
Submitted:
02 March 2025
Posted:
03 March 2025
You are already at the latest version
Abstract
Rattus norvegicus (brown rat), a widely distributed rodent and common biomedical model, serves as a source of many zoonotic pathogens but is not traditionally recognized as a host for influenza A virus (IAV). To assess their susceptibility, we intranasally inoculated Sprague-Dawley rats with different IAV subtypes (H5Nx, H7N9, H9N2, H10N8 and the 2009 pandemic H1N1). All strains productively infected the rats, inducing seroconversion and mild weight reduction without overt clinical signs. Despite differences in replication, all viruses caused significant lung injury with a preferential upper respiratory tract tropism. Investigation of receptor distribution revealed α2,3-linked sialic acid (SA) dominance in the nasal turbinates and trachea, whereas α2,6-linked SA predominated in the lungs, although both receptors coexisted throughout the respiratory tract. This dual receptor expression aligns with the observed tissue-specific replication patterns and broad viral infectivity. Our results demonstrate that the rat is a permissive host for multiple IAV subtypes, challenging its exclusion from the IAV ecology. The asymptomatic yet pathogenic nature of infection, coupled with the global synanthropy of rats, underscores their potential role as cryptic reservoirs in viral maintenance and transmission. These findings highlight the need for expanded surveillance of rodents in influenza ecology to mitigate zoonotic risks.
Keywords:
zoonotic influenza
; host range
; interspecies transmission
; sialic acid receptor
; surveillance
1. Introduction
In recent decades, many influenza A viruses (IAVs) of zoonotic origin have emerged and caused subsequent epidemics or even pandemics in humans, such as the swine-origin 2009 H1N1 pandemic, and the avian-origin H7N9, H10N8, H9N2 and highly pathogenic avian influenza (HPAI) H5Nx outbreaks [1,2,3]. These events have repeatedly highlighted the ecological importance of animals in the genesis and transmission of IAVs.
IAVs have a very wide range of hosts and interspecies spillover is not uncommon in nature. Historically, IAVs have been established in birds, bats, horses, pigs, dogs and hu-mans [4,5]. Sporadic infections in other ungulates and carnivores (e.g., camels, dairy cows, whales, cats, minks, etc.) have also been documented [4,5,6,7,8,9]. While wild waterfowl are thought to serve as a natural reservoir for IAVs, domestic animals, either birds or mammals, may act as intermediate hosts, facilitating the adaptation and emergence of novel variants capable of infecting humans [4,5]. It is therefore crucial to monitor IAVs in different animal species and to better understand the ecology of influenza viruses.
Rodents, including commensal rats, can carry many different zoonotic pathogens but are not generally considered to be natural hosts of IAVs [4,5,10,11], although the presence of IAV nucleic acids or antibodies against viruses such as HPAI H5N1 and H5N6 have been reported [12,13,14,15,16,17]. On the other hand, although laboratory mice (Mus musculus) and rats (Rattus norvegicus) are widely used in biomedical research, and mice are popularly used in influenza research, rats were not generally considered a susceptible host for IAVs. There are limited reports on influenza infection in Rattus species, mostly using adapted viral strains [18,19,20], infant rat models [18,21,22,23], or other non-generalizable conditions [24,25].
Here we propose to experimentally infect rats with different subtypes of IAVs isolated from the field. In particular, the study will focus on prevalent IAV lineages with zoonotic risk and aims to provide direct evidence of the susceptibility of rats to IAVs without prior adaptation. As common synanthropes worldwide, rats are ubiquitous in both rural and urban areas with dense populations [26,27]. They not only share habitats with wild and free-ranging animals, but also have free access to human settlements and live-stock houses. Understanding the role of rats in the ecology of IAVs will aid risk assessment and disease control and prevention.
2. Materials and Methods
2.1. Ethical Statement
All animal experiments were approved and conducted in accordance with institutional guidelines. The design and conduct of the research were endorsed by the Medical Animal Care and Welfare Committee of Shantou University Medical College (Reference No. SUMC2013-111 and SUMC2016-034), and all experiments involving infectious agents were carried out in Biosafety Level 3 (BSL-3) facilities. The research also complied with the Guangdong Provincial Regulations on the Administration of Laboratory Animals and strictly followed the Operational Guidelines for Ethics Committees that Review Biomedical Research issued by the Ministry of Health of China.
2.2. Viruses
Viruses of prevalent and recently emerging subtypes in China were selected for this study. These viruses were initially isolated in 9-day-old embryonated chicken eggs. For avian strains, viral stocks were prepared by propagation in 9-day-old embryonated chicken eggs, while for human isolates, stocks were prepared in Madin-Darby canine kidney (MDCK) cells [28]. All stocks were aliquoted, titrated and frozen at -80°C until use. The representative strains used in this study are summarized in Table 1.
2.3. Animal Experiments
Female specific-pathogen-free (SPF) Sprague-Dawley (SD) rats, 8 to 9 weeks old, were purchased from Beijing Vital River Laboratory Animals Ltd. Co. (Beijing, China) and randomly divided into groups. Prior to inoculation and necropsy, animals were anesthetized by intramuscular injection of sodium pentobarbital (0.2 mg/g). Animals that lost more than 30% of their initial body weight were humanely euthanized and their respiratory tissues and blood were collected where possible.
To test the infectivity of different subtypes of IAVs in SD rats, two groups of three rats and one group of eight rats were inoculated with 100 μl of PBS containing 102, 104 and 106 50% tissue culture infectious dose (TCID50) of each virus strain, respectively. Eight rats were sham-infected with 100 μl PBS as a control. Five rats in the 106 TCID50 dose group were monitored in comparison to the control group (n=5) and weighed throughout the 14-day experimental period, while the remaining rats were necropsied on day 4 post-inoculation (dpi) to collect respiratory tract tissues for virus replication analysis. At 4 dpi, three rats in each virus dose group were euthanized for airway tissue collection. Each tissue was cut into two pieces. One part was placed in cryo-vials containing 1 ml of PBS, homogenized and stored at -80℃ for titration on MDCK cells. The remainder was fixed in 10% neutral buffered formalin for histopathological examination. Meanwhile, blood was also collected for serum preparation.
2.4. Hemagglutination Inhibition Test
Seroconversion was tested at 4 or 21 dpi before the euthanasia of the animals. Rat serum was treated with receptor-destroying enzyme II (RDE II; Denka Seiken, Tokyo, Japan) overnight at 37 °C to inactivate non-specific inhibitors, followed by heat-inactivation at 56 °C for 30 min to remove non-specific agglutinins by adsorption with freshly prepared turkey red blood cells (TRBCs). The sera obtained were tested by hemagglutination inhibition (HAI) assay using the IAVs listed in Table 1, according to the standard protocol of the World Health Organization (WHO) [28]. Briefly, four hemagglutination units per 25 μl of the IAVs listed in Table 1 were added to an equal volume of two-fold serially diluted sera in U-bottom 96-well microtiter plates and incubated for 45 min to 1 h. Finally, 50 μl of 0.5% TRBCs were added to the plates and incubated for 30 min at room temperature. HAI titers were defined as the reciprocal of the highest serum dilution required for complete hemagglutination inhibition.
2.5. Immunohistochemical and Histopathological Studies
Fresh nasal, tracheal and lung tissues were fixed in 10% (vol/vol) neutral buffered formalin for 48 h at room temperature. After fixation, the tissues were dehydrated in grad-ed ethanol solutions and xylene and embedded in paraffin. Finally, 3 μm tissue sections were obtained using a Leica slicer. Serial sections were subjected to standard immuno-histochemical (IHC) staining for viral nucleoprotein (NP) antigen detection and pathological examination by hematoxylin and eosin (H&E) staining as previously described [29,30].
2.6. Lectin Staining Assay
To determine the expression of sialic acid (SA) receptors in the airways of rats, we used biotinylated conjugated lectins from Maackia amurensis agglutinin I and II (MAA I and MAA II, specific for SAα2,3Galβ1,4GalNAc and SAα2,3Galβ1,3GalNAc, respectively; Vector Laboratories, Burlingame, California, USA) and Sambucus nigra agglutinin (SNA; Vector Laboratories, Burlingame, California, USA) to label α2,3-linked and α2,6-linked SA residues, respectively [31,32,33]. Tissues from non-inoculated SPF SD rats were processed using the same methods as described for immunohistochemical and histopathological studies. After blocking, sections were incubated with MAA (1:1500 dilution) or SNA (1:2000 dilution) for 1 h at room temperature. To ensure reliability, we also analyzed the distribution of IAV receptors in the respiratory tract of female 8- to 9-week-old C57BL/6 mice (Beijing Vital River Laboratory Animals Ltd. Co., Beijing, China).
2.7. Statistical Analysis
GraphPad Prism 10.1.0 (GraphPad Software) was used for calculations of area under the curve (AUC) and statistical analyses. Data are presented as mean ± standard deviation unless otherwise stated. Student’s t-test was used for comparisons between two groups, and one-way analysis of variance (ANOVA) was used to compare treatment groups. Statistical significance was set at P < 0.05.
3. Results
3.1. Asymptomatic Infection with IAVs in SD Rats
Clinical signs such as ruffled hair, inactivity, reduced food and water intake and other influenza-like illness were not observed in any of the rats. The body weight of the control rats increased steadily and there were no deaths in any of the inoculated groups (Figure 1). Following inoculation with different virus subtypes, rats in each group experienced no weight loss or a slight weight loss of less than 5% within the first day or two. They then gained weight from baseline over the observation period. However, some HPAI H5 strains (GY5096, SZ2400 and HZ4258) caused a brief weight loss at 8 or 9 dpi, immediately after a 5- or 6-day period of weight gain (Figure 1C,D,F). In contrast, rats in the other groups gained body weight at a stable rate over the 14-day period. Nevertheless, compared to the mock-inoculated control rats, none of the virus-inoculated groups exhibited significant changes in body weight during the infection (Figure 1L, P > 0.05). These results demonstrate that infection with either endemic or emerging IAVs, including HPAI H5 and HPAI H7 viruses, does not cause overt morbidity in the SD rat model, despite the subtle impairment in weight gain.
