
Submitted:
06 March 2025
Posted:
07 March 2025
You are already at the latest version
Abstract
Colpodella species are free-living predatory protists that prey on algae, ciliates and bodonids using myzocytosis. Colpodella species have been reported in human and animal infections. Polymerase chain reaction (PCR) using primers targeting 18S rRNA genes of Cryptosporidium and piroplasms has identified Colpodella species in arthropods, host blood, and feces demonstrating the phylogenetic closeness of Colpodella species to the apicomplexa. However, in both human and animal infections, life cycle stages of Colpodella present in arthropods and infected hosts are unknown. In this review we provide an overview of widespread occurrence of Colpodella species in ticks, and pathogenicity in humans and animals. We discuss methods for culture and microscopy that can aid diagnosis. Phylogenetic tree analysis of Colpodella species identified using 18S rRNA demonstrates that the Colpodella species identified in different geographic regions represent different species and strains that may impact virulence and zoonotic transmission. There is a pressing need to culture Colpodella species, and to stain cells for morphological identification. This will aid molecular investigations aimed at identifying molecular markers of Colpodella spp. facilitating transmission, survival and pathogenesis in hosts, and determining which species and strains to prioritize for the risk of zoonotic infections to humans and for infections in animals.
Keywords:
1. Introduction
1.1. Geographic Distribution of Colpodella Species
1.2. Colpodella Species Infections in Human and Animal Hosts
2. Is Colpodella Species an Opportunistic or Zoonotic Parasite?
2.1. Colpodellosis in the Making
2.2. From Free-Living Predators to Opportunistic Parasites
2.3. Pathogenic Protists Are Vectors for Other Pathogens
2.4. Culturing Colpodella Species for Identification of Virulence Markers
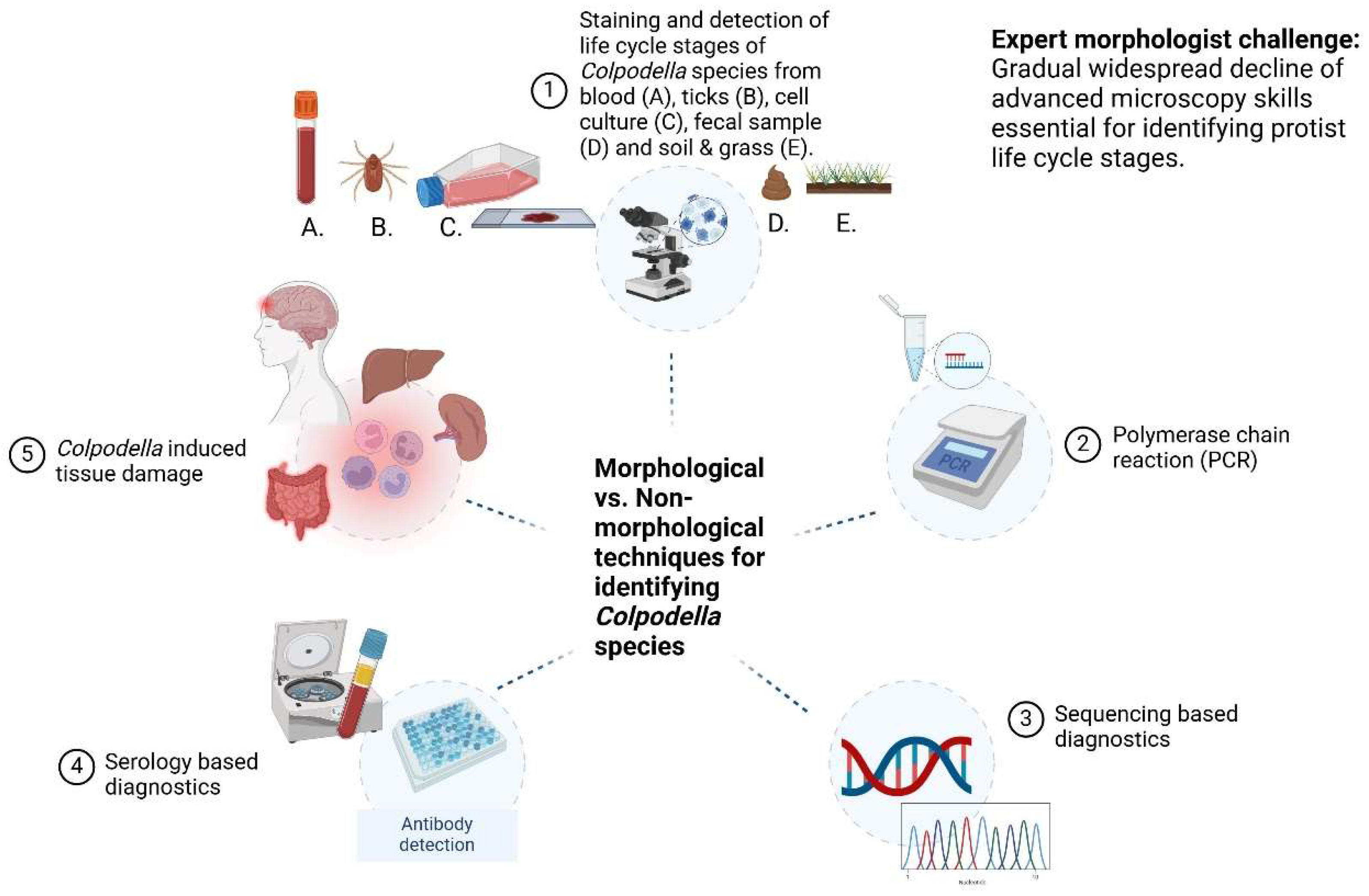
3. Techniques for Detecting Life Cycle Stage Markers of Colpodella spp. in Arthropod and Vertebrate Hosts
3.1. Mechanisms of Pathogenesis in Colpodellosis. What do We Know?
3.2. Microscopy: The Gold Standard for Parasite Identification
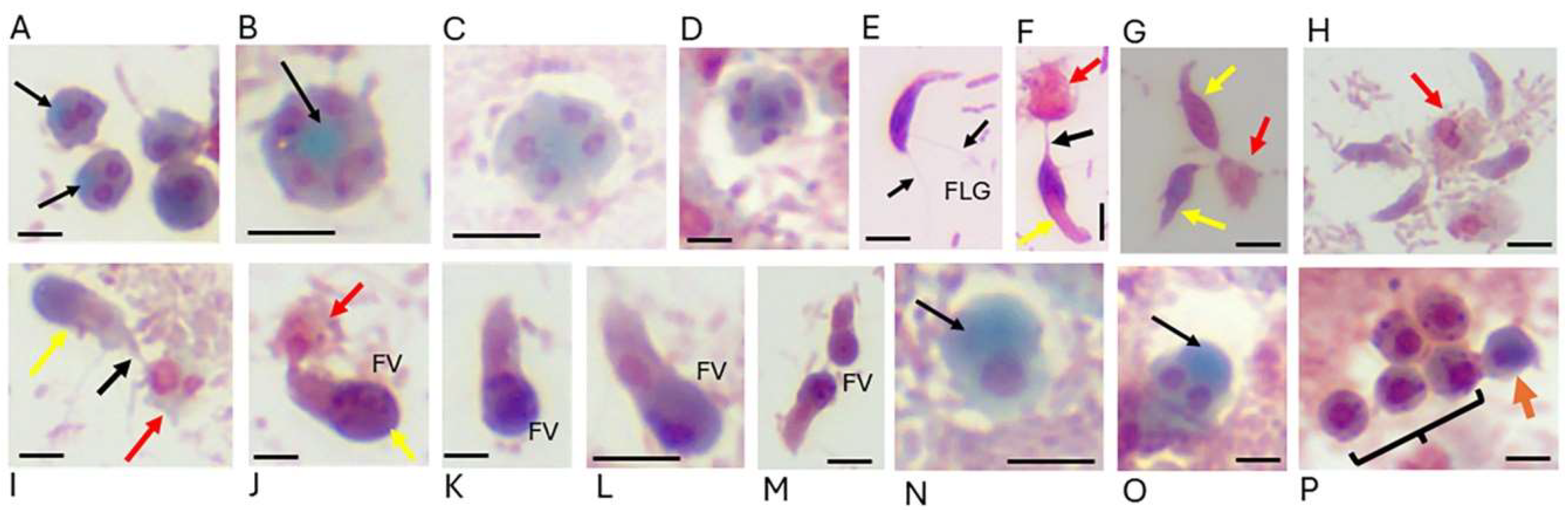
3.3. Morphology-Based Diagnostic Techniques
3.4. Parasite Induced Tissue Damage
3.5. Expert Morphologist Challenge
3.6. Polymerase Chain Reaction (PCR) and Sequencing Based Diagnostics
3.7. Serology Based Diagnostics
3.8. Cost Effectiveness
4. Culture Conditions for Colpodellids and Chromerids
4.1. Culture Media for Cultivating Colpodella Species
4.2. Light, Differential Interference Contrast (DIC) Microscopy and Electron Microscopy (EM)

| Sam-Yellowe’s Trichrome Staining | Application of dyes in the order of incubation |
| Sam-Yellowe’s trichrome A | 0.3 % Methylene blue (1 min) 1% Brilliant green (5 min) 1% Neutral Red (1 min) Distilled water washes were performed in between each dye incubation. After the last wash, smears are air-dried before microscope observation using oil immersion at x1000. |
| Sam-Yellowe’s trichrome D | 1% Crystal violet (30 s) 1% Brilliant green (2 min) 1% Neutral red (1 min) Distilled water washes were performed in between each dye incubation. After the last wash, smears are air-dried before microscope observation using oil immersion at x1000. |
| Sam-Yellowe’s trichrome E | 1 % Crystal violet (30 s) 1% Brilliant green (2 min) 1% Safranin (1 min) Distilled water washes were performed in between each dye incubation. After the last wash, smears are air-dried before microscope observation using oil immersion at x1000. |
