
Submitted:
18 February 2025
Posted:
18 February 2025
You are already at the latest version
Abstract
The Rhox family of homeobox transcription factors are established regulators of gonad function, but their downstream targets have been relatively elusive, particularly in the female reproductive tract. Here we characterize Ins2 as a downstream target of the two granulosa cell-specific factors, Rhox5 and Rhox8 in the ovary. While INS2 is classically produced by islet cells in the pancreas, we found that Ins2 gene expression is present in the mural granulosa cell layer of large antral follicles, and it was not significantly reduced in Rhox5-null mice. This was a surprising finding as we previously validated Ins2 as a direct target of RHOX5 in Sertoli cells, the male counterpart to granulosa cells that serves the germ cell nurse function in the testis. In the ovary, RHOX8 appears to be the major driver of Ins2 expression as evidenced from the maximal activity of Ins2 promoter reporter plasmids when RHOX8 protein was active within granulosa cells in vitro and the down-regulation of endogenous Ins2 in mice with granulosa cell-specific knockdown of RHOX8 in vivo. RHOX5 induces Rhox8 expression in pre-antral granulosa cells and then becomes relatively silent in peri-ovulatory follicles. However, Rhox8 does not peak until after the ovulatory LH surge. Induction of Rhox8 by progesterone, after the normal window of RHOX5 has passed, may explain why Rhox5-null female mice display apparently normal fertility, if RHOX8 is capable of redundant stimulation of target genes which are essential for ovulation.
Keywords:
Ovary
; Granulosa cells
; insulin regulation
; homeobox genes
1. Introduction
Defects in cellular metabolism such as disrupted insulin signaling in diabetic individuals are known to negatively impact fertility in rodents and primates [1]. Insulin II (Ins2) is one of two insulin genes in the mouse genome; it resides on chromosome 7, while Ins1 is located on chromosome 19. The mouse INS2 protein, which differs from mouse INS1 by only 5 amino-acid substitutions and 2 extra amino acids, has been shown to possess virtually identical functions as INS1, including the ability to strongly promote glucose uptake and protein synthesis [2,3]. While the Ins1 and Ins2 genes are best known for being expressed by islet cells in the pancreas (where they are expressed in a 1:2 ratio in the adult mouse pancreas [4]), one or both are also known to be expressed in the adult thymus and brain, as well as some embryonic organs and the yolk sac [5,6]. We and others have recently demonstrated the expression of Ins2, but not Ins1, mRNA in the testes [7,8]. The Akita mouse is a diabetic mouse model that possesses a point mutation in Ins2 that result in accumulation of misfolded INS2 protein that cannot activate insulin receptors and ultimately causes the destruction of insulin producing islet cells in the pancreas [9]. Interestingly, the Akita mouse exhibits a male subfertility phenotype that can be rescued by exogenous insulin treatment [8]. Female Akita mice and mice with streptozotocin induced destruction of INS-producing pancreatic cells exhibit defects in oocyte maturation, development, and granulosa cell growth and survival which lead to suboptimal ovulation and subfertility [10,11].
Successful ovulation requires the coordinated expression of genes which must be turned on and off in the right place at the right time for proper follicle development[12,13]. The developing oocyte is surrounded by granulosa cells which serve as nurse cells to support its maturation and growth. This is achieved in part by secreted hormones and growth factors from mural granulosa cells that act in a paracrine fashion as well as direct transfer between cumulus granulosa cells and the oocyte with the absence of granulosa cell signals leading to ovulation failure [14,15]. While the hormone signals from the pituitary that initiate ovulation are well-characterized, the master control genes that regulate follicle growth within the ovary are not as well known. Homeobox transcription factors (e.g. Hox genes) are master regulators of developmental programs and at least 35 homeobox genes are known to be expressed in the ovary. Studies have clearly demonstrated an essential role for Nobox and Lhx8 in ovulation [16,17,18] and the Iroquois and Obox homeobox gene clusters are differentially regulated during oocyte development, implying that they may be similarly important [19,20,21,22]. However, all of these factors are oocyte-specific.
We became interested in the transcriptional control of insulin signaling when we discovered that RHOX5 directly regulates Ins2 in mouse Sertoli cells [7]. Rhox5 is the founding member of the Reproductive Homeobox X-linked (Rhox) gene cluster which encodes 13 distinct transcription factors in mice, with three genes having undergone tandem duplications to generate a cluster of 42 total genes [23,24]. Primates, rats, hamsters, dogs, and cattle possess Rhox gene orthologs (e.g., RHOXF1, RHOXF2) in the syntenic position on the X chromosome, although the composition of the cluster varies between species [23,25,26]. The function of most RHOX factors is unknown, but they are selectively expressed in the placenta, gonads, and reproductive tract. In the gonads, our analyses have demonstrated that only Rhox5 and Rhox8 are expressed in postnatal Sertoli and granulosa nurse cells [27,28,29,30]. Ablation of these genes results in male subfertility characterized by reduced spermatogenic output and motility defects [31,32]. Interestingly, Rhox5-null mice have a remarkably similar phenotype to Akita mice, suggesting the RHOX5-regulation of Ins2 may underly the Sertoli-cell defect in spermatogenesis. However, Rhox5-null female mice do not exhibit significant fertility complications suggesting that if granulosa INS2 is essential for ovulation that other factors must contribute to its regulation [31]. Rhox5 is induced by follicle stimulating hormone (FSH) in granulosa cells and maintained by factors including GABP, SP1, CREB, and RAS signaling [27,29]. Rhox5 contributes to maximal Rhox8 induction in preantral follicles but is not required once luteinizing hormone (LH) and progesterone signaling are active [27]. We believe that ovulation is not severely compromised in Rhox5-null mice in part because RHOX8 expression is maintained in periovulatory follicles. In this report, we examine the regulation of the Ins2 gene in ovarian granulosa cells by RHOX5 and RHOX8. Our findings demonstrate that Ins2 expression is maintained at least in part by RHOX8 stimulation of its promoter region. However, whether this regulatory relationship is the essential factor that spares Rhox5-null female mice from local metabolic or growth factor signaling defects that compromise ovulation remains to be determined.
2. Materials and Methods
2.1. Mice
All animal experiments were performed in accordance with the National Institutes of Health guidelines and in compliance with the Southern Illinois University Carbondale and the Washington State University Institutional Animal Care and Use Committees. The generation, genotyping, and characterization of Rhox5-null mice has been previously reported [7,31,33,34]. Conditionally activated Rhox8 knockdown TARGATT mice were generated by Applied StemCell. Amhr2-Cre mice were provided by Richard Behringer (MD Anderson Cancer Center). All mice used in this study were maintained on a C57BL6 genetic background. All animals are housed under a 12:12 light-dark cycle at 70% humidity.
2.2. Plasmids and siRNA
To overexpress RHOX8, the Rhox8 coding sequence was cloned into the pCDNA5/FRT vector (Invitrogen-Thermo Fisher, Waltham, MA, USA), which expresses recombinant genes under the control of the cytomegalovirus (CMV) promoter. For stable Rhox8 expression, the pFRT/LacZeo plasmid was introduced in the genome of the rat Spontaneously Immortalized Granulosa Cell (SIGC) line. Subsequently, the Rhox8 transgene was flipped into the FRT site from the pCDNA5/FRT:Rhxo8 expression vector. Expression of Rhox8 was confirmed by RT-PCR and immunofluorescence labeling of RHOX8 protein using a 1:2000 dilution of rabbit polyclonal NBP2-23671 (Novus Biologicals, Minneapolis, MN, USA) which we previously validated in the gonads [27,30]. The Ins2 promoter parental reporter plasmid [35], deletion series, and RHOX binding site mutants were previously generated and characterized in our previous publication [7].
2.3. Superovulation and Granulosa Cell Cultures
For superovulation studies, female mice at postnatal age 21-28 days (PND21-28), selected by mass of at least 15 g for maximal response, were first injected with 5 IU equine chorionic gonadotropin (eCG; Biovendor RP178272, Ashville, NC) as described previously [27,36]. Subsequently, mice were collected for the eCG only group, or 48 h later given a single injection of 4 IU human chorionic gonadotropin (hCG; Sigma C0434, St. Louis, MO). Mice were euthanized 2-24 hours later and their ovaries removed for histological or in vitro analyses. One ovary was homogenized in Trizol (Invitrogen-Thermo Fisher, Waltham, MA, USA) for RNA isolation and one ovary was fixed in 4% paraformaldehyde dissolved in PBS, pH 7.4 for 12-16 hours then processed for embedding in paraffin.
