
Submitted:
21 March 2025
Posted:
24 March 2025
You are already at the latest version
Abstract
Antiretroviral therapy (ART) has significantly improved the prognosis of human immunodeficiency virus type 1 (HIV-1) infection. Although ART can suppress plasma viraemia below detectable levels, it cannot eradicate the HIV-1 DNA (provirus) integrated into the host cell genome. This integration often results in unrepaired DNA damage due to HIV-induced inhibition of DNA repair pathways. Furthermore, HIV in-fection causes telomere attrition in host chromosomes, a critical factor contributing to CD4+ T-cell senescence and apoptosis. HIV proteins can induce DNA damage, block DNA replication, and activate DNA damage responses across various organs. In this review, we explore multiple aspects of the intricate interactions between HIV-1 and the host genome involved in CD4+ T-cell depletion, inflammaging, clonal expansion of infected cells in long-term treated patients, and viral latency. We discuss the molecular mechanisms of DNA damage that contribute to comorbidities in HIV-1-infected individuals and highlight emerging therapeutic strategies targeting the integrated HIV provirus.
Keywords:
HIV-1
; virus–host interaction
; viral DNA integration
; DNA damage
; latency
; LEDGF/p75
; capsid
; CPSF6
; HIV therapies
1. Introduction
Human immunodeficiency virus type 1 (HIV-1) is an RNA virus that belongs to the retrovirus family. Retroviruses are unique among virus families due to their ability to integrate their genetic material into the DNA of host cells. This integration occurs after the production of double-stranded linear viral DNA (vDNA) from viral RNA, a process known as reverse transcription, which is catalyzed by the viral enzyme reverse transcriptase (RT) [1,2,3]. The integration of linear vDNA marks a 'point of no return' for the infected cell, making it a permanent carrier of the viral genome, referred to as a provirus.
Retrovirus infections in mammals over millions of years have introduced proviral sequences into their genomes. These sequences have been transmitted through retroviral infections of the germline. Approximately 8% of the human genome consists of sequences derived from retroviruses, predominantly non-coding regulatory DNA often maintained in a transcriptionally silent state by the host cell's repressive DNA methylation machinery [4]. Mammals have co-opted some endogenous retroviruses for physiological functions; however, these sequences can occasionally have adverse health effects [5]. In contrast to human endogenous retroviruses, HIV-1 proviral integration or components of HIV-1, such as regulatory or accessory proteins, can interact with the human genome, leading to DNA damage or gene dysregulation. Integration can result in DNA breaks [6,7,8], which are often left unrepaired because HIV-1 can inhibit the host DNA repair machinery [9,10]. Proviral integration within or close to specific genes—such as oncogenes and KRAB-ZNF genes—can confer a proliferative advantage, resulting in the clonal expansion of latently infected cells [11,12]. Furthermore, active cellular replication to compensate for the loss of CD4+ T cells, HIV-1-mediated inhibition of DNA repair mechanisms, and the accessory HIV-1 Tat protein can contribute significantly to the shortening of telomeric host DNA, which is a key driver of CD4+ T cell senescence and apoptosis [13,14]. Finally, HIV-1-induced genetic damage in cells across various organs can contribute to the development of comorbidities, including pulmonary arterial hypertension, HIV-1-associated nephropathy, and neurological dysfunctions [15,16,17]. This review examines various aspects of the complex interactions between HIV-1 and the host genome. These interactions are critical in the pathogenesis of CD4+ T cell depletion, inflammation, viral latency, and the clonal expansion of latently infected cells in long-term treated patients. We also explore novel insights into the role of the viral capsid and its inhibitor, Lenacapavir. Finally, we discuss the molecular mechanisms of DNA damage that contribute to comorbidities in individuals living with HIV-1. We highlight emerging therapeutic strategies targeting the integrated HIV-1 provirus, such as the "Shock and Kill" and the "Block and Lock" approach, as well as CRISPR-based gene editing.
