
Submitted:
15 March 2025
Posted:
17 March 2025
You are already at the latest version
Abstract
Population-specific variations in folate metabolism, influenced by evolutionary pressures, challenge the universal application of standardised dietary folate recommendations. This paper examines historical folate intake, the impact of fortification, and the role of a metabolic switch in nucleotide synthesis, proposing that mandatory fortification may pose risks for populations unadapted to high folic acid exposure. Some diets, specifically the diet of Indigenous Australian’s, were historically rich in animal protein, providing sufficient natural folate. The transition to fortified foods, has led to excessive folic acid intake, potentially resulting in toxicity with epigenetic consequences. Additionally, formula-fed infants receive significantly higher folic acid levels than breastfed infants, compounded by maternal supplementation and processes foods being fortified, also raising concerns about excessive exposure. Emerging technologies, including biophysical modelling and deep learning, offer promising avenues for personalised nutrition and precision healthcare.
Keywords:
folic acid fortification
; epigenetics
; neurotoxicity
; personalised
; nutrition
; dietary folate
; metabolic switch
; breastfeeding and supplementation
; one-carbon metabolism
; public health policy
1. Introduction
The study of epigenetics underscores the important role of environment, lifestyle, and diet in gene expression and the biochemical evolution of specific population groups. It is understood that the healthiest populations of animals and humans are relatively void of environmental, dietary and lifestyle pressures. Religious practices are the most common reason for adhering to traditional dietary practices. Kosher food products account for up to a third of supermarket grocery products (Shurtleff W, Aoyagi A 2014).
The biochemical differences between individuals and populations, challenge the standardised "one-size-fits-all" approach to dietary recommendations and food fortification. Yet, the rapid amalgamation of population groups, makes personalised medicine difficult without biochemical and genetic testing (Sedley L 2023)
Genetic testing and its analysis are still in their infancy; we are only beginning to understand the some of the complex mechanisms of the genome and how the epigenome controls its functions (Sedley L 2023). Human evolution could only be partially explained by Darwin’s theory of positive selection (Kim Y, Bertagna F et al 2021), with epigenetic mechanisms being a precursor or evolutionary prerequisite (Sedley L 2023).
The selection of genes responsible for human diversion have been essential for the survival of our species throughout changes to environments, diets, and lifestyles (Hecht S, Shlaer S et al 1942). A mutation that arises in a population providing a survival advantage is known as a selective sweep. Sweeps can be visualised by sequencing the genome of a large portion of a geographical population. The beneficial mutations sweep throµgh populations leaving behind a trail of reduced genetic diversity, with the individuals who carry it, being more likely to survive and reproduce. In populations who depend on cereal grains for sustenance show selection of genes associated with enhanced metabolism of starch (Hancock AM, Witonsky DB et al 2010). Mutational defects in the Human β Globin (HBB) gene are the most common single gene disorders in human (Kiefer CM, Hou C et al 2008). Individuals who carry one copy the mutation are resistant to malaria and confer a selective advantage in malaria endemic regions. The prevalence of this trait is as high as 50% in some African populations, most of which were acquired somewhere between 5000-10000 years ago (Hedrick PW 2011). In contrast, and due to prolonged isolation, the usual trends of selective adaptation are not yet visible in some rural Indigenous Australian populations (Valeggia CR, Snodgrass JJ 2015).
Animal protein as a dietary preference, was a driving force in human evolution, particularly the evolution of the human brain (Armelagos GJ 2014). In contrast, the choice to revert to plant consumption has been associated with devolution and the reversal of changes to facial and dental structures (Hooton EA 1946)
John Harvey Kellogg promoted eugenics through dietary interventions aimed at enhancing the healthy and intelligence, while suppressing undesirable traits (Wilson, 2014). However, evidence suggests that global health and IQ levels have declined, instead (Lynn & Harvey, 2008). Factors such as elective abortion laws (The Editors of the Council on Foreign Relations, 2023), and medical interventions preventing natural foetal termination, may have contributed to this trend (George, 2002).
1.1. Folate
Folate has several therapeutic applications, and dose dependant toxicities have been reported. Folate is well known for its role in modulation of endogenous methylation (CH₃) through one carbon metabolic (1CM) inhibition. However, due to cell-specific epigenetic variations, it remains challenging to predict which organs or tissues may be affected by a reduction in endogenous methylation, potentially leading to pathological consequences due to epigenetic rigidity. While the neurotoxicity of folates is often attributed to its polyglutamate tail, other contributing factors may include purine synthesis feedback inhibition, modulation of the purinergic system, and epigenetic inflexibility (Sedley L 2023).
In 2005, the average daily folic acid intake for women aged 16–44 years, was 108 micrograms (µg) in Australia and 62 µg in New Zealand—both significantly lower than the recommended 400 µg per day (Australian Institute of Health and Welfare 2011).
Today, the recommended daily dose of folic acid for a pregnant or breastfeeding woman is 600-800µg (Organization of Teratology Information Specialists (OTIS) 2024), with standard pregnancy and post-natal supplements containing around 500µg (Elevit 2025).
The rising incidence of neural tube congenital disease (NTD) and spontaneous abortion led to the mandatory fortification of grains with folate, in many countries (Williams J 2015). The risk of NTD is rare; in 2003, New Zealand, 63 pregnancies were affected by NTDs, at a rate of 11.2 per 10,000 births; in 2005, Australia recorded 149 pregnancies affected by NTDs, with an incidence rate of 13.3 per 10,000 births across most Australian regions (Australian Institute of Health and Welfare 2011). In China, 10 in 10,000 births were affected (Avagliano L et al 2019). The global risk of spontaneous abortion was 0.49 in 1,000 (Li et al 2024). Since fortification, the prevalence of NTD in America (Williams et al 2015) and China have reduced over 50% (Avagliano L 2019).
From a bio-unique perspective, the study of epigenetics and the understanding of health management, tends to contradict traditional medicine. For example, The World Health Organisation currently recommends modulation of epigenetic mechanisms by folate, during pregnancy, for the prevention of NTD. However, like all epigenetic processes, we now understand that neural tube closure, requires an infinite fluctuation of epigenetic mechanisms, and an interruption to epigenetic mechanisms, in one cell, during neural development, may result a domino effect of epigenetic pressure in an alternative cell or organ system, and may result in an alternative neurological disease.
The rising incidence of autism spectrum disorder has been proposed to be a result of such mechanisms (Sedley 2020). The process is like applying a band aid to a broken leg, the repair of a major structural disease, is likely to contribute to alternative non-structural adaptations, which may be pathological.
The incidence of autism spectrum disorder is now 1 in 100 live births (World Health Organization 2023).
2. Methods
Folate concentrations of various global food staples and daily rations used throughout history, were extracted from the Australian Food Composition Database, Food Standards Australia and New Zealand (FSANZ 2020), or alternative scholarly databases where necessary. This data was analysed to determine the potential toxic effects of fortification, based on potential evolutionary factors.
2.1. Limitations
Food Standards Australia and New Zealand (FSANZ) (2020) are unable to detect folate levels under 3µg/100g. Therefore, reference values from alternative scholarly databases were utilised instead.