Primary granulosa cells were isolated from eCG-primed mice as described previously [27,29]. Briefly, ovaries were removed and transferred to a 60-mm cell culture dish containing 5 ml of Dulbecco modified Eagle medium/F12 medium (DMEM/F12) supplemented with BSA, Fungizone, and gentamicin. Granulosa cells from multiple ovaries were pooled and treated with 20 µg/ml trypsin for 1 min, and then 300 µg/ml soybean trypsin inhibitor and 160 µg/ml DNase I were added to remove necrotic cells. Cells were cultured at 37°C in 95% air and 5% CO2 for 16 h before transfection. The SIGC line [37] were grown in DMEM -F12 medium supplemented with 5% FBS that was charcoal-stripped of hormones.
Transient transfection of both primary granulosa and SIGC was performed using the AttracteneTM transfection reagent (Qiagen, Germantown, MD, USA), which outperformed Lipofectamine 2000 (Invitrogen-Thermo Fisher, Waltham, MA, USA) and Turbofect (Fermentas-Thermo Fisher, Waltham, MA, USA), as assessed by cotransfection with green fluorescent protein (GFP) expression plasmids. Cell lysates were prepared and used to measure luciferase activity according to the manufacturer’s dual luciferase assay system protocol (Promega, Madison, WI, USA). Relative light units were normalized to the internal control plasmid pRL-TK and expressed as fold-change greater than that of the empty pGL3-basic vector. Transient knockdown of Rhox8 was achieved by transfection of Qiagen’s FlexiTube GeneSolution GS434768 kit for Rhox8. Which included a cocktail of four siRNA designed to block translation of RHOX8 and degrade Rhox8 mRNA.
2.3. Real-time Quantitative RT-PCR (qPCR) Analysis
Total RNA was isolated from ovaries using TRIzol reagent (Invitrogen-Thermo Fisher, Waltham, MA, USA), and then RT was performed using High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Waltham, MA, USA). The quantity and quality of RNA samples were determined by spectrometry and denaturing agarose gel electrophoresis, respectively. Real-time RT-PCR analysis of relative mRNA expression was performed on a MyiQ single-color real-time PCR detection system (Bio-Rad, Hercules, CA, USA) with iQ SYBR Green Supermix (Bio-Rad) according to the manufacturer’s recommendations. Real-time PCR was performed using the following protocol: 2 min at 95°C, 40 - 45 cycles of denaturation (15 sec at 95°C) and annealing/extension (1 min at 60°C), and a final step of melting curve analysis. As an internal control, Rpl19 was used. The relative levels of mRNA were calculated using the 2-ΔΔCt method. The primers for gene amplification have been previously reported [27,30,33].
2.4. In Situ Hybridization
Ovaries were sectioned at 5 µm and every 5th section transferred to a microscope slide and stained with hematoxylin and eosin to assess morphology as previously described [13]. In situ hybridization analysis of Ins2 mRNA expression was conducted using methods described previously [27,38]. Briefly, deparaffinized, rehydrated, and deproteinated sections were hybridized with radiolabeled sense or antisense cRNA probes generated from a linearized plasmid DNA template containing the entire Ins2 coding sequence via incorporation of α-35S-uridine 5’-triphosphate. After hybridization, washing, and ribonuclease A digestion, slides were dipped in NTB liquid photograph emulsion (Kodak, Rochester, NY, USA), stored at 4°C for 4-30 days, and developed in D-19 developer. Slides were then counterstained with Gill modified hematoxylin, dehydrated through ethanol series and xylene, and coverslipped. Regions of hybridization were visualized and localized by brightfield and darkfield microscopy.
2.5. Statistical Analysis
The qPCR expression time courses were subjected to one-way ANOVA Prism 9.0 (GraphPad, San Diego, CA, USA). Comparison of means between two groups were conducted using student t tests and differences between individual means of multiple grouped data were tested by a Tukey multiple-range post-test. All data met the necessary criteria for ANOVA analysis including equal variance as determined by Bartlett’s test. All experimental data are presented as mean ± SEM. Unless otherwise indicated, a P value of less than 0.05 was considered statistically significant.
3. Results
3.1. Ins2 Expression Tracks with Rhox8 Expression in the Ovary
As a first step towards examining whether RHOX5 and RHOX8 are candidates to regulate Ins2 in the ovary as was prevously found in the testis, we analyzed the time course of expression for each gene in hyperstimulated mice induced to ovulate with exogenous gonadotropins; equine chorionic gonadotropin (eCG, which mimics FSH in mice) and human chorionic gonadotropin (hCG, which mimics LH) that elicits ovulation 10-12 h after hCG administration. The granulosa cell-specific regulation and expression of RHOX5 and RHOX8 has previously been reported [27,29]. However, the temporal expression of Ins2 during follicle development and ovulation has not been assessed. In agreement with our prior studies, eCG induced the expression of Rhox5 and Rhox8, with Rhox5 peaking at 4 h post-hCG (Figure 1A) and Rhox8 at 8 h post-hCG (Figure 1B). Ins2 transcription was slightly induced by eCG but exhibited robust expression during the periovulatory window, similarly to Rhox8 (Figure 1C). At 4 h post-hCG, Ins2 mRNA was reduced in ovaries from Rhox5-null mice, but the decline was not significant.
The processed INS2 protein is small and secreted making it difficult to determine its cellular source or putative site of action in the ovary by immunohistochemistry with anti-insulin antibodies. In our prior report, we were unsuccessful in demonstrating Sertoli cell specific localization of insulin proteins and neither of the two polyclonal antibodies we tested would have distinguished INS1 vs INS2 if they had worked [7]. However, transcription of the Ins2 gene was robust enough to detect Ins2 mRNA in ovarian follicles by radioactive in situ hybridization. The expression of Ins2 does not appear strong enough to be detected in preantral follicles. However, Ins2 mRNA was localized to the mural granulosa cell layers of large antral follicles at 4 h post-hCG (Figure 2A, yellow arrow). In agreement with our qPCR analysis (Figure 1D), there was no obvious reduction in Ins2 mRNA in the granulosa cell layer of follicles in ovaries from Rhox5-null mice. At 8 h post-hCG, the upregulation of Ins2 observed by qPCR was recapitulated in the mural granulosa cell layer (Figure 2B). However, Ins2 expression in the cumulus granulosa cells layers was below the detection limit (Figure 2B, yellow arrow). The expression levels of Ins2 in periovulatory follicles of Rhox5-null mice were not decreased relative to wild-type control mice. The localization of Ins2 mRNA by the anti-sense probe was highly specific and no signals were detected with the negative control sense probe (Figure 2C).
3.2. Regulation of Ins2 in Spontaneously Immortalized Rat Granulosa Cells (SIGC)
The promoter region responsible for driving Ins2 expression in the rat pancreas was previously characterized [35]. We used modified versions of this Ins2 reporter plasmid to characterize RHOX regulation of Ins2 in mouse and rat Sertoli cells which are the analagous male counterpart to female granulosa cells [7]. The SIGC line was developed from primary rat ovarian preantral granulosa cells and is a good model to examine granulosa cell differentiation [37]. It lacks endogenous nuclear progesterone receptors [39]. Thus, it is not surprising that SIGC lack robust Rhox8 expression (Figure 3A) as we previously identified Rhox8 as a direct target of PGRA [27]. However, Rhox5 is highly expressed in SIGC likely because the line has the established signaling pathways for endogenous Rhox5 regulation [29,40], and is immortalized by overexpresion of TRP53 which activates Rhox5 in most tumor cells lines, regardless of their tissue of origin [23]. RHOX8 protein is barely detectable in parental SIGC cultures (Figure 3B). To employ SIGC cells as a tool for analysis of RHOX8-regulated factors, we generated a stable Rhox8 overexpression SIGC line which exhibited strong nuclear expression of RHOX8 (Figure 3C). This exogenous expression of RHOX8 could be knocked down to endogenous levels (or lower) by the transient transfection of an Rhox8 siRNA inhibitory cocktail (Figure 3D).
