
Submitted:
06 February 2025
Posted:
07 February 2025
You are already at the latest version
Abstract
Two species of moss, representative of the two categories of ecological strategies typical for bryophytes (i.e. stress tolerator and ruderal), were selected for the initial assessment of their capacity to withstand the conditions of the stratosphere. Samples of moss were attached to the balloons and elevated to the stratosphere during both diurnal and nocturnal periods. Observations of morphological changes, chloroplast condition, and chlorophyll autofluorescence post-introduction were conducted to assess survival abilities. The results demonstrated that Orthotrichum anomalum, a stress-tolerant moss, exhibited significantly higher resilience under both tested conditions compared to the ruderal Physcomitrella patens. Consequently, for further assessment of survival abilities of plants under space- or Mars-like conditions, desiccation tolerant stress-tolerant species are recommended.
Keywords:
mosses
; cellular stress
; stratosphere
; UV light exposure
1. Introduction
The survival of certain taxa in the hostile environment of space has attracted significant interest from researchers and the general public, and has been the subject of recent studies. A significant proportion of this research has been dedicated to the survival mechanisms of tardigrades, a group of ecdysozoans that appear to be highly resilient [1]. While the interest in and application of cryptobiotic mechanisms in animals is well-documented, research on the survival of autotrophic organisms (lichens) under low-orbit space conditions has also been conducted [2,3]. The rationale behind the selection of lichens for astrobiological surveys is partly attributable to their documented physiological resemblances with the aforementioned Tardigrades, particularly their capacity to withstand extreme environments, which is more specifically exemplified by their resilience to drought [4]. Furthermore, the term “anhydrobiosis”, employed by zoologists to denote the reversible metabolic state of organisms associated with their almost complete desiccation [5], is synonymous with the term “desiccation tolerance” utilised in the literature concerning lichens and other autotrophic organisms [6]. Anhydrobiosis/desiccation tolerance enables these organisms to survive drought, as well as other extreme conditions such as freezing, high temperatures or exposure to damaging irradiation, in their inactive, dried-out state. Lichens have been shown to withstand the conditions of space for periods of up to two weeks and one and a half years [2,7]. Nevertheless, despite their photoautotrophy, enabled by the autotrophic symbiont (alga or cyanobacterium), lichens are still species of various clades of the Fungi kingdom. Thus, the results of the study of their extremophilic characteristics cannot be fully applied to the physiology of plants. While there has been astrobiological research conducted on desert blue-green alga Chroococcidiopsis recently [8], no studies have yet been performed on the ability of land plants (Plantae sensu strictissimo) to sustain space and/or Martian conditions.
It is evident that plants in general exhibit desiccation tolerance traits. However, among the most widespread terrestrial plants, i.e., tracheophytes, this trait manifests predominantly in their propagules. However, only a small number of species, often referred to as resurrection plants, can be considered desiccation tolerant during their vegetative phase [9]. In contrast to the prevalent belief, a significant degree of vegetative desiccation tolerance is observed in bryophytes (a non-monophyletic group comprising liverworts, hornworts, and mosses), although this is not universal [10]. It has been hypothesised that the trait of vegetative desiccation tolerance was possessed by the common ancestors of all terrestrial plants, and that while bryophytes retained this trait, it was lost by most present-day plants during the course of evolution (with a small number re-evolving it). Bryophytes possess distinctive traits that were arguably characteristic of the earliest plants colonising dry land during the Ordovician period. However, if they are not considered “primitive”, they are considered to be products of the same selection pressure as the rest of life on Earth over the past 400 million years [10]. The scale of their dimensions and the habitats they occupy did not constitute the selection pressure for them to lose those characteristics. Furthermore, they are, in essence, occupying habitats that are quite similar to those present on Earth when the colonisation of the land began [11].
In contrast to the tracheophytic resurrection plants, which require a specific drying regime to survive desiccation, most bryophytes possess the capacity to withstand rapid drying, to the extent that recovery is not possible in tracheophytes [10]. Their capacity to withstand complete desiccation can be augmented by exposure to mild drought and subsequent rehydration, a process referred to as hardening [12]. Furthermore, the level of desiccation tolerance exhibited by particular moss species has been demonstrated to be associated with their tolerance to UV-B radiation. The most desiccation-tolerant species have been observed to be the least susceptible to UV-B damage, or to be completely undamaged [13]. Mosses are considered survival specialists in the ecological sense, as they are represented on the Stress Tolerator – Ruderal axis in the ecological strategy triangle proposed by Grime [14]. This characteristic confers upon certain species of bryophytes the capacity to withstand significant disturbances in their habitat, in addition to their ability to be among the first species to colonise newly exposed habitats. Their capacity to withstand prolonged periods of conditions that would be devastating to other plants is unparalleled. It has been demonstrated that they are capable of resuming metabolism when rehydrated after several years as herbarium specimens [15]. Furthermore, bryophytes have been observed to recover after being air-dried for up to 25 years [16], and there have been reports of regeneration even in mosses trapped under the Little Ice Age glacier for 400 years [17].
Due to their aforementioned ecological strategies and physiological characteristics, bryophytes are present and abundant in most of the Earth’s ecosystems. In some, specifically the harshest ones, they even establish a dominant group (e.g., tundra and Arctic/Antarctic rocks). However, the potential relevance of bryophytes in astrobiological studies has been largely overlooked. To date, the only experiment conducted with mosses in space focused on their growth patterns rather than their survival abilities [18]. In general, the effects of the space environment on plants have been reduced to the effects of microgravity on plant growth and/or gene expressions [19,20,21]. However, tracheophytes have been of the greatest concern.
