Submitted:
11 March 2025
Posted:
11 March 2025
You are already at the latest version
Abstract
Phenology is one of the most obvious biological characteristics for plants affected by external influences. Phenology variations across temperature reflect energy allocation and reproductive strategies, and measures for plants to adapt to seasonal and environmental changes. Previous studies have mainly focused on national and regional vegetation phenology, but endemic vegetation phenology and dynamic variation in Xinjiang remains unclear. Therefore, our study characterized the Betula microphylla phenology in the Xinjiang area of China, over the period of 2009-2019 based on ≥10 ℃ accumulated temperature data and annual precipitation data using phenological data from field observations, explored B.microphylla ecological strategies and drivers. In this study, a total of 18 phenologies of B. microphylla population naturally distributed in the southern margin of the Junggar Basin were observed, divided the climatic year, the relevant climate data and phenological Julian day were used to analyze. Explored the adaptation strategies of B. microphylla to seasonal changes. We found that (1) The phenology of B. microphylla was generally earlier in warmer years and later in colder years, showing a trade-off relationship. (2) The monthly average temperature and acquired accumulated temperature had strong limitation to inflorescence germination in spring, and the monthly average temperature and low temperature accumulation values showed a synergistic relationship when physiological dormancy was broken. The inflorescence has germination strategy of “high low-temperature needs”. (3) In summer, the shallow root system of B. microphylla can replenish the water lost by transpiration. The male inflorescence shows the strategy of quantity victory. (4) From the perspective of autumn, the male inflorescence abscised, and only spread in the top of branches, so as to provide living space for female inflorescences, it is regarded as a "spatial dislocation" strategy. The male inflorescences use a "long dormancy" strategy to cope with the effects of high autumn temperatures. (5) As winter approaches in Xinjiang, all exposed male inflorescence were covered by bud scales.
Keywords:
synergistic
; quantity
; dormancy
; ecological adaptation
1. Introduction
Vegetation phenology is a sensitive indicator of environmental factors change. Phenological change can reflect physiological condition of the plant itself [1]. Phenophases refer to the times during which plant growth, development, and activities respond to seasonal variations, which represents some of the most prominent biological characteristics influenced by external conditions [2]. Meanwhile, various plant organs such as leaf, flower, stem, root have been identified as bioreactor. Among these plant organs, leaf phenology and flowering phenology are critical traits within the field of reproductive biology research [3]. Leaf undergoes various life processes such as bud emergence, growth, and expands, while flower engages in budburst, pollination, dispersal, and selection of suitable habitats for colonization. In the context of global warming, advanced leaf and flowering phenology in trees and shrubs have been generally reported by previous studies, and this phenomenon was more significantly in trees and shrubs compared to herbaceous plants [4], indicating that woody species exhibit a more rapid response ability to external environmental changes. Furthermore, shifts in phenology may affect individual survival rates and population dynamics while potentially leading to differences in reproductive success.
Phenological alteration also provide feedback to the environment condition, various environmental factors have been identified as influencing vegetation phenology, including temperature [5], photoperiod [6], moisture [7]. For humid temperate zones, a comprehensive study demonstrated that three primary factors exert considerable influence on the phenology of dominant tree species within forests: low winter temperatures, light and temperature[8]. In mid-high latitude regions of the Northern Hemisphere, plant phenology responds more swiftly to temperature fluctuations [9], indicated that the response priorities of plants to phenological influencing factors vary across different climate belts. Besides, variations in seasonal temperatures result in marked differences between spring and autumn phenology [10]. For autumn phenological events, distinct dormancy stages possess unique requirements regarding temperature thresholds and accumulated chilling hours [11]. Further, the various climate conditions impose selection pressures on plants, compelling them to adopt diverse ecological adaptation strategies that facilitate their growth and development. These strategies are particularly evident in alterations of the morphological and physiological traits of plants [12], while phenophases variations result in changes across different organs' growth and development. Phenophases serves as a crucial link between the physiological characteristics of plants and environmental fluctuations. Understanding the relationship between phenophases and environmental factors is essential for elucidating the ecological adaptation strategies employed by species, especially in narrowly distributed species. On the basis, plant’s coping strategies to varying climatic conditions, and disturbances is clarified. Similarity, the accuracy of phenological models across various habitats will also be improved [13,14].
In previous study, there is a notable deficiency of long-term and detailed observational data on individual species. Although there have been studies using many target species, but the type of phenophases was also limited [15,16]. Betulaceae plants are acknowledged as valuable forest resources in their native distribution areas. For example, Betula platyphylla, a highly adaptable dominant tree species found in the natural secondary forests of Northeast China, exhibits relative synchrony with various other tree species across different phenological rhythms [17], suggests that habitat conditions impose natural selection pressures on internal biological rhythms. Moreover, its phenology have been consistently conducted, extensive cultivation throughout numerous regions in China [18,19].
B. microphylla as a tall tree within the Betula genus of the Betulaceae family [20], has a restricted distribution solely to Xinjiang, China. Xinjiang is situated in a high-latitude region characterized by significant diurnal temperature variations. Temperature serves as the primary factor influencing crop production and vegetation survival in this area, rendering it an important focus for research concerning arid and semi-arid regions of Northwest China. Additionally, diverse landform types have fostered various habitat types. Through years of field investigations conducted by our team, we have identified that B. microphylla thrives in habitats such as plain wetlands, coniferous forests on slopes, river valleys, and marsh meadows. Hence, it has thus emerged as an essential natural forest resource within Xinjiang. However, no relevant reports have been found its phenological characteristics. In our study, we conducted long-term field observations on a total of 18 phenophases concerning leaf, flower, and fruit organs across the four seasons of the B. microphylla population naturally distributed in the Mushroom Lake Plain Wetland at the southern margin of the Junggar Basin from 2010 to 2019. The specific start and end times of these phenophases were determined, interannual and seasonal variation trends were analyzed, and meteorological data were collected to investigate the relationship between phenophases and temperature. In summary, our aims were (1) to clarify the seasonal pattern of phenology in B. microphylla, (2) to find meteorological factors influence the phenophases of B. microphylla, (3) to understand ecological adaptation strategies manifest in B. microphylla under varying conditions from a phenological perspective.
2. Study Area
The phenological observation area is located in the plain wetland on the southern bank of Mushroom Lake in Shihezi (Figure 1.), at an altitude of approximately 370m. The annual average temperature ranges from 6 to 7°C, with an extreme minimum temperature around -42°C and an extreme maximum temperature reaching about 42°C [21]. This wetland benefits from a substantial influx of modern glacial meltwater originating from the Altai Mountains to the north and the Manas River, providing abundant water resources for both this ecosystem and the Gurbantunggut Desert situated along its southern margin. Consequently, a unique mountain-oasis-desert multi-ecosystem has formed in this region. Annual precipitation averages between 110 and 150 mm, while annual evaporation rates range from approximately 1800 to 2200 mm. The frost-free period lasts for about 160 to 170 days, with sunshine duration recorded at between 2798 and 2839 hours. The cold climate coupled with significant temperature fluctuations has enabled naturally distributed vegetation within this region to adapt successfully to arid conditions. In recent years, due to economic development along the northern slope of the Tianshan Mountains as well as regional agricultural activities, there has been a gradual increase in farmland reclamation surrounding the wetland. This expansion has led to a markable decline in wetland water levels and adversely affected water availability during critical growth periods for local vegetation. Additionally, urban heat island effects associated with major cities within this economic belt have resulted in changes to plant phenology [22]. Since 1980, coinciding with rising temperature, the phenology of representative woody species such as Salix matsudana, Ulmus pumila, and Populus nigra cv. Afghanica has advanced significantly [23].
3. Materials and Methods
3.1. Phenological Period Observation and Data Arrangement
In accordance with the 《China’s National Phenological Observational Criterion》 [24] and other widely adopted observation techniques [25], we selected ten vigorously growing B. microphylla specimens, ensuring a minimum spacing of 50 meters between each plant, to monitor the onset and conclusion of various phenological events from March 2010 to December 2019. Considering the individual growth characteristics of B. microphylla, we conducted overall phenophases observations on a weekly basis, increasing the frequency to twice per week during flowering and fruiting stages. The number of observations was further augmented in response to significant changes observed in these phenomena. The classification of vegetative growth stages for B. microphylla, as well as the specific observational standards for leaf development, male inflorescence flowering and growth phases, female inflorescence growth phases, and fruiting were all aligned with established protocols utilized by our research team [26]. The meteorological data employed in this study were sourced exclusively from the Wusu Agricultural Meteorological Experiment Station, these data are both authentic and reliable.
