
Submitted:
16 January 2025
Posted:
16 January 2025
You are already at the latest version
Abstract
The target of rapamycin (TOR) kinase is an evolutionarily conserved atypical Ser/Thr protein kinase found in yeasts, plants, and mammals. In plants, TOR functions as a central signaling hub, playing a crucial role in the precise orchestration of growth and development. Extensive studies have underscored its significance in plant signaling by highlighting its integral role in these processes. Recent research has further elucidated TOR's multifaceted roles in plant responses to environmental stresses, emphasizing its importance in stress adaptation. In this review, we will discuss recent findings on the involvement of TOR signaling in plant adaptation to various abiotic and biotic stresses. Moreover, mounting evidence indicates TOR’s role in mediating the plant transcriptome. Therefore, we will specifically discuss TOR-regulated metabolome reprogramming in plant response to different stresses.
Keywords:
Target of Rapamycin (TOR)
; plant stress responses
; biotic stress
; abiotic stress
; metabolome
1. Introduction
Plants exist in intricate environments where they are continually subjected to a range of abiotic and biotic stresses, such as extreme temperatures, drought, high salinity, and pathogen attacks [1,2]. Given their immobility, plants have evolved a sophisticated signal transduction network to manage these challenging environmental conditions [1,2]. This network facilitates large-scale transcriptional, translational, and metabolic changes, which are crucial for redirecting the plant’s limited energies and resources from growth and development towards defense mechanisms [3]. Consequently, the plants’ stress responses are intricately linked to these adaptive changes, enabling them to survive and thrive despite adverse conditions.
TOR is a conserved serine/threonine kinase found in eukaryotes and is a member of the phosphoinositide-3 kinase-related protein kinase (PIKK) family [4,5]. Initially identified in the yeast (Saccharomyces cerevisiae), TOR was recognized as the target protein of rapamycin, an antibiotic produced by Streptomyces hygroscopicus and first discovered in 1975 by Vézina et al. [6]. Subsequent studies have demonstrated the presence of TOR across a variety of non-photosynthetic organisms, including mammals, and Drosophila [4,5]. Furthermore, TOR has been identified in plants, expanding its known presence across both photosynthetic and non-photosynthetic organisms [4,5].
The TOR genes, identified across various species, encode large proteins typically comprising 2, 400-2, 500 amino acids, and these proteins exhibit significant similarity among different eukaryotes [4,5]. A typical TOR protein is characterized by several domains arranged sequentially from the N-terminus to the C-terminus: the HEAT tandem repeat sequence, FAT domain, FRB domain, protein kinase domain, and FATC domain [4,5]. The HEAT repeat sequences are crucial for facilitating protein-protein interactions and membrane binding [4,5]. Both the FAT and FATC domains play roles in protein interactions and kinase activation [4,5]. The FRB domain serves as the target for rapamycin, a specific inhibitor of TOR kinase. Rapamycin binds to the FKBP12 protein, forming a complex that specifically attaches to the FRB domain, thereby inhibiting TOR activity [4,5].
TOR operates primarily through two complexes, TORC1 and TORC2, which are distinguished by their composition and function [4,7,8]. TORC1 is composed of TOR, Lethal with Sec Thirteen 8 (LST8 in plants and yeast, mLST8 in mammals), and the Regulatory-Associated Protein of TOR (RAPTOR in mammals and plants, KOG1 in yeast); in contrast, TORC2 includes TOR, LST8/mLST8, Raptor Independent Companion of TOR (RICTOR), and Stress-activated MAP Kinase Interacting Protein 1 (SIN1) [4,9,10,11,12]. In yeast, there are two TOR genes, TOR1 and TOR2, with TOR1 or TOR2 capable of forming a TORC1 complex, whereas only TOR2 can form a TORC2 complex [13]. In mammals, a single TOR gene is involved in both TORC1 and TORC2 complexes [14]. In plants, only the typical TORC1 complex has been identified. However, TORC2 may not be conserved. It is possible that other proteins with similar functions could substitute for RICTOR and SIN1, suggesting the potential existence of another form of TORC2 complex in plants [13]. Notably, the FKBP12-rapamycin complex binds to the conserved FRB domain in TOR kinase, inhibiting TORC1 activity, but it cannot bind to TORC2 complexes due to the spatial occupation by mSIN1 or AVO1 [4].
Over the past two and a half decades since its discovery, extensive research in both yeast and mammalian model systems has revealed TOR’s fundamental role as a central signaling hub [8,15,16,17]. By integrating multiple major signaling pathways, TOR coordinates responses to nutrient availability, cellular energy status, growth factors, and various stressors [8,15,16,17]. Through this comprehensive integration of signals, TOR effectively orchestrates both cellular and organismal physiology, underscoring its essential function in maintaining homeostasis and regulating diverse biological processes [8,15,16,17].
In Arabidopsis, null tor mutants exhibit an embryonic lethal phenotype, and TOR in Arabidopsis has traditionally been regarded as resistant to rapamycin, which has hindered progress in plant TOR research. Some groundbreaking work addressed these challenges by developing a system to detect TOR activity in plants and creating the estradiol-inducible tor mutant, tor-es [18,19]. This innovation facilitated the discovery that plant TOR is indeed sensitive to rapamycin, albeit requiring higher concentrations [18,19]. Subsequently, the introduction of new-generation ATP-competitive chemical inhibitors targeting TOR kinase, such as AZD8055, Torin1, Torin2, and KU0063794, has significantly advanced plant TOR research [20]. TOR has been extensively reported to mediate various aspects of plant growth and development, by regulating translation, transcription, autophagy, and primary and secondary metabolism. Additionally, numerous studies have highlighted TOR’s crucial role in plant responses to both biotic and abiotic stresses [21].
In this review, we will explore recent advances in TOR-mediated stress responses. Specifically, as the significance of TOR-regulated primary and secondary metabolism becomes increasingly recognized, we will examine the specific role of TOR-modulated metabolic reprogramming in plant responses to diverse biotic and abiotic stresses.
2. Insight on TOR-Mediated Stress Responses via Metabolome Reprogramming
2.1. TOR Is Widely Involved in Plants Responses to Various Environmental Stresses
As illustrated in Figure 1a, a typical TOR protein in plants is sequentially arranged from the N-terminus to the C-terminus as follows: HEAT repeats, FAT domain, FRB domain, kinase domain, and FATC domain. The amino acid sequences of TOR kinases across different plant species exhibit high levels of identity. For example, Arabidopsis TOR (Q9FR53) and tomato TOR (A0A3Q7ERV7), both dicotyledonous plants, share an identity of 78.9% and a similarity of 86.9%. In comparison, AtTOR (Arabidopsis TOR) and OsTOR (rice TOR, Q0DJS1), with rice being a monocotyledonous plant, show an identity of 72.9% and a similarity of 82.2%. The high degree of identity and similarity in TOR amino acid sequences among various plant species underscores the strong conservation of its function. To date, only one TOR complex has been identified in plants, consisting of Raptor, LST8, and TOR itself (Figure 1a).
The function of TOR has long been associated with regulating protein translation [22]. However, recent discoveries have revealed that TOR can either directly phosphorylate specific transcription factors, such as E2Fa and E2Fb, or indirectly regulate the stability of others, including EIN3 in ethylene signaling [23,24]. These findings underscore TOR’s crucial role in gene transcription regulation. Furthermore, transcriptome analysis indicates that TOR facilitates significant transcriptomic reprogramming [23,24].
To elucidate the role of TOR in regulating plant responses to stress, we conducted a comparative analysis of genes regulated by TOR (identified through treatment with a TOR inhibitor, Torin2) [25] and those affected by various biotic and abiotic stresses in tomato. Specifically, we pooled genes differentially expressed in response to various kinds of biotic stresses into one set, including Pseudomonas syringae pv. tomato DC3000-, Phytophthora parasitica-, and herbivore-regulated genes (Table S1) [26,27,28]; similarly, genes differentially expressed in response to drought, salt, cold, and heat were consolidated to form the abiotic stress-responsive gene set (Table S2) [29,30,31,32]. Upon analysis of gene expression patterns, we found substantial overlap between TOR-regulated genes and stress-responsive gene sets. Specifically, approximately one-fifth (463/2058) of the TOR-mediated genes showed concordance with biotic stress-responsive genes, while approximately half (924/2058) overlapped with genes regulated under abiotic stress conditions (Figure 1b-c). This significant overlap suggests a broad involvement of TOR in modulating plant stress responses.
To elucidate the biological pathways implicated by these intersecting genes, we conducted a KEGG (Kyoto Encyclopedia of Genes and Genomes) pathway enrichment analysis using the two overlapping gene sets. Notably, when examining the intersection between TOR-regulated genes and those responsive to biotic or abiotic stresses, the “metabolic pathways” and “biosynthesis of secondary metabolites” emerged as the most significantly enriched pathways (Figure 1d-e). Transcriptional changes are relatively direct processes that occur within hours, typically preceding metabolic changes. It is possible that TOR may affect plant stress response through transcriptome reprogramming, followed by metabolome reprogramming, ultimately leading to physiological adaptations to stress conditions.