2. Materials and Methods
2.1. Moss Material
In this study, the survival skills of two species, Orthotrichum anomalum Hedw. and Physcomitrella patens (Hedw.) Bruch and Schimp, respectively, were assessed following exposure to conditions in the stratosphere. These species represent the extremes of stress tolerance within the stress-tolerance-versus-ruderal axis of the aforementioned triangular ecological strategies scheme. Orthotrichum anomalum represents a group of bryophytes that are classified as stress-tolerant, while Physcomitrella patens represents the more ruderal, ephemeral species. O. anomalum has been demonstrated to exhibit desiccation tolerance [16]. Wang et al. [22] concluded that Physcomitrella patens is desiccation tolerant. However, it should be noted that their experiment was based not on rapid, but slow drying. Consequently, it can be deduced that, even if the species is indeed desiccation tolerant, its category of tolerance would not be the highest according to Wood [23]. This assertion is further substantiated by the utilisation of P. patens as a model organism in genetic studies, which has led to its comprehensive understanding of autecology and physiology. In contrast, O. anomalum is less well documented, yet it is pervasive and cosmopolitan, thus representing its ecological category. Dark-green fresh gametophytes of O. anomalum with shoots 1.5 cm tall, with straight leaves 2.5-4 mm long were collected in the Ostravian region (Czechia) in autumn season from its natural habitat on basic rocks. The P. patens strain was procured from the International Moss Stock Center, Freiburg, Germany (Reute-K1 Cat. No. 40040), and the sample consisted of young, 0.5cm tall gametophytes of the light-green moss with star-shaped rosettes of leaves. The plants were washed in distilled water, dried on a paper towel, and then placed separately inside 2 ml plastic eppendorf test vials (Sarstedt) 24 hours prior to the commencement of the experiment. The total number of test vials was six, with three vials allocated to each species. On the day of the experiment, two vials (one per species) of each species were kept at room temperature on the ground (control) and the remaining vials were attached to two balloons, one launched at daytime (12:00) and the other at night (20:00).
2.2. Stratospheric Missions
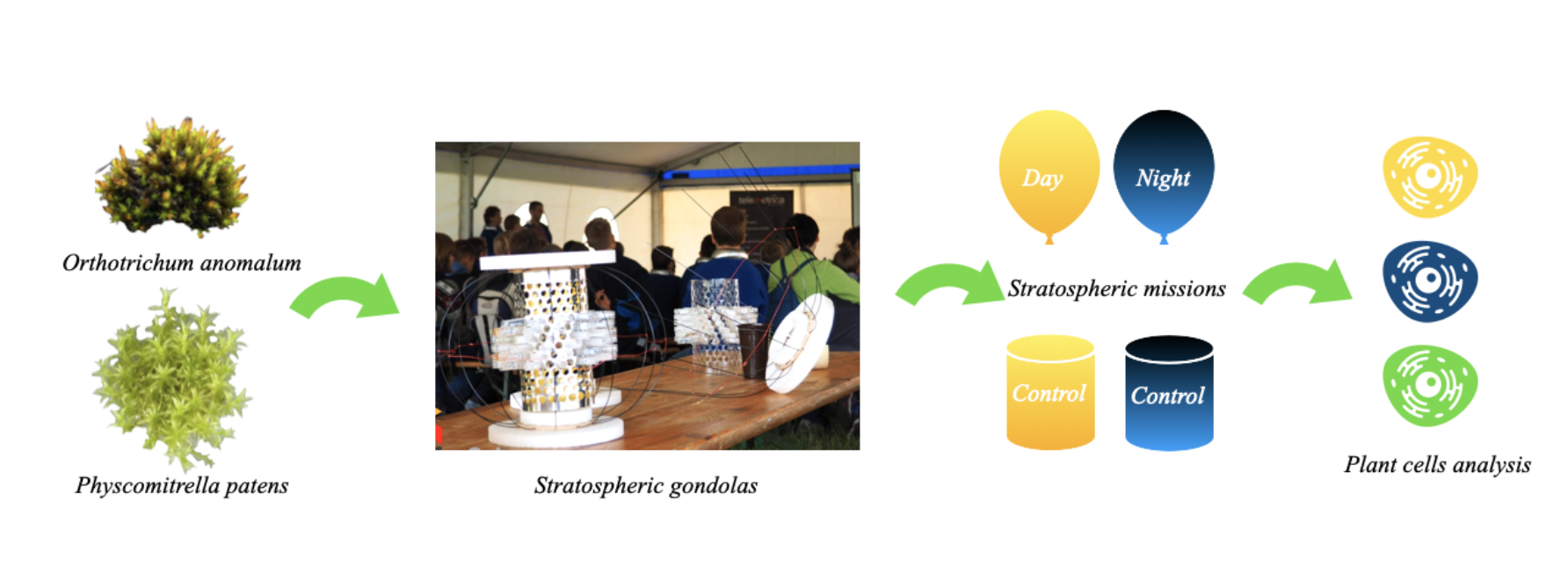
Samples of mosses were launched to the stratosphere in two independent launches on two independent capsules. These capsules were launched in November from the Aerogeodynamic Observatory in Borowiec, Poland (52°16′37.20″N 17°04′28.56″E). The primary compartment of each capsule comprised a cylindrical eppendorf holder, which was shielded from potential mechanical damage by light, flexible carbon rods. The 300g capsules were attached to balloons according to a diagram presented on Figure 1.
Telemetry and environmental parameters were obtained using a Vaisala radiosonde, GPS SpotGen4 and ESP-WROOM-32 controller programmed with the Arduino framework and interfaced with sensors via I2C. An accelerometer ADXL354 was utilised to monitor the effects of spinning and shock waves during the burst. The external conditions were monitored by a BME688 environmental sensor for temperature, humidity, and pressure. UV levels were measured using a GUVA-S12SD sensor (240-370 nm).
