
Submitted:
20 January 2025
Posted:
21 January 2025
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
The lipid nanoparticle (LNP)-enclosed mRNA-containing COVID-19 vaccines were used successfully to vaccinate billions of people during the COVID-19 pandemic. However, as with all medicines, Comirnaty and Spikevax can also cause adverse events (AEs) and complications. Although such events occur in less than 0.5% of vaccinated individuals, the huge scale of global vaccination means that the number of "vaccine injuries" could reach millions worldwide. These AEs have a uniquely broad spectrum affecting multiple organs, with most cases being associated with inflammatory and autoimmune processes. Yet, the clinical significance of AEs is debated, and their cellular and molecular mechanisms are poorly understood. The present review surveys the distinctive structural and functional features of these vaccines linking them to secondary immune effects causing unusual AEs/complications. The discussed unique mRNA-LNP properties include the ribosomal synthesis of the spike protein (SP) fundamentally transforming antigen processing and presentation; the multiple chemical modifications of the mRNA, increasing its stability and translation efficacy; toxicity of the SP casing multiorgan damage; immune stimulation and mRNA transfection by LNP; reduced stability of vaccine nanoparticles in water; immune reactivity and immunogenicity of PEG on the LNP surface; stabilization of the SP by enrichment with proline; and contaminations of the vaccine with plasmid DNA and inorganic elements or complexes. The considered collateral immune effects that may theoretically underlie the AEs are: diversification of the processing and presentation of the SP, innate immune activation, T-cell and antibody-mediated cytotoxicities, dissemination of virus/vaccine hybrid exosomes, somatic hypermutation, reverse transcription, insertion mutagenesis and frameshift mutation. Highlighting these potential processes may help update the risk/benefit ratio of the currently used formulations and make future products of the mRNA-LNP technology platform safer.
Keywords:
LNP
; lipid nanoparticles
; COVID-19 mRNA vaccines
; Comirnaty
; Spikevax
; vaccine complications
; adverse events
; gene therapy
; immunotherapy
; pandemics
1. Introduction
Since the introduction of mRNA-based COVID-19 vaccines in December 2020, Pfizer-BioNTech's Comirnaty and Moderna's Spikevax, >5 billion doses have been administered globally. This success has accelerated the adoption of mRNA-containing lipid nanoparticle (mRNA-LNP) technology for developing novel therapies against infectious diseases, metabolic disorders, cancer and many other diseases. However, aside from their remarkable success and promise, mRNA vaccines also face challenges, such as the emergence of an unusually broad spectrum of adverse events (AEs) and long-term complications, collectively referred to as "post-vaccination syndrome" (PVS) [1,2,3,4,5,6]. A fraction of these AEs, referred to as AEs of “Special Interest,” has been distinguished by the “Brighton Collaboration,” an international team of vaccine experts, as being also typical of COVID-19 and post-COVID syndrome [7,8,9,10,11]. Hence, these mostly severe AEs are often referred to as “Brighton-case AEs,” but this review, based on the commonalities with the mechanism of SARS-CoV-2 infection, makes the nomenclature clearer and calls them “Covid-like” vaccine AEs. Hundreds of studies address the clinical picture, statistics, and the mechanism of Covid-like vaccine AEs, and the issue polarizes opinions far beyond the scientific problem. Many studies focus on a key role of the spike protein (SP) as a common denominator in the virus and vaccine pathology [12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30], but there are much more factors contributing to the problem. There is clearly a need to further investigate the root causes that may explain all Covid-like AEs, broadening the scope to include potential conceptual challenges.
2. COVID-19 mRNA-LNP Vaccines: Collateral Immune Effects Beyond Antiviral Defense
The mRNA-LNPs represent a significant milestone in combining nanotechnology with genetic engineering. Unlike traditional vaccines, which initiate an immune response following immunization with protein or peptide antigens, using LNPs as delivery vehicles for the mRNA vaccines deliver the genetic code for the antigen, specifically, the SP of the SARS-CoV-2 virus. These lipid nanoparticles (NPs) ensure efficient cellular uptake of the mRNA, while stabilizing and protecting it from enzymatic degradation. Once the mRNA-LNPs are captured by antigen-presenting cells (APCs) and escape the endo-lysosomes, the mRNA is translated by ribosomes into the SP, which triggers the anti-viral immune response. This response is amplified relative to other pathogens’ immunogenicity through enhanced and extended production of the antigen, due to the mRNA modification and the LNP’s super-adjuvant, pro-inflammatory actions. Beyond these innovations in immunogenicity, it is not widely recognized that mRNA-LNPs can have further unique immune effects specific to this vaccine, which may sometimes play a role in causing PVS.
One of the potential extra capabilities of mRNA vaccines is the extension of the antiviral immune response against uninfected host cells that also express the SP, which may cause autoimmune AEs. The latter effect is achieved through rapid distribution of the mRNA-LNP over the body and nonspecific transfection of cells that are accessible for LNP fusion. Yet another extra immune activity is a coincidental immune reactivity and immunogenicity of the LNP, itself [31], due to the presence of PEG on the nanoparticle surface. This lends an intrinsic pathogenicity to the mRNA-LNPs. Perhaps the most surprising potential additional action of mRNA-LNPs is the generation of a simplified version of the SARS-CoV-2 virus through the secretion of mRNA-containing SP-expressing exosomes by all mRNA-LNP-transfected secretory cells. These “pseudo-viruses” may potentially spread the infection just like the parent virus. All these extra features make Comirnaty and Spikevax vaccines combo immune modifier nanoparticles with multiple functions beyond the antiviral protection.
Realization of this complexity led some to question the accuracy of reference to COVID-19 mRNA-LNPs as vaccines in the traditional sense, since traditional vaccines do not cause so many different immune effects. Accordingly, alternative terminologies for mRNA vaccines have surfaced, such as “mRNA therapeutics” [32,33,34], however, this term disregards the complex nanoparticle nature the vaccine, as the mRNA is not the only active pharmacological ingredient. The widespread “genetic vaccine” expression also has a glitch in logic, since these vaccines are only partially products of genetic engineering, the LNP component is a creation of nanotechnology. The term "Prophylactic immuno-gene therapy [35] is based on the use of mRNA vaccines in immunotherapy [36] and the overlapping AEs with those of the immune therapies [37,38,39]. The expression highlights the fact that these vaccines are immune function modifiers via genetic manipulation, with a unique benefit of reducing the severity of COVID-19 when the infection with the real virus occurs.
3. The Symptoms, Incidence and Prevalence of Adverse Events and Deaths: Limitations of Statistics
Supplementary Table S1 lists the Covid-like AEs classified by affected organ systems, also giving a non-exhaustive list of related references. The compilation demonstrates the uniquely broad spectrum of symptoms impacting nearly all organ systems.
Regarding the incidence and prevalence of AEs, defined as the number of AE events or reactors related to the overall number of vaccine shots given, while prevalence is the cumulative number of AEs during a certain time-window, a recent snapshot of the Vaccine Adverse Event Reporting System (VAERS) in the United States [40] showed 1,656,138 AE reports of COVID-19 vaccine-related one or more symptoms through November 29, 2024, out of a total of approximately 2.6 million (M) AE reports in the system. This implies that, in less than four years, COVID-19 vaccines accounted for about 63% of all AEs reports registered in VAERS since its inception in 1990. Importantly, in the U.S., 99.9% of administered COVID-19 vaccines were mRNA-based, specifically Comirnaty and Spikevax. However, the VAERS does not give information on the number of vaccine shots, which is provided by another public database, the “Our World in Data (OWID)” [41,42].
According to OWID, 654 M doses of mRNA COVID-19 vaccines were administered in the U.S. as of August 10, 2024 (~402+252 M Pfizer + Moderna) [43]. Assuming 2 vaccinations for all vaccinee, this number of shots suggests ~327 M mRNA vaccine recipients. Disregarding the 4 months’ sampling time difference, the overall AE incidence rate is approximately 0.5 %, or 1 AE in ~200 vaccine recipients. Applying this rate to the 327 M vaccine recipients, an estimated 1.6 M people developed more or less severe AEs in the USA. For the 27 countries of the European Union (EU), using the prevalence rate established for the USA and approximately 826.2 M vaccine doses [43] the cumulative total number of AE-inflicted people in the EU is roughly 2.1 M.
Regarding the suspected lethal outcomes of all Covid-19 vaccines, the VAERS reported 37,544 deaths between 1990 and 2024 April, vs 10,760 cases caused by all other vaccines at the same time, a >3-fold increase [44]. However, comparing only with flu, the increase was >200-fold [35] (see below).
A further consideration is that the VAERS does not stratify the AE reports according to severity, and the symptoms can vary over a broad range from transient inconvenience to death [45]. From a medical aspect, however, it is important to distinguish the severe, chronic, occasionally disabling COVID-19 like AEs among all AEs, contributing to the clinical picture of PVS. According to a statistic from the Paul Erlich Institute in Germany, the incidence of severe AEs is about 0.02 % of all COVID-19 vaccine doses [46], which brings down the prevalence of COVID-like severe AEs by a factor of 25, i.e., ~66,000 and ~83,000 in the US and Europe, respectively.
It should be pointed out regarding the above VAERs-based calculations that there are at least 3 critical, yet uncertain factors influencing these estimations. One is the variation of AE incidence rates, depending on vaccine version, batch, and timing of administration. For example, the VAERS-reported rate was 0.03% as of March 2021, that rose to 0.17% as of May 2023 [35], and further to 0.5%, as mentioned above for November 2024. A second is the ratio of vaccinated people to the administered vaccine doses, in the range of 1 and 1/4. The 0.5% incidence rate estimate used ½, i.e., 2 vaccines for all vaccinees. A third uncertainty is the underreporting factor (URF) in VAERS. Studies have shown different sensitivities for detecting different AEs, varying between 12% and 76% with URFs ranging from 1.3 to 8.3. For COVID-19 vaccines, the UEF was estimated much higher; specifically, 41. For all these uncertainties, the above Figures should be viewed as rough estimates. What they prove is that COVID-19 vaccines, due to the immense number of injections, can be associated with a very high number of AEs in non-COVID-19 infected, mostly healthy people.
One of the greatest challenges in analyzing statistics for vaccine doses, which account for roughly 65% of the global population [5.3/8.2 B], is that the small percentage of incidence rates and the vast prevalence Figures are beyond what can be reasonably comprehended. In particular, it is not possible to see whether these AEs are expectable, implying tolerability, or unusual, implying extra risk. For this reason, a comparison of mRNA vaccine AE incidence with that of flu vaccines, also used again a coronavirus in billions of people, may be more relevant. Such analysis was recently performed by comparing the cumulative AE reports for mRNA vaccines over the past 4 years with that of 12 flu vaccines combined [35]. The data revealed an average 26-fold increase in AEs associated with Covid-19 mRNA vaccines, the disparity being particularly significant, >100-fold in the case of Covid-like AEs. For example, the mRNA-LNP-associated rise of myocarditis, thrombosis, death, myocardial infarction, tachycardia, dyspnea and hypertension were 1,152x, 455x, 226x, 218x, 162x, 152x), and 131x, respectively, where "x" means x-fold higher incidence rate with mRNA vaccines compared to flu [35].
Excess mortality in the fully vaccinated, developed countries over the past 4 years is one of the most contentious issues regarding the COVID-19 vaccine campaigns, debating on the question whether mRNA vaccines contributed to this phenomenon. There are many estimates in this regard [47,48,49,50,51], but in the absence of stratifying the post-vaccination death reports according to age and comorbidity, it is very difficult, if possible at all, to establish direct causality. As with the AE prevalence, an alternative, more palpable quantitative indicator of the vaccine’s lethality is the ratio of VAERS-listed COVID-19 vaccine-related death (~37,400) to that caused by all other vaccines over the past 4 years, which ratio is, according to Rhodes et al., [52], 28-fold. As mentioned above, the average increase of incidence rate of all AEs of COVID-19 vaccines compared to flu, was 26-fold. This remarkable coincidence with the 28-fold increase of death rate highlights the correlation between AEs and fatal outcome of vaccination, also providing indirect support for a causal relationship between vaccine-induced AEs and excess death.
4. Unique Structural Features of Comirnaty and Spikevax
In January 2020 Chinese scientists released the genetic sequence of SARS-CoV-2, and both Comirnaty and Spikevax were developed and achieved emergency use authorization in the same year. They have essentially similar dense spherical LNPs in the 60-120 nm diameter range, and in both, the payload mRNA codes for the full-length, prefusion-stabilized SP of the virus. Nevertheless, their final formulations are not identical [53]. In particular, the proprietary ionizable lipid components (e.g., ALC-0315 in Comirnaty and SM-102 in Spikevax) differ in their pKa and hydrophobic tail length and saturation, which impact their stability and, hence, storage requirements. In the PEGylated lipids, the PEG chain is covalently attached to a proprietary lipid (ALC-0159) in Comirnaty, while in Spikevax, the PEG is attached to 1,2-dimyristoyl-sn-glycero-3-phosphoethanolamine (DMG), a natural phospholipid. The buffer systems also differ: Comirnaty uses a phosphate-based buffer, while Spikevax uses a tromethamine-based buffer. These formulation differences may influence the stability and metabolism of these vaccines. However, the most important difference is likely the mRNA dose. Spikevax contains 3.3 times more mRNA (100 µg) compared to Comirnaty (30 µg in adult doses), which is likely the most important cause of differences in efficacy and toxicity between the two vaccines.
Focusing on Comirnaty, its nanoscale structure, like that of Spikevax, has several unique properties. Unlike bilayer liposomes, the outer coat of the vaccine nanoparticles is variable. Some studies show monolayers, others one or multiple bilayers [54], and yet others intermittent mono and bilayers [55]. Moreover, the LNPs tend to acquire a dual-lobed appearance due to bleb formation [54,56,57], a sign of partition of internal content. This variability implies instability, one cause of AEs, as discussed later.
The discontinuous outer phospholipid lipid layer with its loosely bound PEG lipid may be major factors lending instability to the mRNA-LNPs, which is reflected in the short shelve-live of injectable water-diluted stock (12 h) on room temperature, and the need for ultra-low temperature (-90oC to -60oC) for up to 1.5-year storage. Consistent with the instability, an AFM study showed Comirnaty to contain soft, highly compliant unstable nanoparticles [55]. The possible role of instability in AEs will be discussed later.
Regarding the internal, electrodense core of the LNPs [58], the spatial relationship of lipids and the mRNA has not been conclusively established. The available schematic visualizations of mRNA-LNPs, often mistaken with the model of small inhibitory (si)RNA-containing LNPs, suggest that the polyanionic mRNA binds to the positively charged ionizable lipids (ALC-0315) inside reverted micelles formed by these bipolar lipids (Figure 1A). Yet, in alternative portrayals, the lipids cover the undulant mRNA as sheaths (Figure 1B). or the mRNA is randomly “diluted” in a lipid emulsion (Figure 1 C). However, the mRNA’s size of ~1,414 kDa consisting of 4,284 nucleotides with an extended length of ~1,500 nm cannot be reconciled with a linear or undulant polymer; the mRNA must wind up like thread balls to fit the available internal space of the LNP (Figure 1D). This requirement is met in a recent alternative Comirnaty structure model (Figure 1E), based on electron micrographs showing 2−5 nm electron-dense spots in the LNP's interior aligned into strings, semicircles, or labyrinth-like networks (Figure 1F). It was proposed that ionizable lipid stacks crosslink the winding mRNA loops in a granular mesh via hydrogen bonds rather than ionic forces [55]. This proposal was supported Unruh et al. who found Comirnaty LNPs in a “liquid state” [59]. Figure 1G and H show the disintegration of the mRNA-LNPs upon storage, giving rise to a variety of degradation products including fused and aggregated LNPs and even lipoplexes as they protrude from the LNPs (Figure 1G and H). Consistent with the transfection capability of the mRNA-LNPs, the lively, worm- or snake- or sperm-like appearances of these degradation products suggests their likely involvement in the transmembrane passage of the mRNA. Generally, it can be hypothesized that disintegration of the mRNA-LNP multiplies the nanoparticle numbers and overall surface, thus enhancing their proinflammatory and mRNA transfection potential.
5. Preclinical Pharmacokinetic, Tissue Distribution and Toxicities
The Pk, biodistribution and toxicity of the mRNA-LNPs were studied well before their use for vaccination, mostly using luciferase mRNA and different ionizable and PEGylated lipids [60], yet this information may have not been heavily weighted in the context of human vaccinations. Following their i.m. injection in mice, LNP-carried luciferase was detected beyond the injection site up to 9 days, but it was also seen in liver up to 2 days, which disappeared in 48 hours.
Regarding preclinical toxicological studies of mRNA vaccines, it is the two novel lipids in Comirnaty that was critical for the clinical studies, as prior direct evaluations were not available with these lipids. ALC-0315 is an ionizable lipid which has positive charge at low pH. It is essential for the integration of the mRNA to the LNP, and it also plays a key role in the endo-lysosomal escape of mRNA after entry into the cell. ALC-0315 has numerous toxicities partly due to its immune stimulating, proinflammatory effects, and partly to its fusogenic effect [61,62,63]. The other new lipid used in the Comirnaty formulation is ALC-0159, a 2K-PEGylated lipid (not phospholipid) whose primary function is to form a protective hydrophilic layer that improves colloidal stability, enhances solubility, storage stability, and reduces nonspecific binding to proteins [64]. The short 14-carbon alkyl chains that anchor the molecule to the nanoparticle represent a relatively weak attachment, which is in keeping with the substantial divergence of the clearance of ALC-0315 and ALC-0159: the plasma t½ of ALC-0159 and ALC-0315 in rats were 73 h and 139 h, respectively [65]. Nevertheless, it remains unknown how they are transported in plasma, i.e., whether they bind to plasma proteins or lipoproteins.
As for the toxicity assessment of the novel lipids in Comirnaty, their preclinical evaluation claimed no adverse effects on the basis that the siRNA-containing LNP product patisiran (OnpattroTM), which contains similar (but not identical) lipid excipients, were approved in the US, Europe and Canada for the treatment of hereditary transthyretin-mediated amyloidosis (hATTR amyloidosis) by infusion every 3 weeks [65].
6. The Distinct Structural Characteristics of mRNA-LNPs That May Be Linked to Adverse Events and Complications
Table 1 compiles a list of structural features and chemical characteristics that can, in theory, be linked to the AEs reported for mRNA vaccines. They are inherent properties of Comirnaty and Spikevax, they may or may not apply to other mRNA-LNP constructs. The unique properties are listed in the 1st column of Table 1 and linked to experimentally and/or clinically demonstrated adverse immune processes in column 2. These abnormal immune reactions are then associated with different AEs in column 3 based on cell biology and immune pathology extrapolations.
To avoid misinterpretations of the information in Table 1, it needs to be strongly stressed that these data do not suggest that all included adverse processes occur in all vaccine recipients or even in adverse reactors. The table intends to be a compilation of theoretically possible reasons that may explains the variety of symptoms. It is certain that the Covid-like vaccine AEs rising in a few out of hundreds of vaccine recipients are multicausal; each can arise from one or more of the listed adversarial pathways, acting independently or simultaneously, in an additive or synergistic fashion. The rise and extent of AEs depend on many individually varying genetic and epigenetic factors, as well as external and internal conditions.
6.1. Ribosomal Synthesis of the Spike Protein Fundamentally Transforms Antigen Processing and Presentation
Among the features of mRNA vaccines that present a risk for AEs, it is the principle of using mRNA-LNPs for vaccination that needs to be considered first, since ribosomal synthesis of the SP fundamentally transforms the normal process of antigen processing and presentation. Immunization with currently used traditional vaccines involves the uptake of vaccine (nano)particles by antigen presenting cells (APCs; dendritic cells, macrophages, and B cells) at the site of the injection and the infiltrating lymph and lymph nodes via phagocytosis, processing the antigen via enzymatic degradations and other modifications during their passage via cell organelles, namely the phagosomes (endosomes), phagolysosomes, endoplasmic reticulum (ER), and Golgi apparatus. At the end, the small (12-26 amino acid [66] immunogenic peptides are presented to T helper (Th) and B lymphocytes in narrow recesses (grooves) on the surface of MHC (Major Histocompatibility Complex) Class-II molecules (shortly: Class-II).
The above textbook information is cited to help realizing, how differently the immunogenicity of mRNA-LNP-coded SP proceeds after immunization with mRNA vaccines. According to the mechanism flowchart shown in Figure 2, after the standard first step, i.e., phagocytosis followed by phagolysosome formation, the intact mRNA molecules escape from the phagolysosomes to the ribosomes in the cytoplasm to code for the synthesis of the SP (steps 1 and 2 in Figure 2.). This is a major deviation from the above-delineated, evolution-refined and highly controlled antigen degradation to antigen synthesis on ribosomes, which is influenced by numerous factors that have nothing to do with immunogenicity [68,69,70,71]. These include the levels of rRNA and ribosomal proteins at the time of vaccination, the functioning of open reading frames and chaperons that determine the quaternary structure of the antigen, and the intensity of mTOR (mechanistic Target of Rapamycin) pathway, which is a central regulator of ribosome biogenesis [72]. The mTOR is sensitive to various environmental signals, such as nutrient availability, energy status, hormonal and growth factors, cellular stress and up- or downregulates ribosomal protein synthesis [72]. Finally, post-translational modification of the SP may also occur, and all these controlling factors taken together give reason for doubts that the situ synthesis of an antigen on ribosomes can be controlled, most importantly by the vaccine dose.
Unlike structural or secretory proteins, which have distinct, well-defined functions, the immune response to protein antigens involves a complex chain of molecular and cellular effects, each influenced by individually varying control factors. In addition, immune responses may have different health effects in different people. Thus, uncontrollable input signal, i.e., antigen quality and amount, may entail unpredictable variations in ultimate health effects. In essence, deviation from natural processes of immunogenicity may increase the risk for abnormalities in immune response [73].
6.2. Multiple Chemical Modification of the mRNA Increases Its Stability and Translation Efficacy
Beyond the above modifications of antigen synthesis, processing and presentation, a profound deviation from natural immunogenicity is achieved by multiple chemical modification of the mRNAs. Their combined effect is a substantial increase in the biological stability and translation activity of the mRNA relative to native mRNAs. The specific modifications are discussed below.
