
Submitted:
13 January 2025
Posted:
14 January 2025
You are already at the latest version
Abstract
ZADT is a significant oil sunflower variety in Northwest China, renowned for its robust resistance to salt and drought. However, the lack of an optimal genetic transformation system has greatly hindered the elucidation of its superior traits and the advancement of molecular breeding in sunflowers. Establishing a straightforward and efficient transient transformation system could effectively address these limitations. We utilized Agrobacterium tumefaciens to transform the backbone parent ZADT retention line. The sunflower transient expression system was successfully established and optimized using infiltration, injection, and ultrasound-vacuum methods. Among these methods, an Agrobacterium growth concentration of OD600 0.8 and the surfactant Silwet-77 favored transient transformation. In the infiltration method, the highest transformation efficiency was achieved after 2 hours of infiltration. For the injection method, the optimal conditions were determined to be when seedlings were grown for 4-6 days and kept in dark culture for 3 days. The ultrasound-vacuum method was most effective with 40 kHz ultrasound for 1 minute followed by 0.05 kPa vacuum filtration for 5-10 minutes. Gene expression in the sunflower transient system was maintained for at least 1 week using all three methods. Additionally, the infiltration method was applied to preliminarily identify the salt tolerance function of the genes HaMYB108 and HaWRKY40 in sunflower. A highly efficient transient transformation platform for Agrobacterium-mediated ZADT was successfully established using three distinct methods. This platform was subsequently employed to preliminarily confirm the functions of the candidate gene. It offers significant support for conducting molecular biology studies such as gene function analysis, subcellular localization, gene interactions, and ChIP-seq.
Keywords:
oil sunflower
; the retention line of backbone parent ZADT
; transient transformation
; infiltrationi method
; injection method
; ultrasonic-vacuum method
; condition optimisation
1. Introduction
The sunflower is an important oil crop, contributing 9.0% to global edible oil production [1]. It is also highly salt-tolerant and widely cultivated [2]. However, sunflowers still face numerous challenges, including harsh abiotic and biotic stresses, and low yield. Therefore, improving sunflower performance through breeding is essential, particularly by enhancing stress tolerance, oil production quality, and yield. Compared to traditional breeding, molecular breeding offers the advantages of simpler operation and shorter development time for high-quality sunflower varieties.
Identification of gene function is essential for pinpointing important trait genes and represents a crucial step in molecular breeding. Gene function identification within the context of plant ontogeny is considered more reliable than through heterologous systems. Both stable and transient transformation techniques can be used for gene function identification. However, the stable transformation system in many plants is hampered by regeneration difficulties and low transformation rates [3]. For instance, despite over 30 years of research, a fully implemented stable transformation system for sunflower remains elusive [4]. In contrast, transient transformation techniques can identify gene functions in a short period without requiring prolonged sterile growth conditions [5,6,7]. With the advancement of transient transformation systems, reported methods now include Agrobacterium-mediated gene transformation, particle bombardment, electroporation, and polyethylene glycol-mediated transformation [8,9,10,11,12,13]. Agrobacterium-mediated gene transformation is widely utilized due to its have high gene transfer efficiency, simplicity and cost-effectiveness compared to other methods. Yucheng Wang’s team has developed an innovative Agrobacterium-mediated transient transformation-based gene expression system applicable to herbaceous plants (e.g., tobacco and Arabidopsis thaliana) and trees (e.g., birch, poplar, tamarisk, cork, willow, and aralia) [14]. This technology has been successfully applied to Osmanthus fragrans [15], Arabidopsis thaliana [16], and Brassica napus [17]. Therefore, it is necessary and feasible to establish a transient transformation system in sunflower.
Functional characterization of genes has been achieved using transient transformation systems. The established safflower transient expression system has successfully analyzed the function of genes involved in flavonoid biosynthesis [18]. This system also demonstrated that the transient expression of the CiDREB1C gene enhances tolerance to drought and salt stress while reducing sensitivity to ABA in Caragana intermedia [19]. Additionally, transient overexpression and gene silencing of SgCPR1 and SgCPR2 revealed their association with the synthesis of secondary metabolites such as mogrol in S. grosvenorii [20]. Consequently, establishing a simple and efficient transient transformation system holds significant value for crop breeding.
In this study, we established an efficient and versatile Agrobacterium-mediated transient transformation system for ZADT using GUS as the reporter gene. The salt tolerance functions of the HaMYB108 and HaWRKY40 genes were preliminarily identified using the infiltration method. This transient transformation system is advantageous for identifying key gene functions in sunflower and provides essential materials for molecular breeding, thereby advancing the field of plant molecular biology.
2. Results
2.1. Optimisation of Optimal Conditions for the Infiltration Method in the ZADT Transient Transformation System
We evaluated and optimized the transformation conditions for three infiltration methods: surfactant type, infiltration time, and Agrobacterium growth concentration. Firstly, optimizing the surfactant type revealed that Silwet L-77 infiltrated seedlings exhibited deeper GUS staining, a 44.44% higher relative expression of GUS genes compared to Triton X-100, and a transformation efficiency as high as 90% (Figure 1a-b). Secondly, prolonged immersion of seedlings in the infiltration solution, with sucrose as the main component, caused tissue and organ damage, particularly to the root system. The results indicated the roots were partially necrosis after 4 hours of immersion and complete necrosis after 6-8 hours (Figure 1e-f). Seedlings showed light GUS staining and the lowest relative gene expression when immersed for 1 hour. Taken together, the optimum infiltration time for ZADT was determined to be 2 hours (Figure 1e-h). Finally, the penetration capacity of Agrobacterium varied at different growth phases, necessitating optimization of the concentration conditions for Agrobacterium growth. At OD600 0.4, GUS staining was lightest, the relative expression of the GUS gene was lowest, and the transformation efficiency was only 60%. In contrast, at OD600 0.8, the infiltration effect was optimal, with a 62.96% increase in the relative expression of the GUS gene (Figure 1i-l). In conclusion, using Silwet L-77 as the surfactant, an infiltration time of 2 hours, and an OD600 0.8 were determined to be the optimal conditions for the infiltration method.
2.2. Optimal Conditions for Screening Injection Methods
The injection method involves directly injecting the Agrobacterium suspension into the plant using a syringe. Four conditions were optimized: surfactant type, Agrobacterium growth concentration, seedling growth time, and dark incubation time. Firstly, it was determined that the highest transformation efficiency was achieved using Silwet L-77 as the surfactant and an Agrobacterium growth concentration of OD600 0.8, which was similar to the optimization results of the infiltration method (Figure 2a, d-h). Among these conditions, the GUS staining was most intense at OD600 1.2, with a 72.92% increase in relative GUS expression compared to OD600 0.4. However, the cotyledon injection area was almost necrotic. Therefore, OD600 0.8 was concluded to be the optimal condition (Figure 2d, g-h). Secondly, cotyledon freshness varies across different growth stages, leading to varying effects of infestation. Cotyledons that had unfolded for 2 days were the least effective for injection due to the closely-walled epidermis and mesophyll adherence. Seedlings at 4-6 days exhibited improved infiltration, with deep GUS staining and an average increase of 88.0% in relative GUS gene expression in both groups (Figure 2b, k-l). Finally, the dark incubation time was optimized, revealing that injected sunflower seedlings required a few days of dark incubation to promote the expression of exogenous genes carried by Agrobacterium. One day of dark culture was insufficient, as indicated by light GUS staining and low relative gene expression levels. After 5 days of dark culture, the cotyledons became necrotic due to prolonged ‘starvation’ and excessive Agrobacterium infestation. Thus, 3 days of dark culture was determined to be optimal, and the relative expression of GUS genes was increased by 56.5% compared with 1 day of dark culture (Figure 2c, i-j). In conclusion, the optimal conditions for the injection method were Silwet L-77 as the surfactant, 4-6 days of seedling growth, OD600 0.8, and 3 days of dark culture.