Based on the conclusions drawn from the transcriptome analysis, we will first summarize recent findings concerning TOR-regulated plant responses to abiotic and biotic stresses. Subsequently, we will specifically discuss the role of TOR-mediated metabolic changes in plant stress adaptation.
2.2. TOR-Regulated Plant Responses to Abiotic Stresses
Throughout their life cycle, plants face a range of abiotic stresses, including cold, heat, drought, and salt [1]. To survive and thrive under these fluctuating environmental conditions, plants must swiftly respond to diverse stress signals [1]. A pivotal component of this adaptive response is the TOR signaling pathway, which is instrumental in regulating plant reactions to these various abiotic stresses [4].
Abnormal temperatures have a profound impact on biological processes in plants, influencing growth, development, metabolism, protein translation, and gene expression [33,34]. Recent studies indicate that TOR plays a crucial role in mediating the effects of both low and high temperatures.
In Arabidopsis, exposure to low temperatures initially causes a rapid reduction in TOR activity within the first hour, followed by a recovery after a longer duration [35,36,37]. This fluctuation pattern in TOR activity is also observed in tomato, suggesting that it represents a conserved mechanism across different species [25]. Furthermore, cold stress not only affects TOR activity at the post-translational level but also influences its expression levels. Specifically, cold exposure induces TOR expression between 1- and 3-hour post-treatment, whereas prolonged exposure results in the repression of its expression [35]. These findings collectively indicate that cold stress modulates TOR both at the post-translational and transcriptional levels.
TOR plays a complex role in regulating cold responses in Arabidopsis. Research by Wang et al. indicates that TOR negatively regulates cold responses, whereas findings by Dong et al. suggest that TOR promotes these responses, with Thyroid Adenoma Associated (THADA) acting upstream of TOR in this process [37,38]. Additionally, the FERONIA-ROP2-TOR module has been shown to enhance root hair growth at low temperatures, underscoring TOR’s multifaceted role in plant cold resistance [36]. In tomatoes, low temperatures rapidly and transiently suppress TOR activity, which subsequently activates the transcriptional activity of its direct substrate, PGH1 [25]. This activation leads to increased expression of the C-REPEAT-BINDING FACTOR 1 (CBF1) gene, a key player in cold acclimation and stress response [25]. This, in turn, activates the expression of cold stress response-related genes and regulates metabolic reprogramming, enhancing the accumulation of cold-resistant metabolites, and thereby improving tomato cold resistance [25]. These findings provide comprehensive insights into the intricate regulatory mechanisms of TOR in plant cold stress response.
Elevated temperatures also adversely affect grain crop yield and quality. In perennial ryegrass (Lolium perenne), TOR expression is upregulated under high-temperature stress [37]. In Arabidopsis, glucose via TOR governs the transcriptome reprogramming of a large number of genes involved in heat stress protection [39,40,41]. The E2Fa (E2 PROMOTER BINDING FACTOR a) transcription factor, activated by glucose-TOR module, is shown to activate root apical meristem and promote root growth [23]. Meanwhile, recent studies also indicate that the glucose-TOR-activated E2Fa also binds to the promoters of HSF genes and enhance the expression of heat-responsive genes [39,40,41]. Specifically, the Glc-TOR-E2Fa module activates the expression of HIKESHI-LIKE PROTEIN1 (HLP1), which binds to the promoters of glucose-regulated HS-responsive genes [39,40,41]. Furthermore, Glc-TOR-E2Fa activates the expression of ARABIDOPSIS TRITHORAX 1 (ATX1), encoding an H3K4 methyltransferase already shown to regulate H3K4me3 levels at the promoters of HS recovery genes. Also, glucose through TOR promotes recruitment of histone H3 acetylation marks at the promoters of HS genes to induce their transcription, leading to thermotolerance. This Glc-TOR-mediated histone acetylation is facilitated through HAC1 [39,40,41].
Salt and osmotic stress induce physiological and biochemical changes, leading to metabolic irregularities and growth cessation [42,43]. High NaCl concentration increases TOR activity, while mannitol-induced drought stress inhibits TOR activity [35]. Constitutive overexpression of TOR enhances salt and osmotic stress tolerance in Arabidopsis thaliana, showing improved performance in root growth, fresh weight, and lateral root density [44,45]. YAK1 in Arabidopsis acts as a positive regulator of ABA-mediated drought response. Under favorable growth conditions, TOR inhibits YAK1 activity, thereby negatively regulating ABA signaling transduction and ABA-mediated drought response [46,47]. Furthermore, oxidative stress is reported to enhance TOR activity during short-time treatment, while inhibits TOR activity [35].
Abscisic acid (ABA) is a crucial stress phytohormone that plays a significant role in plant responses to various abiotic stresses, including cold, salt, osmotic, and drought conditions [48]. A sophisticated reciprocal regulation exists between ABA and TOR signaling, which balances stress adaptation and plant growth [49,50]. Under stress conditions, ABA-activated SnRK2 kinases directly interact with and phosphorylate RAPTOR, a core component of the TOR complex (TORC1) [49,50]. This interaction leads to the dissociation of TORC1, inhibition of TOR kinase activity, and subsequent repression of plant growth [49,50]. Conversely, under favorable conditions, activated TOR kinase phosphorylates PYLs, preventing ABA from binding to these receptors [49,50]. This inactivation of SnRK2 promotes plant growth [49,50]. Salicylic acid (SA) is another essential stress phytohormone involved in various abiotic stress responses. A recent study conducted on tomatoes demonstrated that inhibiting TOR under cold stress conditions significantly increased the levels of SA and its derivatives [25].
Autophagy serves as a fundamental degradation and recycling pathway for cytoplasmic substances in eukaryotes and is induced by various stressors, with a significant connection to the TOR signaling pathway [51]. In plants, inhibition of TOR signaling can initiate autophagy, while TOR overexpression can prevent it under nutrient deficiencies, salt, and drought stress, though not under oxidative or endoplasmic reticulum stress [52,53,54,55]. SNF1-related kinase 1 (SnRK1) plays a crucial role in regulating autophagy upstream of TOR during nutrient deficiency, osmotic pressure, and salt stress. In contrast, the regulation of autophagy by oxidative and endoplasmic reticulum stress is dependent on SnRK1 rather than TOR [53,54,55]. Although the role of plant TOR as a regulator of autophagy in response to nutrients and abiotic stress conditions is established [52,53,54,55,56], the precise connection between TOR and the core components in autophagy remains unclear. Notably, recent interactome and phosphoproteomics analyses have identified several autophagy-related proteins as potential substrates of TOR kinase, suggesting a more intricate involvement of TOR in autophagy regulation [21,57].
2.3. TOR-Regulated Plant Responses to Biotic Stress
In addition to various abiotic stresses, plants are also threatened by a range of biotic stresses, including viruses, bacteria, fungi, nematodes, and insects. To counter these biotic threats, plants have developed a sophisticated immune system comprising two layers of defense: pattern-triggered immunity (PTI) and effector-triggered immunity (ETI) [58,59]. PTI is activated by molecular patterns associated with microbes, pathogens, nematodes, herbivores, and parasites, known as MAMPs, PAMPs, NAMPs, HAMPs, and ParAMPs, respectively. In contrast, pathogen- and parasite-secreted proteins, termed as effectors, are perceived by host-derived immunogenic molecules to trigger ETI [58,59]. This dual-layered immune system enables plants to effectively recognize and respond to a wide array of biotic stressors. In this review, plant defense responses against pathogens are a major focus of discussion.
The mechanisms by which plant TOR responds to various pathogens remain largely unexplored. However, recent studies have begun to elucidate potential pathways of regulation. For instance, it has been demonstrated that biotic stresses may influence TOR at the transcriptional level. Specifically, a study on tomatoes revealed that the transcription factor SlMYC2 directly binds to the promoter region of SlTOR, thereby activating its transcription [60]. Given that jasmonic acid (JA) is a crucial phytohormone involved in biotic stress responses and that MYC2 is the central transcription factor in JA signaling, it is plausible that certain biotic stresses increase JA levels, which in turn activate MYC2 and subsequently enhance TOR transcription [60,61]. In contrast, at the translational level, the Pseudomonas effector AvrRpm1 has been found to suppress TOR expression [62]. Furthermore, the type III effector AWR5, when expressed in yeast, exhibits a function similar to that of rapamycin, suggesting a potential role in TOR regulation [63]. These findings indicate a complex interplay between biotic stressors and TOR signaling, warranting further investigation.