3.2. Phenological Observation and Division of Climatic Years
This study primarily investigates the phenology of leaf, flowers, and fruits (Figure 2.). The initial stage of leaf phenology for B. microphylla is characterized by the leaf unfolding stage. Previous research on the phenophases of woody plants in Shihezi City has indicated a weak correlation with temperatures from January to February. However, a significant correlation was observed with temperatures from March to April [27]. Consequently, this paper adopts the average temperature during March and April as the criterion for distinguishing between cold years (Table 1.), warm years, and normal years. Analyzing the variations in average temperatures from March to April between 2010 and 2019 reveals that only 2010 was classified as a cold year, while normal years included 2011, 2014, and 2017. A gradual warming trend became evident in subsequent years (2012, 2013, 2015, 2016, 2018, and 2019). A comparison of average temperatures during these months showed that cold years experienced an average temperature that was approximately 3.8°C lower than that of normal years. Similarity, warm years had an average temperature about 4.85°C higher than normal. The thermal requirements for organisms are based on temperature. Accumulated temperature plays a vital role in agricultural production as well as in understanding plant growth and development dynamics. Currently utilized accumulated temperature thresholds include ≥5°C or ≥10°C. Given the growth habits of B. microphylla, it has been determined that its growth onset occurs in late April. Notably, over the past decade (from data collected), all starting dates for ≥10°C accumulated temperature within April. Therefore, data regarding accumulated temperatures ≥10°C alongside monthly average temperatures from the period spanning from 2010 to 2019 were compiled. Accumulated temperature data served as critical information for investigating both phenology and thermal responses related to B. microphylla within this article.
To present the phenological variations on a year-by-year basis in a more intuitive manner, we first convert the phenological dates into Julian Day format, where April 1st is designated as Day 1; April 2nd as Day 2; and so forth, following this counting method [28]. Secondly, to visually represent the relationship between phenological Julian Days and temperature, we establish the starting dates of phenological Julian Days for different plant organs based on average values derived from multi-year observations: (1) For leaf phenology, leaf flush and young leaf stages occur on April 1st, leaf unfolding stage on May 1st, leaf maturity stage are observed by June 1st, leaf yellowing stage by August 1st, and leaf abscission occurs on September 1st. (2) In terms of female inflorescence, budburst and young fruit stages commence on April 1st, while fruit maturity and abscission occurs by June 1st. (3) Regarding male inflorescence growth phenology, budburst is noted on May 1st, elongation occurs by June 1st, shaping takes place by September 1st, and dormancy begins on October 1st. (4) For male inflorescence flowering phenology, budburst stage, elongation stage, and pollen dispersal stage on April 1st, withering stage and abscission stage are observed by May 1st.
4. Results
4.1. Accumulated Temperature and Sunshine Hours Across Different Years
The start and end dates of the ≥10℃ accumulated temperature across different years exhibit considerable variation, as do the corresponding accumulated temperature values. When classifying climatic year, 2012 stands out as a relatively warm year, with the initiation date for the ≥10℃ accumulated temperature recorded on March 31st. The duration between the start and end dates was 198 days, ranking second only to the highest value (210 days) observed among all studied years. Furthermore, this year experienced an impressive total accumulated temperature of ≥10℃ amounting to 4141.3℃, despite having an annual precipitation of merely 72.6mm. Similarly, another notably warm year, 2016, the start date for the ≥10℃ accumulated temperature occurred even earlier than any other year within the ten-year observation period, specifically on March 24th. The interval from start to end lasted for 195 days, while annual precipitation reached its peak among all ten years at 213.4mm. In 2014, the latest starting point for ≥10℃ accumulating temperatures, was recorded on April 26th, this resulted in a shorter duration at just 166 days and yielded the lowest ≥10℃ accumulated temperature at only 3519.5℃. Additionally, precipitation levels in 2014 were also found to be minimal compared to other observed years at just 65.8mm. In summary, there exists a significant positive correlation between the values of ≥10℃ accumulated temperatures and the number of days spanning from their respective start to end dates over this study period. Notably, an early or late start date for ≥10℃ accumulated temperatures correlates with higher or lower annual precipitation levels respectively.
4.2. The Leaf Phenology of B. microphylla
Based on the observational data collected by our team, the terminal and axillary buds of B. microphylla begin to sprout in Mid-April. The emergence of fresh, green young leaf at the apex of the leaf buds indicates a successful transition into the young leaf stage. Both the leaf flush stage and the young leaf stage demonstrate favorable responses in both cold and warm years. Although average temperatures from March to April may meet criteria for a warm year (Figure 3.), a later onset date for ≥10℃ accumulated temperatures compared to previous years can lead to delays in both leaf flush and young leaf stage (as observed in 2014). Moreover, sudden weather events represent an external factor that significantly influence plant phenology. In 2014, precipitation levels plummeted to their lowest recorded value of 65.8 mm among all observed years, coinciding with a strong cold front that swept across Xinjiang, this was accompanied by widespread rain, snow, fresh gale, sandstorm, abrupt cooling event, and frost conditions. As a result, both leaf flush and young leaf stages for B. microphylla were markable delayed when compared to other typical years (such as 2011 and 2017) within our observation timeframe. Immediately after, approximately nine days later than usual marked the beginning of the leaf unfolding stage, which lasts roughly fifty days. It is noteworthy that leaf unfolding stage tends to be postponed in colder years, compared to normal or warmer years. During warm years such as 2013 and 2015, there was no significant advancement in leaf unfolding stage when juxtaposed with adjacent normal years. A comparative analysis revealed that average temperatures played a crucial role, April's average temperature in 2013 was recorded at 14.1℃, 2℃ lower than that of its preceding year. Similarity, low average temperature were noted for April 2015 at just 13.5℃, a decrease of 7.1℃ from those experienced in April 2014. These findings underscore that specific changes in leaf phenology continue to exhibit a high degree of synchrony with monthly average temperature fluctuations.
The synthesis of chlorophyll involves a series of enzymatic reactions, with the optimal temperature ranging from 20℃ to 30℃. The leaf mainly absorbs blue purple light and appear green. In addition, other types of pigments are present in plants, such as lutein and carotenoids, an increase in temperature not only accelerates the pigment synthesis process but also leads to the decomposition of certain pigments, thereby revealing the colors of other pigments. From July to August, the leaf edges of B. microphylla begin to turn yellow, marking the onset of the leaf yellowing stage, which lasts approximately 20 days. The causes behind phenological fluctuations are multifaceted. Firstly, the average temperature in August for both 2013 (22.7℃) and 2015 (22.6℃) were indicative of warmer years. Conversely, August 2014 experienced an average temperature significantly lower at just 16.3℃ during a regular year. So, the earlier onset of the leaf yellowing stage in both 2013 and 2015 compared to that in 2014 may be attributed to various factors including pest infestations and disease outbreaks. Secondly, during these warmer years, specifically in 2013 and 2015, the total hours of sunshine recorded were notably higher at 2031.2h and 1825.3h respectively when compared with only 1813.4h observed in 2014. Under cooler temperature, leaf photosynthesis within B. microphylla is inhibited, to trigger non-photochemical quenching wherein chlorophyll undergoes decomposition as a means to minimize photosynthetic loss. Thirdly, at lower temperatures chlorophyll molecules become increasingly fragile and more susceptible to damage. The reduced average monthly temperatures coupled with fewer sunlight hours experienced in August 2014 resulted in diminished chlorophyll content along with functional impairment, consequently leading those leaf into an earlier transition into their leaf yellowing stage than observed during other years. In 2013 and 2015, the relatively favorable temperatures facilitated a greater accumulation of chlorophyll in the leaf, leading to a delayed onset of the leaf yellowing stage. The leaf of B. microphylla began to abscise in Mid-September, with nearly all leaf having fallen by the end of October. Consequently, the growth phenology of B. microphylla, which spans approximately 200 days throughout the year, comes to an end.