2. Integration and Post-Integration Stages in HIV-1 Infection
2.1. HIV-1 Provirus Integration
HIV-1 infection begins with the interaction of the viral envelope (Env) glycoproteins with the CD4 receptor and the CCR5/CXCR4 coreceptors expressed on the plasma membrane of the target immune cells. The Env glycoprotein Gp160 is a viral surface protein encoded by the HIV-1 env gene. This glycoprotein is cleaved by a host furin-like protease into the receptor-binding fragment gp120 and the fusion fragment gp41. The interaction between gp120 and the CD4 receptor results in a conformational change of gp120 that allows a secondary interaction of gp120 with the cellular coreceptor CCR5 or CXCR4 and the insertion of the N-terminal of gp41 into the cell membrane. Subsequently, gp41 undergoes a conformational change that brings the virus closer to the cell, allowing the viral membrane to fuse with the cell membrane and enter the viral capsid inside the cell [18]. The reverse transcription follows the capsid entry. The enzyme RT converts the single-stranded RNA genome into an integration-competent double-stranded DNA transported into the nucleus and integrated into the host DNA through the viral enzyme integrase [18].
Many concepts about the early stages of HIV-1 infection have changed based on recent studies that have demonstrated the importance of the capsid in regulating reverse transcription, nuclear entry, integration, viral assembly, and maturation [19]. Inside the capsid, the vDNA molecule exists in the form of a preintegration complex (PIC) constituted by the viral proteins Vpr, matrix proteins, integrase, host proteins (Barrier-to-autointegration factor 1), and the vDNA [20]. Matrix proteins and Vpr both contain nuclear localization signals (NLS), so previous studies indicated that PIC could play a role in nuclear import [21,22,23,24]. However, more recent studies have shown that the capsid is responsible for the nuclear import of PIC [25,26,27]. Capsid is transported through the cytoplasm towards the nucleus by the motor proteins kinesin and dynein. When the capsid arrives at the nuclear pore complex (a large protein complex of the nuclear pore), it is directed toward the central channel of the nuclear pore via interaction with Nup358 (nucleoporin 358). Subsequently, capsid interacts with other essential proteins: Nup153 and CPSF6 (Cleavage and polyadenylation specificity factor subunit 6). These proteins allow the translocation of capsid into the nucleoplasm, where reverse transcription is completed. CPSF6 not only functions in nuclear import but also directs the viral complex enclosed in the capsid to the nuclear speckles, where integration of the viral DNA into the host cell genome occurs [28].