In Solanaceae plants such as tomato and tobacco, the inhibition of TOR activity, either through the use of the TOR inhibitor Torin2 or via Virus-Induced Gene Silencing (VIGS), has been shown to enhance plant defense against various pathogens. In tomatoes, specifically, the repression of TOR activity strengthens defense mechanisms against Botrytis cinerea, Alternaria alternata and Xanthomonas euvesicatoria [64]. Similarly, in tobacco, silencing TOR increases resistance to Xanthomonas euvesicatoria and the tobacco mosaic virus [64]. The study indicates that TOR-regulated plant immunity is dependent on SA [64]. In tomatoes, TOR coordinates cytokinin (CK) and gibberellin (GA) signaling, which mediates both development and defense [65]. The effectiveness of disease resistance mediated by TOR inhibition varies with the developmental stage, being absent in highly morphogenetic leaves but most potent in mature, differentiated ones [65]. Marash’s research demonstrated that inoculating tomatoes with pathogens at different CK/GA ratios revealed that higher CK/GA levels suppressed TOR activity and enhanced resistance, whereas lower CK/GA levels had the opposite effect [65]. This suggests that the differential regulation of TOR could modulate the development-defense trade-off in plants. In rice, TOR negatively regulates both SA- and JA-dependent immune responses and pathogen-triggered immunity (PTI) [66]. SA and JA are known to have antagonistic effects in plant immunity; SA positively regulates immunity against biotrophic and hemibiotrophic pathogens, while JA is effective against necrotrophic pathogens [67]. This study provides valuable insights into how TOR may mitigate the competing effects of these two plant hormones, thereby regulating plant responses to a diverse array of pathogens. Furthermore, the research in rice established a sophisticated model illustrating that TOR balances plant growth and defense, promoting growth while repressing defense responses. In Arabidopsis, TOR inhibition via a TOR inhibitor increased resistance to Fusarium graminearum [68], and the suppression of TOR expression enhanced resistance to Pseudomonas syringae and oomycete pathogens [62]. Additionally, viruses can hijack TOR signaling to facilitate their replication [69], and TOR silencing or inhibition has been shown to promote resistance against the watermelon mosaic virus [70].
2.4. TOR-Modulated Metabolic Reprogramming Contributes to Plant Stress Responses
Mammalian mTOR-mediated metabolic reprogramming is extensively studied, and its dysregulation is implicated in severe human diseases like cancer, type 2 diabetes/obesity, and neurodegenerative disorders [71].
Similarly in plants, multiple independent studies have demonstrated that TOR tightly regulates the plant metabolome [72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89]. Of particular interest is the relationship between TOR-regulated metabolites and plant stress responses, as it has been clearly shown that stress triggers numerous metabolic changes in plants [90]. To systematically investigate this connection, we analyzed the metabolomic data from a comprehensive study by Song et al. in Arabidopsis [84]. Their analysis of an inducible tor mutant (tor-es) identified 141 differentially accumulated metabolites (DAMs). Through extensive literature review of these DAMs’ biological functions, we found that 25 metabolites were associated with abiotic stress responses, 11 with biotic stress responses, and 3 metabolites (3-Methylmalic acid, amentoflavone, and caffeine) were involved in both response pathways, highlighting the significant role of TOR in plant stress metabolism. The functions of the 33 DAMs involved in plant stress responses are summarized in Table 1. Based on the table, TOR appears to regulate plant stress responses by controlling the synthesis and accumulation of stress-response metabolites.
More direct evidence for TOR-mediated metabolic reprogramming in stress resistance comes from a study in tomato. In tomato, cold stress inhibits TOR kinase activity, which triggers a cascade of molecular responses that enhance cold resistance through metabolic reprogramming [25]. This inhibition prevents TOR kinase from phosphorylating the transcription factor PGH1, and the resulting dephosphorylated PGH1 exhibits increased transcriptional activity, leading to enhanced expression of the core cold stress response gene CBF1 [25]. Through this TOR-PGH1-CBF1 axis, extensive transcriptional reprogramming occurs, particularly affecting genes encoding key metabolic enzymes [25]. Specifically, while genes involved in amino acid synthesis are downregulated, those crucial for protective compound synthesis are upregulated, including CS2 and ICS1 in the shikimate synthesis pathway and OTC, ADC1, and ADC2 in the putrescine synthesis pathway [25]. Metabolomic analyses confirm that TOR inhibition under cold stress conditions substantially alters the metabolic landscape [25]. These alterations manifest as reduced amino acid synthesis coupled with enhanced production of cryoprotective compounds, including salicylic acid and its derivatives, flavonoids, and putrescine [25]. This metabolic shift suggests that TOR functions as a central regulator in cold stress response by redistributing carbon skeletons from primary metabolism toward the synthesis of cryoprotective compounds, thereby enabling plants to adapt to cold conditions [25].
3. Discussion
Plants face continuous challenges from diverse environmental stresses, which has led to the evolution of sophisticated signaling networks for survival and adaptation. The TOR has emerged as a central regulatory component within these networks, playing crucial roles in stress response mechanisms. Extensive research from multiple laboratories has demonstrated TOR’s widespread involvement in mediating plants’ responses to various environmental stresses. Despite this substantial body of work, there remains a significant gap in our understanding of how TOR-regulated metabolic alterations connect to TOR-mediated stress responses. Therefore, investigating TOR-orchestrated stress responses from a metabolic perspective offers promising new insights into deciphering these complex regulatory mechanisms.
TOR has been demonstrated to influence the accumulation of diverse metabolites in plants, though the underlying mechanisms remain largely unexplored. TOR may exert both direct and indirect control over metabolite biosynthesis. In terms of indirect regulation, TOR can trigger comprehensive transcriptome reprogramming, leading to the modulation of stress-resistant compound accumulation [25]. This often occurs at the expense of growth- and development-related metabolites, reflecting a resource allocation strategy through which plants maintain a delicate balance between defense and growth under limited energy resources [25]. Additionally, TOR can directly regulate metabolic pathways by phosphorylating key enzymatic components. This direct regulatory mechanism is exemplified in a recent tomato study, where TOR was found to phosphorylate PGH1, a crucial glycolytic enzyme that also functions as a transcription factor in CBF1 expression regulation [25]. Future studies exploring additional TOR substrates involved in metabolic pathways will help elucidate how TOR regulates metabolite accumulation through direct phosphorylation of key biosynthetic enzymes.
Under natural conditions, plants are exposed to multiple concurrent environmental stresses rather than experiencing individual stressors in isolation. The TOR signaling pathway demonstrates a remarkable capacity to orchestrate multiple stress responses through its regulation of diverse compounds, as comprehensively illustrated in Table 1. These compounds are integral components of various stress response mechanisms, positioning TOR as an ideal molecular framework for investigating the complex interplay of multiple environmental stresses and their combined effects on plant physiology.
The mechanism by which TOR regulates stress responses through metabolic reprogramming is illustrated in Figure 2.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Table S1: List of genes overlapping between SlTOR-regulated and biotic stress-responsive genes; Table S2: List of genes overlapping between SlTOR-regulated and abiotic stress-responsive genes.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, L.F.; data curation, L.Y., R.Z., H.Z., L.F; writing—original draft preparation, L.Y., R.Z., H.Z., L.F.; writing—review and editing, L.Y., R.Z., H.Z., Y.Y., L.F.; funding acquisition, L.F. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the National Natural Science Foundation of China (grant no. 32170308 to L.F.) and the funding from Shanghai Jiao Tong University (grant no. WH220415006 to L.F.).
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Zhang, H., et al., Abiotic stress responses in plants. Nature Reviews Genetics, 2022. 23(2): p. 104-119. [CrossRef]
- Jones, J.D.G., B.J. Staskawicz, and J.L. Dangl, The plant immune system: From discovery to deployment. Cell, 2024. 187(9): p. 2095-2116. [CrossRef]
- Monson, R.K., et al., Coordinated resource allocation to plant growth-defense tradeoffs. New Phytol, 2022. 233(3): p. 1051-1066. [CrossRef]
- Liu, Y. and Y. Xiong, Plant target of rapamycin signaling network: Complexes, conservations, and specificities. Journal of Integrative Plant Biology, 2022. 64(2): p. 342-370. [CrossRef]
- Meng, Y., et al., TOR kinase, a GPS in the complex nutrient and hormonal signaling networks to guide plant growth and development. Journal of Experimental Botany, 2022. 73(20): p. 7041-7054. [CrossRef]
- Heitman, J., N.R. Movva, and M.N. Hall, Targets for cell cycle arrest by the immunosuppressant rapamycin in yeast. Science, 1991. 253(5022): p. 905-909. [CrossRef]
- Shi, F. and S. Collins, Regulation of mTOR signaling: Emerging role of cyclic nucleotide-dependent protein kinases and implications for cardiometabolic disease. Int J Mol Sci, 2023. 24(14): p. 11497.