| Sam-Yellowe’s trichrome J | 0.3 % Methylene blue (1 min) 0.5 % Fast green in alcohol (5 min) 1% Neutral Red (1 min) Distilled water washes were performed in between each dye incubation. After the last wash, smears are air-dried before microscope observation using oil immersion at x1000. |
5. Colpodella spp. in Coinfections
5.1. Colpodella spp. Detected in Blood
5.2. Colpodella spp. Detected in Blood and Ticks from Cattle and Goats
5.3. Colpodella spp. Detected in Blood from Dogs, Cats and Ticks
5.4. Colpodella spp. Detected in Camel Ticks, Cattle and Wildlife
5.5. Colpodella spp. Identified in Fecal Samples from Sheep, Goats, Cattle, Duck and Eurasian Coot
5.6. Colpodella spp. Detected in Pangolin Ticks
6. Colpodella spp. in Single Infections
6.1. Colpodella spp. in Human Infections
6.2. Colpodella spp. Detected in Fecal Samples from Large Cats
7. Conclusions and Recommendations
Acknowledgments
Conflicts of Interest
References
- Kuvardina, O.N.; Leander, B.S.; Aleshin, V.V.; Myl'Nikov, A.P.; Keeling, P.J.; Simdyanov, T.G. The Phylogeny of Colpodellids (Alveolata) Using Small Subunit rRNA Gene Sequences Suggests They are the Free-living Sister Group to Apicomplexans. J. Eukaryot. Microbiol. 2002, 49, 498–504. [CrossRef]
- Simpson, A.; Patterson, D. Ultrastructure and the identification of the predatory flagellate Colpodella pugnax Cienkowski (Apicomplexa) with a description of Colpodella turpis n. sp. and a review of the genus. Syst, Parasitol., 1996, 33, 187-198.
- Brugerolle, G. Colpodella vorax: ultrastructure, predation, life-cycle, mitosis, and phylogenetic relationships. Eur. J. Protistol. 2002, 38, 113–125. [CrossRef]
- Mylnikov, A.P. Ultrastructure and phylogeny of colpodellids (Colpodellida, Alveolata). Biol. Bull. 2009, 36, 582–590. [CrossRef]
- Mylnikov, A.P.; Mylnikova, Z.M. Feeding spectra and pseudoconoid structure in predatory alveolate flagellates. Inland Water Biol. 2008, 1, 210–216. [CrossRef]
- Cavalier-Smith, T.; Chao, E. Protalveolate phylogeny and systematics and the origins of Sporozoa and dinoflagellates (phylum Myzozoa nom. nov.). Eur. J. Protistol. 2004, 40, 185–212. [CrossRef]
- Sam-Yellowe, T. Y.; Getty, T. A.; Addepalli, K.; Walsh, A. M.; Williams-Medina, A. R.; Fujioka, H.; Peterson, J. W. Novel life cycle stages of Colpodella sp. (Apicomplexa) identified using Sam-Yellowe's trichrome stains and confocal and electron microscopy. Int Microbiol. 2022, 25, 669-678.
- Getty, T.A.; Peterson, J.W.; Fujioka, H.; Walsh, A.M.; Sam-Yellowe, T.Y. Colpodella sp. (ATCC 50594) Life Cycle: Myzocytosis and Possible Links to the Origin of Intracellular Parasitism. Trop. Med. Infect. Dis. 2021, 6, 127. [CrossRef]
- Matsimbe, A.M.; Magaia, V.; Sanches, G.S.; Neves, L.; Noormahomed, E.; Antunes, S.; Domingos, A. Molecular detection of pathogens in ticks infesting cattle in Nampula province, Mozambique. Exp. Appl. Acarol. 2017, 73, 91–102. [CrossRef]
- Jimale, K.A.; Bezerra-Santos, M.A.; Mendoza-Roldan, J.A.; Latrofa, M.S.; Baneth, G.; Otranto, D. Molecular detection of Colpodella sp. and other tick-borne pathogens in ticks of ruminants, Italy. Acta Trop. 2024, 257, 107306. [CrossRef]
- Li, B.; Zhai, J.-Q.; Wu, Y.-J.; Shan, F.; Zou, J.-J.; Hou, F.-H.; Que, T.-C.; Chen, W. Molecular identification of tick-borne Rickettsia, Anaplasma, Ehrlichia, Babesia, and Colpodella in confiscated Malayan pangolins. PLOS Neglected Trop. Dis. 2024, 18, e0012667. [CrossRef]
- Chiu, H.-C.; Sun, X.; Bao, Y.; Fu, W.; Lin, K.; Chen, T.; Zheng, C.; Li, S.; Chen, W.; Huang, C. Molecular identification of Colpodella sp. of South China tiger Panthera tigris amoyensis (Hilzheimer) in the Meihua Mountains, Fujian, China. Folia Parasitol. 2022, 69. [CrossRef]
- Soliman, A.M.; Mahmoud, H.Y.; Hifumi, T.; Tanaka, T. Discovery of Colpodella spp. in ticks (Hyalomma dromedarii) infesting camels in southern Egypt. Ticks Tick-borne Dis. 2024, 15, 102352. [CrossRef]
- 14.
