
Submitted:
30 January 2025
Posted:
31 January 2025
You are already at the latest version
Abstract
The aquatic macrophyte Brasenia schreberi J. F. Gmel., widely cultivated in China, faces challenges including damage from pests such as Galerucella birmanica Jacoby. This study investigated the population dynamics and spatial distribution of G. birmanica in a B. schreberi aquaculture system from May to November. G. birmanica occurred for several generations, the first population peaked in July, with abundant eggs, larvae, pupae and adults all being recorded. The beetles were nearly absent during the peak summer heat in August, but the population rebounded extraordinary in September, with eggs accounting for 85.4% of the whole population. The distribution of G. birmanica exhibited a conspicuous aggregation phenomenon, with severely chewed area having an average G. birmanica abundance 21.6 times higher than that of the non or lightly chewed area. Further study revealed that there are four special volatile compounds with discrepancy in amount released from intact and damaged leaves, cis-3-hexenyl acetate, 2-phenylethyl isothiocyanate, undecane and methyl salicylate, among these, 2-phenylethyl isothiocyanate was more abundant in damaged leaves and significantly attracted G. birmanica adults, indicating that the damaged B. schreberi leaves may sacrifice themselves by attracting G. birmanica to small regions, thus protect the whole plant population from pest disturbance.
Keywords:
Aquatic vegetable
; Brassica schreberi
; Galerucella birmanica
; herbivore-induced plant volatiles
; 2-phenylethyl isothiocyanate
; plant-insect interaction
1. Introduction
The relationships between insects and their hosts are complicated. The generalists, like Spodoptera exigua and Myzus persicae usually have a wide range of hosts for feeding and oviposition [1,2], while the specialists, like Aphis glycines and Brevicoryne brassicae select only one or several limited hosts [3,4]. For the later, humans may make use of such behavior to control the invasive species [5], or mitigate the pest damages on agricultural crops [6]. Insects usually locate their hosts through visual cues mediated by plant morphological characteristics [7], olfactory cues mediated by volatile organic compounds [8], and gustatory cues mediated by secondary metabolites [9]. Coevolution between insects and their hosts can contribute to the development of plant defense against herbivory [10]. Conversely, it can also facilitate the insects to find their hosts easier [11].
The watershield, Brasenia schreberi J. F. Gmel., is a monotypic genus in the Nymphaeaceae family (also listed in the Cabombaceae family in some references). The perennial, floating-leaved aquatic macrophyte widely but sporadically distributes in lakes and ponds across East Asia, Australia, West Indies, Africa and North and Central America [12,13], The submerged shoots and leaves, especially young buds of the plants are coated with thick mucilage containing multiple polysaccharides like D-galactose, D-glucose, L-rhamnose, L-fucose, etc., and are considered for both nutritional and medical values [14,15,16]. Hence young buds of the B. schreberi have been served as popular aquatic vegetable for a long history in China, Japan and some other eastern Asian countries, though only in China, B. schreberi are regularly farmed and harvested in shallow areas of ponds, lakes and irrigated fields with water depth maintained 30-80 cm [17,18].
The leaf-beetle Galerucella birmanica Jacoby (Coleoptera: Chrysomelidae) is a holometabolous insect with four stages: egg, larva, pupa and adult. It feeds on Trapa natans L. (Trapaceae) and B. schreberi, showing preference on T. natans between the two hosts [19,20]. The larvae experience a period of overeating before pupation, and adults consume even more [20,21,22]. Multiple generations of G. birmanica occur each year with overlapping generations. In central and southern China, 6-7 generations can occur annually [21,23]. G. birmanica overwinters as adult in weed residues and soil crevices on banks, starting activity in early April and moving to the overwintering sites in late October. Detailed relationships between G. birmanica and its host T. natans have been reported, as from late April to early May, when T. natans leaves emerge from the water, the beetles migrate to the leaves, feeding on the leaf tissue, entering a mating peak and laying eggs after 3-4 days [21]. Damaged T. natans leaves exhibit numerous holes, and in severe cases, only the veins remain [23]. In North America, G. birmanica has been considered as a potential biocontrol agent against the T. natans, which is a major invasive species in North America [20,22,24]. Earlier research by [22] demonstrated that G. birmanica oviposited and was able to complete development on both Trapa spp. and B. schreberi in no-choice experiments, and in field choice tests, strong preference for T. natans and only occasional “spill-over” onto B. schreberi were found. Recent report from [20] also suggested that potential risk of G. birmanica towards B. schreberi scored low.
While carefully designed behavioral experiments have been conducted in the lab, few studies have been focused on the “natural” relationship between G. birmanica and B. schreberi in wild fields. Especially in large-scale aquaculture systems where B. schreberi is the only crop cultivated and no T. natans coexists, will the beetle accept B. schreberi as a substitute, and to what extent will this occur? With our field investigation in Taihu Lake basin where B. schreberi has been widely cultivated, the answer is yes. In this paper, we try to record the monthly dynamics of G. birmanica in a human-maintained B. schreberi mono-cultivation system where no T. natans coexists. Additionally, analyzing the volatile compounds emitted from B. schreberi leaves, possible dynamics for the aggregation of G. birmanica in B. schreberi field has been explored.
2. Materials and Methods
2.1. Field Description
The research field is located in Changshu Ecological Agricultural District of Taihu Lake basin in east China (120°33′~121°03′ E, 31°33′~31°50′ N), where has the longest history of B. schreberi cultivation and remains one of the four major B. schreberi production areas of the country. Typical subtropical monsoon humid climate dominates in the area, with the annual average temperature being 15.4 ℃, from extreme minimum temperature of -11.3 ℃ to extreme maximum temperature of 40.2 ℃, and the annual average precipitation being 1135.6 mm. A human maintained B. schreberi cultivation pond in the center of the district is selected for survey of G. birmanica, with toxic pesticides being forbidden for years before the study and only limited organic fertilizers are used. The pond is irrigated by Wangyu River which connects the third largest freshwater lake, Taihu, and the longest river, Yangzte River, in China. The water depth in the pond is maintained at about 30 cm in early spring to help crop rejuvenation and about 60 cm in the rest seasons for the growth and overwintering. The total area of the pond is approximately 2400 m2.
2.2. Field Surveys on Monthly Occurrence of Galerucella birmanica in Brasenia schreberi Pond
The presence and density of G. birmanica in B. schreberi field were investigated by both direct inspection on leaf surface and yellow board sticky trapping (25 cm × 20 cm, double-sided adhesive), once a month covering the growth season of the crop from April to November in 2023. Three parallel transects with a distance no less than 10 m between one and another were set in the B. schreberi pond, along each transect, 12 sampling sites with 4 meters apart were marked, resulting in a total of 36 sites. At each site, the numbers of insects with different developmental stages within a 0.5 m × 0.5 m quadrat were first recorded by visual inspection on leaf surface. In detail, the numbers of eggs were recorded by every clutch, the numbers of larvae, pupae and adults were recorded by every individual, and the total numbers of beetles were countered as clutches of eggs plus individuals of the rests.
As a supplement for the flying adults of G. birmanica, a yellow sticky trap was placed in each sampling site for an additional log of the number of G. birmanica. The trap was positioned as close to the water surface as possible, and after 24 hours, the adults on the trap was counted.
Besides the beetles, the numbers of intact leaves and chewed leaves of B. schreberi were also counted, respectively, in each 0.5 m × 0.5m quadrat, thus leaf chewed rate was calculated as
Then seven leaves that exhibited the most severe chewed damage were selected to calculate the leaf damaged level.
The mean value of these calculations was used to represent the maximum extent of herbivory-induced damage at each sampling site.
2.3. Comparison of Galerucella birmanica Occurrence in Relatively Intact and Severely Chewed Areas
As visible aggregation of G. birmanica in the B. schreberi pond was notified, two additional groups of quadrats, one in areas with relatively intact B. schreberi leaves and the other in areas where the leaves were severely chewed, were randomly marked in the pond in November of the same year. In each group, 42 quadrats, were marked with an interval of at least 4 m, then the numbers of G. birmanica on leaf surface were counted. As the total amount of beetles were reduced in November, the quadrats area here were enlarged to 2 m x 2 m.
2.4. Two-Choice Tests of Adult Galerucella birmanica Towards Volatiles from Intact and Chewed Brasenia schreberi Leaves
To investigate the possible behavioral significance of adult G. birmanica in response to intact and chewed B. schreberi leaves, the volatiles from the two types of leaves were collected and extended to a two-choice test for the beetles.
For the collection of leaf volatiles, a headspace adsorption method was used[25,26]. Intact leaves (non-chewed) and severely chewed leaves of G. birmanica were harvested from field and took back to the lab. They were kept in moisture and wrapped in a sampling bag made of heat-resistant nylon resin. The inlet of the sampling bag was connected to an air pump with a glass scrubber filled with 20 / 50 mesh activated carbon adsorbent. The outlet of the sampling bag was connected to an adsorption column with 80 / 100 mesh Porapak Q (Supelco, USA). Teflon tubes were used for accessory passages. The volatile collection for each sample was maintained for 24 hours, with gas flow rate at 400 ml. min-1. Immediately after collection, the volatile sample on column was eluted with chromatographic grade hexane and the eluent was then concentrated to 200-300 μl with nitrogen. 200 μg of octane was added to each eluent as an internal standard. The collection for each type of leaves were repeated for five times, each sampling with 15 leaves.