2.3. Transient Conversion of ZADT Using Ultrasonic Vacuum Method
The ultrasound-vacuum method employs an ultrasonic cleaner to create micro-wounds and vacuum filtration to facilitate Agrobacterium infiltration into ZADT. Initially, ZADT seedlings exhibited an 87.1% increase in relative GUS gene expression at OD600 0.8 compared to OD600 0.4 (Figure 3a-d). Subsequently, ultrasonic treatment caused significant damage to the seedlings, with extensive necrosis of cotyledons observed at 3 minutes of treatment (Figure 3e-h). Vacuum treatment enhanced the uptake of Agrobacterium by the plant. Transiently transformed sunflower seedlings exhibited 100% transformation efficiency and the most intense GUS staining after 5-10 minutes of filtration. Relative gene expression was increased by 66.67% compared to the 1 minute treatment. (Figure 3i-l). In conclusion, OD600 0.8, ultrasonic treatment for 1 minute, and vacuum treatment for 5-10 minutes were identified as the optimal conditions for the ultrasound-vacuum method.
2.4. Three Methods to Express the Duration of the ZADT Transient Transformation System
The ZADT transient transformation expression system was further evaluated. Firstly, prolonged exposure of hydroponically grown seedlings to low-light conditions using the infiltration method can result in unfavorable elongation of seedling embryonic axes, affecting subsequent experiments. Therefore, seedlings infiltrated with bacterial strains were analyzed for GUS staining 2-6 days post-infiltration. It was observed that GUS expression remained high from days 2 to 4 (Figure 4a). Additionally, GUS staining results from days 3 to 9 were assessed using both the injection method and the ultrasonic vacuum method. GUS expression in both methods remained elevated from days 3 to 5, persisting until day 9 (Figure 4b, c).
2.5. Preliminary Functional Validation of Salt Tolerance Candidate Genes Was Conducted Through Transient Transformation of ZADT Using the Infiltration Method
In the previous stage, we identified two potential sunflower salt tolerance candidate genes, HaMYB108 and HaWRKY40, through transcriptomics (data not published). Preliminary rapid identification of gene function was performed using the infiltration method in the ZADT seedling transient transformation system. This method offers the advantages of ease of operation and whole-seedling infiltration, compared to the injection and ultrasonic vacuum methods. Under untreated conditions, the phenotypes of pBI121 empty vector (1 and 3) and overexpressed seedlings (2 and 4) remained unchanged (Figure 5a). qRT-PCR analysis revealed that the relative expression of HaMYB108 and HaWRKY40 was approximately 5-fold higher than that of the empty vector (EV) after 36 hours of infiltration. The highest difference in relative expression of the two genes was observed at 72 hours, with a 15-fold increase compared to EV. The expression remained higher than that of EV at 96 hours (Figure 5b). Subsequently, it was confirmed that the overexpression of HaMYB108 and HaWRKY40 genes was translated and expressed in ZADT. Protein expression was detected at 72 hours of stress treatment by Western Blot (Figure 5d). Therefore, 350 mM salt stress treatment was performed 72 hours after infiltration. The cotyledons of EV groups showed extensive blackening and death, while seedlings overexpressing HaMYB108 and HaWRKY40 genes remained highly active and exhibited a distinct salt-tolerant phenotype (Figure 5c). These results indicate that the candidate genes’ functions were preliminarily verified using the ZADT transient transformation technique.
3. Discussion
Agrobacterium-mediated genetic transformation is influenced by various factors, including the methods of treatment [22], Agrobacterium concentration [23], and infection time [24], and so on. In this study, we examined the effects of three treatments—immersion method, injection method, and ultrasound-vacuum method—on the transient transformation of ZADT. In order to obtain a transient transformation system with high transformation rate and high expression of target genes in ZADT seedlings, three methods were optimized. First, Agrobacterium density and infestation time directly determine the transient transformation efficiency of plants [25]. Agrobacterium tumefaciens growth concentration OD600 0.9 with 5 d of co-culture has been reported for Gossypium hirsutum transient transformation [20]; Maesa Lanceolata [26] OD600 1.0 with 5 d after infection was the optimal time for co-culture for gene function study; Paeonia lactiflora [27] determined that the concentration of the bacterial solution and the time of infiltration was OD600 1.2 with 3 d of co-culture; and for Pinus tabuliformis [28] it was OD600 0.8, with a co-cultivation for 3d, which is consistent with our results. We found that Agrobacterium growth concentration OD600 0.8 (Figure 1ij, Figure 2d, Figure 3ab), co-cultivated for 3 d (Figure 2c) was favorable for the three transient transformation systems of ZADT. Surfactants have been found to significantly reduce the surface tension at different air-liquid interfaces, thereby facilitating Agrobacterium invasion and increasing plant cell permeability. A study revealed that the use of 0.04% Silwet L-77 facilitated the transient transformation of Cinnamomum camphora [29]. In cotton transient transformation compared Triton-20 and Silwet L-77, and it was found that Silwet L-77 significantly induced GUS gene expression [20]. There are also experimental results that suggest Triton X-100 or Tween-20 are more suitable for the plants they studied, such as Artemisia annua [30] and Paeonia lactiflora [28]. Our study found that infestation of ZADT required the addition of 0.02% Silwet L-77 to the Agrobacterium suspension (Figure 1ab, Figure 2a).
Among the optimisation factors specific to the three transient transformation methods, the infiltration method utilises a high concentration of infiltration solution for a long period of time to immerse seedlings, which can lead to complete necrosis of the roots not conducive to transformation. 2 h is the optimal time for infiltration of ZADT seedlings (Figure 1ef); and the length of plant growth significantly affects transiently transformed plants in the injection method [31]. ultrasound [28] and vacuum [32] could significantly increase the efficiency of transient transformation. ultrasound creates a slight wound on the surface of the seedling, and vacuum filtration can create negative pressure to press Agrobacterium into the plant, resulting in a significant increase in transient expression rate [33]. This was confirmed in the establishment of transient transformation systems in Diospyros kaki Thunb [34] and Medicago sativa [35]. Our results showed that sonication for 1 min and vacuum treatments for 5-10 min significantly improved the transient transformation efficiency of ZADT (Figure 3ef, ij).