To to identify the specific promoter elements responsible for the regulation of Ins2 expression in granulosa cells, we examined the established 375 nucleotide (nt) Ins2 promoter promoter for putative activating transcription factors known to be expressed in the ovary (Figure 4A). We generated a series of deletion constructs from the full-length promoter luciferase reporter construct and transfected them into parental and RHOX8 overexpresing SIGC cells. We discovered that reduction of the promoter to only 103 nt did not elicit a significant drop in promoter activity (Figure 4B). SIGC cells with elevated RHOX8 exhibited a 4-fold increase in Ins2 promoter activity relative to parental SIGC cells. However, shortening the promoter to 72 nt, which eliminated the RHOX binding site we previously characterized in Sertoli cells [7], resulted in substantial downregulation of Ins2 promoter activity. The SP1 transcription factor is known to stimulate gene expression, including Rhox5 [29], in granulosa cells and further truncation of its binding site reduced reporter activity to background levels (Figure 4B). The putative homeobox binding site contains a consensus CTTAATG core binding site that when mutated to CTccATG in the context of the 103, 260, and 375 nt promoters, resulted in diminished activity (Figure 4C). When this “RHOX” binding site was intact, the promoter activity was ~2-3 fold higher in parental SIGC cells and 5-fold higher Rhox8 stable expressing cell lines indicating homeodomain transcription factors are a major driver of maximal Ins2 expression in granulosa cells and that RHOX8 is likely to provide greater stimulation than RHOX5.
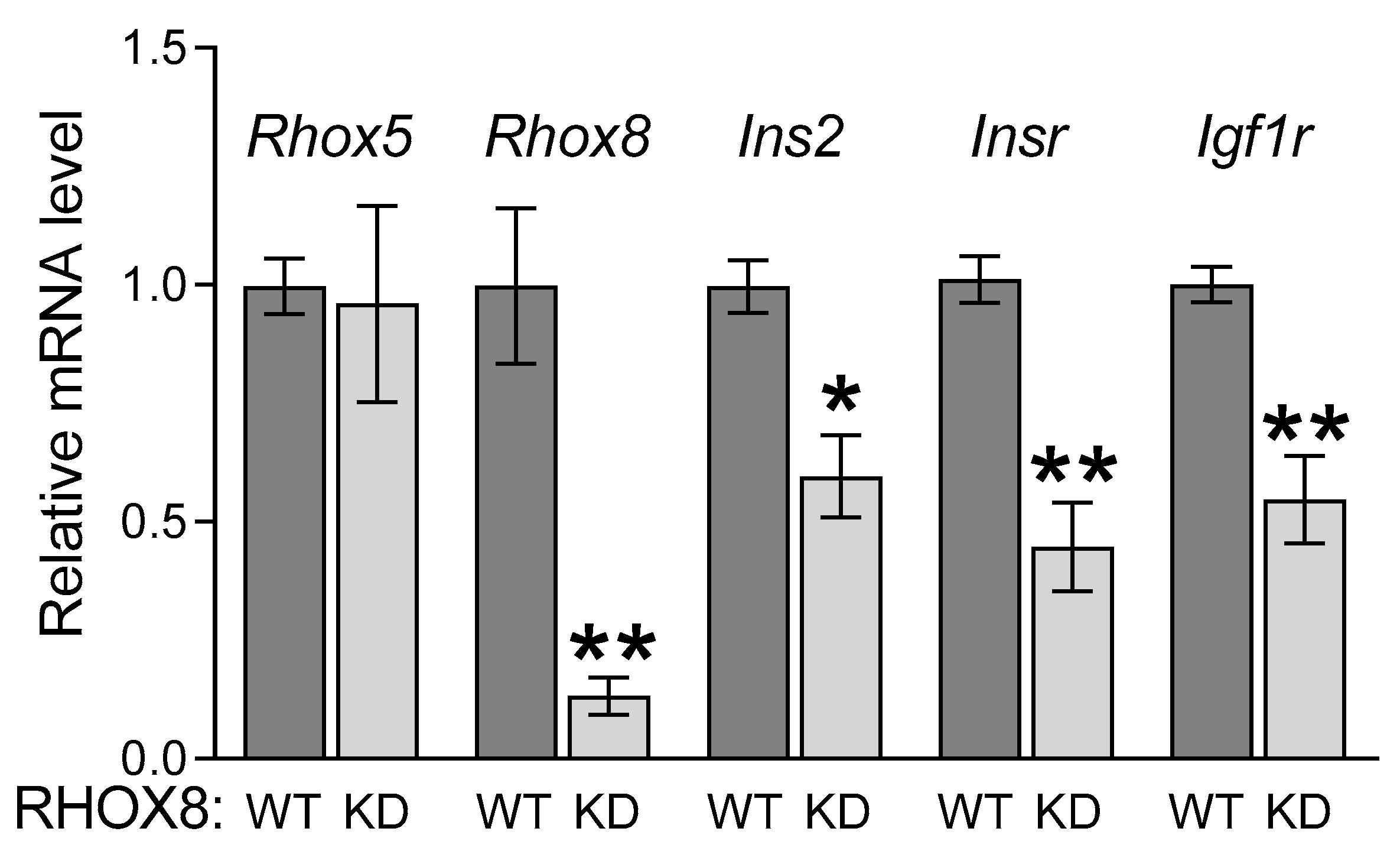
To determine if RHOX8 might activate Ins2 transcription in mouse granulosa cells, we created an inducible Rhox8 shRNA transgenic mouse (with the same validated targeting sequence shown in Figure 4D) that could be conditionally activated using Cre/Lox. These mice utilized Applied Stem Cells’ TARGATT system [43,44]. We chose this system because we had previously shown RHOX8 could be knocked down in vivo by RNA interference (RNAi) [30], and RNAi transgenes can act in a dominant fashion from a random genome integration site. This is necessary to examine Rhox5/Rhox8 double knockouts as the two genes are only 40 kb apart on the X chromosome and breeding of two single knockouts to eliminate both genes was unlikely. The Rhox8 targeting sequence was driven by the ubiquitous U6 Pol III promoter [45,46], that is activated by CRE recombinase removal of a stop cassette leading to transcription of the Rhox8-shRNA transgene and RHOX8 knockdown. We used the anti-Müllerian hormone type 2 receptor (Amhr2)-Cre to activate the RNAi transgene as it turns on in granulosa cells when Rhox5 and Rhox8 are initially induced [47]. The Amhr2-induced shRNA was effective and led to a 7-fold reduction in Rhox8 mRNA levels (Figure 5) in total RNA prepared from whole ovaries collected at 8h post-hCG. However, Rhox5 mRNA levels were unchanged in RHOX8-KD mice suggesting the knockdown effect was specific. The expression of Ins2 was diminished 2-fold after knockdown of RHOX8. We have recently examined the conditional ablation of the insulin receptor encoding genes Insr and Igf1r with Amhr2-Cre [48] or Pgr-Cre [36] and found that loss of insulin signaling results in suboptimal ovulation. Thus, we were curious if RHOX8 might regulate these receptors in vivo. The in vivo knockdown of RHOX8 resulted in a significant decrease in both Insr and Igf1r mRNA expression (Figure 5). However, whether the ~2-fold decrease is sufficient to alter insulin signaling enough to impact successful ovulation was not determined.
4. Discussion
In the present study, we examined the regulation of the primary rodent insulin hormone gene Ins2 by the two members of the Rhox gene cluster that are expressed in ovarian granulosa cells, Rhox5 and Rhox8. To date, the majority of RHOX studies have focused on male reproduction, mostly due to the fact that Rhox5 is a hallmark androgen-regulated gene and the spermatogenesis defects observed in Rhox5-null mice [24,49]. We previously demonstrated that in Sertoli cells, RHOX5 is the primary driver of Ins2 expression and it is one of the few direct targets for RHOX factors that has been validated by chromatin immunoprecipitation [7]. The Akita mouse which has a similar subfertility phenotype in males as Rhox5-null mice harbors a mutation in the INS2 protein that develops systemic diabetes that in turn affects fertility [8,50]. Thus, it has been our expectation that disrupted INS2-dependent signaling in our knockout mice has been a primary contributer to the germ cell loss in the testis, and potentially motility defects arising in the testis or epididymis, that we have previously described [7,31,33]. However, the Akita mouse also exhibits severe defects in ovulation [51], that were not found in Rhox5-null mice, suggesting that if Ins2 expression locally in the ovary is essential for ovulation, there must be additional factors that govern its expression.
