.; Gharieb, R. Molecular screening of SARS-CoV-2 in dogs and cats from households with infected owners diagnosed with COVID-19 during Delta and Omicron variant waves in Iran. Vet Med Sci 2023, Jan, 9(1), 82-90. [CrossRef]
- Agüero, B.; Berrios, F.; Pardo-Roa, C.; Ariyama, N.; Bennett, B.; Medina, R.A.; Neira, V. First detection of Omicron variant BA.4.1 lineage in dogs, Chile. Vet Q 2024, Dec, 44(1), 1-10. [CrossRef]
- Tinto, B.; Revel, J.; Virolle, L.; Chenet, B.; Reboul Salze, F.; Ortega, A.; Beltrame, M.; Simonin, Y. Monitoring SARS-CoV-2 Seroprevalence in Domestics and Exotic Animals in Southern France. Trop Med Infect Dis 2023, Aug 25, 8(9), 426. [CrossRef]
- IDEXX. Leading veterinary diagnostic company sees no COVID-19 cases in pets. www.idexx.com/en/about-idexx/news/no-covid-19-cases-pets (accessed 31 March 2020).
- Johansen, M.D., Irving, A., Montagutelli, X. et al. Animal and translational models of SARS-CoV-2 infection and COVID-19. Mucosal Immunol 2020, 13, 877–891. [CrossRef]
- Almendros, A.; Gascoigne, E. Can companion animals become infected with Covid-19? Vet Rec 2020, Apr 4, 186(13), 419-420. [CrossRef]
- Michael, H.T.; Waterhouse, T.; Estrada, M.; Seguin, M.A. Frequency of respiratory pathogens and SARS-CoV-2 in canine and feline samples submitted for respiratory testing in early 2020. J Small Anim Pract 2021, May, 62(5), 336-342. [CrossRef]
- Piewbang, C.; Poonsin, P.; Lohavicharn, P.; Wardhani, S.W.; Dankaona, W.; Puenpa, J.; Poovorawan, Y.; Techangamsuwan, S. SARS-CoV-2 Transmission from Human to Pet and Suspected Transmission from Pet to Human, Thailand. J Clin Microbiol 2022, Nov 16, 60(11), e0105822. [CrossRef]
- Shi, J.; Wen, Z.; Zhong, G.; Yang, H.; Wang, C.; Huang, B.; Liu, R.; He, X.; Shuai, L.; Sun, Z.; Zhao, Y.; Liu, P.; Liang, L.; Cui, P.; Wang, J.; Zhang, X.; Guan, Y.; Tan, W.; Wu, G.; Chen, H.; Bu, Z. Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS-coronavirus 2. Science 2020, May 29, 368(6494), 1016-1020. [CrossRef]
- Andreeva, A.V.; Nikolaeva, O.N. New coronavirus infection (COVID-19) in animals. Veterinarian 2021, 2, 4–11. [in Russ.]. [CrossRef]
- Temmam, S., Vongphayloth, K., Baquero, E. et al. Bat coronaviruses related to SARS-CoV-2 and infectious for human cells. Nature 2022, 604, 330–336. [CrossRef]
- Sparrer, M.N.; Hodges, N.F.; Sherman, T.; VandeWoude, S.; Bosco-Lauth, A.M.; Mayo, C.E. Role of Spillover and Spillback in SARS-CoV-2 Transmission and the Importance of One Health in Understanding the Dynamics of the COVID-19 Pandemic. J. Clin. Microbiol 2023, e0161022. [CrossRef]
- Brüssow, H. Viral infections at the animal–human interface-Learning lessons from the SARS-CoV-2 pandemic. Microb. Biotechnol. 2023, 1-15. [CrossRef]
- Chan, J.F.W.; Siu, G.K.H.; Yuan, S.; Ip, J.D.; Cai, J.P.; Chu, A.W.H.; Chan, W.M.; Abdullah S.M.U.; Luo, C.; Chan, B.P.C.; et al. Probable Animal-to-Human Transmission of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Delta Variant AY.127 Causing a Pet Shop-Related Coronavirus Disease 2019 (COVID-19) Outbreak in Hong Kong. Clin. Infect. Dis. 2022, 75:e76–e81. [CrossRef]
- Yen, H.L.; Sit, T.H.C.; Brackman, C.J.; Chuk, S.S.Y.; Gu, H.; Tam, K.W.S.; Law, P.Y.T.; Leung, G.M.; Peiris, M.; Poon, L.L.M. Transmission of SARS-CoV-2 delta variant (AY.127) from pet hamsters to humans, leading to onward human-to-human transmission: A case study. Lancet. 2022,399:1070–1078. [CrossRef]
- Wei, C.; Shan, K.J.; Wang, W.; Zhang, S.; Huan, Q.; Qian, W. Evidence for a mouse origin of the SARS-CoV-2 Omicron variant. J. Genet. Genom. 2021, 48:1111–1121. [CrossRef]