- Qi, Y.; Wang, J.; Lu, N.; Qi, X.; Yang, C.; Liu, B.; Lu, Y.; Gu, Y.; Tan, W.; Zhu, C.; et al. Potential novel Colpodella spp. (phylum Apicomplexa) and high prevalence of Colpodella spp. in goat-attached Haemaphysalis longicornis ticks in Shandong province, China. Ticks Tick-borne Dis. 2024, 15, 102328. [CrossRef]
- Phetkarl, T.; Fungwithaya, P.; Udompornprasith, S.; Amendt, J.; Sontigun, N. Preliminary study on prevalence of hemoprotozoan parasites harbored by Stomoxys (Diptera: Muscidae) and tabanid flies (Diptera: Tabanidae) in horse farms in Nakhon Si Thammarat province, Southern Thailand. Veter- World 2023, 16, 2128–2134. [CrossRef]
- Janouškovec, J.; Tikhonenkov, D.V.; Burki, F.; Howe, A.T.; Kolísko, M.; Mylnikov, A.P.; Keeling, P.J. Factors mediating plastid dependency and the origins of parasitism in apicomplexans and their close relatives. Proc. Natl. Acad. Sci. 2015, 112, 10200–10207. [CrossRef]
- Hussain, S.; Li, X.; Bukhari, S.M.; Zhou, M.; Ahmad, S.; Javid, A.; Guan, C.; Hussain, A.; Ali, W.; Khalid, N.; et al. Cross-genera amplification and identification of Colpodella sp. with Cryptosporidium primers in fecal samples of zoo felids from northeast China. Braz. J. Biol. 2023, 83, e247181. [CrossRef]
- Elochukwu, C. V.; Nnabuife, H. E.; Nicodemus, M.; Ogo, N. I.;Sylvanus, O. S.; Cornelius, J. O.; Kamani, J.; Maxwell, O. N. Molecular detection of Colpodella sp. using Cryptosporidium primers in faecal samples of small ruminants in FCT and Plateau State , Nigeria. J. Vet. Biomed. Sci. 2024, 6, 112-122.
- Hasapis, K.A.; Charalambidou, I.; Phanis, C.O.; Kazamia, S.; Kassinis, N.; Schou, C.; Karanis, P. First Detection and Molecular Characterization of Colpodella in Goats, Foxes, and Birds. Acta Parasitol. 2025, 70, 1–6. [CrossRef]
- Neupane, S.; Saski, C.; Nayduch, D. House fly larval grazing alters dairy cattle manure microbial communities. BMC Microbiol. 2021, 21, 1–14. [CrossRef]
- Myshrall, K.L.; Mobberley, J.M.; Green, S.J.; Visscher, P.T.; Havemann, S.A.; Reid, R.P.; Foster, J.S. Biogeochemical cycling and microbial diversity in the thrombolitic microbialites of Highborne Cay, Bahamas. Geobiology 2010, 8, 337–354. [CrossRef]
- Yuan, C. L.; Keeling, P. J.; Krause, P.J. et al. Colpodella spp.–like Parasite Infection in Woman, China. Emerg Infect Dis. 2012, 18, 125-127.
- Jiang, J.-F.; Jiang, R.-R.; Chang, Q.-C.; Zheng, Y.-C.; Jiang, B.-G.; Sun, Y.; Jia, N.; Wei, R.; Liu, H.-B.; Huo, Q.-B.; et al. Potential novel tick-borne Colpodella species parasite infection in patient with neurological symptoms. PLOS Neglected Trop. Dis. 2018, 12, e0006546. [CrossRef]
- Xu, M.; Hu, Y.; Qiu, H.; Wang, J.; Jiang, J. Colpodella sp. (Phylum Apicomplexa) Identified in Horses Shed Light on Its Potential Transmission and Zoonotic Pathogenicity. Front. Microbiol. 2022, 13, 857752. [CrossRef]
- Wheatley, M.A.; Shamoun, J.; Maggi, R.; Breitschwerdt, E.B.; Sommer, S.L.; Cullen, J.M.; Stowe, D.M. Eosinophilic pericardial effusion and pericarditis in a cat. J. Feline Med. Surg. Open Rep. 2023, 9. [CrossRef]
- Neculicioiu, V. S.; Colosi, I. A.; Toc, D. A.; Lesan, A.; Costache, C. When a Ciliate Meets a Flagellate: A Rare Case of Colpoda spp. and Colpodella spp. Isolated from the Urine of a Human Patient. Case Report and Brief Review of Literature. Biology (Basel). 2021, 10, 476.
- Solarz, W.; Najberek, K.; Wilk-Woźniak, E.; Biedrzycka, A. Raccoons foster the spread of freshwater and terrestrial microorganisms—Mammals as a source of microbial eDNA. Divers. Distrib. 2020, 26, 453–459. [CrossRef]
- Stoeck, T.; Kasper, J.; Bunge, J.; Leslin, C.; Ilyin, V.; Epstein, S. Protistan Diversity in the Arctic: A Case of Paleoclimate Shaping Modern Biodiversity?. PLOS ONE 2007, 2, e728. [CrossRef]
- Solon, A.J.; Mastrangelo, C.; Vimercati, L.; Sommers, P.; Darcy, J.L.; Gendron, E.M.S.; Porazinska, D.L.; Schmidt, S.K. Gullies and Moraines Are Islands of Biodiversity in an Arid, Mountain Landscape, Asgard Range, Antarctica. Front. Microbiol. 2021, 12. [CrossRef]
- Heidelberg, K.B.; Nelson, W.C.; Holm, J.B.; Eisenkolb, N.; Andrade, K.; Emerson, J.B. Characterization of eukaryotic microbial diversity in hypersaline Lake Tyrrell, Australia. Front. Microbiol. 2013, 4, 115. [CrossRef]
- Egizi, A.; Morin, P.J.; Fonseca, D.M. Unraveling microbe-mediated interactions between mosquito larvae in a laboratory microcosm. Aquat. Ecol. 2014, 48, 179–189. [CrossRef]
- Rahman, M. T.; Sobur, M. A.; Islam, M. S.; Ievy, S.; Hossain, M. J.; El Zowalaty, M. E.; Rahman, A. T.; Ashour, H. M. Zoonotic Diseases: Etiology, Impact, and Control. Microorganisms. 2020, 8, 1405.
- Sam-Yellowe, T.Y.; Roy, A.; Nims, T.; Qaderi, S.; Peterson, J.W. Detection of Kelch13 and Coronin Genes in Colpodella sp. ATCC 50594. Parasitologia 2025, 5, 5.