The two-choice tests were conducted in a self-designed three-chamber olfactometer (Figure 1). One beetle was first placed in the middle chamber to adapt to the environment for 10 minutes. Then 10 µl of the eluent from intact / chewed leaves of B. schreberi was applied to a 2 cm2 filter paper, which was in sequence placed into the left / right chamber of the olfactometer. Clean air at a flow rate of 500 ml.min-1 was pumped into the chamber in each side of the olfactometer to make sure the beetle in the middle could perceive the odors from both sides. The temperature in the olfactometer was maintained at about 25°C during the tests. If the beetle remained in the selected chamber for more than 30 seconds, it was recorded as having made a choice for the volatiles in that chamber. If the beetle remained in the initial middle chamber for 15 minutes without making a choice, it was considered to have no response and excluded.
The two-choice tests were repeated until 26 adult beetles were recorded to have made choice. To eliminate possible position effect, every 5 beetles that had made choice were treated as one group, after each group, the positions of the filter papers with volatiles from intact / chewed leaves were swapped between the two sides of the olfactometer. The system was ventilated for 30 minutes each time when swapping to remove possible residue. An additional blank assay was also carried out before the test to determine whether there was any difference in the beetle’s preference when both sides of the olfactometer contained only clean air.
2.5. GC-MS Analysis for Volatiles from Intact and Chewed B. schreberi Leaves
Gas chromatography-mass spectrometry (GC-MS, Agilent 6890 GC and 5975C MS) was used to analyze the volatile components from intact and chewed B. schreberi leaves, referencing the conditions described by Zhang et al. [25,26]. Specifically, 1 μl of the eluent was injected into the inlet. The temperature was initially set at 40 ℃ for 5 minutes, increased to 185 ℃ at a rate of 5 ℃ min-1, further increased to 280 ℃ at a rate of 30 ℃ min-1,then remained for 1 minute. High purity nitrogen was used as the carrier gas at a flow rate of 20 ml min-1. The inlet temperature of GC-MS was set at 250 ℃. Volatile components were identified by comparison with NIST 2.0 library. Qualitative Analysis 10.0 was used to analyze the peak areas of various peaks in the GC-MS results. The amounts of special volatiles with remarkable differences between the intact and chewed leaf samples were calculated based on the ratio of the peak area comparing to that of the internal standard octane.
2.6. Verification of Volatile Substance that Attracting Galerucella birmanica
Four standard chemical solutions, cis-3-hexenyl acetate, 2-phenylethyl isothiocyanate, undecane and methyl salicylate in hexane of 1 µg·µl-1, representing each of the volatile that had exhibited remarkable discrepancy in amounts between intact and chewed leaves, were prepared to further verify the choice preference of G. birmanica. Similarly, 10 µl of standard solution or hexane (as control) on filter paper was placed in either side of the three-chamber olfactometer for selection of the beetle in the middle. Each standard solution was repeated until 26 beetles had made choice.
2.7. Statistical Analysis
Spearman correlation analysis was performed on the degree of leaf damage and the number of G. birmanica per hundred leaves. One-way analysis of variance (ANOVA) was used to compare the occurrence of G. birmanica between intact and severely chewed areas in B. schreberi pond. Chi-square test was used to compare the two-choice tests results of G. birmanica between volatiles of intact and chewed B. schreberi leaves and between regents representing separated volatiles.
3. Results
3.1. Monthly Occurrence of Galerucella birmanica in Brasenia schreberi Pond
Four typical developmental stages of G. birmanica, the egg, larva, pupa and adult, were found on the leaf surface of B. schreberi in the investigated pond (Figure 2). Their monthly occurrence on leaf surface was shown in Figure 3a. B. schreberi in the research site started to rejuvenate in April, while no G. birmanica was found at the first month when only sporadical leaves of the plants were floated on the water surface. In May, when leaves of the crop reached a coverage more than 80%, the beetles on leaves were first recorded, with relatively more larvae, followed by pupae and adults, but no eggs. The total number of beetles on leaves kept increasing, until in August, when the beetles almost disappeared in the hottest weather condition. In September, the total beetle number on leaves rebounded remarkably to be 3.4 times that in July, with 96.0% being eggs. The beetles reduced quickly in the following October, and kept with limited occurrence in November when the plants started to senesce. As for the different stages of beetles, besides the remarkable peak of eggs in September, a small peak of eggs also appeared in June. Relatively abundant larvae, pupae and adults appeared in July.
As an additional record for the emergence of G. birmanica in B. schreberi cultivation system, the numbers of G. birmanica trapped by yellow sticky boards showed a different trend compared to the direct record on leaf surface. Only adult G. birmanica were trapped on sticky boards, and significantly high amount of the beetles was recorded in August (Figure 3b). Small amounts of beetles were also trapped in June and November, but almost no beetles were trapped in May and July.
3.2. Leaf Damage Condition of Brasenia schreberi and Its Correlation with Galerucella birmanica Occurrence
Since no pesticide was applied in the cultivation system, intact elliptic leaves of B. schreberi could only been found in April when they first expanded on water surface, as subsequent leaves exhibited more or less chewed in the subsequent seasons. By careful calculation, 70.2% of the leaves were marked being chewed in May (Figure 4a). The leaf chewed rate generally increased, peaking at 91.9% in September. October was an exception, as only 25.9% of the leaved were chewed. The chewed rate rebounded over 70% in November.
Although most of the leaves showed signs of chewing damage, the averaged damaged level of each leaf was not so severe. As shown in Figure 4b, the leaf damaged level was lower than 2% in May, which increased to 3.7% in July, and then decreased to its lowest point as 0.6% in August. The highest leaf damaged level of the leaf occurred in September, as 6.3% of the leaf area was damaged. The data decreased to be less than 2% in October and November.
With Spearman Two-tailed tests, the damage on B. schreberi leaves was significantly correlated with the occurrence of G. birmanica. Positive correlations were found between leaf chewed rate, leaf damaged level and number of beetles on leaves, including eggs, larvae, pupae and total amount (p < 0.05, Table 1), only that no significant correlation was found between the number of adult beetles on leaf and leaf chewed rate (p > 0.05). The number of adult beetles trapped on board was found positively correlated with leaf chewed rate but negatively correlated with leaf damaged level (p < 0.05). As for an additional calculation, the number of beetles per hundred leaves on the leaf surface was significantly correlated with chewed damage (p < 0.05).
Typical correlation between leaf damaged level of B. schreberi and G. birmanica occurrence on leaf surface was also shown in Figure 5. The trends of beetle numbers on leaf surface through the growth season, both per m2 and per 100 leaves, closely mirrored the trend of leaf damaged levels. Visible but less remarkable correlation between leaf chewed rate and beetle numbers was also found.
3.3. Aggregation of Galerucella birmanica in Brasenia schreberi Pond
Field investigation also found that the occurrence of G. birmanica in B. schreberi pond was not uniform, but inclined to aggregate in certain area. An additional investigation in November showed that, the number of G. birmanica at each insect stage was significantly higher in areas where B. schreberi leaves were severely chewed compared to those in areas where the leaves were relatively intact (p < 0.05), which not only further proved the correlation between the number of beetles and leaf damage, but also indicates certain mechanism for the aggregation of G. birmanica (Figure 6).
3.4. Volatiles Differed Between Intact and Chewed Brasenia schreberi Leaves
GC-MS analysis was conducted to identify the components of volatile samples collected from intact and severely chewed leaves of B. schreberi. Four distinct substances that exhibited discrepancy in amount between the two types of leaves were identified: cis-3-hexenyl acetate, 2-phenylethyl isothiocyanate, methyl salicylate and undecane. Intact leaves emitted more methyl salicylate, while severely chewed leaves emitted more cis-3-hexenyl acetate, 2-phenylethyl isothiocyanate and undecane (Figure 7).
3.5. Two-Choice Test of Volatiles from Brasenia schreberi by Adult Galerucella birmanica
The results of the two-choice test of eluents of volatiles from intact and severely chewed leaves by adult G. birmanica showed a strong preference for volatiles from the latter (Figure 8). Out of 26 adults, 21 of them selected the chamber with volatiles from severely chewed leaves, showing a statistically significant difference between the two types of leaves, by Chi-Square test (p < 0.01).
For the four components that showing remarkable discrepancy in amounts between volatiles from intact and severely chewed leaves, adult G. birmanica showed different performance (Figure 8). Using solvent hexane as control, 13 out of the 26 tested adults chose the chamber containing cis-3-hexenyl acetate (p > 0.05), 18 out of the 26 tested adults chose the chamber containing 2-phenylethyl isothiocyanate (p < 0.01), 11 out of the 26 tested adults chose the chamber containing methyl salicylate (p > 0.05), and 13 out of the 26 tested adults chose the chamber containing undecane (p > 0.05).
4. Discussion
Environmental factors strongly influence the development and behavior of insects, thereby contribute to marked seasonal variations in their population dynamics. Temperature, in particular, plays a crucial role in the development and behavior of insects [27,28]. Detailed effect of temperature on the demography of G. birmanica has been reported by Zheng et al. [29]. In our investigation, the population density of G. birmanica in B. schreberi agricultural system exhibited significant seasonal variations. From May to July, the population of G. birmanica on leaf surface gradually increased with the temperature, the data decreased dramatically in August, rebounded to the peak in September, and then decreased again in October and November. Whereas relatively higher amount of flying adult G. birmanica were trapped by sticky board in August. Since the average temperature in our research site reached 29.2 ℃,and the daily highest temperature generally surpassed 30 ℃ in August in our research site, we suggested such high temperature might stimulate the mating and oviposition of G. birmanica but inhibit the egg development or impaired egg viability, leading to the disappearing of G. birmanica in August but quantitative eggs in September on leaf surface. Similar patterns have been observed in other insects, for instance, when the temperature reaches 33 ℃, the larval survival, pupation, and adult emergence rates of Diorhabda rybakowi all decrease significantly [30]. Additionally, low temperature can also inhibit the development of many insects [31]. For instance, the fecundity of Mythimna separata and Harmonia axyridis were reduced, and their preoviposition period were extended under low temperature [32,33]. Similarly, the feeding amount and fecundity of Liriomyza huidobrensis were decreased significantly under low temperature conditions [34]. The decrease of G. birmanica in October and November in our research site might be caused by lower temperature, afterwards, G. birmanica began its overwintering process.