- Tiwari, R.; Dhama, K.; Sharun, K.; Iqbal Yatoo, M.; Malik, Y.S.; Singh, R.; Michalak, I.; Sah, R.; Bonilla-Aldana, D.K.; Rodriguez-Morales, A.J. COVID-19: animals, veterinary and zoonotic links. Vet Q 2020, Dec,40(1),169-182. [CrossRef]
- Salajegheh Tazerji, S.; Magalhães Duarte, P.; Rahimi, P.; Shahabinejad, F.; Dhakal, S.; Singh Malik, Y.; Shehata, A.A.; Lama, J.; Klein, J.; Safdar, M.; Rahman, M.T.; Filipiak, K.J.; Rodríguez-Morales, A.J.; Sobur, M.A.; Kabir, F.; Vazir, B.; Mboera, L.; Caporale, M.; Islam, M.S.; Amuasi, J.H.; Gharieb, R.; Roncada, P.; Musaad, S.; Tilocca, B.; Koohi, M.K.; Taghipour, A.; Sait, A.; Subbaram, K.; Jahandideh, A.; Mortazavi, P.; Abedini, M.A.; Hokey, D.A.; Hogan, U.; Shaheen, M.N.F.; Elaswad, A.; Elhaig, M.M.; Fawzy, M. Transmission of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) to animals: an updated review. J Transl Med 2020, Sep 21,18(1),358. [CrossRef]
- Salajegheh Tazerji, S.; Gharieb, R.; Ardestani, M.M.; Akhtardanesh, B.; Kabir, F.; Vazir, B.; Duarte, P.M.; Saberi, N.; Khaksar, E.; Haerian, S.; Fawzy, M. The risk of pet animals in spreading severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) and public health importance: An updated review. Vet Med Sci 2024, Jan,10(1),e1320. [CrossRef]
- Reagan, K.L.; Sykes, J.E. Canine Infectious Respiratory Disease. Vet Clin North Am Small Anim Pract 2020, Mar,50(2),405-418. [CrossRef]
- Mitchell, J.A.; Cardwell, J.M.; Leach, H.; Walker, C.A.; Le Poder, S.; Decaro, N.; Rusvai, M.; Egberink, H,; Rottier, P.; Fernandez, M.; Fragkiadaki, E.; Shields, S.; Brownlie, J. European surveillance of emerging pathogens associated with canine infectious respiratory disease. Vet Microbiol 2017, Dec,212,31-38. [CrossRef]
- Decaro, N.; Mari, V.; Larocca, V.; Losurdo, M.; Lanave, G.; Lucente, M.S.; Corrente, M.; Catella, C.; Bo, S.; Elia, G.; Torre, G.; Grandolfo, E.; Martella, V.; Buonavoglia, C. Molecular surveillance of traditional and emerging pathogens associated with canine infectious respiratory disease. Vet Microbiol 2016, Aug 30,192,21-25. [CrossRef]
- Erles, K.; Dubovi, E.J.; Brooks, H.W.; Brownlie, J. Longitudinal study of viruses associated with canine infectious respiratory disease. J Clin Microbiol 2004, Oct,42(10),4524-9. [CrossRef]
- Erles, K.; Brownlie, J. Investigation into the causes of canine infectious respiratory disease: antibody responses to canine respiratory coronavirus and canine herpesvirus in two kennelled dog populations. Arch Virol 2005, Aug,150(8),1493-504. [CrossRef]
- Schulz, B.S.; Kurz, S.; Weber, K.; Balzer, H.J.; Hartmann, K. Detection of respiratory viruses and Bordetella bronchiseptica in dogs with acute respiratory tract infections. Vet J 2014, Sep,201(3),365-9. [CrossRef]
- Sowman, H.R.; Cave, N.J.; Dunowska, M. A survey of canine respiratory pathogens in New Zealand dogs. N Z Vet J 2018, Sep,66(5),236-242. [CrossRef]
- Piewbang, C.; Rungsipipat, A.; Poovorawan, Y.; Techangamsuwan, S. Cross-sectional investigation and risk factor analysis of community-acquired and hospital-associated canine viral infectious respiratory disease complex. Heliyon 2019, Nov 14,5(11),e02726. [CrossRef]
- Maboni, G; Seguel, M.; Lorton, A.; Berghaus, R.; Sanchez, S. Canine infectious respiratory disease: New insights into the etiology and epidemiology of associated pathogens. PLoS One 2019, Apr 25,14(4),e0215817. [CrossRef]
- Bryan, H.M.; Darimont, C.T.; Paquet, P.C.; Ellis, J.A.; Goji, N.; Gouix, M.; Smits, J.E. Exposure to infectious agents in dogs in remote coastal British Columbia: Possible sentinels of diseases in wildlife and humans. Can J Vet Res 2011, Jan, 75(1), 11-7. https://www.wellbeingintlstudiesrepository.org/zoodis/1/ accessed on 29 November 2024.
