
Submitted:
29 December 2024
Posted:
30 December 2024
You are already at the latest version
Abstract
Human Parvovirus B19 (B19V) is a widespread virus, causing erythema infectiosum in children and several clinical manifestations from acute to persistent infections in adults. In early 2024, ECDC reported an increased human parvovirus B19 circulation in 14 European countries. A Parvovirus B19 outbreak was also reported in Tuscany, Italy, prompting a detailed investigation of its genetic characteristics. This study is the first Italian report on this outbreak, which carries out a phylogenetic analysis based on the complete coding B19V genomes of 23 positive samples. Phylogenetic clustering assigned all isolates to the G1a genotype, although with some mutations in NS1, VP1 and VP2, compared to the Reference strains.
Keywords:
Parvovirus B19
; outbreak
; G1a genotype
1. Introduction
Human parvovirus B19 (B19V) is a single-stranded DNA virus (ssDNA), belonging to the Parvoviridae family. It is the aetiologic agent of the erythema infectiosum (fifth disease) in children and it can infect adults, particularly those who are immunosuppressed or have chronic hematological diseases, causing a wide range of clinical manifestations [1]. Although most of the patients are asymptomatic or with mild symptoms, when B19V affects adults, it can lead to more serious conditions, such as arthropathy, aplastic crises and hydrops fetalis in pregnant women [2,3]. HB19V can be transmitted via respiratory secretions, hand-to-mouth contact, blood transfusion or transplacental transmission [4]. The incubation period varies from 4 to 14 days after exposure but it can last up to 3 weeks [5]. B19V infections usually follow a seasonal pattern, with incidence peaks typically occurring in late spring or early summer, especially in temperate countries [6]. Larger epidemics have been reported every 3-4 years, with fluctuation in the number of cases across different seasons [7]. Since the first few months of the 2024 and, especially during late spring-early summer, an unexpected surge in B19V was noted in a lot of European countries [8,9] and consequently in Tuscany, in Siena area. While only three cases were recorded in our laboratory in 2023, a lot of requests arrived at the Virology Unit and various samples were positively diagnosed from January to September 2024.
B19V genome is a 5596-nucleotide long ssDNA molecule, encoding for several structural and non-structural proteins. Its genome is composed of two main regions (Figure 1): the non-coding region, formed by two inverted terminal repeats (ITR) and mainly involved in viral replication and encapsidation process, and the coding region for four non-structural proteins (NS1, 7.5-, 9- and 11-kDa) and two structural proteins (VP1 and VP2). The non-structural 1 protein (NS1) is involved in viral replication, cytotoxicity, and host immune response. The structural VP proteins include VP1, containing a unique N-terminus (VP1u) and critical for virus entry via phospholipase A2 (vPLA2) domain, and VP2, which contributes to the viral capsid and is involved in virus-receptor binding [10,11].
Based on the phylogenetic analysis of the NS1-VP1u region, B19V was classified into three genotypes: genotype 1, genotype 2, and genotype 3 [12,13]. Genotype 1 is prevalent worldwide and can be divided into two subgroups (genotype1a and 1b); genotype 2 is commonly harbored in tissues of elderly people, but it is only sporadically detected as a circulating virus, while genotype 3 is only found in restricted areas, such as western Africa [14,15]. This last genotype shows a wider genetic variability in comparison with the other genotypes, and, for this reason, it may show a more limited geographical distribution.
This study is the first Italian report on the 2024 B19V outbreak; it presents an analysis of the epidemiological trends on B19V from January 2023 to September 2024 with a particular emphasis on the 2024 outbreak. Our aim is to present an epidemiological overview of positive cases and to genotypically characterize them.