Besides the temperature, rainfall might also affect the G. birmanica dynamics in B. schreberi system. The decrease of G. birmanica in August might be also explained by the rainfall. Our research site just experienced a rainy season prior to the survey of the month, heavy rain likely washed away many of the individuals previously presented on the leaves. Lethal effect of rainfall on G. birmanica has been also reported by [21].
Based on the field observations of G. birmanica population fluctuation and leaf damaged condition of B. schreberi, we inferred that he first generation of adults likely originated from overwintering beetles near the pond, with the first batch of eggs laid in late April. Over the whole growth season of B. schreberi, there should be 9 generations of G. birmanica occurred in the system, as follows: 1: late April—early June; 2: early June—early July; 3: early July—late July; 4: late July—early August; 5: early August—late August; 6: late August—early September; 7: early September—late September; 8: late September—mid October; 9: mid October—mid November
As for the discrepancy of G. birmanica peaks on leaf surface and sticky board, we suppose such phenomenon was caused by the mating activity of the beetles. High temperature may stimulate the female or male G. birmanica to release sex pheromones. The beetles, in search of the pheromone source, would fly upwind along the pheromone gradient [35]. Consequently, a significant number of eggs were founded in the next month.
The peak of leaf damage of B. schreberi both on chewed rate and damaged level appeared in September. The trends of leaf damage, especially the damaged level, through the whole growth season of B. schreberi, corresponded to the fluctuations in the population of G. birmanica. Previous research by Ding et al.[22] demonstrated that B. schreberi was only attacked when G. birmanica adults spilt over from T. natans. While our findings confirmed that in mono-culture of B. schreberi ecosystem, G. birmanica population was significantly correlated with leaf damage of B. schreberi. Evidently, in the absence of T. natans, G. birmanica poses a severe threat to B. schreberi, therefore careful consideration should be given before introducing G. birmanica as a biological control agent to areas where T. natans is considered as an invasive species.
Our observations showed that the distribution of G. birmanica in B. schreberi pond was not even, the beetles inclined to aggregate in areas where the plant leaves were severely damaged. Similar aggregation scenario of G. birmanica has been also reported by Zheng et al. [36] with T. natans system, and the authors suggested it may due to the biological characteristics of the beetles and the growth status of the plants. In our study, behavioral experiments and analysis of B. schreberi volatiles revealed that damaged leaves emitted significant amounts of 2-phenylethyl isothiocyanate, which exhibited a strong attractant effect on G. birmanica (p < 0.01). Therefore, we suppose that the aggregation of G. birmanica is influenced by the plant volatile promoted by chewing, specifically 2-phenylethyl isothiocyanate.
Herbivore-induced plant volatiles (HIPVs) play a pivotal role in plant-insect interaction. In nature, the diverse array of HIPVs contributes to the complexity of plant defense and signaling processes [37]. When plants are attacked by insects, they release a variety of HIPVs that can indirectly defend against herbivores by attracting their natural enemies or parasitoids [38,39,40], or facilitate plant-to-plant communication, thereby activating the defensive mechanisms of neighboring plants [41].
As a typical representative of HIPVs, isothiocyanates (ITCs) exhibit multiple ecological functions in plant-insect interactions. These secondary metabolites produced by plants are generated from glucosinolates through the action of their hydrolyzing enzyme, myrosinase, when plant tissues are damaged by insect pests or mechanical injury [42]. These compounds generally play a crucial role in protecting plants from various pests and microorganisms [43]. For general herbivores, glucosinolates and/or their ITCs products act as deterrents and toxins against them [44]. For specialist herbivores, such as Pieris rapae, ITCs, in a dose-dependent manner, impair the herbivore’s survival and growth, and increase their development time [45]. However, some specialist herbivores have also evolved corresponding adaptation mechanisms, for example, to develop strategies to prevent the formation of ITCs [46]. For some specialist herbivores, glucosinolates can even be sequestered for defense against predatory insects [47,48].
In certain exceptional cases, ITCs emitted from plants can attract herbivores. Earlier reports suggested that the herbivore Ceutorhynchus assimilis was attracted to ITCs in oilseed rape (Brassica napus L.) volatiles [49], and 3-butenyl, 4-pentenyl and 2-phenylethyl isothiocyanates might take function as attractants to pollen beetle Meligethes aeneus [50]. In our research, 2-phenylethyl isothiocyanate promoted in severely chewed B. schreberi leaves acted as an attractant for the G. birmanica, indicating that G. birmanica has evolved to overcome the defense of B. schreberi through ITCs. We suppose that G. birmanica may track the 2-phenylethyl isothiocyanate as a signal of food resource, mating partner or suitable habitat, thus aggregate in severely chewed area in the B. schreberi pond. Similar attractant mechanism of other HIPVs have been also found as the leafminer Tuta absoluta preferred infested tomato plants with up-regulated β-caryophyllene and tetradecane, supposed that infested plants might be more suitable for future larvae, or might result in a decreased likelihood of predation or parasitism [51].
Methyl salicylate is another type of HIPVs that is frequently detected in plant-insect interaction, synthesized through the salicylic acid pathway and activated when plants are under insect attack [52]. This volatile has been reported to trigger defense mechanisms both in the emitting plant and in neighboring plants [53,54]. Relatively higher level of methyl salicylate was also detected in severely chewed B. schreberi leaves, whereas no preference was found for G. birmanica in two-choice tests. Similar results were also found in the volatile cis-3-hexenyl acetate, which was also promoted in severely chewed leaves but showed no preference in two-choice tests. Such results for these two types of HIPVs in our research may be caused by specific gustatory perception of G. birmanica. Healthy leaves may also generate certain secondary metabolites that function as feeding deterrents to the beetles [55,56,57], meaning these compounds may cause the beetles to perceive bitter-tasting or pungency, thereby discouraging them from feeding [58], which we haven’t explored with B. schreberi yet. Undecane was found decreased in severely chewed leaves in our research, but still, with no preference for G. birmanica.
On the population or community level, herbivore damage to some leaves may promote defensive advantages to the entire plant even neighboring plants, with the latter being both conspecific and interspecific [59]. Our study indicates that through the emission of 2-phenylethyl isothiocyanate, severely chewed B. schreberi leaves may sacrifice themselves to mitigate the harm of G. birmanica to other intact leaves. Only that the neighboring defensive advantages are usually promoted by repelling signals that either resisting the herbivore directly [60] or attracting natural enemies [61], whereas in our study, the signal component 2-phenylethyl isothiocyanate is an attractor to the herbivore, make the herbivore-plant interaction more complicated. Since B. schreberi is a clonal species and its rhizome in silt can extend for several meters, we still lock the evidences to verify if such defensive advantage only happens within leaves of the same individual plant or also happens within neighboring plants. As G. birmanica has been reported to prefer T. natans to B. schreberi when co-exist [22], it will be also interesting to explore if 2-phenylethyl isothiocyanate exhibits further role in the interspecific neighboring effect between B. schreberi and T. natans.
Nevertheless, as only 4 discrepancy volatiles were detected between attacked and intact leaves of B. schreberi, there might be other physiological or signaling pathway to help the plants to defense feeding from G. birmanica. Also mixed compounds and their relative ratio may contribute to the differences on the repelling / attracting effects on herbivore of HIPVs [49,62,63]. Further research is still needed to elucidate the complex mechanisms of herbivore-plant interaction.
On the view of B. schreberi aquaculture, artificially synthesized regent 2-phenylethyl isothiocyanate may be considered as a potential attractant to control the burst of G. birmanica. HIPVs have been nominated in push-pull strategies to repel herbivores from crop plants while simultaneously attracting them to nearby trap plants [64]. Here we suppose that the artificially synthesized regent may play a similar role as natural HIPVs in aquaculture management. Compared with traditional pesticides that have been frequently used in B. schreberi aquaculture, like cypermethrin and permethrin, with notable toxicity that polluting the food as well as cultivation environment, 2-phenylethyl isothiocyanate, with low dose and high attracting efficiency, can be a relatively safer replacer. Our research with the population peaks of G. birmanica that burst alone the growth period of B. schreberi has indicated a proper time schedule for the use of pest controller, while further experiments are still expected for the detailed dose of 2-phenylethyl isothiocyanate that needed in open cultivation system.
5. Conclusions
Through our investigation, we identified the population dynamics of G. birmanica in B. schreberi aquaculture system throughout the entire growth season of the crops, setting up a temporal benchmark for the pest control. We observed that G. birmanica tends to aggregate in areas with severe leaf damage, indicating the beetles have evolved the ability to overcome the direct defense of B. schreberi through physiological metabolites. Also, driven by the herbivore-induced 2-phenylethyl isothiocyanate released from the damaged leaves, the attacked plants of B. schreberi sacrifice themselves to protect their intact healthy neighbors, exhibiting a group defense strategy adopted by the crops to preserve population stability and to reduce the cost of individual plant defense against pests.