2.2. Post-Integrational DNA Repair
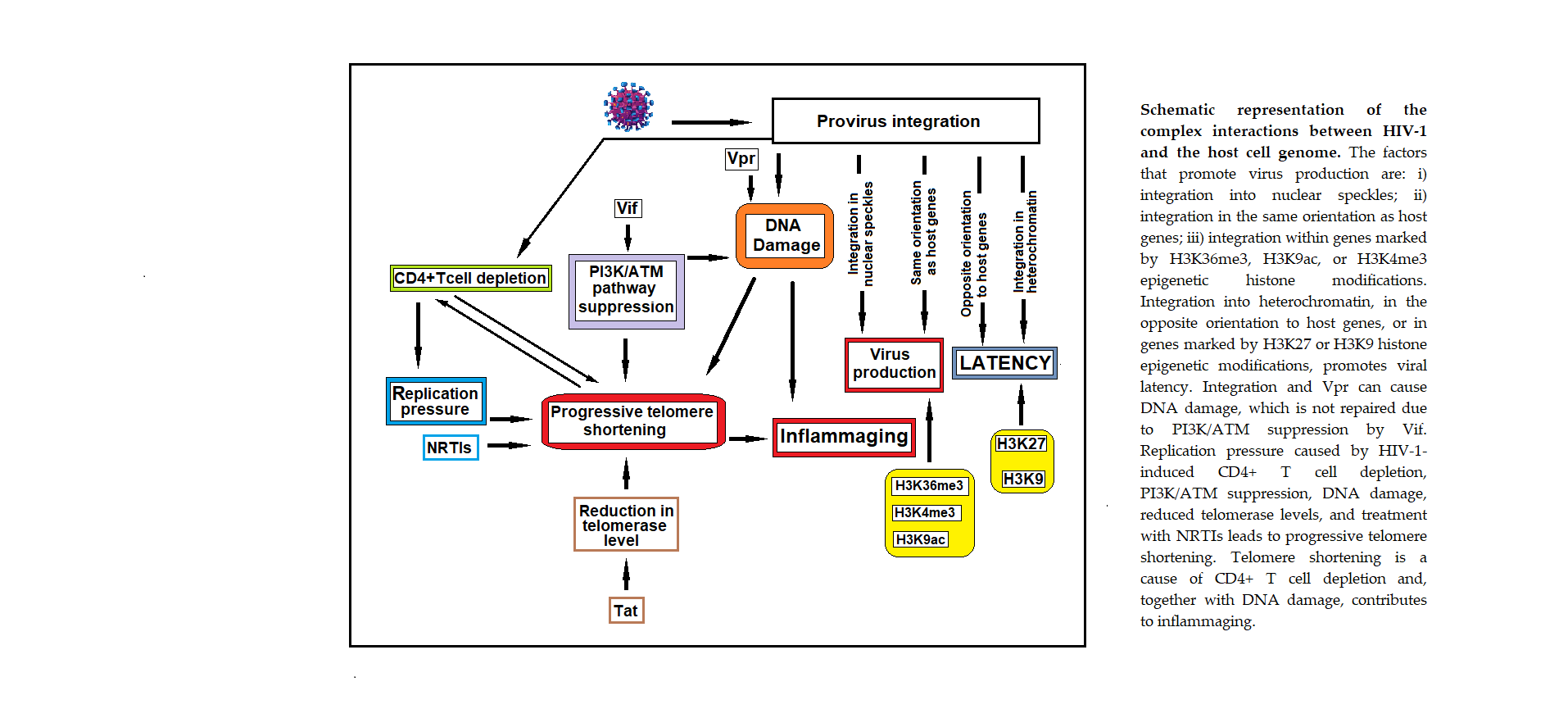
Integration of retroviral DNA by viral integrase creates a discontinuity in the chromatin of the host cell, and repair of this damage is necessary to complete the integration process. Before integration, viral integrase hydrolyses a phosphodiester bond at each vDNA end, excising a dinucleotide and exposing 3’-OH groups on cytosine-adenine dinucleotides. The 3’-vDNA ends are then inserted into the host genome, creating five-nucleotide gaps flanking the integrated vDNA [29]. The host DNA repair machinery subsequently fills these gaps and removes the remaining dinucleotides at the 5' ends of the integrated vDNA. [29,30,31,32]. Several studies have demonstrated the involvement of DNA-dependent protein kinase (DNA-PK) in HIV-1 post-integration DNA repair [30,33,34]. Given that DNA-PK primarily functions as a sensor in DNA double-strand break (DSB) repair, its role in HIV-1 post-integration repair was initially considered unexpected, as the HIV-1 integration intermediate does not contain DSBs. This involvement was clarified when the formation of the Ku70–integrase complex was observed during the DNA damage response (DDR) in HIV-1 post-integration repair [30,34]. Ku is a dimeric protein complex constituted by two polypeptides, Ku70 and Ku80, that binds to DNA DSB ends and is required for the non-homologous end joining (NHEJ) pathway of DNA repair [35]. In HIV-1-infected cells, the viral integrase recruits Ku70 at the site of integration. This polypeptide then attracts Ku80, DNA-PK, and ATM (ataxia-telangiectasia mutated). ATM is a serine/threonine protein kinase that phosphorylates CHK2 (checkpoint kinase 2) and H2AX, two key proteins that initiate activation of the DNA damage checkpoint (Figure 1). The serine/threonine protein kinase CHK2 inhibits CDC25 preventing the entry of the cell into the S and M phases. H2AX is a histone protein from the H2A family that contributes to nucleosome formation, chromatin remodeling, and DNA repair. The phosphorylated form of H2AX is a scaffold that attracts repair factors to DNA damage sites and retains them until the damage is removed. As we will report in the following sections, components of HIV-1 can inhibit ATM-directed DNA repair processes and CHK2, causing DNA repair failure and leading to dysregulation and apoptosis of infected cells.