2.3. Imaging
The microscopic images were analysed and processed in Image J Software, version 1.44g [24] using a Wacom Bamboo tablet. Cell surface measurements were performed on three separate leaves in each moss sample to indicate levels of dehydration. Each measurement consisted of 30 randomly chosen cells taken from the middle part of the left side of the leaf. Subsequent measurements were conducted by the same researcher, utilising the same equipment and software, and repeated on several occasions over a period of several days. Autofluorescence of chlorophyll was measured simultaneously, with surface measurements expressed in levels of mean grey values.
2.4. Statistical Analysis
The analysis of the data was conducted using one-way and two-way analyses of variance in the R version 3.1.2.[25]. Subsequent to the identification of a significant difference by ANOVA, the Tukey honest significant difference test was applied.
3. Results
3.1. Environmental Parameters in the Stratosphere
Both balloons have successfully reached the stratosphere, where samples of mosses were subjected to changes in altitude ranging from 0 km to 30 km (33 km in the case of the nocturnal launch). Temperatures varied from 27°C (during the day) and 3°C (during the night) at ground level to -36°C and -60°C at the highest altitudes respectively. The duration of each mission was approximately one hour and 30 minutes of ascent (5 m/s-1), followed by approximately 30 minutes of descent. The capsules were retrieved approximately 15 min. after landing. The position of the balloons was tracked during the flight using common services: www.aprs.fi, www.sondehub.org and GPS online tracker www.findmespot.com. Following the return of the capsules to Earth, the moss samples were maintained under identical conditions and subsequently transported back to the laboratory for further analysis (a duration of 24 hours). The obtained data revealed changes in the intensity of UV radiation in dependence of altitude. The ground level of UV sensor activity was recorded as 54 units in the presence of sunlight and 2 units in shadow. In the stratosphere, a maximum UV sensor activity of 396 units was recorded in the Sun, and 189 units in shadow. Following an eight-minute flight period, the UV sensor activity in the Sun had already exceeded 250 units. Due to the landing location being in a cloudy area, a faster decrease in UV activity units was observed before landing (32 minutes before landing). The total duration of increased UV light exposure (exceeding 250 units in the Sun) was 1 hour and 49 minutes (Figure 2).
3.2. Cell Size and Morphology
In the aftermath of the day and night stratospheric missions, alterations in the cellular morphology of mosses were observed in both O. anomalum and P. patens (Figure 3). Leaves in the third row from the top of each plant were cut and photographed with an Axio Vision camera under the microscope. The detection of cellular damage and cell wall deformations was conducted under the illumination of a bright-field Zeiss microscope (20x lens). Changes in chloroplast morphology and chlorophyll autofluorescence were visualised using a 100x oil immersion objective lens under the Leica fluorescent microscope (exciting light 546 nm). In control samples of both species, leaves exhibited the most intense green colouration (Figure 3. OC and PC), indicating that no damage was sustained during the experiment’s fundamental procedure involving extended incubation in eppendorf vials prior to and following the stratospheric test. However, the night (Beta) mission exposed the plants to conditions that resulted in significant chloroplast degradation and cellular damage (Figure 3. OB and PB). However, a notable observation was the elongation of O. anomalum cells, which exhibited no signs of cellular damage. This is in contrast to the response of P. patens samples, which demonstrated cracking and degradation under similar environmental stress conditions. In the Alpha mission, chloroplasts and cells exhibited reduced levels of damage compared to those observed in the Beta mission. (Figure 3. OA and PA). Furthermore, in the P. patens control, chloroplasts were observed to be in close proximity to the cellular wall; however, following both treatments, they were relocated to the centres of the cells.
The measurement of cell surfaces revealed that both O. anomalum and P. patens control sample cells had significantly higher surface area than their counterparts subjected to the night mission (p = 0.0241901, p = 0.0081632, respectively). However, no significant differences were observed in the day mission measurements. The results of the cell surface measurement are presented in Figure 4.
3.3. Chlorophyll Autofluorescence
A significant disparity in chlorophyll autofluorescence intensity was detected between O. anomalum and P. patens, both in their initial state and subsequent to treatment (Figure 5).
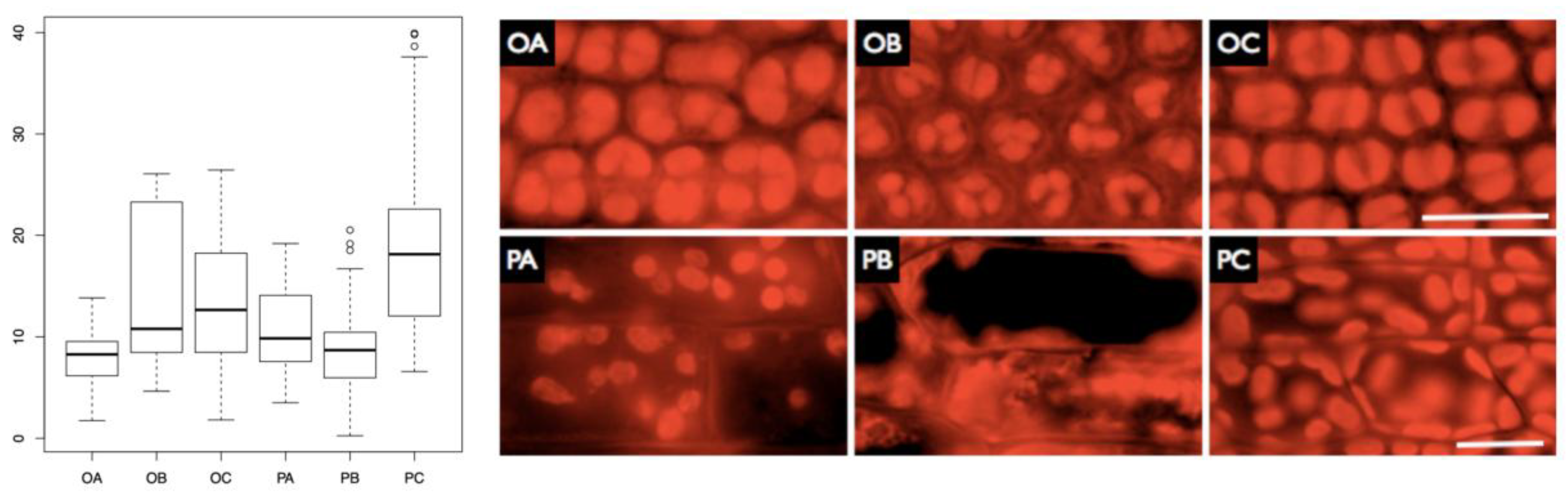
In P. patens, a significantly lower chlorophyll autofluorescence was observed in the samples subjected to both night and day missions (p < 0.001 in both cases compared to the control), with no significant difference between the samples from particular missions. Conversely, in O. anomalum, a significant decrease in autofluorescence was observed only in the sample subjected to the day mission (p < 0.001). Consequently, P. patens samples from both missions exhibited reduced autofluorescence compared to the O. anomalum control and the night mission sample, despite the initial state. This indicates that the decline in fluorescence after exposure to stratospheric conditions is more pronounced in P. patens. The outcomes of the autofluorescence analysis are consistent with the aforementioned results, indicating the most detrimental effect on both chloroplasts and cells in the night mission (Figure 5. OB, PB). In the case of Orthotrichum, it was observed that chloroplasts were swollen compared to the control in the Alpha mission (Figure 5. OC), whilst in the Beta mission, the size of chloroplasts was considerably smaller. In the case of Physcomitrella, smaller but still viable chloroplasts were observed in cells from the Alpha mission (Figure 5. PA), whereas in the night mission, the sample exhibited advanced cellular degradation (Figure 5. PB). Two-way ANOVA confirmed that there was a significant effect of treatment on autofluorescence (p < 0.001). The interaction of treatment and species factors was also found to be significant (p < 0.001), though the factor of species was not found to be significant (p = 0.0867).