4.3. The Phenology of Female Inflorescences of B. microphylla
The inflorescence type of B. microphylla is a catkin, and it is classified as a monoecious plant. Ensuring the consistency of pollen release timing is crucial for the efficient completion of pollination in monoecious species. Consequently, the female inflorescence, serving as the reproductive organ for one growing season, must maintain a relatively rapid growth rate. We observed that various life processes associated with female inflorescences, such as budburst, elongation, shaping, and the formation of both young and mature fruiting structures, all occurred successively from mid-April to late May. As illustrated in Figure 4, there was a significant delay in the phenology of female inflorescence budburst to young fruit stage until late April during colder years. In contrast, this process advanced to varying degrees during warmer years compared to adjacent normal years. Notably, in 2013 and 2015, delays were recorded which may have been influenced by average or accumulated temperatures in April. By conducting linear regression analysis on monthly average temperature data alongside phenological day sequences (Figure 5), we found that during dormancy release stage when B. microphylla experienced high accumulated temperatures, the budburst for female inflorescences was delayed due to insufficient low-temperature accumulation (notably observed in 2010, 2013, and 2017). Furthermore, when monthly average temperatures remained approximately consistent over multiple years (specifically from 2017-2019), early budburst of female inflorescences was noted. This indicates that as a reproductive organ functioning within an annual cycle it responds positively to stable interannual variations in monthly average temperature. Finally, part of the accumulated temperature acquired by these plants contributes towards breaking dormancy, thus in certain years characterized by elevated accumulated temperature values some energy is allocated toward promoting meristematic development leading to an earlier onset of female inflorescence budburst (2012 and 2016). However, in years marked by lower accumulated temperature above process tends to be delayed resulting in postponed entry into germinative phenology ( 2015).
Following the pollination of male flowers, the female inflorescences undergoing a transformation to fruit clusters. After receiving adequate sunlight and absorbing substantial nutrients, these female fruit-clusters reach maturity. The samaras align systematically, upon accumulating sufficient temperature exposure, begin to flutter down with the wind, thus completing the process of seed dissemination. The above stages are referred to as the maturity and abscission stage of the female fruit-clusters. Due to the relatively synchronized timing of life activities among female flowers, they tend to enter the maturity and fruit abscission stage later in colder years or when monthly average temperatures are lower during corresponding months (Figure 5.). In warmer years such as 2014 (with an average temperature of 24.5°C in June), there is an acceleration in fruit shedding leading to an earlier onset of the fruit abscission stage. In summary, the influence of monthly average temperature on both the maturity and abscission stages of female fruit-clusters aligns consistently with previous observations regarding phenological variations within these clusters.
4.4. The Phenology of Male Inflorescences of B. microphylla
B. microphylla is generally a wind pollinated plant, small nut with membranous wings. Its male inflorescences serve as reproductive organs that endure through the winter months. Moreover, the phenological development of these inflorescences begins in late May, with flowering not occurring until April of the following year. By mid to late April, the flower buds initiate budburst, these buds crack and elongate within 4-5 days, followed by a peak period of pollen dispersion lasting 2-3 days (Figure 6.). Thus, the entire process of pollen dissemination is completed within 8-10 days. Overall, the variations in flowering phenology (Figure 7.), specifically during the budburst stage, elongation stage, and pollen dispersion stage are closely associated with climatic classifications such as warm years, cold years, and normal years. In warmer years, this phenology occurs significantly earlier than in normal years, conversely, it is delayed during colder years. As May approaches, male inflorescences enter their budburst stage, a pattern that mirrors the developmental timeline from budburst to young fruit stage observed in female inflorescences. Following pollen dispersion, male flowers begin to wither until they eventually abscise. The withering process may be postponed only during colder years when average monthly temperatures are lower. Otherwise, it typically concludes by mid to late April in other conditions. Elevated temperatures tend to further expedite both withering and abscission processes for male inflorescences. In June, rising temperatures stimulate growth phenology among male inflorescences, the elongation stage commences currently. Over nearly three months' duration thereafter, these organs continue to extend until they reach maturity by early September. Notably lower average monthly temperatures recorded in adjacent years can also contribute to relatively later elongation stage observed during warmer seasons such as those experienced in 2013 and 2015.
When the male inflorescence reaches a specific length, width, and defined shape, it enters a critical phase of growth phenology known as the dormancy stage at the end of October. During this time, the organism suspends complex life activities and accumulates energy for the budburst of flower buds in the following year. Successfully navigating this dormancy stage is an essential prerequisite for both the budburst of flower buds in the subsequent year and the timely onset of the next flowering phenology. Firstly, by comparing the day number of male inflorescence budburst phenology observed from 2010 to 2019 with both the monthly average temperature during its dormancy stage from the previous year and ≥10℃ accumulated temperatures (Figure 8), it becomes evident that there is a strong correlation between these factors. Specifically, variations in monthly average temperature during dormancy are closely linked to timing patterns in subsequent years' budburst stages. Secondly, ≥10℃ accumulated temperatures provide a complementary effect on mitigating low values recorded for monthly average temperatures during male inflorescence dormancy. This relationship is particularly notable between 2016's dormancy stage and 2017's budburst stage. In this instance, while January through March averages indicated a lower mean temperature of 5.9℃ during dormancy in 2016 compared to 8.9℃ recorded for similar months in 2015, an impressive total heat accumulation reached approximately 4033.4℃ over a span of just under seven months (195 days). However, the release of physiological dormancy in plants necessitates a certain accumulation of low temperatures. The inadequate accumulation of low temperatures in 2016 resulted in a delay in the budburst stage of male inflorescence during the spring of 2017. Ultimately, there exists a threshold value for the complementary effect between ≥10°C accumulated temperatures and the monthly average temperature throughout the dormancy stage of male inflorescence. In 2017, the monthly average temperature during this dormancy stage of male inflorescence was recorded at 6.9°C, while the ≥10°C accumulated temperature amounted to 3680.6°C. In contrast, although both the monthly average temperature and ≥10°C accumulated temperature increased in 2018 compared to 2017, no complementary relationship was observed that year. However, this complementary relationship re-emerged in subsequent years, thus indicating that there is indeed a minimum threshold necessary for such an interaction between accumulated temperatures and monthly average temperatures to occur. It is only when the monthly average temperature meets or exceeds this minimum threshold that this complementary effect becomes evident.
5. Discussion
5.1. Ecological Adaptation Strategies of B. microphylla in Spring
Leaf serves as a key organ for the direct observation of vegetation phenology, and extreme environmental changes are often reflected in the foliage of plants. Research conducted on pure forests within urban areas of Shanghai has explored how environmental variations influence the leaf unfolding phenology of trees [29]. The findings indicate that different tree species exhibit varying degrees of sensitivity to precipitation and temperature factors, with this disparity being particularly pronounced within the same species. Specifically, identical tree species can display markedly different phenological rhythms across diverse ecological environments. Phenological observations of Betula pendula distributed throughout the Irtysh River Basin reveal that fluctuations in temperature indicators during equivalent periods can lead to variations in the phenological events [30]. Moreover, there is a correlation between phenology and temperature across different habitats. Some researchers have initiated investigations into how abnormal climatic conditions affect the spring accumulated temperature requirements for various plant species. Their results suggest that the first leaf dates for all tested plants tends to be advanced or delayed during warmer or colder years, respectively [31]. However, differences observed between distinct climatic year are not statistically significant, indicating that accumulated temperature requirements also vary among species. Alterations in ecological adaptation strategies exhibited by plants under different thermal conditions have manifested through their respective phenophases.
The flowering phenology of plants primarily encompasses three key aspects: flowering date, the duration of the flowering, and the synchrony within the flowering period [32]. These factors can provide valuable insights into the reproductive status of plant organisms in advance. On one hand, stable temperatures during early spring are crucial for initiating plant growth phenology. On the other hand, sunlight duration significantly influence the flowering phenology of inflorescences. Phenological observations conducted on Betula albosinensis in Xi'an reveal a negative correlation between the blooming stage duration and hours of sunlight [33]. Specifically, as altitude increases and sunlight duration extends, there is a corresponding reduction in the length of the blooming stage. A comparison with relevant data from this study area indicates that both 2012 and 2016 experienced starting dates for ≥10℃ accumulated temperatures at the end of March, significantly earlier than previous years. Furthermore, total sunlight hours recorded were 2200.6h and 1924.6h respectively. Sunlight has a notable advance effect in budburst stage for female and male inflorescences' budburst stage. Consequently, all observed phenological events occurred earlier compared to other years. Finally, based on these observations, it was noted that pollen dispersal for B. microphylla occurs prior to its pollination stage. Moreover, pollination takes place within this pollen dispersal timeframe, indicating a high degree of synchrony between male and female inflorescences regarding their respective pollen dispersal and pollination behaviors overall.