- Shi, Y.; Tao, J.; Li, B.; Shen, X.; Cheng, J.; Liu, H. The Gut Viral Metagenome Analysis of Domestic Dogs Captures Snapshot of Viral Diversity and Potential Risk of Coronavirus. Front Vet Sci 2021, Jul 7, 8 695088. [CrossRef]
- Yondo, A.; Kalantari, A.A.; Fernandez-Marrero, I.; McKinney, A.; Naikare, H.K.; Velayudhan, B.T. Predominance of Canine Parainfluenza Virus and Mycoplasma in Canine Infectious Respiratory Disease Complex in Dogs. Pathogens 2023, Nov 15, 12(11), 1356. [CrossRef]
- Nikonenko, T.B.; Baryshnikov, P.I.; Novikov, N.A. Microbiocenoses in viral intestinal infections of dogs in the Baikal region. Bulletin of the Altai State Agrarian University 2021. 1 (195), 83-88. URL: https://cyberleninka.ru/article/n/mikrobiotsenozy-pri-virusnyh-kishechnyh-infektsiyah-sobak-v-usloviyah-pribaykalya accessed on 12 November 2024 [in Russ.].
- Woolhouse, M.E.; Gowtage-Sequeria, S. Host range and emerging and reemerging pathogens. Emerg Infect Dis 2005, Dec,11(12),1842-7. [CrossRef]
- McMahon, B.J.; Morand, S.; Gray, J.S. Ecosystem change and zoonoses in the Anthropocene. Zoonoses Public Health 2018, Nov, 65(7), 755-765. [CrossRef]
- Pellegrini, F.; Omar, A.H.; Buonavoglia, C.; Pratelli, A. SARS-CoV-2 and Animals: From a Mirror Image to a Storm Warning. Pathogens 2022, Dec 12, 11(12), 1519. [CrossRef]
- Shamenova, M.; Lukmanova, G.; Klivleyeva, N.; Glebova, T.; Saktaganov, N.; Baimukhametova, A.; Vetrova, E. Interspecies Transmission and Adaptation of Influenza Virus to Dogs. Microbiology and Virology 2024, 1(44), 22–41. https://imv-journal.kz/index.php/mav/article/view/215. [CrossRef]
- Liu, B.; Zhao, P.; Xu, P.; Han, Y.; Wang, Y.; Chen, L.; Wu, Z.; Yang, J. A comprehensive dataset of animal-associated sarbecoviruses. Sci Data 2023, Oct 7, 10(1), 681. [CrossRef]
- Yesilbag, K.; Aytogu, G. Coronavirus host divergence and novel coronavirus (Sars-CoV-2) outbreak. Clin Exp Ocul Trauma Infect 2020, 2(1), 139–147. https://dergipark.org.tr/tr/download/article-file/1081346 accessed on 11 November 2024.