4. Discussion
Exposure to conditions found in the stratosphere had a significant effect on the bryophyte samples, as evidenced by changes in cell size, overall morphology, and autofluorescence. The observed effects manifested in a pattern that was treatment- and species-dependent. In both the tested species, a significant decrease in cell size was observed in the samples subjected to the conditions of the Beta (night) mission. This phenomenon can be attributed primarily to the sudden temperature decline experienced by the samples. This effect was not observed in the Alpha mission samples, where the amplitude of temperature (and the lowest recorded temperature) did not reach such extreme levels. Such a decrease in cell size in plants has been extensively documented, with the process of osmotic shrinkage [26] being a well-documented phenomenon. This can be induced by freezing-related dehydration, as well as mechanical deformation of cells caused by the inevitable occurrence of extracellular ice [27]. This decrease in cell size is not necessarily detrimental, especially in desiccation-tolerant species, and can be reversible, depending on the extent to which the cell’s internal mechanisms are preserved. While O. anomalum cells under the Beta mission conditions exhibited no discernible damage or changes in morphology other than elongation, P. patens cells under identical conditions underwent rupture of the cell walls and complete disintegration of their chloroplasts. It is hypothesised that the deleterious effect would be less prominent if the moss were subjected to low temperatures gradually. It is known that P. patens can withstand low temperatures when the drop in temperature is not sudden [28]. Conversely, O. anomalum cells have been demonstrated to exhibit temperature resilience, maintaining the integrity of their chloroplasts during sudden temperature fluctuations.
P. patens cells exhibited the initial indications of degradation even when subjected to the conditions of the Alpha mission suggesting possible sensitivity to UV radiation. The chloroplasts underwent a relocation process within the cell. In the control cells, the chloroplasts were initially organised in alignment with the cell wall, whereas in the Alpha mission cells, they became scattered and moved towards the cell centres. This finding suggests that the P. patens cytoskeleton underwent degradation, while O. anomalum cells exhibited no discernible morphological alterations under the day stratospheric conditions.
The autofluorescence measurements revealed a significant decrease in chlorophyll fluorescence in all the samples except for the Beta mission O. anomalum sample, which showed no significant change. However, even in instances where a decline in autofluorescence was observed in both the tested species, the decline in P. patens cell fluorescence was considerably more pronounced. It has been established that chlorophyll fluorescence is directly related to the stress level in plants, and that it can be considerably lowered after exposure to elevated doses of UV B radiation [29]. This phenomenon can be attributed to the observed decrease in chlorophyll fluorescence in the Alpha mission samples of both species. However, the decline in chlorophyll fluorescence of the Beta mission P. patens cells is evidently associated with their overall compromised state and the deterioration of their chloroplasts.
As previously stated, O. anomalum cells exhibited no decline in fluorescence and were the only cells to demonstrate elongation. It has been demonstrated that exposure to conditions of stress and darkness can induce a process of rapid elongation in bryophytes [30]. This phenomenon is not necessarily indicative of biomass growth or proliferation; rather, it represents a strategy employed by bryophyte plants to cope with sudden darkness. Conversely, the absence of damage to bryophyte cells from UV-B radiation in the dark can be attributed to the protective effect of darkness, which prevents such damage. The observed absence of this protective effect in P. patens Beta mission cells can be interpreted as a result of irreparable damage and subsequent death of these cells.
Overall, the Orthotrichum anomalum samples exhibited a significantly higher degree of stress resilience in the presence of unfavourable stratospheric conditions in comparison to those of Physcomitrella patens. The most deleterious effects on the moss samples were observed to be the temperature in the Beta mission and UV radiation in the Alpha mission; both of these factors had significantly lower impact on the O. anomalum species samples. These findings lend further support to the distinctive ecological strategies of the studied species as outlined in the ecological strategy triangle scheme [14], and moreover, they underscore the potential of the Stress Tolerator category species to be of particular interest in future astrobiological bryophyte studies. It is therefore plausible that Stress Tolerator bryophytes, able to acclimate to extreme environments such as polar regions (either as species or individuals), would be particularly suitable for assessing bryophytes under simulated Cosmic or Mars-like conditions. In the following studies, it is recommended to focus on the chlorophyll fluorescence as a measurement of immediate stress of the plant together with its reversion to pre-treatment levels, and to add a measurement of reactive oxygen species production to assess the particular damage sustained quantitatively.