The geographical element of B. microphylla is associated with its distribution in Central Asia. Considering the typical vegetation types characteristic of temperate grassland regions within this study area, the spring phenology of leaf exhibits a significant correlation with monthly average temperatures, and developmental trends across various leaf phenologies are consistent. Plant germination progresses through several stages: bud formation, establishment of dormancy, release from dormancy, and temperature-driven promotion prior to germination [34]. The response patterns at each stage to external temperature fluctuations vary. Additionally, differing energy allocation strategies influenced by temperature can affect the timing of phenological events [35]. Firstly, the female inflorescence enters the budburst stage during spring. A comprehensive analysis that considers factors such as accumulated temperature requirements, developmental stage types, and energy allocation phases indicates that there exists a low-temperature requirement for the budburst of female inflorescences. This represents a strategic trade-off made by B. microphylla in response to the perennial cold climate prevalent in this region. Secondly, the flower buds of B. microphylla are encased in leaf-like bud scales, and this species is characterized as a wind-pollinated plant that flowers prior to the unfolding of its leaf. This flowering strategy not only minimizes interference from foliage but also maximizes pollen availability for female inflorescences, thereby expediting the pollination process. Thirdly, a stable average monthly temperature during the germination stage across adjacent years significantly accelerates this process. This observation indicates that the phenology of female inflorescence budburst in B. microphylla is strictly regulated by temperature, and exhibits clear adaptive evolution in response to environmental conditions. Fourthly, B. microphylla completes both pollen dispersion and pollination before its leaf reach full maturity. The maturity of female fruit-clusters occurs between late May and mid-June, suggesting that B. microphylla employs a "rapid strategy" to swiftly complete pollination and fruiting under favorable conditions, which can mitigate adverse effects on fruit development and seedling establishment caused by unfavorable environments. Ultimately, these plants accumulate thermal energy to sustain their growth processes. Furthermore, energy allocation strategy of B. microphylla is notably distinctive, when accumulated temperatures are high, priority is given to utilizing available energy for breaking dormancy. Any remaining energy supports meristem development and facilitates entry into the female inflorescence budburst stage. Surprisingly, in years with low accumulated temperatures, there is a delay in female inflorescence budburst phenology as heat accumulation occurs during spring to ensure successful completion of various life processes within B. microphylla.
5.2. Ecological Adaptation Strategies of B. microphylla in Summer
The temperature rise in summer expedites the cell division process of B. microphylla. Leaf maturity stage and the elongation stage of male inflorescences persist throughout the entire summer and extend across different months, aligning with the climatic year variation trend. During the leaf yellowing stage, light influence the synthesis processes of enzymes and chlorophyll and other substances within plant leaf, either advancing or delaying the leaf yellowing stage. Generally speaking, the leaf phenology throughout the year is conspicuously affected by the climate. According to our division results, warmer year implies a higher average temperature in March and April, indicating a higher heat initiation value for that year. Hence, the female inflorescences enter the maturity and abscission stage earlier. Nevertheless, in a normal year, if the average temperature of a specific phenological month exhibits a markable high value, it will also accelerate the process of entering the fruit abscission stage. This result is consistent with Pei [36] that with a 1°C increase in temperature in March, the flowering phenology will be advanced by 2 days. Notably, the advancement of the flowering period and the maturity process of the fruit of B. microphylla, similar to some species of the same genus, have an obvious heat requirement.
The elevated temperatures experienced during summer, combined with continuous illumination and stable air flow velocities, result in the transpiration process reaching its zenith. However, B. microphylla, as a hygrophilous species has developed specific adaptation strategies to cope with these conditions. Firstly, the leaf of B. microphylla effectively captures light energy for photosynthesis, facilitating rapid leaf growth throughout the summer months. The surplus products of photosynthesis, after fulfilling the plant's respiratory demands are subsequently transported to storage organs. Secondly, the relatively shallow root system of B. microphylla allows it to efficiently utilize surface water resources to compensate for water loss due to transpiration. Such mechanism creates a water gradient that helps maintain a stable temperature within the plant. Finally, male inflorescences enter a critical phase of growth phenology. At this stage, male inflorescences are uniformly distributed along the branches. That is to say, a "quantity-dominant" strategy is evident, which involves maximizing essential resources such as light and heat for growth purposes in order to produce a greater number of male inflorescences available for pollination and subsequent fruit bearing.
5.3. Ecological Adaptation Strategies of B. microphylla in Autumn
The decline in temperature during autumn results in a decrease in the activity of enzymes associated with chlorophyll synthesis, consequently slowing down the rate of chlorophyll production while accelerating its decomposition. We observed the peak period for leaf abscission in B. microphylla occurs in early to mid-October, with most leaf being abscised by the end of October. Dormancy serves as a survival strategy employed by plants to conserve energy and withstand cold northern winters. However, the specific organs that enter dormancy can vary among species. Typically, leaf buds also undergo dormancy, which represents a temporary cessation of bud growth. In contexts such as controlled cultivation environments, leaf budburst is often utilized as an indicator for assessing dormancy release. Furthermore, for B. microphylla, the autumn temperature plays a crucial role in determining the dormancy phenology of male inflorescences.
Previous studies have focused on the native Torreya grandis and the non-native Carya illinoinensis in subtropical regions, revealing that elevated autumn temperatures exacerbate the dormancy depth of leaf buds in T. grandis. Consequently, this phenomenon leads to a postponement in budburst phenology [37,38]. The Betula is distributed across various climatic zones in China, with primitive and evolutionary groups primarily found in the southwestern region. This study also identified that under high autumn temperatures, the male inflorescence of B. microphylla delays its budburst onset for the next year. Our finding indicates that male inflorescences of B. microphylla adopt a "long dormancy" strategy to adapt to insufficient cold requirements resulting from a reduced range of cold exposure. Xinjiang serves as one of the differentiation centers for the Betula. [39], exhibiting a climate distinct from both the humid northeastern region and mountainous southwestern areas. This unique climate provides favorable environmental conditions for inflorescence growth. In Xinjiang, female inflorescences of B. microphylla acquire pollen within a brief timeframe to become fruit-clusters, and abscission. Additionally, this study observed an intriguing phenomenon: during the shaping stage of male inflorescences, some male inflorescences will abscise while remains at branch tips. In that respect, nutrient allocation towards remaining male inflorescences becomes more concentrated. More importantly, this growth pattern facilitates greater spatial availability for female flower buds along branches, demonstrating what can be termed as a "spatial dislocation effect". From the perspective of the unique reproductive patterns exhibited by male inflorescences, a complementary phenomenon arises between ≥10°C accumulated temperatures and low average monthly temperatures during the dormancy stage of these inflorescences. Elevated autumn temperatures lead to a delay in the onset and end-date of low-temperature accumulation, which can also be interpreted as a complementary effect exerted by average monthly temperatures on accumulated temperature.
5.4. Ecological Adaptation Strategies of B. microphylla in Winter
During winter, all perennial trees in northern regions enter a state of dormancy [40], and B. microphylla is no exception. This dormancy results in decreased water content within the plant, which prevents intracellular water from freezing and thereby protects the cell nucleus from damage. The presence of anti-freeze proteins (AFPs) equip these plants with enhanced resilience against extreme cold conditions, improving their tolerance to low temperatures and safeguarding cells from cold-induced injuries. Recent studies have successfully isolated and identified AFPs from various cold-resistant species. Comparative analyses indicate that the recrystallization inhibitory activity of AFPs found in plants surpasses that observed in insects [41]. The manifestation of winter dormancy behavior in B. microphylla, along with its successful completion of growth phenology in subsequent years, suggests an underlying regulatory mechanism involving AFPs within this species.