Author Contributions
Conceptualization, Y.W. and C.Z.; methodology, Y.W. and C.Z.; software, Y.W.; validation, Y.W. and Y.W.; formal analysis, Y.W.; investigation,Y.W. and Y.W.; resources, C.Z.; data curation, Y.W.; writing—original draft preparation, Y.W.; writing—review and editing, Y.W., Y.W. and C.Z.; visualization, Y.W.; supervision, C.Z.; project administration, C.Z.; funding acquisition, C.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Jiangsu Agriculture Science and Technology Innovation Fund (JASTIF) (CX(20)2018), Fundamental Research Plan (Modern Agriculture) of Jiangsu Province (BE2015357), and Emerging Agricultural Education Research and Innovation program of Chinese Ministry of Education (2020265).
Data Availability Statement
The data presented in this study are available on request from the
corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Zhang, Z.; Song, X.; Hu, H.; Wang, D.; Chen, J.; Ma, Y.; Ma, X.; Ren, X.; Ma, Y. Knockdown of UDP-N-acetylglucosamine pyrophosphorylase and chitin synthase A increases the insecticidal efficiency of Lufenuron to Spodoptera exigua. Pestic Biochem Physiol 2022, 186, 105178. [Google Scholar] [CrossRef]
- Dong, Y.; Li, T.; Hou, Y.; Wilson, K.; Wang, X.; Su, C.; Li, Y.; Ren, G.; Xu, P. Densovirus infection facilitates plant-virus transmission by an aphid. New Phytol 2024, 243, 1539–1553. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Bi, Y.D.; Liu, M.; Li, W.; Liu, M.; Di, S.F.; Yang, S.; Fan, C.; Bai, L.; Lai, Y.C. Identification and expression profiles analysis of odorant-binding proteins in soybean aphid, Aphis glycines (Hemiptera: Aphididae). Insect Sci 2020, 27, 1019–1030. [Google Scholar] [CrossRef] [PubMed]
- Inayat, R.; Khurshid, A.; Boamah, S.; Zhang, S.; Xu, B. Mortality, Enzymatic Antioxidant Activity and Gene Expression of Cabbage Aphid (Brevicoryne brassicae L.) in Response to Trichoderma longibrachiatum T6. Front Physiol 2022, 13, 901115. [Google Scholar] [CrossRef] [PubMed]
- Blumenthal, D.M. Interactions between resource availability and enemy release in plant invasion. Ecol Lett 2006, 9, 887–895. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, X.; Wang, Z.; Chen, T.; Zhou, S.; Chen, J.; Pang, L.; Ye, X.; Shi, M.; Huang, J.; et al. Symbiotic bracovirus of a parasite manipulates host lipid metabolism via tachykinin signaling. PLoS Pathog 2021, 17, e1009365. [Google Scholar] [CrossRef]
- Pinero, J.C.; Souder, S.K.; Vargas, R.I. Vision-mediated exploitation of a novel host plant by a tephritid fruit fly. PLoS One 2017, 12, e0174636. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wei, J.; Yi, T.; Li, Y.Y.; Xu, T.; Chen, L.; Xu, H. A green leaf volatile, (Z)-3-hexenyl-acetate, mediates differential oviposition by Spodoptera frugiperda on maize and rice. BMC Biol 2023, 21, 140. [Google Scholar] [CrossRef]
- Zhang, Z.J.; Zhang, S.S.; Niu, B.L.; Ji, D.F.; Liu, X.J.; Li, M.W.; Bai, H.; Palli, S.R.; Wang, C.Z.; Tan, A.J. A determining factor for insect feeding preference in the silkworm, Bombyx mori. PLoS Biol 2019, 17, e3000162. [Google Scholar] [CrossRef]
- Karssemeijer, P.N.; de Kreek, K.A.; Gols, R.; Neequaye, M.; Reichelt, M.; Gershenzon, J.; van Loon, J.J.A.; Dicke, M. Specialist root herbivore modulates plant transcriptome and downregulates defensive secondary metabolites in a brassicaceous plant. New Phytol 2022, 235, 2378–2392. [Google Scholar] [CrossRef]
- Guo, M.; Du, L.; Chen, Q.; Feng, Y.; Zhang, J.; Zhang, X.; Tian, K.; Cao, S.; Huang, T.; Jacquin-Joly, E.; et al. Odorant Receptors for Detecting Flowering Plant Cues Are Functionally Conserved across Moths and Butterflies. Mol Biol Evol 2021, 38, 1413–1427. [Google Scholar] [CrossRef] [PubMed]
- Schneider, E.L.; Carlquist, S. Vessels in Brasenia (Cabombaceae): New perspectives on vessel origin in primary XYLEM OF ANGIOSPERMS. American Journal of Botany 1996, 83, 1236–1240. [Google Scholar] [CrossRef]
- Feng, S.; Luan, D.; Ning, K.; Shao, P.; Sun, P. Ultrafiltration isolation, hypoglycemic activity analysis and structural characterization of polysaccharides from Brasenia schreberi. Int J Biol Macromol 2019, 135, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Feng, S.; Yan, J.; Luan, D.; Sun, P.; Shao, P. Antidiabetic potential of polysaccharides from Brasenia schreberi regulating insulin signaling pathway and gut microbiota in type 2 diabetic mice. Curr Res Food Sci 2022, 5, 1465–1474. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Yu, X.; Liu, J.; Li, J.; Ai, T.; Yin, C.; Liu, H.; Qin, R. A special polysaccharide hydrogel coated on Brasenia schreberi: preventive effects against ulcerative colitis via modulation of gut microbiota. Food Funct 2023, 14, 3564–3575. [Google Scholar] [CrossRef]
- Wang, Y.; Zou, Y.; Fang, Q.; Feng, R.; Zhang, J.; Zhou, W.; Wei, Q. Polysaccharides from Brasenia schreberi with Great Antioxidant Ability and the Potential Application in Yogurt. Molecules 2023, 29, 150. [Google Scholar] [CrossRef]
- Xie, C.; Li, J.; Pan, F.; Fu, J.; Zhou, W.; Lu, S.; Li, P.; Zhou, C. Environmental factors influencing mucilage accumulation of the endangered Brasenia schreberi in China. Sci Rep 2018, 8, 17955. [Google Scholar] [CrossRef]
- Zhou, C.; Zhong, Z. Watershield (Brasenia schreberi J. F. Gmel.), from Popular Vegetable to Endangered Species. World Journal of Agriculture and Soil Science 2021, 6, 2020. [Google Scholar] [CrossRef]
- Ding, J.; Wang, Y.; Jin, X. Monitoring populations of Galerucella birmanica (Coleoptera: Chrysomelidae) on Brasenia schreberi and Trapa natans (Lythraceae): Implications for biological control. Biological Control 2007, 43, 71–77. [Google Scholar] [CrossRef]
- Simmons, W.; Blossey, B. Host plant phylogeny does not fully explain host choice and feeding preferences of Galerucella birmanica, a promising biological control herbivore of Trapa natans. Biological Control 2023, 180, 105201. [Google Scholar] [CrossRef]
- Lu, Z.; Zhu, J.; Zhu, S.; Chen, Z. Preliminary studies on the beetle (Galerucella Birmanica Jacoby) - an insect pest of waterchestnut and watershield. Scientia Agricultura Sinica 1984, 73–76. [Google Scholar]
- Ding, J.; Blossey, B.; Du, Y.; Zheng, F. Galerucella birmanica (Coleoptera: Chrysomelidae), a promising potential biological control agent of water chestnut, Trapa natans. Biological Control 2006, 36, 80–90. [Google Scholar] [CrossRef]
- Han, Q.; Huang, M.; Li, S.; Zeng, X.; Hu, G.; Tang, C. Occurrence and integrated control measures of Galerucella birmanica Jacoby. Journal of Anhui Agricultural Sciences 2015, 43, 80–82. [Google Scholar] [CrossRef]
- Pemberton, R.W. Natural enemies of Trapa spp. in Northeast Asia and Europe. Biological Control 1999, 14, 168–180. [Google Scholar] [CrossRef]
- Zhang, J.; Bisch-Knaden, S.; Fandino, R.A.; Yan, S.; Obiero, G.F.; Grosse-Wilde, E.; Hansson, B.S.; Knaden, M. The olfactory coreceptor IR8a governs larval feces-mediated competition avoidance in a hawkmoth. Proc Natl Acad Sci U S A 2019, 116, 21828–21833. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Komail Raza, S.A.; Wei, Z.; Keesey, I.W.; Parker, A.L.; Feistel, F.; Chen, J.; Cassau, S.; Fandino, R.A.; Grosse-Wilde, E.; et al. Competing beetles attract egg laying in a hawkmoth. Curr Biol 2022, 32, 861–869 e868. [Google Scholar] [CrossRef] [PubMed]
- Ekholm, A.; Faticov, M.; Tack, A.J.M.; Berger, J.; Stone, G.N.; Vesterinen, E.; Roslin, T. Community phenology of insects on oak: local differentiation along a climatic gradient. Ecosphere 2021, 12, e03785. [Google Scholar] [CrossRef]
- Owens, C.H.; Lee, M.J.; Grim, M.; Schroeder, J.; Young, H.S. Interactions between temperature and predation impact insect emergence in alpine lakes. Ecosphere 2023, 14, e4619. [Google Scholar] [CrossRef]
- Zheng, F.S.; Du, Y.Z.; Wang, Z.J.; Xu, J.J. Effect of temperature on the demography of Galerucella birmanica (Coleoptera: Chrysomelidae). Insect Science 2008, 15, 375–380. [Google Scholar] [CrossRef]
- Liu, N.; Gou, W.; Ma, W.; Tang, L.; Hu, G.; Sun, Y. Effects of temperature on the growth, development and reproduction of Diorhabda rybakowi (Coleoptera: Chrysomelidae). Plant Protection 2023, 49, 220–226. [Google Scholar] [CrossRef]