- Emmerstorfer-Augustin, A. and J. Thorner, Regulation of TORC2 function and localization in yeast. Annu Rev Cell Dev Biol, 2023. 39: p. 363-389. [CrossRef]
- Wullschleger, S., R. Loewith, and M.N. Hall, TOR signaling in growth and metabolism. Cell, 2006. 124(3): p. 471-84. [CrossRef]
- Shi, L., Y. Wu, and J. Sheen, TOR signaling in plants: conservation and innovation. Development, 2018. 145(13). [CrossRef]
- Liu, G.Y. and D.M. Sabatini, mTOR at the nexus of nutrition, growth, ageing and disease. Nature Reviews Molecular Cell Biology, 2020. 21(4): p. 183-203. [CrossRef]
- Inoki, K., J. Kim, and K.L. Guan, AMPK and mTOR in cellular energy homeostasis and drug targets. Annu Rev Pharmacol Toxicol, 2012. 52: p. 381-400. [CrossRef]
- Burkart, G.M. and F. Brandizzi, A tour of TOR complex signaling in plants. Trends in Biochemical Sciences, 2021. 46(5): p. 417-428. [CrossRef]
- Saha, S., et al., mTORC1 and SGLT2 inhibitors-a therapeutic perspective for diabetic cardiomyopathy. Int J Mol Sci, 2023. 24(20): p. 15078.
- He, L., S. Cho, and J. Blenis, mTORC1, the maestro of cell metabolism and growth. Genes Dev, 2025. 39(1-2): p. 109-131. [CrossRef]
- Ragupathi, A., C. Kim, and E. Jacinto, The mTORC2 signaling network: Targets and cross-talks. Biochemical Journal, 2024. 481(2): p. 45-91. [CrossRef]
- Foltman, M. and A. Sanchez-Diaz, TOR complex 1: Orchestrating nutrient signaling and cell cycle progression. Int J Mol Sci, 2023. 24(21): p. 15745.
- Xiong, Y. and J. Sheen, Rapamycin and glucose-target of rapamycin (TOR) protein signaling in plants. Journal of Biological Chemistry, 2012. 287(4): p. 2836-2842. [CrossRef]
- Xiong, Y., et al., Glucose-TOR signalling reprograms the transcriptome and activates meristems. Nature, 2013. 496(7444): p. 181-186. [CrossRef]
- Xiong, Y. and J. Sheen, The role of target of rapamycin signaling networks in plant growth and metabolism. Plant Physiology, 2014. 164(2): p. 499-512. [CrossRef]
- Fu, L., P. Wang, and Y. Xiong, Target of rapamycin signaling in plant stress responses. Plant Physiology, 2020. 182(4): p. 1613-1623. [CrossRef]
- Xiong, Y. and J. Sheen, Moving beyond translation: glucose-TOR signaling in the transcriptional control of cell cycle. Cell Cycle, 2013. 12(13): p. 1989-90. [CrossRef]
- Xiong, Y., et al., Glucose-TOR signalling reprograms the transcriptome and activates meristems. Nature, 2013. 496(7444): p. 181-6. [CrossRef]
- Fu, L., et al., The TOR-EIN2 axis mediates nuclear signalling to modulate plant growth. Nature, 2021. 591(7849): p. 288-292. [CrossRef]
- Li, Z., et al., TOR balances plant growth and cold tolerance by orchestrating amino acid-derived metabolism in tomato. Hortic Res, 2024. 11(12): p. uhae253. [CrossRef]
- Hu, C., et al., Ethylene response factors 15 and 16 trigger jasmonate biosynthesis in tomato during herbivore resistance. Plant Physiol, 2021. 185(3): p. 1182-1197. [CrossRef]
- Naveed, Z.A. and G.S. Ali, Comparative transcriptome analysis between a resistant and a susceptible wild tomato accession in response to Phytophthora parasitica. Int J Mol Sci, 2018. 19(12). [CrossRef]
- Hu, Z., et al., High CO2- and pathogen-driven expression of the carbonic anhydrase βCA3 confers basal immunity in tomato. New Phytol, 2021. 229(5): p. 2827-2843. [CrossRef]
- Ikuyinminu, E., et al., Transcriptome, biochemical and phenotypic analysis of the effects of a precision engineered biostimulant for inducing salinity stress tolerance in tomato. Int J Mol Sci, 2023. 24(8). [CrossRef]
- Huang, Y., et al., HSFA1a modulates plant heat stress responses and alters the 3D chromatin organization of enhancer-promoter interactions. Nat Commun, 2023. 14(1): p. 469. [CrossRef]
- Liu, M., et al., Profiling of drought-responsive microRNA and mRNA in tomato using high-throughput sequencing. BMC Genomics, 2017. 18(1): p. 481. [CrossRef]
- Lin, R., et al., CALMODULIN6 negatively regulates cold tolerance by attenuating ICE1-dependent stress responses in tomato. Plant Physiol, 2023. 193(3): p. 2105-2121. [CrossRef]
- Ding, Y., Y. Shi, and S. Yang, Regulatory networks underlying plant responses and adaptation to cold stress. Annu Rev Genet, 2024. 58(1): p. 43-65. [CrossRef]
- Kan, Y., et al., The molecular basis of heat stress responses in plants. Molecular Plant, 2023. 16(10): p. 1612-1634. [CrossRef]
- Pereyra, C.M., et al., Target of rapamycin signaling is tightly and differently regulated in the plant response under distinct abiotic stresses. Planta, 2019. 251(1): p. 21. [CrossRef]
- Pacheco, J.M., et al., Cell surface receptor kinase FERONIA linked to nutrient sensor TORC signaling controls root hair growth at low temperature linked to low nitrate in Arabidopsis thaliana. New Phytol, 2023. 238(1): p. 169-185. [CrossRef]
- Wang, L., et al., The inhibition of protein translation mediated by AtGCN1 is essential for cold tolerance in Arabidopsis thaliana. Plant Cell Environ, 2017. 40(1): p. 56-68. [CrossRef]
- Dong, Y., et al., The Arabidopsis THADA homologue modulates TOR activity and cold acclimation. Plant Biol (Stuttg), 2019. 21 Suppl 1: p. 77-83. [CrossRef]
- Sharma, M., et al., Glucose-regulated HLP1 acts as a key molecule in governing thermomemory. Plant Physiol, 2019. 180(2): p. 1081-1100. [CrossRef]
- Sharma, M., et al., Arabidopsis target of rapamycin coordinates with transcriptional and epigenetic machinery to regulate thermotolerance. Front Plant Sci, 2021. 12: p. 741965. [CrossRef]
- Sharma, M., et al., A glucose-target of rapamycin signaling axis integrates environmental history of heat stress through maintenance of transcription-associated epigenetic memory in Arabidopsis. J Exp Bot, 2022. 73(20): p. 7083-7102. [CrossRef]
- Wang, C.F., et al., Plant salinity sensors: Current understanding and future directions. Front Plant Sci, 2022. 13: p. 859224. [CrossRef]
- Zhao, S., et al., Regulation of plant responses to salt stress. Int J Mol Sci, 2021. 22(9). [CrossRef]
- Deprost, D., et al., The Arabidopsis TOR kinase links plant growth, yield, stress resistance and mRNA translation. EMBO Rep, 2007. 8(9): p. 864-70. [CrossRef]
- Bakshi, A., et al., Target of Rapamycin (TOR) negatively regulates chlorophyll degradation and lipid peroxidation and controls responses under abiotic stress in Arabidopsis thaliana. Plant Stress, 2021. 2: p. 100020. [CrossRef]
- Kim, D., V.O. Ntui, and L. Xiong, Arabidopsis YAK1 regulates abscisic acid response and drought resistance. FEBS Lett, 2016. 590(14): p. 2201-2209. [CrossRef]
- Forzani, C., et al., Mutations of the AtYAK1 kinase suppress TOR deficiency in Arabidopsis. Cell Reports, 2019. 27(12): p. 3696-3708.e5. [CrossRef]
- Kavi Kishor, P.B., et al., Abscisic acid and its role in the modulation of plant growth, development, and yield stability. Trends Plant Sci, 2022. 27(12): p. 1283-1295. [CrossRef]
- Zhu, C., et al., How long should a kiss last between a kinase and its substrate? J Integr Plant Biol, 2022. 64(4): p. 789-791. [CrossRef]
- Wang, P., et al., Reciprocal regulation of the TOR kinase and ABA receptor balances plant growth and stress response. Mol Cell, 2018. 69(1): p. 100-112.e6. [CrossRef]
- Jamsheer K, M., P. Awasthi, and A. Laxmi, The social network of target of rapamycin complex 1 in plants. Journal of Experimental Botany, 2022. 73(20): p. 7026-7040. [CrossRef]
- Mugume, Y., Z. Kazibwe, and D.C. Bassham, Target of rapamycin in control of autophagy: Puppet master and signal integrator. Int J Mol Sci, 2020. 21(21). [CrossRef]
- Soto-Burgos, J. and D.C. Bassham, SnRK1 activates autophagy via the TOR signaling pathway in Arabidopsis thaliana. PLoS One, 2017. 12(8): p. e0182591. [CrossRef]
- Pu, Y., J. Soto-Burgos, and D.C. Bassham, Regulation of autophagy through SnRK1 and TOR signaling pathways. Plant Signal Behav, 2017. 12(12): p. e1395128. [CrossRef]
- Pu, Y., X. Luo, and D.C. Bassham, TOR-dependent and -independent pathways regulate autophagy in Arabidopsis thaliana. Front Plant Sci, 2017. 8: p. 1204. [CrossRef]
- Liu, Y. and D.C. Bassham, Autophagy: Pathways for self-eating in plant cells. Annu Rev Plant Biol, 2012. 63: p. 215-37. [CrossRef]
- Van Leene, J., et al., Capturing the phosphorylation and protein interaction landscape of the plant TOR kinase. Nat Plants, 2019. 5(3): p. 316-327. [CrossRef]
- Jhu, M.Y. and N.R. Sinha, Parasitic plants: An overview of mechanisms by which plants perceive and respond to parasites. Annu Rev Plant Biol, 2022. 73: p. 433-455. [CrossRef]
- Shelake, R.M., et al., Heat stress and plant-biotic interactions: Advances and perspectives. Plants (Basel), 2024. 13(15): p. 2022.