2. Materials and Methods
2.1. Clinical Samples
B19V positive samples, processed for diagnostic routine, were collected at the Microbiology and Virology Unit of ‘S. Maria delle Scotte’ Hospital in Siena (Italy). The nucleic acid extraction was performed with the automatic extractor EZ1 and the DNA Blood kit (Qiagen GmbH, Germany). The quantity of the total nucleic acid was then assessed with Qubit DNA high sensitivity assay kit and Qubit 3.0 fluorometer (Thermo Fisher Scientific). Ethical approval was obtained from the local Ethical Committee for clinical trials (Protocol BIOBANCA VIROMICRO-2023, approval no. 25836, January 15th, 2024) (Comitato Etico Regionale per la Sperimentazione Clinica della Toscana - sezione AREA VASTA SUD EST) in terms of General Data Protection and Regulation (GDPR) upon written informed consent, signed by all subjects prior to participating in this study.
2.2. Serological and Molecular Analysis
Serological IgG analysis was carried out with the LIAISON™ Biotrin Parvovirus B19 IgG kit (Diasorin S.p.A.; Italy), according to the manufacturer’s instructions.
Real-time PCR was carried out with the Parvovirus B19 R-Gene™ kit (Biomerieux, Italy), according to the manufacturer’s instructions.
2.3. Next Generation Sequencing
Library preparation was carried on with the Viral Surveillance Panel (Illumina S.r.l. Milan, Italy) according to the manufacturer’s instructions. Libraries were then pooled and denatured according to Illumina’s instructions and charged on a V2 micro MiSeq 300 cycles flow cell (Illumina S.r.l. Milan, Italy). FastQ Files were analyzed with the BaseSpace™ platform by Illumina with the Dragen targeted microbial app (Illumina S.r.l. Milan, Italy). All obtained sequences were uploaded on Genbank (https://www.ncbi.nlm.nih.gov/nucleotide/) with the following accession numbers: PQ660663; PQ660664; PQ660665; PQ660666; PQ660667; PQ660668, PQ660669; PQ660670; PQ660671; PQ660672; PQ660673; PQ660674; PQ660675; PQ660676; PQ660677; PQ660678; PQ660679; PQ660680; PQ660681; PQ660682; PQ660683; PQ660684; PQ324621.1.
2.4. Phylogenetic Analysis
Sequences were aligned using the Geneious software with the MUSCLE algorithm. The phylogenetic tree was generated using the neighbor-joining algorithm. Bootstrapping and reconstitution were carried out with 1000 replicates to obtain the confidence level of the phylogenetic tree. Sequence homology comparison was carried out with reference sequences representing the three main B19V genotypes from the Gen Bank genetic sequence database: genotype1A: NC_00083.2; AF162273.1; genotype 1B: DQ357065.1; genotype 2: AY044266.2; AY064475.1; genotype 3: AY083234.1; AX003421.1.
3. Results and Discussion
From the beginning of 2023 to September 2024, a total of 550 requests for B19V diagnosis, both with serological and molecular methods, came to the attention of the Microbiology and Virology Unit at ‘S. Maria delle Scotte’ Hospital, based in Siena, Tuscany. Among them, 99 samples were positive for B19V (18%) of which, only three were collected in 2023 (two in April and one in October 2023).
Our data show an increased number of positive samples since the first months of 2024 (Figure 2), reaching the highest peak in late spring-early summer 2024, according to the reported scientific literature [9]. A higher number of female subjects tested positive (63 out of 99), while the remaining 36 were male subjects. Twenty-four samples were from pediatric patients (0-14 years old), whereas the remaining 75 samples were from adults (>15 years old).
This increase in the number of B19V positive samples was partially attributed to the impact of COVID-19 restrictions both on viral circulation and immunity patterns. Studies and reports highlight that strict public health measures during the pandemic, such as social distancing, school closures and reduced social interactions, significantly disrupted the transmission of various infectious diseases, including B19V. These measures led to a decline in population level immunity, particularly among children, who missed typical exposures to common pathogens [16]. Our study showed females were more affected than males. This may represent a problem, since non-immune pregnant women are at risk for fetal infections by B19V, leading to greater complications, especially if the infection occurs in the first or second trimester. It has been widely documented that Parvovirus infection during pregnancy can be passed to the fetus in 30-50% of cases, with a high risk to develop anemia, fetal hydrops and in some cases pre-birth death [17]. It is not known if pregnant women were present in our cohort, since clinical data were not available.