Vitamins in food products are known to degrade following exposure to light and heat. This may affect dietary estimations.
2.2. Estimated Servicing Sizes and Justification
2.2.1. Potato
Extraordinary soil fertility in 18th century Ireland, provided potato as a staple for the population. As a result of a famine, this produce was rationed and consumed by much of the population (Pensary LA 1903). To gain insight into folate intakes throughout history, the folate concentration of a large potato was quantified per meal.
2.2.2. Milk
Rios-Leyvraz M, Yao Q (2023) suggests an infant can consume anywhere from 600 to 800ml of milk per day, depending on the size of the infant’s stomach. Folate values for a daily serving of 624ml of breastmilk, which is suitable for a 1-month-old, was compared to the recommended 2 scoops of (Karicare Infant Formula, 0-6 months) in 100ml of water; 6.5 servings per day; for the same age infant.
2.2.3. Whole Grain and Hulled Wheat, Rice and Maize
Grain staples have been feeding populations for tens of thousands of years, with cultivation of wheat occurring over 10,000 years ago (Haas et al (2018). Hulled and wholegrains were compared, as polished rice only became a staple in Asia in the 1930’s (World Health Organisation 1999). Portion sizes were based on a bowl or 250g of cooked grain, which was calculated to equate to 60g of dry mass, per meal.
2.2.4. Animal Protein Estimates
Evidence of the Indigenous Australian’s diet was documented in 1803; animal produce was highly preferred; consuming very little seasonal roots and fruit (John Morgan 1979). Like the Inuit population, they consumed a high calorie diet which included a large portion of meat to sustain significant daily physical activity (Kommissionen for Ledelsen af de Geologiske og Geografiske Undersøgelser i Grønland, Meddelelser om Grønland 1915). These semi-carnivorous diets were used to evaluate the folate content of animal meat.
Although these populations were likely to have consumed a daily portion size greater; 600g of animal meat, was used to compare. This portion size can be comparable to animal meat consumed by the active carnivorous population, today (Australian Carnivores 2025). A variety of animal meats where compared, kangaroo, chicken, beef, lamb to get an average.
Given that approximately 10% of an animal is composed of offal or organs, hunter’s may have consumed an equal portion of offal, and therefore a value 540g animal protein and 60g of lamn offal was included.
Rations of offal were spared for the peasants throughout the renaissance (Albala K 2002), and despite the poor evolutionary adaptation and high prevalence of haemochromatosis, high iron offal-based meals, like black pudding, haggis and umble pie (The Editors of the Online Etymology Dictionary 2023), are considered national dishes in Northern European countries. This is important to consider, since many expatriate Australians, are derived from this region. Although likely to have consumed less through famine, 400g per day of offal was used to compare.
2.2.5. Breads and Fortified Foods
In 19th century Australia, Church missions were established on land granted to them, to house and train Indigenous Australians on the Christian/Adventist or Presbyterian way of life (Shannon C 2002) (Clapham N 1985) (Wyndham C, H et al 1964), promoting vegetarianism and providing staples of flour and sugar (Sherriff S, Kalucy D et al 2022). Today, many Indigenous Australians consider white bread, and hot potato chips their cultural foods. Food insecurity effects between 20 and 25% of Indigenous Australians; many feeding their families solely on a loaf of fortified bread (Sherriff S, Kalucy D et al 2022).
Fortified bread was calculated with the same mass as wholegrains; 60g of dry Wonder White bread equates to two slices per meal. Adult daily portion sizes equate to 180g of dry mass. The use of commonly consumed yeast spread, Vegemite, was calculated at 5g per meal.
2.2.6. Cereal
Again in the19th century, an American Presbyterian temperance preacher Sylvester Graham promoted a strict grain based vegetarian diet to aid in reducing human animalistic instincts (Graham S 1877) (Wilson BC 2014). John Harvey Kellogg promoted Graham’s whole wheat products at his Battle Creek Adventist wellness facility, Sanitarium, for the same reason (Wilson BC 2014). In renaissance physiology, nutrition was directly associated with sexual desire (Albala K 2002). John Harvey Kellogg believed animal produce also contributed to dental decay which would eventually lead to a toothless race (Kellogg JH 1899). With that, Kellogg promoted the substitution of an animal-based breakfast, with cereal (Wilson BC 2014).
The Sanitarium's dietetic nurse Lenna Cooper, went on to cofound the American Dietetic Association (Cooper LF 1913), and Kellogg’s publication “The New Dietetics” set the standards for the association (Kellogg JH 1927). This is why cereal grains are recommended as a healthy breakfast and a toddler’s first food.
A toddler’s primary first foods, were calculated at 60g of fortified cereal, with 500ml of cow milk, as recommended by New South Wales Ministry of Health (2020).
Other foods, such as 1 large whole fish, uncooked, unscale, and not deboned, 600g, and baby spinach, raw, 60g, were added in daily portion sizes for comparison and to gain a perspective of various dietary types.
2.2.7. Beer
Beer is said to contain B vitamin’s due to the fermentation processes with yeast and grain (Koren et al 2017). 570ml of standard Australian beer; the equivalent of two standard drinks, was used to compare.
3. Analysis
3.1. Compartmentalisation
Cellular compartmentalisation is essential for maintaining the efficiency and specificity of biochemical reactions. By confining metabolic pathways to distinct organelles or cellular regions, cells can regulate processes such as deoxyribose nucleic acid (DNA) replication, energy production, and protein synthesis with precision. Liquid-liquid phase separation distinguishes the nucleoplasm from the cytoplasm (Sedley 2023). This spatial organisation prevents unwanted interactions, enhances reaction rates, and enables precise control over metabolite availability.
3.2. Uracil
Uracil is a pyrimidine nucleotide that is primarily incorporated into ribonucleic acid (RNA), where it pairs with adenine during transcription. Its demand increases during periods of high gene expression, as cells actively produce messenger RNA (mRNA), transfer RNA (tRNA), and ribosomal RNA (rRNA) to support protein synthesis. Unlike DNA, which requires thymine, RNA polymerases selectively incorporate uracil, making it a critical component of cellular function and gene regulation. However, when uracil is mistakenly incorporated into DNA, it can lead to genomic instability, faulty replication, mutation, and impaired repair mechanisms (Sedley L 2023).
3.3. Thymine
Thymine is essential for DNA replication. Unlike RNA, which uses uracil. Thymine is synthesised through the methylation of uracil. Proper nucleotide balance is essential for maintaining cellular integrity and function (Sedley L 2023).
3.4. Purine and Pyrimidine Synthesis
Purine and pyrimidine metabolism is tightly regulated, much like a scale. Accumulation of substrates or supplementation with nucleotide analogues, lead to feedback inhibition of the pathway (Baggott JE, Vaµghn WH et al 1986). B vitamins contain nucleotide moieties, or have similar binding affinities to nucleotides, and thus, can regulate nucleotide synthesis (Sedley L 2023). Therefore, nucleotides have demonstrated management of epigenetic mechanisms such as like one carbon metabolism 1-CM (Ducker GS, Rabinowitz JD 2015), and B vitamins have demonstrated modulation of nucleotide synthesis (Sedley L 2023).