2.3. HIV-1 Provirus Integration Sites
HIV-1 provirus integration is not entirely random but is influenced by a preference for more accessible areas of the genome, often related to a more open and active chromatin structure and transcriptional factors that facilitate viral replication (Figure 2) [36,37,38,39,40]. The number of integration sites varies depending on several factors, such as the class of patient (treated versus untreated, acute versus chronic infection, or untreated aviremic elite controller versus treated aviremic patients), the type of cells examined (activated or resting), the kind of treatment, and the experimental techniques used. No univocal data are available on the exact number of integration sites [41,42,43,44].
Integration has been observed in intronic and exonic regions of active genes. These regions are prevalently involved because they contain the necessary machinery for transcription and splicing, which HIV-1 may exploit for replication. HIV-1 provirus can also integrate into intergenic regions between genes that are generally not transcriptionally active and contain regulatory elements [45,46].
The actively transcribed genes in which HIV-1 preferentially integrates are often located in speckle-associated and speckle-proximal chromatin subnuclear domains (Figure 2). These are regions of chromatin organized near nuclear speckles, which are membrane-less nuclear structures enriched with RNA splicing factors and transcription-related proteins. Genes located into or near nuclear speckles are often transcribed and processed more efficiently, as the high concentrations of transcription and RNA processing factors are more readily accessible to these genes [47]. These regions of chromosomes usually show epigenetic modifications of histones that favor integration [48,49]. Histone epigenetic modifications observed in HIV-1 integration sites are: i) histone H3 lysine 4 trimethylation (H3K4me3), which marks actively transcribed gene promoters and enhancers, and the sites of RNA Polymerase II recruitment; ii) histone H3 lysine 9 acetylation (H3K9ac) and H3 lysine 27 acetylation (H3K27ac), found in active chromatin and enhancer regions, are histone epigenetic modifications associated with open euchromatin that facilitates HIV-1 DNA integration; iii) histone H3 lysine 36 trimethylation (H3K36me3), found in actively transcribed gene bodies, is an epigenetic modification of histone H3 which is a key protein in DNA packaging. SETD2 histone methyltransferase catalyses trimethylation of H3K36 in H3K36me3. In vitro integration assays have shown that trimethylation of lysine 36 of histone H3 significantly increases the integration of HIV-1 PIC into nucleosomal substrates, indicating that this epigenetic modification is significant for the integration of the virus [50,51]. However, SETD2 or H3K36me3 are not strictly required for HIV-1 infection; instead, they influence the location of integration sites and play a key role in regulating HIV-1 post-integration expression and latency [52]. In contrast, latent HIV-1 reservoirs often reside in regions that undergo epigenetic silencing, marked by H3K9me3 and H3K27me3 (repressive marks) [53].
Two host proteins, Pol II (RNA polymerase II) and LEDGF/p75 (Lens Epithelium-Derived Growth Factor), play key roles in integrating HIV-1 provirus into the human genome, directly contributing to the positions and efficiency of integration. Pol II plays an indirect but essential role in integrating the HIV provirus into the host genome. Pol II-associated transcriptional activity creates a favorable environment for integration by loosening chromatin. It generates transcription-associated chromatin marks (e.g., histone modifications like H3K36me3) that are recognized by the HIV integration machinery, specifically by the LEDGF/p75 [54]. LEDGF/p75 binds to HIV-1 integrase and directs the integrase complex to the integration sites, interacting with epigenetically modified histones, especially H3K36me3 [55,56,57,58,59,60].