6.2.1. Replacement of mRNA Uridine with Pseudouridine (ψ)
Extracellular unmodified mRNAs have strong immune stimulatory effect due to their binding by pattern recognition receptors, such as Toll-like receptor-3 (TLR3), TLR7 and TLR8, or the retinoic acid-inducible gene I (RIG-I) receptor [67]. This effect, along with the instability, poor translation efficacy and high cost of mRNA synthesis prevented the therapeutic use of mRNAs prior to 2010s. It was therefore a milestone in mRNA research that Kariko et al., replaced uridine with pseudouridin (ψ) in the mRNA nucleotide chain, to reduce its immune reactivity and increase its biological stability and translation efficacy [74]. In their experiment the transfection of mice with pseudo-urinated luciferase mRNA using lipofectin as mRNA carrier resulted a 78-fold increase in splenic luciferase translation 24 h after the injection, showing the feasibility of lasting mRNA transfection and translation using a complex of ψ-mRNA with a fusogenic lipid [75]. Subsequent studies [76,77] further improved the efficacy of gene translation, particularly with N(1)-methyl-pseudouridine (1m-ψ), which outperformed ψ-mRNA in protein expression [78].
6.2.2. Codon Optimization
Codon optimization implies changing bases in the polynucleotide sequence in the mRNA to match the human codon for amino acid tRNA, thus improve the stability and translation efficacy of the SP. This molecular method ensures high levels of SP expression in human cells for robust immune response, while maintaining the stability and functionality of the mRNA [79].
6.2.3. Methylation of the 5’ Cap
Methylation of the 5’ cap in mRNA vaccines refers to a chemical modification of the mRNA molecule at the 5' end. The addition of a methyl group improves mRNA stability by resisting exonucleases and enhances the recognition of the mRNA by the host cell's ribosomes, leading to efficient translation into the target protein, i.e., SP. The 5’ Cap methylation also minimizes unwanted immune responses against the synthetic mRNA, thus improving vaccine efficacy [80,81,82,83].
6.2.4. UTR Stabilization
UTR stabilization involves modifying the untranslated regions (UTRs) of mRNA to enhance its stability and translational efficiency in vaccines. Thus, Comirnaty utilizes a 5' UTR derived from the human alpha-globin gene and a 3' UTR combined from the amino-terminal enhancer of split (AES) and mitochondrial ribosomal RNA 1 (mtRNR1) genes. Spikevax adopted a 3′ UTR from globins. The stability and efficient translation of the mRNA enhance the production of the target antigen, resulting in a stronger immune response [33,73,84].
6.2.5. 3′ Poly(A) Tail Optimization
The 3' poly(A) tail optimization in mRNA vaccines refers to the engineering of the polyadenylated tail at the 3' end of the mRNA to enhance its stability, efficiency of translation, and overall efficacy in SP expression [85]. The poly(A) tail containing 100–150 adenine residues protect the mRNA from degradation by exonucleases in the cytoplasm, thereby extending its half-life. It can also interact with the 5' cap of the mRNA to form a closed-loop structure, facilitating efficient ribosome binding and protein synthesis. The tail also helps in nuclear export of mRNA and proper localization in the cytoplasm [33,73,84].
6.2.6. GC Enrichment
The enrichment with GC refers to increasing the guanine (G) and cytosine (C) content in the mRNA to enhance its overall functionality. GC-rich regions form stronger hydrogen bonds compared to AU (adenine-uracil) base pairs, increasing the thermodynamical stability of the mRNA, protecting it from degradation. Enrichment with GC also increases the mRNA’s compatibility with the host cell's translation machinery, leading to more efficient protein synthesis. Furthermore, GC-rich sequences can help avoiding secondary structures that interfere with ribosome access and the binding of translation initiation factors. GC enrichment, just as the uridine-ψ exchange, can also reduce the inflammatory response to the mRNA [86,87,88,89].
6.3. The Biological Consequences of Ribosomal Antigen Synthesis and mRNA Modification
After fulfilling their role in the ribosomal synthesis of proteins, mRNAs are usually cleaved by deadenylation, endonucleases, ribozymes and spontaneous hydrolysis of the sugar backbone on a time scale of hours to days [90]. This process is under close regulation that ensures protein homeostasis in the cells. The detouring the SP synthesis to the ribosomes, increasing the stability of the mRNA and boosting its translation to SP may lead to increased SP production which can lead to beneficial as well as adversarial immune effects, as detailed below.
6.3.1. Enhancement of Immunogenicity
The beneficial, highly renown impact of the above discussed two innovations in immunogenicity is a significant enhancement of antiviral antibody response. The SP neutralizing antibody titers in mRNA vaccine recipients often exceeded the titers seen in natural infections up to 4-fold, which established the success of this technology [91,92,93,94]. However, some less beneficial consequences also surfaced, that could contribute to the broad spectrum of AEs.
6.3.2. Uncontrollable Cytoplasmic Accumulation with Diversification of the Processing and Presentation of the SP
The immunologically uncontrolled SP synthesis may alter the cytoplasmic fate of SP, as its direct entry to the natural antigen processing pathways -as occurs after phago-lysosomal digestion of antigens-, is impossible. The full-length SP trimer is more than a hundred times larger (~1,273 amino acids) than the peptides typically presented on MHC molecules (7-13 amino acids), and if alternative clearance mechanism cannot cope with the intense SP production, cytoplasmic accumulation of the antigen may occur. The alternative options for antigen processing and presentation include at least 4 pathways, each of which has a risk for abnormal consequences. These are: (i) digestion in proteasomes that can result in cross presentation of digested peptides on MHC-Class-I molecules invoking CD8-cytotoxic T cell attack; (ii) expression on the plasma membrane leading to antibody-mediated cellular and complement-mediated cytotoxicity; (iii) secretion into the extracellular space, causing toxemia; and (iv) exosomal propagatation of the SP, its mRNA and micro-mRNAs, partially mimicking viral infection.
6.3.3. Digestion of the SP by the Proteasomes Resulting in Cross Presentation on MHC-Class-I Molecules with Autoimmune Damage
Proteasomes are cytoplasmic enzyme complexes hydrolyzing intracellular proteins to MHC-I presentable small peptides, thus they play a primary role in processing the SP for CD8+ cytotoxic T cell (Tc) recognition. At an early age, during T cell development in the thymus, Tc-s with high affinity for self-antigens presented on MHC Class-I molecules are deleted (clonal deletion), the mechanism of central tolerance. Outside of the thymus, the Tc-s that escaped deletion respond with anergy to self-antigens, except during infection, when the co-stimulatory signals override tolerance and activate these T cells for an effective cytotoxic response against the foreign antigen harboring cells, causing apoptosis. This is the mechanism via which the immune system detects and eliminates Class-I presenting cells afflicted by intracellular infection or cancer.
In the case of mRNA vaccines, as a result of booster vaccination, presentation of the SP peptides on MHC Class-I molecules, called cross-presentation [95] may lead to an autoimmune attack against the very APCs that initially educated the Tcs for recognizing and killing SP-peptide-presenting infected cells (Figure 3). Preconditions for co-stimulatory stimulation may also exist, due to the proinflammatory effect of the LNPs (discussed later). Also reviewed later, the mRNA LNPs can transfect essentially all body cells, leading to cross-presentation that can induce a variety of autoimmune phenomena, mentioned in Section 2 and Section 3.
6.3.4. Expression of the SP on the Plasma Membrane Leading to Antibody-Mediated Cellular and Complement-Mediated Humoral Cytotoxicities
As shown in Figure 2 (step 2), another possible fate of intracellularly accumulated SP is plasma membrane insertion, which may also cause autoimmune damage in the presence of SP-binding neutralizing antibodies. Therefore, this type of cytotoxicity can arise after the first or more boosters, or in people also infected with SARS-CoV-2. There are 2 independent pathways of antibody-dependent cytotoxicity, shown in Figure 4A and B. One called antibody-dependent cellular toxicity (ADCC), a capability of NK cells, macrophages and neutrophil leukocytes (Figure 4A) whose Fc receptors bind to the Fc portion of SP-bound antibodies and release apoptotic effector molecules. The other is complement activation (Figure 3B), due to the binding of C1Q to the Fc portion of anti-SP antibodies, initiating the autocatalytic cascade via the classical pathway.
6.3.5. The "Seneca Effect"
The above delineated Tc (Figure 3) and antibody-mediated ADCC (Figure 4) and complement-mediated cytotoxicity on APCs (Figure 4B) represent potentially deadly attacks against the very cells that “taught these T and B cells for specific recognition of the SP. As a stretched, but perhaps useful metaphor, the phenomenon resembles the Seneca-Nero legend, where Seneca, the teacher, was compelled by his student, Nero, to commit suicide. Therefore, the term "Seneca effect" could serve as a memorable descriptor for APC-targeted autoimmunity. It may contribute to the dysregulation of the T regulatory (Treg) and CD4+ and CD8+ effector T cells (Teff) homeostatic cell balance after repeated vaccinations, leading to elevated IgG4 with consequent immune suppression. The clinical consequence is a plethora of autoimmune diseases and promotion of aggressive cancer [73,96] or reduced anti-COVID-19 immunity after booster vaccinations [97].
6.3.6. Secretion of SP into the Extracellular Space for SP Reuptake and Systemic Dissemination
As a further consequence of cytoplasmic accumulation of the SP and subsequent diversification of antigen processing, the SP can undergo secretion to the extracellular space (Figure 2, step 2). This may have an essential role in the anti-SP immunogenicity of the vaccine, since a part of the secreted SP can undergo phagocytic reuptake and processing through the standard phagolysosome-ER-Golgi-Class-II pathway (step 5 in Figure 2). However, since the SP is a facultative toxin, its entry into the systemic circulation implies toxemia, the next addressed major inherent cause of vaccine-induced AEs.
6.3.7. The Autophagy Pathway of Antigen Processing and Presentation
The secretion and phagocytic uptake of the SP, which is the natural process of antigen processing (step 5 in Figure 2), may be one of two pathways through which Class-II proteins are fed with SP fragments. However, these processes, i.e., exocytosis and autocrine endocytosis, are under metabolic (ATP dependence) and other controls which lend individual variation to Class II-presentation. Considering the increased antibody response to the vaccine (5.3.1) it is a question whether such convoluted antigen processing would be the only mechanism of Class-II mediated immunogenicity. There is an alternative mechanism of SP processing, namely autophagy (not shown in the Figure 2) whereupon the SP released from the ribosomes is engulfed by cytoplasmic bilayer vesicles called autophagosomes, which turn into auto-phagolysosomes after fusing with lysosomes. Just like phagolysosomes, they feed the ER sacs and reach Class-II molecules, as an alternative, possibly more efficient pathways of T and B cell education for SP recognition.
6.3.8. Exosomal Dissemination of Vaccine and Viral Elements Propagating Inflammation and Transfection: The Dumbed-Down Virus/LNP Chimera Concept
Exosomes are small extracellular nanovesicles (30–150 nm) that play a role in intercellular communication and carry biomolecules such as lipids, proteins, and nucleic acids, including mRNA. They are secreted by cells and involved in intercellular communication. They can carry proteins, lipids, and nucleic acids from the parent cell throughout the organism. In the case of mRNA vaccine-exposed cells, they can carry the SP on their surface, as well as the whole mRNA and its immune stimulatory small fragments called miRNAs (microRNAs) [73,96,98]. Exosomes propagate from cells in a manner similar to how newly formed virus particles emerge from SARS-CoV-2-infected cells, the only difference is that the nucleocapsid is replaced with SP-crowned plasma membrane and the mRNA does not code all virus proteins [98]. Since the lipids from LNPs may incorporate into SP-expressing exosomes, these vesicles may actually represent dumbed-down virus/LNP chimeras. They can contribute to the immunogenicity of the SP by presenting antigens to APCs just as the vaccines do [98], providing rationale to use exosomes as an alternative to LNP-based vaccines [99,100]. The demonstrated efficacy of this approach for inducing immunogenicity attests to the exosomes’ capability for transfecting distant tissues and cells and thus contributing to the inflammatory and/or autoimmune complications [96,98].
6.3.9. Excessive Somatic Hypermutation in B Cells
Somatic hypermutation is a phenomenon whereby the immunoglobulin genes encoding the variable region of the B-cell receptors (BCRs) undergo especially high rate of mutation to produce slightly different versions of the antibodies after the B cells are activated though interaction with APCs and helper T cells (Figure 2, step 6). The B cells that produce high-affinity antibodies against the antigens survive and undergo affinity maturation, while those with lower-affinity antibodies die off. The process gives rise of plasma cells, which secrete antibodies that are more effective at neutralizing the target antigen, as well as production of long-lived memory B cells, which ensure quick production of high-affinity antibodies at a later encounter with the pathogen. Thus, somatic hypermutation enhances the effectiveness of the antibody response, helping the immune system better combat the pathogen targeted by a vaccine.
The mRNA vaccines are potent inducers of follicular helper and germinal center B cell responses in the spleen and lymph nodes, driving further acceleration of somatic hypermutation and greater level of affinity maturation [101]. Such germinal center activation persisted for at least 3-weeks in rats injected with mRNA-LNPs, which could also be associated with fever [65]. This process contributes to the superior antibody response to mRNA vaccines as discussed in section 6.1., however, it also carries a risk for diversification in the variable region of BCRs, leading to the production of antibodies with cross reactivity with host cell antigens. Thus, the autoimmune complications after mRNA vaccination may be explained with an overdrive of somatic hypermutation, at least in part [102,103,104,105,106,107].
6.3.10. Reverse Transcription of the mRNA with Insertion Mutagenesis May Prolong the Risk of Harm
The extended intracellular presence of stabilized mRNA increases the risk of its transit into the nucleus and undergo reverse transcription to insert into the DNA, i.e., cause insertion mutagenesis. Regarding the possible mechanism of this highly debated phenomenon, it is known that certain nucleic acid sequences in DNA, called transposons, have endonuclease activity, enabling changes in their position within the double helix, and make a variety of sequence changes, including the insertion of reverse transcribed mRNA fragments. The operation of this mechanism in the case of SARS-CoV-2 mRNA was shown by Zhang et al., in a study wherein “L endo-transposons” have inserted reversed transcribed SARS-CoV-2 mRNA into in cultured human cells which could be re-transcribed to SARS-COV-2 mRNA, explaining the positive PCR tests in some long Covid patients [108]. The authors also found evidence of viral-host chimeric transcripts. For mRNA vaccines, Alden et al., reported rapid insertion of reverse transcribed Comirnaty mRNA sequence into the DNA of cultured human liver tumor cells [109]. Regarding the mechanism of reverse transcription, the human enzyme DNA polymerase theta (Polθ, EC No 2.7.7.7) was proposed, which effectively performs RNA-dependent DNA repair [110].
Although there are many criticisms of the theory of reverse transcription of vaccine mRNA, for example by Merchant et al. [111], there is an indirect evidence supporting this possibility. Notably, in light of the fact that the longest circulating proteins in blood, IgG, has a half-life of about 21-28 days, it is difficult to explain the detection of the SP in blood up to 187 days [112], or up to 180 days in cardiac and skeletal muscle cells at sites of inflammation and fibrosis [113]. There are many more studies reporting more or less extended presence of SP in blood or other organs whose reason remain unclear despite the increased stability of the mRNA and/or the protein (27, 112,Castruita, 2023 #8357, 113-122). Obviously, conserving the code of the SP in the DNA over a lifetime or passing it to next generations is a huge concern that needs to be excluded via thorough investigations.
6.3.11. Frameshift Mutation upon mRNA Translation Causing SP Polymorphism
Another adverse consequence of the potential buildup of the SP in the cytoplasm of APCs and other transfected cells is frameshift mutation. It is due to incorrect identification of the base triplets by transfer RNAs in the mRNA, partly because of the presence of 1m-ψ. It may lead to SP polymorphism, with the formation of undefined peptide products and paraproteins with unknown antigenic and auto-immune potential.
The experimental evidence for frameshift mutation includes a study by Mulroney et al. who have shown the formation of 2 additional SP bands over the in-frame expectable product [123]. In another study, Boros et al. found that vaccination with 1mψ-mRNA can elicit cellular immunity to peptide antigens produced by +1 ribosomal frameshifting in major histocompatibility complex-diverse people {Boros, 2024 #8642}. The translation of 1-mψ-mRNA was shown by liquid chromatography tandem mass spectrometry to contain 6 in-frame and 9 chimeric SP peptides [124].
The complications of frameshift mutation include autoimmunity, neurotoxicity and weakening of humoral immunity [97,125,126]. The link between frameshift mutation and Tc-mediated autoimmunity was shown by the significantly increased IFNγ response to frameshifted antigens which was observed only in individuals vaccinated with Comirnaty [124]. Another adverse consequence is a highly significant increase in heart muscle 18-flourodeoxyglucose uptake, which was detected only in vaccinated patients up to half a year [124]. G-quadruplex formation, which makes the RNA secondary structures particularly stable and is implicated in post-vax neurotoxicity [127], is also thought to be a consequence of frameshift mutation.
6.4. The Spike Protein Can Be Toxic
6.4.1. The Structure and Cellular Secretion of the Spike Protein
The SP is a trimer glycoprotein, that forms the well-known spikes (peplomers) of the SARS-CoV-2 virus. It is essential for the binding of virions to ACE-2 and the later identified CD147 receptors, facilitating subsequent fusion and the release of viral mRNA into the host cell. The existence of vaccine-induced Covid-like AEs has led to a consensus that the common element in both the virus and vaccine-induced pathologies is the SP.
As to the question, how could intact SP insert into the bilayer membrane, and enter the bloodstream following immunization, Figure 2, which presents the fates of SP after synthesis on ribosomes, does not give a clue. It is basic knowledge in cell biology that insertion of proteins into bilayers, or their secretion into the external medium occurs via fusion of protein-loaded membrane vesicles with the cell membrane. However, the SP-loaded autophagosomes, the best candidates for such activity in case of ribosome-synthesized SP (Figure 2), destroy the SPs, rather than carry them to the cell surface. There must be another mechanism for SP secretion whereupon the integrity of the SP is maintained. The answer to this puzzle probably lies in the distinction of free and ER-bound ribosomes, the latter known to make the ER rough (rough ER). Despite their location differences, both free and bound ribosomes are structurally identical and function similarly in protein synthesis.
Freely floating cytoplasmic ribosomes primarily synthesize proteins that function within the cytoplasm, as presented in Figure 2. In contrast, ER-bound ribosomes synthesize proteins that are destined for insertion into different membranes or secretion from the cell of intact, functional proteins. Since the LNP-delivered mRNA, released into the cytoplasm of APCs (Step 1 in Figure 2) is unlikely to differentiate between free and bound ribosomes, processing of functional SP is most easily rationalized by synthesis on rough ER. Thus, the question of how intact SP can enter rapidly into the blood may be answered by an inherent property of all cells: synthesize proteins on rER-bound ribosomes.
6.4.2. Clinical Manifestations of SP Toxicity
The process of SP secretion by APC or other cells has already been discussed in relation to the origin and fate of free SPs. From a clinical point of view, it has been recognized from the beginning of the pandemics that the SP has intrinsic toxicities. Therefore, its secretion by APCs, or by any other transfected cells, implies toxicosis or, as proposed below, toxoidemia with sporadic toxicity. This phenomenon is puzzling since the whole SP molecule or its fragments were detected in the blood of AE-inflicted vaccinees months after the injection, which suggests binding to a blood component. The SP has many more unique features discussed below under separate subtitles.
Table 2 compiles a list of potential SP toxicities, underlying the pan-toxicity of the protein. Unlike most viral surface extensions, the SP appears to fundamentally impact essential cellular functions. This unique property is a subject of intense research, as evidenced by >2400 records on Medline (National Library of Medicine) when searching (by EndNote in December, 2024) for the combined “Any Field” search terms “spike protein”, “Covid-19” and “mRNA vaccine”.
6.4.3. The Superantigen-Like Activity of the Spike Protein
One of the hypotheses on the mechanism of SP toxicity is the “superantigen” hypothesis, based on common features of the SP and superantigen toxicities. Superantigens are bacterial toxins produced by, among others, streptococci and staphylococci [132]. They are extremely strong immune activators causing, among others, shock syndrome in streptococcus-infected children [133,134], just as SARS-CoV-2 infection does. Superantigens cause cytokine release and storm, as does the Covid-19 virus and the mRNA vaccines can occasionally do. According to Li et al, up to 20% of the body's T cells can be activated by superantigens, compared to the usual 0.0001-0.001% [135].
As further support for the superantigen hypothesis, a polypeptide domain in the superantigen enterotoxin B of Staphylococci, called "superantigen motif," has been shown to be present in the SP adjacent to its furin cleavage site and the positively charged multi-basic section (PRRAR), which is critical in the high affinity binding of the SARS-CoV-2 to cell membranes near their binding receptors[136,137,138] Valleriani, 2024 #8508;Li,2023 #8510}. This raises the possibility that that the SP can bind to cells with a mechanism that is shared with superantigens. As a further indirect evidence, beyond linking the APC and T cells together via their MHC Class-II and TCR receptors, superantigens also link other cells together, for example MHC and B cells, and indeed, the SP can also do the same, induce red cell aggregation (rouleau formation) [29,139,140] or bind erythrocytes and platelets [141,142,143,144,145]. Specific binding of the SP to non-ACE-2 receptors, e.g., CD147 on lymphocytes and other immune cells [30,146,147,148,149,150]. In lack of specific surface anchors, nonspecific electrostatic attraction between the positively charged region of the SP and negatively charged cell surfaces [141] could also contribute to the superantigenicity of the SP.
Based on all above information Figure 5 provides a schematic chart on the mechanism of the superantigen-like behavior of the SP.