- Zhang, Y., et al., SlMYC2 interacted with the SlTOR promoter and mediated JA signaling to regulate growth and fruit quality in tomato. Front Plant Sci, 2022. 13: p. 1013445. [CrossRef]
- Farooq, M.A., et al., Jasmonic acid mediates Ca2+ dependent signal transduction and plant immunity. Plant Sci, 2024. 348: p. 112239. [CrossRef]
- Meteignier, L.V., et al., Translatome analysis of an NB-LRR immune response identifies important contributors to plant immunity in Arabidopsis. J Exp Bot, 2017. 68(9): p. 2333-2344. [CrossRef]
- Popa, C., et al., The effector AWR5 from the plant pathogen Ralstonia solanacearum is an inhibitor of the TOR signalling pathway. Sci Rep, 2016. 6: p. 27058. [CrossRef]
- Marash, I., et al., TOR inhibition primes immunity and pathogen resistance in tomato in a salicylic acid-dependent manner. Mol Plant Pathol, 2022. 23(7): p. 1035-1047. [CrossRef]
- Marash, I., et al., TOR coordinates cytokinin and gibberellin signals mediating development and defense. Plant Cell Environ, 2024. 47(2): p. 629-650. [CrossRef]
- De Vleesschauwer, D., et al., Target of rapamycin signaling orchestrates growth-defense trade-offs in plants. New Phytol, 2018. 217(1): p. 305-319. [CrossRef]
- Zhang, H., et al., Different viral effectors suppress hormone-mediated antiviral immunity of rice coordinated by OsNPR1. Nat Commun, 2023. 14(1): p. 3011. [CrossRef]
- Aznar, N.R., et al., TOR signaling downregulation increases resistance to the cereal killer Fusarium graminearum. Plant Signal Behav, 2018. 13(2): p. e1414120. [CrossRef]
- Schepetilnikov, M., et al., Viral factor TAV recruits TOR/S6K1 signalling to activate reinitiation after long ORF translation. Embo j, 2011. 30(7): p. 1343-56. [CrossRef]
- Ouibrahim, L., et al., Potyviruses differ in their requirement for TOR signalling. J Gen Virol, 2015. 96(9): p. 2898-2903. [CrossRef]
- Szwed, A., E. Kim, and E. Jacinto, Regulation and metabolic functions of mTORC1 and mTORC2. Physiol Rev, 2021. 101(3): p. 1371-1426. [CrossRef]
- Li, L., et al., Target of rapamycin controls hyphal growth and pathogenicity through FoTIP4 in Fusarium oxysporum. Mol Plant Pathol, 2021. 22(10): p. 1239-1255. [CrossRef]
- Calderan-Rodrigues, M.J., et al., Proteogenic dipeptides are characterized by diel fluctuations and target of rapamycin complex-signaling dependency in the model plant Arabidopsis thaliana. Front Plant Sci, 2021. 12: p. 758933. [CrossRef]
- da Silva, V.C.H., et al., Shedding light on the dynamic role of the "Target of Rapamycin" kinase in the fast-growing C4 species Setaria viridis, a suitable model for biomass crops. Front Plant Sci, 2021. 12: p. 637508. [CrossRef]
- Busche, M., et al., TOR coordinates nucleotide availability with ribosome biogenesis in plants. Plant Cell, 2021. 33(5): p. 1615-1632. [CrossRef]
- Mubeen, U., P. Giavalisco, and C. Caldana, TOR inhibition interrupts the metabolic homeostasis by shifting the carbon-nitrogen balance in Chlamydomonas reinhardtii. Plant Signal Behav, 2019. 14(11): p. 1670595. [CrossRef]
- Scarpin, M.R., S. Leiboff, and J.O. Brunkard, Parallel global profiling of plant TOR dynamics reveals a conserved role for LARP1 in translation. Elife, 2020. 9. [CrossRef]
- Roustan, V. and W. Weckwerth, Quantitative phosphoproteomic and system-level analysis of TOR inhibition unravel distinct organellar acclimation in Chlamydomonas reinhardtii. Front Plant Sci, 2018. 9: p. 1590. [CrossRef]
- Han, C., et al., TOR promotes guard cell starch degradation by regulating the activity of β-AMYLASE1 in Arabidopsis. Plant Cell, 2022. 34(3): p. 1038-1053. [CrossRef]
- Moreau, M., et al., Mutations in the Arabidopsis homolog of LST8/GβL, a partner of the target of Rapamycin kinase, impair plant growth, flowering, and metabolic adaptation to long days. Plant Cell, 2012. 24(2): p. 463-81. [CrossRef]
- Li, D., et al., Target of rapamycin (TOR) regulates the response to low nitrogen stress via autophagy and hormone pathways in Malus hupehensis. Hortic Res, 2022. 9: p. uhac143. [CrossRef]
- Ren, M., et al., Target of rapamycin signaling regulates metabolism, growth, and life span in Arabidopsis. Plant Cell, 2012. 24(12): p. 4850-74. [CrossRef]
- Li, L., et al., Functional characterization of target of rapamycin signaling in Verticillium dahliae. Front Microbiol, 2019. 10: p. 501. [CrossRef]
- Song, L., et al., The RALF1-FERONIA complex interacts with and activates TOR signaling in response to low nutrients. Mol Plant, 2022. 15(7): p. 1120-1136. [CrossRef]
- Mubeen, U., et al., Target of rapamycin inhibition in Chlamydomonas reinhardtii triggers de novo amino acid synthesis by enhancing nitrogen assimilation. Plant Cell, 2018. 30(10): p. 2240-2254. [CrossRef]
- Caldana, C., et al., Systemic analysis of inducible target of rapamycin mutants reveal a general metabolic switch controlling growth in Arabidopsis thaliana. Plant J, 2013. 73(6): p. 897-909. [CrossRef]
- Forzani, C., et al., Mutations of the AtYAK1 kinase suppress TOR deficiency in Arabidopsis. Cell Rep, 2019. 27(12): p. 3696-3708.e5. [CrossRef]
- O'Leary, B.M., et al., Metabolite regulatory interactions control plant respiratory metabolism via target of rapamycin (TOR) kinase activation. Plant Cell, 2020. 32(3): p. 666-682. [CrossRef]
- Artins, A., et al., Sensing and regulation of C and N metabolism-novel features and mechanisms of the TOR and SnRK1 signaling pathways. Plant J, 2024. 118(5): p. 1268-1280. [CrossRef]
- Ding, Y. and S. Yang, Surviving and thriving: How plants perceive and respond to temperature stress. Dev Cell, 2022. 57(8): p. 947-958. [CrossRef]
- Jancewicz, A.L., N.M. Gibbs, and P.H. Masson, Cadaverine's functional role in plant development and environmental response. Front Plant Sci, 2016. 7: p. 870. [CrossRef]
- Ozmen, S., S. Tabur, and S. Oney-Birol, Alleviation role of exogenous cadaverine on cell cycle, endogenous polyamines amounts and biochemical enzyme changes in barley seedlings under drought stress. Scientific Reports, 2023. 13(1): p. 17488. [CrossRef]
- You, J., et al., Transcriptomic and metabolomic profiling of drought-tolerant and susceptible sesame genotypes in response to drought stress. BMC Plant Biol, 2019. 19(1): p. 267. [CrossRef]
- Barqawi, A.A. and A.A. Abulfaraj, Salt stress-related mechanisms in leaves of the wild barley Hordeum spontaneum generated from RNA-Seq datasets. Life (Basel), 2023. 13(7). [CrossRef]
- Brocker, C., et al., Aldehyde dehydrogenase 7A1 (ALDH7A1) attenuates reactive aldehyde and oxidative stress induced cytotoxicity. Chem Biol Interact, 2011. 191(1-3): p. 269-77. [CrossRef]
- Masclaux-Daubresse, C., et al., Stitching together the multiple dimensions of autophagy using metabolomics and transcriptomics reveals impacts on metabolism, development, and plant responses to the environment in Arabidopsis. Plant Cell, 2014. 26(5): p. 1857-1877. [CrossRef]
- Huh, S.U., et al., ATAF2, a NAC transcription factor, binds to the promoter and regulates NIT2 gene expression involved in auxin biosynthesis. Mol Cells, 2012. 34(3): p. 305-13. [CrossRef]
- Kwon, Y.S., et al., Proteomic analyses of the interaction between the plant-growth promoting rhizobacterium Paenibacillus polymyxa E681 and Arabidopsis thaliana. Proteomics, 2016. 16(1): p. 122-35. [CrossRef]
- Luo, K., et al., Indole-3-acetic acid in Fusarium graminearum: Identification of biosynthetic pathways and characterization of physiological effects. Fungal Biol, 2016. 120(9): p. 1135-45. [CrossRef]
- Alsherif, E.A., et al., How carbon nanoparticles, arbuscular mycorrhiza, and compost mitigate drought stress in maize plant: A growth and biochemical study. Plants (Basel), 2022. 11(23). [CrossRef]
- Hijaz, F. and N. Killiny, Exogenous GABA is quickly metabolized to succinic acid and fed into the plant TCA cycle. Plant Signal Behav, 2019. 14(3): p. e1573096. [CrossRef]
- Zhou, H., et al., Potassium indole-3-butyric acid affects rice's adaptability to salt stress by regulating carbon metabolism, transcription factor genes expression, and biosynthesis of secondary metabolites. Front Plant Sci, 2024. 15: p. 1416936. [CrossRef]
- Yang, L., et al., Hydroxycoumarins: New, effective plant-derived compounds reduce Ralstonia pseudosolanacearum populations and control tobacco bacterial wilt. Microbiol Res, 2018. 215: p. 15-21. [CrossRef]
- Wang, Y., et al., 5-ALA, DTA-6, and nitrogen mitigate NaCl stress by promoting photosynthesis and carbon metabolism in rice seedlings. Metabolites, 2024. 14(3): p. 142.