- Lorusso, A.; Calistri, P.; Petrini, A.; Savini, G.; Decaro, N. Novel coronavirus (SARS-CoV-2) epidemic: a veterinary perspective. Vet Ital 2020, 56(1), 5–10. [CrossRef]
- Klivleyeva, N.; Lukmanova, G.; Glebova, T.; Shamenova, M.; Ongarbayeva, N.; Saktaganov, N.; Baimukhametova, A.; Baiseiit, S.; Ismagulova, D.; Kassymova, G.; Rachimbayeva, A.; Murzagaliyeva, A.; Xetayeva, G.; Isabayeva, R.; Sagatova, M. Spread of Pathogens Causing Respiratory Viral Diseases Before and During CoVID-19 Pandemic in Kazakhstan. Indian J Microbiol 2023, February. [CrossRef]
- Klivleyeva, N.G.; Ongarbayeva, N.S.; Korotetskiy, I.S.; Glebova, T.I.; Saktaganov, N.T.; Shamenova, M.G.; Baimakhanova, B.B.; Shevtsov, A.B.; Amirgazin, A.; Berezin, V.E.; et al. Coding-Complete Genome Sequence of Swine Influenza Virus Isolate A/Swine/Karaganda/04/2020 (H1N1) from Kazakhstan. Microbiol. Resour. Announc. 2021, 10, e0078621. [CrossRef]
- Saktaganov, N.T.; Klivleyeva, N.G.; Ongarbayeva, N.S.; Glebova, T.I.; Lukmanova, G.V.; Baimukhametova, A.M. Study on Antigenic Relationships and Biological Properties of Swine Influenza a/H1n1 Virus Strains Isolated in Northern Kazakhstan in 2018. Sel'skokhozyaistvennaya Biol. 2020, 55, 355–363. [CrossRef]
- Klivleyeva, N.G.; Glebova, T.I.; Shamenova, M.G.; Saktaganov, N.T. Influenza A viruses circulating in dogs: A review of the scientific literature. Open Veterinary Journal 2022, 12(5), 676–687. [CrossRef]
- Licitra, B.N.; Duhamel, G.E.; Whittaker, G.R. Canine enteric coronaviruses: emerging viral pathogens with distinct recombinant spike proteins. Viruses 2014, 6, 3363–3376. [CrossRef]
- Esposito, A.M.; Esposito, M.M.; Ptashnik, A. Phylogenetic Diversity of Animal Oral and Gastrointestinal Viromes Useful in Surveillance of Zoonoses. Microorganisms 2022, Sep 10, 10(9), 1815. [CrossRef]
- Klivleyeva N, Saktaganov N, Glebova T, Lukmanova G, Ongarbayeva N, Webby R. Influenza A Viruses in the Swine Population: Ecology and Geographical Distribution. Viruses 2024, 16(11), 1728. [CrossRef]
- Glebova, T.I.; Klivleyeva, N.G.; Saktaganov, N.T.; Shamenova, M.G.; Lukmanova, G.V.; Baimukhametova, A.M.; Baiseiit, S.B.; Ongarbayeva, N.S.; Orynkhanov, K.A.; Ametova, A.V.; Ilicheva, A.K. Circulation of influenza viruses in the dog population in Kazakhstan (2023–2024), Open Veterinary Journal 2024, 14(8), 1896-1904 . [CrossRef]
- Lukmanova, G.; Klivleyeva, N.; Glebova, T.; Ongarbayeva, N.; Shamenova, M.; Saktaganov, N.; Baimukhametova, A.; Baiseiit, S.; Ismagulova, D.; Ismailov, E.; et al. Influenza A Virus Surveillance in Domestic Pigs in Kazakhstan 2018-2021. Ciênc. Rural 2024, 54(10), e20230403. [CrossRef]
- Glebova T.I., Klivleyeva N.G., Baimukhametova A.M., Saktaganov N.T., Lukmanova G.V., Ongarbayeva N.S., Shamenova M.G., Baimakhanova B.B. Circulation of influenza viruses in the epidemic season of 2018–2019 among people residing in Northern and Western Kazakhstan. Infekc. bolezni (Infectious diseases). 2021, 19(2), 70–75. [in Russ]. [CrossRef]
- Khalil, A.M.; Martinez-Sobrido, L.; Mostafa, A. Zoonosis and zooanthroponosis of emerging respiratory viruses. Front Cell Infect Microbiol 2024, Jan 5, 13, 1232772. [CrossRef]
- Vlasova A.N., Diaz A., Damtie D., Xiu L., Toh T.-H., Soon-Yit Lee J., Saif L. J, Gray G.C., Novel Canine Coronavirus Isolated from a Hospitalized Patient With Pneumonia in East Malaysia, Clinical Infectious Diseases, 2022, 74(3), 446–454, . [CrossRef]
- Lednicky, J.A.; Tagliamonte, M.S.; White, S.K.; Blohm, G.M.; Alam, Md. M.; Iovine, N.M.; Salemi, M.; Mavian, C.; Morris G. J. Isolation of a Novel Recombinant Canine Coronavirus From a Visitor to Haiti: Further Evidence of Transmission of Coronaviruses of Zoonotic Origin to Humans, Clinical Infectious Diseases, 2022, 75(1), e1184–e1187, . [CrossRef]
- Domańska-Blicharz, K.; Woźniakowski, G.; Konopka, B.; Niemczuk, K.; Welz, M.; Rola, J.; Socha, W.; Orłowska, A.; Antas, M.; Śmietanka, K.; et al. Animal Coronaviruses in the Light of COVID-19. J. Vet. Res. 2020, 64, 333–345. [CrossRef]
- Pratelli, A. Genetic evolution of canine coronavirus and recent advances in prophylaxis. Vet Res. 2006, 37, 191–200. [CrossRef]
- USDA Confirmation of COVID-19 in Pet Dog in New York. Available online: https://content.govdelivery.com/accounts/USDAAPHIS/bulletins/28ead4f accessed on 1 September 2021.