3.4. Folate Analogues, Compartmentalisation and Nucleotide Synthesis for DNA Replication
Folate analogues are originally synthesised from the purine guanine, and its degradation contributes the guanine pool (cited in Sedley L 2023). Folate analogue and crucial intermediate of 1-CM, 10- formyltetrahydrofolate, is dose dependently directed into the pathway of purine synthesis (Anderson DD, Woeller CF et al 2012). Folate also plays a very important role in the regulation of pyrimidine synthesis, and therefore ,folate deficiency and excess can contribute to excessive uracil incorporation into DNA and RNA, both of which may have a variety of health implications, poor including viral immunity (Blount BC, Mack MM et al 1997) (Paone A, Marani M et al 2014) (Kisliuk RL, Gaumont Y et al 1974).
3.5. Thymine Synthesis from One Carbon Metabolism
Thymidylate Synthase (TYMS) transfers the methyl to uracil from the substrate, 5,10-methyltetrahydrofolate (5,10-MTHF) forming 5-methyltetrahydrofolate (5-MTHF) and thymine, for incorporation into DNA (MacFarlane AJ, Anderson DD et al 2011). A precise level of 5,10-MTHF is essential for thymine synthesis during replication and DNA repair.
5-MTHF, a commonly prescribed vitamin, regulates thymine synthesis via feedback inhibition of methylenetetrahydrofolate reductase (MTHFR), in a dose dependant manner, regulating the abundance of the SHMT substrate 5,10-MTHF, for uracil methylation (Sedley L 2020).
Various folate analogues inhibit TYMS through feedback mechanisms which can also lead to increased uracil incorporation into DNA or RNA (Kisliuk RL, Gaumont Y et al 1974).
3.6. Compartmentalisation of Thymine Synthesis
Thymine for DNA incorporation is compartmentally regulated (MacFarlane AJ, Anderson DD et al 2011). In the cytosol, serine is the substrate for the enzyme Serine Hydroxymethyltransferase (SHMT), which uses tetrahydrofolate (THF) as a catalyst to form glycine and 5,10 MTF. Dihydrofolate reductase (DHFR) contributes to the pool of THF for uracil methylation through the reduction of dihydrofolate (Visentin M, Zhao R et al 2012).
In a cell cycle dependant manner DHFR, TYMS and SHMT are SUMOylated and translocated to the nucleus; (MacFarlane AJ, Anderson DD et al 2011) Here, SHMT derived 5,10-MTHF is adjoined to TYMS promoting uracil methylation for replication (Anderson DD, Woeller CF et al 2012) (Figure 1).
3.7. A Metabo-Switch Between Nucleotide Synthesis, Cell Cycle and Methylation.
In the cytosol, when 5-MTHF is abundant, it is sequestered by SHMT, preventing homocysteine remethylation in 1-CM, thereby slowing down epigenetic methylation in the G2 phase of the cell cycle. This was discovered early, when 5-MTHF supplementation was shown to inhibit 1-CM (Sedley L 2020). SHMT is found to be overexpressed in tissue samples of lung cancer, and knock-down of SHMT results in cell cycle arrest, apoptosis and the accumulation of uracil in nuclear DNA (Paone A, Marani M et al 2014).
Due to a change in the cell phase, the global reduction in gene expression reduces the need for uracil, which is otherwise incorporated into RNA. This creates a competitive gateway between 1-CM and nucleotide synthesis preventing uracil incorporation into DNA (MacFarlane AJ, Anderson DD et al 2011).
3.8. Light Dependant Epigenetics and the Metabo-Switch
Quantum Biology or biophysics is an emerging field of research, which combines the study of physics with classical biology and light inducing endogenous biochemical reactions (Sedley 2025).
The circadian rhythm has been shown to play a vital role in cell division; disruption to the circadian rhythm has been implicated in several cancers with key genes responsible for cell division driven by circadian genes (Sedley L 2023).
Circadian genes also function as transcription factors for the regulation of 1-CM. Mat2A carries a binding site for transcription factor CLOCK and plays a role in the metabo-switch that separates 1-C metabolism's methylation from thymine synthesis in replication (Genome Reference Consortium 2022). It has been demonstrated that 1-CM for methylation purposes, requires light energy. The MTHFR protein requires the dietary derived riboflavin containing, light and heat sensitive cofactor, flavin adenine dionucelotide (FAD) (Rosenblatt DS, Erbe RW 1977) (Hühner J, Ingles- Prieto Á et al 2015) (Igari S, Ohtaki A et al 2011), which has shown to play an important role in circadian rhythm (Sedley 2025).
The product of MTHFR, 5-MTHF is a quencher of photosensitisers known to inhibit DNA strand breaks, and its synthetic counterpart folic acid, promotes double stranded DNA breaks in a dose dependant manner (Offer T, Ames BN et al 2007). As discussed previously, 5-MTHF is sequestered, and translocated to the nucleus by SHMT following SUMOylating. SUMOlyation is responsive to UV radiation, and SHMT is expressed primarily in light sensitive tissues. SHMT translocation and protein levels are increased 12-24 hours in response to UV treatment, however its expression remains stable. SHMT 5’ untranslated region contains an internal ribosome entry site (IRES) which is activated in response to non - lethal UV treatment stimulating protein translation and DNA repair. (Fox JT, Shin WK et al 2009).
3.9. Early Folate Quantification
3.9.1. In-Vitro
Due to low anaerobic conditions, bacterial species of the GIT are obligate anaerobes or facultative anaerobes (Laiño JE, LeBlanc JG et al 2012). Lactobacillus is an obligate anaerobe incapable of growth in folate deficient conditions (Laiño JE, LeBlanc JG et al 2012). Utilisation of folic acid by the genus, kick starts their own endogenous folate synthesis through up-regulation of gene expression, providing a source of carbon for replication and nucleotide synthesis. Essentially, their symbiotic environment initiates their cyclic survival.
The growth of Lactobacillus casei was used to determine the presence of folic acid in a substance (Shane B 2011). Researchers analysing the folate concentration in animal tissue induce folate deficiency using sulphonamide folate inhibitors to suppress the growth of folate producing bacteria in the animal’s gastrointestinal tracts. At the time, they were unaware that Lactobacillus casei and other bacteria of the microbiome are resistant to sulphonamides (Sharma C, Gulati S et al 2017) where inhibition of DHFR (Capasso C, Supuran TC 2019) only encourages diversion via an alternative pathway increasing folate analogue such as 5-MTHF, and speeding up the rate of replication, (Kanehisa M 2000), ultimately resulting in an inaccurate over-quantification of folate levels. (Fehling C, Jägerstad M et al 1976). Therefore, folate quantification data should be used with caution.
Facultative anaerobes on the other hand can grow and produce folate in folate deficient conditions. This is essential for the metabolic switch which enables their anaerobicity. Yet, growth is reduced when exogenous folate is available, this suggests that folic acid is dose dependant and an excess can modulate the metabo-switch and divert 1-CM from promoting nucleotide synthesis and replication to methylation and epigenetic regulation as discussed above (Laiño JE, LeBlanc JG et al 2012). Therefore, folic acid supplementation or fortification of foods will interfere directly with the human gastrointestinal microbiome, influencing growth, fermentation rates and species evolution.