- Rhaman, M.S., et al., 5-aminolevulinic acid-mediated plant adaptive responses to abiotic stress. Plant Cell Rep, 2021. 40(8): p. 1451-1469. [CrossRef]
- Tan, S., et al., Advances in 5-aminolevulinic acid priming to enhance plant tolerance to abiotic stress. Int J Mol Sci, 2022. 23(2). [CrossRef]
- El-Shora, H.M., et al., Alleviation of lead stress on sage plant by 5-aminolevulinic acid (ALA). Plants (Basel), 2021. 10(9). [CrossRef]
- Yang, Y., et al., Copper stress in grapevine: Consequences, responses, and a novel mitigation strategy using 5-aminolevulinic acid. Environ Pollut, 2022. 307: p. 119561. [CrossRef]
- Yang, Y., et al., Drought stress in 'Shine Muscat' grapevine: Consequences and a novel mitigation strategy-5-aminolevulinic acid. Front Plant Sci, 2023. 14: p. 1129114. [CrossRef]
- Helaly, M.N., et al., 5-aminolevulinic acid and 24-epibrassinolide improve the drought stress resilience and productivity of banana plants. Plants (Basel), 2022. 11(6): p. 743.
- Yu, J., et al., Metabolic pathways involved in carbon dioxide enhanced heat tolerance in bermudagrass. Front Plant Sci, 2017. 8: p. 1506. [CrossRef]
- Lei, S., et al., Metabolic regulation of 5-oxoproline for enhanced heat tolerance in perennial ryegrass. Stress Biol, 2024. 4(1): p. 46. [CrossRef]
- Wang, Y., et al., Metabolite profiling in two contrasting Tibetan hulless barley cultivars revealed the core salt-responsive metabolome and key salt-tolerance biomarkers. AoB Plants, 2019. 11(2): p. plz021. [CrossRef]
- Bruňáková, K., et al., Does phenotyping of Hypericum secondary metabolism reveal a tolerance to biotic/abiotic stressors? Front Plant Sci, 2022. 13: p. 1042375. [CrossRef]
- Wu, Z., et al., Study of dandelion (Taraxacum mongolicum Hand.-Mazz.) salt response and caffeic acid metabolism under saline stress by transcriptome analysis. Genes (Basel), 2024. 15(2). [CrossRef]
- Ramzan, M., et al., Potential of kaempferol and caffeic acid to mitigate salinity stress and improving potato growth. Sci Rep, 2024. 14(1): p. 21657. [CrossRef]
- Li, J., et al., Strigolactone enhances tea plant adaptation to drought and Phyllosticta theicola petch by regulating caffeine content via CsbHLH80. Plant Physiol Biochem, 2024. 216: p. 109161. [CrossRef]
- Yoo, Y., et al., Caffeine produced in rice plants provides tolerance to water-deficit stress. Antioxidants (Basel), 2023. 12(11). [CrossRef]
- Zhang, N., et al., Constitutive camalexin production and environmental stress response variation in Arabidopsis populations from the Iberian Peninsula. Plant Sci, 2014. 225: p. 77-85. [CrossRef]
- Xiao, P., et al., Transcriptome and metabolome atlas reveals contributions of sphingosine and chlorogenic acid to cold tolerance in Citrus. Plant Physiol, 2024. 196(1): p. 634-650. [CrossRef]
- Piasecka, A., et al., Phenolic metabolites from barley in contribution to phenome in soil moisture deficit. Int J Mol Sci, 2020. 21(17). [CrossRef]
- Dias, M.C., et al., Phenolic and lipophilic metabolite adjustments in Olea europaea (olive) trees during drought stress and recovery. Phytochemistry, 2021. 185: p. 112695. [CrossRef]
- Lacrampe, N., et al., Nitrogen-mediated metabolic patterns of susceptibility to Botrytis cinerea infection in tomato (Solanum lycopersicum) stems. Planta, 2023. 257(2): p. 41. [CrossRef]
- Silambarasan, S., et al., Co-application of citric acid and Nocardiopsis sp. strain RA07 enhances phytoremediation potentiality of Sorghum bicolor L. Environ Sci Pollut Res Int, 2023. 30(36): p. 86244-86254. [CrossRef]
- Menhas, S., et al., Citric acid-driven cadmium uptake and growth promotion mechanisms in Brassica napus. Chemosphere, 2024. 368: p. 143716. [CrossRef]
- Cay, S., Assessment of tea saponin and citric acid-assisted phytoextraction of Pb-contaminated soil by Salvia virgata Jacq. Environ Sci Pollut Res Int, 2023. 30(17): p. 49771-49778. [CrossRef]
- Kaya, C., et al., Citric acid and hydrogen sulfide cooperate to mitigate chromium stress in tomato plants by modulating the ascorbate-glutathione cycle, chromium sequestration, and subcellular allocation of chromium. Environ Pollut, 2023. 335: p. 122292. [CrossRef]
- Tahjib-Ul-Arif, M., et al., Citric acid-mediated abiotic stress tolerance in plants. Int J Mol Sci, 2021. 22(13). [CrossRef]
- Święcicka, M., et al., Changes in benzoxazinoid contents and the expression of the associated genes in rye (Secale cereale L.) due to brown rust and the inoculation procedure. PLoS One, 2020. 15(5): p. e0233807. [CrossRef]
- Zhang, X., et al., Mechanisms of resistance to spot blotch in Yunnan iron shell wheat based on metabolome and transcriptomics. Int J Mol Sci, 2022. 23(9). [CrossRef]
- Rao, M.J., et al., LC-MS/MS-based metabolomics approach identified novel antioxidant flavonoids associated with drought tolerance in citrus species. Front Plant Sci, 2023. 14: p. 1150854. [CrossRef]
- Kim, S.E., et al., Overexpression of 4-hydroxyphenylpyruvate dioxygenase (IbHPPD) increases abiotic stress tolerance in transgenic sweetpotato plants. Plant Physiol Biochem, 2021. 167: p. 420-429. [CrossRef]
- Mata-Pérez, C., et al., Protein tyrosine nitration during development and abiotic stress response in plants. Front Plant Sci, 2016. 7: p. 1699. [CrossRef]
- Mishra, S., A. Sharma, and A.K. Srivastava, Ascorbic acid: a metabolite switch for designing stress-smart crops. Crit Rev Biotechnol, 2024. 44(7): p. 1350-1366. [CrossRef]