- Liew, A.Y.; Carpenter, A.; Moore, T.A.; Wallace, R.M.; Hamer, S.A.; Hamer, G.L.; Fischer, R.S.B.; Zecca, I.B.; Davila, E.; Auckland, L.D. Clinical and epidemiologic features of SARS-CoV-2 in dogs and cats compiled through national surveillance in the United States. J. Am. Vet. Med. Assoc. 2023, 261, 480–489. [CrossRef]
- USDA Confirmed Cases of SARS-CoV-2 in Animals in the United States. Available online: https://www.aphis.usda.gov/aphis/dashboards/tableau/sars-dashboard. accessed on 15 May 2023.
- Hamer, S.A.; Ghai, R.R.; Zecca, I.B.; Auckland, L.D.; Roundy, C.M.; Davila, E.; Busselman, R.E.; Tang, W.; Pauvolid-Corrêa, A.; Killian, M.L. SARS-CoV-2 B.1.1.7 variant of concern detected in a pet dog and cat after exposure to a person with COVID-19, USA. Transbound. Emerg. Dis. 2022, 69, 1656–1658. [CrossRef]
- Wendling, N.M.; Carpenter, A.; Liew, A.; Ghai, R.R.; Gallardo-Romero, N.; Stoddard, R.A.; Tao, Y.; Zhang, J.; Retchless, A.C.; Ahmad, A. Transmission of SARS-CoV-2 Delta variant (B.1.617.2) from a fully vaccinated human to a canine in Georgia, July 2021. Zoonoses Public Health 2022, 69, 587–592. [CrossRef]
- Doerksen, T.; Lu, A.; Noll, L.; Almes, K.; Bai, J.; Upchurch, D.; Palinski, R. Near-Complete Genome of SARS-CoV-2 Delta (AY.3) Variant Identified in a Dog in Kansas, USA. Viruses 2021, 13, 2104. [CrossRef]
- Sila, T.; Sunghan, J.; Laochareonsuk, W.; Surasombatpattana, S.; Kongkamol, C.; Ingviya, T.; Siripaitoon, P.; Kositpantawong, N.; Kanchanasuwan, S.; Hortiwakul, T.; et al. Suspected Cat-to-Human Transmission of SARS-CoV-2, Thailand, July–September 2021. Emerg. Infect. Dis. 2022, 28, 1485–1488. [CrossRef]
- Yadav, T.; Saxena, S.K. Transmission Cycle of SARS-CoV and SARS-CoV-2. In Coronavirus disease 2019 (COVID-19): Epidemiology, pathogenesis, diagnosis, and therapeutics., S. K. Saxena Eds.; Springer Science and Business Media, LLC, Singapore, 2020, 33–42. [CrossRef]
- Plowright, R.K.; Parrish, C.R.; McCallum, H.; Hudson, P.J.; Ko, A.I.; Graham, A.L.; et al. Pathways to zoonotic spillover. Nat Rev Microbiol 2017, 15, 502–10. [CrossRef]
- Jelinek, H.F.; Mousa, M.; Alefishat, E.; Osman, W.; Spence, I.; Bu, D.; Feng, S.F.; Byrd, J.; Magni, P.A.; Sahibzada, S.; Tay, G.K.; Alsafar, H.S. Evolution, Ecology, and Zoonotic Transmission of Betacoronaviruses: A Review. Front Vet Sci 2021, May 20, 8, 644414. [CrossRef]
- Fang, R.; Yang, X.; Guo, Y.; Peng, B.; Dong, R.; Li, S.; Xu, S. SARS-CoV-2 infection in animals: Patterns, transmission routes, and drivers. Eco Environ Health 2023, Oct 13, 3(1), 45-54. [CrossRef]
- Colina, S.E.; Serena, M.S.; Echeverría, M.G.; Metz, G.E. Clinical and molecular aspects of veterinary coronaviruses. Virus Res 2021, May, 297, 198382. [CrossRef]
- Daszak, P.; Cunningham, A.A.; Hyatt, A.D. Emerging infectious diseases of wildlife-threats to biodiversity and human health. Science 2000, Jan 21, 287(5452), 443-9. [CrossRef]
- Wang, J.; Zhu, Q.; Xing, X.; Sun, D. A Mini-Review on the Common Antiviral Drug Targets of Coronavirus. Microorganisms 2024, Mar 17, 12(3)m 600. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