- Lacoume, S.; Bressac, C.; Chevrier, C. Sperm production and mating potential of males after a cold shock on pupae of the parasitoid wasp Dinarmus basalis (Hymenoptera: Pteromalidae). J Insect Physiol 2007, 53, 1008–1015. [Google Scholar] [CrossRef]
- Qin, J.; Zhang, L.; Cheng, Y.; Luo, L.; Lei, C.; Jiang, X. Age-stage two-sex life table for laboratory populations of oriental armyworm Mythimna separata (Walker) under different temperatures. Acta Phytophylacica Sinica 2017, 44, 729–736. [Google Scholar] [CrossRef]
- Zhao, J.; Chen, Z.; Qu, J.; Zhang, F.; Yin, X.; Xu, Y. Responses of Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae) adults to cold acclimation and the related changes of activities of several enzymes in their bodies. Acta Entomologica Sinica 2010, 53, 147–153. [Google Scholar] [CrossRef]
- Huang, L.H.; Chen, B.; Kang, L. Impact of mild temperature hardening on thermotolerance, fecundity, and Hsp gene expression in Liriomyza huidobrensis. J Insect Physiol 2007, 53, 1199–1205. [Google Scholar] [CrossRef]
- Rao, S.; Cossé, A.A.; Zilkowski, B.W.; Bartlelt, R.J. Aggregation pheromone of the cereal leaf beetle: Field evaluation and emission from males in the laboratory. Journal of Chemical Ecology 2003, 29, 2165–2175. [Google Scholar] [CrossRef] [PubMed]
- Zheng, F.; Zheng, Y.; Lu, Y.; Qiang, C.; Ding, J. Spatial distribution pattern of water chestnut beetle (Galerucella birmanica Jacoby). Chinese journal of eco-agriculture 2006, 14, 171–175. [Google Scholar]
- Turlings, T.C.J.; Erb, M. Tritrophic Interactions Mediated by Herbivore-Induced Plant Volatiles: Mechanisms, Ecological Relevance, and Application Potential. Annual Review of Entomology 2018, 63, 433–452. [Google Scholar] [CrossRef] [PubMed]
- Maselou, D.A.; Anastasaki, E.; Milonas, P.G. The Role of Host Plants, Alternative Food Resources and Herbivore Induced Volatiles in Choice Behavior of an Omnivorous Predator. Frontiers in Ecology and Evolution 2019, 6, 241. [Google Scholar] [CrossRef]
- Wang, B.; Dong, W.; Li, H.; D'Onofrio, C.; Bai, P.; Chen, R.; Yang, L.; Wu, J.; Wang, X.; Wang, B.; et al. Molecular basis of (E)-beta-farnesene-mediated aphid location in the predator Eupeodes corollae. Curr Biol 2022, 32, 951–962. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Xu, L.; Jiang, J.; Zhang, S.; Liu, T.; Xu, Y. Host Plant Species of Bemisia tabaci Affect Orientational Behavior of the Ladybeetle Serangium japonicum and Their Implication for the Biological Control Strategy of Whiteflies. Insects 2020, 11, 434. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Felton, G.W. Priming of antiherbivore defensive responses in plants. Insect Sci 2013, 20, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yan, X. The role of glucosinolates in plant-biotic environment interactions. Acta Ecologica Sinica 2007, 27, 2584–2593. [Google Scholar]
- Ahuja, I.; Rohloff, J.; Bones, A.M. Defence mechanisms of Brassicaceae: implications for plant-insect interactions and potential for integrated pest management. A review. Agronomy for Sustainable Development 2010, 30, 311–348. [Google Scholar] [CrossRef]
- Renwick, J.A.A. The chemical world of crucivores: lures, treats and traps. Entomologia Experimentalis et Applicata 2003, 104, 35–42. [Google Scholar] [CrossRef]
- Agrawal, A.A.; Kurashige, N.S. A role for isothiocyanates in plant resistance against the specialist herbivore Pieris rapae. Journal of Chemical Ecology 2003, 29, 1403–1415. [Google Scholar] [CrossRef]
- Wittstock, U.; Agerbirk, N.; Stauber, E.J.; Olsen, C.E.; Hippler, M.; Mitchell-Olds, T.; Gershenzon, J.; Vogel, H. Successful herbivore attack due to metabolic diversion of a plant chemical defense. Proceedings of the National Academy of Sciences of the United States of America 2004, 101, 4859–4864. [Google Scholar] [CrossRef] [PubMed]
- Müller, C.; Agerbirk, N.; Olsen, C.E.; Boevé, J.-L.; Schaffner, U.; Brakefield, P.M. Sequestration of host plant glucosinolates in the defensive hemolymph of the sawfly Athalia rosae. Journal of Chemical Ecology 2001, 27, 2505–2516. [Google Scholar] [CrossRef]
- Bruce, T.J.A. Glucosinolates in oilseed rape: secondary metabolites that influence interactions with herbivores and their natural enemies. Annals of Applied Biology 2014, 164, 348–353. [Google Scholar] [CrossRef]
- Bartlet, E.; Blight, M.M.; Hick, A.J.; Williams, I.H. The responses of the cabbage seed weevil (Ceutorhynchus assimilis) to the odor of oilseed rape (Brassica napus) and to some volatile isothiocyanates. Entomologia Experimentalis Et Applicata 1993, 68, 295–302. [Google Scholar] [CrossRef]
- Smart, L.E.; Blight, M.M. Response of the pollen beetle, Meligethes aeneus, to traps baited with volatiles from oilseed rape, Brassica napus. Journal of Chemical Ecology 2000, 26, 1051–1064. [Google Scholar] [CrossRef]
- Rodrigo, F.; Burgueno, A.P.; Gonzalez, A.; Rossini, C. Better Together: Volatile-Mediated Intraguild Effects on the Preference of Tuta absoluta and Trialeurodes vaporariorum for Tomato Plants. J Chem Ecol 2023, 49, 725–741. [Google Scholar] [CrossRef]
- Heil, M.; Ton, J. Long-distance signalling in plant defence. Trends Plant Sci 2008, 13, 264–272. [Google Scholar] [CrossRef]
- Gong, Q.; Wang, Y.; He, L.; Huang, F.; Zhang, D.; Wang, Y.; Wei, X.; Han, M.; Deng, H.; Luo, L.; et al. Molecular basis of methyl-salicylate-mediated plant airborne defence. Nature 2023, 622, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.; Du, L.; Liu, Q.; Hu, X.; Ye, W.; Turlings, T.C.J.; Li, Y. Stemborer-induced rice plant volatiles boost direct and indirect resistance in neighboring plants. New Phytol 2023, 237, 2375–2387. [Google Scholar] [CrossRef]
- Engelberth, J.; Alborn, H.T.; Schmelz, E.A.; Tumlinson, J.H. Airborne signals prime plants against insect herbivore attack. Proceedings of the National Academy of Sciences of the United States of America 2004, 101, 1781–1785. [Google Scholar] [CrossRef]
- Yang, K.; Gong, X.L.; Li, G.C.; Huang, L.Q.; Ning, C.; Wang, C.Z. A gustatory receptor tuned to the steroid plant hormone brassinolide in Plutella xylostella (Lepidoptera: Plutellidae). Elife 2020, 9, e64114. [Google Scholar] [CrossRef]
- Hao, X.; Wang, S.; Fu, Y.; Liu, Y.; Shen, H.; Jiang, L.; McLamore, E.S.; Shen, Y. The WRKY46-MYC2 module plays a critical role in E-2-hexenal-induced anti-herbivore responses by promoting flavonoid accumulation. Plant Commun 2024, 5, 100734. [Google Scholar] [CrossRef]
- Wari, D.; Aboshi, T.; Shinya, T.; Galis, I. Integrated view of plant metabolic defense with particular focus on chewing herbivores. J Integr Plant Biol 2022, 64, 449–475. [Google Scholar] [CrossRef]
- Holmes, K.D.; Agrawal, A.A. Induced resistance mitigates the effect of plant neighbors on susceptibility to herbivores. Ecosphere 2021, 12, e03334. [Google Scholar] [CrossRef]
- Foba, C.N.; Shi, J.H.; An, Q.Q.; Liu, L.; Hu, X.J.; Hegab, M.; Liu, H.; Zhao, P.M.; Wang, M.Q. Volatile-mediated tritrophic defense and priming in neighboring maize against Ostrinia furnacalis and Mythimna separata. Pest Manag Sci 2023, 79, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Choh, Y.; Ozawa, R.; Takabayashi, J. Do plants use airborne cues to recognize herbivores on their neighbours? Exp Appl Acarol 2013, 59, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Blassioli-Moraes, M.C.; Michereff, M.F.F.; Magalhães, D.M.; Morais, S.D.; Hassemer, M.J.; Laumann, R.A.; Meneghin, A.M.; Birkett, M.A.; Withall, D.M.; Medeiros, J.N.; et al. Influence of constitutive and induced volatiles from mature green coffee berries on the foraging behaviour of female coffee berry borers, Hypothenemus hampei (Ferrari) (Coleoptera: Curculionidae: Scolytinae). Arthropod-Plant Interactions 2018, 13, 349–358. [Google Scholar] [CrossRef]
- Liu, X.L.; Zhang, J.; Yan, Q.; Miao, C.L.; Han, W.K.; Hou, W.; Yang, K.; Hansson, B.S.; Peng, Y.C.; Guo, J.M.; et al. The Molecular Basis of Host Selection in a Crucifer-Specialized Moth. Curr Biol 2020, 30, 4476–4482. [Google Scholar] [CrossRef] [PubMed]
- Gurr, G.M.; Wratten, S.D.; Landis, D.A.; You, M. Habitat Management to Suppress Pest Populations: Progress and Prospects. Annu Rev Entomol 2017, 62, 91–109. [Google Scholar] [CrossRef]
Figure 1.