3.9.2. In-Vivo
Rats fed a carnivorous diet of protein and fat are considered folate deficient for laboratory studies. Deficient rats have skeletal muscle containing only 0.00002µg/g of folates. A dietary increase of 26µg daily, increases muscle tissue to 0.05µg/g (Ward GJ, Nixon P F 1990) . Therefore, natural folates in animal tissues are directly proportionate to dietary intake, and range from 0.12µg/g in canned fish, to 3µg/g in liver. All other meat and fish products contain very minimal or non- detectable levels in their skeletal muscle (Kumar A 2003). Organ meats are said to be the most abundant source of folate in food products, however, like muscle tissue, animal organs can double in the folate concentration upon fortification, distributing folate to organs at a unique rate (Fehling C, Jägerstad M et al 1976), suggesting fortified animals, produce far more folate for consumption than earlier times.
Interestingly, mice fed butter are significantly more resistant to infection (Webster LT, Pritchett IW 1924), this may be due to proinflammatory constitutes. Moreover, rodent models depleted of folate are protected against viruses. The folate deficient rats in the study ate primarily protein and fat, whereas the folate sufficient animals consumed only milk and bread (Haas VH, Stewart SE et al 1957). This is likely due to a dysregulated metabo-switch, ensuring uracil availability for viruses.
4. Results
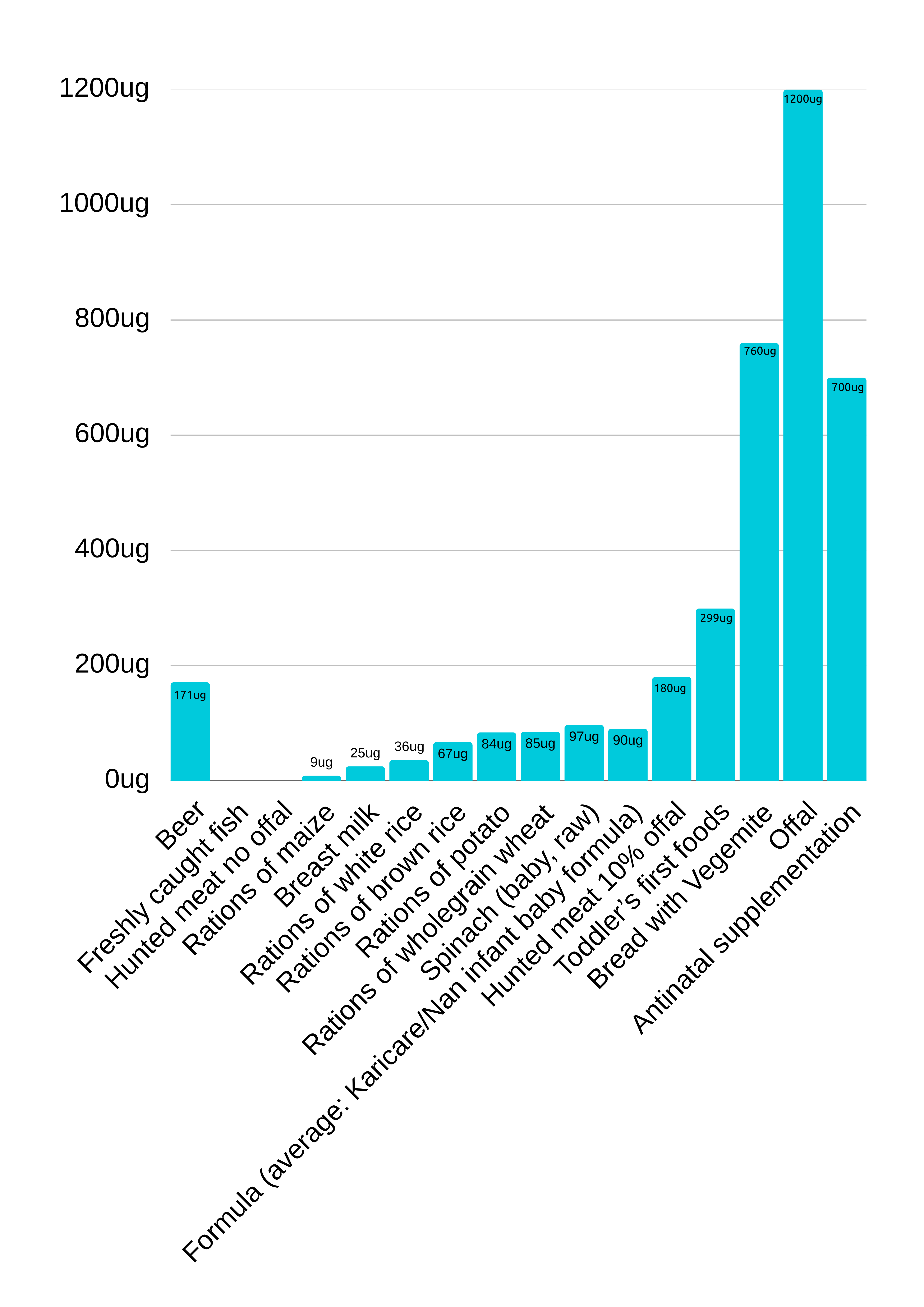
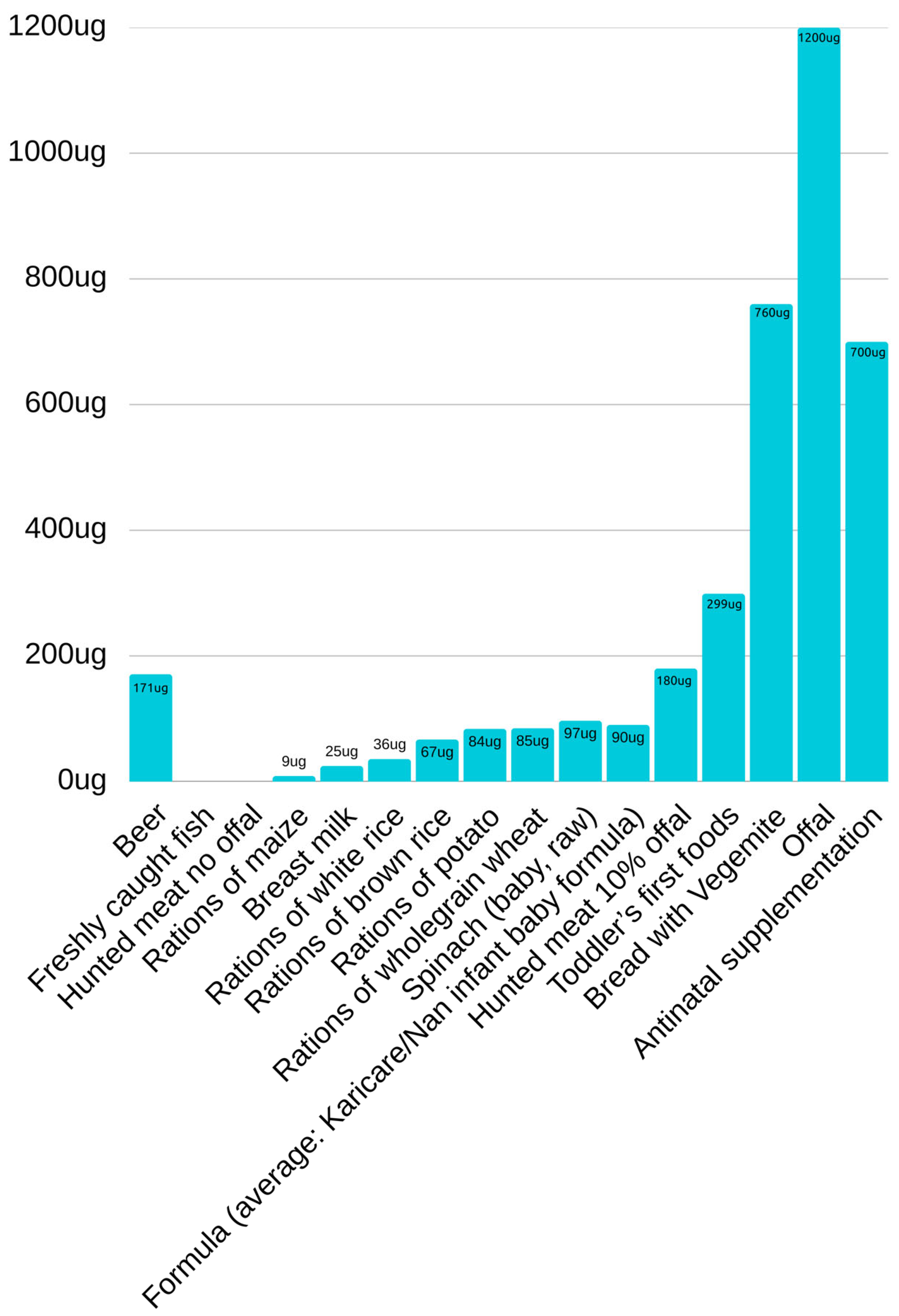
For animal meat and fish, folate levels were not input by FSANZ (2021), this may suggest levels were under the detection limitation of 3µg per 100mg. Non-detectable levels are consistent with data produced by Kumar A (2003) and therefore a value of zero was documented (figure 2).
