
Submitted:
11 March 2025
Posted:
12 March 2025
You are already at the latest version
Abstract
The growing demand for sustainable production of high-value compounds, such as biofuels, lipids, and pigments like carotenoids and phycobilin, has become the subject of numerous investigations. Furthermore, this fact has led to the exploration of re-newable methods utilizing microalgae as feedstock to mitigate the challenges associ-ated with producing these valuable compounds. Nevertheless, despite the numerous advantages of microalgae, the development of a microalgal biorefinery that employs sustainable, environmentally friendly, and economically efficient technologies remains a necessity. To address this challenge, the bio-flocculation process and more specially the self-flocculation ability, is presented as a cost-effective and energy-efficient solu-tion. This method is as easy and effective as chemical flocculation, which is applied at industrial scale, however in contrast, it is sustainable and cost-effective as no costs are involved for pre-treatment of the biomass for oil extraction and for pre-treatment of the medium before it can be re-used. In addition, microalgae possess molecular tools that would allow increasing the efficiency of these processes. In the present review, we summarize microalgal harvesting technologies, with a particular focus on bio- and self-flocculation processes and identify the improvement technologies that could en-hance the production of high added-value compounds while simultaneously reducing costs in microalgae biorefinery.
Keywords:
microalgae
; biorefinery
; bio-flocculation
; self-flocculation
; high added-value compounds
; lipids
; carotenoids
; recombinant proteins
1. Introduction
The combustion of fossil fuels releases considerable quantities of CO₂ into the atmosphere, a major greenhouse gas, with only 40% of this naturally absorbed [1]. Researchers indicate that it is essential to maintain CO₂ levels below 450 ppm to prevent severe environmental disruption. This requires a reduction in emissions by 44.2 million metric tons on an annual basis. Despite the discovery of limited reserves, increasing demand may exhaust fossil fuel resources by 2066 [2,3,4]. In order to address the energy crisis and climate change, renewable energy sources, particularly biofuels, are gaining global support. Many countries are increasing biofuel production and integrating them into transportation for reducing fossil fuel dependency [5]. In this context, biorefineries represent a sustainable alternative to fossil fuel-based industries by utilizing biological resources to produce fuels, chemicals, and other high added-value compounds, thereby reducing greenhouse gas emissions [6]
Among the biorefinery types, microalgae-based biorefineries are particularly promising due to the rapid growth rate of microalgae, their high photosynthetic efficiency, and their ability to capture CO₂, making them highly efficient for carbon mitigation. Unlike terrestrial crops, microalgae do not compete with food production and can be cultivated on non-arable land with minimal water and nutrient input, which greatly enhances their environmental sustainability [7,8]. Furthermore, microalgae contain valuable lipids, carbohydrates, and proteins, which can be converted into biofuels, animal feed, and high value biochemicals, adding economic viability to their use in biorefineries [9]. Thus, microalgae biofuel productions are recognized as third-generation biofuels, positioning these organisms as one of the most promising feedstocks [10,11]. Many sustainable methods for utilizing microalgae in bioenergy production have been explored [12]. There are several benefits associated with using microalgae for biodiesel generation: i) microalgae are autotrophic organisms that harness solar energy to produce triglycerides, which can be transesterified into biodiesel [13]; ii) cultivated microalgae serve as valuable feed in aquaculture due to their high levels of essential polyunsaturated fatty acids, alongside a variety of lipids, hydrocarbons, and other complex oils [14]; iii) the use of microalgae for biodiesel production does not compromise food security and generates minimal greenhouse gas emissions, making it an environmentally friendly and sustainable energy source [7,15] and finally, iv) microalgae can yield up to 136,900 liters of oil per hectare, which is three times more than other oil crops. Given these advantages, microalgae appear to offer a promising solution to the ongoing energy demand and the development of cost-effective processes in microalgae biorefineries could provide a cleaner, renewable energy source and a range of bioproducts while significantly reducing the ecological footprint of fuel production [16]. However, the recovery of algal cells from base water typically necessitates external energy and chemicals due to their low concentration in culture, small size and negative surface charge [17,18,19]. In addition, various morphological and physiological traits of algal cells, including their shape, cell wall structure, and extracellular organic matter, can vary significantly based on nutritional and environmental factors, such as medium composition, light exposure, temperature, pH levels, culture duration, and the type of bioreactor used [2]. Indeed, one of the primary challenges is the efficient harvesting of microalgae from the medium during downstream processes. High biomass concentrations cause microalgae cells to clump together, leading to reduced productivity. To maintain a high concentration of biomass, it is essential to harvest the microalgae [20,21]. Consequently, substantial volumes of water must be extracted from production systems during the harvesting process, which necessitates energy consumption and financial investment. It is estimated that harvesting costs represent over 30% of the total production costs [22]. This high value is largely due to the small size of microalgae cells, their colloidal stability in suspension, and their growth in dilute conditions [23,24].
To date, a diversity of technologies has been developed for harvesting microalgae which includes physical, chemical, biological, and electrical techniques. In some instances, combinations of two or more methods have been employed to achieve optimal biomass recovery [9,25,26]. Centrifugation is one such method used for microalgae harvesting. Based on a mechanical gravitational force that allows for efficient harvesting of suspended cells in a short time; it is rapid and highly efficient in recovery. However, it is also significant energy-intensive, and the generated shear force can damage the cells [25]. On the other hand, gravity sedimentation and filtration, while cost-effective and easy to operate, demonstrate insufficient productivity for effectively separation biomass from the bulk culture [26,27]. In contrast to the aforementioned methods, air flotation is based on the generation of up-rising gas bubbles that bind the algal cells, offering several advantages, including a smaller footprint, flexibility, and potential for industrial scalability. Nonetheless, it requires a higher initial investment and is also energy-intensive [28,29].
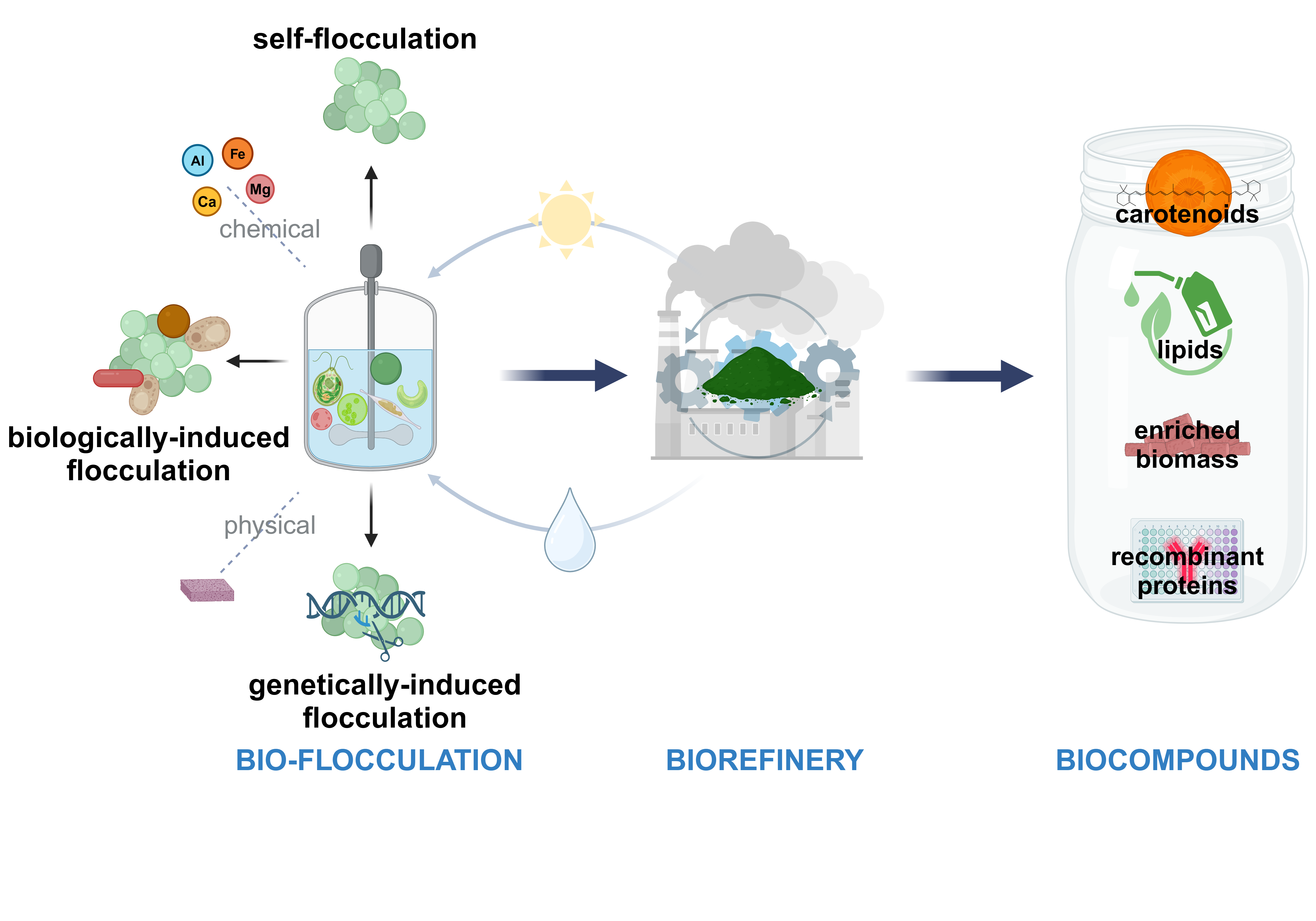
Flocculation is a commonly used technique for separating coarse dispersions and colloidal substances in water, and it has been utilized for the separation and collection of microalgae since the 1980s [30]. Flocculation involves the aggregation of small and unstable particles, facilitated by the neutralization of surface charges, electrostatic patching, and bridging after the introduction of flocculants (Figure 1). The formation of flocs allows for separation or recovery through methods like gravity settling or other conventional separation techniques [31]. This process has been studied as an effective method for harvesting various algal species [18,26].
Considering the prevailing advancements and the rapid technological development currently underway, it is essential to undertake continuous bibliographic reviews of this research topic to facilitate the acceleration and enhancement of ongoing research efforts. The objective of this review is to provide a comprehensive overview of recent flocculation technologies, emphasizing the intricacies of microalgal bio-flocculation and its potential for developing a sustainable, feasible, and environmentally friendly biorefinery. The work also examines promising research pathways for utilizing microalgae as bio-factories and details how bio-flocculation techniques have been applied to enhance the industrial production of high-value compounds such as carotenoids, lipids, and enriched biomass. The review concludes with an outline of future directions and key insights for further investigation.
2. Flocculation Methods
2.1. Physical Flocculation
Utilizing physical flocculation for extraction of microalgae is an effective method to minimize contamination. This process can be accomplished through various techniques, including ultrasound, electro-flocculation, and magnetic separation. For example, ultrasonic irradiation-coagulation demonstrated effective alga recovery under optimal conditions. On the other hand, electro-flocculation presents a more cost-efficient and scalable approach for harvesting microalgae. In this method, negatively charged microalgal cells migrate toward the anode and lose their charge, leading to the formation of aggregates or flocs. The bubbles generated at the anode rise to the surface, capturing microalgal aggregates or flocs, which can be easily removed. There are many studies demonstrating that different metal ions treatment enhances electro-coagulation process in various microalgae [32,33,34]. Another treatment is magnetic nanoparticles that have been investigated as a potential alternative process in microalgae harvesting, where magnetic nanoparticles adhere directly to the surfaces of microalgal cells, promoting flocculation in the presence of a magnetic field. Different researchers have proved the marine microalga harvesting using nanoparticles and they have shown a rapid flocculation rate and high efficiency [35]. Moreover, the effectiveness of nanoparticles binding on different microalgal species appears to be species-specific, with improved adsorption along Chlorella vulgaris, Chlamydomonas reinhardtii and Phaeodactylum tricornutum observed when the nanoparticles were coated with canonic polymers or silica [36,37,38,39]. A notable advantage of utilizing magnetic nanoparticles for microalgae harvesting is their ability to be efficiently reused without the need for pH adjustments [40]. However, it is important to highlight that effective elution strategies are critical for nanoparticle regeneration, and obtaining biomass free of nanoparticles is generally preferred [41]. A significant limitation of this technology is the high cost associated with magnetic nanoparticles and the specialized equipment required for their recycling.
2.2. Chemical Flocculation
Chemical flocculation in microalgae is typically facilitated by three primary categories of flocculants: inorganic flocculants (including metal salts and ammonia), inorganic polymers, and organic polymers. Several studies have demonstrated the effective use of chemical flocculation for harvesting various microalgae species [42,43,44,45,46,47,48,49,50]. The high flocculation efficiency and convenience it offers for microalgae harvesting has positioned chemical flocculants as a viable option for large-scale microalgae biomass production. However, the introduction of inorganic or organic pollution from these flocculants can lead to secondary pollution, and residual chemicals (such as aluminum) in the microalgal biomass, may hinder its application in food and animal feed [51]. The presence of chemical residues, including aluminum, in microalgae can affect the composition of fatty acid methyl esters. This is due to the fact that these residues are also present in lipids extracted from harvested microalgae by chemical flocculation processes [46,52]. The accumulation of these chemical residues can cause cellular damage. Additionally, the quality of pigments in microalgae, particularly chlorophyll, may be affected by the presence of ferric salts [51,53]. In contrast, using aqueous ammonia for harvesting microalgae typically has minimal impact on the distribution of metabolites such as chlorophyll, protein, and lipid [54]. Conversely, chitosan has the potential to facilitate downstream dewatering processes, such as centrifugation and filtration, by reducing time and costs [53,55,56]. Furthermore, although organic flocculants are typically regarded as safe and biodegradable, their utilization is frequently constrained by pH sensitivity [57,58]. It is also important to note that the cost of organic polymers is typically higher than that of inorganic flocculants. Consequently, the utilization of pH-based flocculation techniques, employing either acids or alkalis, has considerably expanded the range of methodologies available for microalgae harvesting. The ability of microalgal cells to maintain a stable suspension due to their negatively charged surfaces has made flocculation via pH adjustment an effective method [59]. The alteration of the proton and hydroxyl ratio and the presence of magnesium in the medium can disrupt the electrostatic interactions between anionic algae. Moreover, the utilization of chemical flocculation could be entirely avoided throughout the aforementioned process, thereby markedly reducing the risk of chemical contamination and secondary pollution. Recently, several microalgae species have been successfully harvested through pH-induced flocculation without the addition of any chemical agents. For instance, a flocculation of 95% was achieved for Nannochloropsis oculata during the late exponential phase when the culture medium was adjusted to pH 10 with Ca(OH)2 [60,61]. Similar results were noted when the pH of culture media for Scenedesmus quadricauda, Dunaliella viridis and Phaeodactylum tricornutum were adjusted to 11.5, 10 and 10.5, respectively, using NaOH or through CO2 regulation [62,63]. Moreover, the addition of a relatively small quantity of magnesium ions to a medium with a high pH value has been demonstrated to facilitate the flocculation of Chlorella vulgaris and Chlorococcum infusionum, as well as some cyanobacteria strains [64,65,66]. It is notable that the flocculation of microalgae at high biomass concentrations by decreased pH demonstrated performance that was comparable to that observed with chemical flocculants. In this instance, the process was driven by proton ions rather than magnesium, and the estimated harvesting cost was less than one dollar per kilogram of microalgal biomass, despite the limited acidic pH range [55,67].