Three-box Olfactometer.
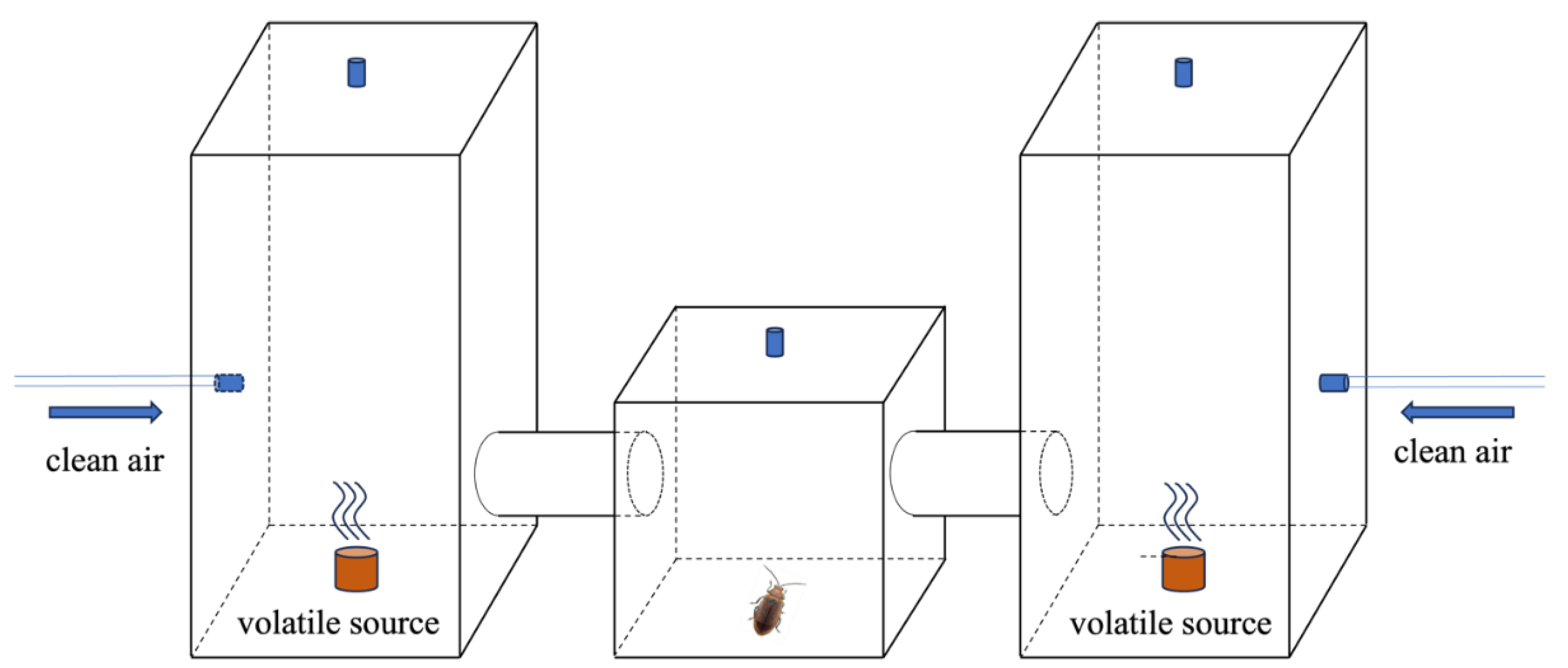
Figure 2.
Galerucella birmanica at different stages.

Figure 3.
Abundance of Galerucella birmanica in different months. Bars represent standard error of total beetles (a) or adult beetles (b), n=36. Different letters represent statistically significant difference at the level p < 0.05 of total beetles (a) or adult beetles (b).
Figure 3.
Abundance of Galerucella birmanica in different months. Bars represent standard error of total beetles (a) or adult beetles (b), n=36. Different letters represent statistically significant difference at the level p < 0.05 of total beetles (a) or adult beetles (b).
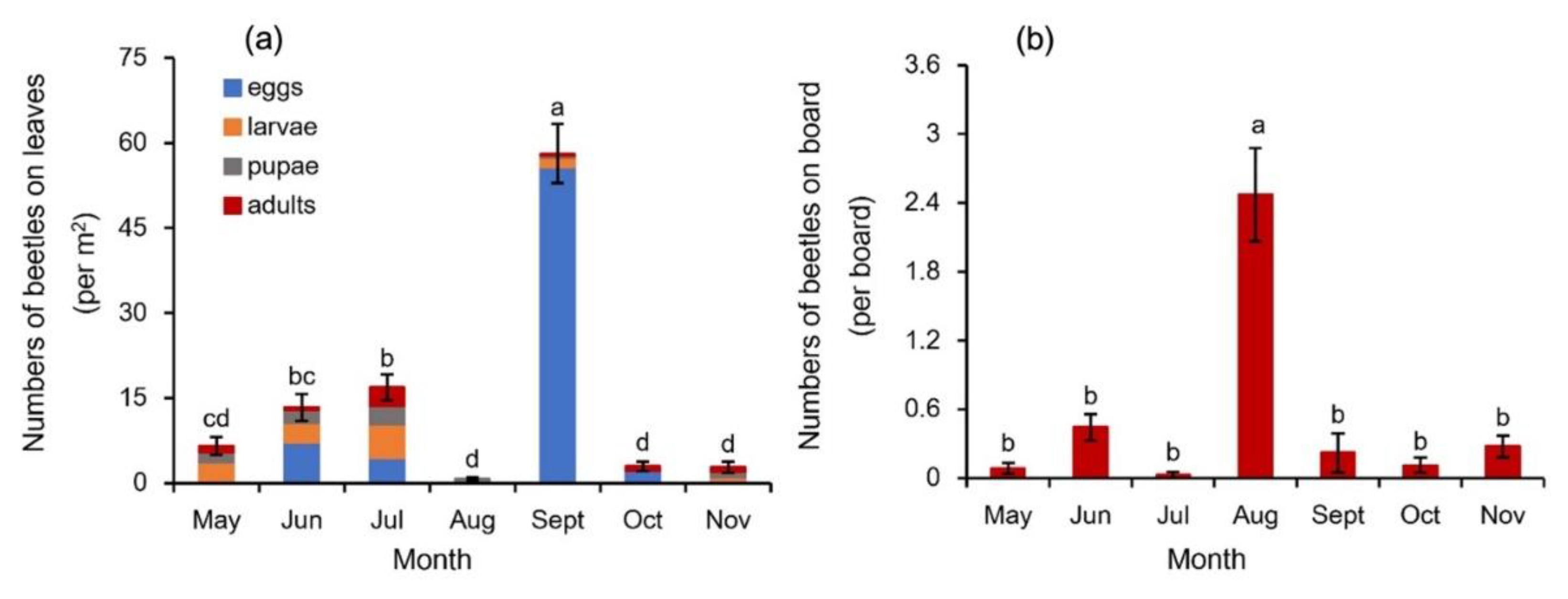
Figure 4.
Leaf damaged condition of Brasenia schreberi. (a) Leaf chewed rate of Brasenia schreberi; (b) Leaf dmaged level of Brasenia schreberi. Bars represent standard error, n=36. Different letters represent statistically significant difference at the level p < 0.05.
Figure 4.
Leaf damaged condition of Brasenia schreberi. (a) Leaf chewed rate of Brasenia schreberi; (b) Leaf dmaged level of Brasenia schreberi. Bars represent standard error, n=36. Different letters represent statistically significant difference at the level p < 0.05.