This high number of positive cases allowed us to characterize the circulating B19Vgenotype circulating in this outbreak.
As mentioned before, three different B19V genotypes were defined: genotype 1, divided in two subgroups (1a and 1b), genotype 2 and genotype 3. Although genotype 1a is the most frequent worldwide, it appears to have been partially substituted by genotype 2 in the last 50 years, but nowadays it is sporadically detected only in rare cases and usually in elderly people. Genotype 3 is less frequent and it is generally associated to peculiar, restricted areas [13,18].
For this purpose, 23 samples with PCR-Ct between 25 and 36 were selected for a Next Generation Sequencing (NGS) analysis. The following bioinformatic analysis with the Dragen Microbial Enrichment Plus software (IlluminaTM) identified B19V in 100% of sequenced samples. The mean coverage was 98.5% in all samples with a mean depth of 115X. The consensus sequences were generated and aligned with CLUSTAL and MUSCLE algorithm with a minimum GC content of 50%. All aligned sequences were used to conduct a phylogenetic analysis; the three genotype references were taken from Genbank (gen1a: AF162273.1; gen 1b: DQ357065.1; gen2: AY064475.1; AY044266.2; gen3: AX003421.1; AY083234.1). The phylogenetic tree was constructed basing on the neighbor-joining method with 1,000 Bootstrap repetitions (cutoff 50). The resulting phylogenetic tree (Figure 3) shows that all samples were clustered under genotype 1a and were closely related with each other (d=0.008; d.s=0.0062) with a distance of 0.147 against genotype 3, 0.128 against genotype 2 and 0.058 against genotype 1b; 0.2% of nucleotide divergence and 3% of amino acid divergence.
All detected mutations were point mutations, and the analysis revealed a broad polymorphic distribution, with frequencies ranging from 13% to 100%. Of these, 40.4% (59/146) were located in the NS1 gene, 8.9% (13/146) were exclusive to the VP1 gene, and the remaining 50.68% (74/146) in the VP2 gene, which shares its sequence with VP1, but differs by 227 amino acids (Supplementary material 1). These data (Table 1) demonstrate a greater variability in the VP1/VP2 genes according to the published literature [19].
Further studies are urgently needed to assess the distribution of these variants over time. The cohort must certainly be expanded to better understand the distribution and role of these variants in the long term. The translation of the sequences revealed the presence of non-synonymous mutations (listed in Table 2) within our cohort of samples. Among these changes, 5 were present in NS1 protein, 3 in VP1 and 3 in VP1/VP2.
It has been documented that variations in the VP1/VP2 capsid proteins may influence the virus interaction with host immune responses and cellular receptors, potentially enhancing its ability to evade immunity and spread more efficiently within the population [20]. These mutations were randomly dispersed among the analyzed genomes and never present in all samples. However, while most of the changes occurred among amino acids similar for physicochemical properties, the replacement of positively charged lysine with negatively charged (K14E) aspartic acid in VP1u could affect the protein. Indeed, this mutation is located in the N-terminal 1-80 aa of VP1u, which is rich in neutralizing epitopes [21]. In this regard, a deficient immune response to VP1u has been associated with persistent infections, emphasizing the important role of the immune response against VP1u in clearing the virus [22,23]. However, in this study, it was not possible to correlate this mutation with a more severe or prolonged disease, because samples were collected for diagnosis without knowing the subjects’ clinical status. The other recorded F554S amino acid change, localized in the carboxy-terminal part of NS1 gene, detected in 9/23 subjects, had previously been detected in other B19V isolates. This region, being highly polymorphic, was identified as exposed to positive selective pressure [25,26].