Acknowledgments
This project was supported by funding from the Jagiellonian University within the SET project (co-financed by EU), „Generation of the Future” - the program of Ministry of Science and Higher Education within a framework of operational program of Innovative Economy and the project New creative teams in priorities of scientific research, reg. no. CZ.1.07/2.3.00/30.0055, supported by Operational Programme Education for Competitiveness and co-financed by the European Social Fund and the state budget of the Czech Republic. Experimental work has been conducted within the “Pan Stanisław” educational project by www.copernicus-project.org. Authors thank prof. K. Turnau from the Institute of Environmental Sciences at the Jagiellonian University for consultation and kind help in obtaining data from fluorescent microscopy.
References
- Jönsson, K.I. , Rabbow E., Schill R.O., Harms- Ringdahl M. and Rettberg P. Tardigrades survive exposure to space in low Earth orbit. Current Biology 2008, 18, R729–R731. [Google Scholar] [CrossRef] [PubMed]
- Sancho, L.G. , de la Torre R., Horneck G., Ascaso C., de Los Rios A., Pintado, Wierchos J. and Schuster M. Lichens survive in space: results from the 2005 LICHENS experiment. Astrobiology 2007, 7, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Cottin, H. , Guan Y.Y., Noblet A., Poch O., Saiagh K., Cloix M., Macari F., Jérome M., Coll P., Raulin F., Stalport F., Szopa C. Bertrand M., Chabin A., Westall F., Chaput D. and Demets R. The PROCESS Experiment: An astrochemistry laboratory for solid and gaseous organic samples in low-Earth Orbit. Astrobiology 2012, 12, 412–425. [Google Scholar] [PubMed]
- Kranner, I. , Beckett R., Hochman A. and Nash T.H.III. Dessication-tolerance in lichens: A review. The Bryologist 2008, 111, 576–593. [Google Scholar] [CrossRef]
- Hengherr, S. , Heyer A. G., Köhler H. R. and Schill R. O. Trehalose and anhydrobiosis in tardigrades: evidence for divergence in response to dehydration. FEBS Journal 2008, 275, 281–288. [Google Scholar] [CrossRef]
- Alpert, P. The limits and frontiers of desiccation tolerant life. Integrative and Comparative Biology 2005, 45, 685–695. [Google Scholar] [CrossRef]
- Onofri S, de la Torre R, de Vera J. P., Ott S, Zucconi L, Selbmann L, Scalzi G, Vankateswaran K.J., Rabbow E, Sánchez Iñigo F.J. and Horneck G Survival of rock-colonizing organisms after 1.5 years in outer space. Astrobiology 2012, 12, 508–516. [Google Scholar] [CrossRef]
- Baqué, M. , Scalzi G., Rabbow E., Rettberg P. and Billi D. Biofilm and planktonic lifestyles differently support the resistance of the desert Cyanobacterium Choococcidiopsis under space and Martian simulations. Origins of Life and Evolution of the Biosphere 2013, 43, 377–389. [Google Scholar] [CrossRef]
- Farrant, J.M. (2007). Mechanisms of desiccation tolerance in angiosperm resurrection plants. In Plant desiccation Tolerance, edited by Jenks, M.A. and Wood, A.J. Blackwell Publishing. pp. 51–90.
- Proctor, M.C.F. , Oliver M.J., Wood A.J., Alpert P., Stark L.R., Cleavitt N.L. and Mishler B.D. Dessication-tolerance in bryophytes: a review. The Bryologist 2007, 110, 595–621. [Google Scholar] [CrossRef]
- Oliver, M.J. , Velten J. and Mishler B.D. Desiccation tolerance in bryophytes: A reflection of the primitive strategy for plant survival in dehydrating habitats? Integrative and Comparative Biology 2005, 45, 788–799. [Google Scholar]
- Proctor, M.C.F. and Pence, V. C. (2002). Vegetative Tissues: Bryophytes, vascular resurrection plants, and vegetative propagules. In Desiccation and survival in plants: Drying without dying, edited by Black, M. and Pritchard, H.W., CABI Publishing,, pp. 207–237.
- Turnbull J., D. , Leslie S. J. and Robinson S. A. Desiccation protects Antarctic mosses from ultraviolet-B induced DNA damage. Functional Plant Biology 2009, 36, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Grime, J.P. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. The American Naturalist 1977, 982, 1169–1194. [Google Scholar] [CrossRef]
- Crum H. (1983). Mosses of the Great Lakes Forest. University of Michigan Press, Ann Arbor, MI, 417 pp.
- P. Alpert and M.J. Oliver. (2002). Drying without dying. In Desiccation and Survival in Plants: Drying without Dying, edited by M. Black and H. W. Pritchard CABI Publishing., pp. 1–43.