- Olmo, J. L.; Esteban, G. F.; Finlay, B. J. New records of the ectoparasitic flagellate Colpodella gonderi on non-Colpoda ciliates. J Int Microbiol. 2011, 14, 207-211.
- Sam-Yellowe, T. Y.; Asraf, M. M.; Peterson, J. W.; Fujioka, H. Fluorescent Nanoparticle Uptake by Myzocytosis and Endocytosis in Colpodella sp. ATCC 50594. Microorganisms 2023, 11, 1945.
- Wu, S.; Meng, J.; Yu, F.; Zhou, C.; Yang, B.; Chen, X.; Yang, G.; Sun, Y.; Cao, W.; Jiang, J.; Wu, J.; Zhan, L. Molecular epidemiological investigation of piroplasms carried by pet cats and dogs in an animal hospital in Guiyang, China. Front Microbiol. 2023, 14, 1266583.
- Huggins, L. G.; Colella, V.; Koehler, A. V.; Schunack, B.; Traub, R. J. A multipronged next-generation sequencing metabarcoding approach unearths hyperdiverse and abundant dog pathogen communities in Cambodia. Transbound Emerg Dis. 2022, 69, 1933-1950.
- Squarre, D.; Nakamura, Y.; Hayashida, K.; Kawai, N.; Chambaro, H.; Namangala, B.; Sugimoto, C.; Yamagishi, J. Investigation of the piroplasm diversity circulating in wildlife and cattle of the greater Kafue ecosystem, Zambia. Parasit Vectors. 2020, 13, 599.
- Salazar-Ardiles, C.; Paredes Valencia, K.; Andrade, D. C. Amoebas: the omnipotent organism and silent assassin. Mol Biol Rep. 2025, 52, 160.
- Mathison, B. A.; Sapp, S. G. H. An annotated checklist of the eukaryotic parasites of humans, exclusive of fungi and algae. ZooKeys. 2021, 1069, 1–313.
- Contreras-Ferro, R.; Trueba, J. M.; Sánchez-Mora, P.; Escudero, R.; Sánchez-Seco, M. P.; Montero, E.; Negredo, A.; González, L. M.; Dashti, A. Llorente, M. T.; Gil-Zamorano, J.; Vázquez, A.; Jado, I.; González-Barrio, D. Why an Integrated Approach to Tick-Borne Pathogens (Bacterial, Viral, and Parasitic) Is Important in the Diagnosis of Clinical Cases. Trop Med Infect Dis. 2024, 9, 272.
- Chen, F.; Fu, S.; Jiang, J. F.; Feng, H.; Liu, Z.; Sun, Y.; Li, M. Persistent human babesiosis with low-grade parasitemia, challenges for clinical diagnosis and management. Heliyon. 2024, 10, e39960.
- Baneth, G.; Nachum-Biala, Y.; Dvorkin, A.; Arogeti, I.; Amiel, S.; Soueid, Y.; Shwartz, D.; Mumcuoglu, K. Y.; Salant, H. Description of Babesia galileei sp. nov. A Piroplasmid species causing severe disease in domestic cats. Parasit Vectors. 2024, 17, 297.
- Olano, J. P.; Weller, P. F.; Guerrant, R. L.; Walker, D. H. Principles of parasitism: host–parasite interactions. Tropical infectious diseases: Principles, pathogens and practice, 2011, p.1-7.
- Roberts, L. S.; Janovy, J. Jr.; Nadler, S. Gerald D. Schmidt & Larry S. Roberts Foundation of Parasitology, 9th edition, McGraw Hill Publishers, 2012.
- Jones, A. W. Introduction to Parasitology, Addison Wesley, Publishing Company, 1967.
- Zeeshan, I.; Ijaz, A.; Maghsi, I. A.; Qasim, M.; Amin, S. B. Navigating Naegleria fowleri: Understanding pathogenesis, causes and preventive measures. Medi Clin Case Rep J. 2023, 1, 166-167.
- Cope, J. R.; Ratard, R. C.; Hill, V. R.; Sokol, T.; Causey, J. J.; Yoder, J. S.; Mirani, G.; Mull, B.; Mukerjee, K. A.; Narayanan, J.; Doucet, M.; Qvarnstrom, Y.; Poole, C. N.; Akingbola, O. A.; Ritter, J. M.; Xiong, Z.; da Silva, A. J.; Roellig, D.; Van Dyke, R. B.; Stern, H.; Xiao, L.; Beach, M. J. The first association of a primary amebic meningoencephalitis death with culturable Naegleria fowleri in tap water from a US treated public drinking water system. Clin Infect Dis. 2015, 60, e36-42.
- Aurongzeb, M.; Fatima, S. Z.; Hussain, S. I.; Rashid, Y.; Aziz, T.; Alhomrani, M.; Alsanie, W. F.; Alamri, A. S. Detection and identification of Naegleria species along with Naegleria fowleri in the tap water samples. BMC Med Genomics. 2025, 18, 6.
- Aykur, M.; Dagci, H. Evaluation of molecular characterization and phylogeny for quantification of Acanthamoeba and Naegleria fowleri in various water sources, Turkey. PLoS One. 2021, 16, e0256659.
- Dereeper, A.; Allouch, N.; Guerlais, V.; Garnier, M.; Ma, L.; De Jonckheere, J. F.; Joseph, S. J.; Ali, I. K. M.; Talarmin, A.; Marcelino, I. Naegleria genus pangenome reveals new structural and functional insights into the versatility of these free-living amoebae. Front Microbiol. 2023, 13, 1056418.
- Phung, N. T. N.; Pham, H. T.; Tran, T. T.; Dinh, V. H.; Tran, N. M.; Tran, N. A. N.; Ngo, M. Q. N.; Nguyen, H. T. T.; Tran, D. K.; Le, T. K. T.; Quek, C.; Pham, V. H.; Pham, S. T. Naegleria fowleri: Portrait of a Cerebral Killer. Diagnostics (Basel). 2025, 15, 89.
- Aykur, M.; Dirim Erdogan, D.; Selvi Gunel, N.; Guler, A.; Biray Avci, C.; Celebisoy, N.; Gunduz, C.; Dagci, H. Genotyping and Molecular Identification of Acanthamoeba Genotype T4 and Naegleria fowleri from Cerebrospinal Fluid Samples of Patients in Turkey: Is it the Pathogens of Unknown Causes of Death? Acta Parasitol. 2022, 67, 1372-1383.
- Borecka, A.; Bielawska-DrÓzd, A.; Skotarczak, B.; Adamska, M.; CieŚlik, P.; Antos-Bielska, M.; SkopiŃska-RÓŻewska, E.; Donskow-Łysoniewska, K. Acanthamoeba - pathogen and vector of highly pathogenic bacteria strains to healthy and immunocompromised individuals. Cent Eur J Immunol. 2020, 45, 228-232.
- Sousa-Ramos, D.; Reyes-Batlle, M.; Bellini, N. K.; Rodríguez-Expósito, R. L.; Piñero, J. E.; Lorenzo-Morales, J. Naegleria australiensis isolated from a wastewater treatment station in Santiago Island, Cape Verde. J Water Health. 2023, 21, 443-450.