Our in vitro and in vivo analyses herein indicates that RHOX8 could be one of these factors. Initially, RHOX5 may be able to stimulate Ins2 in the absence of RHOX8. Indeed, SIGC cells which lack endogenous RHOX8 have abundant expression of Rhox5, and parental SIGC cells do exhibit robust activation of the Ins2 promoter when the homeobox protein binding site (72 to 103 nt upstream of the Ins2 transcription starte site) that we have deemed the “RHOX” binding site is present. This suggests that in the absence of RHOX8, RHOX5 may supply at least basal Ins2 activation, but RHOX8 is a more potent activator of Ins2 in granulosa cells. It is possible that RHOX8 may synergize with RHOX5 to increase activation to maximal levels, or alternatively RHOX5 and/or RHOX8 may induce the expression of another homeodomain containing transcription factor which binds this element and maintains Ins2 expression throughout ovulation. Given the timing of expression, it cannot be ruled out that LH or progesterone could directly influence the expression of Ins2 in vivo. However, it is unlikely that Ins2 is a direct target of progesterone receptors as the reporter construct is highly active in SIGC cells which lack nuclear progesterone receptors (PGR), although a role of the membrane bound receptor isoform can not be ruled out. Indeed, the lack of nuclear PGR is likely why the parental SIGC lack endogenous Rhox8 expression.
In our prior report, we hypothesized that testes generate local insulins to either circumvent potential blocade by the blood-testis-barrier or more likely to keep spermatogenic output at a maximum when dietary state of males is in poor quality. However, it is a mystery why females would require local insulin as there is no barrier to oocyte access from the granulosa cells. Further, women with poor metabolic state have pathways in place to limit conception and implantation when there is not enough energy to support a successful pregnancy, so granulosa cell production of INS2 would seem counterintuitive. In addition to the control of glucose availability in cells, insulin signaling through its cognate receptors are known to initiate signaling cascades responsible for many developmental programs [52]. We have begun to dissect the insulin-dependent pathways that are downstream of INSR and IGF1R using conditional knockout models that preserve insulin secretion systemically (i.e., the mice are not diabetic), but lack the ability to respond to insulin ligands in granulosa cells. In the first report [36], we used Pgr-Cre which resulted in ablation of insulin receptors primarily in the same periovulatory window where we observed Ins2 expression. The fertility complications of these mice were primarly due to dysregulation of uterine proliferation and implantation failure [53]. However, mice lacking both INSR and IGF1R had a 50% reduction in ovulation with oocytes becoming trapped in corpora lutea.
Progesterone production by these abnormal corpora lutea was diminished, which sugests that INS2 might serve as part of a feedback loop in response to progesterone-regualted Rhox8 that folliculogenesis is proceeding normally. We have similarly used Amhr2-Cre to ablate Insr and Igf1r in granulosa cells of follicles transitioning from pre-antral to antral stage [48]. We found that Amhr2-Cre was insufficient to elicit a significant impact on ovarian function as ovulation rates were normal and the mice exhibited typical estrous cycles. Immunohistochemistry for both receptors indicated that mosiac expression was present at the time of ovulation. We are not certain if that was due to suboptimal or non-uniform activity of the Cre or whether INSR and IGF1R that was present in secondary follicles at the time of Amhr2-Cre activation simply did not turn over until follicles reached the ability to skip the block in antral formation. The half-life of insulin receptors in ovarian cells has not be examined, but it's likely that both factors may contribute to the minor impact on ovulation. In this report, we used Amhr2-Cre to activate an inhibitory transgene to knockdown RHOX8. We did observe a ~50% decline of Ins2, Insr, and Igf1r in total ovarian RNA from knockdown ovaries. This provides further evidence that the RHOX8 regulation of Ins2 expression we found in our promoter analysis may be relevant in vivo. The potential co-regulation of INS2 receptors is also interesting as RHOX8 may optimize the ability of follicles to respond to growth promoting and differentiation promoting insulin-dependnt factors. We did not formally quantitate residual RHOX8 expression in this model, but ~20% of mural granulosa cells still had nuclear RHOX8 at 8 h post-hCG, which matches the amount of Rhox8 mRNA that could still be detected by qPCR and in the future we will examine Rhox8-null mice for dysregulation of these genes with enhanced rigor.
5. Conclusions
In contrast to male mice where RHOX5 is a primary driver of Ins2 expression in the testis, we found that while Rhox5 may be permissive for INS2 secretion, it is RHOX8 that is the likely primary driver of Ins2 expression in granulosa cells of the ovary.
Author Contributions
K.H. and J.A.M. conceptualized the study; methodology, K.H. and J.A.M.; software, J.A.M.; validation, J.A.M.; formal analysis, K.H. and J.A.M.; investigation, K.H. and J.A.M.; writing J.A.M; project administration and funding acquisition, J.A.M. Both authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Institutes of Health, Eunice Kennedy Shriver National Institute of Child Health and Human Development grant HD065584 to J.A.M.II.
Data Availability Statement
The original contributions presented in the study are included in the article and supplementary materials, further inquiries can be directed to the corresponding author (J.A.M.II).
Acknowledgments
The authors would like to thank former trainees Raquel Brown and Cassandra Showmaker (Burke) for their assistance in animal management and sample collection. The authors thank Dr. Robert Burghardt (Texas A&M University) for supplying the rat SIGC line.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study, collection and analysis of data, the drafting of the manuscript, and the decision to publish the findings presented herein.
Abbreviations
The following abbreviations are used in this manuscript:
| SIGC | Spontaneously Immortalized Granulosa Cells |
| PGR | Progesterone receptor |
| LH | Luteinizing hormone |
| qPCR | Quantitative real-time reverse transcription polymerase chain reaction |
References
- Neirijnck, Y.; Papaioannou, M.D.; Nef, S. The Insulin/IGF System in Mammalian Sexual Development and Reproduction. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef]
- Wentworth, B.M.; Schaefer, I.M.; Villa-Komaroff, L.; Chirgwin, J.M. Characterization of the two nonallelic genes encoding mouse preproinsulin. Journal of molecular evolution 1986, 23, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.I.; Clemmons, D.R. Insulin-like growth factors and their binding proteins: biological actions. Endocrine reviews 1995, 16, 3–34. [Google Scholar] [PubMed]