3.2. Replication and Distribution of Different IAVs in the Rat Respiratory Tract
To examine virus replication in the rat respiratory tract, groups of three rats were administered different doses of each virus (102, 104 and 106 TCID50) and necropsied at 4 dpi to determine the presence and titer of virus. At the 102 TCID50 dose, limited replication of CA7, the pandemic 2009 H1N1 (pdmH1N1) prototype virus, was detected in the nasal turbinate and trachea (Figure 2C). However, nasal viral replication increased to 104 TCID50/ml and 105 TCID50/ml in the groups challenged with 104 TCID50 and 106 TCID50 doses of CA7, respectively (Figure 2A,B). Among the HPAI H5 viruses, only VN1203 (H5N1 clade 1.0) was detected in the respiratory tract after inoculation with the low 102 TCID50 dose, and viral replication titers were low (< 102 TCID50/ml, Figure 2C). Even at the 104 TCID50 dose, only VN1203 and SP378 were detected in the airways (Figure 2B). These two viruses replicated at moderate levels, with the highest titers observed in the nasal turbinates of the rats. All HPAI H5 viruses were detected after inoculation with the 106 TCID50 dose (Figure 2A), but in general, the nasal titers of the H5 groups were lower compared to other subtypes. For H7N9, AH1 replicated to over 104 TCID50/ml in the nasal turbinate of the 102 TCID50 group, which was approximately two logs higher than SH2 and SP440, and was also detected in the lungs (Figure 2C). At other doses, nasal titers for SH2 and SP440 augmented remarkably with the increased inoculation doses, reaching a level closer to that of AH1 (Figure 2A,B). We observed that WZ598 (H9N2) and JX346 (H10N8) displayed comparable replicative capacity to SH2 or SP440 (H7N9) when inoculated at the same dose, and their nasal titers also elevated with increasing inoculation dose (Figure 2A–C). In summary, for most of the viruses tested, titers were much higher in the nasal turbinate than in the trachea and lung, indicating a preference for replication of these viruses in the upper respiratory tract of rats.
Blood samples were collected for serological testing when the rats were euthanized. We performed the HAI assay and observed successful seroconversion at 14 dpi after virus challenge in this rat model (Figure 3, Table S1), but we were unable to detect antibodies in the sera of rats euthanized at 4 dpi. Although the rats did produce antibodies against HPAI H5 and HPAI H7N9 viruses by 14 dpi, the titers were significantly lower than in groups infected with other viruses we tested. In the HPAI groups, all five rats inoculated with VN1203 (H5N1 clade 1.0) and SP440 (H7N9) became seropositive and produced antibodies with titers ranging from 40 to 320, which were lower than those observed for the low pathogenic pdmH1N1, H7N9, H9N2 and H10N8 (all above 320 in each individual). For the other HPAI H5 virus groups (i.e., clade 2.3.4.4 virus groups), some animals remained seronegative and antibodies in seropositive rats were at low or even marginal levels, with titers as low as 10 to 40.
Overall, our findings demonstrated that the viruses tested in our study were ready to infect rats without prior adaptation and replicated productively in this species, but their antibody production levels varied from strain to strain.
3.3. Pneumonia and Histopathological Changes Induced by IAVs in SD Rats
Pneumonia was not observed in the control group (Figure 3D). Although the gross view of the excised lungs appeared normal and no visible lesions were found in the inoculated rats, H&E staining revealed obvious lung injury and comparable pathological changes among the different virus groups at the same dose (Figure 3B,E–O), but distinct between the different doses (Figure 3A–C). The virus-infected rat lungs exhibited typical histopathological changes compared to the mock controls. Alveolitis, widened alveolar septum, inflammatory infiltration, and erythrocyte extravasation were commonly observed across all three doses (Figure 3). At the 102 TCID50 dose, rat pneumonia was relatively mild, and intact alveolar cells could still be observed, but there was obvious widening of the alveolar septum (Figure 3A). In the other two higher dose groups, alveolar cells were destroyed, and there was marked exudation due to inflammation (Figure 3B,C,E–O). Alveolar fusion and markedly thickened alveolar walls, necrosis, and loss of bronchial epithelial cells were also observed (Figure 3). Rats inoculated with 106 TCID50 IAVs displayed the most severe pneumonia, with pulmonary consolidation occurring in most parts of the lung, characterized by complete filling of the alveolar space with exudates and inflammatory factors instead of alveolar air (Figure 3C). To provide a comprehensive picture of the IAV-induced pneumonia in rats, here we shown the representative pathological changes in the lungs of the 104 TCID50 groups for all viruses (Figure 3E–O). Collectively, all IAVs tested here were able to elicit an inflammatory response and lung injury in SD rats, indicating a productive infection with pathogenicity caused by these viruses.
To confirm productive infection and the presence of IAVs in the respiratory tract of SD rats, we performed IHC staining to detect viral NP antigen in the nasal turbinate and lung tissues. Staining for IAV NP revealed significant differences between viruses. In the nasal turbinate, substantial NP antigen from pdmH1N1 was observed in rats inoculated with CA7 at the dose of 106 TCID50 (Figure 4A). Although we had challenged groups of SD rats with different virus doses, only a few NP proteins from the HZ4258 group were detected in one rat in the 106 group among the five HPAI H5 virus strains (Figure 4B, green arrows). Conversely, NP antigens of all H7N9 strains were detected in the nasal turbinate after inoculation (Figure 4C–J). AH1 antigens were detected in the nasal turbinate of the 102 and 104 TCID50 groups (Figure 4C,D), but not in the 106 TCID50 group. For the other two H7N9 viruses, SH2 and SP440, viral antigens were present in all three groups, as shown in Figure 4E–G and Figure 4H–J, respectively. In the H9N2 and H10N8 groups, viral NP from WZ598 and JX346 were detected in nasal tissues from the 106 TCID50 group (Figure 4K) and the 104 TCID50 group (Figure 4L), respectively. On the other hand, our results revealed negative staining for viral NP in the lungs of most groups (Figure 4M). In fact, only NP antigens from WZ598 (H9N2) and AH1 (H7N9) were recorded in a single rat of each group (Figure 4O,N). In the H9N2-positive lung section, bronchiolar epithelial cells were focally stained positive for WZ598 in one rat from the 104 TCID50 group (Figure 4O). For H7N9, stained antigens were only observed in the lung of one rat inoculated with 102 TCID50 of AH1 (Figure 4N), although each H7N9 strain was detected in the lung tissues as indicated previously by virus titration (Figure 2B). Of the NP-positive nasal and lung sections, we found that viral NP was mainly distributed in the ciliated mucosal epithelial cells.
Taken together, our immunohistochemical staining results revealed a predominant distribution of viral NPs in the rat nasal cavity, consistent with the replication pattern of higher viral titers in the nasal turbinate compared to the lungs. These findings suggest that the IAVs tested tend to replicate preferentially in the upper respiratory tracts of SD rats, providing further evidence of viral replication and pathogenesis.
3.4. Expression of Sialic Acid-Linked Receptors in the Respiratory Tract of SD Rats
The attachment and entry of IAVs into host cells are dependent on the presence of specific SA-linked receptors. To investigate the expression of these receptors in the airways of rodents, we first examined the presence of α2,3- and α2,6-linked SA receptors in the airways of mice, using positive staining for both MAAs and SNA (Figure S1). In contrast to the weak MAA II staining observed in mice (Figure S1B, S1E, S1H), SD rats exhibited significantly stronger expression of α2,3-linked SA throughout the upper and lower respiratory tracts (Figure 6B,E,H). This indicates a notable difference in the presence of SAα2,3Galβ1,3GalNAc in the airways between these two rodent species, as MAA II is specific for the detection of this “traditional” avian influenza receptor [31].
In the nasal turbinate of SD rats (Figure 6A–C), both MAAs and SNA bound to ciliated epithelial cells, indicating the presence of α2,3- and α2,6-linked SA receptors, respectively. However, MAAs (Figure 6A,B) exhibited much denser staining in these nasal epithelial cells compared to SNA (Figure 6C). Additionally, cells in the basal and connective tissues were exclusively stained by MAAs but not by SNA, suggesting the presence of α2,3-linked SA receptors and the absence of α2,6-linked SA receptors in these areas. In the lower respiratory tract, staining for both MAA isoforms was observed in the trachea of rats (Figure 6D,E) and mice (Figure S1D,E). However, we found less SNA-positive staining in the epithelial cells of the rat trachea (Figure 6F) compared to MAAs, and there was minimal staining noted in mice (Figure S1F). In the lungs of SD rats (Figure 6G–I), α2,6-linked SA receptors were more abundant in the bronchi and bronchioles, as indicated by denser SNA staining compared to MAAs. In the alveoli, the staining patterns of MAA I and SNA were similar due to their diffuse distribution, although MAA II staining was significantly reduced. These results demonstrate that α2,3-linked SA receptors are predominantly expressed in the nasal turbinate, trachea and alveoli, whereas α2,6-linked SA receptors are more prevalent in the lungs of the SD rat model. This differential expression may provide insight into the mechanisms underlying the variable replication and transmission of IAVs in this model.