2.3. Bio-Flocculation
Bio-flocculation is a biological process whereby microorganisms, including bacteria, yeast and microalgae, aggregate to form structures known as flocs. This process is most prevalent in aquatic environments, such as wastewater treatment systems and aquaculture. During bio-flocculation, suspended particles in the water cluster together form larger flocs, which are then more readily separable from the water. This method is fundamental to the improvement of water quality, as it reduces the concentration of suspended particles and contaminants and, it is recognized as a sustainable flocculation method for microalgae [68,69]. Furthermore, it facilitates the recycling of essential nutrients within cultivation systems, such as those employed in aquaculture. The role of microalgae bio-flocculation in sustainable water treatment and bio-compounds recovery is an increasingly studied area, as it has the potential to minimize the use of chemical additives and support the principles of circular resource management [70,71,72].
Research efforts have primarily focused on enhancing its efficiency and applicability for microalgae biorefinery. However, most strategies have only been validated at the laboratory scale, with no successful large-scale applications for microalgal harvesting to date. The current investigations into microalgae bio-flocculation have focused on four principal approaches: flocculation by microbial bio-flocculants; fungi-mediated flocculation; bacteria-mediated flocculation; algal-mediated flocculation; and self-flocculation mechanisms.
2.3.1. Flocculation by Bio-Flocculant Molecules
Effective bio-flocculation is typically initiated by other microorganisms or the extracellular substances they produce, which act as flocculants to aggregate the target microalgae [73]. These polymers can include polysaccharides, proteins, or various other bio-flocculant agents secreted by bacteria, fungi, and algae, thereby promoting the sedimentation of microalgae. The efficiency of flocculation is influenced by the species of microalgae involved. Each species synthesizes distinct extracellular polymers, resulting in varying settling rates and efficiencies. Table 1 provides a summary of the principal species of microalgae in which bio-flocculants have been employed for their harvesting.
2.3.2. Fungi-Mediated Flocculation
Fungi have the ability to form filaments or, in some cases, undergo self-pelletization, which can be leveraged for the harvesting of microalgae cells. Certain fungi from the Basidiomycetes, Aspergillus and Phanerochaete species can facilitate coagulation by forming pellets or aggregates through the presence of spores. In contrast, non-coagulative fungi from the Mucor, Rhizopus and Penicillium species can aid in trapping algal cells by developing extensive hyphae. To promote bio-flocculation, fungi and microalgae can sometimes be co-cultivated. However, the effectiveness of this process is often uncertain, as fungi may outgrow microalgae due to their faster growth rates and the concurrent competition for available carbon sources. Fungal exopolysaccharides are common and serve various biological functions, such as acting as viscosity stabilizers in the food industry, enhancing oil recovery, and exhibiting antioxidant, antiviral, and antitumor activities [81,82]. The production of exopolysaccharides by fungi can also promote cell aggregation and is influenced by culture conditions and medium composition [83]. Typically, exopolysaccharide-producing fungal strains are either aerobic or facultative anaerobic, suggesting that low oxygen levels are not conductive to exopolysaccharide synthesis. Certain fungal strains have demonstrated effective flocculation in the microalga Chlorella vulgaris, resulting in minimal settling times. Fungal-mediated bio-flocculation of microalgae can be initiated either by adding fungal cultures or through algal-fungal co-culture. Generally, fungi can flocculate microalgae by charge neutralization and ionic bridging [84]. In several studies have identified that algal and fungal aggregation occurred through ionic bridge formation, enhancing the flocculation efficiency by metal ions [10]. Results reported that the microalga Chlorella vulgaris increased the aggregation to 100% co-cultivating together with Aspergillus niger, Aspergillus flavus, Aspergillus versicolor and Leucogyraphana arizona [48].
2.3.3. Bacteria-Mediated Flocculation
The presence of bacterial cells is unavoidable in outdoor conditions and is often considered a contaminant. However, the interaction between algae and bacteria can provide nutritional and environmental advantages for the bacteria. Conversely, this association can be detrimental to microalgal growth, as bacteria typically present fast growth rate compared to microalgae and thus proliferate quickly. The interaction between bacteria and microalgae is specific to both species and strains. Bacteria can produce glutamic acid as bio-flocculant promoters in microalgae. This production is primarily essential for synthesizing amino acids and factors involved in the Krebs cycle pathway. Glutamic acid can be transformed into polyglutamic acid, which serves as an effective bio-flocculant. Some bacterial strains have been observed to induce agglomeration in microalgal cultures during their growth. The underlying mechanisms of adhesion are normally due to charge neutralization, bridging, and electrostatic patching [85]. The green algae Asterococcus limneticus WL2 and the phytohormone-producing Streptomyces rosealbus MTTC12951 have been cultivated together in a co-cultivation process to measure the production of indole-3-acetic acid and its precursor. The results showed that within 24h of resting time, early deceleration phase co-cultivated cells demonstrated significant bio-flocculation and lipid production efficiency. Thus, the production and harvesting expenses of in vitro grown microbial biomass as well as energy yield suggest co-cultivation, and a two-stage co-cultivation strategy has the potential to improve cost efficiency and energy conservation [86]. Bacterial extracellular secretions can lead to charge neutralization on microalgal cell surfaces, resulting in microalgal flocculation [87,88]. Paenibacillus sp. AM49 has been identified as an effective flocculant for Chlorella vulgaris. This efficiency significantly surpasses that of chemical flocculants, such as polyacrylamide and aluminum. Additionally, Paenibacillus sp. AM49 has been shown to be notable flocculant for other species, including Botriococcus braunii, Scenedesmus quadricauda and Selenastrum capricornutum [89]. There are studies that have utilized bacteria such as Cobetia marina L03 as bio-flocculant for Chlorella vulgaris evading the two necessary steps to successfully harvest microalgae: flocculation and filtration [90]. Another study investigated the flocculation activity of Shinella albus xn-1 with Chlorella vulgaris. This bacterium, commonly found in waste compost, sludge, or soil, participates in nitrate reduction [91,92,93] and the assimilation of methylsulfolano [94]. The flocculation efficiency increased when using the supernatant obtained after solid component separation from the bacterial culture as the flocculant. Several microalgae as Nanochloropsis oculata and three species of Microcystis aeruginosa were treated with bio-flocculant provided from extracellular products derived from the fermentation broth. The results showed that the biomass recovered was lower than that for Chlorella vulgaris, indicating species specificity of the bio-flocculant [85,90].
2.3.4. Alga-Mediated Flocculation
While some species of microalgae exhibit natural self-flocculation, others can be induced to flocculate by altering the pH or introducing cationic additives. Nonetheless, the use of cationic flocculants or acid/base treatments for self-flocculation can lead to contamination of the biomass with residual flocculants [95], complicating downstream processing and biomass utilization [9]. This section focuses on algal-algal bio-flocculation, which involves harvesting through self-aggregating algae or their extracellular polymeric substances. Self-flocculation has been documented in various strains of Chlorella vulgaris, Dunaliella salina and Scenedesmus quadrucauda. Throughout their cell cycle, microalgae secrete extracellular polymeric substances into their environment, creating a biofilm layer. The extracellular substances are primarily made up of proteins, polysaccharides, lipids, and nucleic acids. The dominant monosaccharide components in algal include hexose, uronic acid, pentose, methyl pentose, and sulfates [96]. Proteins involved in cell adhesion are typically lectins, which bind with carbohydrates, amyloids, or other cell surface proteins [97]. In green algae, glycoproteins represent a significant portion of the extracellular substances [98] and these extracellular proteins play a key role in regulating CO2 metabolism in these organisms. The conditions under which algae are cultivated can influence extracellular substances helping with the aggregation of microalgae that are typically non-aggregate and so, with the bio-flocculation process. Some studies have demonstrated improved recovery by algal-algal bio-flocculation of Picochlorum sp. QUCCM130 with low-settling capacity [99]. Results shown that self-settlement capacity increased with cell size and can be indirectly related to a decreased zeta potential of larger cells which enhances the Van der Waals attractive forces in Picochlorum sp. QUCCM130, Nannochloris sp. QUCCM31 and Tetraselmis sp. QUCCM50 [99]. In other study conducted by Salim and co-workers [100], the researchers demonstrated that cell-to-cell adhesion mechanisms vary when a consortium of flocculating algae is employed to harvest the non-flocculating Chlorella vulgaris. Specifically, in Ankistrodesmus falcatus, floc formation occurred through bridging, attributed to the positively charged extracellular polymeric substances, while in Tetraselmmis suecica and Scenedesmus obliquus, flocculation resulted from the principle of patch formation. When the self-flocculating microalgae Ettlia texensis, Scenedesmus obliquus and Ankistrodesmus falcatus were introduced to Chlorella vulgaris, the flocculation efficiencies increased. In contrast, the addition of Tetraselmis suecica as a flocculant to Neochloris oleoabundans at the same ratio resulted in an increase in biomass recovery [101]. The flocculant derived from the self-flocculating Chlorella vulgaris JSC-7 was primarily composed of carbohydrates, with glucose, mannose, and galactose identified as the sugar residues [102]. Using this alga as a flocculant for harvesting the non-flocculating Chlorella vulgaris CNW11 and Scenedesmus obliquus led to improved flocculation efficiencies. When the self-flocculation Ettlia texensis acted as a flocculant for non-flocculating Chlorella vulgaris, it was found that glycoproteins on the cell surfaces played a crucial role in flocculation [103]. Co-culturing flocculating and non-flocculating algae also proved beneficial for harvesting. For instance, when Desmodesmus sp. ZFY was co-cultured with Monoraphidium sp. QLY-1, the flocculation efficiency reached 85.33% with a settling time of 4 hours, compared to mono-culture efficiencies of 57.98% and 32.45%, respectively. The extracellular polymeric substances from this co-culture were notably rich in proteins and carbohydrates. Furthermore, co-culturing enhanced the metabolite content of the biomass [104].
2.3.5. Self-Flocculation
Cellullar self-flocculation is the process by which cells aggregate and adhere to one another in liquid culture, facilitated by specific properties of their surfaces. This phenomenon is common among microorganisms, with several self-flocculating microalgae identified, such as Chlorella vulgaris, Scenedesmus obliquus, Ankistrodesmus falcatus, Phaeodactylum tricornutum and Ettlia texensis [101,102,105,106] or flocculating related genes present in Chlamydomonas reinhardtii, Tetradesmus dimorphus and Microcystis aeruginosa [107]. The recovery of biomass through the sedimentation of flocculating yeast has been utilized in several industries, particularly in brewing and bioethanol production [108]. Flocculation presents a cost-effective and simple method for cell collection, making it an appealing area of research, especially in the context of harvesting microalgal cells. A key benefit of self-flocculation is the elimination of chemical additives, resulting in a sustainable process that avoids chemical contamination. Investigating the biochemical and genetic foundations of flocculation is essential for engineering tis trait. Although the genetic principles related to flocculation are well-established in brewing yeast, where a set of FLO genes promotes cell adhesion [108,109], the molecular mechanisms in other microbial strains, particularly microalgae, are not well understood. Research on Chlorella vulgaris and Scenedesmus obliquus has indicated that polysaccharides produced by these trains play a crucial role in self-flocculation. Additionally, recent studies suggest that glycoproteins may also contribute to the flocculation of Ettlia texensis [103]. Self-flocculation is believed to occur when flocculating agents produced by microalgae link nearby cells or form bridges through charge neutralization, thereby enhancing the flocculation process. Moreover, self-flocculation encompasses not only the spontaneous sedimentation of certain microalgal strains but also the aggregation induced by external factors, such as the addition of alkalis or the consumption of CO2, which raises the medium’s pH [25,110]. This process leads to the precipitation of carbonates and the co-precipitation of magnesium and Calcium ions as pH increases [111]. Another species such as Scenedesmus quadrucaula can produce extracellular polymeric substances that facilitate effective self-flocculation [112,113]. The potential applications of self-flocculating microalgae extend to wastewater treatment [114], where certain strains can remove nutrients while producing lipids [115]. For instance, specific microalgae isolated from undiluted piggery wastewater demonstrates impressive nutrient removal efficiencies. Additionally, microalgae like Tribonema viride and Synechocystis sp. PCC6803 have proven effective in eliminating ammonia nitrogen, total phosphorus, and chemical oxygen demand, underscoring the potential for microalgae [116]. For further advance the capabilities of microbial strains, recent developments in genetic manipulation systems have opened new lines for exploring flocculation agent biosynthesis. Concerns regarding the stability of transgenic microalgae can be alleviated through the integration of target genes into the nuclear genome [105,117,118]. Furthermore, controlling the timing of flocculation can help reduce potential growth declines that may arise from the energy and nutrient demands associated with synthesizing flocculating agents. Techniques such as utilizing ethanol-induced promoters in yeast may be similarly adapted to enhance elements, hence providing a sustainable approach to microalgal cultivation and harvesting.