This research area is specifically located in the semi-arid typical steppe region of the Altai Mountains in Xinjiang. The terrain and climate here significantly differ from the northeast and southwest. Low temperatures and limited sunlight duration during winter are among the factors contributing to plant dormancy. Bud scales (Figure 9.), which are modified leaf that encase buds for protective purposes. They are thickened and coated with wax, often enabling most perennial woody plants to withstand extreme climatic conditions such as severe cold and drought. During winter, although male inflorescences of B. microphylla remain intact without shedding, the emergence of bud scales signifies its initial adaptation to local cold and dry climatic conditions, placing it in an "adaptive evolution" stage. For plant organisms, not all trade-offs manifest above ground. Indeed, the developed root traits of B. microphylla optimizes water resource utilization provided by wetlands while also preventing soil erosion and mitigating soil and water loss [42].
6. Conclusions
Based on nearly 10 years of phenology observations, we analyzed the relationship between temperature and 18 phenophases of B. microphylla located in the southern margin of the Junggar Basin and have conducted a systematic study on the ecological adaptation strategies of B. microphylla to seasons variation. Our study also revealed the differences between the phenological response of warmer years and colder years to temperature. The results indicated that (1) The seasonal phenological patterns of B. microphylla exhibit distinct variations. Moreover, the types of phenophases across different seasons remain relatively stable while their durations show minimal variation. (2) The primary influencing factor for phenological variations is the monthly average temperature during specific phenological events. Throughout the year, leaf phenology is predominantly influenced by the monthly average temperature as well as the differences in climatic year. In spring, the budburst of female inflorescences is associated with accumulated temperature, the type of budburst stage, and energy allocation stages. Conversely, high temperatures experienced during plant’s dormancy stage may delay the budburst of male inflorescences. (3) A comprehensive and scientific theory regarding ecological strategies for B. microphylla has been clearly articulated based on trait analysis and climatic data. The impact of climate on the phenology of B. microphylla can be attributed to five key strategies: a "multiple low-temperature requirements" strategy for inflorescence budburst; a "preceding leaf growth" strategy concerning inflorescences; a "quantity victory" strategy for male inflorescences; a "long dormancy" strategy applicable to male inflorescences under elevated autumn temperatures; and a "spatial dislocation" strategy pertaining to both male and female inflorescences. Xinjiang, located in the northwest and remote from the ocean, exhibits an exceptionally intense continental climate characterized by terrains such as grasslands and deserts. Currently, the rational exploitation, utilization, and protection of native natural forest resources in Xinjiang hold paramount importance. Our findings will effectively provide valuable insights into research on the response of B.microphylla to climate change. We hope to draw attention to the significance of narrow-distributed endemic species through a case study of B. microphylla phenology and adaptation strategies.
Author Contributions
Conceptualization, Y.W.; software, Y.W.; writing-original draft preparation, Y.W.; writing-review and editing, Y.W.; data curation, X.L.; supervision, Z.D., G.H., W.X., Z.M., and P.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Natural Science Foundation of China (Grant No. 32160317) and Foundation of Shihezi University (Grant No. ZZZC201930B).
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to privacy.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ryu, Y.; Sonnentag, O.; Toomey, M. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system. Agricultural and Forest Meteorology, 2013, 169, 156-173. [CrossRef]
- Tao, Z.X.; Ge, Q.S.; Xu, Y.J.; Wang, H.J. Comparison of changes in flowering phenology of woody plants and temperature sensitivity between Xi′an and Baoji. Acta Ecologica Sinica, 2020, 40, 3666-3676.
- Elsa, E.C.; Isabelle, C.; Annette, M.; Harold, A.M.; Mark, D.S. Shifting plant phenology in response to global change. Trends in Ecology & Evolution, 2007, 22, 357-365.
- Fang, X.Q.; Chen, F.H. Plant phenology and climate change. SCIENCE CHINA Earth Sciences, 2015, 58, 1043-1044.
- Menzel, A.; Sparks, T.H.; Estrella, N.; Koch, E.; Aasa, A.; Ahas, R.; Alm-Kübler, K.; Bissolli, P.; Braslavská, O.g.; Briede, A.;et al. European phenological response to climate change matches the warming pattern. Glob. Chang. Biol. 2006, 12, 1969–1976.
- Laube, J.; Sparks, T.; Estrella, N.; Höfler, J.; Ankerst, D.; Menzel, A. Chilling outweighs photoperiod in preventing precocious spring development. Glob. Chang. Biol. 2014, 20, 170–182. [CrossRef]
- Peng, J.; Wu, C.; Zhang, X.; Wang, X. Satellite detection of cumulative and lagged effects of drought on autumn leaf senescence over the Northern Hemisphere. Glob. Chang. Biol. 2019, 25, 2174–2188. [CrossRef]
- Korner, C.; Basler, D. Phenology Under Global Warming. Science, 2010, 327, 1461-1462. [CrossRef]
- Schwartz, M.D.; Rein, A.; Aasa, A. Onset of spring starting earlier across the northern hemisphere. Glob Change Biology, 2006, 12, 343-351. [CrossRef]
- Deborah, Z.; Thomas, W.C.; Lidong, M.; Susanne, S.R.; Constantin, M. Increased growing-season productivity drives earlier autumn leaf senescence in temperate trees. Science, 2020, 370, 1066-1071.
- Basler, D.; Körner, C. Photoperiod and temperature responses of bud swelling and bud burst in four temperate forest tree species. Tree Physiol, 2014, 34, 377-88. [CrossRef]
- McIntyre, S.; Lavorel, S.; Landsberg, J.; Forbes, T.D.A. Disturbance response in vegetation towards a global perspective on functional traits. JOURNAL OF VEGETATION SCIENCE, 1999, 10, 621-630. [CrossRef]
- Norby, R.J.; Jackson, R.B. Root dynamics and global change: seeking an ecosystem perspective. THE NEW PHYTOLOGIST, 2000, 147, 3-12. [CrossRef]
- Westoby, M.; Falster, D.S.; Moles, A.T.; Vesk, P.A.; Wright, I.J. Plant ecological strategies: Some leading dimensions of variation between species. ANNUAL REVIEW OF ECOLOGY AND SYSTEMATICS, 2002, 33, 125-159. [CrossRef]
- Zhao, Y.P.; Liu, J.F.; Wang, Q.; Huang, R.Z.; Sun, J.Y.; Nie, W.; Yang, S.W. Differences in the response of different vegetation types to frost event and their driving forces. Acta Ecologica Sinica, 2024, 44, 1-14.
- Liao, Y.Y.; Zhang, X.L. Differences and influencing factors between xylem growth phenology and vegetation phenology in Saihanba area. Forestry and Ecological Sciences, 2024, 39, 153-160+168.
- Zhu, N.; Jiang, Hong.; Jin, Y.Y. A phenology study on the common tree species of natural secondary forests in northeast China. Acta phytoecologica et geobotanica sinica, 1990, 14, 336-349.
- Luan, K.Z.; Fan, X.F.; Wang, W.F. Difference in Leaf-Fall phenology between Betula platyphylla and Populus davidiana and its ecological significance. 2012, 40, 20-22.
- Xu, K.J. Identifying regional dominant forest tree species based on multi-scale spatial and temporal characteristics of Chinese high-resolution earth observation satellite data[D]. Nanjing: Nanjing University, 2019.
- Huang, G.; Du, Z.Z.; Sun, X.N.; Cao, T.; Yan, P. Life Table and Survival Analysis of Betula microphylla Population on the Southern Margin of Junggar Basin. Jiangsu Agricultural Sciences. 2012, 40, 310-313.
- Huang, G.; Yan, P.; Du, Z.Z.; Cao, T.; Xu, W.B. Inflorescence growth phenology and ecological adaptation of Betula microphylla. Acta Botanica Boreali-Occidentalia Sinica, 2014, 34, 2569-2575.
- Ahmed, G.; Zan, M. The influence of heat island effect on vegetation phenology in Major Urban Cluster in the Tianshan northslope economic belt of Xinjiang. Journal of ecology and rural environment, 2022, 38, 872-881.
- Wang, Y. Relationship between flowering phenology and atmospheric environmental factors of woody plants in Xinjiang[D]. Urumqi: Xinjiang University, 2021.