Plant transient expression usually peaks at 2–4 d of infection, followed by a decrease in the number of expressing cells and the level of expression per transformed cell. However, this kinetic trend ultimately depends on the host plant genotype [36]. We observed that the transcript levels of genes expressed in ZADT peaked at 2–5 d of infection in vivo. Among them, GUS gene expression in ZADT was high in vivo at 2–4 d post-infection using the infiltration method, and at 3–5 d post-infection using the injection and ultrasound-vacuum methods (Figure 4). This period can be utilized for subsequent studies such as gene function.
Transient transformation systems have been reported to be applied in various pathways. Tobacco transient transformation technology is commonly used for gene subcellular localization and gene interaction validation experiments [37]. The subcellular localization of short hair cotton genes and their corresponding proteins was investigated using a transient expression system [38]. The Agrobacterium- mediated transient transformation method successfully detected the efficiency of sgRNA in the CRISPR systems of sorghum and birch [39,40]. Artemisia annua used the ChIP-seq transient expression system of AaHY5 (a positive regulator of artemisinin biosynthesis) to identify AaWRKY14 as a potential transcription factor linking AaHY5 to the regulation of artemisinin biosynthesis genes [41]. Tamarix hispida transiently transformed with the ThSCL32 (SCARECROW-like 32) gene was identified by RNA-seq and ChIP-PCR as a transcription factor involved in salt tolerance through enhanced expression of ThPHD3 (prolyl-4-hydroxylase domain 3 protein) [42]. We functionally phenotyped the salt tolerance genes HaMYB108 and HaWRKY40 using the ZADT transient transformation system established by Agrobacterium infiltration, and preliminarily demonstrated their resistance function under salt stress (Figure 6). This indicates that transient transformation technology has gained increasing applications in biological research.
4. Materials and Methods
4.1. Preparation of Experimental Materials
Seeds of the backbone parent line ZADT were provided by the Institute of Economic Crops at the Xinjiang Academy of Agricultural Sciences. ZADT was transformed using Agrobacterium strain EHA105 carrying the pBI121-GUS vector.
4.2. Agrobacterium-Mediated Transient Transformation of Sunflower
The Agrobacterium-mediated transient transformation system for sunflower comprises three methods: infiltration, injection, and ultrasonic-vacuum. The primary experimental procedures include seedling preparation, preparation of the infiltration solution, bacterial infection, and culture. The detailed workflow is illustrated in Figure 6.
4.2.1. Infiltration
The infiltration method was evaluated and optimized for three factors: surfactant type (Silwet L-77 and Triton X-100), Agrobacterium growth concentration (OD600 of 0.4, 0.8, and 1.2), and infiltration time (1, 2, 4, 6, and 8 hours). To assess the relative expression of the candidate genes HaMYB108 and HaWRKY40 in transiently transformed seedlings, samples were collected at 36, 72, and 96 hours post-infiltration. The samples were collected in liquid nitrogen and stored in a -80℃ freezer.
4.2.2. Cotyledonary Injection
The cotyledonary injection method was evaluated and optimized for four factors: surfactant type (Silwet L-77 and Triton X-100), Agrobacterium growth concentration (OD600 0.4, 0.8, 1.2), seedling growth duration (5, 7, 9 days), and dark incubation period (1, 3, 5 days).
4.2.3. Ultrasonic-Vacuum Infiltration
The ultrasonic-vacuum method was evaluated and optimized by examining three factors: Agrobacterium growth concentration (OD600 0.4, 0.8, 1.2), sonication time (1, 3, 5 minutes), and vacuum time (1, 5, 10 minutes).
A univariate experimental design was employed to assess the instantaneous conversion efficiency of these factors. Each experiment was replicated at least three times with ten seedlings per replication. Conversion efficiency = number of GUS stained seedlings / total number of seedlings.
4.3. Detection of Transient Transformation Efficiency and Duration
After conducting the three infestation methods, the sunflower seedlings were transferred to their respective environmental conditions for cultivation. The seedlings were then subjected to GUS staining and counted. The infiltration method was observed and counted every 2 days until day 6. The injection method and ultrasonic-vacuum method were observed and counted every 3 days until day 9.
4.4. β-glucuronidase (GUS) Staining
Seedlings were stained at 100 rpm in a shaker at 37 ℃ for 16 hours in darkness. The staining solution was then washed away with water. The samples were rinsed with 50%, 70%, and 100% ethanol for 5 minutes each, and the excess liquid was discarded. The samples were subsequently soaked in 70% ethanol at 75 ℃ until all chlorophyll was removed. Sites stained blue indicate GUS expression.
4.5. qRT-PCR Experiment
Total RNA was extracted from 100 mg of sample using the Plant RNA Kit (Omega BioTek, USA), following the manufacturer’s instructions. After DNase treatment, RNA was reverse transcribed into cDNA using SuperScript™ II Reverse Transcriptase (Takara, Japan) and a poly(dT)18 primer. qRT-PCR was conducted on a CFX96 Touch Real-Time PCR System (Bio-Rad, USA) with three biological replicates per sample. HaEF-1 and HaActin served as internal reference genes. Relative gene expression was calculated using the 2-ΔΔCT method [21]. Primer information for all genes used in the qRT-PCR experiments is provided in Supplementary Table 1.
4.6. Construction of Recombinant Plasmids
To verify the function of the ZADT candidate gene using the Agrobacterium-mediated transient transformation method, the coding sequences (CDS) of the candidate genes HaMYB108 (HanXRQr2_Chr17g0827521) and HaWRKY40 (HanXRQr2_Chr17g0785961) were cloned from the cDNA of ZADT roots. The ORF regions of these two genes were inserted downstream of the CaMV35S promoter in the binary expression vector pCAMBIA2300-GFP (the reorganization vector was purchased from Wuhan Transduction Biological Laboratory Co.), and recombinant plasmids HaMYB108::GFP and HaWRKY40::GFP were constructed through homologous recombination. The primers used for vector construction are listed in Supplementary Table 1.
4.7. Plant Protein Extraction and Western Blotting Experiments
Total protein containing GFP, GFP-HaMYB108, or GFP-HaWRKY40 was extracted from ZADT tissues using the P-PER® Plant Protein Extraction Kit (Thermo Fisher Scientific, USA). The blot was incubated overnight at 4°C with a 1:500 dilution of monoclonal mouse antibodies against GFP and β-actin (Abcam, China). The goat anti-mouse IgG secondary antibody conjugated to peroxidase was used at a 1:5000 dilution for incubation at room temperature. The development and fixation were performed using the standard procedure with SuperSignal® West Dura Extended Duration Substrate (Thermo Fisher Scientific, USA).
4.8. Data Analysis and Graph Construction
GraphPad Prism 8.0.2 and SPSS statistical software were utilized for data analysis. Experimental results were analyzed using one-way analysis of variance (ANOVA). Treatment differences were presented as mean ± standard deviation (SD) and compared at p<0.05 using Tukey’s least significant difference (LSD) test. Image processing was conducted using Adobe Photoshop software.