2.3.5.1. Genetically Improvement of Self-Flocculation
In recent years, there has been a significant increase in the use of genetic and metabolic engineering in the field of microalgae. The main goal is to achieve more efficient and sustainable microalgal technology, as well as increased productivity and yield in the production processes of high value-added compounds derived from these photosynthetic microorganisms. However, these genetic tools have only been successfully demonstrated in a limited number of species, mainly due to their metabolic versatility, the lack of unique and strong promoters that unify the protocols, the low performance in the expression of heterologous genes and the instability of nuclear expression [119]. Nevertheless, in certain biological processes, the restricted understanding of their molecular bases makes this task especially challenging. This is exemplified by the mechanisms that regulate self-flocculation in microorganisms, with a particular focus on microalgae. Furthermore, there is a limited number of research groups globally that are dedicated to this topic, and a similarly limited number of published studies [86,120]. The fact that self-flocculation has been demonstrated to be a profitable and environmentally sustainable mechanism for microalgal biorefineries highlights the necessity to investigate its molecular mechanisms in greater depth in order to enhance yields through the utilisation of genetic engineering and to establish this mechanism as an effective solution to the reduction of chemical usage in microalgae biomass harvesting processes. With regard to the induction of self-flocculation in microalgae through genetic engineering, the existing bibliography is notably scarce. The majority of studies have focused on the examination of the dominant agents involved in this process, both in microalgae and in highly flocculent microorganisms that are widely utilized in industrial applications, such as bacteria and fermentation yeasts. As previously describe in 2.3.5, some authors have investigated the molecules involved in the flocculation mechanism of two self-flocculant microalgae from the Chlorella and Scenedesmus genus. In 2014, M. A. Alam and colleagues investigated the flocculation mechanism of Chlorella vulgaris JSC-7, [102] identifying cell wall polysaccharides as flocculating agents. Their crude extracts were observed to induce flocculation in both C. vulgaris CNW11 and Scenedesmus obliquus FS. Alternatively, a study by the authors Guo S et al., 2013 [121] on Scenedesmus obliquus AS-6-1 demonstrated that the self-flocculation of this microalgae was mediated by polysaccharides associated with the cell wall. These polysaccharides were found to comprise glucose, mannose, galactose, rhamnose and fructose in a molar ratio of 8:5:3:2:1. The cell wall polysaccharides may be the subject of genetic studies with the objective of inducing their expression in the cell walls of other microalgae and reorganising them to achieve a more adequate composition for flocculation. In a further investigation conducted by Lam and co-workers, the glycoproteins present in the cell wall and their interactions in the self-flocculent microalga E. texenis were elucidated, thereby opening the door to possible genetic modifications or integration of these glycoproteins and their interactions in other species of non-flocculating microalgae An alternative approach was the induction of self-flocculation in the model green microalga Chlamydomonas reinhardtii by expressing a flocculin gene from the highly flocculant yeast Saccharomyces bayanus (FLO5), which is responsible for the flocculation process in these microorganisms. The genetically engineered Chlamydomonas species exhibited self-flocculation capacities that were 2-3.5 times higher than those observed in the wild-type strain [122]. On the other hand, the ongoing advancement of technology and the emergence of novel tools such as genetic editing, synthetic biology, computational biology and artificial intelligence could facilitate the development of sustainable and viable genetic engineering techniques for the induction of self-flocculation. In this context, the utilization of CRISPR-Cas gene editing technology represents a promising approach for the development of genetically enhanced microalgae strains with enhanced self-flocculation capacity [123,124]. The application of CRISPR-Cas technology would facilitate the modification of specific genes that influence cell surface properties and the production of exopolysaccharides, which are pivotal in the flocculation process [33,125]. Such modifications could optimize the natural tendency of microalgae to group together in flocs, thereby significantly enhancing their self-flocculation ability. This would facilitate the harvesting and collection of algal biomasses, while reducing the costs associated with separating algae from the culture medium. Moreover, the generation of highly flocculent strains using CRISPR-Cas technology would allow the engineering of microalgae to enhance other beneficial properties, such as increased efficiency in CO₂ fixation [126] or high added-value compounds production [127].
Despite the lack of research exploring the use of CRISPR-Cas technology to induce self-flocculation in microalgae, the advances made in other areas demonstrate the potential of CRISPR technology in modifying microalgae to improve certain metabolic and biomolecule accumulation characteristics relevant to industry [128,129]. For instance, CRISPR has been utilized to increase the accumulation of lipids and other metabolites with potential utility in biofuels as in the case of Chlamydomonas reinhardtii [130], Nannochloropsis gaditana [131] or Chlorella spp. [132]. Furthermore, the use of CRISPR has enabled the manipulation of microalgal genes resulting in an increased yield of interesting industrial carotenoids as for example: astaxanthin in Chlamydomonas reinhardtii [133], β-carotene in Dunaliella salina [134] or even the marine carotenoid diatoxanthin in the diatom Phaeodactylum tricornutum [135]. Furthermore, the potential of CRISPR technology is being investigated in other areas of interest for microalgae, including its use for a sustainable aquaculture and an efficient biological treatment of wastewater [136]. In summary, the utilisation of genetic editing systems in microalgae has the potential to expedite the development of more efficient microalgal strains as a biological platform to produce high added-value molecules. This, in turn, could facilitate a feasible and suitable biomass concentration and harvesting for microalgal biorefineries.
3. Valuable Microalgae Compounds and Bio-Flocculation Implications
Microalgae are commonly referred to as ’sustainable bio-factories’ because of their ability to both reduce atmospheric carbon dioxide and produce a wide range of valuable compounds. However, the key to successful large-scale use of microalgae as bio-factories lies in the selection of the right microalgae and ideal growth conditions. A screening process should be established to select the best microalgae strains based on their growth, strength and metabolite production, thanks to the diverse range of microalgae species [137]. Current opportunities in microalgae technology are shifting towards the production of valuable bio-compounds such as lipids, carotenoids and other compounds for pharmaceutical, nutraceutical and cosmetic uses. However, the economic costs associated with microalgal biomass harvesting have emerged as a significant barrier to the market’s expansion for microalgae-derived products. In this context, bio-flocculation has garnered attention as a promising solution to mitigate these costs, offering a more environmentally friendly alternative. Despite the growing interest in this area, research efforts have been limited, with a scarcity of publications addressing bio-flocculation processes.
The subsequent sections offer a overview of the most relevant examples found in the literature on the implications of bio-flocculation for the production of high value-added compounds of major industrial relevance in microalgae biorefinery.
3.1. Lipids
The lipids produced by microalgae are a chemically diverse group of compounds, with the predominant characteristic of being insoluble in water and soluble in non-polar organic solvents. These lipids originate primarily in the plasma membrane, the endomembrane system, chloroplasts, and lipid bodies, where they are stored in the form of triacylglycerides and free fatty acids. Microalgal lipids can be classified into two main categories: neutral lipids and polar lipids. Neutral lipids, such as triglycerides (TAGs) and sterols, function as energy reserves, while polar lipids, including phospholipids and glycolipids, play essential structural roles within the cell. In addition to these classes, microalgae also produce waxes and hydrocarbons, which serve diverse functions. Notably, microalgae are capable of producing fatty acids with up to 24 carbons, and they are unique photosynthetic microorganisms in the production of omega-3 compounds, which are essential for cardiovascular health and cognitive development [138,139,140]. TAGs, in turn, not only serve as an energy reserve during adverse conditions, protecting the photosynthetic process, but are also key in the production of advanced biofuels due to their high productivity and photosynthetic efficiency.
Polyunsaturated fatty acids (PUFAs), such as eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA), found in microalgae, are highly regarded due to their health benefits as food supplements, and they lack plant-based alternatives. These characteristics position microalgae as a valuable resource for both the biofuel industry and the nutraceutical sector[139]. From an ecological perspective, microalgae lipids play a crucial role in aquatic food webs. These lipids serve as a primary source of essential fatty acids for marine organisms, including zooplankton, fish, and crustaceans. Furthermore, microalgae lipids are involved in regulating cell buoyancy, allowing microalgae to optimize their exposure to light for photosynthesis. In environmental terms, microalgae lipids have garnered attention as a sustainable alternative for the production of biofuels, particularly biodiesel, due to their high fatty acid content [141,142]. In addition, some hydrocarbons and waxes produced by microalgae can serve as precursors for bioplastics and other environmentally friendly bioproducts. Furthermore, microalgae’s capacity to sequester carbon dioxide during their growth makes them a promising solution for climate change mitigation given that microalgae contribute to the reduction of greenhouse gases while generating high-energy compounds [143].
Oleaginous microalgae are photosynthetic microorganisms that can accumulate substantial amounts of lipids, rendering them a promising source for the production of biofuels and other high-added-value compounds [144]. Their capacity to synthesize fatty acids under diverse environmental conditions renders them particularly attractive for applications in the energy and nutraceutical industries. Notable species include Botryococcus braunii, Nannochloropsis sp., Chlorella vulgaris, Neochloris oleoabundans, and Crypthecodinium cohnii, which exhibit lipid accumulation ranging from 20% to 80% of their dry weight [145]. These microalgae have been extensively studied due to their high oil production efficiency and their potential to substitute for fossil fuels in the production of sustainable energy. Despite the prevalence of mechanical or chemical processes in the current biomass collection level, there is an emerging body of research exploring bio-flocculation as a promising and sustainable alternative [146,147]). Some authors have explored the use of fungal culture-assisted bioflocculation, which has been demonstrated to improve the flocculation performance of microalgae species of interest. In 2015, Nazim Muradov and his team evaluated 33 fungal strains against microalgae species commonly utilized for biodiesel production, including the heterotrophic freshwater microalgae Chlorella protothecoides and the marine microalgae Tetraselmis suecica [148]. In addition, other authors have investigated the bio-flocculation of oleaginous microalgae using the natively adapted filamentous fungi Aspergillus niger, isolated from wastewater and its subsequent hydrothermal liquefaction for biofuel production [149]. Additionally, the lipid-rich cellulolytic fungus Aspergillus terreus, which was recently isolated, was evaluated as a bio-flocculant to enhance the collection yield of the oleaginous microalga Chlorella sp.. In this instance, lipids extracted from the Chlorella-Aspergillus co-culture were utilized for the production of biodiesel through methylation, yielding a substantial percentage of C18:1,2 unsaturated fatty acids, which is regarded as a suitable fraction for biodiesel production [75]. Conversely, other researchers have explored microalgae-bacteria co-cultures as a bioflocculation methodology. In this regard, Manogaran Lakshmikandan and colleagues examined the efficacy of co-cultivation of Asterococcus limneticus WL2 and Streptomyces rosealbus MTTC 12951 in both the harvesting processes and the production of lipids of interest for biodiesel [150]. In another sense, some authors have explored the use of naturally occurring flocculant molecules (bio-based flocculants) for the flocculation of microalgae and the production of biofuels. In 2018, Liandong Zhu et al. delved into the use of chitosan as a bio-flocculant compound on Chlorella vulgaris [151]. A substantial number of studies have been conducted on natural coagulant compounds and natural polymers, predominantly protein, polysaccharide, carbohydrate, or polyphenol compounds derived from various plant sources, including Moringa species, Strychnos potatorum, Plantago ovata or Jatropha curcu; from bacteria such as Bacillus megaterium SP1, lactic-acid bacteria or Arthrobacter humicola; fungi such as Aspergillus or Rhizopus; and the microalgae themselves [152]. In relation to this final point, in 2021 the self-flocculation capability of the oleaginous microalgae of interest for biodiesel, Auxenochlorella protothecoides UTEX 2341, was analyzed [153].
The implementation of bio-flocculation processes has been demonstrated to enhance the efficiency of biomass collection and to contribute to the economic and environmental viability of lipid production from microalgae biorefinery. This, in turn, supports the sustainable development of biofuels.
3.2. Carotenoids
Carotenoids are terpenoid pigments derived from tetraterpenes (C40). These hydrocarbons are classified into two principal groups: firstly, carotenes, which contain solely carbon and hydrogen atoms (e.g., α-carotene, β-carotene, γ-carotene, phytoene and lycopene), and secondly, xanthophylls, which possess a carbon, hydrogen and oxygen molecule structure [154].
Photosynthetic organisms, as well as some non-photosynthetic bacteria and fungi, are capable of synthesizing carotenoids and significantly microalgae are among the most prolific producers of naturally occurring carotenoids [155] (Figure 2).
Carotenoids obtained from microalgae constitute a sustainable and valuable source of bioactive compounds, exhibiting applications in diverse industries, including food, pharmaceutical, and cosmetics. These pigments, notably astaxanthin from Haematococcus pluvialis, β-carotene from Dunaliella salina, or lutein from Chlorella vulgaris, possess potent antioxidant, anti-inflammatory, and photoprotective properties, rendering them essential in nutritional supplements and sunscreens [128]. Moreover, the extraction of these pigments from microalgae within a biorefinery enables the optimization of biomass utilization, integrating the production of bioenergy, proteins, and other high-added-value compounds in a circular economy model. The increasing demand for natural pigments, in conjunction with limitations on synthetic dyes, has prompted the development of more efficient and sustainable extraction and purification technologies for carotenoids, thereby solidifying microalgae as a promising alternative in the context of biorefinery [156]. In this perspective, bio-flocculation is presented as an efficient and sustainable strategy for harvesting microalgae biomass in biorefineries aimed at producing carotenoids without the need to add external chemical agents.
Despite the limited availability of research on the application of bio-flocculation processes for collecting microalgal biomass exclusively for the extraction of carotenoids, some studies have reported the potential use of these methods for this purpose. For example, in 2022, Hui Liu et al. stated the application of co-cultures of filamentous fungi with microalgae for the treatment of wastewater and its association with the production of biomass enriched with high-added-value compounds, such as carotenoids [157]. The use of co-cultures of Aspergillus sp., Ganoderma lucidum, or Trichoderma reesei was studied primarily with microalgae such as Chlorella sp., Scenedesmus sp., or Haematococcus pluvialis, which are rich in lutein and astaxanthin. Moreover, a more recent study by Goswami R et al. reported the utilization of fungi-assisted bio-flocculation for the harvesting of the microalgae Picochlorum sp. using Aspergillus niger and, the potential subsequent production of astaxanthin and β-carotene [158]. On the other hand, certain studies have reported the use of co-cultures of flocculent microalgae and non-flocculent microalgae to increase the harvest efficiency of the latter and the yields of carotenoids production such as β-carotene. For instance, the co-cultures of Coelastrum cf. pseudomicroporum and Scenedesmus sp. have been examined [159] as well as Skeletonema sp. with Euglena sp. [160] and the co-culture of the marine microalgae Nannochloropsis oculata and Phaeodactylum tricornutum [161]. Finally, some authors have explored the self-flocculant capacity of certain microalgae species, studying the most beneficial abiotic conditions for an optimal increase in said process with applicability in an efficient and sustainable harvesting of microalgal biomass and the production of carotenoids, such as β-carotene, lutein, violaxanthin, canthaxanthin, and astaxanthin. In 2022, Malik and his team characterized a newly self-flocculant algal strain identified as Bracteacoccus pseudominor BERC09, which revealed its promising potential to produce carotenoids and lipids [162]. And, in 2024 Galan et al. focused on the study of culture parameters that could influence the self-flocculation of Haematococcus pluvialis with the aim of improving its harvesting yields for a further production of astaxanthin among other valuable industrial carotenoids [163].
3.3. Proteins
Microalgae have garnered heightened interest among food technologists due to their distinctive advantages over traditional protein sources. These organisms exhibit rapid growth, are not contingent on arable land, require minimal water, and can produce a diverse array of bioactive compounds. Notably, species such as Chlorella vulgaris, Arthrospira platensis, Dunaliella salina and Haematococcus pluvialis are particularly noteworthy for their exceptional protein composition, which can account for up to 70% of their dry weight [164]. These microorganisms offer a sustainable source of protein and possess an intriguing amino acid profile, rich in essential amino acids, similar to some animal protein sources, such as egg [165]. Consequently, microalgae are regarded as promising protein sources due to their distinctive amino acid profile and the presence of numerous bioactive peptides derived from species such as Porphyridium, Nostoc commune, Chlorella pyrenoidosa and Arthrospira platensis [166]. This renders them ideal for application in the food, cosmetics, and pharmaceutical industries. Existing studies have reported a substantial increase in the protein content of various food products, following the incorporation of microalgae biomass. However, the utilization and development of microalgae for protein-rich foods for humans is in the early stages, and thus, they are considered an alternative protein source for the future [167]. Microalgae proteins have important applications, such as emulsifying, foaming, and gelling properties, which are crucial in determining the quality and texture of foods. Furthermore, microalgae peptides have been shown to possess bioactivity with various health-promoting effects, including antioxidant, antidiabetic, and antihypertensive properties, as well as antibacterial, osteogenic, anticancer, and anti-aging activities [164,168].