FSANZ (2021), state that folate in beer was imputed as zero, suggesting the quantification has not been performed, yet they did have detectable levels of riboflavin, again suggesting levels may have been below detectable limits. Given the growing incidence of boutique beers, alternative data was used, for comparison. Hungarian Beer reports to contain up to 400µg per litre of folate, therefore a value of 171µg for two standard drinks per day was documented.
A 1-month-old baby fed 624ml of Breast milk receives approximately 25µg of folic acid per day, and a formula fed baby of the same age, receives 90µg. A toddler who is commencing first foods, fed a fortified cereal grain would consume approximately 300µg of folic acid.
A daily dose of folic acid from a diet of fortified bread and Vegemite could have up to 760µg of folic acid.
Offal contains the most folic acid, with a portion size containing up to 1200µg of folic acid. A daily diet of 10% off and 90% animal skeletal muscle meat would contain up to 180µg of folate.
Of the grains, maize has only 9µg of folic acid, white rice 36µg, and brown rice contains 67µg.
Potato contains 84µg and spinach contains the most folate of any vegetable, 1.6µg per gram, a dose 97µg per day 60g.
5. Discussion
Alongside current knowledge of potential folate toxicities, this research has made it clear that 5-MTHF, is a regulator of cell cycle initiation of thymine synthesis for DNA replication, and the consequence of over supplementation of this substance and other folic acid analogues could result in excess uracil accumulation in the DNA or RNA. An excess substrate may result in an epigenetic lock, preventing the natural fluctuation of the metabo-switch, that regulates 1-cm and methyl group synthesis for active gene expression and requirement for uracil, as opposed to the flexible cell cycle transition to DNA replication, when necessary.
5.1. Public Health Implications and Future Directions
This research suggests that current folate recommendations are overestimated, therefore what may be considered a deficiency, is more likely to be levels closer to normality, prior to the fortification and supplementation of animals and people.
The implications pertaining to folate deficiency and viral immunity are plausible if we consider an excess of folates in the cytoplasm, compared to natural endogenous protein/transporter levels. This may be due circadian dysfunction, insufficient light exposure or a lack of active light regulated cofactors, like FAD. Excess folate may be shunted towards cytoplasmic 1-CM, methyl synthesis for maintenance of the G2 cell phase, where viral replication can utilise excess uracil.
The contrast is also possible. If there is excessive light exposure and an excess of folates, light activated proteins or cofactors in the cytoplasm, 5-MTHF is transported to the nucleus stimulating unnecessary replication, which is an essential feature of cancer.
The effects of light exposure on the development of skin and other cancers is gaining traction, (Sedley L 2025), and more research is needed to determine the combined effects of light exposure, and folates in viral immunity, and the initiation of cell cycle phase changes.
Today, based on the information within this report, and since most processed foods are fortified, more research is needed to determine true folate intakes. Moreover, most consumable feeds for animals are fortified, therefore more research is needed to determine the accuracy of folate recommendations for domesticated animals.
In contrast to organic beef, despite mass production of folates during fermentation in the gastrointestinal tract of ruminants, folic acid supplementation is used to increase body mass, and modulation of other metabolic pathways (Do, H et al 2024). Earlier offal estimations like those described by Kumar (2003), are over 50% lower than estimations provided by Food Standards Australia and New Zealand (2020). True levels of skeletal and organ folate in consumable animals require re-assessment and wild animals could be used for comparison.
Similarly, plant estimates such as wholegrain/wholemeal wheat, had levels ranging between 1-3µg/g, much higher than estimates today, again highlighting inaccuracy (Kumar A 2003).
Long term evolutionary adaption has been largely explored, showing the requirements for folate vary between populations, as evidenced by different DNA methylation patterns and positive selection of key genes with one carbon metabolism. Higher methylation levels are observed in African American and Han-Chinese Americans compared to Caucasian-Americans, which may be a result of dietary selection and preference for unfortified foods A genetic drift model identified the MTHFR variant rs1801133 as positively selected, in high-altitude Tibet, likely due to increased UV exposure (Sedley L 2020).. Short term epigenetic adaption to dietary changes and the presence of a cellular metabolic lock at the metabo-switch, in different population groups require further investigation.
Due to limited environmental pressures, populations who maintain traditional practices tend to be the healthiest. Residents of isolated blue zone regions who maintain traditional agricultural practices, which are embedded in their history, culture, and values have exceptional longevity. (Engelbrecht HR, Merrill SM et al 2022). Indigenous Australian’s were blue zoned for over 75,000 years, but have suffered rapid health deterioration since British colonisation, due to environmental and dietary pressures (Valeggia CR, Snodgrass JJ 2015). Given the diet of indigenous Australians, only 250 years ago was documented to be primarily animal protein, therefore the population was used to demonstrate the importance of bio-individuality. The data pertaining to hunted animal meat and 10% offal was used to demonstrate the potential for folate toxicity. A transition from this diet to a diet of fortified bread and Vegemite would result in a potential daily increase of over 4.4 times more folic acid, thereby treating a deficiency that never existed, with potential toxic effects.