- Biswas, S., A.K. Biswas, and B. De, Influence of sodium chloride on growth and metabolic reprogramming in nonprimed and haloprimed seedlings of blackgram (Vigna mungo L.). Protoplasma, 2020. 257(6): p. 1559-1583. [CrossRef]
- Li, X., et al., Integrative physiological, metabolomic, and transcriptomic analysis reveals the drought responses of two apple rootstock cultivars. BMC Plant Biol, 2024. 24(1): p. 219. [CrossRef]
- Zeier, J., New insights into the regulation of plant immunity by amino acid metabolic pathways. Plant Cell Environ, 2013. 36(12): p. 2085-103. [CrossRef]
- Zou, Y.N., et al., Metabolomics reveals arbuscular mycorrhizal fungi-mediated tolerance of walnut to soil drought. BMC Plant Biol, 2023. 23(1): p. 118. [CrossRef]
- Wang, Y., et al., Transcriptomics and metabolomics revealed that phosphate improves the cold tolerance of alfalfa. Front Plant Sci, 2023. 14: p. 1100601. [CrossRef]
- Castro-Vázquez, L., et al., Pressurized extraction as an opportunity to recover antioxidants from orange peels: Heat treatment and nanoemulsion design for modulating oxidative stress. Molecules, 2021. 26(19). [CrossRef]
- Lwalaba, J.L.W., et al., Transcriptome analysis reveals the tolerant mechanisms to cobalt and copper in barley. Ecotoxicol Environ Saf, 2021. 209: p. 111761. [CrossRef]
- Jin, S., et al., Accumulation of hydroxycinnamic acid amides in winter wheat under snow. Biosci Biotechnol Biochem, 2003. 67(6): p. 1245-9. [CrossRef]
- Neamțu, A.A., et al., A comprehensive view on the impact of chlorogenic acids on colorectal cancer. Curr Issues Mol Biol, 2024. 46(7): p. 6783-6804. [CrossRef]
- Huo, X., et al., Effect of S-Allyl-L-Cysteine on nitric oxide and Cadmium processes in Rice (Oryza sativa L. sp. Zhongzao35) seedlings. Toxics, 2024. 12(11). [CrossRef]
- Cheng, L.L., et al., Mechanism of S-allyl-L-cysteine alleviating Cadmium stress in seedling roots and buds of rice seedlings. Huan Jing Ke Xue, 2021. 42(6): p. 3037-3045. [CrossRef]
- Owusu, A.G., et al., Transcriptomic and metabolomic analyses reveal the potential mechanism of waterlogging resistance in cotton (Gossypium hirsutum L.). Front Plant Sci, 2023. 14: p. 1088537. [CrossRef]
- Zhuang, Q., et al., Joint transcriptomic and metabolomic analysis reveals the mechanism of low-temperature tolerance in Hosta ventricosa. PLoS One, 2021. 16(11): p. e0259455. [CrossRef]
- Zhou, C., et al., Novel finding on how melatonin and nanoselenium alleviate 2,4-D butylate stress in wheat plants. J Agric Food Chem, 2023. 71(35): p. 12943-12957. [CrossRef]
- Dong, X., et al., Multiomics analyses reveal MsC3H29 positively regulates flavonoid biosynthesis to improve drought resistance of autotetraploid cultivated alfalfa (Medicago sativa L.). J Agric Food Chem, 2024. 72(25): p. 14448-14465. [CrossRef]
- Kasthuri, T., et al., Proteomic profiling spotlights the molecular targets and the impact of the natural antivirulent umbelliferone on stress response, virulence factors, and the quorum sensing network of Pseudomonas aeruginosa. Front Cell Infect Microbiol, 2022. 12: p. 998540. [CrossRef]
- Beesley, A., et al., Engineered coumarin accumulation reduces mycotoxin-induced oxidative stress and disease susceptibility. Plant Biotechnol J, 2023. 21(12): p. 2490-2506. [CrossRef]
- Zhou, Y., et al., Transcriptome sequencing and metabolome analysis reveal the molecular mechanism of Salvia miltiorrhiza in response to drought stress. BMC Plant Biol, 2024. 24(1): p. 446. [CrossRef]
Figure 1.
TOR regulates a large number of stress-responsive genes. (a) Structure and complex of TOR kinase in plants. (b) Overlapping genes between TOR-regulated and biotic stress-responsive genes. (c) Overlapping genes between TOR-regulated and abiotic stress-responsive genes. (d) KEGG analysis of overlapping genes in (b). (e) KEGG analysis of overlapping genes in (c).
Figure 1.
TOR regulates a large number of stress-responsive genes. (a) Structure and complex of TOR kinase in plants. (b) Overlapping genes between TOR-regulated and biotic stress-responsive genes. (c) Overlapping genes between TOR-regulated and abiotic stress-responsive genes. (d) KEGG analysis of overlapping genes in (b). (e) KEGG analysis of overlapping genes in (c).
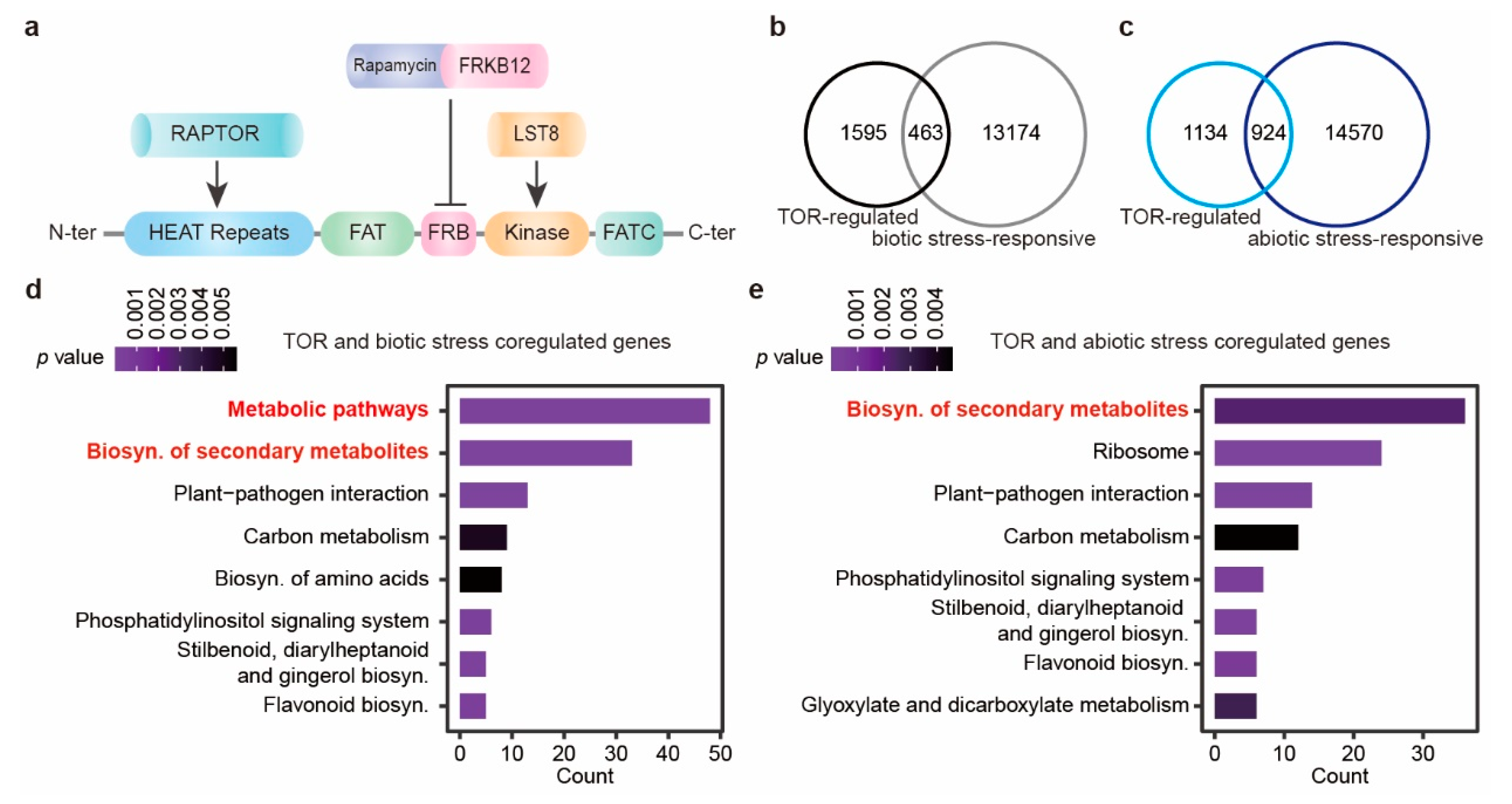
Figure 2.
TOR-mediated metabolic reprogramming enhances plant resilience against diverse abiotic and biotic stresses. Plants, as sessile organisms, are continuously exposed to a wide spectrum of environmental challenges. These challenges encompass both abiotic stresses, including salinity, drought, oxidative conditions, and extreme temperatures, and biotic stresses from various organisms such as microbes, pathogens, nematodes, herbivores, and parasites. Within the complex plant signaling network, TOR functions as a central regulatory hub that orchestrates metabolic reprogramming to enhance stress resilience. This master regulator integrates diverse upstream stress signals and modulates plant metabolism through multiple mechanisms. Specifically, TOR regulates transcriptional responses to enhance the production of stress-resistant metabolites, directly modifies the activity of key metabolic enzymes across various pathways, and employs additional regulatory mechanisms. Consequently, the accumulated stress-resistant compounds establish a protective biochemical “shield”, thereby significantly enhancing plant resilience against multiple environmental stressors.