- Visvesvara, G. S.; Moura, H.; Schuster, F. L. Pathogenic and opportunistic free-living amoebae: Acanthamoeba spp., Balamuthia mandrillaris, Naegleria fowleri, and Sappinia diploidea. FEMS Immunol Med Microbiol. 2007, 50, 1-26.
- Walochnik, J.; Wylezich, C.; Michel, R. The genus Sappinia: history, phylogeny and medical relevance. Exp Parasitol. 2010, 126, 4-13.
- Otero-Ruiz, A.; Gonzalez-Zuñiga, L. D.; Rodriguez-Anaya, L. Z.; Lares-Jiménez, L. F.; Gonzalez-Galaviz, J. R.; Lares-Villa, F. Distribution and Current State of Molecular Genetic Characterization in Pathogenic Free-Living Amoebae. Pathogens. 2022, 11, 1199.
- Rojo, J. U.; Rajendran, R.; Salazar, J. H. Laboratory Diagnosis of Primary Amoebic Meningoencephalitis. Lab Med. 2023, 54, e124-e132.
- Shaukat, A.; Khaliq, N.; Riaz, R.; Munsab, R.; Ashraf, T.; Raufi, N.; Shah, H. Noninvasive diagnostic biomarkers, genomic profiling, and advanced microscopic imaging in the early detection and characterization of Naegleria fowleri infections leading to primary amebic meningoencephalitis (PAM). Ann Med Surg (Lond). 2024, 86, 2032-2048.
- El-Sayed, N. M.; Hikal, W. M. Several staining techniques to enhance the visibility of Acanthamoeba cysts. Parasitol Res. 2015, 114, 823-30.
- Gunarathna, N.; Amarasinghe, A.; Wijesundara, S.; Iddawela, D.; Wickramasinghe, S. Isolation, molecular characterization and phylogeny of Naegleria species in water bodies of North-Western Province, Sri Lanka. PLoS One. 2021, 16, e0248510.
- Philippe, N.; Shukla, A.; Abergel, C.; Bisio, H. Genetic manipulation of giant viruses and their host, Acanthamoeba castellanii. Nat Protoc. 2024, 19, 3-29.
- Guimaraes, A. J.; Gomes, K. X.; Cortines, J. R.; Peralta, J. M.; Peralta, R. H. Acanthamoeba spp. as a universal host for pathogenic microorganisms: One bridge from environment to host virulence. Microbiol Res. 2016, 193, 30-38.
- Arthofer, P.; Panhölzl, F.; Delafont, V.; Hay, A.; Reipert, S.; Cyran, N.; Wienkoop, S.; Willemsen, A.; Sifaoui, I.; Arberas-Jiménez, I.; Schulz, F.; Lorenzo-Morales, J.; Horn M. A giant virus infecting the amoeboflagellate Naegleria. Nat Commun. 2024, 15, 3307.
- Sam-Yellowe, T.Y.; Yadavalli, R.; Fujioka, H.; Peterson, J.W.; Drazba, J.A. RhopH3, rhoptry gene conserved in the free-living alveolate flagellate Colpodella sp. (Apicomplexa). Eur. J. Protistol. 2019, 71, 125637. [CrossRef]
- Birnbaum, J.; Scharf, S.; Schmidt, S.; Jonscher, E.; Hoeijmakers, W.A.M.; Flemming, S.; Toenhake, C.G.; Schmitt, M.; Sabitzki, R.; Bergmann, B.; et al. A Kelch13-defined endocytosis pathway mediates artemisinin resistance in malaria parasites. Science 2020, 367, 51–59. [CrossRef]
- Behrens, H. M.; Schmidt, S.; Spielmann, T. The newly discovered role of endocytosis in artemisinin resistance. Med Res Rev. 2021, 41, 2998-3022.
- Sharma, A. I.; Shin, S. H.; Bopp, S.; Volkman, S. K.; Hartl, D. L.; Wirth, D. F. Genetic background and PfKelch13 affect artemisinin susceptibility of PfCoronin mutants in Plasmodium falciparum. PLoS Genet. 2020, 16, e1009266.
- Siddiqui, F. A.; Boonhok, R.; Cabrera, M.; Mbenda, H. G. N.; Wang, M.; Min, H.; Liang, X.; Qin, J.; Zhu, X.; Miao, J.; Cao, Y.; Cui, L. Role of Plasmodium falciparum Kelch 13 Protein Mutations in P. falciparum Populations from Northeastern Myanmar in Mediating Artemisinin Resistance. mBio. 2020, 11, e01134-19.
- Gnädig, N. F.; Stokes, B. H.; Edwards, R. L.; Kalantarov, G. F.; Heimsch, K. C.; Kuderjavy, M.; Crane, A.; Lee, M. C. S.; Straimer, J.; Becker, K.; Trakht, I. N.; Odom John, A. R.; Mok, S.; Fidock, D. A. Insights into the intracellular localization, protein associations and artemisinin resistance properties of Plasmodium falciparum K13. PLoS Pathog. 2020, 16, e1008482.
- Demas, A. R.; Sharma, A. I.; Wong, W.; Early, A. M.; Redmond, S.; Bopp, S.; Neafsey, D. E.; Volkman, S. K.; Hartl, D. L.; Wirth, D. F. Mutations in Plasmodium falciparum actin-binding protein coronin confer reduced artemisinin susceptibility. Proc Natl Acad Sci U S A. 2018, 115, 12799-12804.
- Bane, K. S.; Lepper, S.; Kehrer, J.; Sattler, J. M.; Singer, M.; Reinig, M.; Klug, D.; Heiss, K.; Baum, J.; Mueller, A. K.; Frischknecht, F. The Actin Filament-Binding Protein Coronin Regulates Motility in Plasmodium Sporozoites. PLoS Pathog. 2016, 12:e100571.
- Chan, K. T.; Creed, S. J.; Bear, J. E. Unraveling the enigma: progress towards understanding the coronin family of actin regulators. Trends Cell Biol. 2011, 21, 481-8.
- Sam-Yellowe, T. Y.; Fujioka, H.; Peterson, J. W. Ultrastructure of Myzocytosis and Cyst Formation, and the Role of Actin in Tubular Tether Formation in Colpodella sp. (ATCC 50594). Pathogens, 2022, 11, 455.
- Nakada-Tsukui, K.; Nozaki, T. Trogocytosis in Unicellular Eukaryotes. Cells. 2021, 10, 2975.
- Xiang, C.; Li, Y.; Jing, S.; Han, S.; He, H. Trichomonas gallinae Kills Host Cells Using Trogocytosis. Pathogens. 2023, 12, 1008.
- Salti, M. I.; Sam-Yellowe, T. Y. Are Colpodella Species Pathogenic? Nutrient Uptake and Approaches to Diagnose Infections. Pathogens. 2024, 13, 600.
- Vandersea, M. W.; Birkenheuer, A. J.; Litaker, R. W.; Vaden, S. L.; Renschler, J. S.; Gookin, J. L. Identification of Parabodo caudatus (class Kinetoplastea) in urine voided from a dog with hematuria. J Vet Diagn Invest. 2015, 27, 117-20.