Expat Australians who may have evolved to consume more organ meats, may be less likely to be negatively affected by a highly fortified diet. However, due to greater wealth, the fortified rationed diet, is less likely to be consumed by this population, and future technologies will help to determine, how evolution has influenced the folate pathways in this population and if current folate intake is adequate.
This research suggests that other than a high organ meat diet, all other staple diets or rations throughout history have contained much less daily folates than the current daily recommendation of 400µg.
To gain greater perspective, a person on a plant-based would need to consume 720g of spinach to reach folate levels consumed by those consuming early rations of offal. And a pregnant woman would be required to consume 370g of spinach a day to reach current pregnancy dietary recommendations. Although it is said that spinach was domesticated over 2000 years ago (Ribera, A et al 2021), a diet this high in leafy greens, is unlikely due to the toxic effects of oxalates (Noonan and Savage 1999). Similarly, a pregnant woman, would have to consume 1200g per day of wheat grain to reach current dietary recommendations. Together, current dietary folate recommendations are not replenishing a deficiency but are rather a standardised medicine to treat a rare congenital disease, which may have negative effects in people who aren’t susceptible to this disease. A future that sees advanced targeted folate therapies are likely to reduce the incidence of NTD, whilst reducing the prevalence of alternative disease; increasing health and cognitive metrics.
Formula fed infants are receiving 3.6 times more folic acid than a breast-fed baby, however, this doesn’t take into consideration the breastfeeding mother is also consuming fortified food and is likely to be taking antenatal supplements, which suggests that breast milk was traditionally much lower in folates. Without adequate evolution, this level of folic acid could be toxic to infants and requires further investigation.
It must also be considered that the folic acid concentrations of food are directly influenced by light exposure. Fortified and non-fortified food products such as bread and milk that have been exposed to light, produce toxic metabolites that may contribute to neurological disease and dietary intolerances (Sedley 2025 and Sedley 2025). This requires further investigation as to advance personalised medicine.
Due to knowledge of unique epigenetic mechanisms between populations, it is clear, mandatory fortification, and dietary folate recommendations puts most population groups at risk of toxic levels of folic acid. The evolution a population group has undergone to determine appropriate folic acid levels in the diet is difficult due to rapid amalgamation and migration of populations, however technological advances will help us overcome this obstacle. The future of nutritional medicine will see computer models and deep learning of unique biochemistry, gene expression, rate of evolution, epigenetic patterns, compared to dietary consumption and the emergence of cellular targeted folate therapies, to enable precision healthcare.
6. Conclusion
Due to early discrepancies in folate quantification, many studies that report on a requirement for folates should be re-evaluated. Historical dietary patterns suggest that current folate recommendations may not align with natural intake levels, with plant-based sources requiring impractically high consumption to meet modern dietary guidelines, making the potential for toxicity high. For the sake of public safety, I call for a re-evaluation folate requirement, in humans, animals and pregnancy.
Variations in folate metabolism across populations, influenced by evolution and epigenetics, also highlight the need for reassessment of mandatory fortification. Future advancements in biophysics, deep learning, pre-screening and personalised medicine will enable a more precise understanding of folate requirements, mitigating potential health risks while optimising nutritional strategies.
References
- Albala, K. Eating right in the renaissance; University of California Press; Los Angeles, 2002. [Google Scholar]
- Anderson, D. D.; Woeller, C. F.; et al. Serine hydroxymethyltransferase anchors the de novo thymidylate synthesis pathway to the nuclear lamina for DNA synthesis. Journal of Biological Chemistry 2012, 287(10), 7051–7062. [Google Scholar] [CrossRef] [PubMed]
- Armelagos, G. J. Brain evolution, the determinants of food choice, and the omnivore’s dilemma. Critical Reviews in Food Science and Nutrition 2014, 54(10), 1330–1341. [Google Scholar] [CrossRef]
- Carnivores, Australian. Carnivore diet: Meals and portion sizes. n.d. Available online: https://www.australiancarnivores.com.au/meals-portion-sizes.
- Australian Institute of Health and Welfare. Mandatory folic acid and iodine fortification in Australia and New Zealand: Baseline report for monitoring. AIHW. 2011. Available online: https://www.aihw.gov.au/getmedia/1a4bb10d-dba2-479e-b99f-abefcfaf62d1/10787.pdf.
- Avagliano, L.; Massa, V.; George, T. M.; Qureshy, S.; Bulfamante, G. P.; Finnell, R. H. Overview on neural tube defects: From development to physical characteristics. Birth Defects Research 2019, 111(19), 1455–1467. [Google Scholar] [CrossRef] [PubMed]
- Baggott, JE; Vaughn, WH. Inhibition of 5-aminoimidazole-4-carboxamide ribotide transformylase adenosine deaminase and 5’-adenylate deaminase by polyglutamates of methotrexate and oxidized folates and by 5-aminoimidazole-4-carboxamide riboside and ribotide. Biochem J 1986, 236. [Google Scholar]
- Blount, B. C.; Mack, M. M.; et al. Folate deficiency causes uracil misincorporation into human DNA and chromosome breakage: Implications for cancer and neuronal damage. Medical Sciences 1997, 94. [Google Scholar]
- Capasso, C; Supuran, TC. Bonev, BB, Brownnm, Eds.; Bacterial Resistance to Antibiotics–From Molecules to Man, Dihydropteroate synthase (sulfonamides) and dihydrofolate reductase inhibitors; Wiley Online Library, 2019. [Google Scholar]
- Clapham, N. Seventh-day Adventists in the South Pacific 1885-1985: Australia New Zealand South Sea Islands, SignsdoPub C, 1985.
- Cooper, LF. The new cookery: A book of recipes most of which are in use at the Battle Creek Sanitarium; The Good Health Pub Co, 1913. [Google Scholar]
- Do, H.; Takemoto, S.; Tomonaga, S. Folic acid supplementation during the fattening period affects growth and nutritional metabolism in Japanese Black beef cattle. Scientific Reports 2024, 14, 24653. [Google Scholar] [CrossRef]
- Ducker, G. S.; Rabinowitz, J. D. ZMP: A master regulator of 1-CM. Molecular Cell 2015, 57(2), 203–204. [Google Scholar] [CrossRef]
- Elevit. Elevit products. Elevit. 2025. Available online: https://www.elevit.com.au/products/elevit.
- Engelbrecht, HR; Merrill, SM. (2022) Sex differences in epigenetic age in Mediterranean high longevity regions. Front Aging 3. [CrossRef]
- Editors of Council on Foreign Relations Abortion law: Global comparisons;Council on Foreign Relations, 2023.
- Australian Dietary Guidelines. In Editors at Nutrition Australia; Nutrition Australia, 2013.
- Editors of Food Standards Australia and New Zealand Australian Food Composition Database–Release, 2020.
- FSANZ. Available online: https://www.foodstandards.gov.au/science/monitoringnutrients/afcd.