6.4.4. The Spike Protein May Act Like a Toxoid with Sporadic Pathogenicity
Due to the unavoidable secretion of toxic SP after vaccination, the mRNA vaccines bear resemblance to attenuated live virus- or toxoid-based vaccines which rarely cause symptoms of the very disease against which they were developed. This analogy is based on the overlapping of COVID-19 and post-COVID symptoms with those of Covid-like vaccine AEs, and the fact that the vaccine-caused AEs are rare, just as the pathogenicity of attenuated live virus- or toxoid-based vaccines. A historical example for vaccine pathogenicity is paralytic poliomyelitis, where the weakened poliovirus in the oral formulation regains virulence and causes polio symptoms [151]. Likewise, the diphtheria and tetanus toxoids in the DTaP vaccines can occasionally revert to be toxic [152,153,154,155,156], casing the symptoms of infection. Thus, although the mechanisms are entirely different, the mRNA vaccines have a risk to act like SARS-Cov-2, i.e., become a sporadic pathogen. Support for this concept came recently, when the Phase I clinical trial with Moderna’s new, mRNA-LNP-based respiratory syncytial virus (RSV) vaccine was halted due to severe respiratory diseases that it caused.
6.5. The LNP Is Pharmacologically Active Immune Stimulant and mRNA Transfectant
Another essential innovation in using mRNA for vaccination is the use of LNPs as mRNA carriers. The introduction of LNP technology [157,158] solved many unresolved problems in the mRNA gene therapy, a critical step in the development of LNP-mRNA-LNP vaccines. In addition to protecting and carrying the mRNA, the LNPs are strong stimulators of the innate immune system and thus serve as adjuvants to the vaccine that enables its potent immunogenicity. This adjuvant effect seems to be particularly effective, since the LNPs cause many-fold lymphocyte and plasma cell proliferation in the germinal centers of lymph node follicles, where the antibodies are formed [101,159,160]. Although the exact mechanism of this remarkable immune stimulation is not clear, one likely contributor is that fact that the LNPs can simultaneously trigger activation of both the cellular and humoral the arms of the innate immune systems, specifically, leukocytes and the C system. The two activations can amplify each other in a positive feedback loop.
6.5.1. Activation of the Cellular Arm of Innate Immunity
The mRNA-LNPs are strong activators of innate cellular immunity. In Comirnaty, the primary contributing factor is the ionizable aminolipid ALC-0315, which accounts for 46% of the lipid content [53]). Experiments on mice and monkeys have shown that the LNP can bind to Toll-like receptors of immune cells, most importantly TLR2 and TLR4, as well as to other danger signal receptors. This leads to NF-κB pathway of Th1-dominant proinflammatory cytokine and chemokine secretion, including IL-1β, IL-2, IL-6, IL-18, IFN-γ, TNF-α and GM-CSF [65,161,162]. These LNPs also cause NLRP3 inflammosome activation that entails caspase-1-mediated apoptosis [162]. The stimulation of cellular immunity is further enhanced by the anaphylatoxins C3a and C5a [163,164], products of the concurrent stimulation of the C system by mRNA-LNPs.
6.5.2. Triggering of Humoral Immune Response: Complement Activation
The fact that liposomes and many other nanoparticulate drugs and agents can activate the C system have been known for decades [165,166,167,168], but for mRNA-LNPs it was shown only in a 2022 pig study [169]. This study investigated the mechanism of the allergic and anaphylactic reactions to Comirnaty that occurred shortly after the start of vaccinations in December 2020 [53,170]. The symptoms of these reactions were very similar to the infusion reactions caused by intravenous injection of liposomes, which were shown earlier to be related to C activation [171]. The study by Dezsi et al. provided evidence that Comirnaty can cause C activation-related pseudoallergy (CARPA) [169], suggesting that the human vaccine reactions could be attributed, at least in part, to CARPA [53]. This theory was supported later by Barth et al. [172] and Barta et. al. [173].
A further adverse consequence of C activation by the LNPs is accelerated disintegration in the lymph and blood due to the formation of the membrane attack complex (MAC, SC5b-9) resulting membrane damage with in premature release of payload [174,175,176,177]. Due to the above adverse impacts of anti-PEG antibodies on PEGylated drug products, the FDA recommends screening for both anti-protein and anti-PEG antibodies [178].
Regarding the mechanism of C activation by mRNA-LNPs, an in vitro study revealed that it proceeds via the alternative pathway [179]. It was also shown in human peripheral blood mononuclear cells (PBMCs) that C activation may play a causal role in the vaccine-induced secretion of IL-1β and TNF-α [179].
In addition to the alternative pathway activation, the LNPs exposure to high levels of anti-PEG antibodies in plasma leads to classical pathway activation. Further pig experiments showed that such activation can cause anaphylactic shock [173], explaining the increased incidence of anaphylaxis after mRNA vaccination [169]. However, unlike the constitutive C action by the LNP lipids, the substantial inter- and intraindividual variation of anti-PEG antibody levels in blood makes this type of antibody-dependent C activation sporadic. It should also be noted that the SP is also a C activator via the lectin pathway [12,16]. Thus, at different stages after vaccinations, different C activation pathways can get involved, putting the body under constant inflammatory pressure.
6.5.3. The LNP Is a Superadjuvant
Adjuvants are vaccine ingredients that enhance the antigen-specific immune responses. The mRNA vaccines do not contain any added adjuvant, yet they are very efficient inducers of anti-SP response, which needs to be understood. The above delineated simultaneous activation of the cellular and humoral innate immunity by the LNPs which may be additive and/or synergistic in immune stimulation may explain the “superadjuvancy” of the LNP.
Among the experimental evidences for such splendid function, the best known is the robust proliferation of follicular Th and germinal B cells, B memory cells and plasma cells in the in the lymph nodes of mice, as well as the release of proinflammatory cytokines and chemokines after immunization of mice with LNPs [101,180,181,182]. The explosion of B cell activity may have another contributor mechanism as well: the phenomenon of autoboosting. In this scenario the B cells, educated to recognize the SP on virus infected cells, also recognize the SP on the APC, an interaction that normally occurs when the already primed B cells encounter the antigen at a second time, after the booster injection. Thus, the distinction of memory B cells may become questionable, the naïve B cells may readily transform to plasma cells. Hence, in the context of mRNA-LNP vaccines, the LNP may be considered as a pharmacologically active ingredient [183], a "super adjuvant”.
6.5.4. The LNP Is a Fusogenic Transfecting Agent
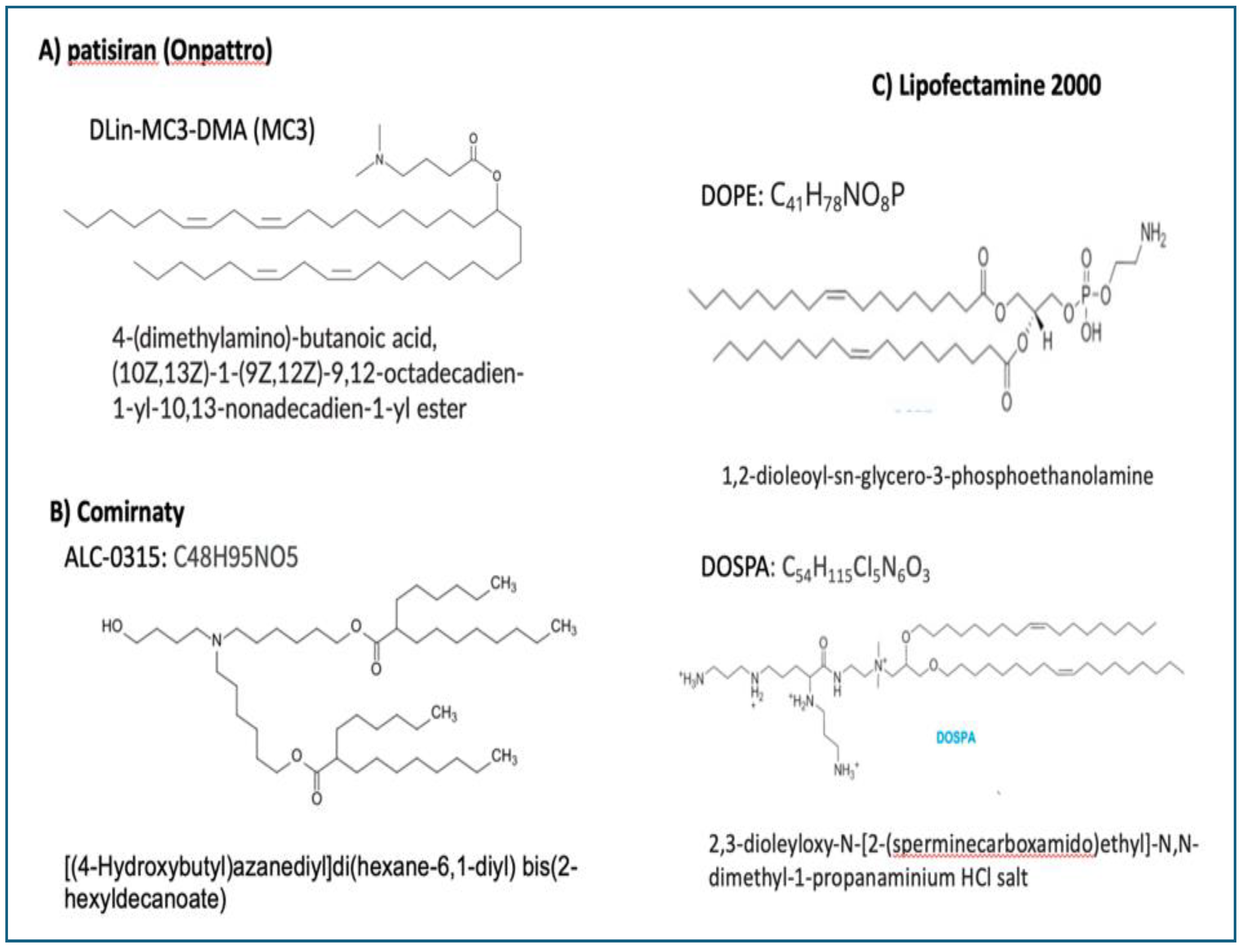
Another major adverse impacts of ionizable lipids is their fusogenic activity. Since the original mission of the type of LNP used for vaccinations was transfection of cells in gene therapy, the selection of ionizable lipid component was based, among others, on its fusogenic potential. Actually, the clinical success of liver-targeted patisiran (Onpattro), the first FDA-approved gene therapy against amyloidosis which used a fusogenic lipid in the LNP [184] was an important contributor to the selection of ALC 0315 for Comirnaty, although the latter was not intended to directly target the liver or other organs. As shown in Figure 3, the ionizable fusogenic lipids in Onpattro, Comirnaty and DOPE/DOSPA, components of the best-known transfection agent, lipofectamine [185], are very similar in having an ionizable amino group at one end, and multiple membrane-affine fatty acid chains, at the other. It is therefore not surprising that Cominaty transfects cells beyond the APCs in the lymph nodes draining the deltoid muscle.
The capability of LNPs to deliver mRNA in different body cells via fusion was shown, among others, by Pardi et al. in 2015 [60] that 24 min after deep muscle injection of luciferase mRNA-LNP in mice, intense fluorescence emerged in the liver. It was also observed that superficial muscle injection entailed less protein translation in the liver, suggesting that the injection site and depth are critical variables in LNP spreading [60]. More relevantly to the vaccine campaign, in a preclinical study by Pfizer/BioNTech, tritiated lipid marker was used to explore the biodistribution of Comirnaty-equivalent luciferase-mRNA-LNP in rats [65]. The study showed 2.8% of radioactivity in the plasma 15 min after the LNP injection, peaking between 1-4 h and distribution mainly into the liver, adrenal glands, spleen and ovaries in >2% over 48 h [65]. Hoverer <2% radioactivity was also seen in 12 other organs. Yet in further studies, the Ψ-mRNA was detected in the brain, heart, liver, spleen, ovaries, testes, and bone marrow and blood after vaccination of rats with mRNA-LNPs. Filling the gap in LNP tissue distribution studies in large animals, Ferraresso et al. [186] found that exogenous protein expression occurred in all major organs in swine when injected i.v. with a relatively low dose of mRNA encapsulated in a clinically relevant LNP formulation. Exogenous protein was detected in the liver, spleen, lung, heart, uterus, colon, stomach, kidney, small intestine, and brain, as well as in circulating white blood cells and platelets, and bone marrow megakaryocytes and hematopoietic stem cells. These results showed that nearly all major organs are “viable targets for mRNA therapies” [186], while the coins’ other side is that it may be difficult, if possible at all, to exclusively target certain cells with LNPs without off-target effects.
These and other data, reviewed by Pateev [187], attest to rapid multiorgan distribution and transfection of body cells with the spike protein-mRNA. The consequence is the same as described for APC above, proteasome processing of translated spike protein triggering Cytotoxic T cell damage with cell injury or death. This process explains the multiorgan autoimmune phenomena.
Figure 6.
Chemicals structures of ionizable lipids in A) Onpattro, B) Comirnaty and C) Lipofectamine 3000, containing DOSPA and DOPE at 3/1 mol ratio [185]. It should be noted that the pKa of DOSPA is much higher than that of the ionizable cationic lipids used in mRNA-LNPs.
Figure 6.
Chemicals structures of ionizable lipids in A) Onpattro, B) Comirnaty and C) Lipofectamine 3000, containing DOSPA and DOPE at 3/1 mol ratio [185]. It should be noted that the pKa of DOSPA is much higher than that of the ionizable cationic lipids used in mRNA-LNPs.
6.6. The PEG on the LNP Surface Is Immune Reactive and Immunogenic
While the intrinsic C activation by the vaccine lipids is likely to be a key contributor to the LNPs’ super adjuvant effect, the mRNA-LNPs can activate C via another way, too, depending on an extrinsic factor: access to anti-PEG antibodies. This activation proceeds via the classical pathway and may synergize with the constitutive activation by the lipids via the C3 amplification loop [188]. The main consequences of such occasional C activation are discussed below.
6.6.1. True and Pseudoallergic Reactogenicity
The first alarming AEs, observed shortly after the vaccination campaign began in 2020 December, was a cumulation of allergic reactions to the vaccine, some escalating to anaphylaxis [170]. Similar reactions and were reported in many other vaccination centers in different countries, leading to new guidelines for vaccine eligibility, excluding people with severe allergies [189]. Nevertheless, the incidence rate of mRNA-induced anaphylaxis is still more common than observed with other vaccines, e.g., 64-fold increase relative to flu vaccines. Since these reactions cannot be linked to specific allergy against any vaccine ingredient, the most likely mechanism is C activation-related pseudoallergy (CARPA) [167,168,169,173]. Solid evidence for a causal role of anti-PEG antibodies in C activation-related anaphylactic shock was recently obtained in pigs [173].
6.6.2. Anti-PEG Immunogenicity
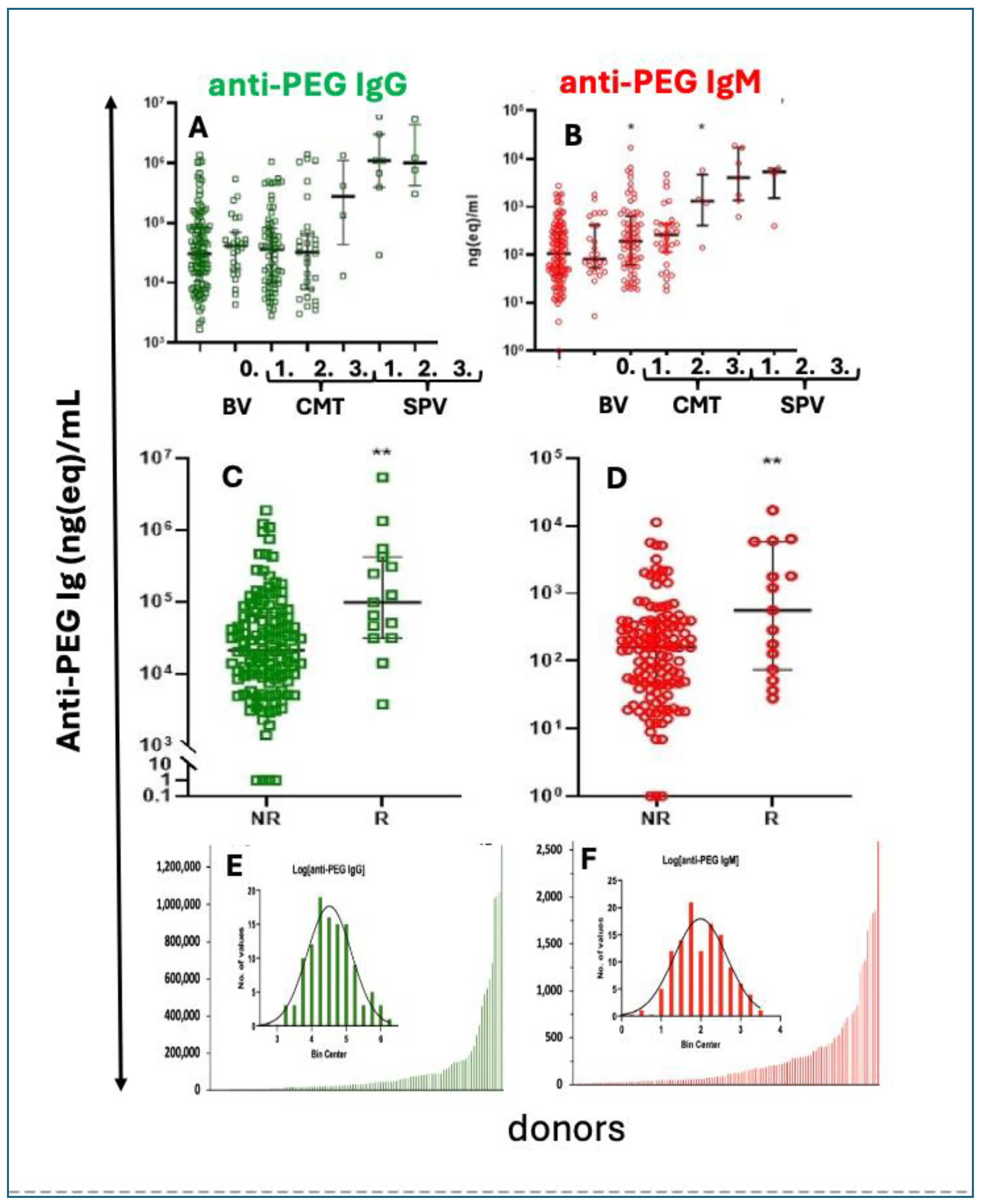
Considering the bridging role of C activation between innate and adaptive immunity [190,191], C activation by the LNPs is likely to play a causal or co-stimulatory role in the anti-PEG immunogenicity of mRNA vaccines. Such activity was recently shown by Kozma et al., in recipients of Comirnaty and Spikevax, after the second and third booster injection. As shown in Figure 4A and B, the levels of anti-PEG IgG and IgM rose over the pre-vaccination baseline with an order of magnitude in the case of Spikevax, while Comirnaty caused a smaller but significant rise of anti-PEG IgM after the second booster injection (Figure 4B). The connection between blood anti-PEG antibody levels and allergic reactions is demonstrated in the significantly higher anti-PEG IgG and IgM in AE reactors compared to non-reactors (Figure 4C, D). This study also revealed the presence and highly left-skewed distribution of preexisting anti-PEG IgG and IgM in 98-99 % of 116-118 healthy unvaccinated blood donors, among whom 3–4% were ‘anti-PEG Ab supercarriers” displaying extremely high antibody levels (Figure 4 E, F) [192]. The greater immunogenicity of Spikevax compared to Comirnaty [192] is consistent with the results of Carreno et al and Ju et al., [193,194] and the finding of Comirnaty’s immunogenicity (Figure 4B) was confirmed by Bavli et al. [195] by showing significant increases in mean anti-PEG IgG from 7.8 to 17.5 lg/mL three weeks after immunization with Comirnaty [195].
Figure 7.
Anti-PEG antibody levels and changes after vaccination with mRNA-LNP vaccines. A, B. by mRNA vaccines Pre-vaccination and postvaccine levels and correlation with HSRs/anaphylaxis. anti-PEG antibody concentrations in a mixed population of blood donors. Anti-PEG-IgG (A, green) and anti-PEG-IgM (B, red) values were sorted in growing order. The bottom bars (which look like lines) are absolute Ab concentrations. The inserted probability distribution histograms are made from the log of absolute Ab levels, grouped into bins on the abscissa.
Figure 7.
Anti-PEG antibody levels and changes after vaccination with mRNA-LNP vaccines. A, B. by mRNA vaccines Pre-vaccination and postvaccine levels and correlation with HSRs/anaphylaxis. anti-PEG antibody concentrations in a mixed population of blood donors. Anti-PEG-IgG (A, green) and anti-PEG-IgM (B, red) values were sorted in growing order. The bottom bars (which look like lines) are absolute Ab concentrations. The inserted probability distribution histograms are made from the log of absolute Ab levels, grouped into bins on the abscissa.
6.7. The LNP Is Unstable in Water
The nanoparticles in mRNA-LNP vaccines are not as stable as the traditional therapeutic liposomes. At the pH of human blood or tissue fluids, the ionic interaction between the ionizable lipid (BNT-0315) and the negatively charged mRNA might lose dominance, and the complex may be held together, at least in part, by hydrogen bonds [55]. Also, as revealed by electron microscopic images, the phospholipid bilayer coat of Comirnaty NPs lacks continuity, mono- and bilayers alter, implying less packaging and barrier functions compared to liposomes. A part of the LNPs are bicompartmental with mRNA-free blebs budding out, which was associated with increased the transfection potency [196].
Nano-mechanically, the Comirnaty NPs are soft, compliant structures, that were shown by multiple experimental approaches to be in a “liquid state” [59]. These unique features of mRNA-LNPs may contribute to the inherent instability of vaccine NPs whose shelf-life, after opening the thawed vials, is 24h [197]. An electron microscopic study showed that during long storage at 4oC the Comirnaty NPs are prone for disintegration, yielding snake-like winding nano-segments hypothetically identified as mRNA lipoplexes [55]. This is illustrated in Figure 6, which shows an electron microscopy image of Comirnaty LNPs stained with uranyl acetate after 7-day storage at 4°C. While other therapeutic liposomes show no morphological changes during such storage time, most of the nanoparticles in Comirnaty display major transformations, mainly fusion and disintegration, with the appearance of worm-, or snake-like structures presumably mRNA-lipoplexes.