In conclusion, this is the first Italian report on 2024 B19V outbreak, including the genotyping of the current circulating strain. Although the genotype does not seem to have particularly changed, further studies, both on a larger court and on the protein conformation variability, could be needed to understand the possible impact of these mutations on proteins and their level of involvement in virus virulence and transmissibility.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. FigureS1: Illustration of the nucleotide variation frequencies detected in NS1, VP1 and VP2 genes.
Author Contributions
methodology, writing-original draft preparation, formal analysis, G.B.; conceptualization, validation, supervision, writing-review and editing, funding acquisition, M.G.C.
Funding
This study was supported by the Italian Ministry of Health- CUP B93C2200121000, CCM 2022 ‘SURVEID’.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the local Ethics Committee (Comitato Etico Regionale per la Sperimentazione Clinica della Toscana - sezione AREA VASTA SUD EST) (Protocol BIOBANCA VIROMICRO-2023, approval no. 25836, 15/01/2024) for studies involving humans.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Datasets analyzed or generated during the study can be provided, upon reasonable request, by contacting the corresponding author (mariagrazia.cusi@unisi.it).
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of this study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Servey, J.T.; Brian V Reamy B., V.; Hodge, J. Clinical presentations of parvovirus B19 infection. Am Fam Physician. 20007, 75, 373–376. [Google Scholar]
- Badrinath, A.; Gardere, A.; Samantha, L. Palermo S.L.; Campbell K.S.; and Kloc A. Analysis of Parvovirus B19 persistence and reactivation in human heart layers. Front. Virol. 2024, 4, 1304779. [Google Scholar] [CrossRef]
- Lehmann, T. , von Landenberg P.; Modrow S. Parvovirus B19 and Autoimmunity: A Review of Current Findings in Adult Infections. Autoimmun Rev. 2003, 2, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Dittmer, F.P.; De Moura Guimarães, C.; Alberto Borges Peixoto, A.; Monezi Pontes, K.F.; Bonasoni, M.P.; Tonni, G.; Araujo, E. Júnior. Parvovirus B19 Infection and Pregnancy: Review of the Current Knowledge. Journal of Personalized Medicine,.
- De Cnc Garcia, R.; Leon, L.A. Human Parvovirus B19: A Review of Clinical and Epidemiologial Aspects in Brazil. Future Microbiology 2021, 16, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Gallinella, G. Parvoviridae. In Encyclopedia of Infection and Immunity; Rezaei, N., Ed.; Elsevier: Oxford, UK, 2022; pp. 259–277. [Google Scholar]
- Enders, M.; Weidner, A. & Enders, G. Current epidemiological aspects of human parvovirus B19 infection during pregnancy and childhood in the western part of Germany. Epidemiol. Infect, 2007, 135, 563–569. [Google Scholar]
- Vyse, A. J.; Andrews, N. J.; Hesketh, L. M. & Pebody, R. The burden of parvovirus B19 infection in women of childbearing age in England and Wales. Epidemiol. Infect. 2007, 135, 1354–1362. [Google Scholar]
- d’Humières, C.; Fouillet, A.; Verdurme, L. ; Lakoussan S-B.; Gallien Y.; Coignard C.; Hervo M.; Ebel A.; Visseaux B.; Maire B.; et al. An unusual outbreak of parvovirus B19 infections, France, 2023 to 2024. Euro Surveill. 2400, 29, pii=2400339. [Google Scholar]
- Ganaie Safder, S. , Qiu J. Recent Advances in Replication and Infection of Human Parvovirus B19. Front Cell Infect Microbiol. 2018, 8, 166. [Google Scholar]
- Yong Luo, Y.; Jianming Qiu, J. Human parvovirus B19: a mechanistic overview of infection and DNA replication. Future Virol. 2015, 10, 155–167. [Google Scholar]