Analysis of integration sites has shown that HIV-1 integration occurs prevalently in specific genes encoding proteins involved in cell proliferation and cancer (such as BACH2, MKL2, and HN1L) [61,62,63,64,65], in genes encoding proteins involved in chromatin modification (such as KANSL1) [61,66], and pre-mRNA splicing (such as PRPF6) [61,67]. Several authors have reported that the most frequent gene integration sites are the MKL2 and BACH2 [11,68,69,70,71]. In these genes, the HIV-1 provirus is often found in the same transcriptional orientation, a condition that can promote the expression of host genes. MKL2 and BACH2 are known human proto-oncogenes [72,73,74], and integration into these genes may be responsible for the clonal expansion of infected CD4+ T cells and, in some cases, the development of tumors. [75]. Other genes that are frequent sites of integration are ARIH2 (a cancer-related gene involved in acute myeloid leukemia and gastric cancer) [61,76,77], MKL1 (a gene related to MKL2), MROH1 (encoding large multiple HEAT-repeat-containing proteins) [61,78], and XPO6 (encoding an importin-beta protein that mediates nuclear export of profilin-actin complexes) [61,79].
In contrast to actively infected cells, integration sites of provirus in resting cells are often found in suboptimal regions for proviral gene expression, which seems to promote the latency of HIV-1 [44,68,80,81,82]. Latency is the major obstacle to HIV-1 infection cure. If ART treatment is interrupted, the virus starts replicating from reservoirs of latently infected cells, so ART must be taken for life to suppress viral replication continually. Sites of integration in patients treated over time with ART or in elite controller (individuals with natural control of HIV-1) have been found in heterochromatin, non-genic regions, and an opposite orientation to the host gene, suggesting that a selection of intact proviruses with viral latency characteristics occurs during prolonged antiretroviral therapy or in subjects able to control viremia without ART [83,84,85]. Latency is favored by specific epigenetic modifications of histones, including histone methylation and acetylation. Histone 3 methylation of lysine 27 (H3K27) and histone 3 methylation of lysine 9 (H3K9) are frequently observed in resting cells [86]. These modifications alter chromatin accessibility and have an inhibitory effect on the transcriptional activity of the integrated provirus [86,87].
Histone modifications play a key role in latency; however, several studies suggest that latency primarily depends on a combination of factors. In addition to epigenetic modifications of histones, these include low levels of key activating transcription factors, provirus orientation, and integration into specific genes, such as KRAB-ZNF genes. [53,88,89,90].
2.4. HIV-1 Provirus Orientation
When the provirus integrates into a host gene, its orientation concerning the transcriptional mechanism of the host gene can influence its expression and viral latency (Figure 2). This effect is primarily due to transcriptional interaction and interference between host gene transcription and the integrated provirus. Researchers inserted HIV-1 genomes into the intron of an active host gene in both orientations to assess the impact of host gene readthrough transcription. They found that when the provirus was in the same orientation as the host gene, readthrough transcription enhanced HIV-1 gene expression. Conversely, in the opposite (convergent) orientation, readthrough transcription inhibited HIV-1 expression. This orientation had a more than tenfold effect on HIV-1 gene expression [91].