- Bengtsson, M.; Stahlberg, A.; Rorsman, P.; Kubista, M. Gene expression profiling in single cells from the pancreatic islets of Langerhans reveals lognormal distribution of mRNA levels. Genome research 2005, 15, 1388–1392. [Google Scholar] [CrossRef] [PubMed]
- Deltour, L.; Leduque, P.; Blume, N.; Madsen, O.; Dubois, P.; Jami, J.; Bucchini, D. Differential expression of the two nonallelic proinsulin genes in the developing mouse embryo. Proceedings of the National Academy of Sciences of the United States of America 1993, 90, 527–531. [Google Scholar] [CrossRef]
- Heath, V.L.; Moore, N.C.; Parnell, S.M.; Mason, D.W. Intrathymic expression of genes involved in organ specific autoimmune disease. J Autoimmun 1998, 11, 309–318. [Google Scholar] [CrossRef]
- MacLean, J.A., 2nd; Hu, Z.; Welborn, J.P.; Song, H.W.; Rao, M.K.; Wayne, C.M.; Wilkinson, M.F. The RHOX homeodomain proteins regulate the expression of insulin and other metabolic regulators in the testis. The Journal of biological chemistry 2013, 288, 34809–34825. [Google Scholar] [CrossRef]
- Schoeller, E.L.; Albanna, G.; Frolova, A.I.; Moley, K.H. Insulin rescues impaired spermatogenesis via the hypothalamic-pituitary-gonadal axis in Akita diabetic mice and restores male fertility. Diabetes 2012, 61, 1869–1878. [Google Scholar] [CrossRef]
- Izumi, T.; Yokota-Hashimoto, H.; Zhao, S.; Wang, J.; Halban, P.A.; Takeuchi, T. Dominant negative pathogenesis by mutant proinsulin in the Akita diabetic mouse. Diabetes 2003, 52, 409–416. [Google Scholar] [CrossRef]
- Chang, A.S.; Dale, A.N.; Moley, K.H. Maternal diabetes adversely affects preovulatory oocyte maturation, development, and granulosa cell apoptosis. Endocrinology 2005, 146, 2445–2453. [Google Scholar] [CrossRef]
- Wang, Q.; Moley, K.H. Maternal diabetes and oocyte quality. Mitochondrion 2010, 10, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Jo, M.; Brannstrom, M.; Akins, J.W.; Curry, T.E., Jr. New insights into the ovulatory process in the human ovary. Human reproduction update 2025, 31, 21–47. [Google Scholar] [CrossRef]
- Kumar, T.R. Rerouting of follicle-stimulating hormone secretion and gonadal function. Fertil Steril 2023, 119, 180–183. [Google Scholar] [CrossRef] [PubMed]
- Ackert, C.L.; Gittens, J.E.; O'Brien, M.J.; Eppig, J.J.; Kidder, G.M. Intercellular communication via connexin43 gap junctions is required for ovarian folliculogenesis in the mouse. Developmental biology 2001, 233, 258–270. [Google Scholar] [CrossRef] [PubMed]
- Chang, R.J.; Cook-Andersen, H. Disordered follicle development. Mol Cell Endocrinol 2013, 373, 51–60. [Google Scholar] [CrossRef]
- Pangas, S.A.; Choi, Y.; Ballow, D.J.; Zhao, Y.; Westphal, H.; Matzuk, M.M.; Rajkovic, A. Oogenesis requires germ cell-specific transcriptional regulators Sohlh1 and Lhx8. Proceedings of the National Academy of Sciences of the United States of America 2006, 103, 8090–8095. [Google Scholar] [CrossRef]
- Qin, Y.; Choi, Y.; Zhao, H.; Simpson, J.L.; Chen, Z.J.; Rajkovic, A. NOBOX homeobox mutation causes premature ovarian failure. Am J Hum Genet 2007, 81, 576–581. [Google Scholar] [CrossRef]
- Rajkovic, A.; Pangas, S.A.; Ballow, D.; Suzumori, N.; Matzuk, M.M. NOBOX deficiency disrupts early folliculogenesis and oocyte-specific gene expression. Science 2004, 305, 1157–1159. [Google Scholar] [CrossRef]
- Jorgensen, J.S.; Gao, L. Irx3 is differentially up-regulated in female gonads during sex determination. Gene Expr Patterns 2005, 5, 756–762. [Google Scholar] [CrossRef]
- Rajkovic, A.; Yan, C.; Yan, W.; Klysik, M.; Matzuk, M.M. Obox, a family of homeobox genes preferentially expressed in germ cells. Genomics 2002, 79, 711–717. [Google Scholar] [CrossRef]
- Lee, H.S.; Kim, E.Y.; Kim, K.H.; Moon, J.; Park, K.S.; Kim, K.S.; Lee, K.A. Obox4 critically regulates cAMP-dependent meiotic arrest and MI-MII transition in oocytes. FASEB journal : official publication of the Federation of American Societies for Experimental Biology 2010, 24, 2314–2324. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Kim, K.H.; Kim, E.Y.; Lee, S.Y.; Ko, J.J.; Lee, K.A. Obox4-silencing activated STAT3 and MPF/MAPK signaling accelerate GVBD in mouse oocytes. Reproduction 2016. [Google Scholar] [CrossRef] [PubMed]
- MacLean, J.A. , 2nd. The role of Rhox homeobox factors in tumorigenesis. Front Biosci (Landmark Ed) 2013, 18, 474–492. [Google Scholar] [CrossRef]
- MacLean, J.A., 2nd; Wilkinson, M.F. The Rhox genes. Reproduction 2010, 140, 195–213. [Google Scholar] [CrossRef]
- Wilming, L.G.; Boychenko, V.; Harrow, J.L. Comprehensive comparative homeobox gene annotation in human and mouse. Database (Oxford) 2015, 2015. [Google Scholar] [CrossRef]
- Wayne, C.M.; MacLean, J.A.; Cornwall, G.; Wilkinson, M.F. Two novel human X-linked homeobox genes, hPEPP1 and hPEPP2, selectively expressed in the testis. Gene 2002, 301, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.M.; Davis, M.G.; Hayashi, K.; MacLean, J.A. Regulated expression of Rhox8 in the mouse ovary: evidence for the role of progesterone and RHOX5 in granulosa cells. Biology of reproduction 2013, 88, 126. [Google Scholar] [CrossRef]
- Daggag, H.; Svingen, T.; Western, P.S.; van den Bergen, J.A.; McClive, P.J.; Harley, V.R.; Koopman, P.; Sinclair, A.H. The rhox homeobox gene family shows sexually dimorphic and dynamic expression during mouse embryonic gonad development. Biology of reproduction 2008, 79, 468–474. [Google Scholar] [CrossRef]
- MacLean, J.A., 2nd; Rao, M.K.; Doyle, K.M.; Richards, J.S.; Wilkinson, M.F. Regulation of the Rhox5 homeobox gene in primary granulosa cells: preovulatory expression and dependence on SP1/SP3 and GABP. Biology of reproduction 2005, 73, 1126–1134. [Google Scholar] [CrossRef]
- Welborn, J.P.; Davis, M.G.; Ebers, S.D.; Stodden, G.R.; Hayashi, K.; Cheatwood, J.L.; Rao, M.K.; MacLean, J.A. , 2nd. Rhox8 Ablation in the Sertoli Cells Using a Tissue-Specific RNAi Approach Results in Impaired Male Fertility in Mice. Biology of reproduction 2015, 93, 8. [Google Scholar] [CrossRef]
- Maclean, J.A., 2nd; Chen, M.A.; Wayne, C.M.; Bruce, S.R.; Rao, M.; Meistrich, M.L.; Macleod, C.; Wilkinson, M.F. Rhox: a new homeobox gene cluster. Cell 2005, 120, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.; Kasu, M.; Bottoms, C.J.; Douglas, J.C.; Sekulovski, N.; Hayashi, K.; MacLean Ii, J.A. Rhox8 homeobox gene ablation leads to rete testis abnormality and male subfertility in micedagger. Biology of reproduction 2023, 109, 520–532. [Google Scholar] [CrossRef] [PubMed]
- MacLean, J.A., 2nd; Hayashi, K.; Turner, T.T.; Wilkinson, M.F. The Rhox5 homeobox gene regulates the region-specific expression of its paralogs in the rodent epididymis. Biology of reproduction 2012, 86, 189. [Google Scholar] [CrossRef]
- Pitman, J.L.; Lin, T.P.; Kleeman, J.E.; Erickson, G.F.; MacLeod, C.L. Normal reproductive and macrophage function in Pem homeobox gene-deficient mice. Developmental biology 1998, 202, 196–214. [Google Scholar] [CrossRef] [PubMed]