6.8. The Spike Protein Is Enriched with Proline
Although the public understanding originally was that the recombinant SP would persist in the body only for a few days or weeks, in reality, clinical studies now report that modified SARS-CoV-2 mRNA routinely persist up to a month from injection and can be detected in cardiac and skeletal muscle at sites of inflammation and fibrosis, while the recombinant SP may persist a little over half a year in blood. This can be explained with double proline substitution (often called the "2P mutation") of two amino acids in the full-length SP encoded by Comirnaty's codon-optimized mRNA.
Proline is a unique amino acid often found in proteins where structural stability is required. It has a distinctive cyclic structure where the amino group is connected to the side chain, making it a "secondary" amine rather than the usual "primary" amine found in other amino acids. This rigidity can cause kinks or bends in protein structures, which influences protein folding, stability, and flexibility. Furthermore, proline and hydroxyproline emerge as prominent deuterium (heavy hydrogen) binding sites in structural proteins lending robust isotopic stability to the protein that resists not only enzymatic breakdown, but virtually all (non)-enzymatic cleavage mechanisms known in chemistry [198,199].
These effects combined stabilizes the SP in the pre-fusion peplomer conformation, which is preferred for immunogenicity [200]. This change also reduces the protein’s metabolism and prolongs its circulation time, potentially contributing to its slow-developing harmful effects and widespread distribution throughout the body, playing a key role in the chronic side effects of the vaccine.
6.9. The Injectable Vaccines May Contain Contaminations with Plasmid DNA and Inorganic Elements
Several studies reported different undeclared contaminants in the Pfizer mRNA vaccine [34,201,202], including segments of the plasmid DNA vector used as the template for in vitro transcription of SP DNA to mRNA. As discussed in the section on turbo cancers, integration of these DNA fragments, including the tumor promoter SV40, into the host DNA, can be shown in vitro, but the evidence for their contribution to “turbo” malignancy in vivo is mostly empirical, heavily argued in the cancer field.
DNA contamination in the vaccine as source of cancerogenic mutagenesis was demonstrated, among others [203] by Kammerer et al., who found large amounts of residual DNA in Comirnaty that included the monkey Simian virus’ cancerogenic promoter/enhancer sequence, known as SV40 [204]. As for other contaminants, Diblasi et al. [34] reported the presence of 55 undeclared chemical elements in 6 different brands of COVID-19 vaccines, including Comirnaty and Spikevax [34]. Except for noble gases, the undeclared elements were from all groups of the periodic table, some with luminescent and magnetic properties. Beyond the cytotoxicity of lanthanides, high concentrations of the heavy metals detected have been associated with toxic effects in humans. The range of detected elements showed substantial variation across different Covid-19 vaccine brands and sampling times, for example Comirnaty, 21-25 different elements were detected in different batches, samples at different times, with only a few declared in the vaccine’s specification. This inconsistency led the authors to the conclusion that the differences arise because of dynamic nature of the nanoscale self-assembling nanostructures over time. In fact, other studies report different exotic sub- or multimicron particles in the mRNA vaccines, such as ribbons, sheets, nanotubes, nano dots and nano scrolls [205]. These represent lipid self-assemblies, mRNA lipoplexes or different ion-lipid complexes and crystals, graphene oxide, calcium carbonate with graphene inclusions, iron oxide, PEG and many more multi-molecular complexes.
Nonetheless, neither the source of these contaminations nor their contribution to AEs were determined and remain to be explored in the future. The fact that they were present in all vaccines points to a systemic issue with the physical and chemical properties of the vaccines, rather than to a fault of any particular brand or process.
In the lack of experimental data or hypotheses regarding the causality between molecular self-assemblies and AEs, one theoretical mechanism is C activation. The causal role of this phenomenon in AEs was discussed in the context of adverse LNP effects (6.4), which also applies to alternative nanoarchitecture including vaccine contaminations. As is known, urate, hydroxyapatite and cholesterol crystals [206,207,208,209,210], organic and inorganic phosphates [211,212], needle-structures [213,214] or other artificial, non-biological surfaces can activate the C system involving both the classical and the alternative pathways. Complement activation, as mentioned earlier, represents a major proinflammatory stimulus.
7. The Issue of Turbo Cancer, Alias Conspiracy Cancer
The close temporal relationship of some haemato-lymphoproliferative and other malignant illnesses with mRNA vaccinations, mainly after booster injections, led to the concept of “turbo cancer” [51,215,216,217,218,219], a highly debated assertion on the rise of rapidly progressing new cancers following vaccinations with mRNA vaccines. It is considered as an anti-vaccination conspiracy theory, that is not supported by mainstream cancer research (Wikipedia). Nevertheless, it has several scientific explanations, one of which is insertion mutagenesis [203,204], i.e., the insertion of replication-competent plasmid DNA sequences into the host DNA. DNA contamination in the vaccine as source of cancerogenic mutagenesis was demonstrated, among others, by Kammerer et al., who found large amounts of residual DNA in Comirnaty that included the monkey Simian virus’ cancerogenic promoter/enhancer sequence, known as SV40 [204]. However, critics of the turbo cancer concept point to lack of credible evidence that the in vitro findings are relevant to humans in vivo, given the huge differences between the in vitro and in vivo conditions of insertional mutagenesis, frameshift mutation or somatic hypermutation, the other causes implicated in vaccine-induced tumorigenesis.
Notwithstanding the claim on lack of credible scientific evidence, there are experimental and clinical data that support the turbo cancer concept. Nacionales et al described ectopic lymphoid tissue proliferation in mice injected with mRNA [220], whose features are reproduced by the substantially increased rate of human malignant B and T cell lymphomas and lymphoid leukemias [219,221]. Thus, Gentilini, et al reviewed 28 reports on malignancies developed soon after mRNA-LNP Covid vaccination, and 26 of them were B- cell and T cell lymphoproliferative disorders [221]. The authors argued that the time correlation between vaccination and symptoms met the Hill's criteria for causation, a group of nine principles establishing epidemiologic evidence of a causal relationship between a presumed cause and an observed effect [222].
Further explanations for turbo cancer, alias “conspiracy cancer” include postvaccination decline of adaptive and innate immune responses [215]; lymphopenia with suppression of type I interferon response [73,223]; inhibition of p53 SP tumor suppressor protein [224], tumorigenic effect of methylated pseudouridine [225,226,227]; the blocking of innate and specific IgG1 antitumor responses by nonspecific IgG4 via inhibition of Fc receptor (called fragment crystallizable gamma receptor IIb, FcγRIIB) mediated activation of B cells, dendritic cells and macrophages [228]; downregulation of tumor suppressor genes among others, by exosome-spread non-coding micro-RNAs (miRNAs, ~22 nucleotides) and G-quadruplexes (four-stranded regions in guanine-enriched mRNAs) [127,216,229,230,231]. Thus, there is a multitude of possible vaccine effects considered to cause turbo cancer, implying “turbo-vaccine-induced cancer”.
8. Outlook
The mRNA-LNP-based vaccines have played a pivotal role in combating the COVID-19 pandemic and remain the leading choice for stimulating immunity against new SARS-CoV-2 variants. This technology represents a revolutionary advancement in vaccine science, offering significant advantages such as simplified, accelerated, and cost-effective production. Its flexibility allows for rapid adaptation to viral mutations and the potential to deliver multiple antigens simultaneously, enabling the development of combined vaccines against multiple viral strains. The assertions by the manufacturers, regulatory agencies (WHO, CDC, FDA), major healthcare institutions and other authorities that these vaccines are effective and safe enabled continued promotion and unforeseen investment into this technology. Accordingly, the US FDA recently approved Moderna's second mRNA vaccine for RSV (mRESVIA, mRNA-1345) [232,233] and in August 2024, it endorsed the use of updated versions of mRNA vaccines (targeting the Omicron variant KP.2 strain) for both adults and children, granting "emergency use" authorization for babies aged 6 months or older [234]. Beyond the fully authorized COVID-19 and RSV vaccines, >50 National Clinical Trial (NCT)-identified Phase I-III clinical trials are in progress against Covid-19, infectious diseases and cancer [33,235,236,237] and over 300 mRNA-LNP-based drugs are in preclinical development by scores of companies. The mRNA-LNP-based antiviral vaccine trials are against influenza, cytomegalovirus, Epstein-Barr virus, varicella-zoster virus, herpes simplex virus, Zika virus, norovirus (stomach flu) and HIV [33,235,236,237]. The global market for mRNA vaccines and therapeutics is already in the multibillion-dollar range and is projected to grow several-fold within a few years [238,239,240]. Clearly, the mRNA-LNPs technology is thought to have remarkable promise for personalized, more effective treatments for a range of unmet medical needs.
However, the picture may not be so crystal clear, as the mRNA vaccines may combine the highly esteemed capability for anti-COVID protection with inflammatory, auto immunogen and -arguably- cancerogenic potencies. Concerns about this unpleasant side of a Janus face stems in more than 2,000 scientific studies, moratorium calls by physician coalitions, legal depositions and compensation lawsuits, intense debates even in top political forums (British Parliament and the U.S. Congress), and the abundance of spoken and written information through public hearings and various media outlets featuring experts, vaccine injured people and concerned public leaders [241,242,243,244,245,246,247].
In light of negative efficacy, it has been questioned whether the booster injections provide any defense against COVID-19 [97], and whether the vaccine’s overall benefits outweigh the potential harms, particularly these days, when the pandemic is over, and the pathogenicity of new strains is not as expressed as that of the early variants [241,242,243,244,245,246,247,248].
However, as mentioned, it is not the breadth of AEs but their low frequency that has been stressed as basis of the “safe” declaration. In fact, the 0.03-0.5 % reported incidence rate represents low risk indicators by pharmacotherapy standards, however, vaccines must meet higher standards since they are given to large populations of healthy people. Quantifying the AE incidence as being “rare” may be subjective without the context and reference point used for the assessment.
Based on these and many other reasons, the problem is spurring calls for temporary moratorium on the currently used mRNA vaccine formulations [241]. An important development along this line is the hold on Moderna’s Phase I vaccine trial for RSV vaccine in children due to severe lower respiratory complications [249]. The fact that mRNA vaccines cause the same disease as the virus against which they were developed supports the contention that these prophylactic immune-gene therapies can act like attenuated virus vaccines.
In summary, the opposing views on the safety of mRNA-LNP vaccines, leading to an unparalleled division of society, underscores the importance of better understanding the AEs of mRNA-LNP-based COVID-19 vaccines and plan the future of this technology platform accordingly [250]. Beyond identifying the biological barriers inherent in the technology and find solutions for their overcoming, a major goal of future research in this field is the identification of people at risk for AEs and complications. The present comprehensive review of facts and hypotheses may contribute to achieving these goals, as well as to avoid the “Semmelweis reflex”, a metaphorical term referring to the human tendency to disregard or downplay new evidence or ideas that contradict established paradigms.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Acknowledgments
The financial support for the author’s research by the European Union Horizon 2020 projects 825828 (Expert), the National Research, Development, and Innovation Office (NKFIH) of Hungary (2020-1.1.6-JÖVŐ-2021- 00013) and the 2022-1.2.5-TÉT-IPARI-KR-2022-00009 are acknowledged.
Conflicts of Interest
The author is affiliated, among others, with an immune toxicology service provider SME, SeroScience LLC.
References
- Krumholz, H.M.; Wu, Y.; Sawano, M.; Shah, R.; Zhou, T.; Arun, A.S.; et al. Post-Vaccination Syndrome: A Descriptive Analysis of Reported Symptoms and Patient Experiences After Covid-19 Immunization. MedRxiv posted November 10, 2023. [CrossRef]
- Palmer, M.; Bhakdi, S.; Hooker, B.; Holland, M.; DesBois, M.; Rasnick, D.; et al. mRNA Vaccine Toxicity. https://d4ce.org/mRNA-vaccine-toxicity/2023.
- Costa, C.; Moniati, F. The Epidemiology of COVID-19 Vaccine-Induced Myocarditis. Adv Med. 2024, 2024, 4470326. [Google Scholar] [CrossRef] [PubMed]
- Novak, N.; Tordesillas, L.; Cabanillas, B. Adverse rare events to vaccines for COVID-19: From hypersensitivity reactions to thrombosis and thrombocytopenia. Int Rev Immunol. 2022, 41, 438–447. [Google Scholar] [CrossRef] [PubMed]
- Oueijan, R.I.; Hill, O.R.; Ahiawodzi, P.D.; Fasinu, P.S.; Thompson, D.K. . Rare Heterogeneous Adverse Events Associated with mRNA-Based COVID-19 Vaccines: A Systematic Review. Medicines (Basel). 2022, 9. [Google Scholar] [CrossRef]
- Padilla-Flores, T.; Sampieri, A.; Vaca, L. Incidence and management of the main serious adverse events reported after COVID-19 vaccination. Pharmacol Res Perspect. 2024, 12, e1224. [Google Scholar] [CrossRef] [PubMed]
- Law, B. Priority List of COVID-19 Adverse events of special interest: Quarterly update December 2020. Safety Platform for Emergency Vaccines (SPEAC). 2021;https://brightoncollaboration.org/wp-content/uploads/2023/08/SO2_D2.1.2_V1.2_COVID-19_AESI-update_V1.3-1.pdf.
- Fraiman, J.; Erviti, J.; Jones, M.; Greenland, S.; Whelan, P.; Kaplan, R.M.; et al. Serious adverse events of special interest following mRNA COVID-19 vaccination in randomized trials in adults. Vaccine. 2022, 40, 5798–5805. [Google Scholar] [CrossRef]
- Faksova, K.; Walsh, D.; Jiang, Y.; Griffin, J.; Phillips, A.; Gentile, A.; et al. COVID-19 vaccines and adverse events of special interest: A multinational Global Vaccine Data Network (GVDN) cohort study of 99 million vaccinated individuals. Vaccine. 2024, 42, 2200–2211. [Google Scholar] [CrossRef]
- Slater, K.M.; Buttery, J.P.; Crawford, N.W.; Cheng, D.R. Letter to the Editor: A comparison of post-COVID vaccine myocarditis classification using the Brighton Collaboration criteria versus (United States) Centers for Disease Control criteria: an update. Commun Dis Intell (2018) 2024, 48. [Google Scholar] [CrossRef]
- Levitan, B.; Hadler, S.C.; Hurst, W.; Izurieta, H.S.; Smith, E.R.; Baker, N.L.; et al. The Brighton collaboration standardized module for vaccine benefit-risk assessment. Vaccine. 2024, 42, 972–986. [Google Scholar] [CrossRef]
- Ali, Y.M.; Ferrari, M.; Lynch, N.J.; Yaseen, S.; Dudler, T.; Gragerov, S.; et al. Lectin Pathway Mediates Complement Activation by SARS-CoV-2 Proteins. Front Immunol. 2021, 12, 714511. [Google Scholar] [CrossRef]
- DeOre, B.J.; Tran, K.A.; Andrews, A.M.; Ramirez, S.H.; Galie, P.A. SARS-CoV-2 Spike Protein Disrupts Blood-Brain Barrier Integrity via RhoA Activation. J Neuroimmune Pharmacol. 2021, 16, 722–728. [Google Scholar] [CrossRef]
- Indes, J.E.; Koleilat, I.; Hatch, A.N.; Choinski, K.; Jones, D.B.; Aldailami, H.; et al. Early experience with arterial thromboembolic complications in patients with COVID-19. J Vasc Surg. 2021, 73, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Robles, J.P.; Zamora, M.; Adan-Castro, E.; Siqueiros-Marquez, L.; Martinez de la Escalera, G.; Clapp, C. The spike protein of SARS-CoV-2 induces endothelial inflammation through integrin alpha5beta1 and NF-kappaB signaling. J Biol Chem. 2022, 298, 101695. [Google Scholar] [CrossRef]
- Perico, L.; Morigi, M.; Galbusera, M.; Pezzotta, A.; Gastoldi, S.; Imberti, B.; et al. SARS-CoV-2 Spike Protein 1 Activates Microvascular Endothelial Cells and Complement System Leading to Platelet Aggregation. Front Immunol. 2022, 13, 827146. [Google Scholar] [CrossRef]
- Forsyth, C.B.; Zhang, L.; Bhushan, A.; Swanson, B.; Zhang, L.; Mamede, J.I.; et al. The SARS-CoV-2 S1 Spike Protein Promotes MAPK and NF-kB Activation in Human Lung Cells and Inflammatory Cytokine Production in Human Lung and Intestinal Epithelial Cells. Microorganisms. 2022, 10. [Google Scholar] [CrossRef]
- Thachil, J.; Favaloro, E.J.; Lippi, G. D-dimers-"Normal" Levels versus Elevated Levels Due to a Range of Conditions, Including "D-dimeritis," Inflammation, Thromboembolism, Disseminated Intravascular Coagulation, and COVID-19. Semin Thromb Hemost. 2022, 48, 672–679. [Google Scholar] [CrossRef]
- Rao, E.; Grover, P.; Zhang, H. Thrombosis after SARS-CoV2 infection or COVID-19 vaccination: will a nonpathologic anti-PF4 antibody be a solution?-A narrative review. J BioX Res. 2022, 5, 97–103. [Google Scholar] [CrossRef]
- Fujimura, Y.; Holland, L.Z. COVID-19 microthrombosis: unusually large VWF multimers are a platform for activation of the alternative complement pathway under cytokine storm. Int J Hematol. 2022, 115, 457–469. [Google Scholar] [CrossRef]
- Huynh, T.V.; Rethi, L.; Lee, T.W.; Higa, S.; Kao, Y.H.; Chen, Y.J. Spike Protein Impairs Mitochondrial Function in Human Cardiomyocytes: Mechanisms Underlying Cardiac Injury in COVID-19. Cells 2023, 12. [Google Scholar] [CrossRef]
- Schwartz, L.; Aparicio-Alonso, M.; Henry, M.; Radman, M.; Attal, R.; Bakkar, A. Toxicity of the spike protein of COVID-19 is a redox shift phenomenon: A novel therapeutic approach. Free Radic Biol Med. 2023, 206, 106–110. [Google Scholar] [CrossRef]
- Vettori, M.; Dima, F.; Henry, B.M.; Carpene, G.; Gelati, M.; Celegon, G.; et al. Effects of Different Types of Recombinant SARS-CoV-2 Spike Protein on Circulating Monocytes' Structure. Int J Mol Sci. 2023, 24. [Google Scholar] [CrossRef]
- Vettori, M.; Carpene, G.; Salvagno, G.L.; Gelati, M.; Dima, F.; Celegon, G.; et al. Effects of Recombinant SARS-CoV-2 Spike Protein Variants on Platelet Morphology and Activation. Semin Thromb Hemost. 2023. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Nishikawa, M.; Kanno, H.; Yang, R.; Ibayashi, Y.; Xiao, T.H.; et al. Long-term effects of Pfizer-BioNTech COVID-19 vaccinations on platelets. Cytometry A. 2023, 103, 162–167. [Google Scholar] [CrossRef]
- Acevedo-Whitehouse, K.; Bruno, R. Potential health risks of mRNA-based vaccine therapy: A hypothesis. Med Hypotheses. 2023, 171, 111015. [Google Scholar] [CrossRef] [PubMed]
- Parry, P.I.; Lefringhausen, A.; Turni, C.; Neil, C.J.; Cosford, R.; Hudson, N.J.; et al. 'Spikeopathy': COVID-19 Spike Protein Is Pathogenic, from Both Virus and Vaccine mRNA. Biomedicines. 2023, 11. [Google Scholar] [CrossRef]
- Niu, C.; Liang, T.; Chen, Y.; Zhu, S.; Zhou, L.; Chen, N.; et al. SARS-CoV-2 spike protein induces the cytokine release syndrome by stimulating T cells to produce more IL-2. Front Immunol. 2024, 15, 1444643. [Google Scholar] [CrossRef] [PubMed]
- Dima, F.; Salvagno, G.L.; Lippi, G. Effects of recombinant SARS-CoV-2 spike protein variants on red blood cells parameters and red blood cell distribution width. Biomed J. 2024, 2024, 100787. [Google Scholar] [CrossRef]
- Shouman, S.; El-Kholy, N.; Hussien, A.E.; El-Derby, A.M.; Magdy, S.; Abou-Shanab, A.M.; et al. SARS-CoV-2-associated lymphopenia: possible mechanisms and the role of CD147. Cell Commun Signal. 2024, 22, 349. [Google Scholar] [CrossRef]
- Lee, Y.; Jeong, M.; Park, J.; Jung, H.; Lee, H. Immunogenicity of lipid nanoparticles and its impact on the efficacy of mRNA vaccines and therapeutics. Exp Mol Med. 2023, 55, 2085–2096. [Google Scholar] [CrossRef]
- Xu, J.; Wu, Z.; Wass, L.; Larson, H.J.; Lin, L. Mapping global public perspectives on mRNA vaccines and therapeutics. NPJ Vaccines. 2024, 9, 218. [Google Scholar] [CrossRef]
- Wang, Y.S.; Kumari, M.; Chen, G.H.; Hong, M.H.; Yuan, J.P.; Tsai, J.L.; et al. mRNA-based vaccines and therapeutics: an in-depth survey of current and upcoming clinical applications. J Biomed Sci. 2023, 30, 84. [Google Scholar] [CrossRef]
- Diblasi, L.; Monteverde, M.; Nonis, D.; Sangorrín, M. At Least 55 Undeclared Chemical Elements Found in COVID-19 Vaccines from AstraZeneca, CanSino, Moderna, Pfizer, Sinopharm and Sputnik, V.; with Precise ICP-MS. International Journal of Vaccine Theory, Practice, and Research 2024, 3, 1367–1393. [Google Scholar] [CrossRef]
- Szebeni, J. Expanded spectrum and incidence of adverse events linked to COVID-19 genetic vaccines: New concepts on prophylactic immuno-gene therapy and iatrogenic orphan disease Preprints Org. 2024;Submittedhttps://www.preprints.org/manuscript/202411.1837/v1.