- Bradbury, R. S.; Sapp, S. G. H.; Potters, I.; Mathison, B. A.; Frean, J.; Mewara, A.; Sheorey, H.; Tamarozzi, F.; Couturier, M. R.; Chiodini, P.; Pritt, B. Where Have All the Diagnostic Morphological Parasitologists Gone? J Clin Microbiol. 2022, 60, e0098622.
- Rosenblatt, J. E. Laboratory diagnosis of infections due to blood and tissue parasites. Clin Infect Dis. 2009, 49, 1103-8.
- Ricciardi, A.; Ndao, M. Diagnosis of Parasitic Infections: What’s Going On? Journal of Biomolecular Screening. 2015, 20, 6-21.
- Leander, B. S.; Kuvardina, O. N.; Aleshin, V. V.; Mylnikov, A. P.; Keeling, P. J. Molecular phylogeny and surface morphology of Colpodella edax (Alveolata): insights into the phagotrophic ancestry of apicomplexans. J Eukaryot Microbiol. 2003, 50, 334-40.
- Li, X.; Dang, Z.; Tang, W.; Zhang, H.; Shao, J.; Jiang, R.; Zhang, X.; Huang, F. Detection of Parasites in the Field: The Ever-Innovating CRISPR/Cas12a. Biosensors (Basel). 2024, 14, 145.
- Sam-Yellowe, T. Y.; Addepalli, K.; Yadavalli, R.; Peterson, J. W. New trichrome stains identify cysts of Colpodella sp. (Apicomplexa) and Bodo caudatus. Int Microbiol. 2020, 23, 303-311.
- Aikawa, M. Variations in structure and function during the life cycle of malarial parasites. Bull World Health Organ. 1977, 55, 139-56.
- Mackenstedt, U.; Brockelman, C. R.; Mehlhorn, H.; Raether, W. Comparative morphology of human and animal malaria parasites. I. Host-parasite interface. Parasitol Res. 1989, 75, 528-35.
- Fuehrer, H. P.; Campino, S.; Sutherland, C. J. The primate malaria parasites Plasmodium malariae, Plasmodium brasilianum and Plasmodium ovale spp.: genomic insights into distribution, dispersal and host transitions. Malar J. 2022, 21, 138.
- Antunes, S.; Rosa, C.; Couto, J.; Ferrolho, J.; Domingos, A. Deciphering Babesia-Vector Interactions. Front Cell Infect Microbiol. 2017, 7, 429.
- Lares-Jiménez, L. F.; Borquez-Román, M. A.; Alfaro-Sifuentes, R.; Meza-Montenegro, M. M.; Casillas-Hernández, R.; Lares-Villa F. Detection of serum antibodies in children and adolescents against Balamuthia mandrillaris, Naegleria fowleri and Acanthamoeba T4. Exp Parasitol. 2018, 189, 28-33.
- Orfano, A. S.; Nacif-Pimenta, R.; Duarte, A. P.; Villegas, L. M.; Rodrigues, N. B.; Pinto, L. C.; Campos, K. M.; Pinilla, Y. T.; Chaves, B.; Barbosa Guerra, M. G.; Monteiro, W. M.; Smith, R. C.; Molina-Cruz, A.; Lacerda, M. V.; Secundino, N. F.; Jacobs-Lorena, M.; Barillas-Mury, C.; Pimenta, P. F. Species-specific escape of Plasmodium sporozoites from oocysts of avian, rodent, and human malarial parasites. Malar J. 2016, 15, 394.
- de Souza, W. Contribution of microscopy to a better understanding of the anatomy of pathogenic protists. Proc Natl Acad Sci U S A. 2024, 121, e2321515121.
- Kuupiel, D.; Bawontuo, V.; Mashamba-Thompson, T. P. Improving the Accessibility and Efficiency of Point-of-Care Diagnostics Services in Low- and Middle-Income Countries: Lean and Agile Supply Chain Management. Diagnostics (Basel). 2017, 7, 58.
- von Philipsborn, P.; Steinbeis, F.; Bender, M. E.; Regmi, S.; Tinnemann, P. Poverty-related and neglected diseases - an economic and epidemiological analysis of poverty relatedness and neglect in research and development. Glob Health Action. 2015, 8, 25818.
- Getty, T. Life Cycle and Morphological Characterization of Colpodella sp. (ATCC 50594) in Hay Medium. Master's Thesis completed December 2020. Cleveland State University, Cleveland Ohio, USA.
- Sam-Yellowe, T. Y.; Yadavalli, R. Voromonas Pontica Identified by Giemsa Staining and AntiRhopH3 Protein Reactivity. J Microbiol Modern Tech. 2019, 4, 103.
- Oborník, M.; Modrý, D.; Lukeš, M.; Cernotíková-Stříbrná, E.; Cihlář, J.; Tesařová, M.; Kotabová, E.; Vancová, M.; Prášil, O.; Lukeš, J. Morphology, ultrastructure and life cycle of Vitrella brassicaformis n. sp., n. gen., a novel chromerid from the Great Barrier Reef. Protist. 2012, 163, 306-23.
- Oborník, M.; Vancová, M.; Lai, D. H.; Janouškovec, J.; Keeling, P. J.; Lukeš, J. Morphology and ultrastructure of multiple life cycle stages of the photosynthetic relative of apicomplexa, Chromera velia. Protist. 2011,162, 115-30.
- Sam-Yellowe, T. Y.; Salti, M. I.; Adeloye, O. E. Sam-Yellowe’s Trichrome Staining Identifies Life Cycle Stages of Free-Living Colpodellids. J Appl Microb Res. 2024, 7, 2 (01-09).
- Füssy, Z.; Masařová, P.; Kručinská, J.; Esson, H. J.; Oborník, M. Budding of the Alveolate Alga Vitrella brassicaformis Resembles Sexual and Asexual Processes in Apicomplexan Parasites. Protist. 2017, 168, 80-91.
- Zhou, X.; Xia, S.; Huang, J. L.; Tambo, E.; Zhuge, H. X.; Zhou, X. N. Human babesiosis, an emerging tick-borne disease in the People's Republic of China. Parasit Vectors. 2014, 7, 509.
- Kumar, A.; O'Bryan, J.; Krause, P. J. The Global Emergence of Human Babesiosis. Pathogens. 2021, 10, 1447.