- Fleenor, D.E.; Freemark, M. Prolactin induction of insulin gene transcription: roles of glucose and signal transducer and activator of transcription 5. Endocrinology 2001, 142, 2805–2810. [Google Scholar] [CrossRef]
- Sekulovski, N.; Whorton, A.E.; Shi, M.; Hayashi, K.; MacLean, J.A. , 2nd. Periovulatory insulin signaling is essential for ovulation, granulosa cell differentiation, and female fertility. FASEB journal : official publication of the Federation of American Societies for Experimental Biology 2020, 34, 2376–2391. [Google Scholar] [CrossRef]
- Stein, L.S.; Stoica, G.; Tilley, R.; Burghardt, R.C. Rat ovarian granulosa cell culture: a model system for the study of cell-cell communication during multistep transformation. Cancer research 1991, 51, 696–706. [Google Scholar]
- Hayashi, K.; Yoshioka, S.; Reardon, S.N.; Rucker, E.B., 3rd; Spencer, T.E.; DeMayo, F.J.; Lydon, J.P.; MacLean, J.A. , 2nd. WNTs in the neonatal mouse uterus: potential regulation of endometrial gland development. Biology of reproduction 2011, 84, 308–319. [Google Scholar] [CrossRef]
- Stein, L.S.; Stein, D.W.; Echols, J.; Burghardt, R.C. Concomitant alterations of desmosomes, adhesiveness, and diffusion through gap junction channels in a rat ovarian transformation model system. Exp Cell Res 1993, 207, 19–32. [Google Scholar] [CrossRef]
- Yu, J.; Yaba, A.; Kasiman, C.; Thomson, T.; Johnson, J. mTOR controls ovarian follicle growth by regulating granulosa cell proliferation. PloS one 2011, 6, e21415. [Google Scholar] [CrossRef]
- Chen, X.; Fang, F.; Liou, Y.C.; Ng, H.H. Zfp143 regulates Nanog through modulation of Oct4 binding. Stem cells (Dayton, Ohio) 2008, 26, 2759–2767. [Google Scholar] [CrossRef]
- Vlaeminck-Guillem, V.; Vanacker, J.M.; Verger, A.; Tomavo, N.; Stehelin, D.; Laudet, V.; Duterque-Coquillaud, M. Mutual repression of transcriptional activation between the ETS-related factor ERG and estrogen receptor. Oncogene 2003, 22, 8072–8084. [Google Scholar] [CrossRef] [PubMed]
- Hippenmeyer, S.; Youn, Y.H.; Moon, H.M.; Miyamichi, K.; Zong, H.; Wynshaw-Boris, A.; Luo, L. Genetic mosaic dissection of Lis1 and Ndel1 in neuronal migration. Neuron 2010, 68, 695–709. [Google Scholar] [CrossRef] [PubMed]
- Tasic, B.; Hippenmeyer, S.; Wang, C.; Gamboa, M.; Zong, H.; Chen-Tsai, Y.; Luo, L. Site-specific integrase-mediated transgenesis in mice via pronuclear injection. Proceedings of the National Academy of Sciences of the United States of America 2011, 108, 7902–7907. [Google Scholar] [CrossRef] [PubMed]
- Shukla, V.; Coumoul, X.; Deng, C.X. RNAi-based conditional gene knockdown in mice using a U6 promoter driven vector. International journal of biological sciences 2007, 3, 91–99. [Google Scholar] [CrossRef]
- Shukla, V.; Coumoul, X.; Wang, R.H.; Kim, H.S.; Deng, C.X. RNA interference and inhibition of MEK-ERK signaling prevent abnormal skeletal phenotypes in a mouse model of craniosynostosis. Nat Genet 2007, 39, 1145–1150. [Google Scholar] [CrossRef]
- Jamin, S.P.; Arango, N.A.; Mishina, Y.; Hanks, M.C.; Behringer, R.R. Genetic studies of the AMH/MIS signaling pathway for Mullerian duct regression. Mol Cell Endocrinol 2003, 211, 15–19. [Google Scholar] [CrossRef]
- Douglas, J.C.; Sekulovski, N.; Arreola, M.R.; Oh, Y.; Hayashi, K.; MacLean, J.A. , 2nd. Normal Ovarian Function in Subfertile Mouse with Amhr2-Cre-Driven Ablation of Insr and Igf1r. Genes (Basel) 2024, 15. [Google Scholar] [CrossRef]
- Le Beulze, M.; Daubech, C.; Balde-Camara, A.; Ghieh, F.; Vialard, F. Mammal Reproductive Homeobox (Rhox) Genes: An Update of Their Involvement in Reproduction and Development. Genes (Basel) 2023, 14. [Google Scholar] [CrossRef]
- Kim, S.T.; Moley, K.H. Paternal effect on embryo quality in diabetic mice is related to poor sperm quality and associated with decreased glucose transporter expression. Reproduction 2008, 136, 313–322. [Google Scholar] [CrossRef]
- Lee, J.; Lee, H.C.; Kim, S.Y.; Cho, G.J.; Woodruff, T.K. Poorly-Controlled Type 1 Diabetes Mellitus Impairs LH-LHCGR Signaling in the Ovaries and Decreases Female Fertility in Mice. Yonsei Med J 2019, 60, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Nandi, A.; Wang, X.; Accili, D.; Wolgemuth, D.J. The effect of insulin signaling on female reproductive function independent of adiposity and hyperglycemia. Endocrinology 2010, 151, 1863–1871. [Google Scholar] [CrossRef] [PubMed]
- Sekulovski, N.; Whorton, A.E.; Shi, M.; Hayashi, K.; MacLean, J.A. , 2nd. Insulin signaling is an essential regulator of endometrial proliferation and implantation in mice. FASEB journal : official publication of the Federation of American Societies for Experimental Biology 2021, 35, e21440. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Rhox8 and Ins2 expression peak during the periovulatory window. (a-c) Immature C57BL/6 mice were superovulated with a single injection of 5 IU eCG followed by an injection of 4 IU hCG 48h later. Intact ovaries were extirpated at 0, 2, 4, 8, 12, 16, and 24 hours after hCG administration and RNA extracted. Relative expression was determined for each gene by qPCR [27]. Values (n= 6 animals per time point) were normalized against ribosomal L19 (Rpl19) mRNA and bars are shown as fold above background (+/- SEM), which was arbitrarily given a value of 1. Letters denote mean values that were significantly different (P < 0.05, one-way ANOVA with Tukey multiple-range post-test). Data is segregated into immature (purple), preovulatory (green), periovulatory (blue) and luteal (orange) phases of follicular development. (d) The expression of Ins2 was assessed in ovaries from superovulated Rhox5-null mice were collected at 4 h post-hCG and 8 h post-hCG and no significant differences in expression were observed (student’s t test) between Rhox5-null animals and wild-type (WT) animals which were normalized against Rpl19 and given a value of 100 (n=6 ovaries per time point and genotype).
Figure 1.
Rhox8 and Ins2 expression peak during the periovulatory window. (a-c) Immature C57BL/6 mice were superovulated with a single injection of 5 IU eCG followed by an injection of 4 IU hCG 48h later. Intact ovaries were extirpated at 0, 2, 4, 8, 12, 16, and 24 hours after hCG administration and RNA extracted. Relative expression was determined for each gene by qPCR [27]. Values (n= 6 animals per time point) were normalized against ribosomal L19 (Rpl19) mRNA and bars are shown as fold above background (+/- SEM), which was arbitrarily given a value of 1. Letters denote mean values that were significantly different (P < 0.05, one-way ANOVA with Tukey multiple-range post-test). Data is segregated into immature (purple), preovulatory (green), periovulatory (blue) and luteal (orange) phases of follicular development. (d) The expression of Ins2 was assessed in ovaries from superovulated Rhox5-null mice were collected at 4 h post-hCG and 8 h post-hCG and no significant differences in expression were observed (student’s t test) between Rhox5-null animals and wild-type (WT) animals which were normalized against Rpl19 and given a value of 100 (n=6 ovaries per time point and genotype).
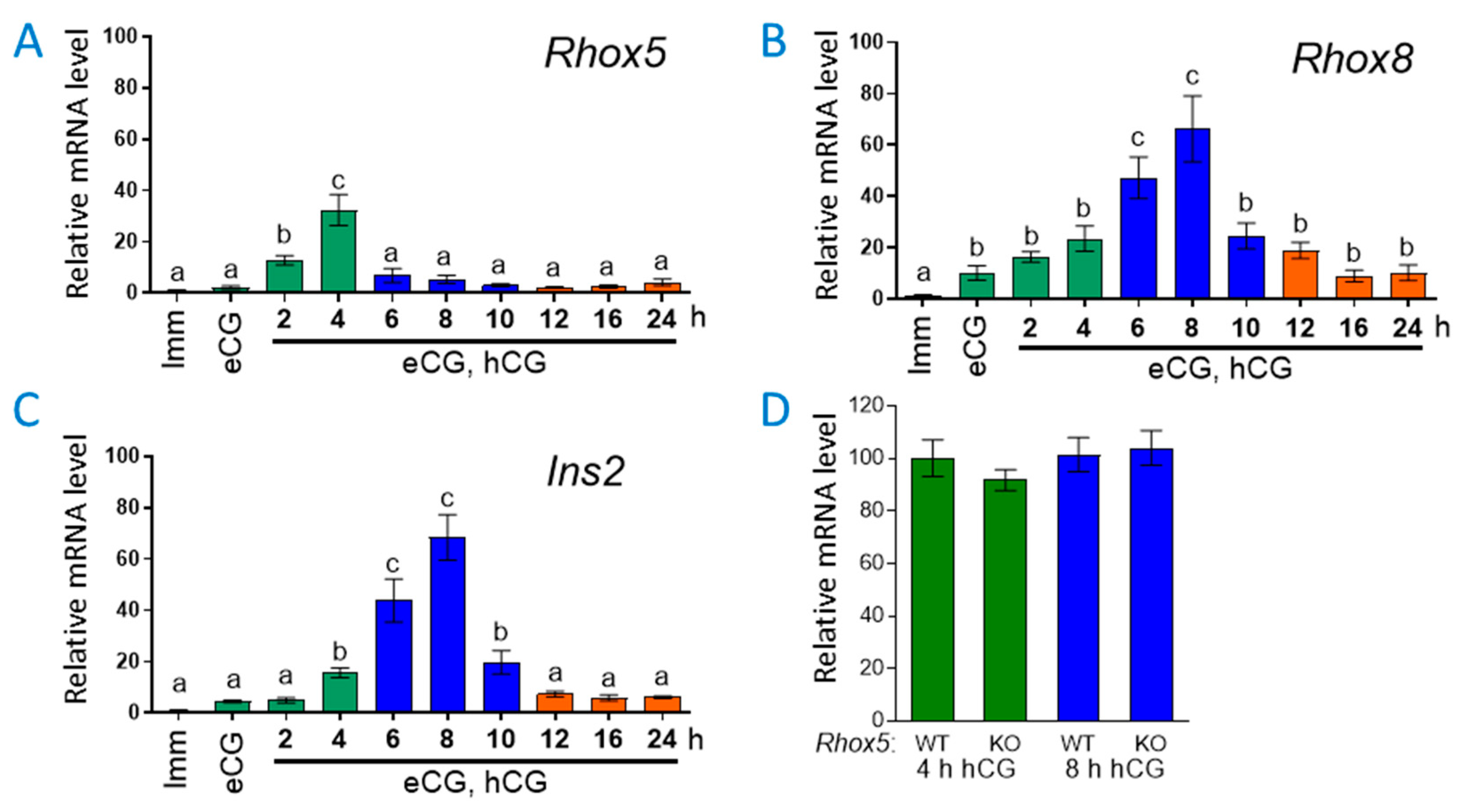
Figure 2.