- Servant, A.; Laperche S,; Lallemand F. ; Marinho V.; De Saint Maur G.; J.F.; Garbarg-Chenon A. Genetic diversity within human erythroviruses: identification of three genotypes. J. Virol. 2002, 76, 9124–9134. [Google Scholar] [CrossRef]
- Hübschen, J.M.; Mihneva, Z.; Mentis, A.F.; Schneider, F.; AboudyY. ; Grossman Z.; Rudich H. et al. Phylogenetic analysis of human parvovirus B19 sequences from eleven different countries confirms the predominance of genotype 1 and suggests the spread of genotype 3b. J. Clin. Microbiol. 2009, 47, 3735–3738. [Google Scholar] [CrossRef] [PubMed]
- Gallinella, G.; Venturoli, S.; Manaresi, E.; Musiani, M.; Zerbini, M. B19 virus genome diversity: Epidemiological and clinical correlations. J. Clin. Virol. 2003, 28, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Eis-Hübinger, A.M.; Reber, U.; Edelmann, A.; Kalus, U.; Hofmann, J. Parvovirus B19 genotype 2 in blood donations. Transfusion. 2014, 54, 54,1682–1684. [Google Scholar] [CrossRef] [PubMed]
- Russcher, A.; van Boven, M.; Benincà, E.; Verweij, E.J.T.; M. W.A.; Zaaijer H.L.; Vossen A. C. T. M.; Kroes A.C.M. Changing epidemiology of parvovirus B19 in the Netherlands since 1990, including its re-emergence after the COVID-19 pandemic. Sci Rep. 2024, 14, 9630. [Google Scholar] [CrossRef]
- de Jong, E.P. , Walther, F.J., Kroes, A.C.M. and Oepkes, D. Parvovirus B19 infection in pregnancy: new insights and management. Prenat. Diagn., 2011, 31, 419–425. [Google Scholar] [CrossRef]
- Jain, A.; Kant, R. Genotypes of erythrovirus B19, their geographical distribution & circulation in cases with various clinical manifestations. Indian J. Med. Res. 2018, 147, 239–247. [Google Scholar]
- Karlin, D.G. Parvovirus B19 and Human Parvovirus 4 Encode Similar Proteins in a Reading Frame Overlapping the VP1 Capsid Gene. Viruses 2024, 16, 191. [Google Scholar] [CrossRef]
- Saikawa, T.; Anderson, S.; Momoeda, M.; Kajigaya, S.; Young, N.S. Neutralizing linear epitopes of B19 parvovirus cluster in the VP1 unique and VP1-VP2 junction regions. J. Virol, 1993, 67, 3004–3009. [Google Scholar] [CrossRef]
- Anderson, S.; Momoeda, M.; Kawase, M.; Kajigaya, S.; Young, N.S. Peptides derived from the unique region of B19 parvovirus minor capsid protein elicit neutralizing antibodies in rabbits. Virology 1995, 206, 626–632. [Google Scholar] [CrossRef]
- Kurtzman, G.J.; Cohen, B.J.; Field, A.M.; Oseas, R.; Blaese, R.M.; Young, N.S. Immune response to B19 parvovirus and an antibody defect in persistent viral infection. J. Clin. Investig. 1989, 84, 1114–1123. [Google Scholar] [CrossRef]
- Kurtzman, G.J.; Cohen, B.J.; Field, A.M.; Oseas, R.; Blaese, R.M.; Young, N.S. Immune response to B19 parvovirus and an antibody defect in persistent viral infection. J. Clin. Investig. 1989, 84, 1114–1123. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, G.; Frickhofen, N.; Kimball, J.; Jenkins, D.W.; Nienhuis, A.W.; Young, N.S. Pure Red-Cell Aplasia of 10 Years’ Duration Due to Persistent Parvovirus B19 Infection and Its Cure with Immunoglobulin Therapy. New Engl. J. Med. 1989, 321, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Stamenković, G.G; Ćirković, V.S.; Šiljić, M.M.; Blagojević, J.V.; Aleksandra, M. Knežević A.M.; Joksić I.D. & Stanojević M.P. Substitution rate and natural selection in parvovirus B19. Sci. Rep. 2016, 6. [Google Scholar]
- Haoran, J.; Qi, Q.; Yangzi, Z.; Yan, Z.; Wenbo X; Cui, Aili; XiaomeL. The epidemiological and genetic characteristics of human parvovirus B19 in patients with febrile rash illnesses in China. Sci. Reports, 2023, 13, 15913. [Google Scholar]
Figure 1.