5. Conclusions
An Agrobacterium-mediated transient transformation system for sunflower was established and optimised using three different methods: infiltration, injectionand ultrasonic vacuum, with a transformation efficiency of 90-100%. The gene expression time of this transient transformation system was at least 1 week. It can be used for molecular biology research such as gene function identification, subcellular localisation, gene interactions and Chip-seq to find downstream regulatory genes.
Funding
This study was financially supported by Major Special Project 4 of Xinjiang Uygur Autonomous Region (No.2022A03004-4), the Natural Science Foundation of Xinjiang Uygur Autonomous Region (No. 2020D01C020), and the Research and Innovation Program for Excellent Doctoral Students at Xinjiang University (XJU2022BS050).
References
- Temme, A.A.; Kerr, K.L.; Masalia, R.R.; Burke, J.M.; Donovan, L.A. Key traits and genes associate with salinity tolerance independent from vigor in cultivated sunflower. Plant Physiol. 2020, 184(2), pp.00873.
- Kumar, A.P.K.; et al. Genetics, genomics and breeding of sunfower. Lewes, Delaware : Excelic Press LLC, 2019.
- Su, W.B.; Xu, M.Y.; Radani, Y.; Yang, L.M. Technological development and application of plant genetic transformation. Int J Mol Sci. 2023, 24(13), 10646. [Google Scholar] [CrossRef] [PubMed]
- Darqui, F.S.; Radonic, L.M.; Beracochea, V.C.; Hopp, H.E.; Bilbao, L.M. Peculiarities of the transformation of Asteraceae family species: The cases of sunflower and lettuce. Front Plant Sci. 2021, 12, e767459. [Google Scholar] [CrossRef] [PubMed]
- Li, M.X.; Wu, Q.Q.; Guo, F.Q.; Ouyang, Y.Z.; Ao, D.Y.; You, S.J.; Liu, Y.Y. A versatile, rapid Agrobacterium-mediated transient expression system for functional genomics studies in cannabis seedling. Planta. 2024, 260, 18. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.J.; Niu, C.; Xie, Q.; Xing, Q.; Qi, H. The recent advances of transient expression system in horticultural plants. Acta Hortic Sin. 2017, 44(9), 1796–1810. [Google Scholar]
- Tyurin, A.A.; Suhorukova, A.V.; Kabardaeva, K.V.; Goldenkova-Pavlova, I.V. Transient gene expression is an effective experimental tool for the research into the fine mechanisms of plant gene function: advantages, limitations, and solutions. Plants. 2020, 9(9), 1187–1206. [Google Scholar] [CrossRef] [PubMed]
- Leissing, F.; Reinstdler, A.; Thieron, H.; Panstrega, R. Gene gun-mediated transient gene expression for functional studies in plant immunity. Methods Mol Biol. 2022, 2523, 63–77. [Google Scholar]
- Wang, Y.; Zhang, Y.; Dong, Y.; Li, D.L.; Shi, S.L.; Li, S.H.; Li, L.Z.; He, Y.J.; Li, J.Y.; Chen, H.Y.; et al. A highly efficient mesophyll protoplast isolation and PEG-mediated transient expression system in eggplant. Sci Hortic. 2022, 304, 111303. [Google Scholar] [CrossRef]
- Gou, Y.J.; Li, Y.L.; Bi, P.P.; Wang, D.J.; Feng, J.Y. Optimization of the protoplast transient expression system for gene functional studies in strawberry (Fragaria vesca). Plant Cell Tissue Organ Cult. 2020, 141, 41–53. [Google Scholar] [CrossRef]
- Ren, R.; Gao, J.; Lu, C.Q.; Wei, Y.L.; Jin, J.P.; Wong, S.M.; Zhu, G.F.; Yang, F.X. Highly efficient protoplast isolation and transient expression system for functional characterization of flowering related genes in cymbidium orchids. Int J Mol Sci. 2020, 21(7), 2264. [Google Scholar] [CrossRef] [PubMed]
- Zottini, M.; Barizza, E.; Costa, A.; Formentin, E.; Ruberti, C.; Carimi, F.; Schiavo, F.L. Agroinfiltration of grapevine leaves for fast transient assays of gene expression and for long-term production of stable transformed cells. Plant Cell Rep. 2008, 27(5), 845–853. [Google Scholar] [CrossRef]
- Tsuda, K.; Qi, Y.P.; Nguyen, L.V.; Bethke, G.; Tsuda, Y.; Glazebrook, J.; Katagiri, F. An efficient Agrobacterium-mediated transient transformation of Arabidopsis. Plant J. 2012, 69, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Liu, G.F.; Meng, X.N.; Li, Y.B.; Wang, Y.C. A versatile Agrobacterium-mediated transient gene expression system for herbaceous plants and trees. Bioch Gene 2012, 50(9-10), 761–769. [Google Scholar] [CrossRef]
- Zhong, S.W.; Dong, B.; Zhou, J.; Miao, Y.F.; Yang, L.Y. Wang, Y.G. Xiao, Z.; Fang, Q.; Wan, Q.Q.; Zhao, H.B. Highly efficient transient gene expression of three tissues in Osmanthus fragrans mediated by Agrobacterium tumefaciens. Sci Hortic. 2023, 310, 111725. [Google Scholar] [CrossRef]
- Guo, Y. Wang, Y.C.; Wang, Z.B. Optimizing transient genetic transformation method on Arabidopsis plants mediated by Agrobacterium tumefaciens. J Northeast Forestryuniversity. 2016, 44, 41–44. [Google Scholar]
- Zhang K, He JJ, Liu L; Xie, R.D.; Qiu, L.; Li, X.C.; Yuan, W.J.; Chen, K.; Yin, Y.T.; Kyaw, M.M.M.; et al. A convenient, rapid and efficient method for establishing transgenic lines of Brassica napus. Plant Methods 2020, 16, 43. [Google Scholar] [CrossRef] [PubMed]
- Xian, B.; Xi, Z.Q.; Ren, C.X.; Yan, J.; Chen, J.; Pei, J. The establishment of transient expression systems and their application for gene function analysis of favonoid biosynthesis in Carthamus tinctorius L. BMC Plant Biol. 2023, 23, 186. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Yang, Q.; Yang, T.R.; Wu, Y.; Wang, G.X.; Yang, F.Y.; Wang, R.G.; Lin, X.F.; Li, G.J. Development of Agrobacterium-mediated transient expression system in Caragana intermedia and characterization of CiDREB1C in stress response. BMC Plant Biol. 2019, 19, 237–250. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.J.; Xie, L.; Shi, H.W.; Cui, S.R.; Lan, F.S.; Luo, Z.L.; Ma, X.J. Development of an efficient transient expression system for Siraitia grosvenorii fruit and functional characterization of two NADPH-cytochrome P450 reductases. Phytochemistry. 2021, 189, 112824. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J. Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25(4), 402–408. [Google Scholar] [CrossRef]
- Jelly, N.S.; Valat, L.; Walter, B.; Maillot, P. Transient expression assays in grapevine: a step towards genetic improvement. Plant Biotechnol J. 2014, 12(9), 1231–1245. [Google Scholar] [CrossRef]