Microalgae have been identified as a promising organism for the production of recombinant proteins due to their unique characteristics and ability to grow under controlled conditions. Species such as Chlamydomonas reinhardtii, Phaeodactylum tricornutum and Nannochloropsis gaditana have been the focus of extensive research for their potential in the production of therapeutic proteins and vaccines. Notably, Chlamydomonas reinhardtii has been utilized to produce recombinant proteins, including EGF (epidermal growth factor) and monoclonal antibodies, which have demonstrated applications in cancer treatment and immunological diseases [169,170]. Additionally, Phaeodactylum tricornutum has demonstrated its efficacy in producing proteins with antiviral activity, which could be pivotal in the development of novel treatments for viral infections [171]. Nannochloropsis gaditana has been utilized to produce bioactive proteins with antioxidant and anti-inflammatory properties, rendering it suitable for applications in the food and cosmetics industry [172]. These investigations underscore the potential of microalgae as biotechnological platforms for the sustainable production of recombinant proteins with diverse industrial and medical applications.
Bio-flocculation has the potential to become a viable process for the harvesting of microalgal proteins. This is due to its efficiency in cell aggregation, its low environmental impact, and its potential cost reduction compared to traditional methods such as centrifugation or filtration. Despite the absence of specific studies on the recovery of microalgal proteins by bio-flocculation, the existing evidence on its efficacy in biomass separation suggests that this method could facilitate the concentration and recovery of proteins without the use of aggressive chemical agents, thus preserving their functionality and nutritional quality. In addition, the possibility of using natural bio-flocculants or those produced by endogenous microorganisms minimizes contamination of the final product, aligning with the principles of sustainable bioprocesses. Consequently, further exploration in this domain has the potential to establish bio-flocculation as a cost-effective and eco-friendly approach for the large-scale production of microalgal proteins.
3.4. Other Valuable Compounds
Microalgae have been identified as a promising source of secondary metabolites, with potential applications in various industrial sectors, including agriculture, food, and pharmacology. These molecules of interest include phytohormones, polysaccharides, carbohydrates such as starch, substances with biocidal activity or micro and macro nutrients.
Phytohormones and Biostimulant Molecules
Several species of microalgae synthesize phytohormones that act as biostimulants in higher plants. For instance, Chlamydomonas reinhardtii, Chlorella vulgaris, Scenedesmus obliquus, Thalassiosira sp., Nannochloropsis oceanica, Haematococcus pluvialis or the cyanobacteria Arthrospira platensis, Synechococcus sp. or Anabaena sp. are known to produce auxins, gibberellins, and cytokines, which have been demonstrated to stimulate plant growth, enhance root development, and fortify plants against abiotic stress. These phytohormones have found application in the formulation of agricultural biostimulants, resulting in enhanced crop yield and quality [173,174,175,176].
Biocidal Molecules
Some microalgae produce compounds with biocidal activity that can replace synthetic agrochemicals, promoting more sustainable agriculture. Chlorella vulgaris and Nannochloropsis oculata have been shown to have antibacterial and antifungal activity against plant pathogens such as Fusarium oxysporum and Pseudomonas syringae. Furthermore, Oscillatoria agardhii and other cyanobacteria of the Nostoc genus have been found to produce compounds with insecticidal and antifungal properties, which underscores their potential as ecological alternatives for the integrated management of agricultural pests and diseases [177,178].
Enriched Biomass and Biofertilization
Microalgal biomass is not only a source of bioactive metabolites but can also be directly used as a biofertilizer. Species such as Scenedesmus obliquus, Chlorella vulgaris and Anabaena cylindrica are rich in nitrogen, phosphorus, and essential micronutrients that can improve soil fertility and plant nutrition. Furthermore, algal biomass has been shown to enhance soil structure and support beneficial microbial activity, thereby contributing to more sustainable and efficient agricultural systems [179,180].
Polysaccharides
Polysaccharides derived from microalgae possess significant bioactive properties. Chlorella vulgaris is recognized for its ability to produce polysaccharides with immunostimulant and antioxidant activities, making them candidates for applications in the food and pharmaceutical industries. Likewise, Porphyridium cruentum and Arthrospira platensis synthesize sulfated polysaccharides with antiviral and anti-inflammatory activity, which has sparked interest in the biomedical field[181,182,183].
Starch and Other Carbohydrates
Species such as Chlamydomonas reinhardtii, Nannochloropsis gaditana, Chlorella vulgaris and Botryococcus brainii accumulate starch and other carbohydrates of industrial interest. These compounds are of interest for use in the production of biofuels and bioplastics, which offer sustainable alternatives to conventional materials. In addition, the ability of some microalgae to modulate carbohydrate synthesis under different environmental conditions makes them ideal candidates for the production of bioenergy and high value-added bioproducts [184,185].
While there is a lack of studies exclusively employing bio-flocculation for the recovery of these minor molecules, its application in other contexts has demonstrated significant advantages, including reduced operating costs, lower environmental impact, and better preservation of bioactive compounds sensitive to more aggressive processes, for using bio-flocculation as an alternative harvesting method for these types of bio-compounds.
4. Conclusion
The metabolic diversity of microalgae and their capacity to produce a wide range of bioactive compounds underscores their potential as biotechnological resources for biorefinery. Pursuing ongoing research in this domain is imperative to enhance their cultivation and extraction, thereby facilitating their incorporation into sustainable industrial applications. The recovery of bioactive molecules produced by microalgae necessitates the implementation of efficient harvesting and extraction methods. Conventional techniques employed include centrifugation, membrane filtration, dissolved air flotation, and chemical or biological flocculation. Centrifugation and filtration are highly effective methods, but they require a significant amount of energy and financial resources. Chemical flocculation, while efficient, can leave unwanted residues in the biomass. In this context, bio-flocculation emerges as a promising and sustainable alternative for the harvesting of microalgal biomass. This method uses metabolites excreted by the microalgae themselves or natural microbial consortia to induce cell aggregation and their subsequent separation. It does not require the use of synthetic chemical agents, which can be a significant environmental benefit. The optimization and scaling of this approach could represent a significant advancement in microalgae biotechnology, aligning with more sustainable and efficient production strategies for obtaining biofertilizers, biostimulants, and biocides of microalgal origin.
5. Future Perspectives
The future of bio-flocculation as a green tool for recovering valuable compounds from microalgae in biorefineries lies in the integration of advanced technologies to enhance efficiency and scalability. Genetic engineering and synthetic biology could be employed to develop highly efficient flocculating microorganisms with tailored properties, improving recovery rates and selectivity. Additionally, the induction of self-flocculation in microalgae through metabolic engineering or controlled environmental triggers could reduce the need for external flocculants, simplifying downstream processing and lowering costs. Artificial intelligence and machine learning algorithms can optimize process parameters, predict flocculation behaviour, and enhance operational efficiency. Furthermore, emerging bioinformatics tools will enable the identification of key genes and metabolic pathways involved in bio-flocculation, facilitating strain improvement and process optimization. By combining these innovative approaches, bio-flocculation can become a more precise, cost-effective, and sustainable strategy, reinforcing its role in circular economy-driven biorefineries.
Author Contributions
ED-S conceived and designed the project. Manuscript writing—original draft preparation, LGH-M, AMG-D and ED-S; writing—editing, ED-S. The manuscript was corrected, revised and approved by all authors. All authors have read and agreed to the published version of the manuscript.
Funding
This project was funded by the own authors of the manuscript.
Institutional Review Board Statement
Not applicable
Data Availability Statement
Not applicable
Acknowledgments
All the figures in the manuscript were created in https://BioRender.com
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Wang, H.; Zhang, Y.; Chen, L.; Cheng, W.; Liu, T. Combined Production of Fucoxanthin and EPA from Two Diatom Strains Phaeodactylum Tricornutum and Cylindrotheca Fusiformis Cultures. Bioprocess Biosyst Eng 2018, 41, 1061–1071. [Google Scholar] [CrossRef] [PubMed]
- Darwesh, O.M.; Matter, I.A.; Eida, M.F.; Moawad, H.; Oh, Y.K. Influence of Nitrogen Source and Growth Phase on Extracellular Biosynthesis of Silver Nanoparticles Using Cultural Filtrates of Scenedesmus Obliquus. Applied Sciences (Switzerland) 2019, 9. [Google Scholar] [CrossRef]
- Caspeta, L.; Buijs, N.A.A.; Nielsen, J. The Role of Biofuels in the Future Energy Supply. Energy Environ Sci 2013, 6, 1077–1082. [Google Scholar] [CrossRef]
- Abas, N.; Kalair, A.; Khan, N. Review of Fossil Fuels and Future Energy Technologies. Futures 2015, 69, 31–49. [Google Scholar] [CrossRef]
- Ray, A.; Banerjee, S.; Das, D. Microalgal Bio-Flocculation: Present Scenario and Prospects for Commercialization. [CrossRef]
- Malik, S.; Khan, F.; Atta, Z.; Habib, N.; Haider, M.N.; Wang, N.; Alam, A.; Jambi, E.J.; Gull, M.; Mehmood, M.A.; et al. Microalgal Flocculation: Global Research Progress and Prospects for Algal Biorefinery. Biotechnol Appl Biochem 2020, 67, 52–60. [Google Scholar] [CrossRef]
- Mathimani, T.; Senthil Kumar, T.; Chandrasekar, M.; Uma, L.; Prabaharan, D. Assessment of Fuel Properties, Engine Performance and Emission Characteristics of Outdoor Grown Marine Chlorella Vulgaris BDUG 91771 Biodiesel. Renew Energy 2017, 105, 637–646. [Google Scholar] [CrossRef]
- Li, T.; Hu, J.; Zhu, L. Self-flocculation as an Efficient Method to Harvest Microalgae: A Mini-review. Water (Switzerland) 2021, 13. [Google Scholar] [CrossRef]
- Milledge, J.J.; Heaven, S. A Review of the Harvesting of Micro-Algae for Biofuel Production. Rev Environ Sci Biotechnol 2013, 12, 165–178. [Google Scholar] [CrossRef]
- Zhu, L.; Nugroho, Y.K.; Shakeel, S.R.; Li, Z.; Martinkauppi, B.; Hiltunen, E. Using Microalgae to Produce Liquid Transportation Biodiesel: What Is Next? Renewable and Sustainable Energy Reviews 2017, 78, 391–400. [Google Scholar] [CrossRef]
- Goh, B.H.H.; Ong, H.C.; Cheah, M.Y.; Chen, W.H.; Yu, K.L.; Mahlia, T.M.I. Sustainability of Direct Biodiesel Synthesis from Microalgae Biomass: A Critical Review. Renewable and Sustainable Energy Reviews 2019, 107, 59–74. [Google Scholar] [CrossRef]
- Chia, S.R.; Ong, H.C.; Chew, K.W.; Show, P.L.; Phang, S.M.; Ling, T.C.; Nagarajan, D.; Lee, D.J.; Chang, J.S. Sustainable Approaches for Algae Utilisation in Bioenergy Production. Renew Energy 2018, 129, 838–852. [Google Scholar] [CrossRef]
- Borges, L.; Morón-Villarreyes, J.A.; D’Oca, M.G.M.; Abreu, P.C. Effects of Flocculants on Lipid Extraction and Fatty Acid Composition of the Microalgae Nannochloropsis Oculata and Thalassiosira Weissflogii. Biomass Bioenergy 2011, 35, 4449–4454. [Google Scholar] [CrossRef]
- Williams, P.J.L.B.; Laurens, L.M.L. Microalgae as Biodiesel & Biomass Feedstocks: Review & Analysis of the Biochemistry, Energetics & Economics. Energy Environ Sci 2010, 3, 554–590. [Google Scholar] [CrossRef]
- Kumar, K.; Mishra, S.K.; Shrivastav, A.; Park, M.S.; Yang, J.W. Recent Trends in the Mass Cultivation of Algae in Raceway Ponds. Renewable and Sustainable Energy Reviews 2015, 51, 875–885. [Google Scholar] [CrossRef]
- Eze, C.N.; Onyejiaka, C.K.; Ihim, S.A.; Ayoka, T.O.; Aduba, C.C.; Ndukwe, J.K.; Nwaiwu, O.; Onyeaka, H. Bioactive Compounds by Microalgae and Potentials for the Management of Some Human Disease Conditions. AIMS Microbiol 2023, 9, 55–74. [Google Scholar] [PubMed]
- Branyikova, I.; Filipenska, M.; Urbanova, K.; Ruzicka, M.C.; Pivokonsky, M.; Branyik, T. Physicochemical Approach to Alkaline Flocculation of Chlorella Vulgaris Induced by Calcium Phosphate Precipitates. Colloids Surf B Biointerfaces 2018, 166, 54–60. [Google Scholar] [CrossRef]
- Lee, Y.C.; Lee, K.; Oh, Y.K. Recent Nanoparticle Engineering Advances in Microalgal Cultivation and Harvesting Processes of Biodiesel Production: A Review. Bioresour Technol 2015, 184, 63–72. [Google Scholar] [CrossRef]
- Li, S.; Hu, T.; Xu, Y.; Wang, J.; Chu, R.; Yin, Z.; Mo, F.; Zhu, L. A Review on Flocculation as an Efficient Method to Harvest Energy Microalgae: Mechanisms, Performances, Influencing Factors and Perspectives. Renewable and Sustainable Energy Reviews 2020, 131, 110005. [Google Scholar] [CrossRef]
- Vandamme, D.; Pontes, S.C.V.; Goiris, K.; Foubert, I.; Pinoy, L.J.J.; Muylaert, K. Evaluation of Electro-Coagulation-Flocculation for Harvesting Marine and Freshwater Microalgae. Biotechnol Bioeng 2011, 108, 2320–2329. [Google Scholar] [CrossRef]