- Fehling, C.; Jägerstad, M.; et al. Reduction of folate levels in the rat: Difference in depletion between the central and peripheral nervous systems. Zeitschrift für Ernährungswissenschaft 1976, 15(1), 1–8. [Google Scholar] [CrossRef]
- Fox, J. T.; Shin, W. K.; et al. A UV-responsive internal ribosome entry site enhances serine hydroxymethyltransferase 1 expression for DNA damage repair. Journal of Biological Chemistry 2009, 284(45), 31097–31108. [Google Scholar] [CrossRef]
- George, L. Plasma folate levels and the risk of spontaneous abortion. JAMA Network Open 2002, 288(15), 1867. [Google Scholar] [CrossRef]
- Genome Reference Consortium. Human Build 38 patch release 14 (GRCh38.p14): Mat2A, 2022.
- Graham, S. Lectures on the science of human life. In The Office of the Health reformer. Battle Creek SR Wells and Company Publishers; New Yor.
- Haas, VH; Stewart, SE. ) Folic acid deficiency and the sparing of mice infected with the virus of lymphocytic choriomeningitis. Virology 1957, 3. [Google Scholar]
- Haas, M.; Schreiber, M.; Mascher, M. Domestication and crop evolution of wheat and barley: Genes, genomics, and future directions. Journal of Integrative Plant Biology 2018, 60(12), 1067–1080. [Google Scholar] [CrossRef]
- Hancock, A. M.; Witonsky, D. B.; et al. Human adaptations to diet, subsistence, and ecoregion are due to subtle shifts in allele frequency. Proceedings of the National Academy of Sciences 2010, 107(2), 8924–8930. [Google Scholar] [CrossRef]
- Hecht, S.; Shlaer, S.; et al. Energy quanta and vision. Journal of General Physiology 1942, 25(6), 819–840. [Google Scholar]
- Hedrick, P. W. Population genetics of malaria resistance in humans. Heredity 2011, 107(4), 283–304. [Google Scholar] [CrossRef]
- Hooton, E. A. The evolution and devolution of the human face. American Journal of Orthodontics and Oral Surgery 1946, 32(41). [Google Scholar]
- Hühner, J; Ingles-Prieto, Á. Quantification of riboflavin flavin mononucleotide and flavin adenine dinucleotide in mammalian model cells by CE with LED-induced fluorescence detection: CE and CEC. Electrophoresis 2015, 36(4), 518–525. [Google Scholar] [CrossRef]
- Igari, S; Ohtaki, A. Properties and crystal structure of methylenetetrahydrofolate reductase from thermus thermophilus HB8. PLos One 2011, 6(8), e23716. [Google Scholar] [CrossRef]
- Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Research 2000, 2(1), 27–30. [Google Scholar] [CrossRef]
- Kellogg, JH. Are we to be a toothless race? Dent Regist 53(3), 135–144. [PubMed]
- Kellogg, JH. The New Dietetics, A guide to scientific feeding in Health and Disease; The Modern Medical Publishing Company; Revised Edition, 1927. [Google Scholar]
- Kiefer, CM; Hou, C. Epigenetics of β-globin gene regulation. Mutat Res 2008, 647(1-2), 68–76. [Google Scholar] [CrossRef]
- Kim, Y.; Bertagna, F.; et al. Quantum biology: An update and perspective. Quantum Reports 2021, 3(1), 80–126. [Google Scholar]
- Kisliuk, RL; Gaumont, Y. ) Polyglutamyl derivatives of folate as substrates and inhibitors of thymidylate synthetase. J Biol Chem 1974, 249(13), 4100–4103. [Google Scholar] [CrossRef]
- Kommissionen for Ledelsen af de Geologiske og Geografiske Undersøgelser i Grønland, Meddelelser om Grønland (1915) København C, A, Reitzels Forlag 49.
- Koren, D; Orbán, C; Galló, N; Kun, S; Vecseri-Hegyes, B; Kun-Farkas, G. Folic acid content and antioxidant activity of different types of beers available in Hungarian retail. J Food Sci Technol 2017, 54(5), 1158–1167. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kumar, A. Extraction and determination of folates in foods and the fortification of rice with folic acid; University of New South Wales, 2003. [Google Scholar] [CrossRef]
- Laiño, JE; LeBlanc, JG. Production of natural folates by lactic acid bacteria starter cultures isolated from artisanal Argentinean yogurts. Can J Microbiol 2012, 5(5), 581–588. [Google Scholar] [CrossRef]
- Li, J.; Fan, W.; Wang, X.; Hou, X.; Chen, Z.; Lv, M. Mental health in early pregnancy and spontaneous abortion risk: A prospective cohort study. Alpha Psychiatry 2024, 25(5), 648–655. [Google Scholar] [CrossRef]
- Lynn, R.; Harvey, J. The decline of the world’s IQ. Intelligence 2008, 36(2), 112–120. [Google Scholar] [CrossRef]
- MacFarlane, A. J.; Anderson, D. D.; et al. Nuclear localization of de novo thymidylate biosynthesis pathway is required to prevent uracil accumulation in DNA. Journal of Biological Chemistry 2011, 286(51), 44015–44022. [Google Scholar]
- Noonan, S. C.; Savage, G. P. Oxalate content of foods and its effect on humans. Asia Pacific Journal of Clinical Nutrition 1999, 8(1), 64–74. [Google Scholar] [PubMed]
- New South Wales Ministry of Health. Starting family foods: Guidelines for introducing solid foods to infants (Publication No. GL2020_012). NSW Health. 2020. Available online: https://www.health.nsw.gov.au/heal/Publications/starting-family-foods.pdf.
- Offer, T; Ames, BN. ) 5-Methyltetrahydrofolate inhibits photosensitization reactions and strand breaks in DNA. FASEB J 2007, 21(9), 2101–2107. [Google Scholar] [CrossRef]
- Organization of Teratology Information Specialists (OTIS)Folic acid Folate. Mother To Baby Fact Sheets. January 2024. Retrieved from. Available online: https://www.ncbi.nlm.nih.gov/books/NBK582717/.
- Paone, A; Marani, M. SHMT1 knockdown induces apoptosis in lung cancer cells by causing uracil misincorporation. Cell Death Dis 2014, 5(11). [Google Scholar]
- Pensary, LA. 1903) A retrospect of the Irish poor law dispensary lunatic asylum and workhouse systems, hospitals for paying patients. Br Med J 1(2192), 27–28.