- Song, J.; Zhang, Y.; Zhou, C.; Zhan, J.; Cheng, X.; Huang, H.; et al. The dawn of a new Era: mRNA vaccines in colorectal cancer immunotherapy. Int Immunopharmacol. 2024, 132, 112037. [Google Scholar] [CrossRef] [PubMed]
- Baldo, B.A. Immune- and Non-Immune-Mediated Adverse Effects of Monoclonal Antibody Therapy: A Survey of 110 Approved Antibodies. Antibodies (Basel). 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Baumeister, S.H.C.; Mohan, G.S.; Elhaddad, A.; Lehmann, L. Cytokine Release Syndrome and Associated Acute Toxicities in Pediatric Patients Undergoing Immune Effector Cell Therapy or Hematopoietic Cell Transplantation. Front Oncol. 2022, 12, 841117. [Google Scholar] [CrossRef] [PubMed]
- Peaytt, R.; Parsons, L.B.; Siler, D.; Matthews, R.; Li, B.; Bell, D.; et al. The impact of early versus late tocilizumab administration in patients with cytokine release syndrome secondary to immune effector cell therapy. J Oncol Pharm Pract. 2023, 29, 45–51. [Google Scholar] [CrossRef]
- OpenVAERS. VAERS COVID Vaccine Adverse Event Reports. https://openvaerscom/covid-data. 2024;https://openvaers.com/covid-data.
- word, CO. Our World in Data. https://ourworldindataorg/grapher/covid-vaccine-doses-by-manufacturer?country=European+Union~HUN~USA. 2024.
- Mathieu, E.; Ritchie, H.; Ortiz-Ospina, E.; Roser, M.; Hasell, J.; Appel, C.; et al. A global database of COVID-19 vaccinations. Nat Hum Behav. 2021, 5, 947–953. [Google Scholar] [CrossRef]
- Our World in Data. covid-vaccine-doses-by-manufacturer. https://ourworldindataorg/grapher/covid-vaccine-doses-by-manufacturer?time=2021-01-12latest&country=European+Union~USA. https://ourworldindataorg/covid-vaccinations. 2024.
- Rhodes, P.; Parry, P.I. Pharmaceutical product recall and educated hesitancy towards new drugs and novel vaccines. International Journal of Risk & Safety in Medicine. 2024, 35, 317–333. [Google Scholar]
- VAERS. VAERS IDs. https://vaershhsgov/data/datasetshtml. 2023.
- Institute, P.E. Safety report. https://wwwpeide/SharedDocs/Downloads/EN/newsroom-en/dossiers/safety-reports/safety-report-27-december-2020-31-march-2022pdf?__blob=publicationFile&v=8. 2022.
- Zhou, F.; Hu, T.J.; Zhang, X.Y.; Lai, K.; Chen, J.H.; Zhou, X.H. The association of intensity and duration of non-pharmacological interventions and implementation of vaccination with COVID-19 infection, death, and excess mortality: Natural experiment in 22 European countries. J Infect Public Health. 2022, 15, 499–507. [Google Scholar] [CrossRef]
- Mostert, S.; Hoogland, M.; Huibers, M.; Kaspers, G. Excess mortality across countries in the Western World since the COVID-19 pandemic: ‘Our World in Data’ estimates of January 2020 to December 2022. BMJ Public Health. 2024, 2, e000282. [Google Scholar] [CrossRef]
- Kayano, T.; Sasanami, M.; Nishiura, H. Science-based exit from stringent countermeasures against COVID-19: Mortality prediction using immune landscape between 2021 and 2022 in Japan. Vaccine X. 2024, 20, 100547. [Google Scholar] [CrossRef]
- Woolf, S.H.; Lee, J.H.; Chapman, D.A.; Sabo, R.T.; Zimmerman, E. Excess Death Rates by State During the COVID-19 Pandemic: United States, 2020‒2023. Am J Public Health. 2024, 114, 882–891. [Google Scholar] [CrossRef] [PubMed]
- Gibo, M.; Kojima, S.; Fujisawa, A.; Kikuchi, T.; Fukushima, M. Increased Age-Adjusted Cancer Mortality After the Third mRNA-Lipid Nanoparticle Vaccine Dose During the COVID-19 Pandemic in Japan. Cureus. 2024, 16, e57860. [Google Scholar]
- Rhodes, P.; Parry, P.I. Pharmaceutical product recall and educated hesitancy towards new drugs and novel vaccines. International Journal of Risk & Safety in Medicine. 2024, 0, 1–17. [Google Scholar]
- Szebeni, J.; Storm, G.; Ljubimova, J.Y.; Castells, M.; Phillips, E.J.; Turjeman, K.; et al. Applying lessons learned from nanomedicines to understand rare hypersensitivity reactions to mRNA-based SARS-CoV-2 vaccines. Nat Nanotechnol. 2022, 17, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Schoenmaker, L.; Witzigmann, D.; Kulkarni, J.A.; Verbeke, R.; Kersten, G.; Jiskoot, W.; et al. mRNA-lipid nanoparticle COVID-19 vaccines: Structure and stability. Int J Pharm. 2021, 601, 120586. [Google Scholar] [CrossRef] [PubMed]
- Szebeni, J.; Kiss, B.; Bozo, T.; Turjeman, K.; Levi-Kalisman, Y.; Barenholz, Y.; et al. Insights into the Structure of Comirnaty Covid-19 Vaccine: A Theory on Soft, Partially Bilayer-Covered Nanoparticles with Hydrogen Bond-Stabilized mRNA-Lipid Complexes. ACS Nano. 2023, 17, 13147–13157. [Google Scholar] [CrossRef] [PubMed]
- Brader, M.L.; Williams, S.J.; Banks, J.M.; Hui, W.H.; Zhou, Z.H.; Jin, L. Encapsulation state of messenger RNA inside lipid nanoparticles. Biophys J. 2021, 120, 2766–2770. [Google Scholar] [CrossRef]
- Wang, M.M.; Wappelhorst, C.N.; Jensen, E.L.; Chi, Y.T.; Rouse, J.C.; Zou, Q. Elucidation of lipid nanoparticle surface structure in mRNA vaccines. Sci Rep. 2023, 13, 16744. [Google Scholar] [CrossRef]
- Leung, A.K.; Hafez, I.M.; Baoukina, S.; Belliveau, N.M.; Zhigaltsev, I.V.; Afshinmanesh, E.; et al. Lipid Nanoparticles Containing siRNA Synthesized by Microfluidic Mixing Exhibit an Electron-Dense Nanostructured Core. J Phys Chem C Nanomater Interfaces. 2012, 116, 18440–18450. [Google Scholar] [CrossRef]
- Unruh, T.; Gotz, K.; Vogel, C.; Frohlich, E.; Scheurer, A.; Porcar, L.; et al. Mesoscopic Structure of Lipid Nanoparticle Formulations for mRNA Drug Delivery: Comirnaty and Drug-Free Dispersions. ACS Nano. 2024, 18, 9746–9764. [Google Scholar] [CrossRef]
- Pardi, N.; Tuyishime, S.; Muramatsu, H.; Kariko, K.; Mui, B.L.; Tam, Y.K.; et al. Expression kinetics of nucleoside-modified mRNA delivered in lipid nanoparticles to mice by various routes. J Control Release. 2015, 217, 345–351. [Google Scholar] [CrossRef]
- Segalla, G. Apparent Cytotoxicity and Intrinsic Cytotoxicity of Lipid Nanomaterials Contained in a COVID-19 mRNA Vaccine. International Journal of Vaccine Theory Practice and Research. 2023, 3, 957–972. [Google Scholar] [CrossRef]
- Xu, Y.; Golubovic, A.; Xu, S.; Pan, A.; Li, B. Rational design and combinatorial chemistry of ionizable lipids for RNA delivery. J Mater Chem B. 2023, 11, 6527–6539. [Google Scholar] [CrossRef] [PubMed]
- Domingo, J.L. A review of the scientific literature on experimental toxicity studies of COVID-19 vaccines, with special attention to publications in toxicology journals. Arch Toxicol. 2024, 98, 3603–3617. [Google Scholar] [CrossRef]
- Kamanzi, A.; Zhang, Y.; Gu, Y.; Liu, F.; Berti, R.; Wang, B.; et al. Quantitative Visualization of Lipid Nanoparticle Fusion as a Function of Formulation and Process Parameters. ACS Nano. 2024, 18, 18191–18201. [Google Scholar] [CrossRef]
- Ltd PAP. Nonclinical Evaluation Report: BNT162b2 [mRNA] COVID-19 vaccine (COMIRNATYTM). https://wwwtgagovau/sites/default/files/foi-2389-06pdf. 2021;https://t.co/Zrhakh7Xgv.
- Sarri, C.A.; Papadopoulos, G.E.; Papa, A.; Tsakris, A.; Pervanidou, D.; Baka, A.; et al. Amino acid signatures in the HLA class II peptide-binding region associated with protection/susceptibility to the severe West Nile Virus disease. PLoS One. 2018, 13, e0205557. [Google Scholar] [CrossRef]
- Chaudhary, N.; Weissman, D.; Whitehead, K.A. mRNA vaccines for infectious diseases: principles, delivery and clinical translation. Nat Rev Drug Discov. 2021, 20, 817–838. [Google Scholar] [CrossRef]
- Granneman, S.; Tollervey, D. Building ribosomes: even more expensive than expected? Curr Biol. 2007, 17, R415–R417. [Google Scholar] [CrossRef]
- Sonenberg, N.; Hinnebusch, A.G. Regulation of translation initiation in eukaryotes: mechanisms and biological targets. Cell. 2009, 136, 731–745. [Google Scholar] [CrossRef]
- Henras, A.K.; Plisson-Chastang, C.; O'Donohue, M.F.; Chakraborty, A.; Gleizes, P.E. An overview of pre-ribosomal RNA processing in eukaryotes. Wiley Interdiscip Rev RNA. 2015, 6, 225–242. [Google Scholar] [CrossRef]
- Brito Querido, J.; Diaz-Lopez, I.; Ramakrishnan, V. The molecular basis of translation initiation and its regulation in eukaryotes. Nat Rev Mol Cell Biol. 2024, 25, 168–186. [Google Scholar] [CrossRef] [PubMed]
- Gentilella, A.; Kozma, S.C.; Thomas, G. A liaison between mTOR signaling, ribosome biogenesis and cancer. Biochim Biophys Acta. 2015, 1849, 812–820. [Google Scholar] [CrossRef] [PubMed]
- Seneff, S.; Nigh, G.; Kyriakopoulos, A.M.; McCullough, P.A. Innate immune suppression by SARS-CoV-2 mRNA vaccinations: The role of G-quadruplexes, exosomes, and MicroRNAs. Food Chem Toxicol. 2022, 164, 113008. [Google Scholar] [CrossRef] [PubMed]
- Kariko, K.; Buckstein, M.; Ni, H.; Weissman, D. Suppression of RNA recognition by Toll-like receptors: the impact of nucleoside modification and the evolutionary origin of RNA. Immunity. 2005, 23, 165–175. [Google Scholar] [CrossRef]
- Kariko, K.; Muramatsu, H.; Welsh, F.A.; Ludwig, J.; Kato, H.; Akira, S.; et al. Incorporation of pseudouridine into mRNA yields superior nonimmunogenic vector with increased translational capacity and biological stability. Mol Ther. 2008, 16, 1833–1840. [Google Scholar] [CrossRef]
- Anderson, B.R.; Muramatsu, H.; Jha, B.K.; Silverman, R.H.; Weissman, D.; Kariko, K. Nucleoside modifications in RNA limit activation of 2'-5'-oligoadenylate synthetase and increase resistance to cleavage by RNase L. Nucleic Acids Res. 2011, 39, 9329–9338. [Google Scholar] [CrossRef]
- Kariko, K.; Muramatsu, H.; Ludwig, J.; Weissman, D. Generating the optimal mRNA for therapy: HPLC purification eliminates immune activation and improves translation of nucleoside-modified, protein-encoding mRNA. Nucleic Acids Res. 2011, 39, e142. [Google Scholar] [CrossRef]
- Andries, O.; Mc Cafferty, S.; De Smedt, S.C.; Weiss, R.; Sanders, N.N.; Kitada, T. N(1)-methylpseudouridine-incorporated mRNA outperforms pseudouridine-incorporated mRNA by providing enhanced protein expression and reduced immunogenicity in mammalian cell lines and mice. J Control Release. 2015, 217, 337–344. [Google Scholar] [CrossRef]
- Lai, C.J.; Kim, D.; Kang, S.; Li, K.; Cha, I.; Sasaki, A.; et al. Viral codon optimization on SARS-CoV-2 Spike boosts immunity in the development of COVID-19 mRNA vaccines. J Med Virol. 2023, 95, e29183. [Google Scholar] [CrossRef]
- Kachaev, Z.M.; Lebedeva, L.A.; Kozlov, E.N.; Shidlovskii, Y.V. Interplay of mRNA capping and transcription machineries. Biosci Rep. 2020, 40. [Google Scholar] [CrossRef]
- Warminski, M.; Depaix, A.; Ziemkiewicz, K.; Spiewla, T.; Zuberek, J.; Drazkowska, K.; et al. Trinucleotide cap analogs with triphosphate chain modifications: synthesis, properties, and evaluation as mRNA capping reagents. Nucleic Acids Res. 2024, 52, 10788–10809. [Google Scholar] [CrossRef] [PubMed]
- Muttach, F.; Muthmann, N.; Rentmeister, A. Synthetic mRNA capping. Beilstein J Org Chem. 2017, 13, 2819–2832. [Google Scholar] [CrossRef] [PubMed]
- Imprachim, N.; Yosaatmadja, Y.; Newman, J.A. Crystal structures and fragment screening of SARS-CoV-2 NSP14 reveal details of exoribonuclease activation and mRNA capping and provide starting points for antiviral drug development. Nucleic Acids Res. 2023, 51, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Zhou, Y.; Zhang, S.; Chen, S.J. mRNA vaccine sequence and structure design and optimization: Advances and challenges. J Biol Chem. 2024, 301, 108015. [Google Scholar] [CrossRef]
- Grandi, C.; Emmaneel, M.; Nelissen, F.H.T.; Roosenboom, L.W.M.; Petrova, Y.; Elzokla, O.; et al. Decoupled degradation and translation enables noise modulation by poly(A) tails. Cell Syst. 2024, 15, 526–543. [Google Scholar] [CrossRef]
- Rauch, S.; Lutz, J.; Kowalczyk, A.; Schlake, T.; Heidenreich, R. RNActive(R) Technology: Generation and Testing of Stable and Immunogenic mRNA Vaccines. Methods Mol Biol. 2017, 1499, 89–107. [Google Scholar]
- Rauch, S.; Lutz, J.; Muhe, J.; Kowalczyk, A.; Schlake, T.; Heidenreich, R. Sequence-Optimized mRNA Vaccines Against Infectious Disease. Methods Mol Biol. 2024, 2786, 183–203. [Google Scholar]
- Kramps, T. Introduction to RNA Vaccines Post COVID-19. Methods Mol Biol. 2024, 2786, 1–22. [Google Scholar]
- Fu, Q.; Zhao, X.; Hu, J.; Jiao, Y.; Yan, Y.; Pan, X.; et al. mRNA vaccines in the context of cancer treatment: from concept to application. J Transl Med. 2025, 23, 12. [Google Scholar] [CrossRef]
- Garneau, N.L.; Wilusz, J.; Wilusz, C.J. The highways and byways of mRNA decay. Nat Rev Mol Cell Biol. 2007, 8, 113–126. [Google Scholar] [CrossRef]
- Assis, R.; Jain, A.; Nakajima, R.; Jasinskas, A.; Kahn, S.; Palma, A.; et al. Distinct SARS-CoV-2 Antibody Responses Elicited by Natural Infection and mRNA Vaccination. bioRxiv. 2021, 2021, 440089. [Google Scholar]
- Cho, A.; Muecksch, F.; Schaefer-Babajew, D.; Wang, Z.; Finkin, S.; Gaebler, C.; et al. Anti-SARS-CoV-2 receptor-binding domain antibody evolution after mRNA vaccination. Nature. 2021, 600, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Abou-Saleh, H.; Abo-Halawa, B.Y.; Younes, S.; Younes, N.; Al-Sadeq, D.W.; Shurrab, F.M.; et al. Neutralizing antibodies against SARS-CoV-2 are higher but decline faster in mRNA vaccinees compared to individuals with natural infection. Journal of Travel Medicine. 2022, 29, taac130. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Muecksch, F.; Schaefer-Babajew, D.; Finkin, S.; Viant, C.; Gaebler, C.; et al. Naturally enhanced neutralizing breadth against SARS-CoV-2 one year after infection. Nature. 2021, 595, 426–431. [Google Scholar] [CrossRef]
- Embgenbroich, M.; Burgdorf, S. Current Concepts of Antigen Cross-Presentation. Front Immunol. 2018, 9, 1643. [Google Scholar] [CrossRef]
- Kyriakopoulos, A.M.; Nigh, G.; McCullough, P.A.; Seneff, S. Autoimmune and Neoplastic Outcomes After the mRNA Vaccination: The Role of T Regulatory Cell Responses. International Journal of Vaccine Theory, Practice, and Research. 2024, 3, 1395–1433. [Google Scholar] [CrossRef]
- Nakatani, E.; Morioka, H.; Kikuchi, T.; Fukushima, M. Behavioral and Health Outcomes of mRNA COVID-19 Vaccination: A Case-Control Study in Japanese Small and Medium-Sized Enterprises. Cureus. 2024, 16, e75652. [Google Scholar] [CrossRef]
- Bansal, S.; Perincheri, S.; Fleming, T.; Poulson, C.; Tiffany, B.; Bremner, R.M.; et al. Cutting Edge: Circulating Exosomes with COVID Spike Protein Are Induced by BNT162b2 (Pfizer-BioNTech) Vaccination prior to Development of Antibodies: A Novel Mechanism for Immune Activation by mRNA Vaccines. J Immunol. 2021, 207, 2405–2410. [Google Scholar] [CrossRef]
- Tsai, S.J.; Atai, N.A.; Cacciottolo, M.; Nice, J.; Salehi, A.; Guo, C.; et al. Exosome-mediated mRNA delivery in vivo is safe and can be used to induce SARS-CoV-2 immunity. J Biol Chem. 2021, 297, 101266. [Google Scholar] [CrossRef]
- Cacciottolo, M.; Nice, J.B.; Li, Y.; LeClaire, M.J.; Twaddle, R.; Mora, C.L.; et al. Exosome-Based Multivalent Vaccine: Achieving Potent Immunization, Broadened Reactivity, and Strong T-Cell Responses with Nanograms of Proteins. Microbiol Spectr. 2023, 11, e0050323. [Google Scholar] [CrossRef]
- Pardi, N.; Hogan, M.J.; Naradikian, M.S.; Parkhouse, K.; Cain, D.W.; Jones, L.; et al. Nucleoside-modified mRNA vaccines induce potent T follicular helper and germinal center B cell responses. J Exp Med. 2018, 215, 1571–1588. [Google Scholar] [CrossRef] [PubMed]
- Zan, H.; Zhang, J.; Ardeshna, S.; Xu, Z.; Park, S.R.; Casali, P. Lupus-prone MRL/faslpr/lpr mice display increased AID expression and extensive DNA lesions, comprising deletions and insertions, in the immunoglobulin locus: concurrent upregulation of somatic hypermutation and class switch DNA recombination. Autoimmunity. 2009, 42, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Smith, D.; Aviszus, K.; Detanico, T.; Heiser, R.A.; Wysocki, L.J. Somatic hypermutation as a generator of antinuclear antibodies in a murine model of systemic autoimmunity. J Exp Med. 2010, 207, 2225–2237. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, H.; Okumura, F.; Miyabe, S.; Sugiura, M.; Joh, T.; Shimozato, K.; et al. Analysis of VH gene rearrangement and somatic hypermutation in Sjogren's syndrome and IgG4-related sclerosing sialadenitis. Scand J Immunol. 2010, 72, 44–49. [Google Scholar] [CrossRef]
- Zuckerman, N.S.; Hazanov, H.; Barak, M.; Edelman, H.; Hess, S.; Shcolnik, H.; et al. Somatic hypermutation and antigen-driven selection of B cells are altered in autoimmune diseases. J Autoimmun. 2010, 35, 325–335. [Google Scholar] [CrossRef]
- Okumura, F.; Sakuma, H.; Nakazawa, T.; Hayashi, K.; Naitoh, I.; Miyabe, K.; et al. Analysis of VH gene rearrangement and somatic hypermutation in type 1 autoimmune pancreatitis. Pathol Int. 2012, 62, 318–323. [Google Scholar] [CrossRef]
- Schroeder, K.; Herrmann, M.; Winkler, T.H. The role of somatic hypermutation in the generation of pathogenic antibodies in SLE. Autoimmunity. 2013, 46, 121–127. [Google Scholar] [CrossRef]
- Zhang, L.; Richards, A.; Barrasa, M.I.; Hughes, S.H.; Young, R.A.; Jaenisch, R. Reverse-transcribed SARS-CoV-2 RNA can integrate into the genome of cultured human cells and can be expressed in patient-derived tissues. Proc Natl Acad Sci U S A. 2021, 118. [Google Scholar] [CrossRef]
- Alden, M.; Olofsson Falla, F.; Yang, D.; Barghouth, M.; Luan, C.; Rasmussen, M.; et al. Intracellular Reverse Transcription of Pfizer BioNTech COVID-19 mRNA Vaccine BNT162b2 In Vitro in Human Liver Cell Line. Curr Issues Mol Biol. 2022, 44, 1115–1126. [Google Scholar] [CrossRef]
- Chandramouly, G.; Zhao, J.; McDevitt, S.; Rusanov, T.; Hoang, T.; Borisonnik, N.; et al. Poltheta reverse transcribes RNA and promotes RNA-templated DNA repair. Sci Adv. 2021, 7. [Google Scholar] [CrossRef]
- Merchant, H.A. Comment on Alden et al. Intracellular Reverse Transcription of Pfizer BioNTech COVID-19 mRNA Vaccine BNT162b2 In Vitro in Human Liver Cell Line. Curr. Issues Mol. Biol. 2022, 44, 1115–1126. [Google Scholar] [CrossRef] [PubMed]
- Brogna, C.; Cristoni, S.; Marino, G.; Montano, L.; Viduto, V.; Fabrowski, M.; et al. Detection of recombinant Spike protein in the blood of individuals vaccinated against SARS-CoV-2: Possible molecular mechanisms. Proteomics Clin Appl. 2023, 17, e2300048. [Google Scholar] [CrossRef] [PubMed]