4. Discussion
The role of rats and other rodents in the influenza ecology is understudied and controversial. The present study provides compelling experimental evidence that SD rats, a representative model of the Rattus species, are susceptible to productive infection by diverse subtypes of contemporary IAVs that pose a major threat to both public health and agriculture, including avian HPAI H5Nx (clades 1.0 and 2.3.4.4a, b, e, g, either human or avian isolates), H7N9 (HPAI and LPAI), H9N2, H10N8, and the mammalian-adapted pandemic 2009 H1N1 viruses (Table 1). Notably, these infections occurred without prior viral adaptation, challenging the conventional assumptions that rats are refractory to IAV infection. Our findings, together with other previous reports [11,12,13,14,15,16,17,34,35,36,37], underscores the need to re-evaluate rodents as potential reservoirs, mechanical vectors, or bridging hosts in the zoonotic transmission of IAVs. The absence of overt clinical signs, despite robust viral replication, seroconversion and histopathological evidence of lung injury (Figure 1, Figure 2, Figure 3, Figure 4 and Figure 5), positions rats as cryptic carriers capable of sustaining IAV infections undetected in natural settings.
A striking feature of IAV-infected SD rats is the dissociation between their subclinical manifestations and significant virological and immunological findings. While mice and ferrets develop observable disease or mortality following experimental IAV challenge [29,30], rats show only transient weight fluctuations (Figure 1) and no influenza-like symptoms, even when infected with HPAI H5 or H7N9 strains. This asymptomatic phenotype is similar to that observed in wild waterfowl, which are natural reservoirs of IAV [4,5]. The absence of disease presentation and the induction of seroconversion in most rats suggests that rats may use effective immune mechanisms to mitigate systemic spread of the virus while permitting local replication in the upper respiratory tract [this study and 19, 20, 34]. This balance may facilitate viral persistence without compromising host survival, allowing rats to act as stealth vectors in ecosystems where they interact with domestic animals, wildlife and humans.
The receptor distribution landscape has been previously reported in BABL/c and C57BL/6J mice and in mouse tracheal epithelial cells (mTEDC) [32,33]. To our knowledge, here we provide the first report on the distribution of α2,3- and α2,6-linked SA in the respiratory tracts of SD rats (Figure 6). The preferential replication of IAVs in the nasal turbinate, as evidenced by higher viral titers and NP antigen localization (Figure 2 and Figure 5), highlights a distinct tissue tropism in rats. This pattern correlates with the predominance of α2,3-linked SA receptors in the upper respiratory tract (Figure 6), which are preferentially recognized by avian-adapted IAVs. However, the concurrent detection of both α2,3- and α2,6-linked SA receptors in the rat lung suggests dual receptor expression that could permit infection of the lower respiratory tract, as observed with H7N9, H9N2, H10N8 and some H5 strains, such as the human isolates VN1203 and SP378 (Figure 2). The discordance between SA receptor distribution (α2,6-SA dominance in the lung with MAA I-positive staining of α2,3-SA) and relatively low pulmonary viral titers (especially for the mammalian-adapted strains like pdmH1N1) implies that additional host factors, such as mucin barriers, antiviral interferon responses, protease availability, and other unknown mechanisms may restrict systemic dissemination. This receptor-replication mismatch warrants further investigation and caution when extrapolating murine data to other rodents, particularly given the contrasting SA expression profiles in mice, which exhibit much weaker α2,3-SA expression compared to rats.
Although viable virus was detected for all virus strains, our results shown significant differences in replication efficiency in rats between IAV strains. This disparity may reflect subtype-, clade- or strain-specific adaptations. Notably, all strains with G57 H9N2-like internal genes (i.e., SZ378, AH1, SH2, SP440, WZ598 and JX346) [38,39,40,41,42,43] replicated robustly in the upper respiratory tract of rats. This may indicate a growth advantage conferred by this specific internal gene cassette. Of note, the efficient replication of H9N2, an enzootic subtype in Eurasia with a broad host range and pandemic potential, and its derivatives suggests that rats may contribute to the evolution of the related viruses. The predominance of the G57 H9N2 genotype in China [38], which is associated with human infections and the emergence of novel avian strains (such as H7N9/H7N7 [39,40], H10N8/H10N6 [41,42], H5N6 [43] and H3N8 [44]), further underlies this risk.
Despite the absence of clinical disease, all IAVs induced significant pulmonary pathology characterized by alveolitis, inflammatory infiltration, and epithelial necrosis (Figure 4). The dose-dependent severity of lung injury implies the potential for subclinical infections to cause covert tissue damage that could predispose rats to secondary infections or alter their respiratory dynamics in natural settings. Of particular concern is the capacity of clade 2.3.4.4 HPAI H5 viruses to replicate in rats without adaptation, albeit with variable viral titers. Although these strains exhibited relatively low nasal titers compared to H7N9 or H9N2 viruses, their detection in the respiratory tract raises questions about the risk of rat-mediated viral persistence or reassortment in regions where HPAI circulates enzootically. Although we have not tested the currently devastating H5N1 clade 2.3.4.4b strains circulating in the Americas, the Animal and Plant Health Inspection Service recently detected this HPAI virus in several black rats (Rattus rattus) in Riverside, California during February 2025, alongside numerous prior reports of infections in deer mice (Peromyscus maniculatus) and house mice (Mus musculus) across the United States [45]. These findings collectively underscore the ever-expanding host spectrum of HPAI H5N1 and emphasize the underappreciated role of murine rodents in influenza ecology, highlighting the critical need to investigate their capacity to sustain viral transmission in natural settings.
The ubiquity of rats in human-dominated ecosystems amplifies their potential role as intermediaries in IAV transmission cycles. As synanthropes, rats have free access to poultry and livestock farms, wet markets, wildlife, and urban garbage dumps, creating opportunities for cross-species virus exchange. Their preferential replication in the upper respiratory and shedding of virus in nasal secretions could facilitate environmental contamination, enabling mechanical transmission to susceptible hosts. Animals that prey on rats or scavenge carcasses, including but not limited to cats, dogs and other carnivores, may also be infected. Although IAV transmission by rats has not been confirmed, animal infections with IAVs through exposure to contaminated environments via virus-laden fomites and consumption of contaminated food and water have been recorded in cats, chickens, mice and SD rats naturally or experimentally [7,35,45,46,47,48,49].
It is also worth noting that the coexistence of α2,3- and α2,6-SA receptors raises the possibility that rats may act as “mixing vessels” for reassortment, particularly when co-infected with avian and human IAVs. While no natural reassortants have been reported in rats, the detection of IAV nucleic acids in wild Rattus norvegicus populations (11.04% in Boston, Massachusetts and 3.47% in southern China) [14,15] suggests ongoing spillover events that merit closer surveillance. Furthermore, the asymptomatic nature of infection complicates surveillance efforts, necessitating active molecular or serological screening in wild rat populations to assess their contribution to IAV maintenance.
This study focused on a laboratory rat model under controlled conditions that may not fully replicate the physiological or immunological state of wild rats. Virus replication outside the respiratory tract was not explored. Natural routes of exposure (e.g., oral, aerosol, fomite, etc.) and co-infections with other pathogens could alter infection outcomes. Additionally, the lack of transmission experiments leaves unresolved whether rats can horizontally transmit IAVs from rat to rat or to the other mammals—a critical question for evaluating their role in viral ecology. While receptor binding specificity has been inferred from previous studies, direct characterization of viral hemagglutinin affinity for rat SA receptors is also needed to clarify tropism mechanisms. Future studies should prioritize in vivo transmission assays, comparative receptor profiling in various wild rodents, and investigations into the molecular determinants of viral fitness in rat respiratory tissues. Additional strains could be included in the experiments to further expands our understanding of IAV infections in this animal, and systematic screening of wild rat populations for IAV RNA and antibodies, particularly in regions with enzootic H5, H7, H9 or H10 viruses, is also warranted to assess the role of rats in influenza A virus transmission and evolution.
5. Conclusions
This study redefines Rattus norvegicus as a permissive host for multiple IAV subtypes prevalent in birds or humans, and highlights its ability to sustain subclinical infections with potential ecological consequences. The convergence of broad viral susceptibility, synanthropic behavior, and dual SA receptor expression in the respiratory tracts positions rats as underrecognized players in influenza ecology. While their role as “mixing vessels” remains speculative, the risk of environmental virus amplification and spillover to domestic animals or humans cannot be dismissed. Strengthening surveillance in rodent populations and integrating rats into One Health frameworks will be essential for mitigating zoonotic threats in an era of escalating avian influenza activity.
Supplementary Materials
Author Contributions
Conceptualization, H.Z.; methodology, L.L., R.C., Y.G. and H.Z.; validation, L.L. and H.Z.; formal analysis, L.L.; investigation, L.L., R.C., Z.Y., Q.C. and H.Z.; resources, Y.G. and H.Z.; data curation, L.L.; writing—original draft preparation, L.L. and H.Z.; writing—review and editing, L.L. and H.Z.; visualization, L.L. and H.Z.; supervision, H.Z.; project administration, H.Z.; funding acquisition, Y.G. and H.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Hong Kong Research Grant Council, grant numbers 17108417, 17118515, T11-705/14-N and T11-712/19-N; National Institutes of Health, contract numbers HHSN272201400006C and 75N93021C00016 and Li Ka Shing Foundation. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the article.
Institutional Review Board Statement
The animal study protocol was approved by the Medical Animal Care and Welfare Committee of Shantou University Medical College (protocol codes SUMC2013-111, 3 April 2013 and SUMC2016-034, 24 March 2016).
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article and supplementary material. Further inquiries can be directed to the corresponding authors.
Acknowledgments
We gratefully acknowledge all the staff of the Guangdong-Hong Kong Joint Laboratory of Emerging Infectious Diseases/ Joint Laboratory for International Collaboration in Virology and Emerging Infectious Diseases/ Joint Institute of Virology (STU/HKU) and SKLEID for their technical support and administrative assistance. This article is a revised and expanded version of a poster entitled “Experimental infection of rats with influenza A viruses: Implications for murine rodents in influenza A virus ecology”, which was presented at 3rd Annual CEIRR Network Meeting, New York Academy of Medicine, New York, July 21-24, 2024 [50].