- El-Baz, F.K.; Gad, M.S.; Abdo, S.M.; Abed, K.A.; Matter, I.A. Performance and Exhaust Emissions of a Diesel Engine Burning Algal Biodiesel Blends. International Journal of Mechanical and Mechatronics Engineering 2016, 16, 151–158. [Google Scholar]
- Van Haver, L.; Nayar, S. Polyelectrolyte Flocculants in Harvesting Microalgal Biomass for Food and Feed Applications. Algal Res 2017, 24, 167–180. [Google Scholar] [CrossRef]
- Sanyano, N.; Chetpattananondh, P.; Chongkhong, S. Coagulation-Flocculation of Marine Chlorella Sp. for Biodiesel Production. Bioresour Technol 2013, 147, 471–476. [Google Scholar] [CrossRef]
- Singh, G.; Patidar, S.K. Microalgae Harvesting Techniques: A Review. J Environ Manage 2018, 217, 499–508. [Google Scholar] [CrossRef]
- Mathimani, T.; Mallick, N. A Comprehensive Review on Harvesting of Microalgae for Biodiesel - Key Challenges and Future Directions. Renewable and Sustainable Energy Reviews 2018, 91, 1103–1120. [Google Scholar] [CrossRef]
- Wan, C.; Alam, M.A.; Zhao, X.Q.; Zhang, X.Y.; Guo, S.L.; Ho, S.H.; Chang, J.S.; Bai, F.W. Current Progress and Future Prospect of Microalgal Biomass Harvest Using Various Flocculation Technologies. Bioresour Technol 2015, 184, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Son, J.; Sung, M.; Ryu, H.; Oh, Y.K.; Han, J.I. Microalgae Dewatering Based on Forward Osmosis Employing Proton Exchange Membrane. Bioresour Technol 2017, 244, 57–62. [Google Scholar] [CrossRef]
- Laamanen, C.A.; Ross, G.M.; Scott, J.A. Flotation Harvesting of Microalgae. Renewable and Sustainable Energy Reviews 2016, 58, 75–86. [Google Scholar] [CrossRef]
- Demir, I.; Besson, A.; Guiraud, P.; Formosa-Dague, C. Towards a Better Understanding of Microalgae Natural Flocculation Mechanisms to Enhance Flotation Harvesting Efficiency. Water Science and Technology 2020, 82, 1009–1024. [Google Scholar] [CrossRef]
- Sukenik, A.; Bilanovic, D.; Shelef, G. Flocculation of Microalgae in Brackish and Sea Waters. Biomass 1988, 15, 187–199. [Google Scholar] [CrossRef]
- Battah, M.; El-Ayoty, Y.; El-Fatah Abomohra, A.; El-Ghany, S.A.; Esmael, A.; Chaudhary, R.; Khattar, J.; Singh, D.; Luísa da Cunha Gonçalves, A.; José Vieira Simões José Carlos Magalhães Pires, M.; et al. Flocculation as a Low-Cost Method for Harvesting Microalgae for Bulk Biomass Production KU Leuven Kulak, Laboratory Aquatic Biology, E. Sabbelaan 53, 8500 Kortrijk, Corresponding Author: Trends Biotechnol 2013, 31, 233–239. [Google Scholar]
- Wong, Y.K.; Ho, Y.H.; Leung, H.M.; Ho, K.C.; Yau, Y.H.; Yung, K.K.L. Enhancement of Chlorella Vulgaris Harvesting via the Electro-Coagulation-Flotation (ECF) Method. Environmental Science and Pollution Research 2017, 24, 9102–9110. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Graf, A.; Hernández, S.; Morales, M. Biomitigation of CO2 from Flue Gas by Scenedesmus Obtusiusculus AT-UAM Using a Hybrid Photobioreactor Coupled to a Biomass Recovery Stage by Electro-Coagulation-Flotation. Environmental Science and Pollution Research 2020, 27, 28561–28574. [Google Scholar] [CrossRef]
- Kim, D.Y.; Lee, K.; Lee, J.; Lee, Y.H.; Han, J.I.; Park, J.Y.; Oh, Y.K. Acidified-Flocculation Process for Harvesting of Microalgae: Coagulant Reutilization and Metal-Free-Microalgae Recovery. Bioresour Technol 2017, 239, 190–196. [Google Scholar] [CrossRef]
- Xu, H.; Tang, Z.; Yang, D.; Dai, X.; Chen, H. Enhanced Growth and Auto-Flocculation of Scenedesmus Quadricauda in Anaerobic Digestate Using High Light Intensity and Nanosilica: A Biomineralization-Inspired Strategy. Water Res 2023, 235, 119893. [Google Scholar] [CrossRef] [PubMed]
- Cerff, M.; Morweiser, M.; Dillschneider, R.; Michel, A.; Menzel, K.; Posten, C. Harvesting Fresh Water and Marine Algae by Magnetic Separation: Screening of Separation Parameters and High Gradient Magnetic Filtration. Bioresour Technol 2012, 118, 289–295. [Google Scholar] [CrossRef]
- Lee, K.; Lee, S.Y.; Na, J.G.; Jeon, S.G.; Praveenkumar, R.; Kim, D.M.; Chang, W.S.; Oh, Y.K. Magnetophoretic Harvesting of Oleaginous Chlorella Sp. by Using Biocompatible Chitosan/Magnetic Nanoparticle Composites. Bioresour Technol 2013, 149, 575–578. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.K.; Chieh, D.C.J.; Jalak, S.A.; Toh, P.Y.; Yasin, N.H.M.; Ng, B.W.; Ahmad, A.L. Rapid Magnetophoretic Separation of Microalgae. Small 2012, 8, 1683–1692. [Google Scholar] [CrossRef]
- Sumathi, Y.; Kumar, P.; Singhania, R.R.; Chen, C.W.; Gurunathan, B.; Dong, C. Di; Patel, A.K. Harnessing Fe3O4 Nanoparticles for Sustainable Harvesting of Astaxanthin-Producing Microalgae: Advancing Industrial-Scale Biorefinery. Sep Purif Technol 2025, 353, 128408. [Google Scholar] [CrossRef]
- Tripathi, G.; Pandey, V.K.; Ahmad, S.; Irum; Khujamshukurov, N. A.; Farooqui, A.; Mishra, V. Utilizing Novel Aspergillus Species for Bio-Flocculation: A Cost-Effective Approach to Harvest Scenedesmus Microalgae for Biofuel Production. Curr Res Microb Sci 2024, 7, 100272. [Google Scholar] [CrossRef]
- Xu, H.; Liu, C.; Wang, A.; Yue, B.; Lin, T.; Ding, M. Microalgae Treatment of Food Processing Wastewater for Simultaneous Biomass Resource Recycling and Water Reuse. J Environ Manage 2024, 369, 122394. [Google Scholar] [CrossRef]
- Chekli, L.; Corjon, E.; Tabatabai, S.A.A.; Naidu, G.; Tamburic, B.; Park, S.H.; Shon, H.K. Performance of Titanium Salts Compared to Conventional FeCl3 for the Removal of Algal Organic Matter (AOM) in Synthetic Seawater: Coagulation Performance, Organic Fraction Removal and Floc Characteristics. J Environ Manage 2017, 201, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Maia, C.; Pôjo, V.; Tavares, T.; Pires, J.C.M.; Malcata, F.X. Surfactant-Mediated Microalgal Flocculation: Process Efficiency and Kinetic Modelling. Bioengineering 2024, 11, 1–14. [Google Scholar] [CrossRef]
- Yudong, N.; Tao, Z.; Haihua, W.; Haixing, C. Upcycling Harmful Algal Blooms into Short-Chain Organic Matters Assisted with Cellulose-Based Flocculant. Bioresour Technol 2024, 397, 130425. [Google Scholar] [CrossRef]
- Papazi, A.; Makridis, P.; Divanach, P. Harvesting Chlorella Minutissima Using Cell Coagulants. J Appl Phycol 2010, 22, 349–355. [Google Scholar] [CrossRef]
- Rwehumbiza, V.M.; Harrison, R.; Thomsen, L. Alum-Induced Flocculation of Preconcentrated Nannochloropsis Salina: Residual Aluminium in the Biomass, FAMEs and Its Effects on Microalgae Growth upon Media Recycling. Chemical Engineering Journal 2012, 200–202, 168–175. [Google Scholar] [CrossRef]
- Beach, E.S.; Eckelman, M.J.; Cui, Z.; Brentner, L.; Zimmerman, J.B. Preferential Technological and Life Cycle Environmental Performance of Chitosan Flocculation for Harvesting of the Green Algae Neochloris Oleoabundans. Bioresour Technol 2012, 121, 445–449. [Google Scholar] [CrossRef]
- Zheng, H.; Gao, Z.; Yin, J.; Tang, X.; Ji, X.; Huang, H. Harvesting of Microalgae by Flocculation with Poly (γ-Glutamic Acid). Bioresour Technol 2012, 112, 212–220. [Google Scholar] [CrossRef]
- Wang, X.; Chen, Y.P.; Guo, J.S.; Fang, F.; Yan, P. Flocculation-Enhanced Photobiological Hydrogen Production by Microalgae: Flocculant Composition, Hydrogenase Activity and Response Mechanism. Chemical Engineering Journal 2024, 485, 150065. [Google Scholar] [CrossRef]
- Tran, D.T.; Le, B.H.; Lee, D.J.; Chen, C.L.; Wang, H.Y.; Chang, J.S. Microalgae Harvesting and Subsequent Biodiesel Conversion. Bioresour Technol 2013, 140, 179–186. [Google Scholar] [CrossRef]
- Swar, S.S.; Boonnorat, J.; Ghimire, A. Algae-Based Treatment of a Landfill Leachate Pretreated by Coagulation-Flocculation. J Environ Manage 2023, 342, 118223. [Google Scholar] [CrossRef]
- Marinho, Y.F.; de Oliveira, A.P.S.; Oliveira, C.Y.B.; Napoleão, T.H.; Guedes Paiva, P.M.; de Sant’Anna, M.C.S.; Malafaia, C.B.; Gálvez, A.O. Usage of Moringa Oleifera Residual Seeds Promotes Efficient Flocculation of Tetradesmus Dimorphus Biomass. Biomass Convers Biorefin 2022. [Google Scholar] [CrossRef]
- Lama, S.; Pappa, M.; Brandão Watanabe, N.; Formosa–Dague, C.; Marchal, W.; Adriaensens, P.; Vandamme, D. Interference of Extracellular Soluble Algal Organic Matter on Flocculation–Sedimentation Harvesting of Chlorella Sp. Bioresour Technol 2024, 411. [Google Scholar] [CrossRef]
- Chen, F.; Liu, Z.; Li, D.; Liu, C.; Zheng, P.; Chen, S. Using Ammonia for Algae Harvesting and as Nutrient in Subsequent Cultures. Bioresour Technol 2012, 121, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Imbimbo, P.; Ferrara, A.; Giustino, E.; Liberti, D.; Monti, D.M. Microalgae Flocculation: Assessment of Extraction Yields and Biological Activity. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef]
- Endrawati, H.; Widianingsih, W.; Nuraini, R.A.T.; Hartati, R.; Redjeki, S.; Riniatsih, I.; Mahendrajaya, R.T. The Effect of Chitosan Concentration on Flocculation Efficiency Microalgae Porphyridium Cruentum (Rhodhophyta). IOP Conf Ser Earth Environ Sci 2021, 919. [Google Scholar] [CrossRef]
- Gani, P.; Apandi, N.M.; Mohamed Sunar, N.; Matias-Peralta, H.M.; Kean Hua, A.; Mohd Dzulkifli, S.N.; Parjo, U.K. Outdoor Phycoremediation and Biomass Harvesting Optimization of Microalgae Botryococcus Sp. Cultivated in Food Processing Wastewater Using an Enclosed Photobioreactor. Int J Phytoremediation 2022, 24, 1431–1443. [Google Scholar] [CrossRef]
- Nguyen, T.D.P.; Tran, T.N.T.; Le, T.V.A.; Nguyen Phan, T.X.; Show, P.L.; Chia, S.R. Auto-Flocculation through Cultivation of Chlorella Vulgaris in Seafood Wastewater Discharge: Influence of Culture Conditions on Microalgae Growth and Nutrient Removal. J Biosci Bioeng 2019, 127, 492–498. [Google Scholar] [CrossRef]
- Pérez, L.; Salgueiro, J.L.; Maceiras, R.; Cancela, Á.; Sánchez, Á. An Effective Method for Harvesting of Marine Microalgae: PH Induced Flocculation. Biomass Bioenergy 2017, 97, 20–26. [Google Scholar] [CrossRef]
- Schlesinger, A.; Eisenstadt, D.; Bar-Gil, A.; Carmely, H.; Einbinder, S.; Gressel, J. Inexpensive Non-Toxic Flocculation of Microalgae Contradicts Theories; Overcoming a Major Hurdle to Bulk Algal Production. Biotechnol Adv 2012, 30, 1023–1030. [Google Scholar] [CrossRef]
- Verfaillie, A.; Blockx, J.; Praveenkumar, R.; Thielemans, W.; Muylaert, K. Harvesting of Marine Microalgae Using Cationic Cellulose Nanocrystals. Carbohydr Polym 2020, 240, 116165. [Google Scholar] [CrossRef]
- Mixson, S.M.; Stikeleather, L.F.; Simmons, O.D.; Wilson, C.W.; Burkholder, J.A.M. PH-Induced Flocculation, Indirect Electrocoagulation, and Hollow Fiber Filtration Techniques for Harvesting the Saltwater Microalga Dunaliella. J Appl Phycol 2014, 26, 1701–1709. [Google Scholar] [CrossRef]
- Spilling, K.; Seppälä, J.; Tamminen, T. Inducing Autoflocculation in the Diatom Phaeodactylum Tricornutum through CO2 Regulation. J Appl Phycol 2011, 23, 959–966. [Google Scholar] [CrossRef]
- ’t Lam, G.P.; Vermuë, M.H.; Olivieri, G.; van den Broek, L.A.M.; Barbosa, M.J.; Eppink, M.H.M.; Wijffels, R.H.; Kleinegris, D.M.M. Cationic Polymers for Successful Flocculation of Marine Microalgae. Bioresour Technol 2014, 169, 804–807. [Google Scholar] [CrossRef]
- Wu, Z.; Zhu, Y.; Huang, W.; Zhang, C.; Li, T.; Zhang, Y.; Li, A. Evaluation of Flocculation Induced by PH Increase for Harvesting Microalgae and Reuse of Flocculated Medium. Bioresour Technol 2012, 110, 496–502. [Google Scholar] [CrossRef]
- An, L.; Xu, X.; Yan, F.; Du, W.; Dai, R. Simultaneous Removal of Cyanobacteria and Algal Organic Matter by Mn(VII)/CaSO3 Enhanced Coagulation: Performance and Mechanism. J Hazard Mater 2025, 485. [Google Scholar] [CrossRef]
- Hu, Y.R.; Wang, F.; Wang, S.K.; Liu, C.Z.; Guo, C. Efficient Harvesting of Marine Microalgae Nannochloropsis Maritima Using Magnetic Nanoparticles. Bioresour Technol 2013, 138, 387–390. [Google Scholar] [CrossRef]
- Van Den Hende, S.; Carré, E.; Cocaud, E.; Beelen, V.; Boon, N.; Vervaeren, H. Treatment of Industrial Wastewaters by Microalgal Bacterial Flocs in Sequencing Batch Reactors. Bioresour Technol 2014, 161, 245–254. [Google Scholar] [CrossRef]
- Kim, D.G.; La, H.J.; Ahn, C.Y.; Park, Y.H.; Oh, H.M. Harvest of Scenedesmus Sp. with Bioflocculant and Reuse of Culture Medium for Subsequent High-Density Cultures. Bioresour Technol 2011, 102, 3163–3168. [Google Scholar] [CrossRef]
- Chen, W.; Wang, T.; Dou, Z.; Xie, X. Microalgae Harvesting by Self-Driven 3D Microfiltration with Rationally Designed Porous Superabsorbent Polymer (PSAP) Beads. Environ Sci Technol 2021, 55, 15446–15455. [Google Scholar] [CrossRef]
- Al-Humairi, S.T.; Lee, J.G.M.; Harvey, A.P.; Salman, A.D.; Juzsakova, T.; Van, B.; Le, P.C.; La, D.D.; Mungray, A.K.; Show, P.L.; et al. A Foam Column System Harvesting Freshwater Algae for Biodiesel Production: An Experiment and Process Model Evaluations. Science of the Total Environment 2023, 862. [Google Scholar] [CrossRef]
- Li, Y.; Ma, Q.; Pan, Y.; Chen, Q.; Sun, Z.; Hu, P. Development of an Effective Flocculation Method by Utilizing the Auto-Flocculation Capability of Phaeodactylum Tricornutum. Algal Res 2021, 58, 102413. [Google Scholar] [CrossRef]
- Gorin, K. V.; Sergeeva, Y.E.; Butylin, V. V.; Komova, A. V.; Pojidaev, V.M.; Badranova, G.U.; Shapovalova, A.A.; Konova, I.A.; Gotovtsev, P.M. Methods Coagulation/Flocculation and Flocculation with Ballast Agent for Effective Harvesting of Microalgae. Bioresour Technol 2015, 193, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.-M.; Lee, S.J.; Park, M.-H.; Kim, H.-S.; Kim, H.-C.; Yoon, J.-H.; Kwon, G.-S.; Yoon, B.-D. Harvesting of Chlorella Vulgaris Using a Bioflocculant from Paenibacillus Sp. AM49; 2001; Vol. 23;
- Ayad, H.I.; Matter, I.A.; Gharieb, M.M.; Darwesh, O.M. Bioflocculation Harvesting of Oleaginous Microalga Chlorella Sp. Using Novel Lipid-Rich Cellulolytic Fungus Aspergillus Terreus (MD1) for Biodiesel Production. Biomass Convers Biorefin. [CrossRef]
- Tripathi, G.; Pandey, V.K.; Ahmad, S. ; Irum; Khujamshukurov, N.A.; Farooqui, A.; Mishra, V. Utilizing Novel Aspergillus Species for Bio-Flocculation: A Cost-Effective Approach to Harvest Scenedesmus Microalgae for Biofuel Production. Curr Res Microb Sci.