- Sanchez-Vicente, S.; Tokarz, R. Tick-Borne Co-Infections: Challenges in Molecular and Serologic Diagnoses. Pathogens. 2023, 12, 1371.
| Organism Name | Source | Animal | Tick/Biting flies | Country |
| 1. Colpodella sp. QYi-2023b | Ticks infesting goat and dog | Goat | Haemaphysalis longicornis | China: Yiyuan county Shangdong |
| 2. Colpodella sp. QYi-2023a | Ticks infesting goat | Goat | Haemaphysalis longicornis | China: Yi yuan county Shangdong |
| 3. Colpodella sp. isolate 103 | Blood | Horse | N/A | China |
| 4. Colpodella sp. isolate 115 | Blood | Horse | N/A | China |
| 5. Colpodella sp. RRJ-2016 isolate T18 | N/A | N/A | N/A | China |
| 6. Colpodella sp. RRJ-2016 isolate T17 | N/A | N/A | N/A | China |
| 7. Colpodella sp. HLJ | Woman with neurological symptoms | N/A | Tick | China |
| 8. Colpodella sp. HLJ | ticks infesting dog | Dog | Haemaphysalis longicornis | China: Yiyuan County, Shandong |
| 9. Uncultured Colpodella | Blood | N/A | N/A | (China) |
| 10. Colpodella sp. | Blood | Dog | N/A | China: Guiyang |
| 11. Colpodella sp. | Blood | Cat | N/A | China: Guiyang |
| 12. Colpodella sp. | N/A | N/A | N/A | China |
| 13. Colpodella sp. | N/A | Panthera tigris altaica (Amur tiger) | N/A | China |
| 14. Colpodella sp. | N/A | Panthera tigris altaica (Amur tiger) | N/A | China |
| 15. Colpodella sp. | N/A | Panthera tigris altaica (Amur tiger) | N/A | China |
| 16. Colpodella sp. | N/A | Panthera tigris altaica (Amur tiger) | N/A | China |
| 17.Colpodella tetrahymenae | N/A | Panthera tigris altaica (Amur tiger) | N/A | China |
| 18. Colpodella sp. | N/A | N/A | Rhipicephalus microplus | China |
| 19. Colpodella sp. | N/A | N/A | Rhipicephalus microplus | China |
| 20.Colpodella tetrahymenae | Wastewater | N/A | Dermacentor everestianus | China: Qinghai |
| 21. Colpodella sp. | N/A | N/A | Dermacentor nuttalli | china: Qinghai |
| 22. Colpodella sp. | N/A | N/A | Dermacentor nuttalli | china: Qinghai |
| 23. Colpodella sp. | N/A | N/A | Dermacentor nuttalli | China: Qinghai |
| 24.Colpodella tetrahymenae | N/A | N/A | Dermacentor everestianus | China: Qinghai |
| 25. Colpodellidae sp. | man with relapsing fever | Homo sapiens | N/A | China |
| 26. Colpodellidae sp. | woman with relapsing Babesia-like illness | Homo sapiens | N/A | China |
| 27. Colpodella sp. clone pangolin 18_Tick2 | Amblyomma javanense | Pangolin | Tick | China: Guangzhou |
| 28. Colpodella sp. clone pangolin 17_Tick1 | Amblyomma javanense | Pangolin | Tick | China: Guangzhou |
| 29. Colpodella sp. clone pangolin 16_Tick2 | Amblyomma javanense | Pangolin | Tick | China: Guangzhou |
| 30. Colpodella sp. clone pangolin 12_Tick1 | Amblyomma javanense | Pangolin | Tick | China: Guangzhou |
| 31. Colpodella sp. clone pangolin 11_Tick2 | Amblyomma javanense | Pangolin | Tick | China: Guangzhou |
| 32. Colpodella sp. clone pangolin 4_Tick1 | Amblyomma javanense | Pangolin | Tick | China: Guangzhou |
| 33. Colpodella sp. | N/A | N/A | Tick | Pakistan |
| 34. Colpodella sp. | N/A | N/A | Tick | Pakistan |
| 35. Colpodella sp. | N/A | N/A | Tick | Pakistan |
| 36. Colpodella sp. | N/A | N/A | Tick | Pakistan |
| 37. Colpodella sp. | N/A | N/A | Tick | Pakistan |
| 38. Colpodella sp. | N/A | N/A | Tick | Pakistan |
| 39. Colpodella sp. | tick P03 collected from cattle | Cattle | Rhipicephalus (Boophilus) microplus; sex: female | Russia |
| 40.Colpodella angusta | N/A | N/A | N/A | Russia |
| 41. Colpodella sp. | N/A | Stomoxys indicus | Thailand: Nakhon Si Thammarat | |
| 42. Colpodella sp. | Blood | Dog | Cambodia | |
| 43. Uncultured Colpodella | N/A | Bos taurus | N/A | Japan |
| 44. Colpodella sp. | Feces | Fox (Vulpes vulpes indutus) | N/A | Nicosia, Cyprus |
| 45. Colpodella sp. | Feces | Duck Anas spp. |
N/A | Nicosia, Cyprus |
| 46. Colpodella sp. | Feces | Duck Anas spp. |
N/A | Nicosia, Cyprus |
| 47. Colpodella sp. | Feces | Eurasian Coot (Fulica atra) | ||
| 48. Colpodella sp. | Feces | Goat (Capra hircus) | N/A | Nicosia, Cyprus |
| 49.Colpodella tetrahymenae | N/A | N/A | N/A | Portugal |
| Colpodella angusta | Identified with Amyloodinium ocellatum (dinoflagellate ectoparasite) | Sea bass Dicentrarchus labrax | N/A | Portugal |
| 50. Uncultured Colpodella clone PL31 | Skin, dried ear fragments, Warta Mouth National Park, western Poland | Procyon lotor | N/A | Poland |
| 51. Uncultured Colpodella clone PL31 | Skin, dried ear fragments, Warta Mouth National Park, western Poland | Procyon lotor | N/A | Poland |
| 52. Uncultured Colpodella | slow sand filter column for wastewater treatment, Leipzig | N/A | N/A | Germany |
| 53.Colpodella angusta | Marine sample | N/A | Germany Helgoland |
|
| 54. Uncultured Colpodella | lake water filtered through 3 um from lake Esch sur Sure, depth 0m | N/A | N/A | Luxembourg |
| 55. Colpodella sp. | Ticks | Cattle and goats | Rhipicephalus bursa | Italy |
| 56. Colpodella sp. | Lake water | France | ||
| 57.Colpodella angusta | Feces of calves with diarrhea | N/A | Turkey: Nevsehir | |
| 58.Colpodella gonderi | Woman with urinary tract infection (identified with Colpoda steinii) | N/A | Romania | |
| 59. Colpodella sp. TT-2023 | whole body of Hyalomma dromedarii | Camel | Hyalomma dromedarii | Egypt:Luxor |
| 60. Colpodella sp. TT-2023 | whole body of Hyalomma dromedarii | Camel | Hyalomma dromedarii | Egypt:Aswan |
| 61. Colpodella sp. TT-2023 | whole body of Hyalomma dromedarii | Camel | Hyalomma dromedarii | Egypt:Luxor |
| 62. Colpodella sp. TT-2023 | whole body of Hyalomma dromedarii | Camel | Hyalomma dromedarii | Egypt:Aswan |
| 63. Uncultured Colpodella | Feces | Sheep | N/A | Nigeria |
| 64. Uncultured Colpodella | Feces | Sheep | N/A | Nigeria |
| 65. Uncultured Colpodella | Feces | Sheep | N/A | Nigeria |
| 66. Uncultured Colpodella | Feces | Sheep | N/A | Nigeria |
| 67. Uncultured Colpodella | Feces | Sheep | N/A | Nigeria |
| 68. Uncultured Colpodella | Feces | Sheep | N/A | Nigeria |
| 69. Colpodella sp. | tick P03 collected from cattle | Cattle | Rhipicephalus (Boophilus) microplus; sex: female | Mozambique |
| 70. Colpodella sp. | Blood | Cattle and wildlife | Zambia | |