- Ribera, A.; van Treuren, R.; Kik, C.; et al. On the origin and dispersal of cultivated spinach (Spinacia oleraceaL) Genetic Resources and Crop Evolution. 2021, 68, 1023–1032. [Google Scholar] [CrossRef]
- Rios-Leyvraz, M.; Yao, Q. The volume of breast milk intake in infants and young children: A systematic review and meta-analysis. Breastfeeding Medicine 2023, 18(3), 188–197. [Google Scholar] [CrossRef] [PubMed]
- Rosenblatt, DS; Erbe, RW. Methylenetetrahydrofolate reductase in cultured human cells, II, Genetic and biochemical studies of methylenetetrahydrofolate reductase deficiency. Pediatr Res 1977, 11(11), 1141–1143. [Google Scholar] [CrossRef]
- Shannon, C. Acculturation: Aboriginal and Torres Strait Islander nutrition. Asia Pac J Clin Nutr 2002, 11. [Google Scholar]
- Sharma, C; Gulati, S. Antibiotic sensitivity pattern of indigenous lactobacilli isolated from curd and human milk samples. 3 Biotech 2017, 7(1), 53. [Google Scholar] [CrossRef]
- Sedley, L. Advances in nutritional epigenetic a fresh perspective for an old idea; lessons learned limitations and future directions. Epigenet Insights 2020, 13. [Google Scholar] [CrossRef]
- Sedley, L. Dinan T, Ed.; Epigenetics. In Nutritional psychiatry: a primer for clinicians; Cambridge University Press; Cambridge, 2023. [Google Scholar]
- Sedley, L. Illuminating the Connection: Breastfeeding, Lactose Intolerance. and Early Life Disease Prevention 2025. [Google Scholar] [CrossRef]
- Sedley, L. The Epigenetics and Biological Clock of Skin Cancer. 2025. [Google Scholar] [CrossRef]
- Shane, B. Folate status assessment history: Implications for measurement of biomarkers in NHANES. Am J Clin Nutr 2011, 94(1), 337S–342S. [Google Scholar] [PubMed]
- Sherriff, S; Kalucy, D; et al. Murradambirra Dhangaang (make food secure) Aboriginal community and stakeholder perspectives on food insecurity in urban and regional Australia. BMC Public Health 2022, 22(1). [Google Scholar]
- Shurtleff, W.; Aoyagi, A. History of Seventh-day Adventist Work with Soyfoods Vegetarianism Meat (1863-2013); Soyinfo Centre; Lafayette, 2014. [Google Scholar]
- Editors of the Online Etymology Dictionary (2023) Online Etymology Dictionary, Humble. Available online: https://www.etymonline.com/word/humble.
- Valeggia, CR; Snodgrass, JJ. Health of indigenous peoples. Annu Rev Anthropol 2015, 44(1), 117–135. [Google Scholar] [CrossRef]
- Visentin, M; Zhao, R. The Antifolates. Hematol Oncol Clin North Am 2012, 26(3), 629–648. [Google Scholar] [CrossRef]
- Ward, G. J.; Nixon, P. F. Modulation of pteroylpolyglutamate concentration and length in response to altered folate nutrition in a comprehensive range of rat tissues. J Nutr 1990, 120(5), 476–484. [Google Scholar]
- Webster, L. T.; Pritchett, I. W. Microbic virulence and host susceptibility in paratyphoid-enteritidis infection of white mice. J Exp Med 1924, 40(3), 397–404. [Google Scholar]
- World Health Organization. Autism. 2023. Available online: https://www.who.int/news-room/fact-sheets/detail/autism.
- Williams, J.; Mai, C. T.; et al. Updated estimates of neural tube defects prevented by mandatory folic acid fortification—United States, 1995–2011. MMWR Morbidity and Mortality Weekly Report 2015, 64(1), 1–5. [Google Scholar] [CrossRef]
- Wilson, B. C. Dr, John Harvey Kellogg and the Religion of Biologic living; Indiana University Press, Bloomington, 2014. [Google Scholar]
- Wyndham, C. H.; McPherson, R. K.; Munro, A. Reactions to heat of Aborigines and Caucasians. J Appl Physiol 1964, 19(6), 1055–1058. [Google Scholar] [CrossRef]
Figure 1.
Schematic Representation of the Folate Metabo-Switch.
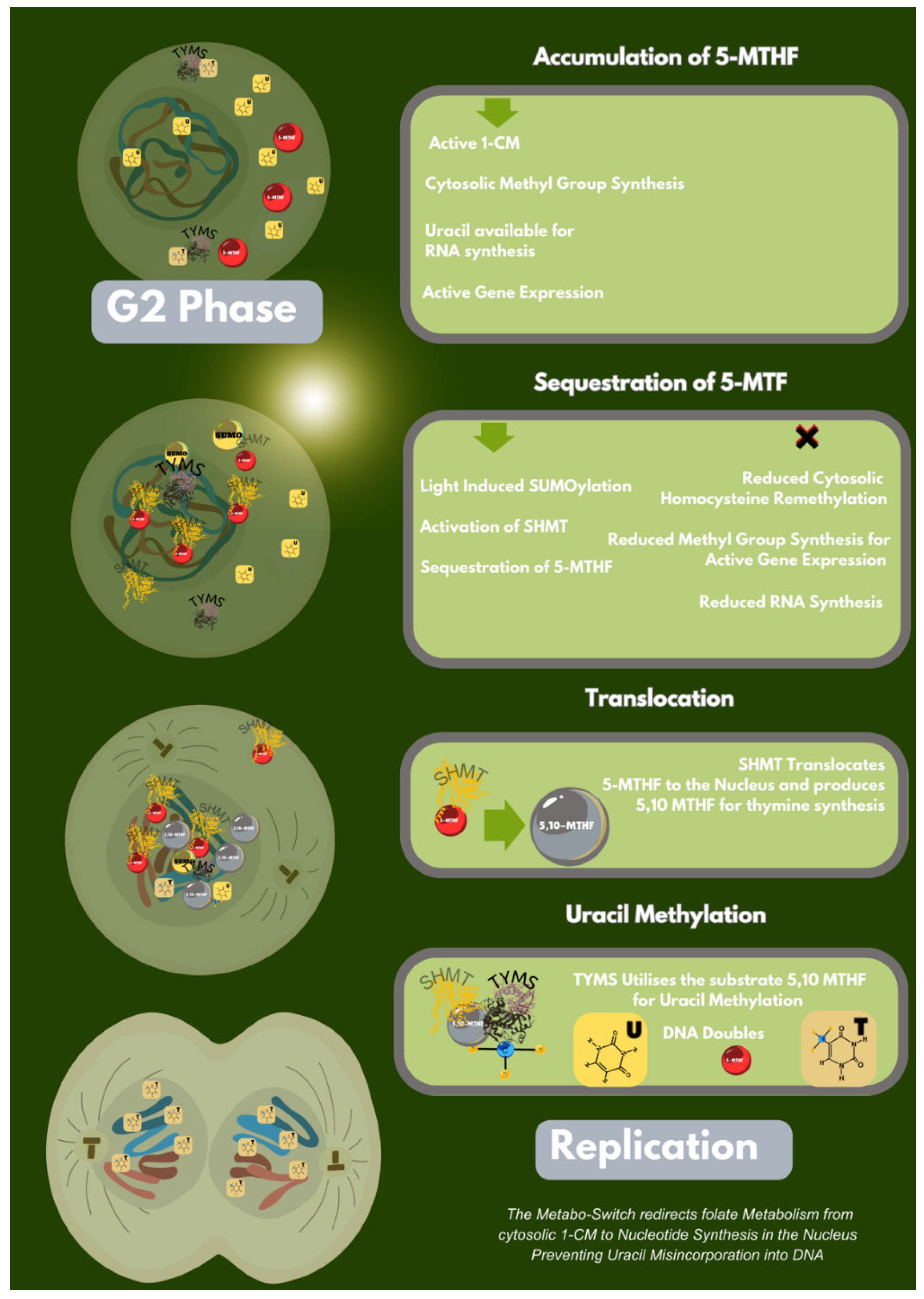
Figure 2.
Folate Composition of Various Food Staples and Rations.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