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| IAV | influenza A virus |
| HPAI | highly pathogenic avian influenza |
| LPAI | low pathogenic avian influenza |
| BSL-3 | Biosafety Level 3 |
| MDCK | Madin-Darby canine kidney |
| CA7 | A/California/07/2009 |
| VN1203 | A/Vietnam/1203/2004 |
| GY5096 | A/Duck/Guiyang/5096/2013 |
| SZ2400 | A/Chicken/Shenzhen/2400/2013 |
| SP378 | A/Shenzhen/SP378/2015 |
| HZ4258 | A/Duck/Huzhou/4258/2013 |
| AH1 | A/Anhui/1/2013 |
| SH2 | A/Shanghai/2/2013 |
| SP440 | A/Guangdong/SP440/2017 |
| WZ598 | A/Chicken/Wenzhou/598/2013 |
| JX346 | A/Jiangxi-Donghu/346/2013 |
| SPF | specific-pathogen-free |
| SD | Sprague-Dawley |
| TCID50 | 50% tissue culture infectious dose |
| dpi | day(s) post-inoculation |
| RDE | receptor-destroying enzyme |
| HAI | hemagglutination inhibition |
| TRBC | turkey red blood cell |
| WHO | World Health Organization |
| IHC | immuno-histochemical |
| H&E | hematoxylin and eosin |
| NP | nucleoprotein |
| SA | sialic acid |
| MAA | Maackia amurensis agglutinin |
| SNA | Sambucus nigra agglutinin |
| AUC | area under the curve |
| ANOVA | one-way analysis of variance |
| pdmH1N1 | pandemic 2009 H1N1 |
| mTEDC | mouse tracheal epithelial cells |
| RNA | ribonucleic acid |
References
- Smith, G.J.; Vijaykrishna, D.; Bahl, J.; Lycett, S.J.; Worobey, M.; Pybus, O.G.; Ma, S.K.; Cheung, C.L.; Raghwani, J.; Bhatt, S.; et al. Origins and evolutionary genomics of the 2009 swine-origin H1N1 influenza A epidemic. Nature 2009, 459, 1122–1125. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Cao, B.; Hu, Y.; Feng, Z.; Wang, D.; Hu, W.; Chen, J.; Jie, Z.; Qiu, H.; Xu, K.; et al. Human infection with a novel avian-origin influenza A (H7N9) virus. N Engl J Med 2013, 368, 1888–1897. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Global Influenza Programme. Available online: https://www.who.int/teams/global-influenza-programme/avian-influenza/monthly-risk-assessment-summary (accessed on 27 February, 2025).
- Webster, R.G.; Bean, W.J.; Gorman, O.T.; Chambers, T.M.; Kawaoka, Y. Evolution and ecology of influenza A viruses. Microbiol Rev 1992, 56, 152–179. [Google Scholar] [CrossRef]
- Yoon, S.W.; Webby, R.J.; Webster, R.G. Evolution and ecology of influenza A viruses. Curr Top Microbiol Immunol 2014, 385, 359–375. [Google Scholar] [CrossRef]
- Chu, D.K.W.; Perera, R.; Ali, A.; Oladipo, J.O.; Mamo, G.; So, R.T.Y.; Zhou, Z.; Chor, Y.Y.; Chan, C.K.; Belay, D.; et al. Influenza A Virus Infections in Dromedary Camels, Nigeria and Ethiopia, 2015-2017. Emerg Infect Dis 2020, 26, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Cruz, C.D.; Icochea, M.E.; Espejo, V.; Troncos, G.; Castro-Sanguinetti, G.R.; Schilling, M.A.; Tinoco, Y. Highly Pathogenic Avian Influenza A(H5N1) from Wild Birds, Poultry, and Mammals, Peru. Emerg Infect Dis 2023, 29, 2572–2576. [Google Scholar] [CrossRef] [PubMed]
- Mok, C.K.P.; Qin, K. Mink infection with influenza A viruses: an ignored intermediate host? One Health Adv 2023, 1, 5. [Google Scholar] [CrossRef]
- Burrough, E.R.; Magstadt, D.R.; Petersen, B.; Timmermans, S.J.; Gauger, P.C.; Zhang, J.; Siepker, C.; Mainenti, M.; Li, G.; Thompson, A.C.; et al. Highly Pathogenic Avian Influenza A(H5N1) Clade 2.3.4.4b Virus Infection in Domestic Dairy Cattle and Cats, United States, 2024. Emerg Infect Dis 2024, 30, 1335–1343. [Google Scholar] [CrossRef]
- Velkers, F.C.; Blokhuis, S.J.; Veldhuis Kroeze, E.J.B.; Burt, S.A. The role of rodents in avian influenza outbreaks in poultry farms: a review. Vet Q 2017, 37, 182–194. [Google Scholar] [CrossRef]
- Strand, T.M.; Lundkvist, Å. Rat-borne diseases at the horizon. A systematic review on infectious agents carried by rats in Europe 1995-2016. Infect Ecol Epidemiol 2019, 9, 1553461. [Google Scholar] [CrossRef]
- Shortridge, K.F.; Gao, P.; Guan, Y.; Ito, T.; Kawaoka, Y.; Markwell, D.; Takada, A.; Webster, R.G. Interspecies transmission of influenza viruses: H5N1 virus and a Hong Kong SAR perspective. Vet Microbiol 2000, 74, 141–147. [Google Scholar] [CrossRef]
- El-Sayed, A.; Prince, A.; Fawzy, A.; Nadra, E.; Abdou, M.I.; Omar, L.; Fayed, A.; Salem, M. Sero-prevalence of avian influenza in animals and human in Egypt. Pak J Biol Sci 2013, 16, 524–529. [Google Scholar] [CrossRef] [PubMed]
- Cummings, C.O.; Hill, N.J.; Puryear, W.B.; Rogers, B.; Mukherjee, J.; Leibler, J.H.; Rosenbaum, M.H.; Runstadler, J.A. Evidence of Influenza A in Wild Norway Rats (Rattus norvegicus) in Boston, Massachusetts. Front Ecol Evol 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Gao, Y.; Wen, Y.; Ke, X.; Ou, Z.; Li, Y.; He, H.; Chen, Q. Detection of Virus-Related Sequences Associated With Potential Etiologies of Hepatitis in Liver Tissue Samples From Rats, Mice, Shrews, and Bats. Front Microbiol 2021, 12, 653873. [Google Scholar] [CrossRef]
- Camp, J.V.; Desvars-Larrive, A.; Nowotny, N.; Walzer, C. Monitoring Urban Zoonotic Virus Activity: Are City Rats a Promising Surveillance Tool for Emerging Viruses? Viruses 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.W.; Zhang, X.L.; Sun, J.; Liu, H.; Chen, J.M. Infection of wild rats with H5N6 subtype highly pathogenic avian influenza virus in China. J Infect 2023, 86, e117–e119. [Google Scholar] [CrossRef]
- Mahmud, M.I.; Maassab, H.F.; Jennings, R.; Potter, C.W. Influenza virus infection of a newborn rats: virulence of recombinant strains prepared from a cold-adapted, attenuated parent. Arch Virol 1979, 61, 207–216. [Google Scholar] [CrossRef]
- Daniels, M.J.; Selgrade, M.K.; Doerfler, D.; Gilmour, M.I. Kinetic profile of influenza virus infection in three rat strains. Comp Med 2003, 53, 293–298. [Google Scholar]
- Marchenko, V.; Zelinskaya, I.; Toropova, Y.; Shmakova, T.; Podyacheva, E.; Lioznov, D.; Zhilinskaya, I.N. Influenza A Virus Causes Histopathological Changes and Impairment in Functional Activity of Blood Vessels in Different Vascular Beds. Viruses 2022, 14. [Google Scholar] [CrossRef]
- Michaels, R.H.; Mahmud, M.I.; Coup, A.J.; Jennings, R.; Potter, C.W. Influenza virus infection in newborn rats: a possible marker of attenuation for man. J Med Virol 1978, 2, 253–264. [Google Scholar] [CrossRef]
- Jennings, R.; Potter, C.W.; Teh, C.Z.; Mahmud, M.I. The replication of type A influenza viruses in the infant rat: a marker for virus attenuation. J Gen Virol 1980, 49, 343–354. [Google Scholar] [CrossRef]
- Teh, C.; Jennings, R.; Potter, C.W. Influenza virus infection of newborn rats: virulence of recombinant strains prepared from influenza virus strain A/Okuda/57. J Med Microbiol 1980, 13, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Dhur, A.; Galan, P.; Hannoun, C.; Huot, K.; Hercberg, S. Effects of iron deficiency upon the antibody response to influenza virus in rats. J Nutr Biochem 1990, 1, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Gaciong, Z.; Alexiewicz, J.M.; Massry, S.G. Impaired in vivo antibody production in CRF rats: role of secondary hyperparathyroidism. Kidney Int 1991, 40, 862–867. [Google Scholar] [CrossRef]
- McKinney, M.L. Urbanization as a major cause of biotic homogenization. Biological Conservation 2006, 127, 247–260. [Google Scholar] [CrossRef]
- Feng, A.Y.T.; Himsworth, C.G. The secret life of the city rat: a review of the ecology of urban Norway and black rats (Rattus norvegicus and Rattus rattus). Urban Ecosystems 2014, 17, 149–162. [Google Scholar] [CrossRef]
- WHO Global Influenza Surveillance Network. Manual for the laboratory diagnosis and virological surveillance of influenza. Available online: https://www.who.int/publications/i/item/manual-for-the-laboratory-diagnosis-and-virological-surveillance-of-influenza (accessed on 27 February, 2025).