- Krauson, A.J.; Casimero, F.V.C.; Siddiquee, Z.; Stone, J.R. Duration of SARS-CoV-2 mRNA vaccine persistence and factors associated with cardiac involvement in recently vaccinated patients. NPJ Vaccines. 2023, 8, 141. [Google Scholar] [CrossRef]
- Ndeupen, S.; Qin, Z.; Jacobsen, S.; Bouteau, A.; Estanbouli, H.; Igyarto, B.Z. The mRNA-LNP platform's lipid nanoparticle component used in preclinical vaccine studies is highly inflammatory. iScience. 2021, 24, 103479. [Google Scholar] [CrossRef]
- Stanworth, S.J.; New, H.V.; Apelseth, T.O.; Brunskill, S.; Cardigan, R.; Doree, C.; et al. Effects of the COVID-19 pandemic on supply and use of blood for transfusion. Lancet Haematol. 2020, 7, e756–e64. [Google Scholar] [CrossRef]
- Chang, L.; Yan, Y.; Wang, L. Coronavirus Disease 2019: Coronaviruses and Blood Safety. Transfus Med Rev. 2020, 34, 75–80. [Google Scholar] [CrossRef]
- Bouhou, S.; Lahjouji, K.; Masrar, A. Blood donor eligibility after COVID-19 vaccination: the current state of recommendations. Pan Afr Med J. 2021, 40, 207. [Google Scholar] [CrossRef]
- Jacobs, J.W.; Bibb, L.A.; Savani, B.N.; Booth, G.S. Refusing blood transfusions from COVID-19-vaccinated donors: are we repeating history? Br J Haematol. 2022, 196, 585–588. [Google Scholar] [CrossRef]
- Hunain, R.; Uday, U.; Rackimuthu, S.; Nawaz, F.A.; Narain, K.; Essar, M.Y.; et al. Effects of SARS-CoV-2 vaccination on blood donation and blood banks in India. Ann Med Surg (Lond). 2022, 78, 103772. [Google Scholar] [CrossRef]
- Roubinian, N.H.; Greene, J.; Liu, V.X.; Lee, C.; Mark, D.G.; Vinson, D.R.; et al. Clinical outcomes in hospitalized plasma and platelet transfusion recipients prior to and following widespread blood donor SARS-CoV-2 infection and vaccination. Transfusion. 2024, 64, 53–67. [Google Scholar] [CrossRef]
- Yonker, L.M.; Swank, Z.; Bartsch, Y.C.; Burns, M.D.; Kane, A.; Boribong, B.P.; et al. Circulating Spike Protein Detected in Post-COVID-19 mRNA Vaccine Myocarditis. Circulation. 2023, 147, 867–876. [Google Scholar] [CrossRef] [PubMed]
- Hanna, N.; De Mejia, C.M.; Heffes-Doon, A.; Lin, X.; Botros, B.; Gurzenda, E.; et al. Biodistribution of mRNA COVID-19 vaccines in human breast milk. EBioMedicine. 2023, 96, 104800. [Google Scholar] [CrossRef]
- Mulroney, T.E.; Poyry, T.; Yam-Puc, J.C.; Rust, M.; Harvey, R.F.; Kalmar, L.; et al. N(1)-methylpseudouridylation of mRNA causes +1 ribosomal frameshifting. Nature. 2024, 625, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Boros, L.G.; Kyriakopoulos, A.M.; Brogna, C.; Piscopo, M.; McCullough, P.A.; Seneff, S. Long-lasting, biochemically modified mRNA, and its frameshifted recombinant spike proteins in human tissues and circulation after COVID-19 vaccination. Pharmacol Res Perspect. 2024, 12, e1218. [Google Scholar] [CrossRef] [PubMed]
- Safra, M.; Tamari, Z.; Polak, P.; Shiber, S.; Matan, M.; Karameh, H.; et al. Altered somatic hypermutation patterns in COVID-19 patients classifies disease severity. Front Immunol. 2023, 14, 1031914. [Google Scholar] [CrossRef]
- Jacob-Dolan, C.; Lifton, M.; Powers, O.C.; Miller, J.; Hachmann, N.P.; Vu, M.; et al. B cell somatic hypermutation following COVID-19 vaccination with Ad26.COV2.S. iScience. 2024, 27, 109716. [Google Scholar] [CrossRef]
- Fay, M.M.; Lyons, S.M.; Ivanov, P. RNA G-Quadruplexes in Biology: Principles and Molecular Mechanisms. J Mol Biol. 2017, 429, 2127–2147. [Google Scholar] [CrossRef]
- Avolio, E.; Carrabba, M.; Milligan, R.; Kavanagh Williamson, M.; Beltrami, A.P.; Gupta, K.; et al. The SARS-CoV-2 Spike protein disrupts human cardiac pericytes function through CD147 receptor-mediated signalling: a potential non-infective mechanism of COVID-19 microvascular disease. Clin Sci (Lond). 2021, 135, 2667–2689. [Google Scholar] [CrossRef]
- Perico, L.; Morigi, M.; Pezzotta, A.; Locatelli, M.; Imberti, B.; Corna, D.; et al. SARS-CoV-2 spike protein induces lung endothelial cell dysfunction and thrombo-inflammation depending on the C3a/C3a receptor signalling. Sci Rep. 2023, 13, 11392. [Google Scholar] [CrossRef]
- Sahin, U.; Kariko, K.; Tureci, O. mRNA-based therapeutics--developing a new class of drugs. Nat Rev Drug Discov. 2014, 13, 759–780. [Google Scholar] [CrossRef]
- Grobbelaar, L.M.; Venter, C.; Vlok, M.; Ngoepe, M.; Laubscher, G.J.; Lourens, P.J.; et al. SARS-CoV-2 spike protein S1 induces fibrin(ogen) resistant to fibrinolysis: implications for microclot formation in COVID-19. Biosci Rep. 2021, 41. [Google Scholar] [CrossRef]
- White, J.; Herman, A.; Pullen, A.M.; Kubo, R.; Kappler, J.W.; Marrack, P. The V beta-specific superantigen staphylococcal enterotoxin B: stimulation of mature T cells and clonal deletion in neonatal mice. Cell. 1989, 56, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Noval Rivas, M.; Porritt, R.A.; Cheng, M.H.; Bahar, I.; Arditi, M. Multisystem Inflammatory Syndrome in Children and Long COVID: The SARS-CoV-2 Viral Superantigen Hypothesis. Front Immunol. 2022, 13, 941009. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, N.; Ratzon, R.; Hazan, I.; Zimmerman, D.R.; Singer, S.R.; Wasser, J.; et al. Multisystemic inflammatory syndrome in children and the BNT162b2 vaccine: a nationwide cohort study. Eur J Pediatr. 2024, 183, 3319–3326. [Google Scholar] [CrossRef]
- Li, H.; Llera, A.; Malchiodi, E.L.; Mariuzza, R.A. The structural basis of T cell activation by superantigens. Annu Rev Immunol. 1999, 17, 435–466. [Google Scholar] [CrossRef]
- Hamdy, A.; Leonardi, A. Superantigens and SARS-CoV-2. Pathogens. 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.; Bhardwaj, N. Super(antigen) target for SARS-CoV-2. Nature Reviews Immunology. 2021, 21, 72. [Google Scholar] [CrossRef]
- Lubinski, B.; Whittaker, G.R. The SARS-CoV-2 furin cleavage site: natural selection or smoking gun? Lancet Microbe. 2023, 4, e570. [Google Scholar] [CrossRef]
- Jung, F.; Connes, P. Morphology and Function of Red Blood Cells in COVID-19 Patients: Current Overview 2023. Life (Basel). 2024, 14. [Google Scholar] [CrossRef]
- Grau, M.; Ibershoff, L.; Zacher, J.; Bros, J.; Tomschi, F.; Diebold, K.F.; et al. Even patients with mild COVID-19 symptoms after SARS-CoV-2 infection show prolonged altered red blood cell morphology and rheological parameters. J Cell Mol Med. 2022, 26, 3022–3030. [Google Scholar] [CrossRef]
- Boschi, C.; Scheim, D.E.; Bancod, A.; Militello, M.; Bideau, M.L.; Colson, P.; et al. SARS-CoV-2 Spike Protein Induces Hemagglutination: Implications for COVID-19 Morbidities and Therapeutics and for Vaccine Adverse Effects. Int J Mol Sci. 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Scheim, D.E.; Vottero, P.; Santin, A.D.; Hirsh, A.G. Sialylated Glycan Bindings from SARS-CoV-2 Spike Protein to Blood and Endothelial Cells Govern the Severe Morbidities of COVID-19. Int J Mol Sci. 2023, 24. [Google Scholar] [CrossRef]
- Wang, R.; Tian, Z.; Zhu, M.; Zhang, B.; Li, Y.; Zheng, Y.; et al. SARS-CoV-2 spike protein potentiates platelet aggregation via upregulating integrin alphaIIbbeta3 outside-in signaling pathway. J Thromb Thrombolysis. 2024. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, C.C.; Basnet, N.; Bodakuntla, S.; Alvarez-Brecht, P.; Nichols, S.; Martinez-Sanchez, A.; et al. Direct Cryo-ET observation of platelet deformation induced by SARS-CoV-2 spike protein. Nat Commun. 2023, 14, 620. [Google Scholar] [CrossRef] [PubMed]
- Luzak, B.; Rozalski, M.; Przygodzki, T.; Boncler, M.; Wojkowska, D.; Kosmalski, M.; et al. SARS-CoV-2 Spike Protein and Neutralizing Anti-Spike Protein Antibodies Modulate Blood Platelet Function. Int J Mol Sci.
- Degenfeld-Schonburg, L.; Sadovnik, I.; Smiljkovic, D.; Peter, B.; Stefanzl, G.; Gstoettner, C.; et al. Coronavirus Receptor Expression Profiles in Human Mast Cells, Basophils, and Eosinophils. Cells. 2024, 13. [Google Scholar] [CrossRef]
- Faghihi, H. CD147 as an alternative binding site for the spike protein on the surface of SARS-CoV-2. Eur Rev Med Pharmacol Sci. 2020, 24, 11992–11994. [Google Scholar]
- Wang, K.; Chen, W.; Zhang, Z.; Deng, Y.; Lian, J.Q.; Du, P.; et al. CD147-spike protein is a novel route for SARS-CoV-2 infection to host cells. Signal Transduct Target Ther. 2020, 5, 283. [Google Scholar] [CrossRef]
- Behl, T.; Kaur, I.; Aleya, L.; Sehgal, A.; Singh, S.; Sharma, N.; et al. CD147-spike protein interaction in COVID-19: Get the ball rolling with a novel receptor and therapeutic target. Sci Total Environ. 2022, 808, 152072. [Google Scholar] [CrossRef]
- Brodowski, M.; Pierpaoli, M.; Janik, M.; Kowalski, M.; Ficek, M.; Slepski, P.; et al. Enhanced susceptibility of SARS-CoV-2 spike RBD protein assay targeted by cellular receptors ACE2 and CD147: Multivariate data analysis of multisine impedimetric response. Sens Actuators B Chem. 2022, 370, 132427. [Google Scholar] [CrossRef]
- Platt, L.R.; Estivariz, C.F.; Sutter, R.W. Vaccine-associated paralytic poliomyelitis: a review of the epidemiology and estimation of the global burden. J Infect Dis. 2014, 210 (Suppl 1), S380–S389. [Google Scholar] [CrossRef]
- Baraff, L.J.; Manclark, C.R.; Cherry, J.D.; Christenson, P.; Marcy, S.M. Analyses of adverse reactions to diphtheria and tetanus toxoids and pertussis vaccine by vaccine lot, endotoxin content, pertussis vaccine potency and percentage of mouse weight gain. Pediatr Infect Dis J. 1989, 8, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Ipp, M.; Goldbach, M.; Greenberg, S.; Gold, R. Effect of needle change and air bubble in syringe on minor adverse reactions associated with diphtheria-tetanus toxoids-pertussis-polio vaccination in infants. Pediatr Infect Dis J. 1990, 9, 291–293. [Google Scholar] [PubMed]
- Golaz, A.; Hardy, I.R.; Glushkevich, T.G.; Areytchiuk, E.K.; Deforest, A.; Strebel, P.; et al. Evaluation of a single dose of diphtheria-tetanus toxoids among adults in Odessa, Ukraine, 1995: immunogenicity and adverse reactions. J Infect Dis. 2000, 181 (Suppl 1), S203–S207. [Google Scholar] [CrossRef] [PubMed]
- Relyveld, E.H.; Bizzini, B.; Gupta, R.K. Rational approaches to reduce adverse reactions in man to vaccines containing tetanus and diphtheria toxoids. Vaccine. 1998, 16, 1016–1023. [Google Scholar] [CrossRef]
- Campbell, N.S.; Kalman, D.D. Successful vaccination with tetanus and diphtheria and acelluar pertussis vaccine after adverse reaction to diphtheria and tetanus toxoids and acellular pertussis vaccine or diphtheria and tetanus vaccine in pediatric patients. Ann Allergy Asthma Immunol. 2019, 123, 409–411. [Google Scholar] [CrossRef]
- Cullis, P.R.; Hope, M.J. Lipid Nanoparticle Systems for Enabling Gene Therapies. Mol Ther. 2017, 25, 1467–1475. [Google Scholar] [CrossRef]
- Cullis, P.R.; Felgner, P.L. The 60-year evolution of lipid nanoparticles for nucleic acid delivery. Nat Rev Drug Discov. 2024. [Google Scholar] [CrossRef]
- Parhiz, H.; Brenner, J.S.; Patel, P.N.; Papp, T.E.; Shahnawaz, H.; Li, Q.; et al. Added to pre-existing inflammation, mRNA-lipid nanoparticles induce inflammation exacerbation (IE). J Control Release. 2022, 344, 50–61. [Google Scholar] [CrossRef]
- Szoke, D.; Kovacs, G.; Kemecsei, E.; Balint, L.; Szotak-Ajtay, K.; Aradi, P.; et al. Nucleoside-modified VEGFC mRNA induces organ-specific lymphatic growth and reverses experimental lymphedema. Nat Commun. 2021, 12, 3460. [Google Scholar] [CrossRef]
- Anchordoquy, T.; Artzi, N.; Balyasnikova, I.V.; Barenholz, Y.; La-Beck, N.M.; Brenner, J.S.; et al. Mechanisms and Barriers in Nanomedicine: Progress in the Field and Future Directions. ACS Nano. 2024, 18, 13983–13999. [Google Scholar] [CrossRef]
- Sharma, P.; Hoorn, D.; Aitha, A.; Breier, D.; Peer, D. The immunostimulatory nature of mRNA lipid nanoparticles. Adv Drug Deliv Rev. 2024, 205, 115175. [Google Scholar] [CrossRef]
- Vogt, W. Anaphylatoxins: possible roles in disease. Complement. 1986, 3, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Schanzenbacher, J.; Kohl, J.; Karsten, C.M. Anaphylatoxins spark the flame in early autoimmunity. Front Immunol. 2022, 13, 958392. [Google Scholar] [CrossRef] [PubMed]
- Alving, C.R.; Kinsky, S.C.; Haxby, J.A.; Kinsky, C.B. Antibody binding and complement fixation by a liposomal model membrane. Biochemistry. 1969, 8, 1582–1587. [Google Scholar] [CrossRef] [PubMed]
- Szebeni, J. Complement activation-related pseudoallergy caused by liposomes, micellar carriers of intravenous drugs and radiocontrast agents. Crit Rev Ther Drug Carr Syst. 2001, 18, 567–606. [Google Scholar] [CrossRef]
- Szebeni, J. Complement activation-related pseudoallergy: a new class of drug-induced acute immune toxicity. Toxicology. 2005, 216, 106–121. [Google Scholar] [CrossRef]
- Szebeni, J. Complement activation-related pseudoallergy: A stress reaction in blood triggered by nanomedicines and biologicals. Mol Immunol. 2014, 61, 163–173. [Google Scholar] [CrossRef]
- Dezsi, L.; Meszaros, T.; Kozma, G.; M, H.V.; Olah, C.Z.; Szabo, M.; et al. A naturally hypersensitive porcine model may help understand the mechanism of COVID-19 mRNA vaccine-induced rare (pseudo) allergic reactions: complement activation as a possible contributing factor. Geroscience. 2022, 44, 597–618. [Google Scholar] [CrossRef]
- de Vrieze, J. Suspicions grow that nanoparticles in Pfizer's COVID-19 vaccine trigger rare allergic reactions. Science. 2021. [Google Scholar] [CrossRef]
- Szebeni, J.; Fontana, J.L.; Wassef, N.M.; Mongan, P.D.; Morse, D.S.; Dobbins, D.E.; et al. Hemodynamic changes induced by liposomes and liposome-encapsulated hemoglobin in pigs: a model for pseudoallergic cardiopulmonary reactions to liposomes. Role of complement and inhibition by soluble CR1 and anti-C5a antibody. Circulation. 1999, 99, 2302–2309. [Google Scholar] [CrossRef]
- Barth, I.; Weisser, K.; Gaston-Tischberger, D.; Mahler, V.; Keller-Stanislawski, B. Anaphylactic reactions after COVID-19 vaccination in Germany. Allergol Select. 2023, 7, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Barta, B.A.; Radovits, T.; Dobos, A.B.; Tibor Kozma, G.; Meszaros, T.; Berenyi, P.; et al. Comirnaty-induced cardiopulmonary distress and other symptoms of complement-mediated pseudo-anaphylaxis in a hyperimmune pig model: Causal role of anti-PEG antibodies. Vaccine X. 2024, 19, 100497. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Lai, S.K. Anti-PEG immunity: emergence, characteristics, and unaddressed questions. Wiley Interdiscip Rev Nanomed Nanobiotechnol. 2015, 7, 655–677. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.; Chen, B.M.; Su, Y.C.; Chang, Y.C.; Cheng, T.L.; Barenholz, Y.; et al. Premature Drug Release from Polyethylene Glycol (PEG)-Coated Liposomal Doxorubicin via Formation of the Membrane Attack Complex. ACS Nano. 2020, 14, 7808–7822. [Google Scholar] [CrossRef]
- Kozma, G.T.; Shimizu, T.; Ishida, T.; Szebeni, J. Anti-PEG antibodies: Properties, formation, testing and role in adverse immune reactions to PEGylated nano-biopharmaceuticals. Adv Drug Deliv Rev. 2020, 154–155, 163–175. [Google Scholar] [CrossRef]
- Gabizon, A.; Szebeni, J. Complement Activation: A Potential Threat on the Safety of Poly(ethylene glycol)-Coated Nanomedicines. ACS Nano. 2020, 14, 7682–7688. [Google Scholar] [CrossRef]
- USFDA. Guidance for Industry: Immunogenicity Assessment for Therapeutic Protein Products. https://wwwfdagov/regulatory-information/search-fda-guidance-documents/immunogenicity-assessment-therapeutic-protein-products. 2014;FDA-2013-D-0092.
- Bakos, T.; Meszaros, T.; Kozma, G.T.; Berenyi, P.; Facsko, R.; Farkas, H.; et al. mRNA-LNP COVID-19 Vaccine Lipids Induce Complement Activation and Production of Proinflammatory Cytokines: Mechanisms, Effects of Complement Inhibitors, and Relevance to Adverse Reactions. Int J Mol Sci. 2024, 25. [Google Scholar] [CrossRef]
- Lederer, K.; Castano, D.; Gomez Atria, D.; Oguin, T.H.; Wang, S., 3rd; Manzoni, T.B.; et al. SARS-CoV-2 mRNA Vaccines Foster Potent Antigen-Specific Germinal Center Responses Associated with Neutralizing Antibody Generation. Immunity. 2020, 53, 1281–1295. [Google Scholar] [CrossRef]
- Alameh, M.G.; Tombacz, I.; Bettini, E.; Lederer, K.; Sittplangkoon, C.; Wilmore, J.R.; et al. Lipid nanoparticles enhance the efficacy of mRNA and protein subunit vaccines by inducing robust T follicular helper cell and humoral responses. Immunity. 2021, 54, 2877–2892. [Google Scholar] [CrossRef]
- Alameh, M.G.; Tombacz, I.; Bettini, E.; Lederer, K.; Sittplangkoon, C.; Wilmore, J.R.; et al. Lipid nanoparticles enhance the efficacy of mRNA and protein subunit vaccines by inducing robust T follicular helper cell and humoral responses. Immunity. 2022, 55, 1136–1138. [Google Scholar] [CrossRef]
- Hemmrich, E.; McNeil, S. Active ingredient vs excipient debate for nanomedicines. Nat Nanotechnol. 2023, 18, 692–695. [Google Scholar] [CrossRef] [PubMed]
- Akinc, A.; Maier, M.A.; Manoharan, M.; Fitzgerald, K.; Jayaraman, M.; Barros, S.; et al. The Onpattro story and the clinical translation of nanomedicines containing nucleic acid-based drugs. Nat Nanotechnol. 2019, 14, 1084–1087. [Google Scholar] [CrossRef] [PubMed]
- Dalby, B.; Cates, S.; Harris, A.; Ohki, E.C.; Tilkins, M.L.; Price, P.J.; et al. Advanced transfection with Lipofectamine 2000 reagent: primary neurons, siRNA, and high-throughput applications. Methods. 2004, 33, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Ferraresso, F.; Badior, K.; Seadler, M.; Zhang, Y.; Wietrzny, A.; Cau, M.F.; et al. Protein is expressed in all major organs after intravenous infusion of mRNA-lipid nanoparticles in swine. Mol Ther Methods Clin Dev. 2024, 32, 101314. [Google Scholar] [CrossRef]
- Pateev, I.; Seregina, K.; Ivanov, R.; Reshetnikov, V. Biodistribution of RNA Vaccines and of Their Products: Evidence from Human and Animal Studies. Biomedicines. 2023, 12. [Google Scholar] [CrossRef]
- Lachmann, P.J. The amplification loop of the complement pathways. Adv Immunol. 2009, 104, 115–149. [Google Scholar]
- lebel, P. COMIRNATY® (COVID-19 Vaccine, mRNA) suspension for injection, for intramuscular use. https://labelingpfizercom/ShowLabelingaspx?id=16351#section-6. 1023.