- La Farge, C. , Williams K.H. and England J.H. Regeneration of Little Ice Age bryophytes emerging from a polar glacier with implications of totipotency in extreme environmemts. Proceedings of the National Academy of Sciences 2013, 110, 9839–9844. [Google Scholar] [CrossRef] [PubMed]
- Kern, V.D. and Sack F.D., Irradiance- dependent regulation of gravitropism by red light in protonemata of the moss Ceratodon purpureus. Planta 1999, 209, 299–307. [Google Scholar] [CrossRef]
- Paul, A.-L. , Zupanska, A.K., Ostrow, D.T., Zhang, Y., Sun, Y., Li, J.L., Shanker, S., Farmerie, W.G., Amalfitano, C.E. and Ferl, R.J. Spaceflight transcriptomes: unique responses to a novel environment. Astrobiology 2012, 12, 40–56. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, M. , Oono Y., Gusev O., Matsumoto T., Yazawa T., Levinskikh M.A., Sychev V.N., Bingham G.E., Wheeler R. and Hummerick M. Genome-wide expression analysis of reactive oxygen species gene network in Mizuna plants grown in long-term spaceflight. BMC Plant Biology 2014, 14, 4. [Google Scholar] [CrossRef]
- Ferl, R.J. , Koh J., Denison F. and Paul A.-L. Spaceflight induces specific alterations in the proteomes of Arabidopsis. Astrobiology 2015, 15, 32–56. [Google Scholar] [CrossRef]
- Wang, X.Q. , Yang P.F., Liu Z., Liu W.Z., Hu Y., Chen H., Kuang T.Y., Pei Z.M., Shen S.H. and He Y.K. Exploring the mechanism of Physcomitrella patens desiccation tolerance through a proteomic strategy. Plant Physiology 2009, 149, 1739–1750. [Google Scholar] [CrossRef]
- Wood, A.J. The nature and distribution of vegetative desiccation-tolerance in hornworts, liverworts and mosses. The Bryologist 2007, 110, 163–177. [Google Scholar] [CrossRef]
- Schulze, K. , López D.A., Tillich U.M. and Frohme M. A simple viability analysis for unicellular cyanobacteria using a new autofluorescence assay, automated microscopy, and ImageJ. BMC Technology 2011, 11, 118. [Google Scholar]
- R Core Team (2014). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: http://www.R-project.org/.
- Towill, L.E. and Mazur P. Osmotic shrinkage as a factor in freezing injury in plant tissue cultures. Plant physiology 1976, 57, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, M. Ide H., Price W.S., Arata Y., Nakamura T. and Kishimoto T. (2009). Freezing behaviours in plant tissues: visualization using NMR micro-imaging and biochemical regulatory factors involved. In Plant Cold Hardiness: From the Laboratory to the Field, edited by Gusta, L., Wisniewski, M. and Tanino, K., CAB International., pp. 19–28.
- Minami, A. Nagao M., Ikegami K., Koshiba T., Arakawa K., Fujikawa S. and Takezawa D. Cold acclimation in bryophytes: low-temperature- induced freezing tolerance in Physcomitrella patens is associated with increases in expression levels of stress-related genes but not with increase in level of endogenous abscisic acid. Planta 2005, 220, 414–423. [Google Scholar] [PubMed]
- Hui, R. , Li X., Zhao R., Liu L., Gao Y. and Wei Y. UV-B radiation suppresses chlorophyll fluorescence, photosynthetic pigment and antioxidant systems of two key species in soil crusts from the Tengger Desert, China. Journal of Arid Environments 2015, 113, 6–15. [Google Scholar] [CrossRef]
- Bakken, S. Effects of nitrogen supply and irradiance on growth and nitrogen status in the moss Dicranum majus from differently polluted areas. Journal of Bryology 1995, 18, 707–721. [Google Scholar] [CrossRef]
Figure 1.
The following diagram illustrates the high-altitude balloon assembly for two flights of the stratospheric mission. During the nocturnal flight, the video capsule was detached, thereby reducing the payload mass and enabling the attainment of greater altitude. The astrobiological capsule has been designed with the objective of being ultralight, with the capacity to expose more than 200 biological samples in 2ml testing eppendorfs.
Figure 1.
The following diagram illustrates the high-altitude balloon assembly for two flights of the stratospheric mission. During the nocturnal flight, the video capsule was detached, thereby reducing the payload mass and enabling the attainment of greater altitude. The astrobiological capsule has been designed with the objective of being ultralight, with the capacity to expose more than 200 biological samples in 2ml testing eppendorfs.

Figure 2.
Data logs from the GUVA-S12SD 240 nm-370 nm sensor revealing oscillations in UV light intensity values indicating rotation movement of the biological capsule. Y axis shows activity sensor units in relation to the 2.5h time of flight duration (X axis).
Figure 2.
Data logs from the GUVA-S12SD 240 nm-370 nm sensor revealing oscillations in UV light intensity values indicating rotation movement of the biological capsule. Y axis shows activity sensor units in relation to the 2.5h time of flight duration (X axis).