- Zhu, H.; Wang, J.; Wang, P.; Song, W.; Zheng, Z.; Chen, R.; Guo, K.; Zhang, T.; Peiris, J.S.; Chen, H.; et al. Substitution of lysine at 627 position in PB2 protein does not change virulence of the 2009 pandemic H1N1 virus in mice. Virology 2010, 401, 1–5. [Google Scholar] [CrossRef]
- Zhu, H.; Wang, D.; Kelvin, D.J.; Li, L.; Zheng, Z.; Yoon, S.W.; Wong, S.S.; Farooqui, A.; Wang, J.; Banner, D.; et al. Infectivity, transmission, and pathology of human-isolated H7N9 influenza virus in ferrets and pigs. Science 2013, 341, 183–186. [Google Scholar] [CrossRef]
- Ibricevic, A.; Pekosz, A.; Walter, M.J.; Newby, C.; Battaile, J.T.; Brown, E.G.; Holtzman, M.J.; Brody, S.L. Influenza virus receptor specificity and cell tropism in mouse and human airway epithelial cells. J Virol 2006, 80, 7469–7480. [Google Scholar] [CrossRef]
- Nicholls, J.M.; Bourne, A.J.; Chen, H.; Guan, Y.; Peiris, J.S. Sialic acid receptor detection in the human respiratory tract: evidence for widespread distribution of potential binding sites for human and avian influenza viruses. Respir Res 2007, 8, 73. [Google Scholar] [CrossRef]
- Ning, Z.Y.; Luo, M.Y.; Qi, W.B.; Yu, B.; Jiao, P.R.; Liao, M. Detection of expression of influenza virus receptors in tissues of BALB/c mice by histochemistry. Vet Res Commun 2009, 33, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Shortridge, K.F.; Zhou, N.N.; Guan, Y.; Gao, P.; Ito, T.; Kawaoka, Y.; Kodihalli, S.; Krauss, S.; Markwell, D.; Murti, K.G.; et al. Characterization of avian H5N1 influenza viruses from poultry in Hong Kong. Virology 1998, 252, 331–342. [Google Scholar] [CrossRef]
- Achenbach, J.E.; Bowen, R.A. Transmission of avian influenza A viruses among species in an artificial barnyard. PLoS One 2011, 6, e17643. [Google Scholar] [CrossRef] [PubMed]
- Hiono, T.; Okamatsu, M.; Yamamoto, N.; Ogasawara, K.; Endo, M.; Kuribayashi, S.; Shichinohe, S.; Motohashi, Y.; Chu, D.H.; Suzuki, M.; et al. Experimental infection of highly and low pathogenic avian influenza viruses to chickens, ducks, tree sparrows, jungle crows, and black rats for the evaluation of their roles in virus transmission. Vet Microbiol 2016, 182, 108–115. [Google Scholar] [CrossRef] [PubMed]
- VanDalen, K.K.; Nemeth, N.M.; Thomas, N.O.; Barrett, N.L.; Ellis, J.W.; Sullivan, H.J.; Franklin, A.B.; Shriner, S.A. Experimental infections of Norway rats with avian-derived low-pathogenic influenza A viruses. Arch Virol 2019, 164, 1831–1836. [Google Scholar] [CrossRef]
- Pu, J.; Wang, S.; Yin, Y.; Zhang, G.; Carter, R.A.; Wang, J.; Xu, G.; Sun, H.; Wang, M.; Wen, C.; et al. Evolution of the H9N2 influenza genotype that facilitated the genesis of the novel H7N9 virus. Proc Natl Acad Sci U S A 2015, 112, 548–553. [Google Scholar] [CrossRef]
- Lam, T.T.; Wang, J.; Shen, Y.; Zhou, B.; Duan, L.; Cheung, C.L.; Ma, C.; Lycett, S.J.; Leung, C.Y.; Chen, X.; et al. The genesis and source of the H7N9 influenza viruses causing human infections in China. Nature 2013, 502, 241–244. [Google Scholar] [CrossRef]
- Lam, T.T.; Zhou, B.; Wang, J.; Chai, Y.; Shen, Y.; Chen, X.; Ma, C.; Hong, W.; Chen, Y.; Zhang, Y.; et al. Dissemination, divergence and establishment of H7N9 influenza viruses in China. Nature 2015, 522, 102–105. [Google Scholar] [CrossRef]
- Ma, C.; Lam, T.T.; Chai, Y.; Wang, J.; Fan, X.; Hong, W.; Zhang, Y.; Li, L.; Liu, Y.; Smith, D.K.; et al. Emergence and evolution of H10 subtype influenza viruses in poultry in China. J Virol 2015, 89, 3534–3541. [Google Scholar] [CrossRef]
- Xiao, C.; Ma, W.; Sun, N.; Huang, L.; Li, Y.; Zeng, Z.; Wen, Y.; Zhang, Z.; Li, H.; Li, Q.; et al. PB2-588 V promotes the mammalian adaptation of H10N8, H7N9 and H9N2 avian influenza viruses. Sci Rep 2016, 6, 19474. [Google Scholar] [CrossRef]
- Bi, Y.; Chen, Q.; Wang, Q.; Chen, J.; Jin, T.; Wong, G.; Quan, C.; Liu, J.; Wu, J.; Yin, R.; et al. Genesis, Evolution and Prevalence of H5N6 Avian Influenza Viruses in China. Cell Host Microbe 2016, 20, 810–821. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Sun, H.; Gao, F.; Luo, K.; Huang, Z.; Tong, Q.; Song, H.; Han, Q.; Liu, J.; Lan, Y.; et al. Human infection of avian influenza A H3N8 virus and the viral origins: a descriptive study. Lancet Microbe 2022, 3, e824–e834. [Google Scholar] [CrossRef]
- Animal and Plant Health Inspection Service, U.S.; Department of Agriculture. Detections of Highly Pathogenic Avian Influenza in Mammals. Available online: https://www.aphis.usda.gov/livestock-poultry-disease/avian/avian-influenza/hpai-detections/mammals (accessed on 27 February, 2025).
- Kuiken, T.; Rimmelzwaan, G.; van Riel, D.; van Amerongen, G.; Baars, M.; Fouchier, R.; Osterhaus, A. Avian H5N1 influenza in cats. Science 2004, 306, 241. [Google Scholar] [CrossRef] [PubMed]
- Songserm, T.; Amonsin, A.; Jam-on, R.; Sae-Heng, N.; Meemak, N.; Pariyothorn, N.; Payungporn, S.; Theamboonlers, A.; Poovorawan, Y. Avian influenza H5N1 in naturally infected domestic cat. Emerg Infect Dis 2006, 12, 681–683. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; Xu, L.; Zhu, H.; Deng, W.; Chen, T.; Lv, Q.; Li, F.; Yuan, J.; Xu, Y.; Huang, L.; et al. Transmission of H7N9 influenza virus in mice by different infective routes. Virol J 2014, 11, 185. [Google Scholar] [CrossRef]
- Jones, J.C.; Sonnberg, S.; Webby, R.J.; Webster, R.G. Influenza A(H7N9) virus transmission between finches and poultry. Emerg Infect Dis 2015, 21, 619–628. [Google Scholar] [CrossRef]
- Li, L.; Chen, R.; Yan, Z. , Cai, Q.; Guan, Y., Zhu, H. Experimental infection of rats with influenza A viruses: Implications for murine rodents in in-fluenza A virus ecology. In Abstract Book, 3rd Annual CEIRR Network Meeting, New York Academy of Medicine, 1216 5th Ave, New York, NY 10029, July 21-24, 2024. [Google Scholar]
Figure 1.
Changes in body weight of Sprague-Dawley (SD) rats inoculated with selected influenza A viruses (see Table 1). To evaluate the pathogenicity of different influenza viruses in rats, 8- to 9-week-old SD rats were inoculated with 106 TCID50 of the indicated viruses (A-K). The animals were weighed 1 to 14 days after inoculation. There were no deaths in any group during this period. Data presented are mean changes of body weight ± standard deviation of five individual rats for the control (grey) and five for each virus-inoculated group (blue). The area under the curve (AUC) for each group was calculated using GraphPad Prism 10.1.0 (L). One-way analysis of variance (ANOVA) of the group-specific AUC and pairwise t-tests between the virus-inoculated groups and the control suggest no statistical significance in the changes in body weight (P > 0.05).
Figure 1.
Changes in body weight of Sprague-Dawley (SD) rats inoculated with selected influenza A viruses (see Table 1). To evaluate the pathogenicity of different influenza viruses in rats, 8- to 9-week-old SD rats were inoculated with 106 TCID50 of the indicated viruses (A-K). The animals were weighed 1 to 14 days after inoculation. There were no deaths in any group during this period. Data presented are mean changes of body weight ± standard deviation of five individual rats for the control (grey) and five for each virus-inoculated group (blue). The area under the curve (AUC) for each group was calculated using GraphPad Prism 10.1.0 (L). One-way analysis of variance (ANOVA) of the group-specific AUC and pairwise t-tests between the virus-inoculated groups and the control suggest no statistical significance in the changes in body weight (P > 0.05).