- Dempsey, P.W.; Allison, M.E.; Akkaraju, S.; Goodnow, C.C.; Fearon, D.T. C3d of complement as a molecular adjuvant: bridging innate and acquired immunity. Science. 1996, 271, 348–350. [Google Scholar] [CrossRef]
- Wills-Karp, M. Complement activation pathways: a bridge between innate and adaptive immune responses in asthma. Proc Am Thorac Soc. 2007, 4, 247–251. [Google Scholar] [CrossRef]
- Kozma, G.T.; Meszaros, T.; Berenyi, P.; Facsko, R.; Patko, Z.; Olah, C.Z.; et al. Role of anti-polyethylene glycol (PEG) antibodies in the allergic reactions to PEG-containing Covid-19 vaccines: Evidence for immunogenicity of PEG. Vaccine. 2023, 41, 4561–4570. [Google Scholar] [CrossRef]
- Carreno, J.M.; Singh, G.; Tcheou, J.; Srivastava, K.; Gleason, C.; Muramatsu, H.; et al. mRNA-1273 but not BNT162b2 induces antibodies against polyethylene glycol (PEG) contained in mRNA-based vaccine formulations. Vaccine. 2022, 40, 6114–6124. [Google Scholar] [CrossRef]
- Ju, Y.; Lee, W.S.; Pilkington, E.H.; Kelly, H.G.; Li, S.; Selva, K.J.; et al. Anti-PEG Antibodies Boosted in Humans by SARS-CoV-2 Lipid Nanoparticle mRNA Vaccine. ACS Nano. 2022, 16, 11769–11780. [Google Scholar] [CrossRef] [PubMed]
- Bavli, Y.; Chen, B.M.; Gross, G.; Hershko, A.; Turjeman, K.; Roffler, S.; et al. Anti-PEG antibodies before and after a first dose of Comirnaty(R) (mRNA-LNP-based SARS-CoV-2 vaccine). J Control Release. 2023, 354, 316–322. [Google Scholar] [CrossRef]
- Cheng, M.H.Y.; Leung, J.; Zhang, Y.; Strong, C.; Basha, G.; Momeni, A.; et al. Induction of Bleb Structures in Lipid Nanoparticle Formulations of mRNA Leads to Improved Transfection Potency. Adv Mater. 2023, 35, e2303370. [Google Scholar] [CrossRef] [PubMed]
- Pfizer, Biontech. Comirnaty Original Omicron BA.4-5. https://labelingpfizercom/ShowLabelingaspx?id=19823. 2023.
- Boros, L.G.; Seneff, S.; Turi, M.; Palcsu, L.; Zubarev, R.A. Active involvement of compartmental, inter- and intramolecular deuterium disequilibrium in adaptive biology. Proc Natl Acad Sci U S A. 2024, 121, e2412390121. [Google Scholar] [CrossRef]
- Korchinsky, N.; Davis, A.M.; Boros, L.G. Nutritional deuterium depletion and health: a scoping review. Metabolomics. 2024, 20, 117. [Google Scholar] [CrossRef]
- Wrapp, D.; Wang, N.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.L.; Abiona, O.; et al. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science. 2020, 367, 1260–1263. [Google Scholar] [CrossRef]
- Raoult, D. Confirmation of the presence of vaccine DNA in the Pfizer anti-COVID-19 vaccine. Hal Open Science. 2024, 04778576. [Google Scholar]
- Lee, M.; Ki-Yeob, J. Foreign Materials in Blood Samples of Recipients of COVID19 Vaccines. International Journal of Vaccine Theory, Practice, and Research. 2022, 2, 249–265. [Google Scholar] [CrossRef]
- Wang, T.J.; Kim, A.; Kim, K. Method of Replication-Competent Plasmid DNA from COVID-19 mRNA Vaccines for Quality Control. Journal of High School Science. 2024, 8, 427–439. [Google Scholar]
- Kämmerer, U.; Schulz, V.; Steger, K. BioNTech RNA-Based COVID-19 Injections Contain Large Amounts Of Residual DNA Including An SV40 Promoter/Enhancer Sequence. Science, Public Health Policy and the Law. 2024, 5, 2019–2024. [Google Scholar]
- CGC. Case briefing document. http://ukcitizen, 2024.
- Doherty, M.; Whicher, J.T.; Dieppe, P.A. Activation of the alternative pathway of complement by monosodium urate monohydrate crystals and other inflammatory particles. Ann Rheum Dis. 1983, 42, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Russell, I.J.; Mansen, C.; Kolb, L.M.; Kolb, W.P. Activation of the fifth component of human complement (C5) induced by monosodium urate crystals: C5 convertase assembly on the crystal surface. Clin Immunol Immunopathol. 1982, 24, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Fields, T.R.; Abramson, S.B.; Weissmann, G.; Kaplan, A.P.; Ghebrehiwet, B. Activation of the alternative pathway of complement by monosodium urate crystals. Clin Immunol Immunopathol. 1983, 26, 249–257. [Google Scholar] [CrossRef]
- Baranyi, L.; Szebeni, J.; Savay, S.; Bodo, M.; Bunger, R.; Alving, C. ; editors. When cholesterol kills: fatal complement activation-mediated circulatory distress provoked by cholesterol-enriched liposomes. FASEB, 2000; 20814-3998. [Google Scholar]
- Baranyi, L.; Szebeni, J.; Savay, S.; Bodo, M.; Basta, M.; Bentley, T.B.; et al. Complement-Dependent Shock and Tissue Damage Induced by Intravenous Injection of Cholesterol-Enriched Liposomes in Rats. Journal of Applied Research. 2003, 3. [Google Scholar]
- Ratajczak, M.Z.; Lee, H.; Wysoczynski, M.; Wan, W.; Marlicz, W.; Laughlin, M.J.; et al. Novel insight into stem cell mobilization-plasma sphingosine-1-phosphate is a major chemoattractant that directs the egress of hematopoietic stem progenitor cells from the bone marrow and its level in peripheral blood increases during mobilization due to activation of complement cascade/membrane attack complex. Leukemia. 2010, 24, 976–985. [Google Scholar]
- Klein, C.P.; de Groot, K.; van Kamp, G. Activation of complement C3 by different calcium phosphate powders. Biomaterials. 1983, 4, 181–184. [Google Scholar] [CrossRef]
- Hasselbacher, P. Binding of immunoglobulin and activation of complement by asbestos fibers. J Allergy Clin Immunol. 1979, 64, 294–298. [Google Scholar] [CrossRef]
- Szebeni, J.; Alving, C.R.; Savay, S.; Barenholz, Y.; Priev, A.; Danino, D.; et al. Formation of complement-activating particles in aqueous solutions of Taxol: possible role in hypersensitivity reactions. Int Immunopharmacol. 2001, 1, 721–735. [Google Scholar] [CrossRef]
- Qin, Z.; Bouteau, A.; Herbst, C.; Igyarto, B.Z. Pre-exposure to mRNA-LNP inhibits adaptive immune responses and alters innate immune fitness in an inheritable fashion. PLoS Pathog 2022, 18, e1010830. [Google Scholar] [CrossRef]
- Valdes Angues, R.; Perea Bustos, Y. SARS-CoV-2 Vaccination and the Multi-Hit Hypothesis of Oncogenesis. Cureus. 2023, 15, e50703. [Google Scholar] [CrossRef]
- Kruger, U. Covid Vaccination and Turbo-Cancer. Multiple Tumors in Multiple Organs. https://wwwglobalresearchca/turbo-cancer-we-have-problem/5789172 2024.
- Wiedmeier-Nutor, J.E.; Iqbal, M.; Rosenthal, A.C.; Bezerra, E.D.; Garcia-Robledo, J.E.; Bansal, R.; et al. Response to COVID-19 Vaccination Post-CAR T Therapy in Patients With Non-Hodgkin Lymphoma and Multiple Myeloma. Clin Lymphoma Myeloma Leuk. 2023, 23, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Sekizawa, A.; Hashimoto, K.; Kobayashi, S.; Kozono, S.; Kobayashi, T.; Kawamura, Y.; et al. Rapid progression of marginal zone B-cell lymphoma after COVID-19 vaccination (BNT162b2): A case report. Front Med (Lausanne). 2022, 9, 963393. [Google Scholar] [CrossRef] [PubMed]
- Nacionales, D.C.; Weinstein, J.S.; Yan, X.J.; Albesiano, E.; Lee, P.Y.; Kelly-Scumpia, K.M.; et al. B cell proliferation, somatic hypermutation, class switch recombination, and autoantibody production in ectopic lymphoid tissue in murine lupus. J Immunol. 2009, 182, 4226–4236. [Google Scholar] [CrossRef] [PubMed]
- Gentilini, P.; Lindsay, J.C.; Konishi, N.; Fukushima, M.; Polykretis, P. A Case Report of Acute Lymphoblastic Leukaemia (ALL)/Lymphoblastic Lymphoma (LBL) Following the Second Dose of Comirnaty®: An Analysis of the Potential Pathogenic Mechanism Based on of the Existing Literature. Preprints Org. 2024. [CrossRef]
- Hill, A.B. The Environment and Disease: Association or Causation? Proc R Soc Med. 1965, 58, 295–300. [Google Scholar] [CrossRef]
- Menetrier-Caux, C.; Ray-Coquard, I.; Blay, J.Y.; Caux, C. Lymphopenia in Cancer Patients and its Effects on Response to Immunotherapy: an opportunity for combination with Cytokines? J Immunother Cancer. 2019, 7, 85. [Google Scholar] [CrossRef]
- Zhang, S.; El-Deiry, W.S. Transfected SARS-CoV-2 spike DNA for mammalian cell expression inhibits p53 activation of p21(WAF1), TRAIL Death Receptor DR5 and MDM2 proteins in cancer cells and increases cancer cell viability after chemotherapy exposure. Oncotarget. 2024, 15, 275–284. [Google Scholar] [CrossRef]
- Stockert, J.A.; Gupta, A.; Herzog, B.; Yadav, S.S.; Tewari, A.K.; Yadav, K.K. Predictive value of pseudouridine in prostate cancer. Am J Clin Exp Urol. 2019, 7, 262–272. [Google Scholar]
- Stockert, J.A.; Weil, R.; Yadav, K.K.; Kyprianou, N.; Tewari, A.K. Pseudouridine as a novel biomarker in prostate cancer. Urol Oncol. 2021, 39, 63–71. [Google Scholar] [CrossRef]
- Han, X.; Wang, M.; Zhao, Y.L.; Yang, Y.; Yang, Y.G. RNA methylations in human cancers. Semin Cancer Biol. 2021, 75, 97–115. [Google Scholar] [CrossRef]
- Raszek, M.; Cowley, D.; Redwan, E.M.; Vladimir, N.; Uversky, V.N.; Rubio-Casillas, A. Exploring the possible link between the spike protein immunoglobulin G4 antibodies and cancer progression. Exploration of immunology. 2024, 4, 267–284. [Google Scholar] [CrossRef]
- Mansoori, B.; Mohammadi, A.; Shirjang, S.; Baradaran, B. Micro-RNAs: The new potential biomarkers in cancer diagnosis, prognosis and cancer therapy. Cell Mol Biol (Noisy-le-grand). 2015, 61, 1–10. [Google Scholar] [PubMed]
- Chang, W.; Chang, Q.; Lu, H.; Liu, S.; Li, Y.; Chen, C. MicroRNA-873-5p suppresses cell malignant behaviors of thyroid cancer via targeting CXCL5 and regulating P53 pathway. Hum Vaccin Immunother. 2022, 18, 2065837. [Google Scholar] [CrossRef] [PubMed]
- Trifylli, E.M.; Kriebardis, A.G.; Koustas, E.; Papadopoulos, N.; Fortis, S.P.; Tzounakas, V.L.; et al. A Current Synopsis of the Emerging Role of Extracellular Vesicles and Micro-RNAs in Pancreatic Cancer: A Forward-Looking Plan for Diagnosis and Treatment. Int J Mol Sci. 2024, 25. [Google Scholar] [CrossRef]
- Wilson, E.; Goswami, J.; Baqui, A.H.; Doreski, P.A.; Perez-Marc, G.; Zaman, K.; et al. Efficacy and Safety of an mRNA-Based RSV PreF Vaccine in Older Adults. N Engl J Med. 2023, 389, 2233–2244. [Google Scholar] [CrossRef]
- Moderna. Prescribing Information: Respiratory Syncytial Virus Vaccine) Injectable suspension, for intramuscular use. https://wwwfdagov/media/179005/download?attachment=&utm_source=chatgptcom. 2024;https://www.fda.gov/media/179005/download?attachment=&utm_source=chatgpt.com.
- FDA. FDA Approves and Authorizes Updated mRNA COVID-19 Vaccines to Better Protect Against Currently Circulating Variants. https://wwwfdagov/news-events/press-announcements/fda-approves-and-authorizes-updated-mrna-covid-19-vaccines-better-protect-against-currently. 2024;https://www.fda.gov/news-events/press-announcements/fda-approves-and-authorizes-updated-mrna-covid-19-vaccines-better-protect-against-currently.
- Zhang, G.; Tang, T.; Chen, Y.; Huang, X.; Liang, T. mRNA vaccines in disease prevention and treatment. Signal Transduct Target Ther. 2023, 8, 365. [Google Scholar] [CrossRef]
- Ladak, R.J.; He, A.J.; Huang, Y.H.; Ding, Y. The Current Landscape of mRNA Vaccines Against Viruses and Cancer-A Mini Review. Front Immunol. 2022, 13, 885371. [Google Scholar] [CrossRef]
- Sayour, E.J.; Boczkowski, D.; Mitchell, D.A.; Nair, S.K. Cancer mRNA vaccines: clinical advances and future opportunities. Nat Rev Clin Oncol. 2024, 21, 489–500. [Google Scholar] [CrossRef]
- Xie, W.; Chen, B.; Wong, J. Evolution of the market for mRNA technology. Nat Rev Drug Discov. 2021, 20, 735–736. [Google Scholar] [CrossRef]
- Application Therapeutics, Research. Lipid Nanoparticles Market Size, Share & Industry Analysis, By Type (Solid Lipid Nanoparticles (SLNs), Nanostructured Lipid Carriers (NLCs), and Others). https://wwwfortunebusinessinsightscom/lipid-nanoparticles-market-106960. 2024;https://www.fortunebusinessinsights.com.
- mRNA Vaccine and Therapeutics Market Size & Share Analysis - Growth Trends & Forecasts (2024 - 2029) https://wwwmordorintelligencecom/industry-reports/mrna-vaccines-and-therapeutics-market. 2024;https://www.mordorintelligence.com/industry-reports/mrna-vaccines-and-therapeutics-market.
- The hope accord. https://thehopeaccordorg/?utm_source=chatgptcom. 2024.
- Housfeld. COVID19 Inquiry: The treatment of the vaccine injured in this country has historically been a source of shame. https://wwwhausfeldcom/en-gb/news/covid19-inquiry-the-treatment-of-the-vaccine-injured-in-this-country-has-historically-been-a-source-of-shame/?utm_source=chatgptcom. 2025.
- Association, B.M. Engagement with the UK Covid-19 Inquiry. Module 4: Vaccines and therapeutics. 2025;https://www.bma.org.uk/advice-and-support/covid-19/what-the-bma-is-doing/bma-engagement-with-the-uk-covid-19-inquiry?utm_source=chatgpt.com(https://www.bma.org.uk/advice-and-support/covid-19/what-the-bma-is-doing/bma-engagement-with-the-uk-covid-19-inquiry?utm_source=chatgpt.com).
- NVIC. Protecting Health and Informed Consent Rights Since 1982. https://wwwnvicorg/ 2025.
- Help. https://www.worldcouncilforhealth.org/. https://wwwworldcouncilforhealthorg/ [Internet]. 2025; https://www.worldcouncilforhealth.org/.
- The Times. State’s ‘fatal strategic flaws’ during Covid are laid bare. The Times https://wwwthetimescom/uk/healthcare/article/states-failure-to-protect-public-during-covid-laid-bare-p2tp93xvd?utm_source=chatgptcom®ion=global. 2024;Thursday July 18 2024.
- UK COVID-19 Inquiry. https://wwwgovuk/government/publications/uk-covid-19-inquiry-terms-of-reference. 2024.
- Litaker, J.R.; Lopez Bray, C.; Tamez, N.; Durkalski, W.; Taylor, R. COVID-19 Vaccine Acceptors, Refusers, and the Moveable Middle: A Qualitative Study from Central Texas. Vaccines (Basel). 2022, 10. [Google Scholar] [CrossRef]
- FDA. Considerations for Respiratory Syncytial Virus (RSV) Vaccine Safety in Pediatric Populations. VRBPAC Briefing Document. 2024;https://www.fda.gov/media/184301/download.
- Igyarto, B.Z.; Jacobsen, S.; Ndeupen, S. Future considerations for the mRNA-lipid nanoparticle vaccine platform. Curr Opin Virol. 2021, 48, 65–72. [Google Scholar] [CrossRef]
Figure 1.
Structural and Functional Insights into Comirnaty LNPs. The figure presents schematic models and transmission electron micrographs (TEM) of Comirnaty lipid nanoparticles (LNPs) to highlight their molecular structure and dynamic behavior. Panels A-D illustrate conceptual depictions of the spatial relationships between the lipids and the mRNA in the LNPs. Panel E introduces the hydrogen bond-crosslinked "thread ball" model. Panels F and H display uranyl acetate-stained TEM images of Comirnaty LNPs. In Panel F, 2−5 nm electron-dense spots are visible, forming string-like, semicircular, or labyrinthine patterns, which support the thread ball hypothesis. In Panel G, yellow arrows point to fusion or aggregation events, while white arrows highlight worm-like “tails” extruding from the LNPs. A closer view of one such extrusion event in Panel H gives the impression of a sperm-like structure, suggesting its potential role in transfection (the delivery of genetic material), figure modified from Ref. [55] with permission.
Figure 1.
Structural and Functional Insights into Comirnaty LNPs. The figure presents schematic models and transmission electron micrographs (TEM) of Comirnaty lipid nanoparticles (LNPs) to highlight their molecular structure and dynamic behavior. Panels A-D illustrate conceptual depictions of the spatial relationships between the lipids and the mRNA in the LNPs. Panel E introduces the hydrogen bond-crosslinked "thread ball" model. Panels F and H display uranyl acetate-stained TEM images of Comirnaty LNPs. In Panel F, 2−5 nm electron-dense spots are visible, forming string-like, semicircular, or labyrinthine patterns, which support the thread ball hypothesis. In Panel G, yellow arrows point to fusion or aggregation events, while white arrows highlight worm-like “tails” extruding from the LNPs. A closer view of one such extrusion event in Panel H gives the impression of a sperm-like structure, suggesting its potential role in transfection (the delivery of genetic material), figure modified from Ref. [55] with permission.
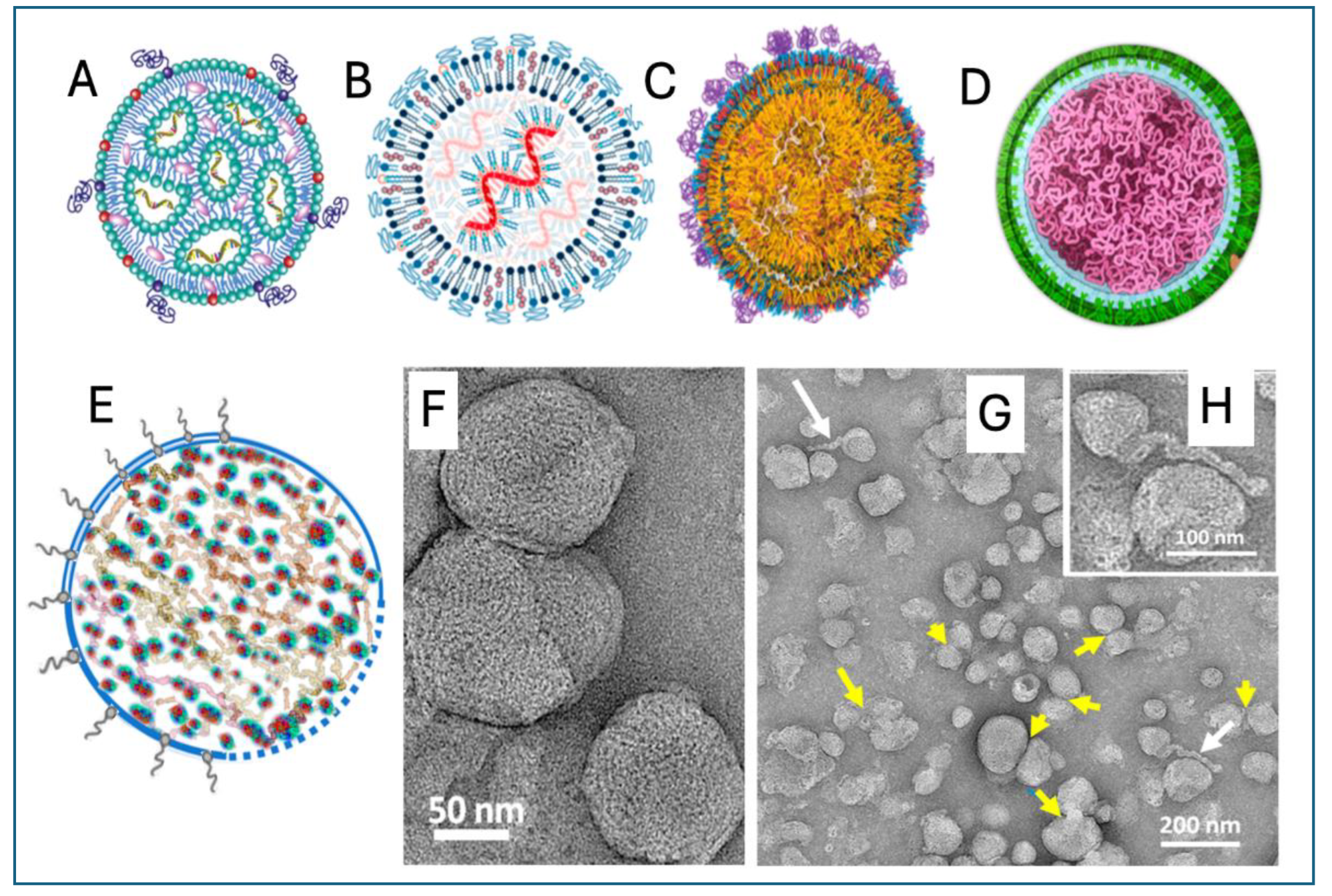
Figure 2.