Figure 2.
TOR-mediated metabolic reprogramming enhances plant resilience against diverse abiotic and biotic stresses. Plants, as sessile organisms, are continuously exposed to a wide spectrum of environmental challenges. These challenges encompass both abiotic stresses, including salinity, drought, oxidative conditions, and extreme temperatures, and biotic stresses from various organisms such as microbes, pathogens, nematodes, herbivores, and parasites. Within the complex plant signaling network, TOR functions as a central regulatory hub that orchestrates metabolic reprogramming to enhance stress resilience. This master regulator integrates diverse upstream stress signals and modulates plant metabolism through multiple mechanisms. Specifically, TOR regulates transcriptional responses to enhance the production of stress-resistant metabolites, directly modifies the activity of key metabolic enzymes across various pathways, and employs additional regulatory mechanisms. Consequently, the accumulated stress-resistant compounds establish a protective biochemical “shield”, thereby significantly enhancing plant resilience against multiple environmental stressors.
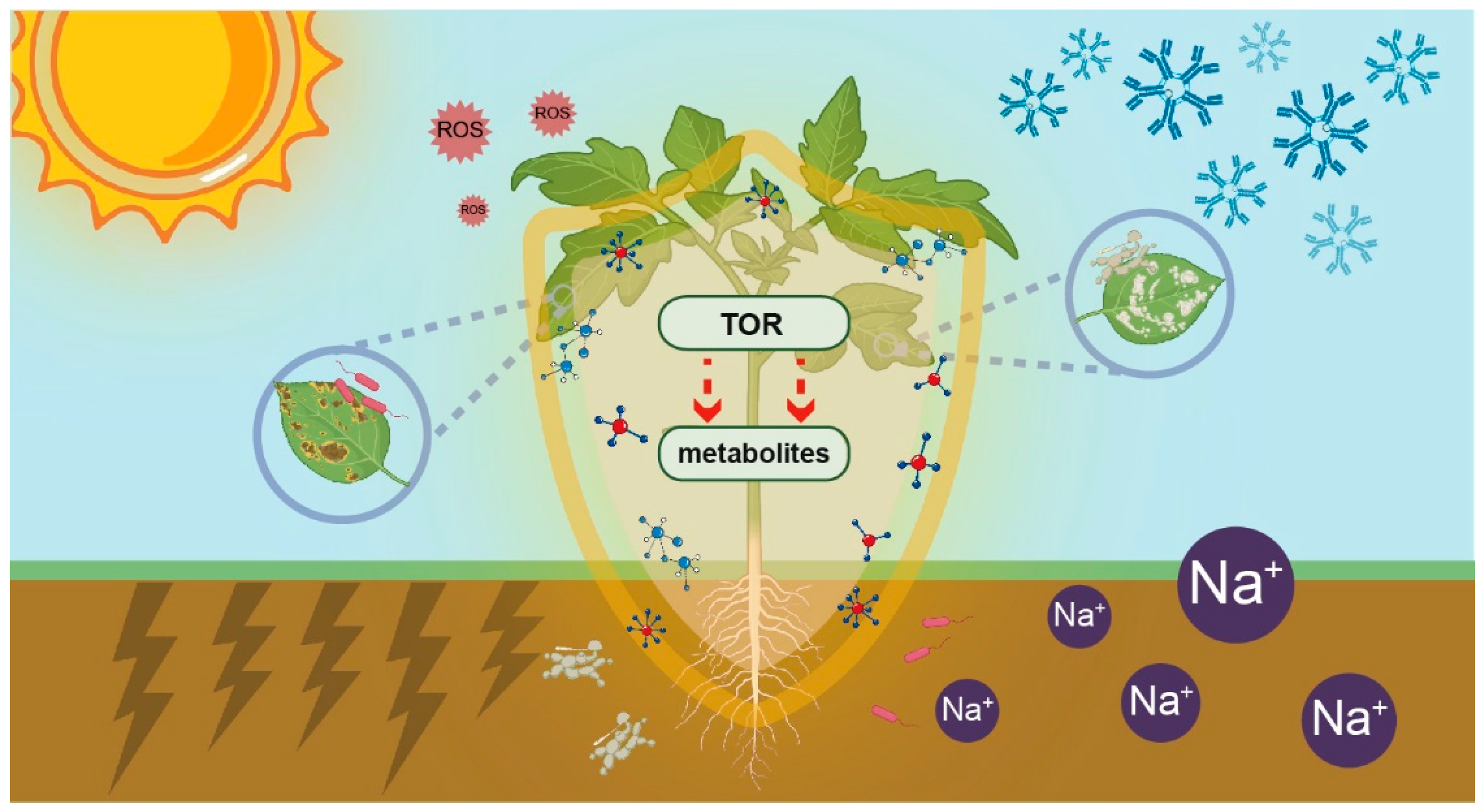

Table 1.
TOR-mediated stress-related DAMs in Arabidopsis. The data were originated from Song et al. [84], and the functions of the 33 stress-related DAMs were determined through comprehensive literature analysis.
Table 1.
TOR-mediated stress-related DAMs in Arabidopsis. The data were originated from Song et al. [84], and the functions of the 33 stress-related DAMs were determined through comprehensive literature analysis.
| Compounds | Class | Up- or Down-regulated in tor-es | Function | References |
|---|---|---|---|---|
| 1,5-Diaminopentane | Phenolamides | up | Drought, oxidative stress, heavy metal stress | [91,92] |
| 2-Aminoadipic acid (L-Homoglutamic acid) | Amino acid and derivatives | up | Oxidative stress, drought, salt | [93,94,95,96] |
| 3-Indoleacetonitrile | Indole derivatives | down | Pathogen attack | [97,98,99] |
| 3-Methylmalic acid | Organic acids and derivatives | up | Salt, drought, biotic stress | [100,101,102] |
| 4-Hydroxycoumarin | Phenylpropanoids | down | Pathogen attack | [103] |
| 5-Aminolevulinate | Organic acids and derivatives | up | Drought, salt, heavy metal | [104,105,106,107,108,109,110] |
| 5-Oxoproline | Amino acid and derivatives | up | Heat stress | [111,112] |
| 6-Aminocaproic acid | Organic acids and derivatives | up | Salt stress | [113] |
| Amentoflavone | Flavone | up | Temperature, light, drought, biotic stress | [114] |
| Caffeic acid | Phenylpropanoids | up | Salt stress | [115,116] |
| Caffeine | Alkaloids | down | Drought, biotic stress | [117,118] |
| Camalexin | Alkaloids | up | Pathogen attack | [119] |
| Chlorogenic acid (3-O-Caffeoylquinic acid) | Organic acids and derivatives | down | Cold stress | [120] |
| Chrysoeriol | Flavone | up | Oxidative stress | [121,122] |
| Citramalate | Organic acids and derivatives | up | Pathogen attack | [123] |
| Citric acid monohydrate | Organic acids and derivatives | up | Heavy metal | [124,125,126,127,128] |
| Citric acid | Organic acids and derivatives | up | Heavy metal | [124,125,126,127,128] |
| DIMBOA glucoside | Others | up | Biotic stress | [129,130] |
| Genistein 7-O-Glucoside (Genistin) | Isoflavone | up | Drought | [131] |
| Homogentisic acid | Organic acids and derivatives | up | Abiotic stress, ABA signalling | [132] |
| L-(-)-Tyrosine | Amino acid and derivatives | up | Abiotic stress | [133] |
| L-Ascorbate | Vitamins and derivatives | up | Abiotic stress | [134] |
| L-Homoserine | Amino acid and derivatives | up | Drought, salt stress | [135,136] |
| L-Pipecolic acid | Amino acid and derivatives | up | Biotic stress | [137] |
| N-Acetyl-L-phenylalanine | Amino acid and derivatives | up | Cold, drought | [138,139] |
| Narirutin | Flavone | up | Heavy metal stress, oxidative stress | [140,141] |
| N-p-Coumaroyl agmatine | Phenolamides | up | Biotic stress | [142] |
| Quinic acid | Organic acids and derivatives | up | Abiotic stress | [143] |
| S-Allyl-L-cysteine | Amino acid and derivatives | up | Heavy metal stress | [144,145] |
| Sinapyl alcohol | Phenylpropanoids | up | Flooding stress, cold stress | [146,147] |
| Tricin | Flavone | down | Cold, drought, salt stress | [148,149] |
| Umbelliferone | Phenylpropanoids | down | Pathogen attack | [150,151] |
| Xanthohumol | Flavanone | up | Drought stress | [152] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