- Wan, M.W.; Liu, X.Z. China’s National Phenological Observational Criterion[M]. Beijing: Science Press, 1979, 15-58.
- Zhu, K.Z.; Wan, M.W. Phenology[M]. Changsha: Hunan Education Publishing House, 1979, 1-58.
- Wan, M.W. How to observe phenology[M]. Beijing: Beijing Publishing house, 1964, 17-33.
- Li, H.Y.; Wang, X.T.; Zheng, X.Q.; Xu, H.Y.; Xiang, D.; Zhu, J.L. Response of woody plants to climate warming during phenological period in No. 8 division of Xinjiang Production and Construction Crops. Chinese Agriculture Science Bulletin, 2021, 37, 58-63.
- Rathcke, B.; Lacey, E.P. Phenological patterns of terrestrial plants. Annual Review of Ecology & Systematics, 1985, 16, 179-214.
- Zhang, Y.R.; Pang, Y.L.; Luo, X.Y.; Xu, J.Y.; Yang, Y.Y.; Da, L.J.; Song, K. Driving factors of leaf-unfolding phenology in deciduous trees in Shanghai. Journal of East China Normal University(Natural Science), 2023, 6, 125-133.
- Liu, X.L.; Zhang, Z.D.; Xue, Z.F.; Zeng, S.J.; Liu, T. Plant diversity characteristics of forests in Haba River Valley, a tributary of Irtysh River. Journal of plant resources and environment, 2024, 33, 89-96.
- Wang, H.J.; Tao, Z.X.; Ge, Q.S. Effects of climate variation on the first leaf dates of 39 woody species and their thermal requirements in Xi’an, China. Chinese Journal of Plant Ecology, 2019, 43, 877-888. [CrossRef]
- Wright, J.W.; Meagher, T.R. Pollination and seed predation drive flowering phenology in Silene latifolia (Caryophyllaceae). Ecology, 2003, 84, 2062-2073. [CrossRef]
- Li, N. Preliminary study on floral phenology, pollen characteristics and hybrid breeding of Betula albosinensis. Yangling, Northwest A&F University, 2023.
- Yang, Q.S.; Gao, Y.H.; Wu, X.Y.; Moriguchi, T.; Bai, S.L.; Teng, Y.W. Bud endodormancy in deciduous fruit trees: Advances and prospects. Horticulture Research, 2021, 8, 1-11. [CrossRef]
- Chen, X.Q.; Wang, L.X.; Inouye, D. Delayed response of spring phenology to global warming in subtropics and tropics. Agricultural & Forest Meteorology, 2017, 234-235, 222-235. [CrossRef]
- Pei, S.X.; Guo, Q.S.; Xin, X.B.; Liu, W.; Hong, M. Response of 4 common broad-leaved arbors phenology to climate change in the northeast China. Scientia Silvae Sinicae, 2011, 47, 181-187.
- Wang, F.C.; Zhang, R.; Lin, J.H.; Zheng, J.B.; Hanninen, H.; Wu, J.S. High autumn temperatures increase the depth of bud dormancy in the subtropical torreya grandis and carya illinoinensis and delay leaf senescence in the deciduous carya. Trees, 2022, 36, 1053-1065. [CrossRef]
- Liu, G.H.; Chen, X.Q.; Zhang, Q.H.; Lang, W.G.; Delpierre, N. Antagonistic effects of growing season and autumn temperatures on the timing of leaf coloration in winter deciduous trees. Glob Change Biology, 2018, 24, 3537–3545. [CrossRef]
- Jiang, J.M. The study of the geographical distribution of the Betula in China. Forest Research, 1990, 3, 55-62.
- Zhang, C.H. The research of breaking dormancy and germination mechanism of Zelkova schneideriana seeds. Taian, Shandong Agricultural University, 2018.
- Dong, X.Y.; Wei, J.P.; Cui, J.M.; Wu, Z.F.; Zheng, G.Q.; Li, H.; Wang, Y.; Tian, H.Y.; Liu, Z.G. Research progress in plant antifreeze protein. Chinese Bulletin of Botany, 2023, 58, 966-981.
- Wang, Y.X.; Liu, J.M., Du, Z.Z.; Xu, W.B.; Ma, Z.C.; Huang, G.; Yan, P. Causes of rapid degradation of Betula microphyla population in the southern margin of Junggar basin and protection strategies. Acta Ecologica Sinica, 2024, 44,1-10.
Figure 1.
Overview of the study sites. (a) Geographical locations of weather station and study area in China. (b) Days of ≥10℃ accumulated temperature start and end date during 2009–2019. (c) ≥10℃ accumulated temperature during 2009–2019. (d) Annual sunshine duration during 2009-2019. (e) Annual precipitation during 2009-2019.
Figure 1.
Overview of the study sites. (a) Geographical locations of weather station and study area in China. (b) Days of ≥10℃ accumulated temperature start and end date during 2009–2019. (c) ≥10℃ accumulated temperature during 2009–2019. (d) Annual sunshine duration during 2009-2019. (e) Annual precipitation during 2009-2019.
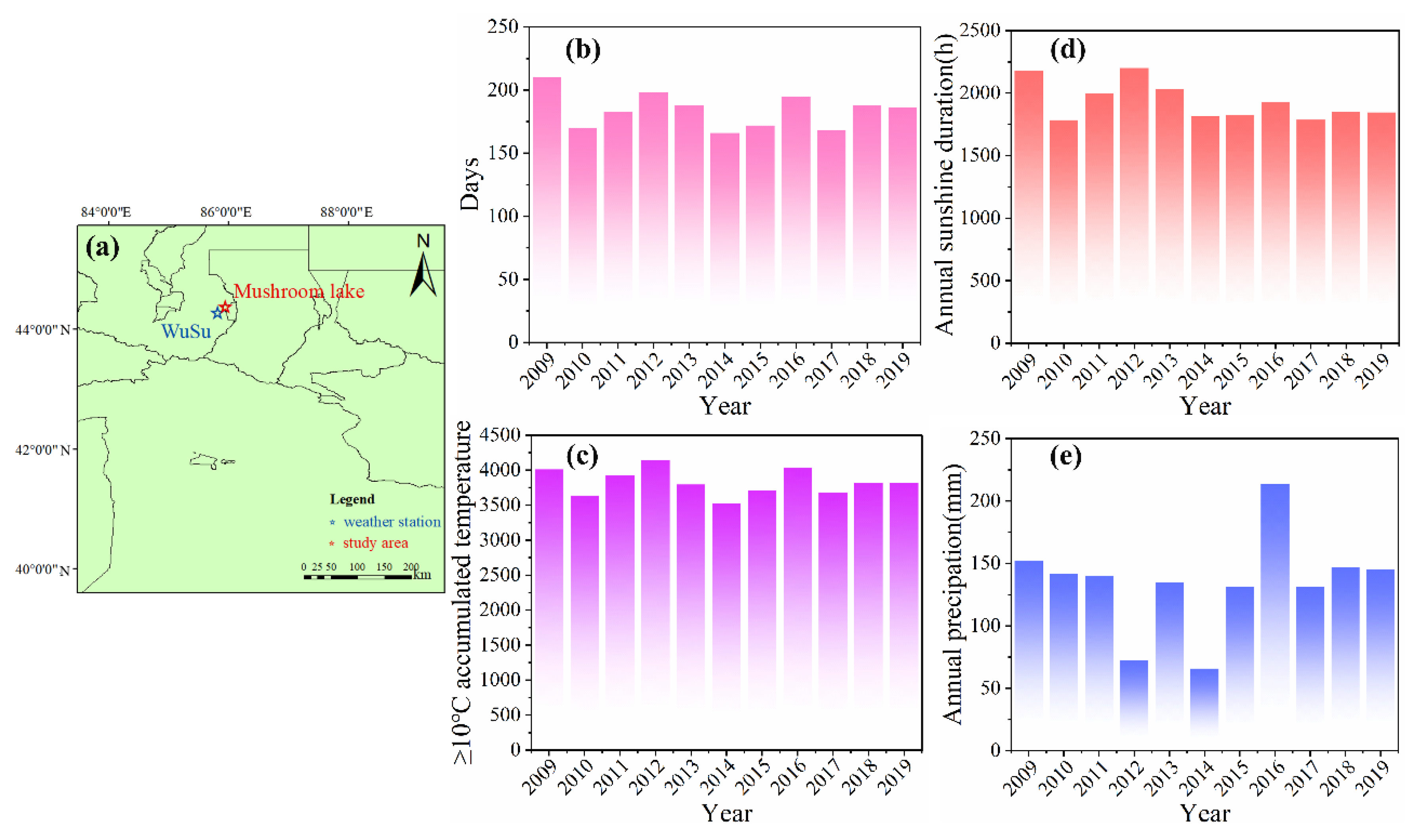
Figure 2.