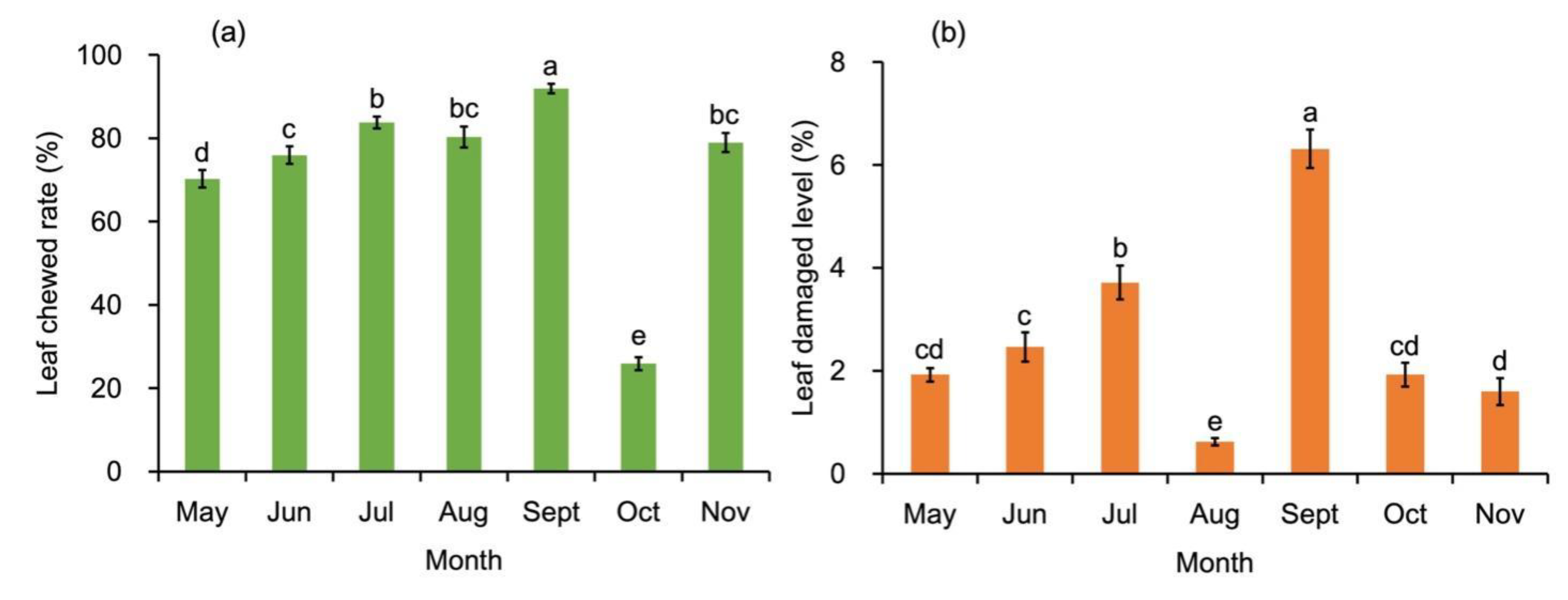
Figure 5.
Correlation between leaf damage and beetle density. Bars represent standard error, n=36.
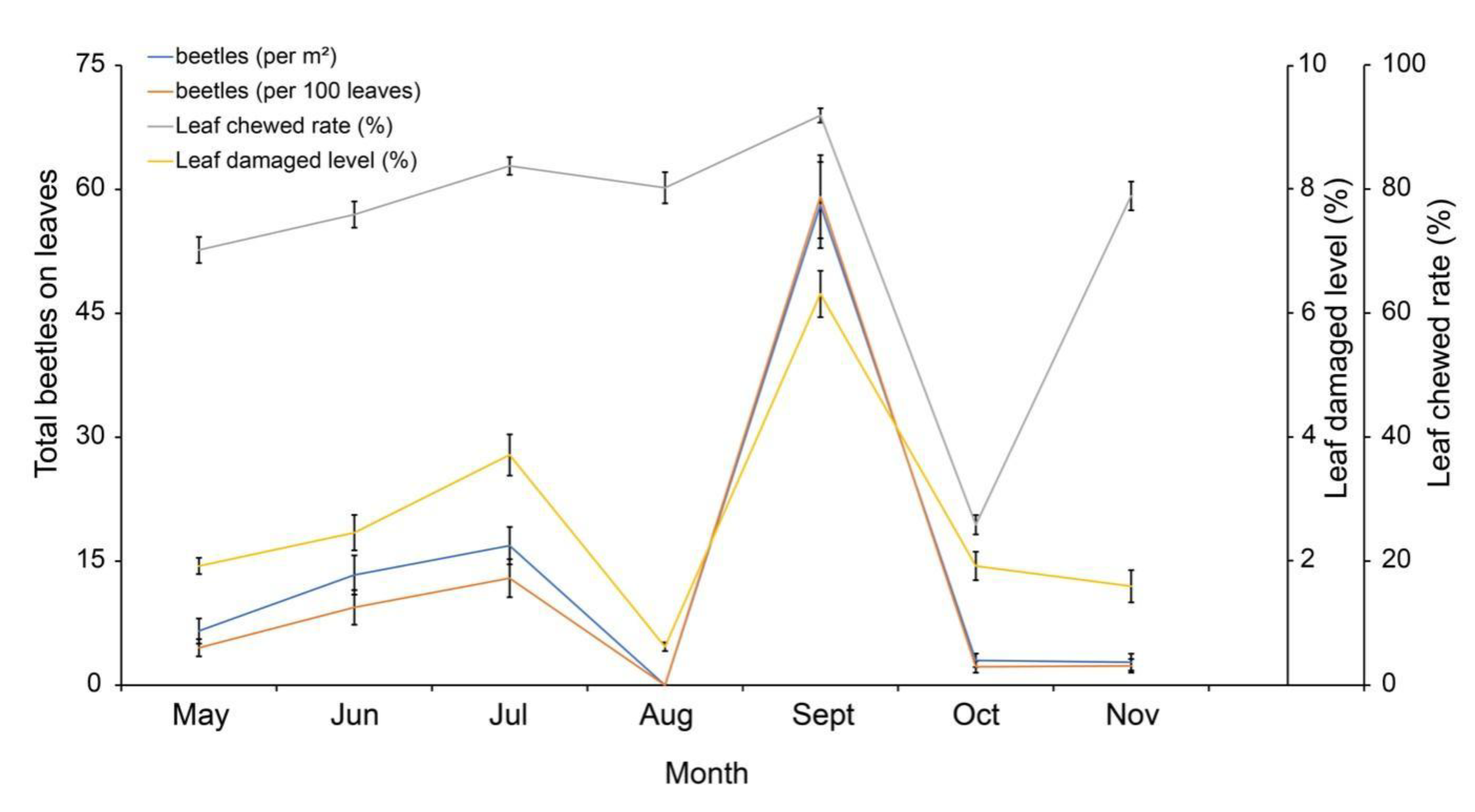
Figure 6.
Beetles in the relatively intact and severely chewed areas. Bars represent standard error, n=42. Different letters represent statistically significant difference between intact / chewed areas at the level p < 0.05.
Figure 6.
Beetles in the relatively intact and severely chewed areas. Bars represent standard error, n=42. Different letters represent statistically significant difference between intact / chewed areas at the level p < 0.05.
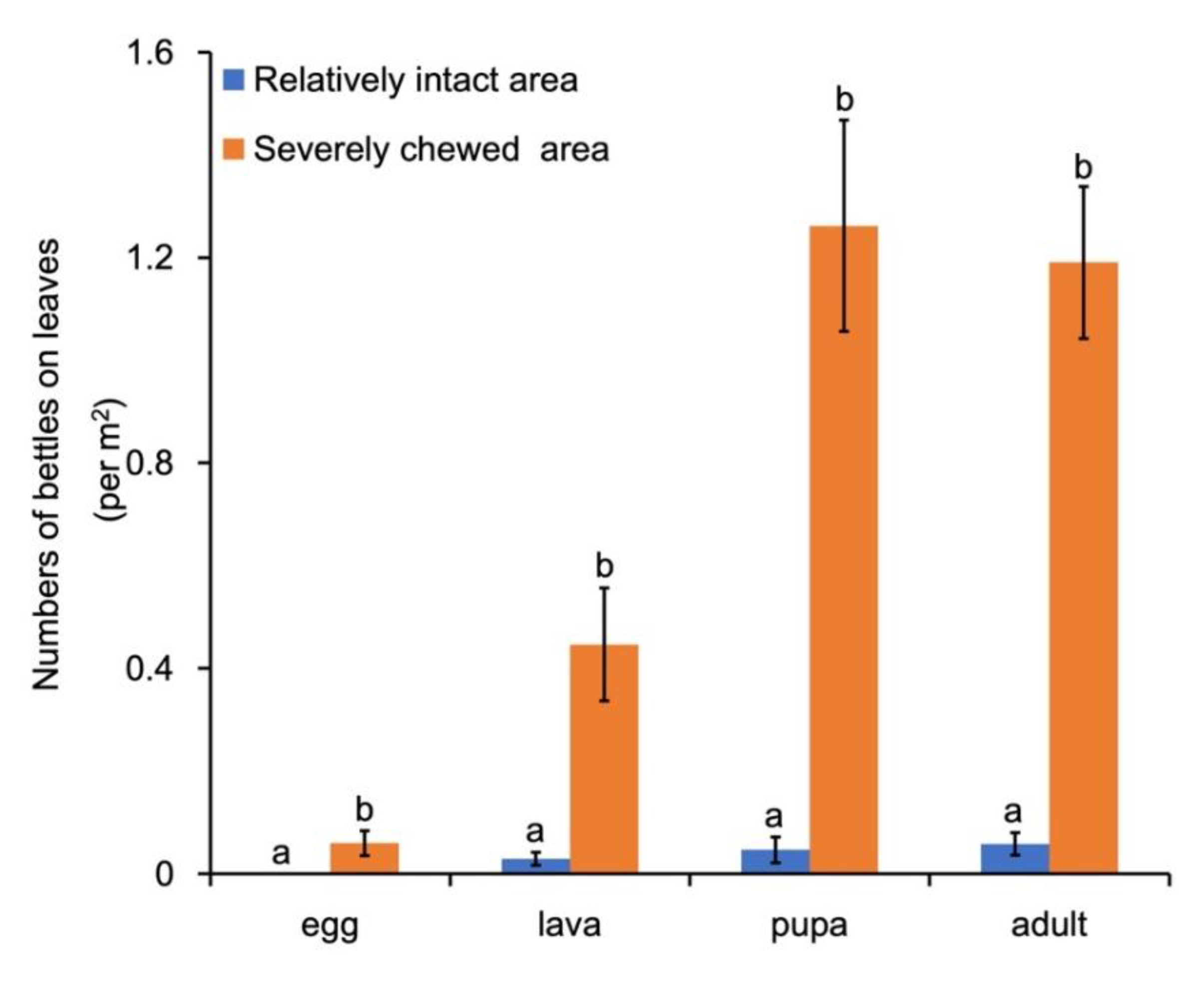
Figure 7.