The mechanism of mRNA-LNP vaccine action. The figure, reproduced from the review by Chaudhary et al., [67] with permission, illustrates the steps via which the vaccine induces immunogenicity against the spike protein (SP) in dendritic or other APCs. The uptake of mRNA-LNPs via phagocytosis (step 1) is followed by the release of the mRNA from the endosomes and translation on ribosomes to the full-length SP trimer (step 2). Thereafter the SP may be expressed on the cell membrane as seen on SARS-2-CoV (top 3 darts), it can be secreted by the cell (bottom dart) or processed by proteasomes to yield antigenic fragments that are presented on MHC I surface receptors (step 3). These teach cytotoxic T cells to recognize the antigen and trigger apoptosis in virus-infected cells (step 4). The SP molecules liberated to the extracellular space get phagocytosed in an autocrine fashion to feed MHC Class-II molecules via endo-lysosomes (step 5, MHC II not labelled), to “educate” Th and B cells for specific neutralizing antibody production (step 6). Abbreviations: BCR, B cell receptor; ER, endoplasmic reticulum; TCR, T-cell receptor; BCR, B-cell receptor. Further details are given in ref. [67].
Figure 2.
The mechanism of mRNA-LNP vaccine action. The figure, reproduced from the review by Chaudhary et al., [67] with permission, illustrates the steps via which the vaccine induces immunogenicity against the spike protein (SP) in dendritic or other APCs. The uptake of mRNA-LNPs via phagocytosis (step 1) is followed by the release of the mRNA from the endosomes and translation on ribosomes to the full-length SP trimer (step 2). Thereafter the SP may be expressed on the cell membrane as seen on SARS-2-CoV (top 3 darts), it can be secreted by the cell (bottom dart) or processed by proteasomes to yield antigenic fragments that are presented on MHC I surface receptors (step 3). These teach cytotoxic T cells to recognize the antigen and trigger apoptosis in virus-infected cells (step 4). The SP molecules liberated to the extracellular space get phagocytosed in an autocrine fashion to feed MHC Class-II molecules via endo-lysosomes (step 5, MHC II not labelled), to “educate” Th and B cells for specific neutralizing antibody production (step 6). Abbreviations: BCR, B cell receptor; ER, endoplasmic reticulum; TCR, T-cell receptor; BCR, B-cell receptor. Further details are given in ref. [67].
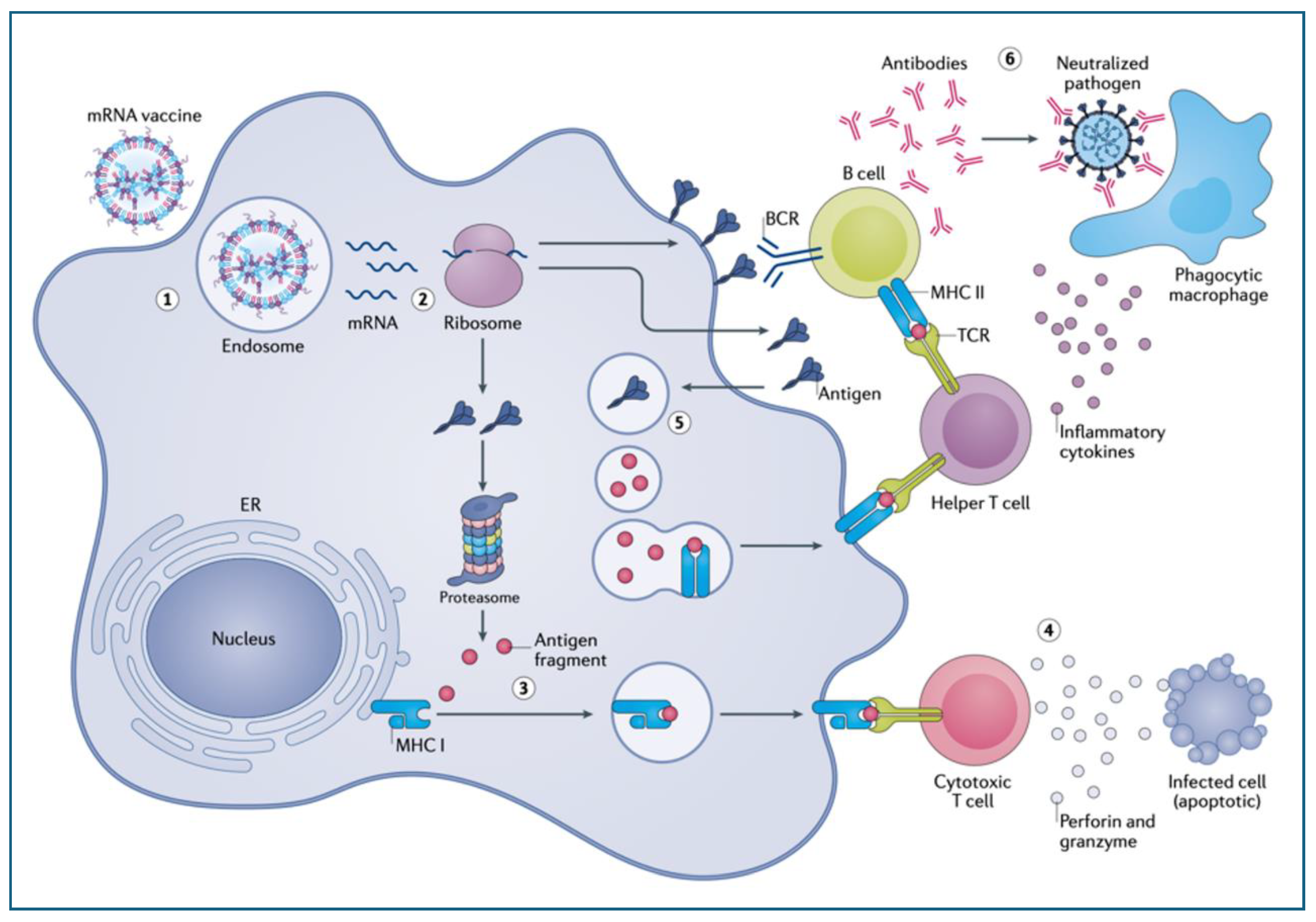
Figure 3.
Potential autoimmune attack against APCs by SP-specific cytotoxic T cells with increasing impact after booster vaccinations. The scheme applies to all body cells transfected with the vaccine and expressing SP fragments on their surface Class-I molecules.
Figure 3.
Potential autoimmune attack against APCs by SP-specific cytotoxic T cells with increasing impact after booster vaccinations. The scheme applies to all body cells transfected with the vaccine and expressing SP fragments on their surface Class-I molecules.
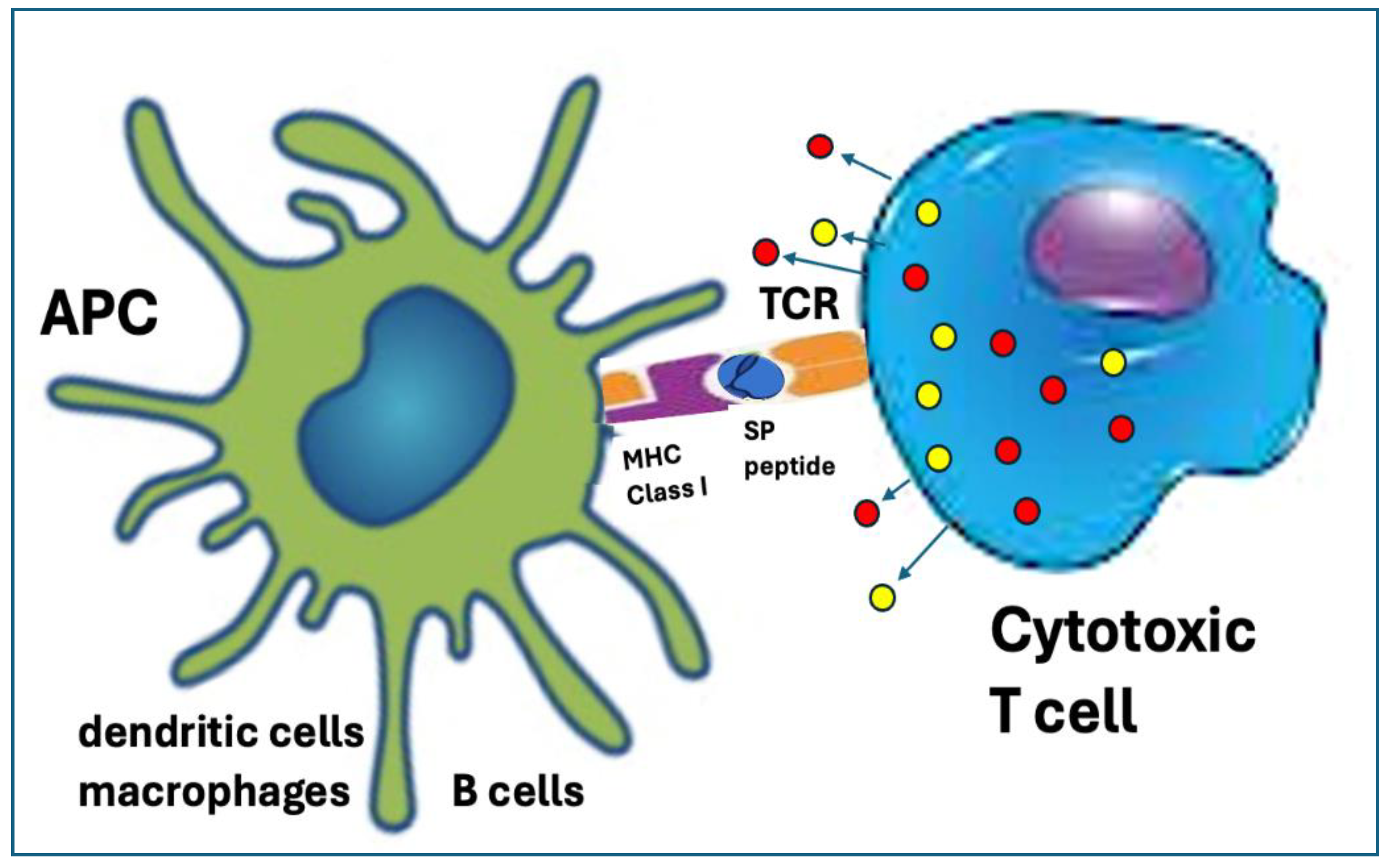
Figure 4.
Potential secondary immune processes triggered by vaccination with mRNA-LNPs, after the rise of specific anti-SP antibodies. A) Anti-SP specific antibody (aSP)-dependent cellular cytotoxicity (ADCC), whereupon natural killer (NK) cells, macrophages or neutrophils recognize the cell-bound antibodies’ Fc portion via their Fc receptor (FcR) and release cytotoxic granules inducing apoptosis. B) Cell-bound antibodies initiate classical pathway C activation starting with the binding of C1q and ending in the formation of the membrane attack complex (MAC), which causes membrane damage. The SP on the APC surface also activates C via the lectin pathway, by binding mannose binding lectin (MBL) or ficolin. The end result is the same: MAC-mediated cytotoxicity.
Figure 4.
Potential secondary immune processes triggered by vaccination with mRNA-LNPs, after the rise of specific anti-SP antibodies. A) Anti-SP specific antibody (aSP)-dependent cellular cytotoxicity (ADCC), whereupon natural killer (NK) cells, macrophages or neutrophils recognize the cell-bound antibodies’ Fc portion via their Fc receptor (FcR) and release cytotoxic granules inducing apoptosis. B) Cell-bound antibodies initiate classical pathway C activation starting with the binding of C1q and ending in the formation of the membrane attack complex (MAC), which causes membrane damage. The SP on the APC surface also activates C via the lectin pathway, by binding mannose binding lectin (MBL) or ficolin. The end result is the same: MAC-mediated cytotoxicity.
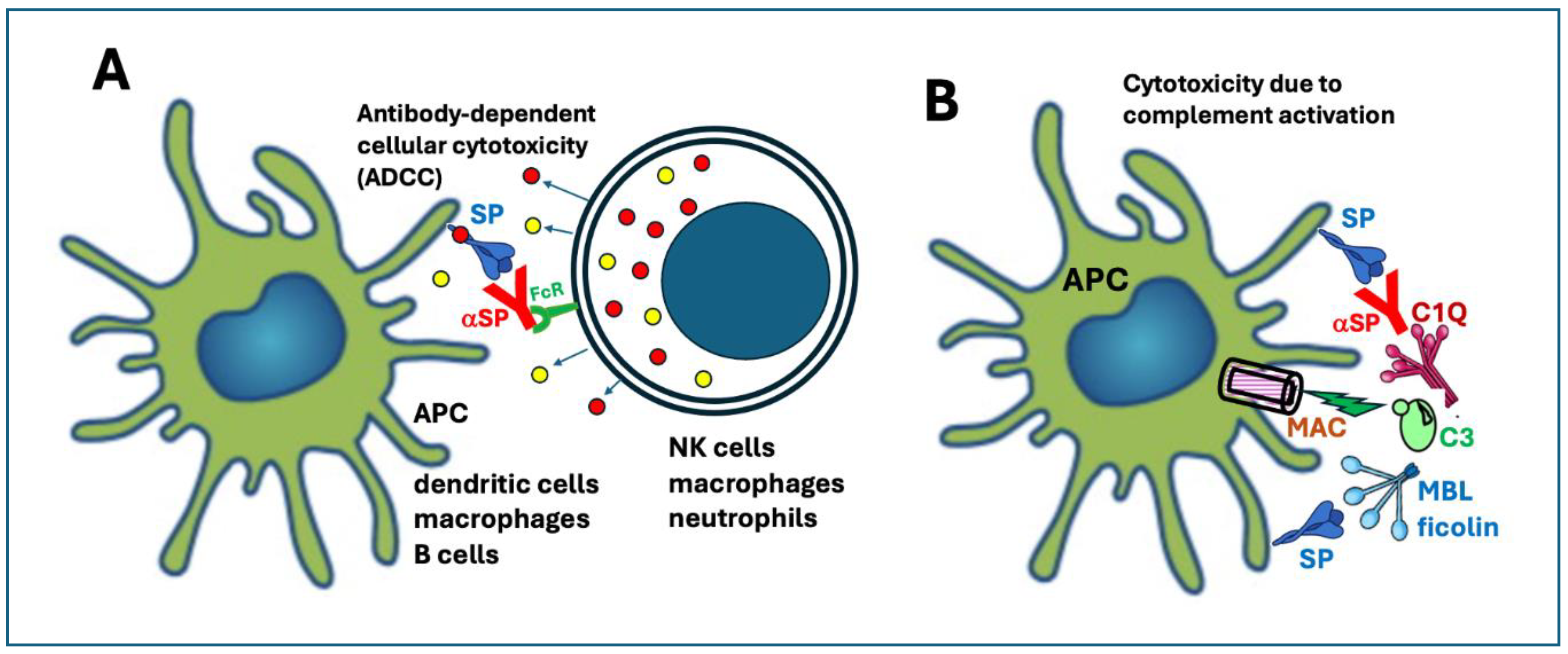
Figure 5.
The superantigen-like activity of the spike protein. A) Illustration of the effects of superantigens. Superantigen of bacterial origin can bind APC MHC Class-II receptor directly to the variable region of T cell receptor b chain, leading to massive nonspecific stimulation of both APC and T cells with cytokine release. B) The superantigen hypothesis raises the possibility that the SP of vaccine origin, crowning the APC, can bind to the TCR vβ chain via the superantigen motif on SP. This results in massive nonspecific immune stimulation with cytokine release or storm.
Figure 5.
The superantigen-like activity of the spike protein. A) Illustration of the effects of superantigens. Superantigen of bacterial origin can bind APC MHC Class-II receptor directly to the variable region of T cell receptor b chain, leading to massive nonspecific stimulation of both APC and T cells with cytokine release. B) The superantigen hypothesis raises the possibility that the SP of vaccine origin, crowning the APC, can bind to the TCR vβ chain via the superantigen motif on SP. This results in massive nonspecific immune stimulation with cytokine release or storm.
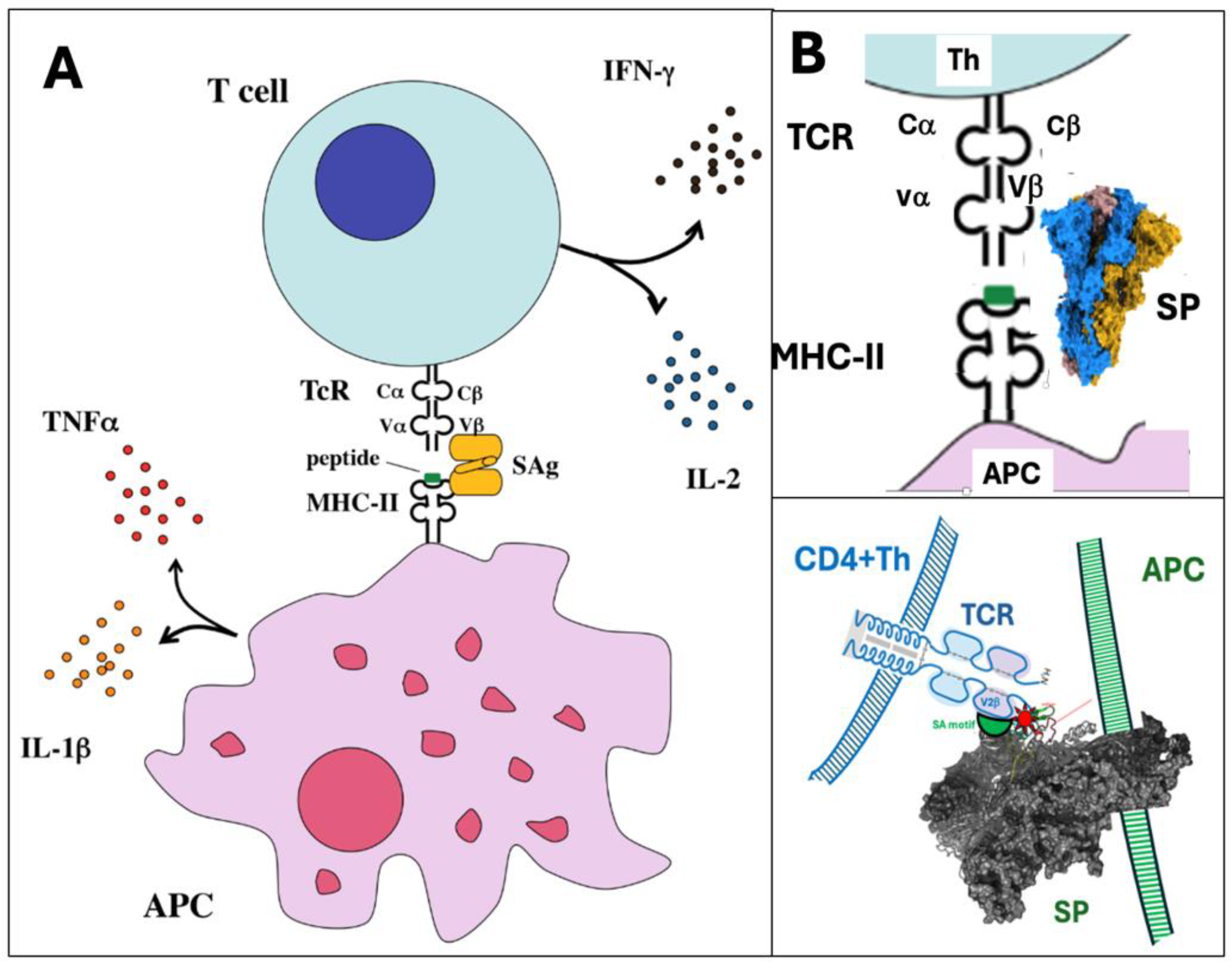

Table 1.
The physical and chemical properties of mRNA-LNP vaccines that can be associated with theoretical risks for AEs.
Table 1.
The physical and chemical properties of mRNA-LNP vaccines that can be associated with theoretical risks for AEs.
| Unique vaccine properties | Sections* | Collateral immune effects | Potential AEs complications |
|---|---|---|---|
| (1) Ribosomal synthesis of the spike protein fundamentally transforms antigen processing and presentation | 6.1. |
|
|
| 2) Multiple chemical modification of the mRNA increases its stability and translation efficacy | 6.2.1-6.2.6 | ||
| 3) The spike protein can be toxic | 6.4.1.-6.4.4 |
|
|
| 4) The LNP is pharmacologically active immune stimulant and mRNA transfectant | 6.5.1.-6.5.4 |
|
|
| 5) PEG on the LNP surface is immune reactive and immunogenic | 6.6.1-6.6.2 |
|
|
| 6) The mRNA-LNP is unstable in water | 6.7. |
|
|
| 7) The spike protein is stabilized by enrichment with proline and G-quadruplexes | 6.8 |
|
|
| 8) The injectable vaccine may contain contaminations with plasmid DNA and inorganic elements or complexes | 6.9 |
|
|
*Sections, Section number where the content is discussed in lengths; SP, spike protein; CARPA, complement activation-related pseudoallergy, PEG, polyethylene glycol.
Table 2.
Systemic changes underlying the pan-toxicity of the SP.
| Systemic toxicities | References |
|---|---|
| Complement activation | [12,16] |
| Endothelial inflammation, microvascular damage | [15,16,121,128,129] |
| Oxidative (mitochondrial) damage | [21,22,130] |
| Cytokine release | [17,28] |
| (Micro)thrombosis | [14,19,20,129] |
| Red blood cell aggregation | [29] |
| White cell activation and/or cytotoxicity | [23,24,30] |
| Platelet activation and/or coagulation abnormalities | [18,25] |
| Fibrin thrombus formation | [131] |
| Blood-brain barrier damage | [13] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