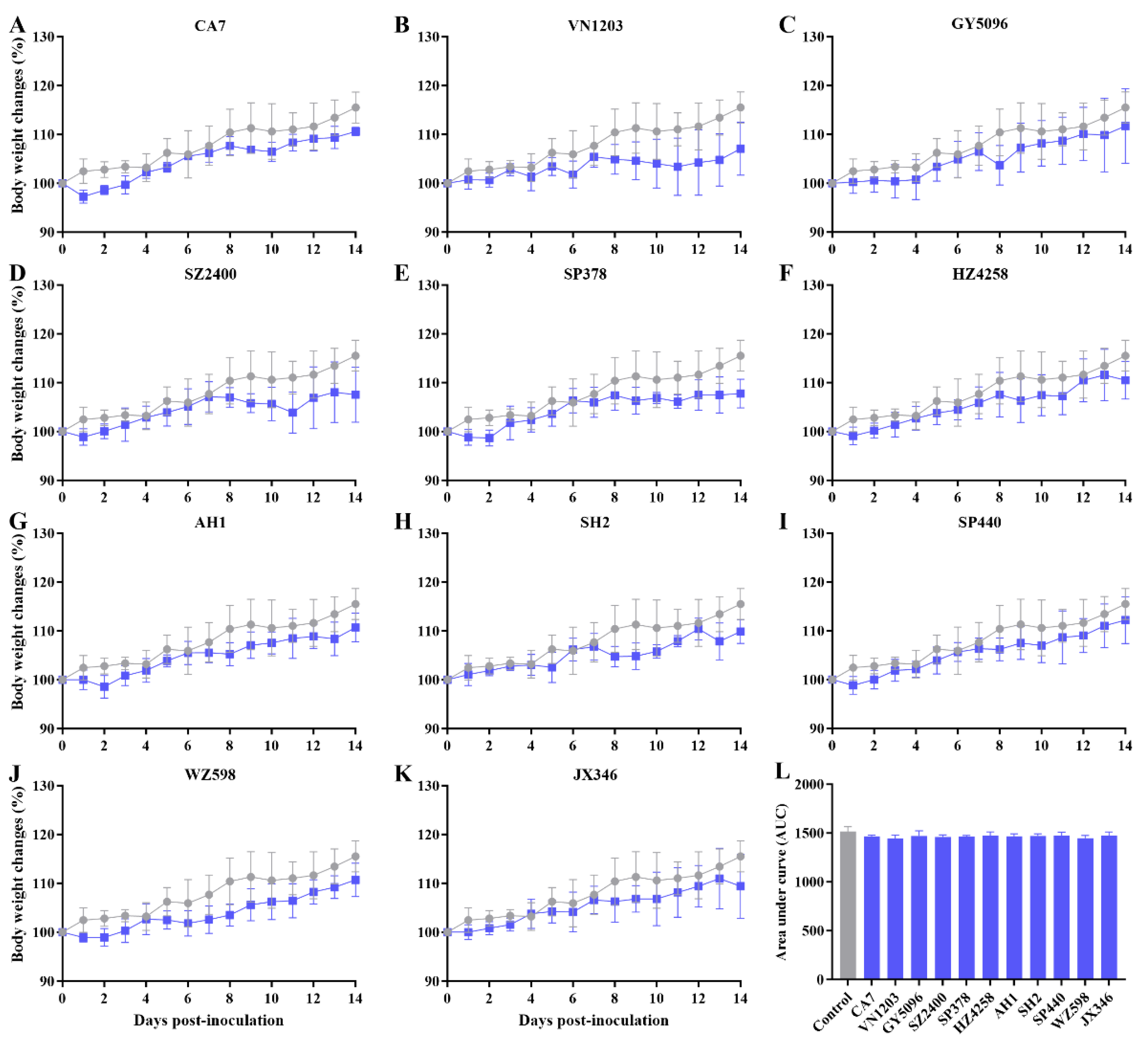
Figure 2.
Replication of selected influenza A viruses in the respiratory tract of SD rats. To evaluate the infectivity of different influenza viruses (see Table 1) in rats, 8- to 9-week-old SD rats were inoculated with doses of 106 (A), 104 (B) and 102 (C) TCID50 of the indicated viruses. Animals were humanely euthanized and lungs harvested for virus titration 4 days after inoculation. Virus titers in nasal turbinate (blue), trachea (green) and lung (red) were determined by titration on MDCK cells and TCID50 calculated by the Reed-Muench method. Log10TCID50/ml is shown, with error bars indicating standard deviations for three individual rats in each virus group. No viable virus was detected in the control group and data are not shown.
Figure 2.
Replication of selected influenza A viruses in the respiratory tract of SD rats. To evaluate the infectivity of different influenza viruses (see Table 1) in rats, 8- to 9-week-old SD rats were inoculated with doses of 106 (A), 104 (B) and 102 (C) TCID50 of the indicated viruses. Animals were humanely euthanized and lungs harvested for virus titration 4 days after inoculation. Virus titers in nasal turbinate (blue), trachea (green) and lung (red) were determined by titration on MDCK cells and TCID50 calculated by the Reed-Muench method. Log10TCID50/ml is shown, with error bars indicating standard deviations for three individual rats in each virus group. No viable virus was detected in the control group and data are not shown.
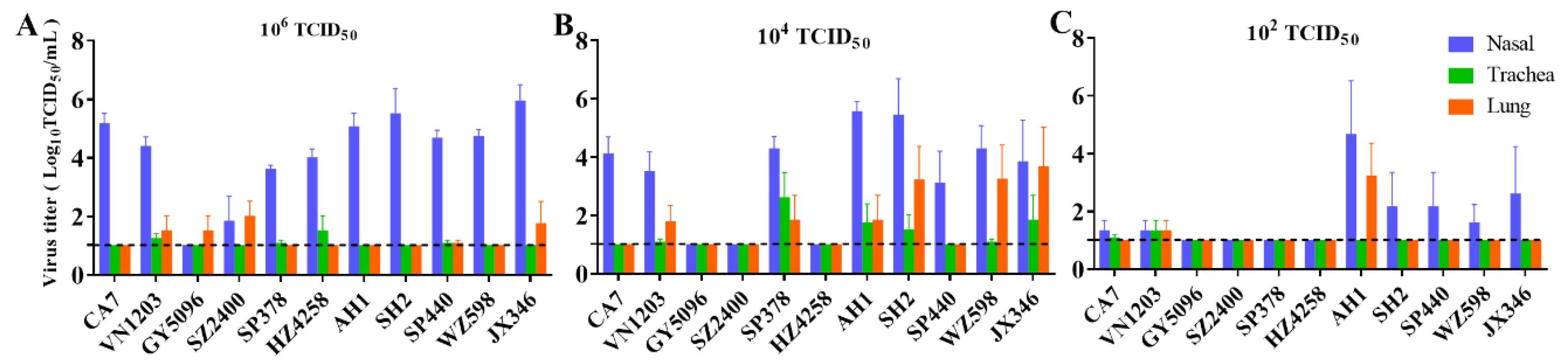
Figure 3.
Serological antibody titers in rats challenged with selected influenza A viruses. Blood from rats inoculated with 106 TCID50 of each virus (see Table 1) was collected for serological analysis at the end of the 14-day experiment (14 dpi). Antibodies to influenza A viruses were titrated by the hemagglutination inhibition (HAI) test. Each solid circle represents one seropositive rat in each virus group. Data from rats that failed to produce antibodies (seronegative) and from the control group are shown in Table S1.
Figure 3.
Serological antibody titers in rats challenged with selected influenza A viruses. Blood from rats inoculated with 106 TCID50 of each virus (see Table 1) was collected for serological analysis at the end of the 14-day experiment (14 dpi). Antibodies to influenza A viruses were titrated by the hemagglutination inhibition (HAI) test. Each solid circle represents one seropositive rat in each virus group. Data from rats that failed to produce antibodies (seronegative) and from the control group are shown in Table S1.
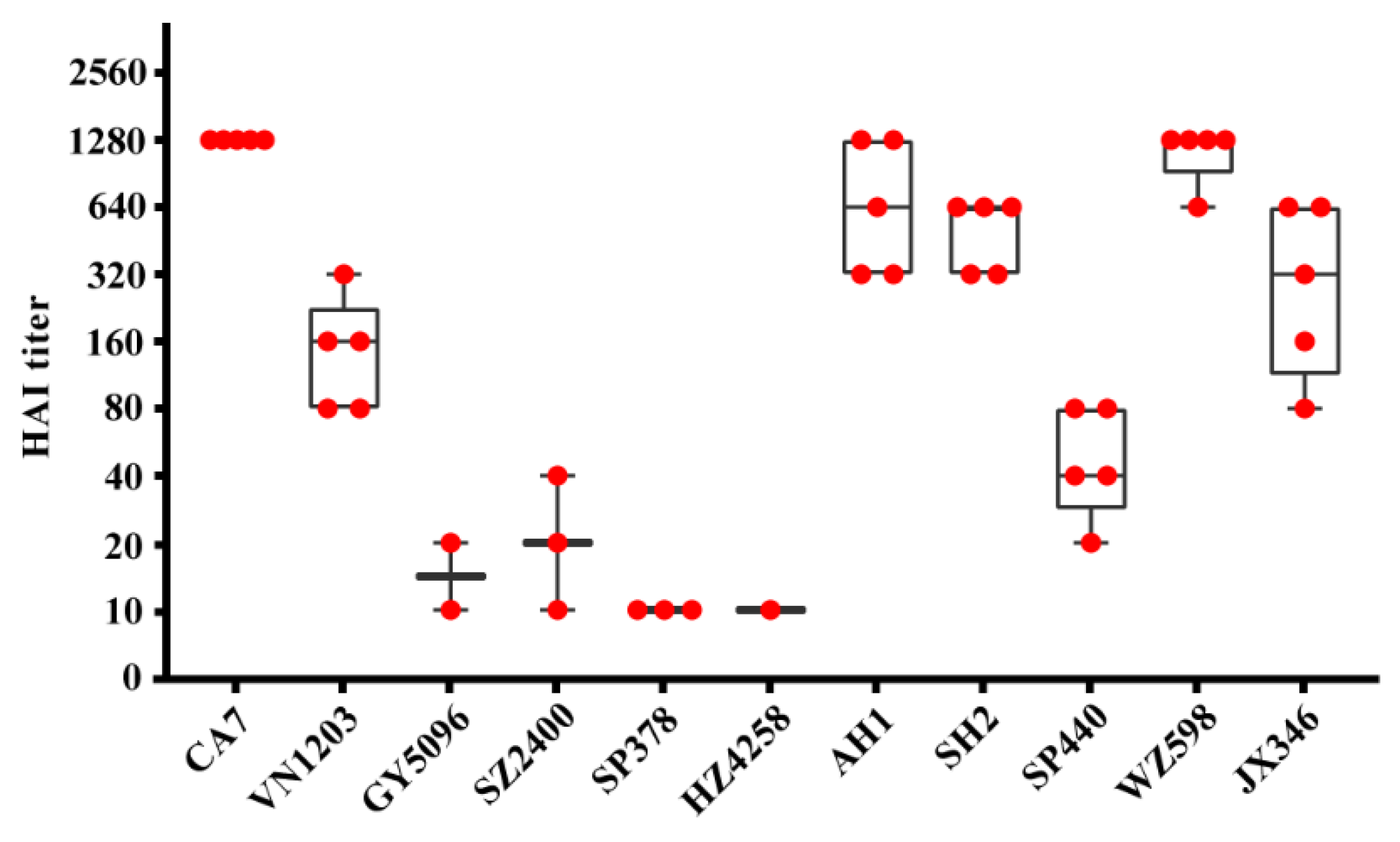
Figure 4.