GC-MS analysis of volatiles from intact and severely chewed leaves. 1, cis-3-Hexenyl acetate; 2, 2-phenylethyl isothiocyanate; 3, Undecane; 4, Methyl salicylate.
Figure 7.
GC-MS analysis of volatiles from intact and severely chewed leaves. 1, cis-3-Hexenyl acetate; 2, 2-phenylethyl isothiocyanate; 3, Undecane; 4, Methyl salicylate.
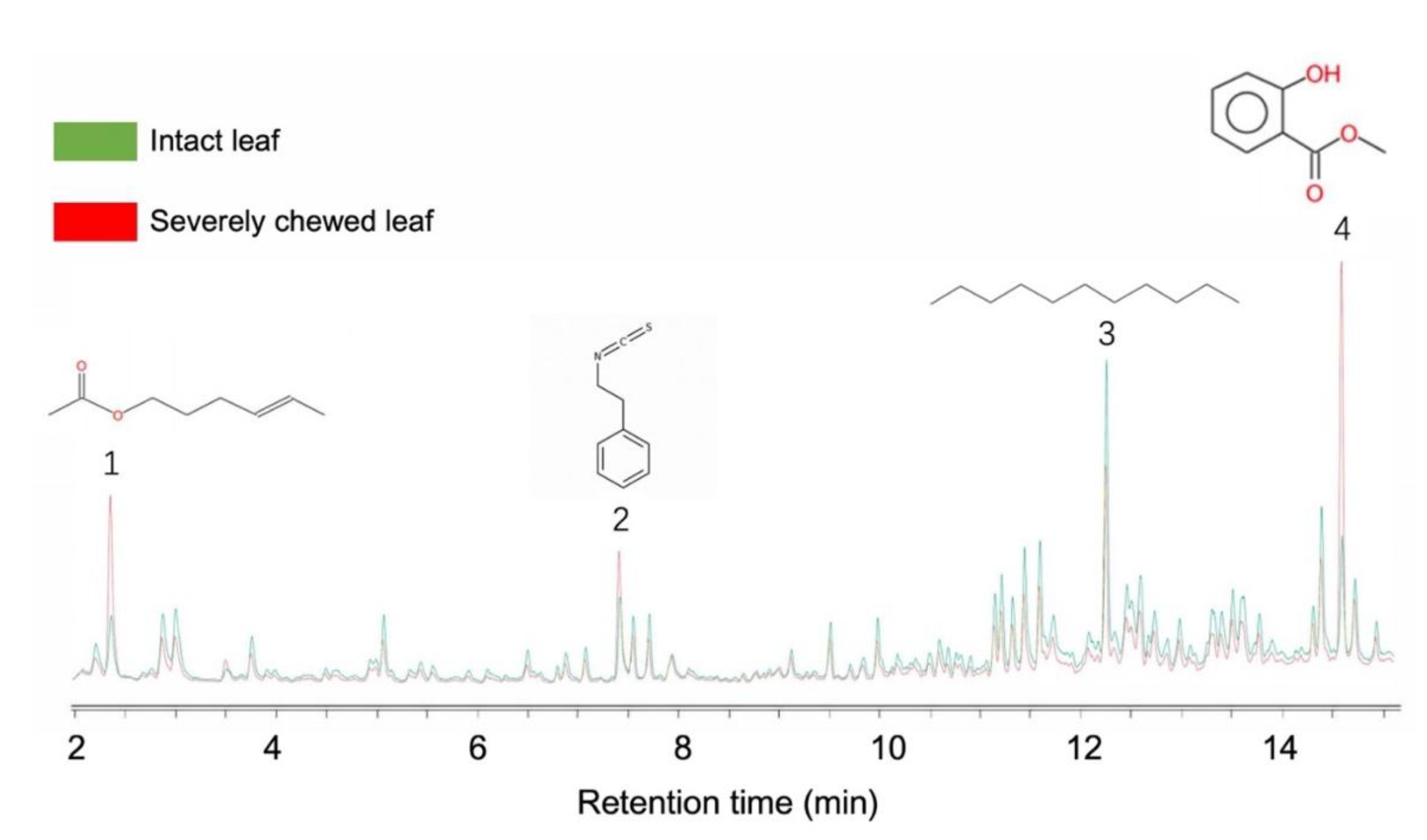
Figure 8.
Two-choice tests of volatiles from Brasenia schreberi by Galerucella birmanica adults. ** represent p < 0.01 by Chi-square tests, n=26.
Figure 8.
Two-choice tests of volatiles from Brasenia schreberi by Galerucella birmanica adults. ** represent p < 0.01 by Chi-square tests, n=26.
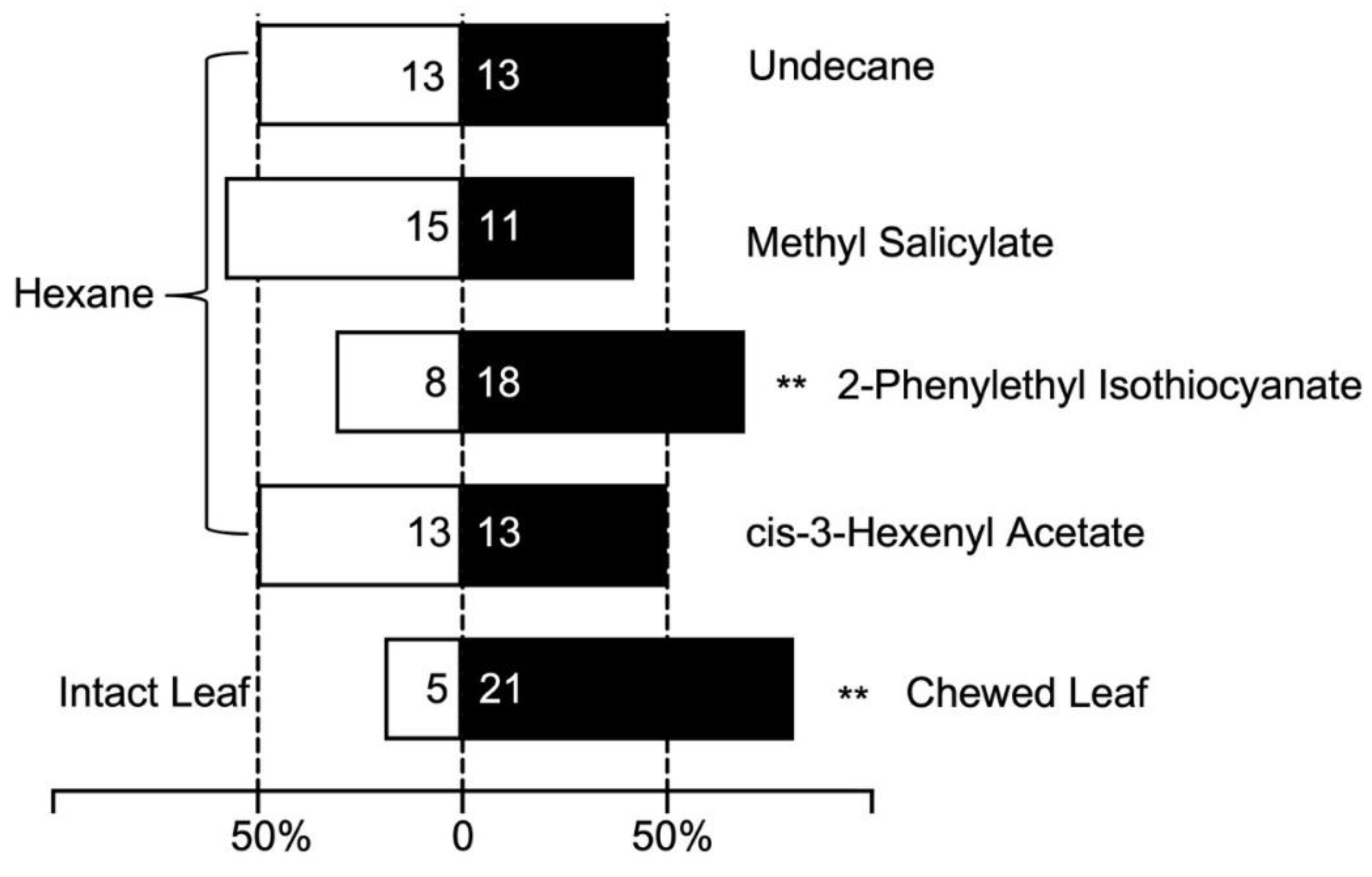

Table 1.
Correlation between Galerucella birmanica abundance and leaf damage of Brasenia schreberi.
| Beetles on leaves (per m2) | Beetles on leaves (per hundred leaves) | Beetles on board | |||||
|---|---|---|---|---|---|---|---|
| egg | larva | pupa | adult | total | |||
| Leaf chewed rate |
0.3846 (0.0000) |
0.2416 (0.0001) |
0.2264 (0.0003) |
0.1011 (0.1095) |
0.4533 (0.0000) |
0.4641 (0.000) |
0.1586 (0.0117) |
| Leaf damaged level |
0.6431 (0.0000) |
0.3919 (0.0000) |
0.3509 (0.0000) |
0.2691 (0.0000) |
0.7809 (0.0000) |
0.7783 (0.0000) |
-0.3034 (0.0000) |
Note: Data shown here are R (p) values, with boldface represents p < 0.05, analyzed by Spearman correlation analysis. Beetles on board only included adults, whereas those on leaves included all development stages.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