- Wang, S.; Ku, S.S.; Ye, X.; He, C.; Kwon, S.Y.; Choi, P.S. Current status of genetic transformation technology developed in Cucumber (Cucumis Sativus L.). J Integr Agr. 2015, 14, 469–482. [Google Scholar] [CrossRef]
- Ebrahimzadegan, R.; Maroufi, A. In vitro regeneration and Agrobacterium-mediated genetic transformation of Dragon’s Head plant (Lallemantia iberica). Sci Rep. 2022, 12(1), 1784. [Google Scholar] [CrossRef] [PubMed]
- Mi, J.K.; Kon, B.; Chung-Mo, P. Optimization of conditions for transient Agrobacterium-mediated gene expression assays in Arabidopsis. Plant Cell Rep. 2009, 28, 1159–1167. [Google Scholar]
- Faizal, A.; Geelen, D. Agroinfiltration of intact leaves as a method for the transient and stable transformation of saponin producing Maesa Lanceolata. Plant Cell Rep. 2012, 31, 1517–1526. [Google Scholar] [CrossRef]
- Guan, S.X.; Kang, X.N.; Ge, J.Y.; Fei, R.W.; Duan, S.Y.; Sun, X.M. An efficient Agrobacterium-mediated transient transformation system and its application in gene function elucidation in Paeonia lactiflora Pall. Front Plant Sci. 2022, 13, 999433. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.W.; Ma, J.J.; Liu, H.M.; Guo, Y.T.; Li, W.; Niu, S.H. An efficient system for Agrobacterium-mediated transient transformation in Pinus tabuliformis. Plant Methods. 2020, 16, 52–61. [Google Scholar] [CrossRef]
- Zhang SY, Hu R, Yang L, Zuo ZJ. Establishment of a transient transformation protocol in Cinnamomum camphora. Forests. 2023, 14(9), 1872. [Google Scholar] [CrossRef]
- Li, Y.P.; Chen, T.T.; Wang, W. Liu, H.; Yan, X.; Wu-Zhang, K.Y.; Qin, W.; Xie, L.H,; Zhang, Y.J. Peng, B.W. et al. A high-efciency Agrobacterium-mediated transient expression system in the leaves of Artemisia annua L. Plant Methods. 2021, 17, 106. [Google Scholar] [CrossRef]
- Wu, S.L. Construction of transients system and function analysis of FT gene in mulberry (Morus alba L.). 2015. [master’s thesis] (Xi’an Shaanxi: Shaanxi Normal University).
- Chen, X.J.; He, S.T.; Jiang, L.N.; Li, X.Z.; Guo, W.L.; Chen, B.H.; Zhou, J.G. ; Skliar,V. An efficient transient transformation system for gene function studies in pumpkin (Cucurbita moschata D.). Sci Horticulturae. 2021, 282, 110028. [Google Scholar] [CrossRef]
- Xia, C.L.; Li, Y.N.; Wen, Z.B. The establishment and preliminary verification of transient transformation system of Salsola Laricifolia. Arid Zone Res. 2020, 37, 1317–1326. [Google Scholar]
- Mo, R.L.; Yang, S.C.; Zhang, Q.L.; Xu, L.Q.; Luo, Z.R. Vacuum infiltration enhances the Agrobacterium-mediated transient transformation for gene functional analysis in persimmon (Diospyros kaki thunb.). Sci Hortic Amsterdam. 2019, 251, 174–180. [Google Scholar] [CrossRef]
- Wu, Y.J.; Leng, Y.R.; Jing, S.L.; Zheng, C.P.; Lang, C.J.; Yang, L.P. Using vacuum infection method to transiently expression of GUS Gene in Medicago sativa L. by vacuum infiltration. Mol Plant Breed. 2021, 20, 859–864. [Google Scholar]
- Lacroix, B.; Citovsky, V. The roles of bacterial and host plant factors in Agrobacterium-mediated genetic transformation. Int J Dev Biol 2013, 57(6-7-8), 467–481. [Google Scholar] [CrossRef]
- Shoji, T. Analysis of the intracellular localization of transiently expressed and fluorescently labeled copper-containing amine oxidases, diamine oxidase and N-methylputrescine oxidase in tobacco, using an Agrobacterium infiltration protocol. Methods Mol Biol. 2017, 1694, 215–223. [Google Scholar]
- Li, H.P.; Li, K.; Guo, Y.T.; Guo, J.G.; Miao, K.T.; Botella, J.R.; Song, C.P.; Miao, Y.C. A transient transformation system for gene characterization in upland cotton (Gossypium hirsutum). Plant Methods. 2018, 14, 50–61. [Google Scholar] [CrossRef]
- Sharma, R.; Liang, Y.; Lee, Y.; Pidatala, V.R.; Mortimer, J.C.; Scheller, H.V. Agrobacterium-mediated transient transformation of sorghum leaves for accelerating functional genomics and genome editing studies. BMC Res Notes. 2020, 13, 116. [Google Scholar] [CrossRef]
- Sun, S.L.; Han, X.; Jin, R.X.; Jiao, J.B.; Wang, J.W.; Niu, S.Y.; Yang, Z.Y.; Wu, D.; Wang, Y.C. Generation of CRISPR-edited birch plants without DNA integration using Agrobacterium- mediated transformation technology. Plant Science. 2024, 342, 112029. [Google Scholar] [CrossRef]
- Zhou, L.M.; Huang, Y.Z.; Wang, Q.; Guo, D.J. AaHY5 ChIP-seq based on transient expression system reveals the role of AaWRKY14 in artemisinin biosynthetic gene regulation. Plant Physiol Bioch. 2021, 168, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.J.; Fang, J.R.; Lv, J.X.; Li, Z.Y. , Liu, Z. Y.; Wang, Y.C.; Wang, C. Gao, C.Q. Overexpression of ThSCL32 confers salt stress tolerance by enhancing ThPHD3 gene expression in Tamarix hispida. Tree Physiol. 2023, 43(8), 1444–1253. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Optimisation of different conditions for transient transformation of ZADT by infiltration. (ab, ef, ij) Graphs of GUS staining results and unstained phenotypes of ZADT seedlings after alcohol decolourisation for different surfactants, infestation times, and Agrobacterium growth concentrations; (c,g,k) Histograms of the relative expression of GUS genes for different optimized conditions of the infiltration method; (d,h,l) Histograms of the transformation efficiencies for different optimized conditions. The data was analysed using one-way analysis of variance (ANOVA), and differences in treatment means were expressed as mean ± SE. Tukey’s LSD test at p<0.05 was used for comparison. Bars=1.0 cm.
Figure 1.
Optimisation of different conditions for transient transformation of ZADT by infiltration. (ab, ef, ij) Graphs of GUS staining results and unstained phenotypes of ZADT seedlings after alcohol decolourisation for different surfactants, infestation times, and Agrobacterium growth concentrations; (c,g,k) Histograms of the relative expression of GUS genes for different optimized conditions of the infiltration method; (d,h,l) Histograms of the transformation efficiencies for different optimized conditions. The data was analysed using one-way analysis of variance (ANOVA), and differences in treatment means were expressed as mean ± SE. Tukey’s LSD test at p<0.05 was used for comparison. Bars=1.0 cm.