Histopathological analysis of the lungs of SD rats inoculated with different subtypes of influenza A virus. Three rats were inoculated at the indicated doses per virus, and the lungs were harvested at 4 dpi. Hematoxylin-eosin staining was performed on 3 μm sections. Whole sections were used for analysis, but only representative images are shown, as pathological changes were comparable among the different virus groups at the same dose. Dose-dependent effects are shown with representative lung sections from rats inoculated with SH2 (H7N9) at 102 TCID50 (A), 104 TCID50 (B) and 106 TCID50 (C). Pathological changes in the lungs of mock-infected SD rats (D) and those inoculated with all tested influenza A viruses are shown with representative lung sections from the 104 TCID50 group (E, CA7, pandemic 2009 H1N1; F, VN1203, H5N1; G, GY5096, H5N1; H, SZ2400, H5N6; I, SP378, H5N6; J, HZ4258, H5N8; K, AH1, H7N9; L, SH2, H7N9; M, SP440, H7N9; N, WZ598, H9N2; O, JX346, H10N8). Scale bars indicate 200 μm.
Figure 4.
Histopathological analysis of the lungs of SD rats inoculated with different subtypes of influenza A virus. Three rats were inoculated at the indicated doses per virus, and the lungs were harvested at 4 dpi. Hematoxylin-eosin staining was performed on 3 μm sections. Whole sections were used for analysis, but only representative images are shown, as pathological changes were comparable among the different virus groups at the same dose. Dose-dependent effects are shown with representative lung sections from rats inoculated with SH2 (H7N9) at 102 TCID50 (A), 104 TCID50 (B) and 106 TCID50 (C). Pathological changes in the lungs of mock-infected SD rats (D) and those inoculated with all tested influenza A viruses are shown with representative lung sections from the 104 TCID50 group (E, CA7, pandemic 2009 H1N1; F, VN1203, H5N1; G, GY5096, H5N1; H, SZ2400, H5N6; I, SP378, H5N6; J, HZ4258, H5N8; K, AH1, H7N9; L, SH2, H7N9; M, SP440, H7N9; N, WZ598, H9N2; O, JX346, H10N8). Scale bars indicate 200 μm.
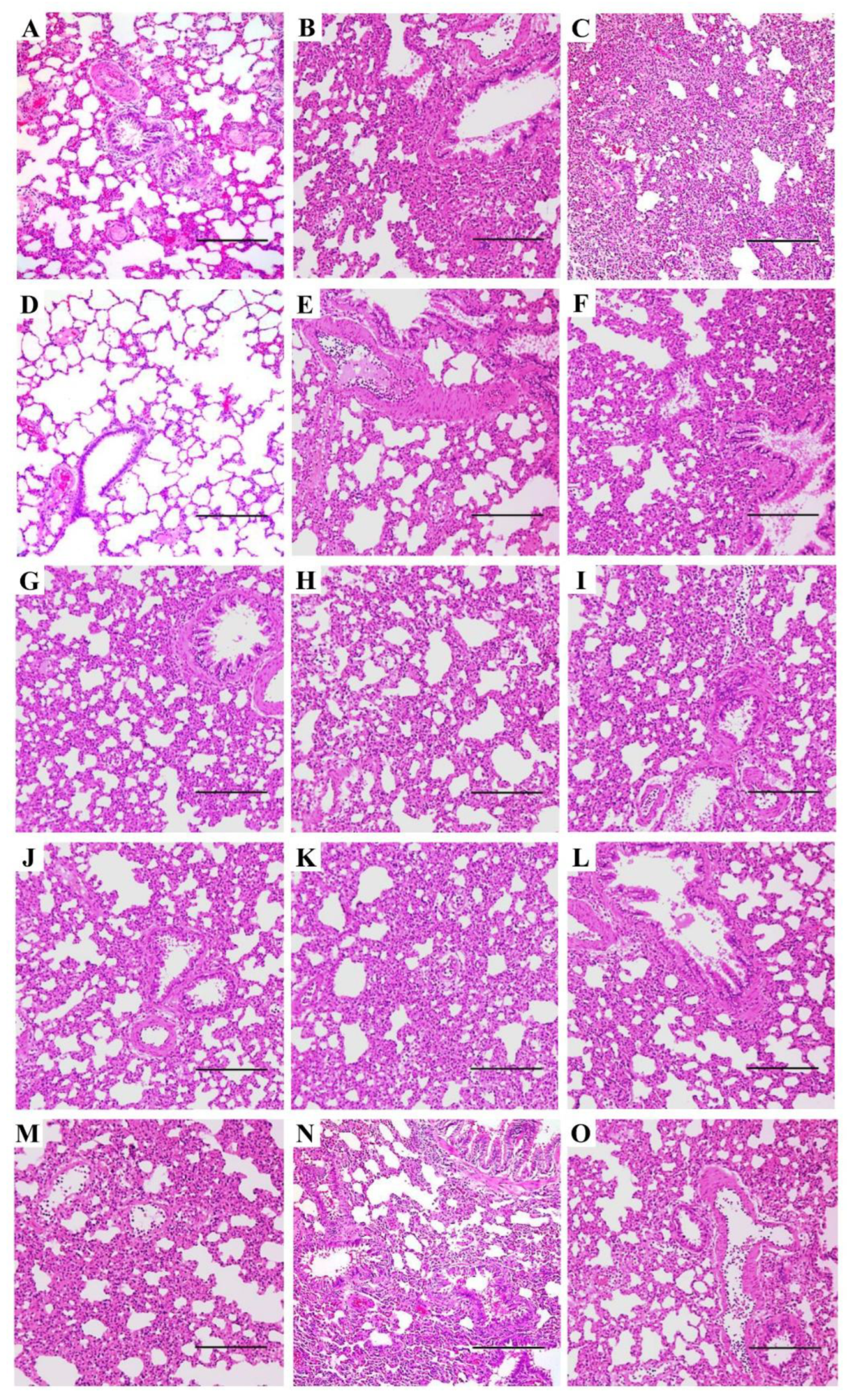
Figure 5.
Immunohistochemical analysis of influenza virus nucleoprotein (NP) in the respiratory tract of SD rats. Airway tissues were harvested at 4 dpi and 3 μm sections were used for immunohistochemical analysis. A representative image was selected for viruses with similar virus distribution patterns. Influenza NP antigens (brown) are shown in the nasal turbinate (A, CA7, pandemic 2009 H1N1, 106 TCID50; B, HZ4258, H5N8, 106 TCID50; C, AH1, H7N9, 102 TCID50; D, AH1, H7N9, 104 TCID50; E, SH2, H7N9, 102 TCID50; F, SH2, H7N9, 104 TCID50; G, SH2, H7N9, 106 TCID50; H, SP440, H7N9, 102 TCID50; I, SP440, H7N9, 104 TCID50; J, SP440, H7N9, 106 TCID50; K, WZ598, H9N2, 106 TCID50; L, JX346, H10N8, 104 TCID50) and lungs (M, VN1203, H5N1, 104 TCID50; N, AH1, H7N9, 102 TCID50; O, WZ598, H9N2, 104 TCID50). Scale bars indicate 200 μm.
Figure 5.
Immunohistochemical analysis of influenza virus nucleoprotein (NP) in the respiratory tract of SD rats. Airway tissues were harvested at 4 dpi and 3 μm sections were used for immunohistochemical analysis. A representative image was selected for viruses with similar virus distribution patterns. Influenza NP antigens (brown) are shown in the nasal turbinate (A, CA7, pandemic 2009 H1N1, 106 TCID50; B, HZ4258, H5N8, 106 TCID50; C, AH1, H7N9, 102 TCID50; D, AH1, H7N9, 104 TCID50; E, SH2, H7N9, 102 TCID50; F, SH2, H7N9, 104 TCID50; G, SH2, H7N9, 106 TCID50; H, SP440, H7N9, 102 TCID50; I, SP440, H7N9, 104 TCID50; J, SP440, H7N9, 106 TCID50; K, WZ598, H9N2, 106 TCID50; L, JX346, H10N8, 104 TCID50) and lungs (M, VN1203, H5N1, 104 TCID50; N, AH1, H7N9, 102 TCID50; O, WZ598, H9N2, 104 TCID50). Scale bars indicate 200 μm.