A representation of B19V genome. The 5596nt genome is composed by 5’ and 3’ inverted terminal repeats, and 3 main genes: the non-structural protein1 (NS1), the viral protein 1 (VP1) and the viral protein 2 (VP2). The two proteins share the whole sequence of VP2, but VP1 has an additional N-terminal domain known as VP1 unique region (VP1u).
Figure 1.
A representation of B19V genome. The 5596nt genome is composed by 5’ and 3’ inverted terminal repeats, and 3 main genes: the non-structural protein1 (NS1), the viral protein 1 (VP1) and the viral protein 2 (VP2). The two proteins share the whole sequence of VP2, but VP1 has an additional N-terminal domain known as VP1 unique region (VP1u).
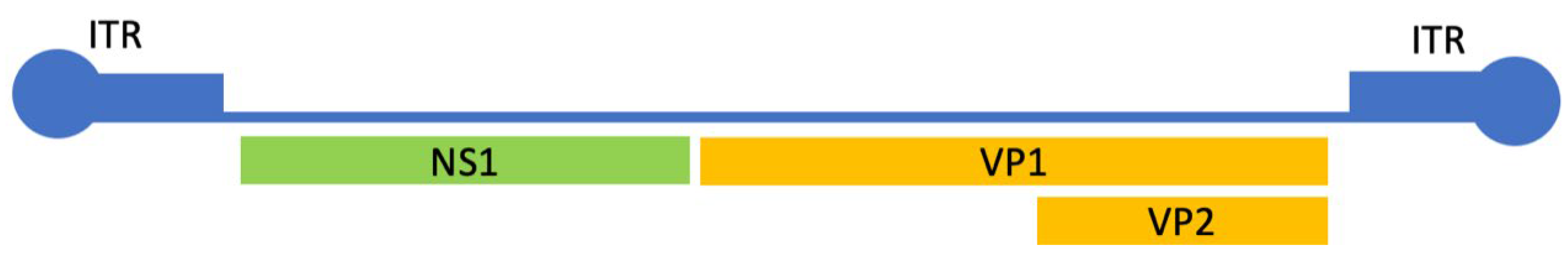
Figure 2.
Parvovirus B19 cases from 2023 to the end of September 2024 in Siena area, Italy. A) Number of positive samples distributed by gender; blue bars are from males and purple bars from females. B) Number of positive samples distributed by age; green bars represent adult patients (>15 years old), yellow bars correspond to children.
Figure 2.
Parvovirus B19 cases from 2023 to the end of September 2024 in Siena area, Italy. A) Number of positive samples distributed by gender; blue bars are from males and purple bars from females. B) Number of positive samples distributed by age; green bars represent adult patients (>15 years old), yellow bars correspond to children.