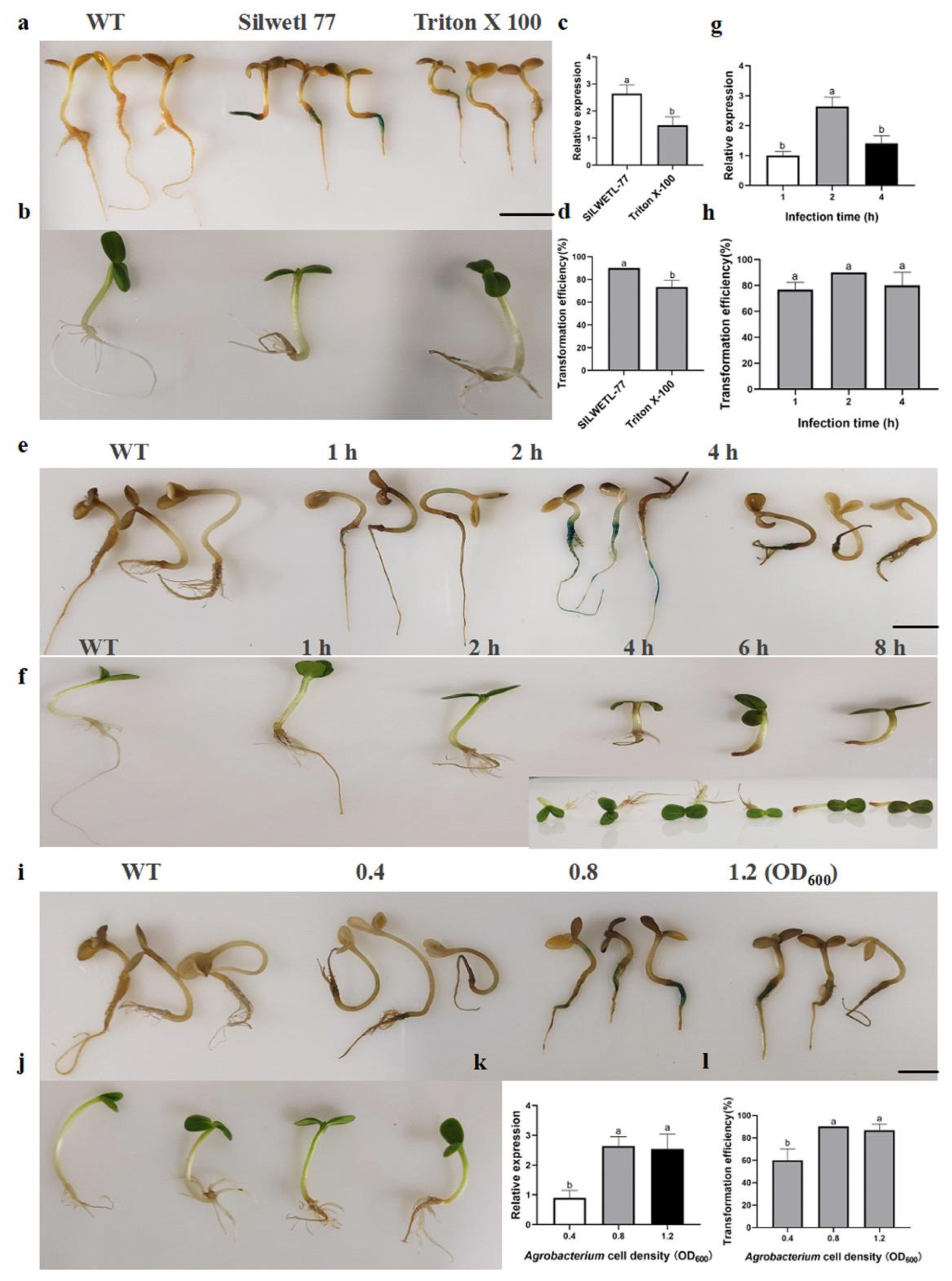
Figure 2.
Optimization of different conditions for transient transformation of ZADT by injection method. (a-d) Phenotypic plots of GUS staining optimized by the injection method for four conditions: surfactant, seedling growth time, Agrobacterium growth concentration and dark incubation time. (e,g,i,k) Relative expression of GUS gene for different optimisation conditions; (f,h,j,l) Histogram of transformation efficiency for different optimisation conditions. The experimental data were analysed by one-way analysis of variance (ANOVA). Differences in treatment means were expressed as mean ± SE, and compared at p<0.05 using Tukey’s LSD test. Bars=1.0 cm.
Figure 2.
Optimization of different conditions for transient transformation of ZADT by injection method. (a-d) Phenotypic plots of GUS staining optimized by the injection method for four conditions: surfactant, seedling growth time, Agrobacterium growth concentration and dark incubation time. (e,g,i,k) Relative expression of GUS gene for different optimisation conditions; (f,h,j,l) Histogram of transformation efficiency for different optimisation conditions. The experimental data were analysed by one-way analysis of variance (ANOVA). Differences in treatment means were expressed as mean ± SE, and compared at p<0.05 using Tukey’s LSD test. Bars=1.0 cm.