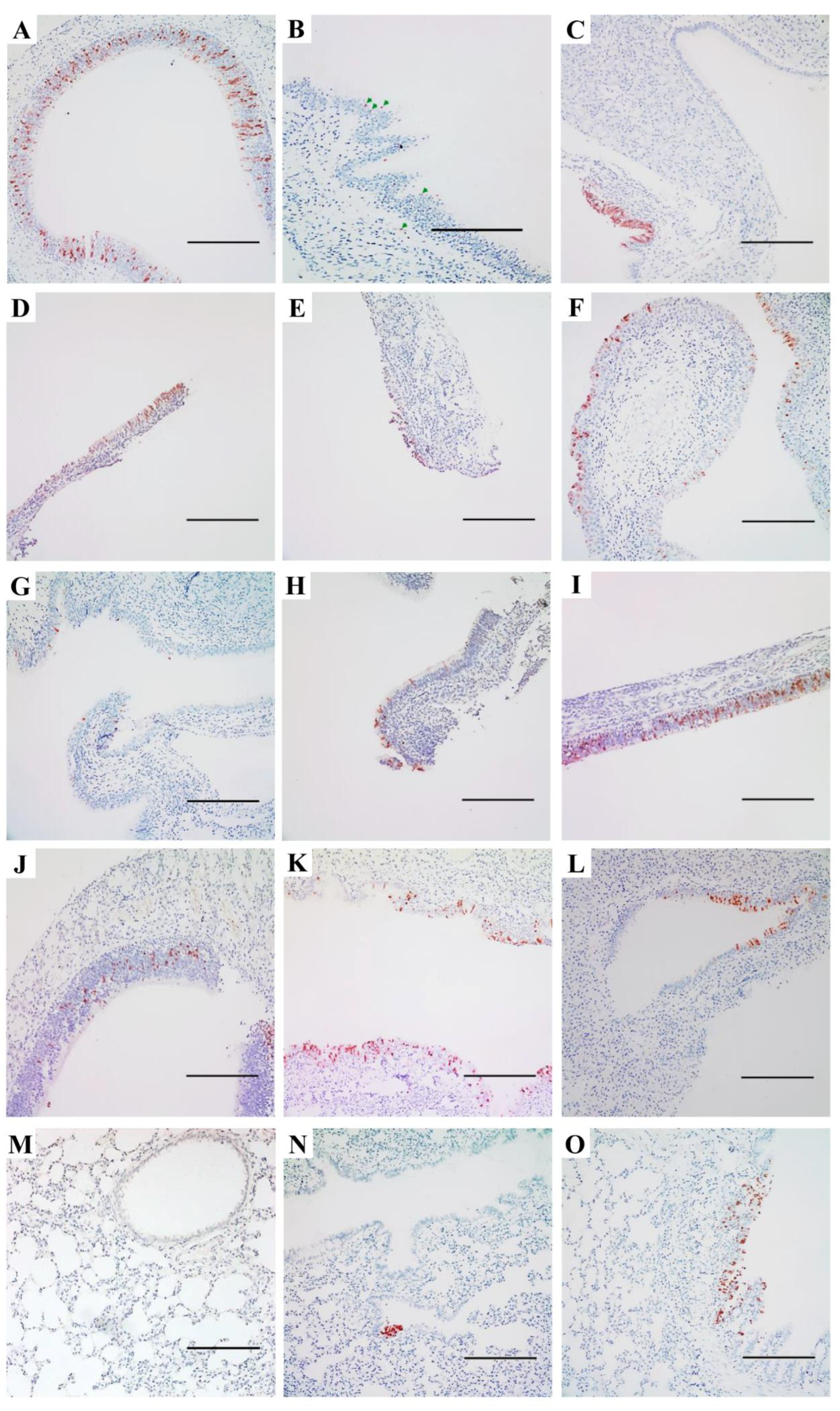
Figure 6.
Distribution of α2,3- and α2,6-linked sialosides in the respiratory tracts of SD rats. The α2,3- and α2,6-linked sialosides in the nasal turbinate (A, B, C), trachea (D, E, F) and lung (G, H, I) of naïve rats were detected with biotinylated Maackia amurensis agglutinin I or II (MAA I, MAA II) for α2,3-linked sialosides or Sambucus nigra agglutinin (SNA) for α2,6-linked sialosides. Both α2,3- and α2,6-linked sialosides were visible in brown. Scale bars indicate 200 μm.
Figure 6.
Distribution of α2,3- and α2,6-linked sialosides in the respiratory tracts of SD rats. The α2,3- and α2,6-linked sialosides in the nasal turbinate (A, B, C), trachea (D, E, F) and lung (G, H, I) of naïve rats were detected with biotinylated Maackia amurensis agglutinin I or II (MAA I, MAA II) for α2,3-linked sialosides or Sambucus nigra agglutinin (SNA) for α2,6-linked sialosides. Both α2,3- and α2,6-linked sialosides were visible in brown. Scale bars indicate 200 μm.
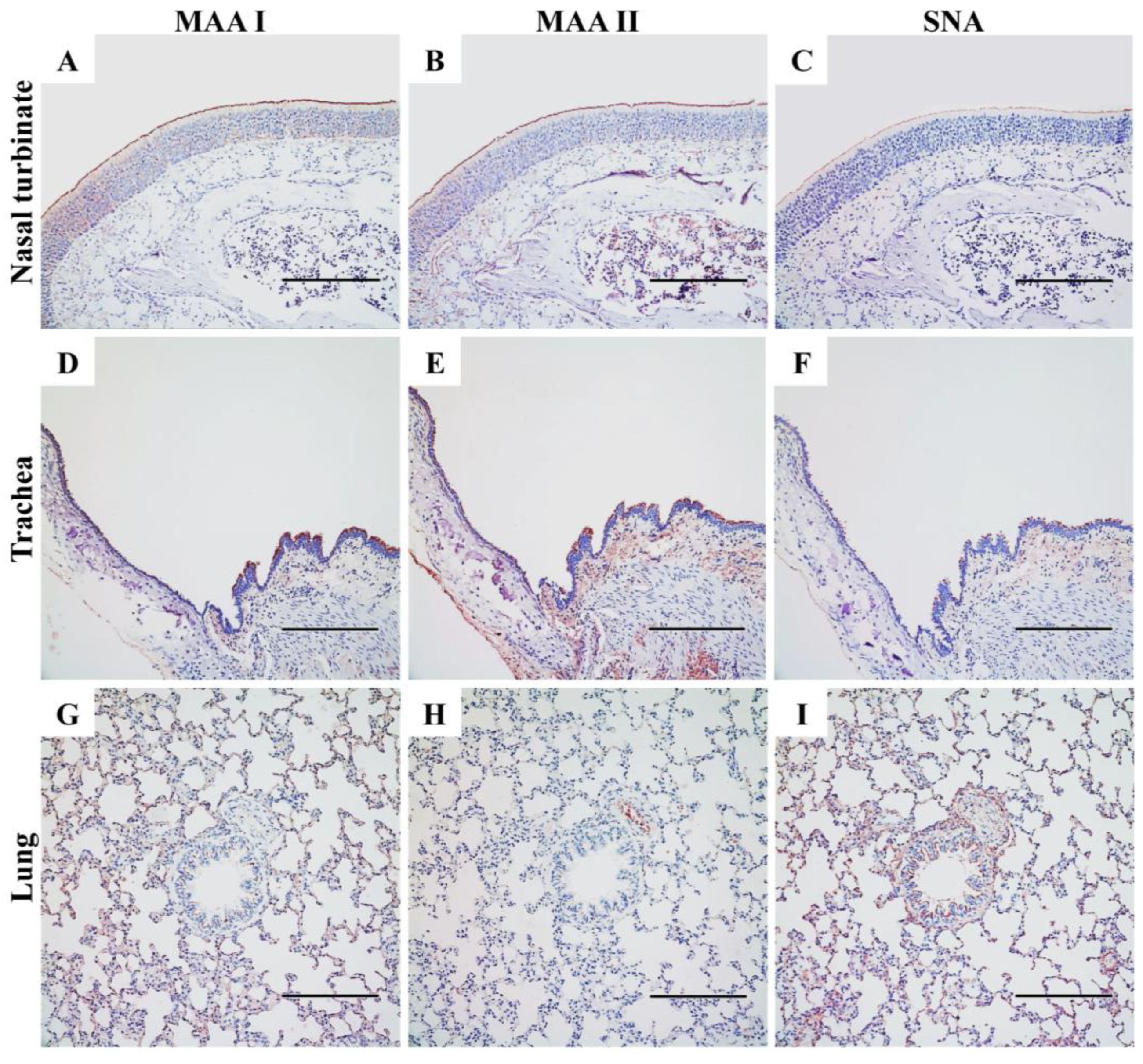

Table 1.
Summary of the influenza A virus strains used in this study.
| Strain Name | Abbreviation | Subtype | Clade1 |
| A/California/07/2009 | CA7 | H1N1 | H1N1/2009 prototype |
| A/Vietnam/1203/2004 | VN1203 | H5N1 | HAPI H5 clade 1.0 |
| A/Duck/Guiyang/5096/2013 | GY5096 | H5N1 | HPAI H5 clade 2.3.4.4g |
| A/Chicken/Shenzhen/2400/2013 | SZ2400 | H5N6 | HPAI H5 clade 2.3.4.4a |
| A/Shenzhen/SP378/2015 | SP378 | H5N6 | HPAI H5 clade 2.3.4.4e |
| A/Duck/Huzhou/4258/2013 | HZ4258 | H5N8 | HPAI H5 clade 2.3.4.4b |
| A/Anhui/1/2013 | AH1 | H7N9 | H7N9 prototype |
| A/Shanghai/2/2013 | SH2 | H7N9 | H7N9 prototype |
| A/Guangdong/SP440/2017 | SP440 | H7N9 | HPAI H7N9 |
| A/Chicken/Wenzhou/598/2013 | WZ598 | H9N2 | H9N2 Y280 lineage |
| A/Jiangxi-Donghu/346/2013 | JX346 | H10N8 | H10N8 prototype |
1 All H5 viruses and the SP440 (H7N9) are highly pathogenic avian influenza (HPAI) strains, while the others are low pathogenic (LPAI).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