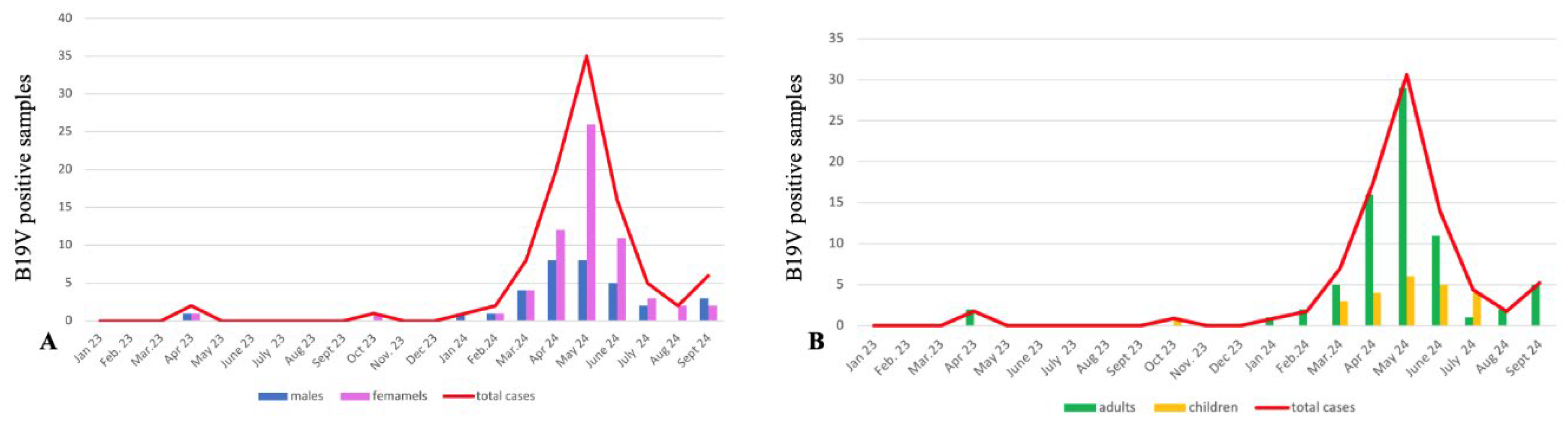
Figure 3.
Phylogenetic tree of NGS samples. The phylogenetic tree shows that all samples were clustered under the genotype1a. No samples were clustered under the genotype1b, genotype2 and genotype3. The used references are highlighted in bold, all of them are accessible in the Genbank database. Genotype 1A: NC_00083.2; AF162273.1; genotype 1B:DQ357065.1; genotype 2: AY044266.2; AY064475.1; genotype 3: AY083234.1; AX003421.1.
Figure 3.
Phylogenetic tree of NGS samples. The phylogenetic tree shows that all samples were clustered under the genotype1a. No samples were clustered under the genotype1b, genotype2 and genotype3. The used references are highlighted in bold, all of them are accessible in the Genbank database. Genotype 1A: NC_00083.2; AF162273.1; genotype 1B:DQ357065.1; genotype 2: AY044266.2; AY064475.1; genotype 3: AY083234.1; AX003421.1.