| 71. Colpodella sp. | N/A | Bovine | N/A | Brazil |
| 72. Colpodella sp. | N/A | Bovine | N/A | Brazil |
| 73. Colpodella sp. | N/A | Bovine | N/A | Brazil |
| 74. Colpodella sp. | N/A | Bovine | N/A | Brazil |
| 75. Colpodella sp. | N/A | Bovine | N/A | Brazil |
| 76. Colpodella sp. | N/A | Bovine | N/A | Brazil |
| 77. Uncultured Colpodella | tropical floodplain lake | N/A | N/A | Brazil |
| 78.Colpodella tetrahymenae | N/A | N/A | N/A | Costa Rica |
| 79.Colpodella angusta | laboratory culture | N/A | N/A | Canada: Vancouver |
| 80.Colpodella angusta | laboratory culture | N/A | N/A | Canada: Vancouver |
| 81.Clopodella angusta | Damp wood chip and surface sand, Locarno beach | N/A | N/A | Canada Vancouver |
| 82.Colpodella angusta | Wood chip on the beach | N/A | N/A | Canada Boundary Bay |
| 83.Colpodella angusta. | Soil from UBC endowment lands | N/A | N/A | Canada Vancouver |
| 84. Uncultured Colpodella | cave and mine | N/A | N/A | New York, USA |
| 85. Colpodella sp. ATCC 50594 | Brown woodland soil, Gambrill Park | N/A | N/A | Maryland, USA |
| 86. Colpodella angusta FL1 | Fresh water laboratory dishes with mosquito larvae, Rutgers University | Mosquito larvae | N/A | New Jersey, USA |
| 87. Colpodella sp. | Cat blood | N/A | N/A | North Carolina, USA |
| 88.Colpodella angusta | Mucus from Acropora formosa, Birch Aquarium | N/A | N/A | San Diego California, USA |
| 89.Colpodella angusta | Anoxic marine sediment, Bolinas Tidal Flat | N/A | N/A | Bolinas, California, USA |
| 90. Uncultured Colpodella | Intertidal thrombolites | N/A | N/A | Florida, USA |
| 91. Uncultured Colpodella | Intertidal thrombolites | N/A | N/A | Florida, USA |
| 92. Uncultured Colpodella | Intertidal thrombolites | N/A | N/A | Florida, USA |
| 93. Uncultured Colpodella | Intertidal thrombolites | N/A | N/A | Florida, USA |
| 94. Colpodella sp. | Cattle manure, identified with Parabodo sp. | N/A | N/A | Kansas, USA |
| 95.Colpodella angusta | Soil, trembling aspen rhizosphere, elevated CO2 conditions | N/A | N/A | Michigan, USA |
| 96. Colpodella sp. | Button and pink thrombolithic mats | N/A | N/A | Bahamas |
| 97. Colpodella sp. | Hypersaline Lake Tyrrell | N/A | N/A | Australia |
| 98.Colpodella angusta | Wastewater | N/A | N/A | Australia |
| 99.Colpodella angusta | Wastewater | N/A | N/A | Australia |
| 100.Colpodella angusta | Wastewater | N/A | N/A | Australia |
| 101. Colpodella sp. | Wastewater | N/A | N/A | Australia |
| 102.Colpodella angusta |
Megalapteryx didinus coprolite, sample 01098a, animal feces/manure Dart River Valley |
N/A | N/A | New Zealand |
| 103. Colpodella sp. | Non crust habitat, Asgard Range | N/A | N/A | Antarctica |
| 104.Colpodella angusta | Soil in front of the Brazilian Antarctic Station | N/A | N/A | Antarctica |
| 105.Colpodella angusta | Oxygen-depleted intertidal marine sediment, upper 2 cm, Greenland | N/A | N/A | Arctic |
| 106.Colpodella angusta | Composting diary manure | N/A | N/A | |
| 107.Colpodella angusta | Marine | N/A | N/A | |
| 108.Colpodella angusta | Animal feces/manure, pig manure storage pit | N/A | N/A |
| Source found with Colpodella spp. | References |
| Ticks | |
| Ixodes persulcatus | [23] |
| Rhipicephalus (Boophilus) microplus | [9] |
| Rh. bursa | [10] |
| Rh. duttoni | [12] |
| Rh. haemaphysaloides | NCBI accession number MH208621 |
| Haemaphysalis longicornis | [12,14] |
| H. flava | [12] |
| H. bispinosa | [12] |
| H. hystricis | [12] |
| Hyalomma dromedarii | [13] |
| Dermacentor everestianus | NCBI accession number MH012047 |
| D. nuttalli | NCBI accession number MH012045 |
| D. andersoni | [12] |
| D. atrosignatus | [12] |
| D. taiwanensis | [12] |
| Amblyomma javanense | [11] |
| Biting fly | |
| Stomoxys indicus | [15] |
| Host tissue and body fluids | |
| Skin | [27] |
| Blood | [22,24,25,36,37,38] |
| Cerebrospinal fluid | [23] |
| Urine | [26] |
| Fecal samples | [17,18,19], NCBI accession number JN245625 |
| Colpodella spp. in humans and animals | Year | Country | Reference |
| Human relapsing fever, non-tick associated blood infection, single infection, female | 2012 | China | [22] |
| Human relapsing fever, non-tick associated, single infection, male | 2017 | China | NCBI accession number MF594625 |
| Cattle, tick associated, co-infection | 2017 | Mozambique | [9] |
| Human tickborne infection, neurological symptoms, single infection, female | 2018 | China | [23] |
| Raccoon, non-tick associated Colpodella spp. in the skin of the ear, co-infection | 2019 | Poland | [27] |
| Cattle, non-tick associated blood infection, co-infection | 2020 | Zambia | [38] |
| Human urinary tract infection associated with Colpodella gonderi and its prey Colpoda steinii, female | 2021 | Romania | [26] |
| Large zoo felids, Colpodella spp. in fecal samples, co-infection | 2021 | China | [17] |
| Dog, non-tick associated blood infection, co-infection | 2021 | Cambodia | [37] |
| Tiger (Panthera tigris amoyensis Hizheimer) in blood and ticks, tickborne Colpodella spp. infection, single infection, multiple organ damage | 2022 | China | [12] |
| Horse non-tick associated blood infection, co-infection | 2022 | China | [24] |
| Cat, non-tick associated blood infection, inflammation, tissue damage, single infection | 2023 | USA | [25] |
| Cats and dogs, non-tick associated blood infection, co-infection | 2023 | China | [36] |
| Horse, Colpopdella spp. in infesting biting fly (Stomoxys indicus), co-infection | 2023 | Thailand | [15] |
| Goats and dogs, Colpodella spp. in ticks | 2024 | China | [14] |
| Camels, Colpodella spp. in infesting ticks | 2024 | Egypt | [13] |
| Cattle and goats, Colpodella spp. in infesting ticks | 2024 | Italy | [10] |
| Goats and sheep, non-tick associated infection, Colpodella spp. in diarrhetic fecal samples, co-infection | 2024 | Nigeria | [18] |
| Pangolins, Colpodella spp. in infesting ticks, co-infection | 2024 | China | [11] |
| Goats, fox, duck, Eurasian Coot, non-tick associated, Colpodella spp. in fecal samples | 2025 | Cyprus | [19] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