Ins2 transcripts are localized to antral follicles and does not appear to be regulated by RHOX5. Representative photomicrographs show that Ins2 mRNA is present in antral follicles of superovulated mice at (a) 4 h post-hCG. Transcripts were abundant in the mural granulosa cell layer, yellow arrows. No differences between wild-type and Rhox5-null mice were observed. (b) Ins2 expression increased at 8 h post-hCG but is not different between wild-type and Rhox5-null mice. The cumulus granulosa cell layer around the oocyte appeard to lack Ins2 transcripts, yellow arrow. (c) No appreciable signal was observed with the Ins2 sense strand probe in any tissue section.
Figure 2.
Ins2 transcripts are localized to antral follicles and does not appear to be regulated by RHOX5. Representative photomicrographs show that Ins2 mRNA is present in antral follicles of superovulated mice at (a) 4 h post-hCG. Transcripts were abundant in the mural granulosa cell layer, yellow arrows. No differences between wild-type and Rhox5-null mice were observed. (b) Ins2 expression increased at 8 h post-hCG but is not different between wild-type and Rhox5-null mice. The cumulus granulosa cell layer around the oocyte appeard to lack Ins2 transcripts, yellow arrow. (c) No appreciable signal was observed with the Ins2 sense strand probe in any tissue section.
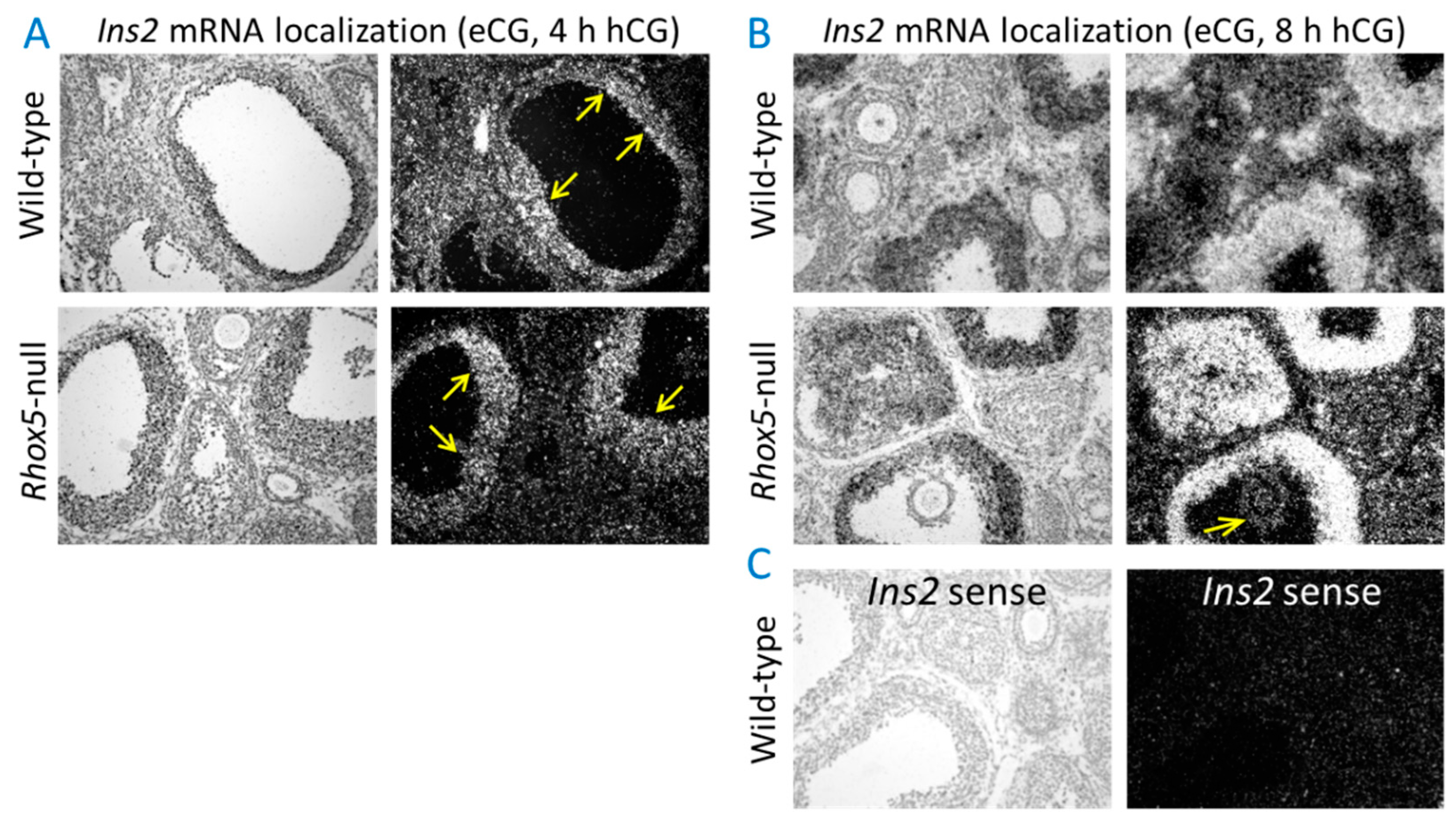
Figure 3.
Generation of a stable RHOX8 expressing SIGC line. (a) The endogenous levels of rat Rhox5 and Rhox8 were determined in parental SIGC cells by qPCR (n=6). (b) SIGC cells lack robust nuclear RHOX8 expression as assessed by immunoflouroresence in two independent SIGC cultures. (c) The stable integration of Rhox8 transgene into the flip-in site of LacZeo competent SIGC cells resulted in uniform abundant nuclear expression of RHOX8. (d) The expression of RHOX8 could be inhibited by the addition of a Rhox8 siRNA cocktail.
Figure 3.
Generation of a stable RHOX8 expressing SIGC line. (a) The endogenous levels of rat Rhox5 and Rhox8 were determined in parental SIGC cells by qPCR (n=6). (b) SIGC cells lack robust nuclear RHOX8 expression as assessed by immunoflouroresence in two independent SIGC cultures. (c) The stable integration of Rhox8 transgene into the flip-in site of LacZeo competent SIGC cells resulted in uniform abundant nuclear expression of RHOX8. (d) The expression of RHOX8 could be inhibited by the addition of a Rhox8 siRNA cocktail.