- Cho, K.; Hur, S.P.; Lee, C.H.; Ko, K.; Lee, Y.J.; Kim, K.N.; Kim, M.S.; Chung, Y.H.; Kim, D.; Oda, T. Bioflocculation of the Oceanic Microalga Dunaliella Salina by the Bloom-Forming Dinoflagellate Heterocapsa Circularisquama, and Its Effect on Biodiesel Properties of the Biomass. Bioresour Technol 2016, 202, 257–261. [Google Scholar] [CrossRef]
- Roselet, F.; Burkert, J.; Abreu, P.C. Flocculation of Nannochloropsis Oculata Using a Tannin-Based Polymer: Bench Scale Optimization and Pilot Scale Reproducibility. Biomass Bioenergy 2016, 87, 55–60. [Google Scholar] [CrossRef]
- Farid, M.S.; Shariati, A.; Badakhshan, A.; Anvaripour, B. Using Nano-Chitosan for Harvesting Microalga Nannochloropsis Sp. Bioresour Technol 2013, 131, 555–559. [Google Scholar] [CrossRef]
- Al-Hothaly, K.A. An Optimized Method for the Bio-Harvesting of Microalgae, Botryococcus Braunii, Using Aspergillus Sp. in Large-Scale Studies. MethodsX 2018, 5, 788–794. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; He, G. Influence of Nutritional Conditions on Exopolysaccharide Production by Submerged Cultivation of the Medicinal Fungus Shiraia Bambusicola. World J Microbiol Biotechnol 2008, 24, 2903–2907. [Google Scholar] [CrossRef]
- Mahapatra, S.; Banerjee, D. Fungal Exopolysaccharide: Production, Composition and Applications. Microbiol Insights 2013, 6, MBI.S10957. [Google Scholar] [CrossRef]
- Civzele, A.; Mezule, L. Fungal – Assisted Microalgae Flocculation and Simultaneous Lignocellulolytic Enzyme Production in Wastewater Treatment Systems. Biotechnology Reports 2025, 45. [Google Scholar] [CrossRef]
- Powell, R.J.; Hill, R.T. Mechanism of Algal Aggregation by Bacillus Sp. Strain RP1137. Appl Environ Microbiol 2014, 80, 4042–4050. [Google Scholar] [CrossRef]
- Wan, C.; Zhao, X.Q.; Guo, S.L.; Asraful Alam, M.; Bai, F.W. Bioflocculant Production from Solibacillus Silvestris W01 and Its Application in Cost-Effective Harvest of Marine Microalga Nannochloropsis Oceanica by Flocculation. Bioresour Technol 2013, 135, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Lakshmikandan, M.; Murugesan, A.G.; Ameen, F.; Maneeruttanarungroj, C.; Wang, S. Efficient Bioflocculation and Biodiesel Production of Microalgae Asterococcus Limneticus on Streptomyces Two-Stage Co-Cultivation Strategy. Biomass Bioenergy 2023, 175, 106886. [Google Scholar] [CrossRef]
- Ndikubwimana, T.; Zeng, X.; Liu, Y.; Chang, J.S.; Lu, Y. Harvesting of Microalgae Desmodesmus Sp. F51 by Bioflocculation with Bacterial Bioflocculant. Algal Res 2014, 6, 186–193. [Google Scholar] [CrossRef]
- Cho, K.; Hur, S.P.; Lee, C.H.; Ko, K.; Lee, Y.J.; Kim, K.N.; Kim, M.S.; Chung, Y.H.; Kim, D.; Oda, T. Bioflocculation of the Oceanic Microalga Dunaliella Salina by the Bloom-Forming Dinoflagellate Heterocapsa Circularisquama, and Its Effect on Biodiesel Properties of the Biomass. Bioresour Technol 2016, 202, 257–261. [Google Scholar] [CrossRef]
- Oh, H.M.; Lee, S.J.; Park, M.H.; Kim, H.S.; Kim, H.C.; Yoon, J.H.; Kwon, G.S.; Yoon, B.D. Harvesting of Chlorella Vulgaris Using a Bioflocculant from Paenibacillus Sp. AM49. Biotechnol Lett 2001, 23, 1229–1234. [Google Scholar] [CrossRef]
- Lei, X.; Chen, Y.; Shao, Z.; Chen, Z.; Li, Y.; Zhu, H.; Zhang, J.; Zheng, W.; Zheng, T. Effective Harvesting of the Microalgae Chlorella Vulgaris via Flocculation-Flotation with Bioflocculant. Bioresour Technol 2015, 198, 922–925. [Google Scholar] [CrossRef]
- Li, Y.; Xu, Y.; Liu, L.; Jiang, X.; Zhang, K.; Zheng, T.; Wang, H. First Evidence of Bioflocculant from Shinella Albus with Flocculation Activity on Harvesting of Chlorella Vulgaris Biomass. Bioresour Technol 2016, 218, 807–815. [Google Scholar] [CrossRef]
- Vogelaar, J.C.T.; De Keizer, A.; Spijker, S.; Lettinga, G. Bioflocculation of Mesophilic and Thermophilic Activated Sludge. Water Res 2005, 39, 37–46. [Google Scholar] [CrossRef]
- Lee, M.; Woo, S.G.; Ten, L.N. Shinella Daejeonensis Sp. Nov., a Nitrate-Reducing Bacterium Isolated from Sludge of a Leachate Treatment Plant. Int J Syst Evol Microbiol 2011, 61, 2123–2128. [Google Scholar] [CrossRef]
- Matsui, T.; Shinzato, N.; Tamaki, H.; Muramatsu, M.; Hanada, S. Shinella Yambaruensis Sp. Nov., a 3-Methy-Sulfolane-Assimilating Bacterium Isolated from Soil. Int J Syst Evol Microbiol 2009, 59, 536–539. [Google Scholar] [CrossRef]
- Lal, A.; Das, D. Biomass Production and Identification of Suitable Harvesting Technique for Chlorella Sp. MJ 11/11 and Synechocystis PCC 6803. 3 Biotech 2016, 6, 1–10. [Google Scholar] [CrossRef]
- Xiao, R.; Zheng, Y. Overview of Microalgal Extracellular Polymeric Substances (EPS) and Their Applications. Biotechnol Adv 2016, 34, 1225–1244. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J. The Biofilm Matrix. Nat Rev Microbiol 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Paulsen, B.S.; Aslaksen, T.; Freire-Nordi, C.S.; Vieira, A.A.H. Extracellular Polysaccharides from Ankistrodesmus Densus (Chlorophyceae). J Phycol 1998, 34, 638–641. [Google Scholar] [CrossRef]
- Saadaoui, I.; Cherif, M.; Siddiqui, S.A.; Al Jabri, H.; Sayadi, S. Algal-Algal Bioflocculation Enhances the Recovery Efficiency of Picochlorum Sp. QUCCCM130 with Low Auto-Settling Capacity. Algal Res 2023, 71, 103038. [Google Scholar] [CrossRef]
- Salim, S.; Bosma, R.; Vermuë, M.H.; Wijffels, R.H. Harvesting of Microalgae by Bio-Flocculation. J Appl Phycol 2011, 23, 849–855. [Google Scholar] [CrossRef]
- Salim, S.; Vermuë, M.H.; Wijffels, R.H. Ratio between Autoflocculating and Target Microalgae Affects the Energy-Efficient Harvesting by Bio-Flocculation. Bioresour Technol 2012, 118, 49–55. [Google Scholar] [CrossRef]
- Alam, M.A.; Wan, C.; Guo, S.L.; Zhao, X.Q.; Huang, Z.Y.; Yang, Y.L.; Chang, J.S.; Bai, F.W. Characterization of the Flocculating Agent from the Spontaneously Flocculating Microalga Chlorella Vulgaris JSC-7. J Biosci Bioeng 2014, 118, 29–33. [Google Scholar] [CrossRef]
- Salim, S.; Kosterink, N.R.; Tchetkoua Wacka, N.D.; Vermuë, M.H.; Wijffels, R.H. Mechanism behind Autoflocculation of Unicellular Green Microalgae Ettlia Texensis. J Biotechnol 2014, 174, 34–38. [Google Scholar] [CrossRef]
- Zhao, F.; Xiao, J.; Ding, W.; Cui, N.; Yu, X.; Xu, J.W.; Li, T.; Zhao, P. An Effective Method for Harvesting of Microalga: Coculture-Induced Self-Flocculation. J Taiwan Inst Chem Eng 2019, 100, 117–126. [Google Scholar] [CrossRef]
- Guo, S.L.; Zhao, X.Q.; Tang, Y.; Wan, C.; Alam, M.A.; Ho, S.H.; Bai, F.W.; Chang, J.S. Establishment of an Efficient Genetic Transformation System in Scenedesmus Obliquus. J Biotechnol 2013, 163, 61–68. [Google Scholar] [CrossRef]
- Li, Y.; Ma, Q.; Pan, Y.; Chen, Q.; Sun, Z.; Hu, P. Development of an Effective Flocculation Method by Utilizing the Auto-Flocculation Capability of Phaeodactylum Tricornutum. Algal Res 2021, 58, 102413. [Google Scholar] [CrossRef]
- Muir, E.; Grossman, A.R.; Chisti, Y.; Fedrizzi, B.; Guieysse, B.; Plouviez, M. Self-Aggregation for Sustainable Harvesting of Microalgae. Algal Res 2024, 83, 103685. [Google Scholar] [CrossRef]
- Zhao, X.Q.; Bai, F.W. Yeast Flocculation: New Story in Fuel Ethanol Production. Biotechnol Adv 2009, 27, 849–856. [Google Scholar] [CrossRef]
- Bauer, F.F.; Govender, P.; Bester, M.C. Yeast Flocculation and Its Biotechnological Relevance. Appl Microbiol Biotechnol 2010, 88, 31–39. [Google Scholar] [CrossRef]
- Alam, A.; Vandamme, D.; Chun, W.; Zhao, X.; Foubert, I.; Wang, Z.; Muylaert, K.; Yuan, Z. Bioflocculation as an Innovative Harvesting Strategy for Microalgae. Rev Environ Sci Biotechnol 2016, 15, 573–583. [Google Scholar] [CrossRef]
- Wu, Z.; Zhu, Y.; Huang, W.; Zhang, C.; Li, T.; Zhang, Y.; Li, A. Evaluation of Flocculation Induced by PH Increase for Harvesting Microalgae and Reuse of Flocculated Medium. Bioresour Technol 2012, 110, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Aljuboori, A.H.R.; Uemura, Y.; Thanh, N.T. Flocculation and Mechanism of Self-Flocculating Lipid Producer Microalga Scenedesmus Quadricauda for Biomass Harvesting. Biomass Bioenergy 2016, 93, 38–42. [Google Scholar] [CrossRef]
- Tan, D.; Zhang, H.; Sun, S.; Dong, F.; Sun, H.; Li, B. Rapid Flocculation-Sedimentation of Microalgae with Organosilane-Functionalized Halloysite. Appl Clay Sci 2019, 177, 37–42. [Google Scholar] [CrossRef]
- Rai, A.; Sirotiya, V.; Mourya, M.; Khan, M.J.; Ahirwar, A.; Sharma, A.K.; Kawatra, R.; Marchand, J.; Schoefs, B.; Varjani, S.; et al. Sustainable Treatment of Dye Wastewater by Recycling Microalgal and Diatom Biogenic Materials: Biorefinery Perspectives. Chemosphere 2022, 305, 135371. [Google Scholar] [CrossRef]
- Chen, Z.; Shao, S.; He, Y.; Luo, Q.; Zheng, M.; Zheng, M.; Chen, B.; Wang, M. Nutrients Removal from Piggery Wastewater Coupled to Lipid Production by a Newly Isolated Self-Flocculating Microalga Desmodesmus Sp. PW1. Bioresour Technol 2020, 302, 122806. [Google Scholar] [CrossRef]
- Cheng, P.; Chen, D.; Liu, W.; Cobb, K.; Zhou, N.; Liu, Y.; Liu, H.; Wang, Q.; Chen, P.; Zhou, C.; et al. Auto-Flocculation Microalgae Species Tribonema Sp. and Synechocystis Sp. with T-IPL Pretreatment to Improve Swine Wastewater Nutrient Removal. Science of the Total Environment 2020, 725, 138263. [Google Scholar] [CrossRef] [PubMed]
- Kilian, O.; Benemann, C.S.E.; Niyogi, K.K.; Vick, B. High-Efficiency Homologous Recombination in the Oil-Producing Alga Nannochloropsis Sp. Proc Natl Acad Sci U S A 2011, 108, 21265–21269. [Google Scholar] [CrossRef] [PubMed]
- Trentacoste, E.M.; Shrestha, R.P.; Smith, S.R.; Glé, C.; Hartmann, A.C.; Hildebrand, M.; Gerwick, W.H. Metabolic Engineering of Lipid Catabolism Increases Microalgal Lipid Accumulation without Compromising Growth. Proc Natl Acad Sci U S A 2013, 110, 19748–19753. [Google Scholar] [CrossRef]
- Valverde, F.; Romero-Campero, F.J.; León, R.; Guerrero, M.G.; Serrano, A. New Challenges in Microalgae Biotechnology. Eur J Protistol 2016, 55, 95–101. [Google Scholar] [CrossRef]
- Liu, Z.; Hao, N.; Hou, Y.; Wang, Q.; Liu, Q.; Yan, S.; Chen, F.; Zhao, L. Technologies for Harvesting the Microalgae for Industrial Applications: Current Trends and Perspectives. Bioresour Technol 2023, 387. [Google Scholar]
- Guo, S.L.; Zhao, X.Q.; Wan, C.; Huang, Z.Y.; Yang, Y.L.; Asraful Alam, M.; Ho, S.H.; Bai, F.W.; Chang, J.S. Characterization of Flocculating Agent from the Self-Flocculating Microalga Scenedesmus Obliquus AS-6-1 for Efficient Biomass Harvest. Bioresour Technol 2013, 145, 285–289. [Google Scholar] [CrossRef]
- Díaz-Santos, E.; Vila, M.; Vigara, J.; León, R. A New Approach to Express Transgenes in Microalgae and Its Use to Increase the Flocculation Ability of Chlamydomonas Reinhardtii. J Appl Phycol 2016, 28, 1611–1621. [Google Scholar] [CrossRef]
- Bhatia, S. ; Pooja; Yadav, S.K. CRISPR-Cas for Genome Editing: Classification, Mechanism, Designing and Applications. Int J Biol Macromol 2023. [Google Scholar]