leaf and inflorescence phenology of B. microphylla. (a) leaf flush stage. (b)young leaf stage. (c)leaf unfolding stage. (d)leaf maturity stage. (e)leaf yellowing stage. (f)leaf abscission stage. (g) male inflorescence budburst stage. (h) male inflorescence elongation stage. (i) male inflorescence shapinging stage. (j) male inflorescence dormancy stage. (k) male inflorescence flowering phenology-budburst stage. (l) female inflorescence budburst stage. (m) male inflorescence flowering phenology-pollination stage. (n) male inflorescence flowering phenology-pollen dispersal stage. (o) young female fruit-cluster stage. (p) male inflorescence flowering phenology-withering stage. (q) female fruit-cluster maturity stage. (r) female fruit-cluster abscission stage. (s) during autumn, male inflorescences will abscise while remains at branch tips. (t) male inflorescences at branch tips.
Figure 2.
leaf and inflorescence phenology of B. microphylla. (a) leaf flush stage. (b)young leaf stage. (c)leaf unfolding stage. (d)leaf maturity stage. (e)leaf yellowing stage. (f)leaf abscission stage. (g) male inflorescence budburst stage. (h) male inflorescence elongation stage. (i) male inflorescence shapinging stage. (j) male inflorescence dormancy stage. (k) male inflorescence flowering phenology-budburst stage. (l) female inflorescence budburst stage. (m) male inflorescence flowering phenology-pollination stage. (n) male inflorescence flowering phenology-pollen dispersal stage. (o) young female fruit-cluster stage. (p) male inflorescence flowering phenology-withering stage. (q) female fruit-cluster maturity stage. (r) female fruit-cluster abscission stage. (s) during autumn, male inflorescences will abscise while remains at branch tips. (t) male inflorescences at branch tips.

Figure 3.
Interannual variation trend of leaf phenology of B. microphylla. (a) leaf flush stage. (b)young leaf stage. (c)leaf unfolding stage. (d)leaf maturity stage. (e)leaf yellowing stage. (f)leaf abscission stage.
Figure 3.
Interannual variation trend of leaf phenology of B. microphylla. (a) leaf flush stage. (b)young leaf stage. (c)leaf unfolding stage. (d)leaf maturity stage. (e)leaf yellowing stage. (f)leaf abscission stage.
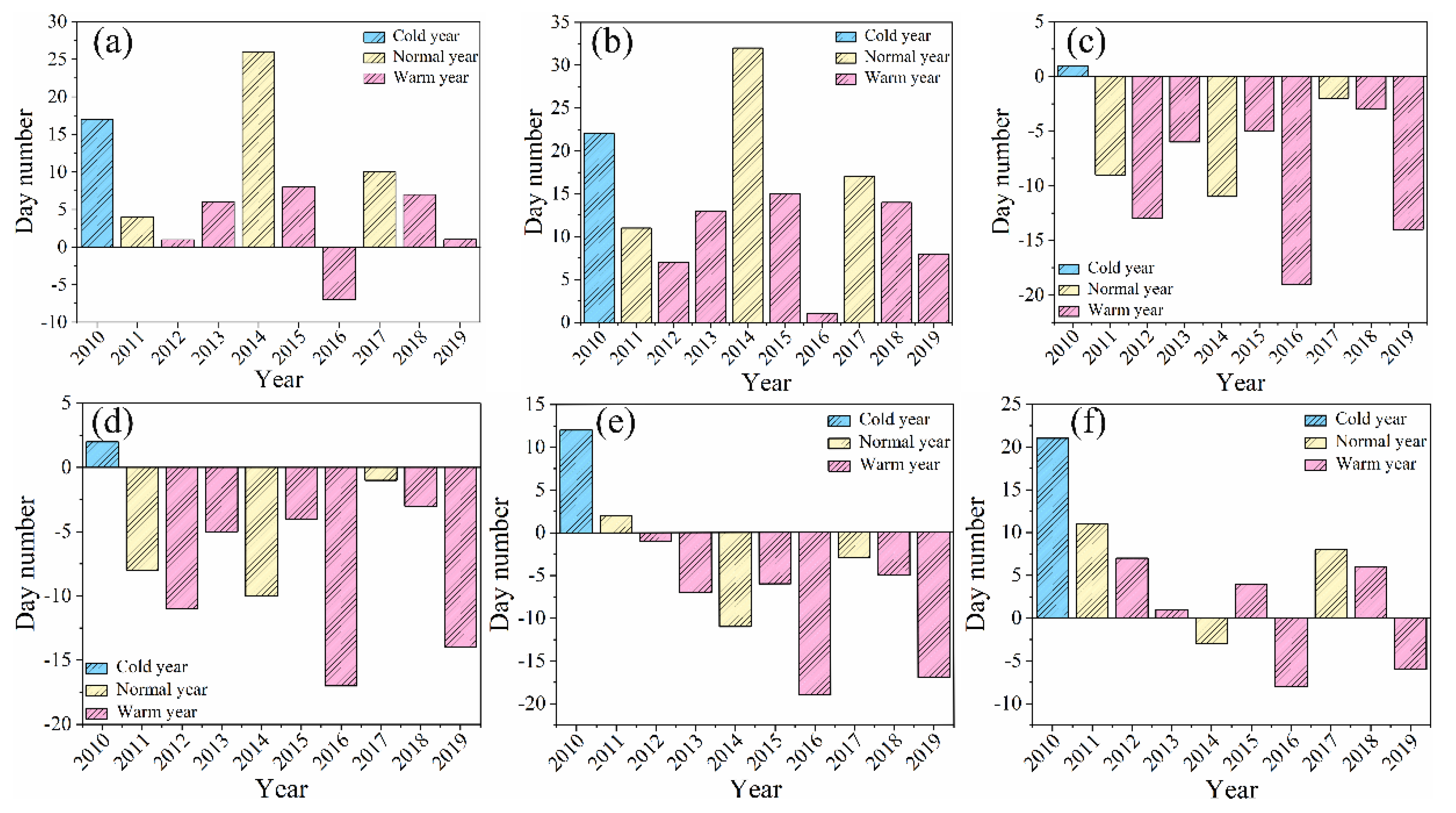
Figure 4.
Interannual variation trend of female inflorescence phenology of B. microphylla. (a) budburst stage-young fruit stage. (b)Female fruit-clusters maturity stage. (c) Female fruit-clusters abscission stage.
Figure 4.
Interannual variation trend of female inflorescence phenology of B. microphylla. (a) budburst stage-young fruit stage. (b)Female fruit-clusters maturity stage. (c) Female fruit-clusters abscission stage.