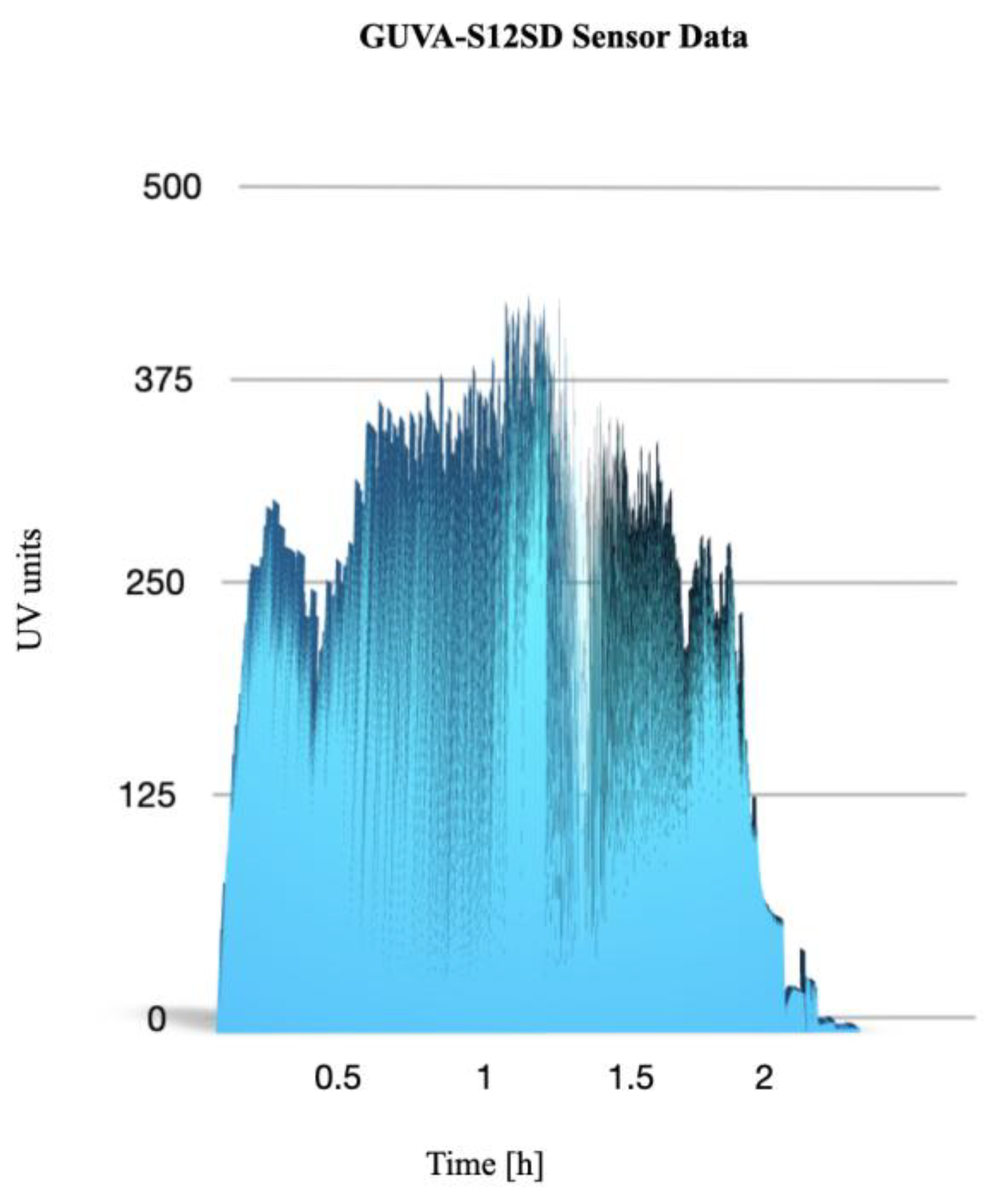
Figure 3.
Microscope images of mosses Orthotrichum anomalum (O) and Physcomitrella patens (P) in day A (Alpha), B night (Beta) and C (Control) conditions. Scale: 25 micrometers.
Figure 3.
Microscope images of mosses Orthotrichum anomalum (O) and Physcomitrella patens (P) in day A (Alpha), B night (Beta) and C (Control) conditions. Scale: 25 micrometers.
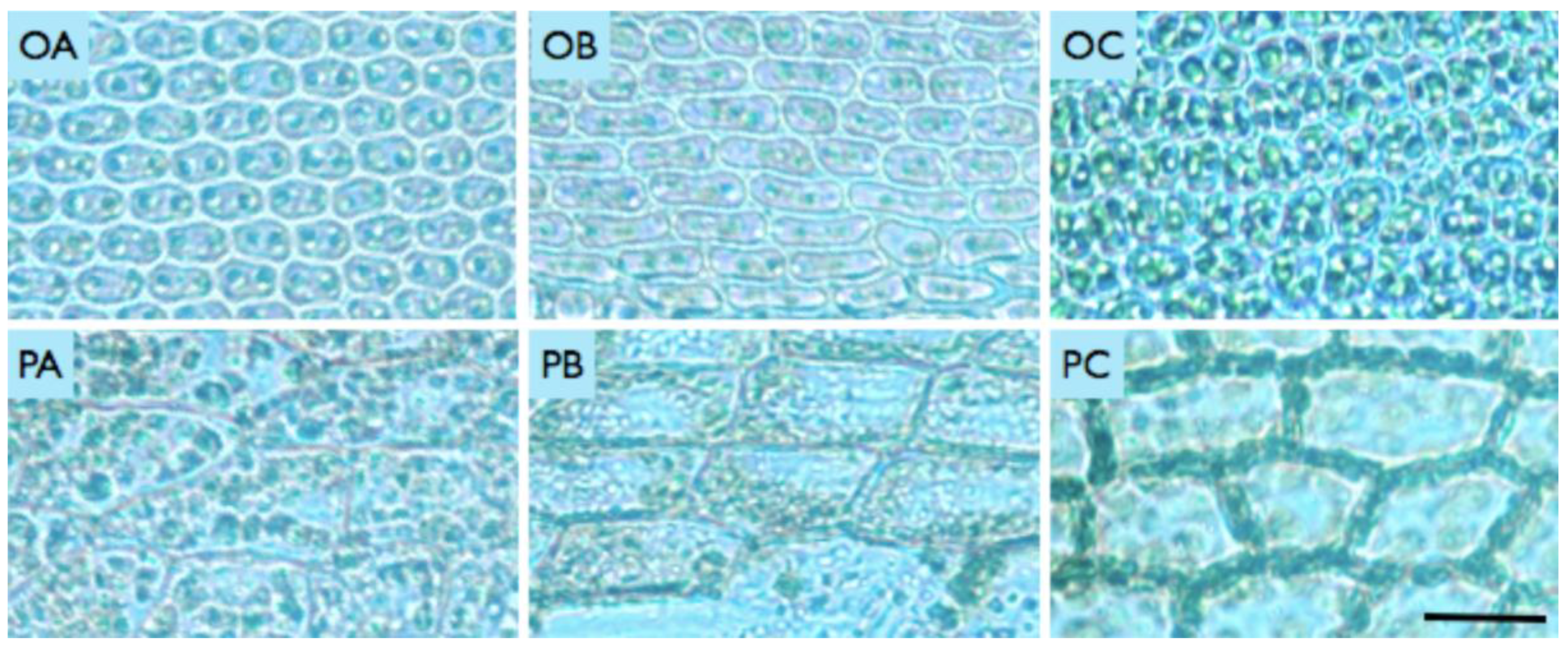
Figure 4.