- Pickar-Oliver, A.; Gersbach, C.A. The next Generation of CRISPR–Cas Technologies and Applications. Nat Rev Mol Cell Biol 2019, 20, 490–507. [Google Scholar]
- Patel, A.K.; Singhania, R.R.; Awasthi, M.K.; Varjani, S.; Bhatia, S.K.; Tsai, M.L.; Hsieh, S.L.; Chen, C.W.; Dong, C. Di Emerging Prospects of Macro- and Microalgae as Prebiotic. Microb Cell Fact 2021, 20, 1–16. [Google Scholar] [CrossRef]
- Lee, T.M.; Lin, J.Y.; Tsai, T.H.; Yang, R.Y.; Ng, I.S. Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) Technology and Genetic Engineering Strategies for Microalgae towards Carbon Neutrality: A Critical Review. Bioresour Technol 2023, 368. [Google Scholar] [CrossRef] [PubMed]
- Dhokane, D.; Shaikh, A.; Yadav, A.; Giri, N.; Bandyopadhyay, A.; Dasgupta, S.; Bhadra, B. CRISPR-Based Bioengineering in Microalgae for Production of Industrially Important Biomolecules. Front Bioeng Biotechnol 2023, 11. [Google Scholar] [CrossRef]
- Aditi; Bhardwaj, R. ; Yadav, A.; Swapnil, P.; Meena, M. Characterization of Microalgal β-Carotene and Astaxanthin: Exploring Their Health-Promoting Properties under the Effect of Salinity and Light Intensity. Biotechnology for Biofuels and Bioproducts 2025, 18, 18. [Google Scholar] [CrossRef]
- Arshad, S.; Qadir, M.L.; Hussain, N.; Ali, Q.; Han, S.; Ali, D. Advances in CRISPR/Cas9 Technology: Shaping the Future of Photosynthetic Microorganisms for Biofuel Production. Functional Plant Biology 2025, 52. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.H.T.; Park, S.; Jeong, J.; Shin, Y.S.; Sim, S.J.; Jin, E.S. Enhancing Lipid Productivity by Modulating Lipid Catabolism Using the CRISPR-Cas9 System in Chlamydomonas. J Appl Phycol 2020, 32, 2829–2840. [Google Scholar] [CrossRef]
- Ajjawi, I.; Verruto, J.; Aqui, M.; Soriaga, L.B.; Coppersmith, J.; Kwok, K.; Peach, L.; Orchard, E.; Kalb, R.; Xu, W.; et al. Lipid Production in Nannochloropsis Gaditana Is Doubled by Decreasing Expression of a Single Transcriptional Regulator. Nat Biotechnol 2017, 35. [Google Scholar] [CrossRef]
- Lin, W.R.; Ng, I.S. Development of CRISPR/Cas9 System in Chlorella Vulgaris FSP-E to Enhance Lipid Accumulation. Enzyme Microb Technol 2020, 133, 109458. [Google Scholar] [CrossRef] [PubMed]
- Kneip, J.S.; Kniepkamp, N.; Jang, J.; Mortaro, M.G.; Jin, E.; Kruse, O.; Baier, T. CRISPR/Cas9-Mediated Knockout of the Lycopene ε-Cyclase for Efficient Astaxanthin Production in the Green Microalga Chlamydomonas Reinhardtii. Plants 2024, 13, 1393. [Google Scholar] [CrossRef]
- Hu, L.; Feng, S.; Liang, G.; Du, J.; Li, A.; Niu, C. CRISPR/Cas9-Induced β-Carotene Hydroxylase Mutation in Dunaliella Salina CCAP19/18. AMB Express 2021, 11. [Google Scholar] [CrossRef]
- Græsholt, C.; Brembu, T.; Volpe, C.; Bartosova, Z.; Serif, M.; Winge, P.; Nymark, M. Zeaxanthin Epoxidase 3 Knockout Mutants of the Model Diatom Phaeodactylum Tricornutum Enable Commercial Production of the Bioactive Carotenoid Diatoxanthin. Mar Drugs 2024, 22, 185. [Google Scholar] [CrossRef]
- Ambily, B.; Limna Mol, V.P.; Sini, H.; Nevin, K.G. CRISPR-Based Microalgal Genome Editing and the Potential for Sustainable Aquaculture: A Comprehensive Review. J Appl Phycol 2024.
- Fernandes, T.; Cordeiro, N. Microalgae as Sustainable Biofactories to Produce High-Value Lipids: Biodiversity, Exploitation, and Biotechnological Applications. Mar Drugs 2021, 19. [Google Scholar] [CrossRef]
- Manning, S.R. Microalgal Lipids: Biochemistry and Biotechnology. Curr Opin Biotechnol 2022, 74, 1–7. [Google Scholar] [CrossRef]
- Xin, Y.; Wu, S.; Miao, C.; Xu, T.; Lu, Y. Towards Lipid from Microalgae: Products, Biosynthesis, and Genetic Engineering. Life 2024, 14, 447. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.; Wang, Y.; Zuo, M.; Wang, X.; Shen, B. Structural Analysis of Isolated Components in Microalgal Lipids: Neutral Lipids, Phospholipids, and Glycolipids. Bioresour Technol Rep 2025, 29, 102083. [Google Scholar] [CrossRef]
- Oliveira, J.; Costa, E.; Maia Dias, J.; Pires, J.C. Biodiesel Production by Biocatalysis Using Lipids Extracted from Microalgae Oil of Chlorella Vulgaris and Aurantiochytrium Sp. Bioenergy Res 2023. [Google Scholar] [CrossRef]
- Kiyani, D.A.; Maryam, S.; Amina, S.J.; Ahmad, A.; Chattha, M.W.A.; Janjua, H.A. Lipid Extraction and Analysis of Microalgae Strain Pectinodesmus PHM3 for Biodiesel Production. BMC Biotechnol 2023, 23. [Google Scholar] [CrossRef]
- Zhang, C.; Singh, R.P.; Yadav, P.; Kumar, I.; Kaushik, A.; Roychowdhury, R.; Mubeen, M.; Singh, S.K.; Kumar, A.; Wang, J. Recent Advances in Biotechnology and Bioengineering for Efficient Microalgal Biofuel Production. Fuel Processing Technology 2025, 270, 108199. [Google Scholar] [CrossRef]
- Mutanda, T.; Naidoo, D.; Bwapwa, J.K.; Anandraj, A. Biotechnological Applications of Microalgal Oleaginous Compounds: Current Trends on Microalgal Bioprocessing of Products. Front Energy Res 2020, 8. [Google Scholar] [CrossRef]
- Wang, M.; Ye, X.; Bi, H.; Shen, Z. Microalgae Biofuels: Illuminating the Path to a Sustainable Future amidst Challenges and Opportunities. Biotechnology for Biofuels and Bioproducts 2024, 17. [Google Scholar]
- Zhang, L.; Tsui, T.-H.; Tong, Y.W.; Liu, R.; Baganz, F. Harvesting of Oleaginous Microbial Cells and Extraction of Microbial Lipids. In Microbial Lipids. In Microbial Lipids and Biodiesel Technologies; Springer Nature Singapore: Singapore, 2025; pp. 223–239. [Google Scholar]
- Zhang, L.; Tsui, T.-H.; Tong, Y.W.; Liu, R.; Aggarangsi, P. Microbial Lipid Technology Based on Oleaginous Microalgae. In Microbial Lipids and Biodiesel Technologies; Springer Nature Singapore: Singapore, 2025; pp. 77–100. [Google Scholar]
- Muradov, N.; Taha, M.; Miranda, A.F.; Wrede, D.; Kadali, K.; Gujar, A.; Stevenson, T.; Ball, A.S.; Mouradov, A. Fungal-Assisted Algal Flocculation: Application in Wastewater Treatment and Biofuel Production. Biotechnol Biofuels 2015, 8. [Google Scholar] [CrossRef]
- Jaiswal, K.K.; Kumar, V.; Gururani, P.; Vlaskin, M.S.; Parveen, A.; Nanda, M.; Kurbatova, A.; Gautam, P.; Grigorenko, A. V. Bio-Flocculation of Oleaginous Microalgae Integrated with Municipal Wastewater Treatment and Its Hydrothermal Liquefaction for Biofuel Production. Environ Technol Innov 2022, 26, 102340. [Google Scholar] [CrossRef]
- Lakshmikandan, M.; Murugesan, A.G.; Ameen, F.; Maneeruttanarungroj, C.; Wang, S. Efficient Bioflocculation and Biodiesel Production of Microalgae Asterococcus Limneticus on Streptomyces Two-Stage Co-Cultivation Strategy. Biomass Bioenergy 2023, 175. [Google Scholar] [CrossRef]
- Zhu, L.; Li, Z.; Hiltunen, E. Theoretical Assessment of Biomethane Production from Algal Residues after Biodiesel Production. Wiley Interdiscip Rev Energy Environ 2018, 7. [Google Scholar] [CrossRef]
- Ogbonna, C.N.; Nwoba, E.G. Bio-Based Flocculants for Sustainable Harvesting of Microalgae for Biofuel Production. A Review. Renewable and Sustainable Energy Reviews 2021, 139, 110690. [Google Scholar] [CrossRef]
- Li, J.; Li, B.; Yang, J. Bio-Flocculation Property Analyses of Oleaginous Microalgae Auxenochlorella Protothecoides Utex 2341. Sustainability (Switzerland) 2021, 13, 1–11. [Google Scholar] [CrossRef]
- Tjandraatmadja, G.; Diaper, C. Sources of Critical Contaminants in Domestic Wastewater - a Literature Review. 2006, 88.
- Rengel, R.; Giraldez, I.; Díaz, M.J.; García, T.; Vigara, J.; León, R. Simultaneous Production of Carotenoids and Chemical Building Blocks Precursors from Chlorophyta Microalgae. Bioresour Technol 2022, 351. [Google Scholar] [CrossRef]
- Tanaka, K.; Lan, J.C.-W.; Kondo, A.; Hasunuma, T. Metabolic Engineering and Cultivation Strategies for Efficient Production of Fucoxanthin and Related Carotenoids. Appl Microbiol Biotechnol 2025, 109, 57. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Chen, L.; Tan, Z.; Deng, Z.; Liu, H. Application of Filamentous Fungi in Microalgae-Based Wastewater Remediation for Biomass Harvesting and Utilization: From Mechanisms to Practical Application. Algal Res 2022, 62. [Google Scholar] [CrossRef]
- Goswami, R.K.; Mehariya, S.; Karthikeyan, O.P.; Verma, P. Fungi-Assisted Bio-Flocculation of Picochlorum Sp.: A Novel Bio-Assisted Treatment System for Municipal Wastewater. Journal of Water Process Engineering 2024, 57. [Google Scholar] [CrossRef]
- Úbeda, B.; Gálvez, J.Á.; Michel, M.; Bartual, A. Microalgae Cultivation in Urban Wastewater: Coelastrum Cf. Pseudomicroporum as a Novel Carotenoid Source and a Potential Microalgae Harvesting Tool. Bioresour Technol 2017, 228, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Indahsari, H.S.; Tassakka, A.C.M.A.R.; Dewi, E.N.; Yuwono, M.; Suyono, E.A. Effects of Salinity and Bioflocculation during Euglena Sp. Harvest on the Production of Lipid, Chlorophyll, and Carotenoid with Skeletonema Sp. as a Bioflocculant. J Pure Appl Microbiol 2022, 16. [Google Scholar] [CrossRef]
- Elshobary, M.E.; Ebaid, R.; Alquraishi, M.; Ende, S.S.W. Synergistic Microalgal Cocultivation: Boosting Flocculation, Biomass Production, and Fatty Acids Profile of Nannochloropsis Oculata and Phaeodactylum Tricornutum. Biomass Bioenergy 2025, 193, 107595. [Google Scholar] [CrossRef]
- Malik, S.; Ashraf, M.U.F.; Shahid, A.; Javed, M.R.; Khan, A.Z.; Usman, M.; Manivannan, A.; Mehmood, M.A.; Ashraf, G.A. Characterization of a Newly Isolated Self-Flocculating Microalga Bracteacoccus Pseudominor BERC09 and Its Evaluation as a Candidate for a Multiproduct Algal Biorefinery. Chemosphere 2022, 304, 135346. [Google Scholar] [CrossRef]
- Galán-González, J.; Quintero-Zapata, I.; Elías-Santos, M.; Galán-Wong, L.J.; López-Chuken, U.J.; Guajardo-Barbosa, C.; Beltrán-Rocha, J.C. Harvest by Autoflocculation, Biomass, and Carotenoid Production in Sequential Batch Culture of Haematococcus Pluvialis Under High Ionic Strength and Macroelements Content. J Mar Sci Technol 2024, 32. [Google Scholar] [CrossRef]
- Nunes, E.; Odenthal, K.; Nunes, N.; Fernandes, T.; Fernandes, I.A.; Pinheiro de Carvalho, M.A.A. Protein Extracts from Microalgae and Cyanobacteria Biomass. Techno-Functional Properties and Bioactivity: A Review. Algal Res 2024, 82, 103638. [Google Scholar] [CrossRef]
- Menon, R.; Raja, T.; Kumar, S.; Gokhale, T. Biorefinery Approach to Obtain Sustainable Biofuels and High-Value Chemicals from Microalgae. Microalgal Biofuels 2025, 109–137. [Google Scholar] [CrossRef]
- Ashraf, A.; Guo, Y.; Yang, T.; Ud Din, A.S.; Ahmad, K.; Li, W.; Hou, H. Microalgae-Derived Peptides: Exploring Bioactivities and Functional Food Innovations. J Agric Food Chem 2025. [Google Scholar] [CrossRef]
- Song, Y.; Hu, Z.; Liu, S.; Luo, S.; He, R.; Yang, X.; Li, S.; Yang, X.; An, Y.; Lu, Y. Utilization of Microalgae and Duckweed as Sustainable Protein Sources for Food and Feed: Nutritional Potential and Functional Applications. J Agric Food Chem 2025.