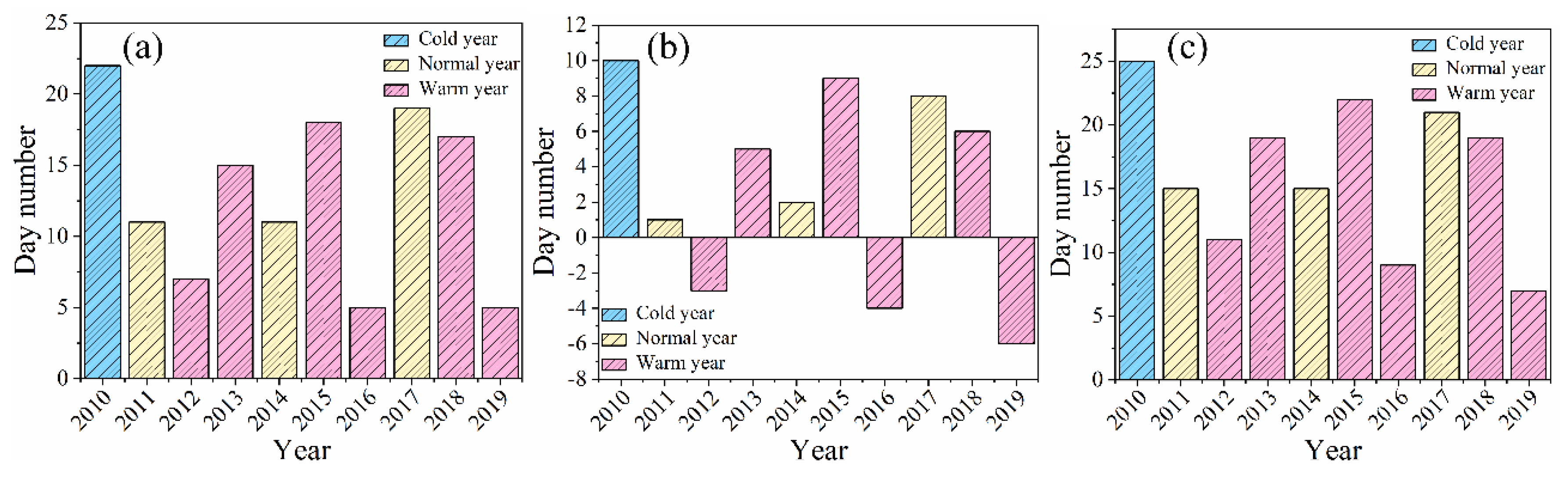
Figure 5.
The relationship between the monthly average of female inflorescence budburst, accumulated temperature and the beginning of female inflorescence budburst.
Figure 5.
The relationship between the monthly average of female inflorescence budburst, accumulated temperature and the beginning of female inflorescence budburst.
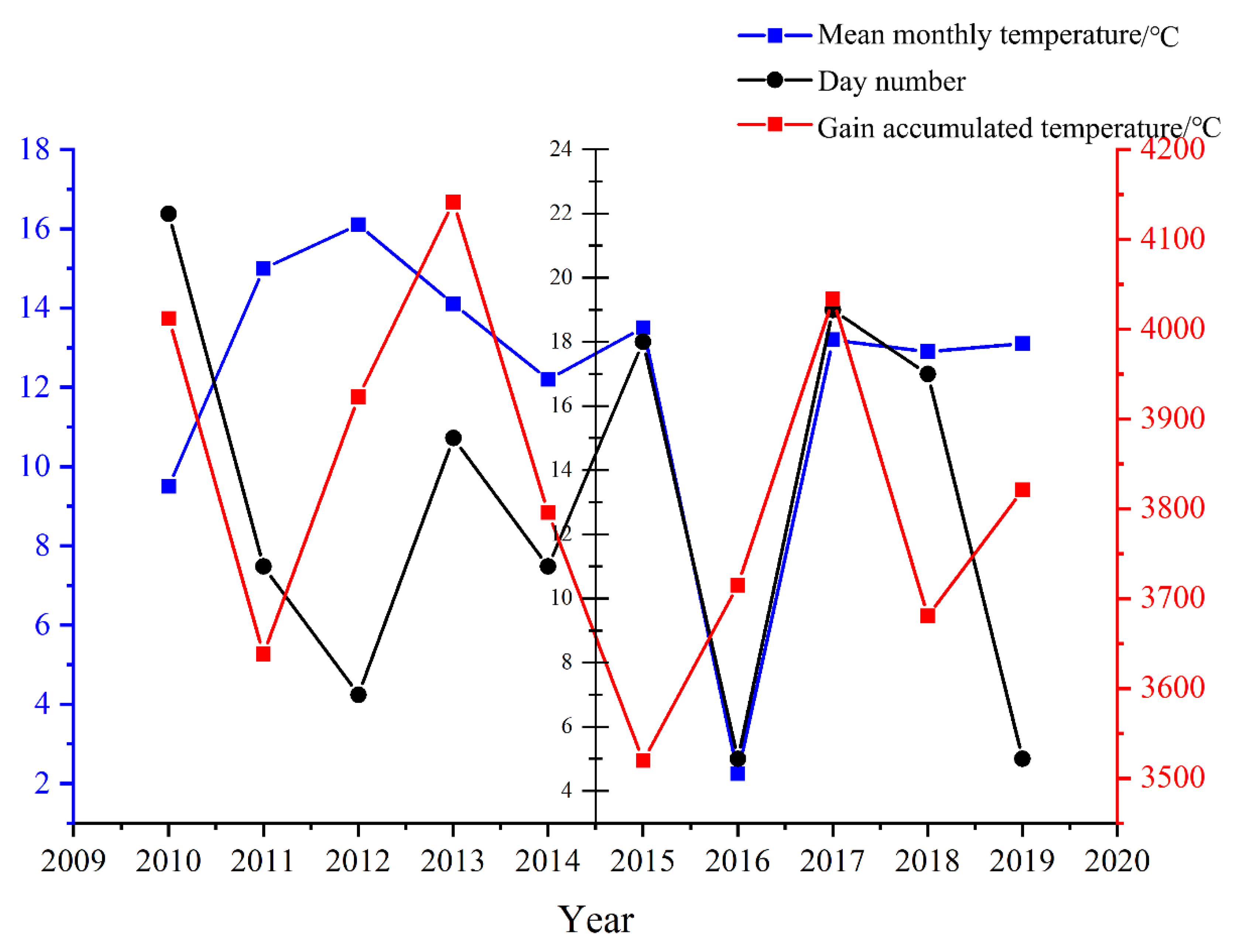
Figure 6.
Interannual variation trend of male inflorescence growth phenology of B. microphylla. (a) male inflorescence budburst stage. (b) male inflorescence elongation stage. (c) male inflorescence shapinging stage. (d) male inflorescence dormancy stage.
Figure 6.
Interannual variation trend of male inflorescence growth phenology of B. microphylla. (a) male inflorescence budburst stage. (b) male inflorescence elongation stage. (c) male inflorescence shapinging stage. (d) male inflorescence dormancy stage.

Figure 7.
Interannual variation trend of male inflorescence flowering phenology of B. microphylla. (a) male inflorescence budburst stage. (b) male inflorescence elongation stage. (c) pollen dispersal stage. (d) withering stage. (e) abscission stage.
Figure 7.
Interannual variation trend of male inflorescence flowering phenology of B. microphylla. (a) male inflorescence budburst stage. (b) male inflorescence elongation stage. (c) pollen dispersal stage. (d) withering stage. (e) abscission stage.
Figure 8.
The relationship between the dormancy monthly temperature of male inflorescences, ≥10℃ accumulated temperature and the budburst stage of male inflorescences.
Figure 8.
The relationship between the dormancy monthly temperature of male inflorescences, ≥10℃ accumulated temperature and the budburst stage of male inflorescences.
Figure 9.
The bud scale of B. microphylla. (a) bud scale on tree branch. (b-c) the bud scale gradually expand, allowing the leaves and inflorescence to appear. (d) growth strategy of inflorescence in B. microphylla-“growth before leaves”.
Figure 9.
The bud scale of B. microphylla. (a) bud scale on tree branch. (b-c) the bud scale gradually expand, allowing the leaves and inflorescence to appear. (d) growth strategy of inflorescence in B. microphylla-“growth before leaves”.


Table 1.
The average temperature from March to April 2010 to 2019 in the study area.
| Year | The average temperature in March(℃) | The average temperature in April(℃) | The average temperature in March-April(℃) | ≥10℃ accumulated temperature |
|---|---|---|---|---|
| 2010 | -5.8 | 9.5 | 1.85 | 3638.3 |
| 2011 | -5 | 15 | 5 | 3924.8 |
| 2012 | 0.8 | 16.1 | 8.45 | 4141.3 |
| 2013 | 5.6 | 14.1 | 9.85 | 3795.7 |
| 2014 | -0.9 | 12.2 | 5.65 | 3519.5 |
| 2015 | 2.1 | 13.5 | 7.8 | 3715 |
| 2016 | 2.25 | 15.3 | 8.775 | 4033.4 |
| 2017 | -2.2 | 13.2 | 5.5 | 3680.6 |
| 2018 | 7.4 | 12.9 | 10.15 | 3820.9 |
| 2019 | 7.9 | 13.1 | 10.5 | 3812.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