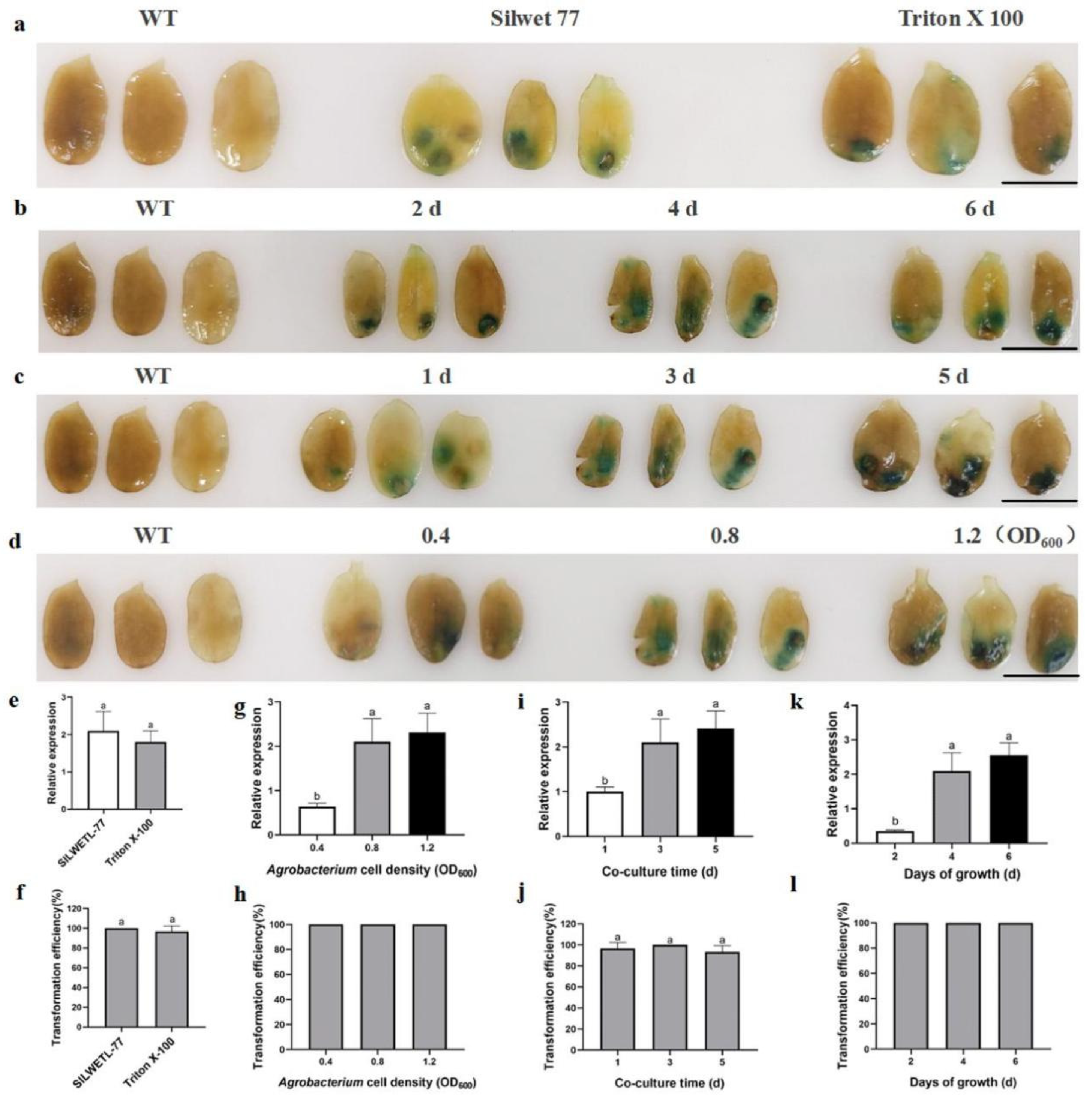
Figure 3.
Optimization of different conditions for transient transformation of ZADT by ultrasound-vacuum method. (ab, ef, ij) GUS staining results of Agrobacterium concentration, ultrasound time and vacuum filtration time after alcohol decolourisation and phenotypic plots of dark-grown ZADT seedlings without decolourization; (c,g,k) Relative expression of GUS genes under different optimized conditions; (d,h,l) Histograms of transformation efficiency under different optimized conditions. The experimental data were analysed by one-way analysis of variance (ANOVA). Differences in treatment means were expressed as mean ±SE and compared at p<0.05 using Tukey’s LSD test. Bars=1.0 cm.
Figure 3.
Optimization of different conditions for transient transformation of ZADT by ultrasound-vacuum method. (ab, ef, ij) GUS staining results of Agrobacterium concentration, ultrasound time and vacuum filtration time after alcohol decolourisation and phenotypic plots of dark-grown ZADT seedlings without decolourization; (c,g,k) Relative expression of GUS genes under different optimized conditions; (d,h,l) Histograms of transformation efficiency under different optimized conditions. The experimental data were analysed by one-way analysis of variance (ANOVA). Differences in treatment means were expressed as mean ±SE and compared at p<0.05 using Tukey’s LSD test. Bars=1.0 cm.
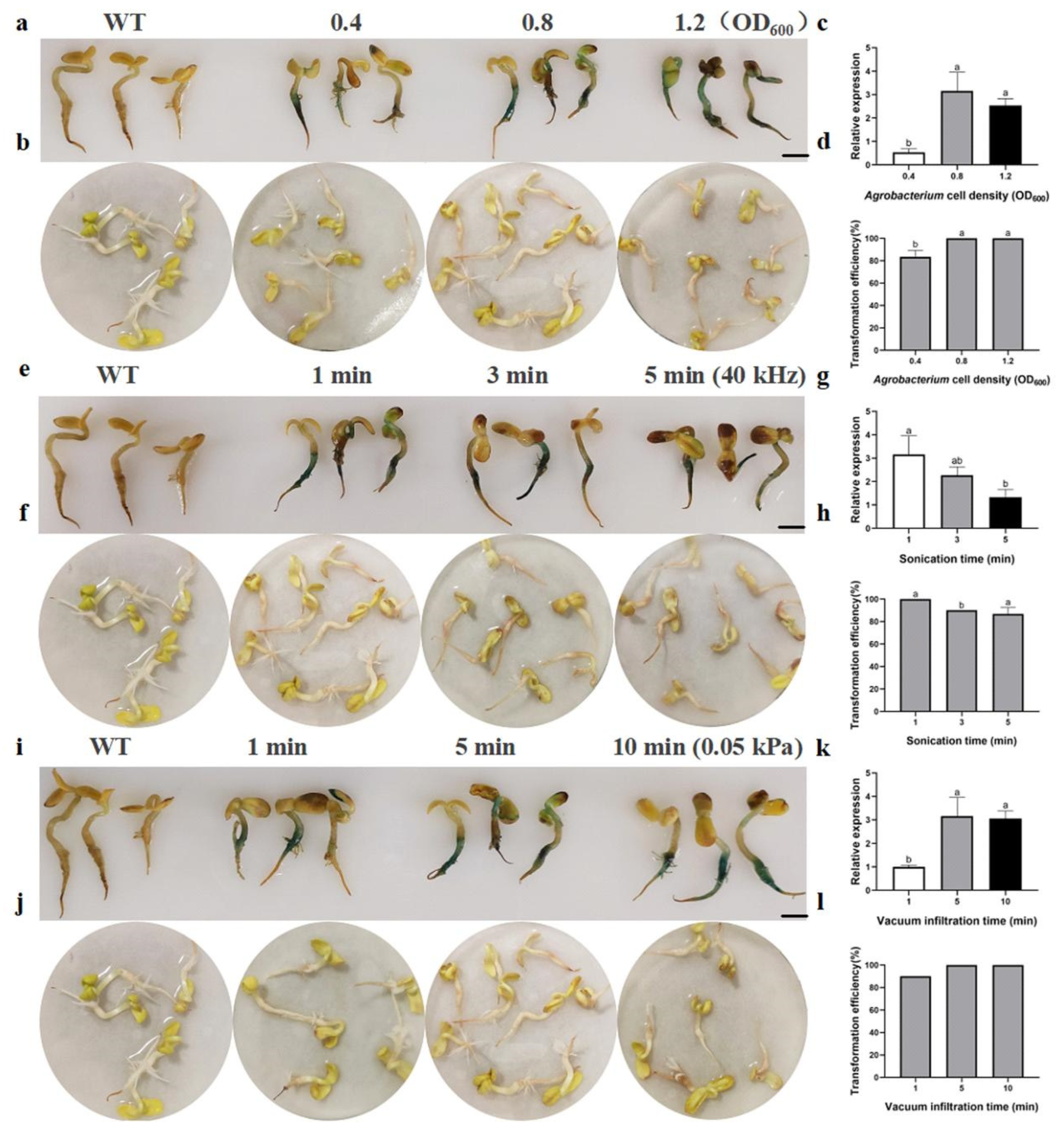
Figure 4.