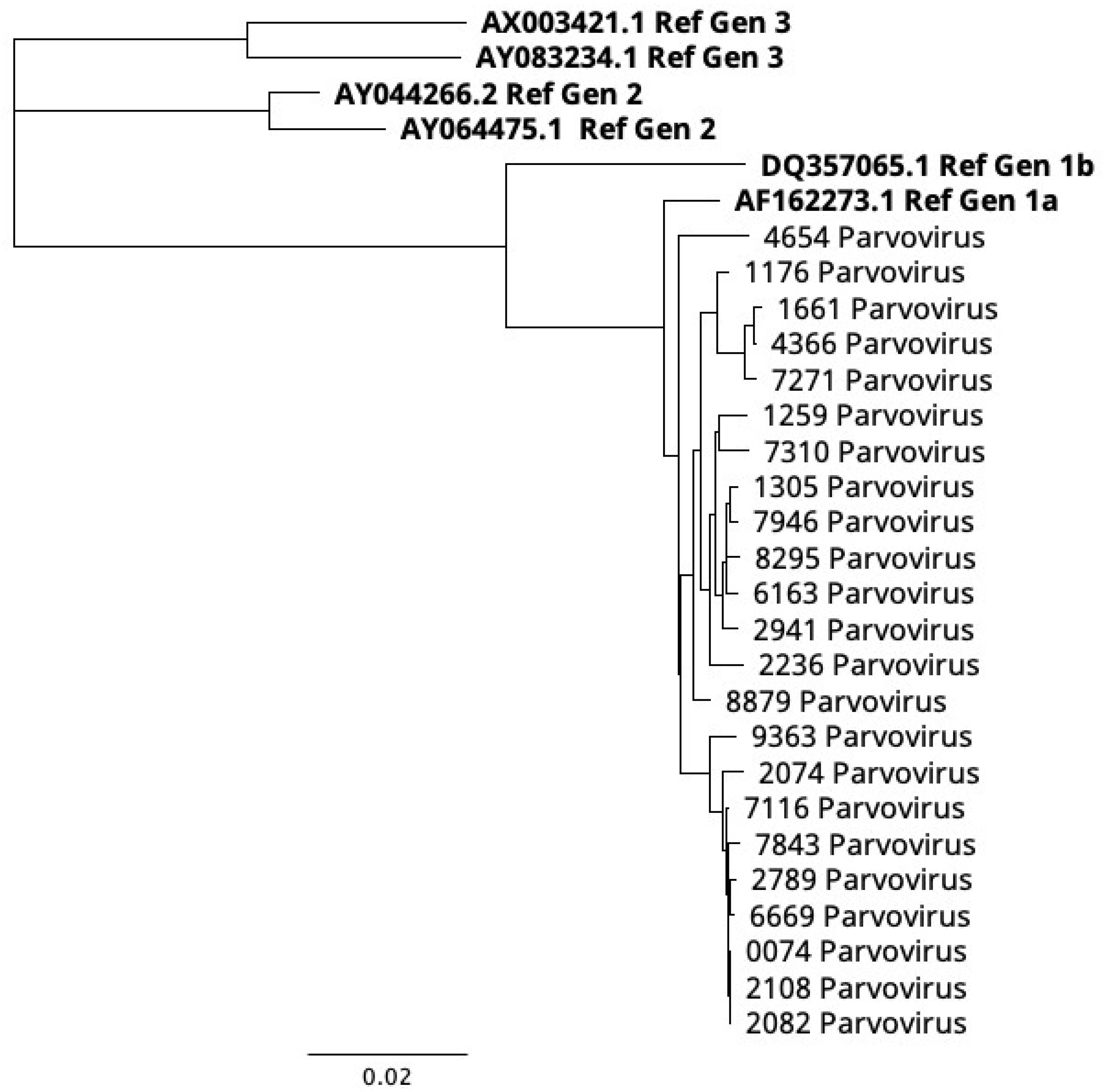

Table 1.
The nucleotide mutations present in all samples analyzed with the NGS approach.
| Gene | Mutations |
|---|---|
| NS1 | 803G>A; 1175 C>T; 1227 A>T, 1430 A>T; 1140 C>T; 1529A>G; 1928 A>C; 2286 T>C |
| VP1/VP2 | 2754 C>T; 3351T>G; 3444 T>A; 3489 C>T; 3531 A>C; 3752 C>G; 3795 A>G; 3894 C>T; 4264C>T; 43069A>T; 4345 T>C; 4346 C>A. |
Table 2.
List of amino acid mutations and their frequency in the B19V sequences.
| Gene | Mutation | Frequency |
|---|---|---|
| NS1 | C17S | 9/23 |
| NS1 | F57L | 4/23 |
| NS1 | V71A | 5/23 |
| NS1 | L111F | 6/23 |
| NS1 | F554S | 9/23 |
| VP1u | K14E | 7/23 |
| VP1 | V30L | 18/23 |
| VP1u | S98N | 17/23 |
| VP1/VP2 | A260T-A33T | 6/23 |
| VP1/VP2 | N533S-N306S | 17/23 |
| VP1/VP2 | G537A-G310A | 9/23 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