The following measurements were taken of the Orthotrichum anomalum cell surface: A, B, C represent the results from the day mission, night mission and control group, respectively. The vertical axis indicates the number of square pixels. The following measurements are presented for Physcomitrella patens. A, B, C – day mission, night mission, control (respectively); vertical axes denote thousands of square pixels.
Figure 4.
The following measurements were taken of the Orthotrichum anomalum cell surface: A, B, C represent the results from the day mission, night mission and control group, respectively. The vertical axis indicates the number of square pixels. The following measurements are presented for Physcomitrella patens. A, B, C – day mission, night mission, control (respectively); vertical axes denote thousands of square pixels.
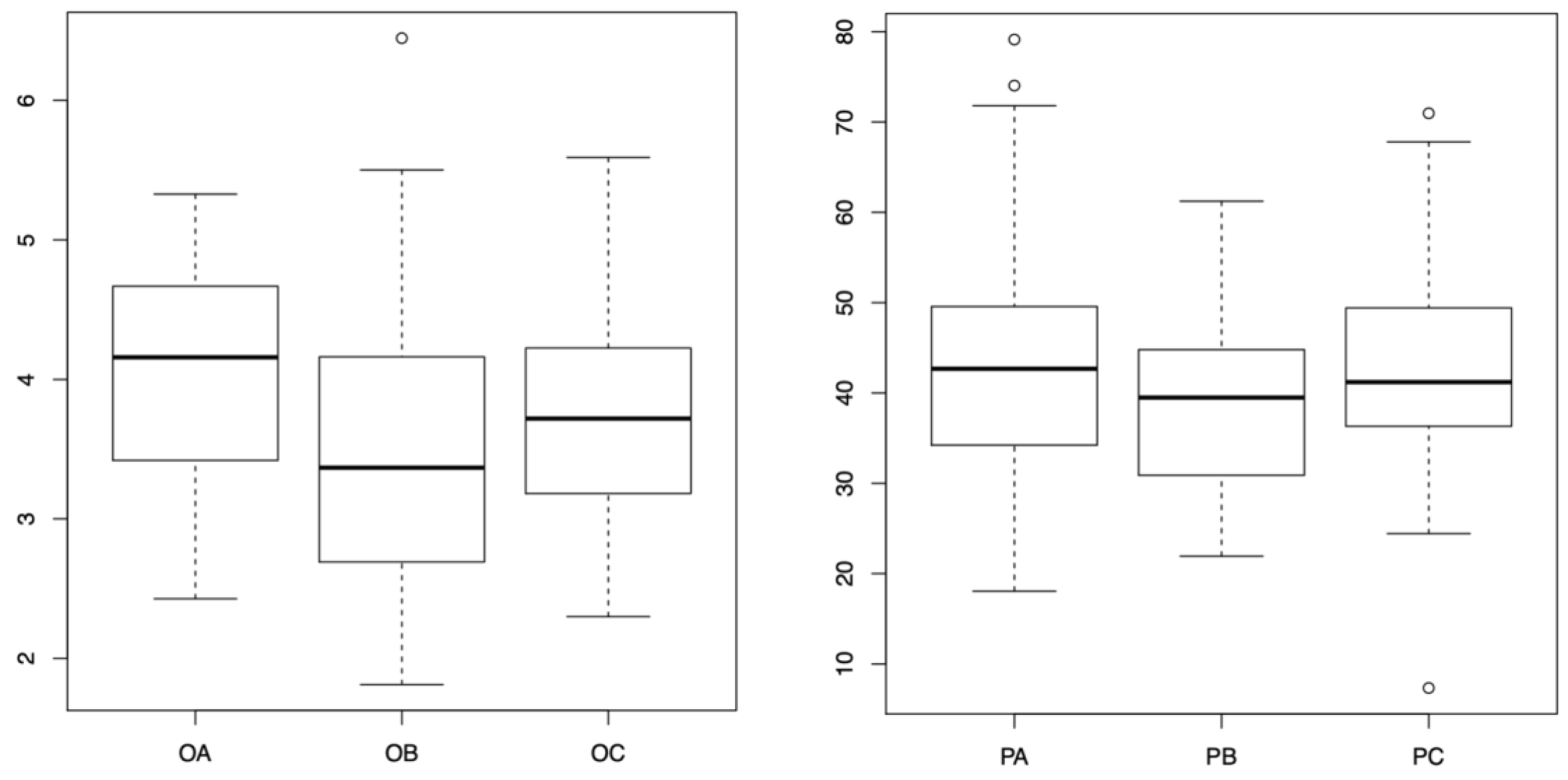
Figure 5.
Measurements of autofluorescence of chlorophyll for two tested moss species in two different stratospheric missions. O – O. anomalum, P – P. patens, A, B, C – day mission, night mission, control (respectively); vertical axis denotes measured mean grey values. Autofluorescence of the chlorophyll in Orthotrichum anomalum (O) and Physcomitrella patens (P) in A (Alpha), B (Beta) and C (Control) conditions. Scales for rows of images: 20 micrometres.
Figure 5.
Measurements of autofluorescence of chlorophyll for two tested moss species in two different stratospheric missions. O – O. anomalum, P – P. patens, A, B, C – day mission, night mission, control (respectively); vertical axis denotes measured mean grey values. Autofluorescence of the chlorophyll in Orthotrichum anomalum (O) and Physcomitrella patens (P) in A (Alpha), B (Beta) and C (Control) conditions. Scales for rows of images: 20 micrometres.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