Determination of GUS gene expression duration for three transient transformation methods of ZADT. (a-c) GUS staining phenotypes of ZADT seedlings transiently transformed were observed after treatment by the infiltration method for 2-6 d, injection method and ultrasound-vacuum method for 3-9 d after treatment. Bars=1.0 cm.
Figure 4.
Determination of GUS gene expression duration for three transient transformation methods of ZADT. (a-c) GUS staining phenotypes of ZADT seedlings transiently transformed were observed after treatment by the infiltration method for 2-6 d, injection method and ultrasound-vacuum method for 3-9 d after treatment. Bars=1.0 cm.

Figure 5.
Preliminary validation of the function of salt tolerance candidate genes based on transient transformation of ZADT by infiltration. (a) Phenotypes of transiently overexpressed ZADT seedlings. 1 and 3 are seedlings transiently transformed with the empty vector pBI121; 2 and 4 are seedlings transiently overexpressing HaMYB108 and HaWRKY40, respectively; (b) The results of transformed and transient overexpression of HaMYB108 and HaWRKY40 relative expression of empty vector pBI121 (EV) were detected by qRT-PCR, respectively; (c) Phenotypes of transiently overexpressed ZADT seedlings stressed by 350 mM NaCl for 3 d; (d) Detection of HaMYB108 and HaWRKY40 proteins in the transient expression system. 1. GFP protein: 26 kDa, 2. HaWRKY40 protein: 30.47 kDa, 3. HaMYB108 protein: 20.338 kDa, 4-6. β-actin protein: 43 kDa. Experimental data were analysed by one-way analysis of variance (ANOVA), and the differences in treatment means were expressed as mean±SE, and compared at p<0.05 using Tukey’s LSD test. Bars=0.5 cm.
Figure 5.
Preliminary validation of the function of salt tolerance candidate genes based on transient transformation of ZADT by infiltration. (a) Phenotypes of transiently overexpressed ZADT seedlings. 1 and 3 are seedlings transiently transformed with the empty vector pBI121; 2 and 4 are seedlings transiently overexpressing HaMYB108 and HaWRKY40, respectively; (b) The results of transformed and transient overexpression of HaMYB108 and HaWRKY40 relative expression of empty vector pBI121 (EV) were detected by qRT-PCR, respectively; (c) Phenotypes of transiently overexpressed ZADT seedlings stressed by 350 mM NaCl for 3 d; (d) Detection of HaMYB108 and HaWRKY40 proteins in the transient expression system. 1. GFP protein: 26 kDa, 2. HaWRKY40 protein: 30.47 kDa, 3. HaMYB108 protein: 20.338 kDa, 4-6. β-actin protein: 43 kDa. Experimental data were analysed by one-way analysis of variance (ANOVA), and the differences in treatment means were expressed as mean±SE, and compared at p<0.05 using Tukey’s LSD test. Bars=0.5 cm.
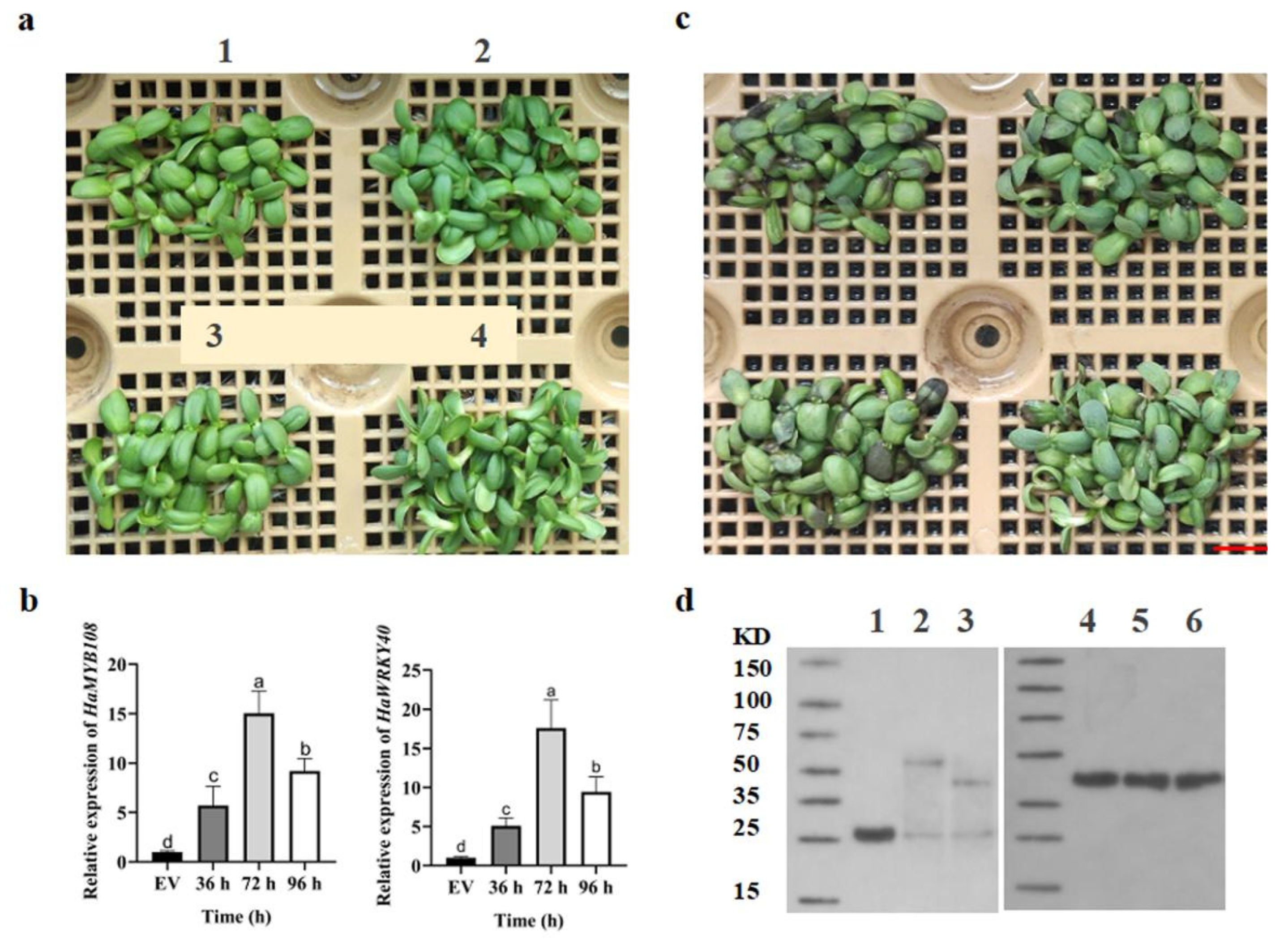
Figure 6.
Flow chart of Agrobacterium-mediated transient transformation system in sunflower.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