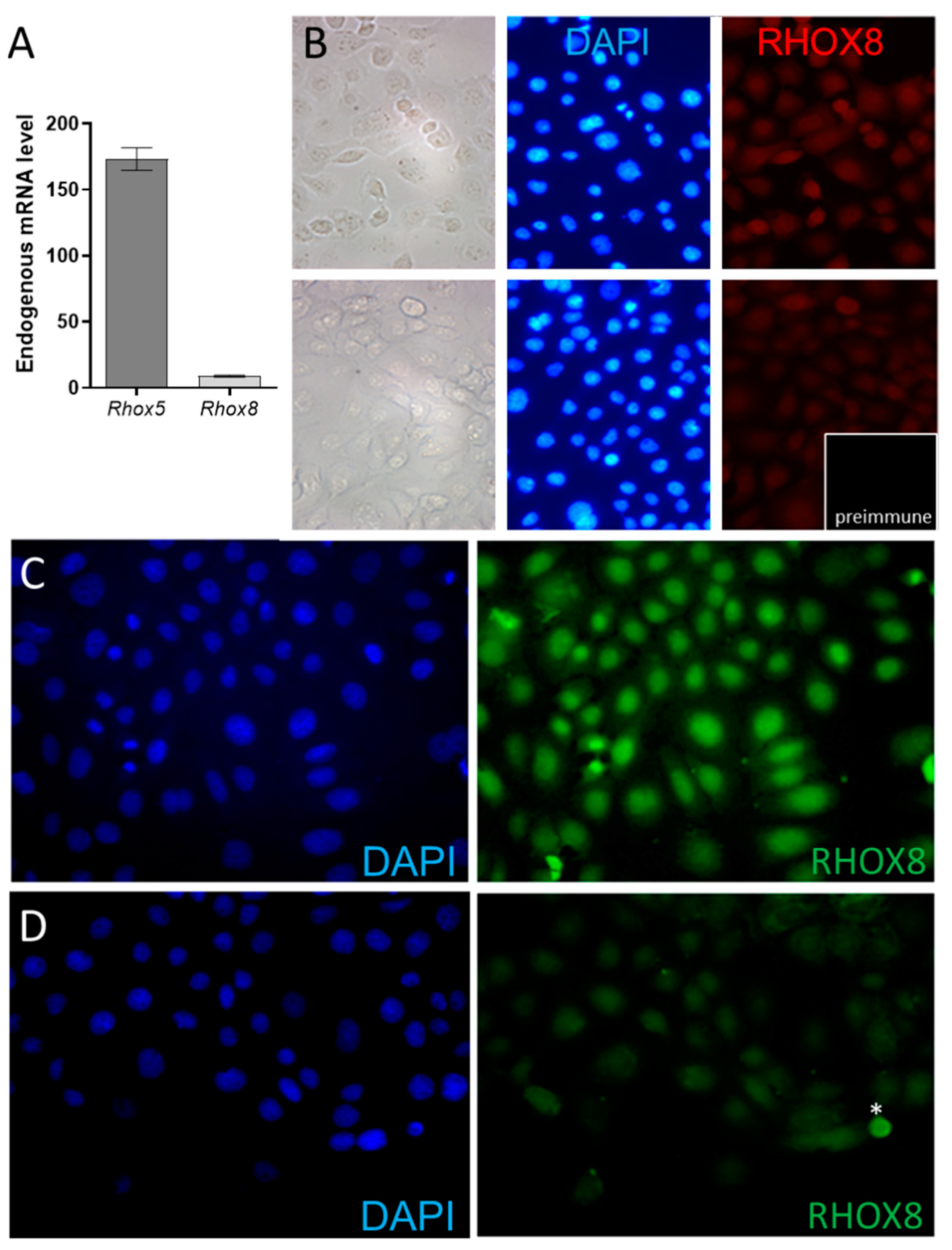
Figure 4.
RHOX8 drives maximal Ins2 promoter activity in SIGC cells. (a) Putative transcription factor binding sites contained within the Ins2 promoter were determined by TESS and TFSEARCH algorithms. Putative positive regulators of Ins2 expression are indicated in green from prior studies [7,29,41,42] and factors of unknown relevance but established granulosa expression in blue. (b) Relative luciferase activity from Ins2 promoter constructs transiently transfected rat SIGC granulosa cells. Values shown are the ratios of Ins2 promoter-dependent firefly luciferase activity normalized by Renilla luciferase activity internal control (pRL-TK) for transfection efficiency and expressed as fold above pGL3-basic vector which was arbitrarily assigned a value of 1. Assays were performed in triplicate with three independent preparations of SIGC and Rhox8 Stable cell clones. Data are shown as mean ± SEM relative promoter activity and significant differences determined by unpaired t-test, **P<0.001. (c) Promoter activity in constructs with mutation in the putative homeobox binding site (*RHOX) were assayed as in panel B.
Figure 4.
RHOX8 drives maximal Ins2 promoter activity in SIGC cells. (a) Putative transcription factor binding sites contained within the Ins2 promoter were determined by TESS and TFSEARCH algorithms. Putative positive regulators of Ins2 expression are indicated in green from prior studies [7,29,41,42] and factors of unknown relevance but established granulosa expression in blue. (b) Relative luciferase activity from Ins2 promoter constructs transiently transfected rat SIGC granulosa cells. Values shown are the ratios of Ins2 promoter-dependent firefly luciferase activity normalized by Renilla luciferase activity internal control (pRL-TK) for transfection efficiency and expressed as fold above pGL3-basic vector which was arbitrarily assigned a value of 1. Assays were performed in triplicate with three independent preparations of SIGC and Rhox8 Stable cell clones. Data are shown as mean ± SEM relative promoter activity and significant differences determined by unpaired t-test, **P<0.001. (c) Promoter activity in constructs with mutation in the putative homeobox binding site (*RHOX) were assayed as in panel B.
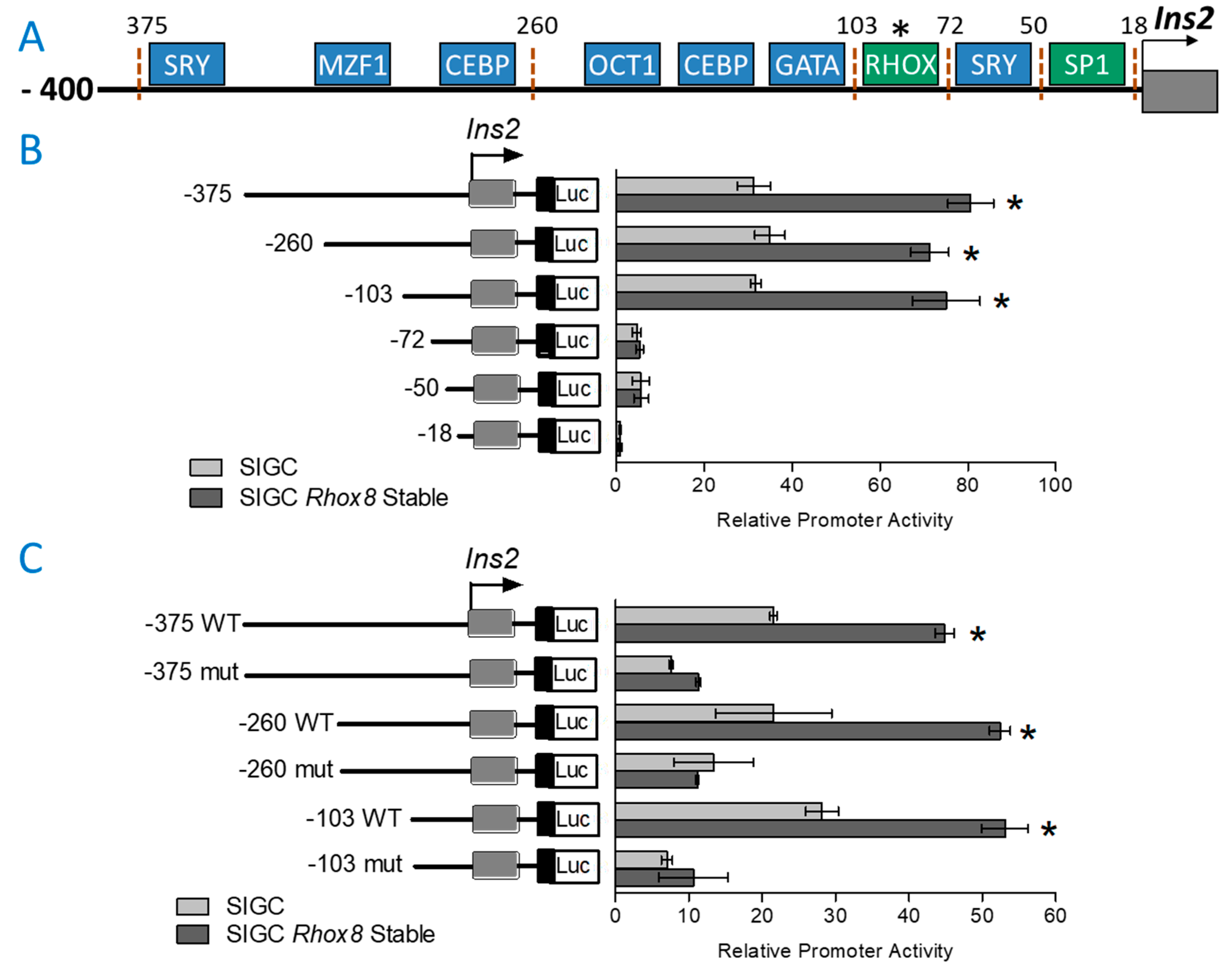
Figure 5.
In vivo knockdown of RHOX8 results in reduced expression of insulin signaling in mouse granulosa cells. Mice were induced to superovulate and relative gene expression determined at 8 h post-hCG as determined in Figure 1. Data are shown as mean ± SEM relative mRNA expression levels and significant differences determined by unpaired t-test, *P<0.01, **P<0.001.
Figure 5.
In vivo knockdown of RHOX8 results in reduced expression of insulin signaling in mouse granulosa cells. Mice were induced to superovulate and relative gene expression determined at 8 h post-hCG as determined in Figure 1. Data are shown as mean ± SEM relative mRNA expression levels and significant differences determined by unpaired t-test, *P<0.01, **P<0.001.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