- Elisha, C.; Bhagwat, P.; Pillai, S. Emerging Production Techniques and Potential Health Promoting Properties of Plant and Animal Protein-Derived Bioactive Peptides. Crit Rev Food Sci Nutr 2024.
- Quezada-Rivera, J.J.; Ponce-Alonso, J.; Davalos-Guzman, S.D.; Soria-Guerra, R.E. Chlamydomonas Reinhardtii for the Production of Recombinant Proteins: Current Knowledge and Perspectives. Fundamentals of Recombinant Protein Production, Purification and Characterization. [CrossRef]
- Ma, K.; Deng, L.; Wu, H.; Fan, J. Towards Green Biomanufacturing of High-Value Recombinant Proteins Using Promising Cell Factory: Chlamydomonas Reinhardtii Chloroplast. Bioresour Bioprocess 2022, 9. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Serrano, B.V.; Cabanillas-Salcido, S.L.; Cordero-Rivera, C.D.; Jiménez-Camacho, R.; Norzagaray-Valenzuela, C.D.; Calderón-Zamora, L.; De Jesús-González, L.A.; Reyes-Ruiz, J.M.; Farfan-Morales, C.N.; Romero-Utrilla, A.; et al. Antiviral Effect of Microalgae Phaeodactylum Tricornutum Protein Hydrolysates against Dengue Virus Serotype 2. Mar Drugs 2024, 22. [Google Scholar] [CrossRef]
- Paterson, S.; Alonso-Pintre, L.; Morato-López, E.; González de la Fuente, S.; Gómez-Cortés, P.; Hernández-Ledesma, B. Microalga Nannochloropsis Gaditana as a Sustainable Source of Bioactive Peptides: A Proteomic and In Silico Approach. Foods 2025, 14. [Google Scholar] [CrossRef] [PubMed]
- Stirk, W.A.; Ordog, V.; Novák, O.; Rolčík, J.; Strnad, M.; Balint, P.; van Staden, J. Auxin and Cytokinin Relationships in Twenty-Four Microalgae Strains. J Phycol 2013, 49, 459–467. [Google Scholar] [CrossRef]
- Stirk, W.A.; Bálint, P.; Tarkowská, D.; Novák, O.; Strnad, M.; Ördög, V.; van Staden, J. Hormone Profiles in Microalgae: Gibberellins and Brassinosteroids. Plant Physiology and Biochemistry 2013, 70, 348–353. [Google Scholar] [CrossRef]
- Lu, Y.; Xu, J. Phytohormones in Microalgae: A New Opportunity for Microalgal Biotechnology? Trends Plant Sci 2015, 20, 273–282. [Google Scholar] [CrossRef]
- Puente-Padilla, B.L.; Romero-Villegas, G.I.; Sánchez-Estrada, A.; Cira-Chávez, L.A.; Estrada-Alvarado, M.I. Effect of Marine Microalgae Biomass (Nannochloropsis Gaditana and Thalassiosira Sp. ) on Germination and Vigor on Bean (Phaseolus Vulgaris L.) Seeds “Higuera.” Life 2025, 15, 386. [Google Scholar] [CrossRef]
- Cock, I.E.; Cheesman, M.J. A Review of the Antimicrobial Properties of Cyanobacterial Natural Products. Molecules 2023, 28. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, R.; Yadav, A.; Sahoo, A.; Kumari, P.; Singh, L.A.; Swapnil, P.; Meena, M.; Kumar, S. Microalgal-Based Sustainable Bio-Fungicides: A Promising Solution to Enhance Crop Yield. Discover Sustainability 2025, 6. [Google Scholar] [CrossRef]
- Parmar, P.; Kumar, R.; Neha, Y.; Srivatsan, V. Microalgae as next Generation Plant Growth Additives: Functions, Applications, Challenges and Circular Bioeconomy Based Solutions. Front Plant Sci 2023, 14. [Google Scholar] [CrossRef]
- Nur, M.M.A.; Mahreni; Murni, S. W.; Setyoningrum, T.M.; Hadi, F.; Widayati, T.W.; Jaya, D.; Sulistyawati, R.R.E.; Puspitaningrum, D.A.; Dewi, R.N.; et al. Innovative Strategies for Utilizing Microalgae as Dual-Purpose Biofertilizers and Phycoremediators in Agroecosystems. Biotechnology Reports 2025, 45, e00870. [Google Scholar] [CrossRef]
- Guehaz, K.; Boual, Z.; Abdou, I.; Telli, A.; Belkhalfa, H. Microalgae’s Polysaccharides, Are They Potent Antioxidants? Critical Review. Arch Microbiol 2024, 206. [Google Scholar]
- Babich, O.; Ivanova, S.; Michaud, P.; Budenkova, E.; Kashirskikh, E.; Anokhova, V.; Sukhikh, S. Synthesis of Polysaccharides by Microalgae Chlorella Sp. Bioresour Technol 2024, 406, 131043. [Google Scholar] [CrossRef]
- Pointcheval, M.; Massé, A.; Floc’hlay, D.; Chanonat, F.; Estival, J.; Durand, M.J. Antimicrobial Properties of Selected Microalgae Exopolysaccharide-Enriched Extracts: Influence of Antimicrobial Assays and Targeted Microorganisms. Front Microbiol 2025, 16. [Google Scholar] [CrossRef]
- Six, A.; Dimitriades-Lemaire, A.; Lancelon-Pin, C.; Putaux, J.L.; Dauvillée, D.; Petroutsos, D.; Alvarez Diaz, P.; Sassi, J.F.; Li-Beisson, Y.; Fleury, G. Red Light Induces Starch Accumulation in Chlorella Vulgaris without Affecting Photosynthesis Efficiency, Unlike Abiotic Stress. Algal Res 2024, 80, 103515. [Google Scholar] [CrossRef]
- Sarkar, P.; Bandyopadhyay, T.K.; Gopikrishna, K.; Nath Tiwari, O.; Bhunia, B.; Muthuraj, M. Algal Carbohydrates: Sources, Biosynthetic Pathway, Production, and Applications. Bioresour Technol 2024, 413, 131489. [Google Scholar] [CrossRef]
Figure 1.
The main events involved in flocs formation in the flocculation process. (Charges neutralization; bridging; patching).
Figure 1.
The main events involved in flocs formation in the flocculation process. (Charges neutralization; bridging; patching).
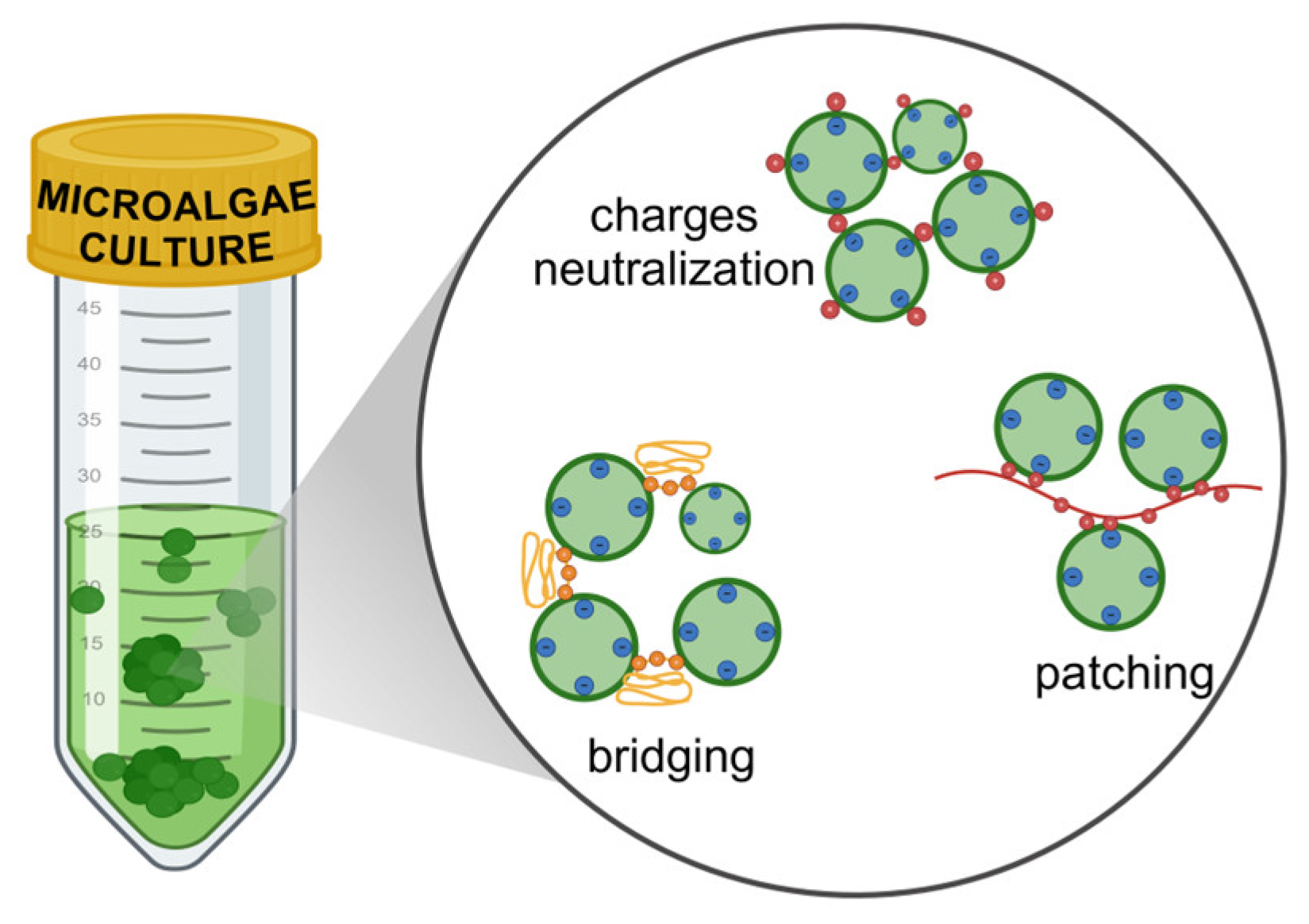
Figure 2.
Some of the most relevant carotenoids produced by microalgae. Chemical structures and example species are illustrated.
Figure 2.
Some of the most relevant carotenoids produced by microalgae. Chemical structures and example species are illustrated.
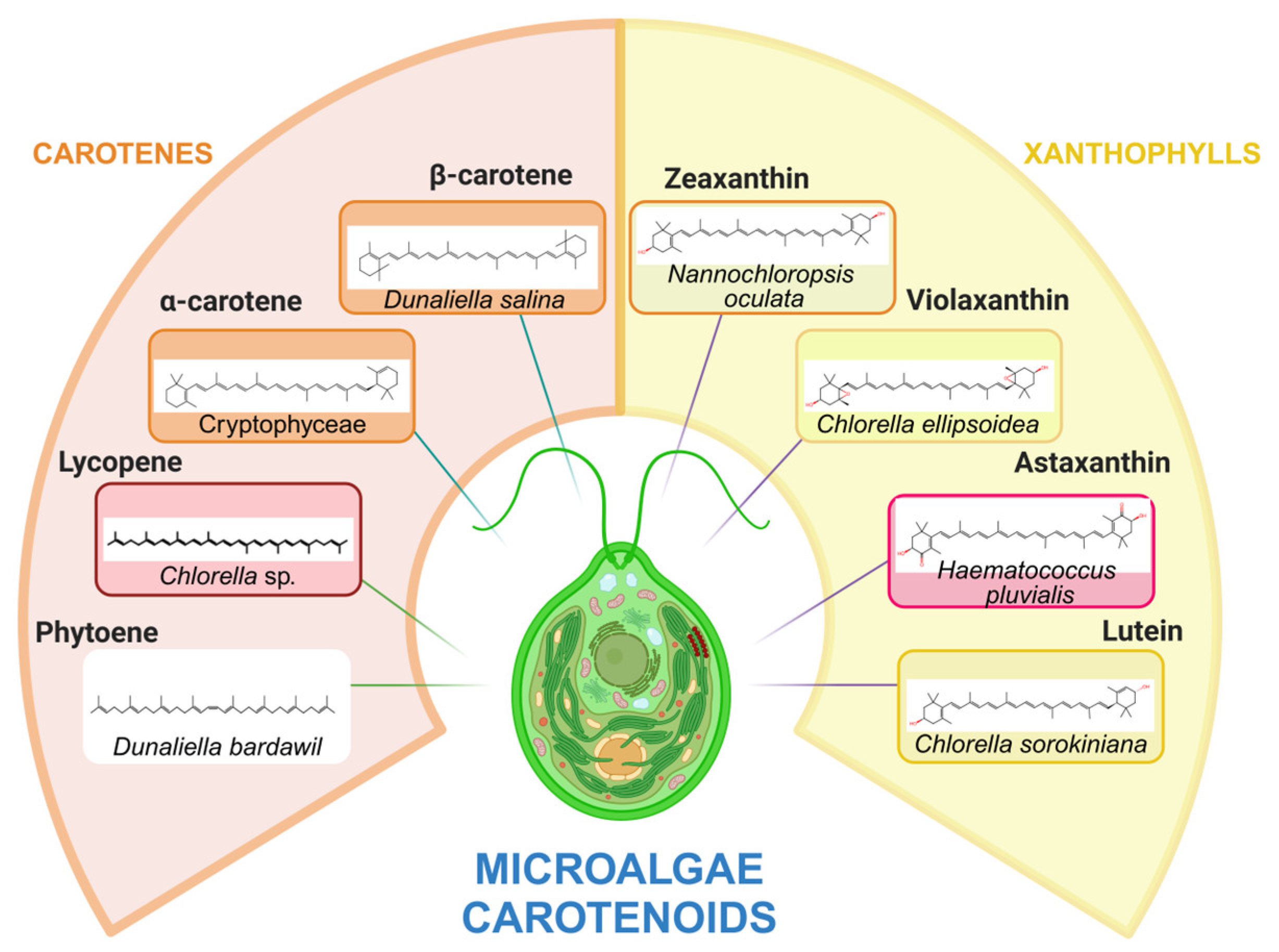

Table 1.
Main microalgae species harvesting by bio-flocculants.
| Microalgae | Bio-flocculant used | Recovery efficiency (%) | Reference |
| Chlorella vulgaris | Paenibacillus sp. AM49 | 90 | [74] |
| Chlorella sp. | Aspergillus terreus | 80-90 | [75] |
| Scenedesmus sp. | Aspergillus sp. | 80-90 | [76] |
| Dunaliella salina | Heterocapsa circularisquama | 80-90 | [77] |
| Nannochloropsis oculata | Tannin-based polymer | 99 | [78] |
| Nannochloropsis sp. | Nano-chitosan | 85 | [79] |
| Botryococcus braunii | Aspergillus sp. | 97 | [80] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