An analysis of HIV-1 integration sites in primary CD4+ T cells revealed that the orientation of the provirus relative to the host gene's transcriptional direction was a critical determinant of latency. Proviruses integrated in the same orientation as host genes were transcriptionally active, while those in the opposite orientation tended toward latency [92]. The orientation of provirus can modulate its expression even when integrated in proximity to active transcriptional units. Einkauf et al. conducted a detailed analysis of the chromosomal positions of many intact and defective proviral sequences in individuals on long-term antiretroviral therapy. The results of their study indicate that prolonged antiretroviral treatment is associated with an accumulation of intact proviruses with deeper latency characteristics, probably caused by immune-mediated selection pressure. They observed that the insertion sites were near active transcriptional units and accessible host chromatin sites and that the intact viral sequences were located in opposite orientations to host genes, supporting the assumption that transcriptional interference between host and proviral gene expression may be a predominant mechanism for maintaining HIV-1 latency, and explaining why HIV-1 can remain transcriptionally silent despite integration into actively transcribed and typically highly expressed host genes [83]. The role of ART in selecting cells with latency characteristics and specific orientation of provirus was studied by Kok et al., who examined the influence of the activation state of CD4+ T cells, the effect of antiretroviral therapy, and the clinical stage of HIV-1 infection on HIV-1 integration site features and selection. They observed a preference for a convergent orientation of the provirus compared to the same orientation relative to the host gene in all HIV-1 integration sites. This was more evident in patients treated with ART compared with untreated patients. Although provirus orientation is probably not the only factor determining HIV-1 latency, understanding these events that occur over time during ART could be essential for developing therapies that aim to reverse latency or eliminate reservoirs of HIV-1.
3. HIV-1-Induced Host Genome Damage
3.1. HIV-1-Induced DNA Breaks
The HIV-1 genome contains nine genes that encode fifteen viral proteins [93]. The mRNAs for viral structural proteins are transcribed by gag, pol, and env viral genes. In addition, HIV-1 encodes for proteins with essential regulatory functions (Tat and Rev) and for proteins with accessory functions, such as Nef, Vpr, Vif, and Vpu. Accessory HIV-1 proteins can interfere with post-integrational repair and induce further strand breaks in the DNA of infected cells. Viral protein R (Vpr) is expressed in five primate lentiviral groups (HIV-1, HIV-2, and SIVs from mandrill, Sykes, and African green monkeys) [94]. It is a multifunctional protein that plays several functions in the viral life cycle, such as increasing the rate of viral replication and accelerating the cytopathic effect of the virus in T cells and macrophages [95]. Several reports indicate that Vpr induces DDR, suggesting that this viral protein may itself cause damage to host DNA [6,7,8,9]. However, the mechanism by which Vpr induces structural alteration of DNA remains unclear. Several mechanisms of Vpr-induced DNA damage and DDR activation have been proposed. Experiments conducted by Li et al. showed that Vpr directly induces DNA damage, blocks DNA replication, and activates ATR (ataxia telangiectasia and Rad3-related protein) [9]. Iijima et al. [8] demonstrated that Vpr unwinds double-stranded DNA by converting it into a relaxed form and induces the ubiquitination of histone H2B. This chromatin remodeling promotes efficient loading of RPA70 (replication protein A), a single-stranded DNA-binding subunit of RPA that activates the ATR-dependent DDR [96,97,98]. In addition, Vpr-induced unwinding of double-stranded DNA resulted in the accumulation of negatively supercoiled DNA and covalent complexes of topoisomerase 1 and DNA, which cause DSBs (Figure 3) [7]. While topoisomerase 1 is responsible for causing single-strand breaks, under certain conditions—such as collisions with replication machinery—these single-strand breaks can be converted into double-strand breaks, which are much more dangerous and can lead to genomic instability. Another mechanism by which Vpr can induce the formation of DSBs in DNA is through the production of ROS, which leads to oxidative stress. This oxidative damage also interferes with cellular repair mechanisms [7]. By causing replication fork stalling, Vpr indirectly leads to the collapse of these forks, a phenomenon associated with the formation of DSBs. This is often compounded by the inability of the cell to resolve stalled forks due to Vpr-mediated inhibition of repair proteins [99]. Vpr-induced DNA damage and activation of DDR signaling can alter cellular transcription through modulation of NF-kB. This primary transcription factor family regulates innate and adaptive immune response and inflammation genes. Following DSBs induced by Vpr, DNA damage activates ATM, which results in NEMO activation and IKK-mediated degradation of IkB. This causes the translocation of the NF-kB class II type RelA into the nucleus. RelA binds to genes containing NF-kB sites, promoting their transcription. Since the HIV-1 genome contains multiple NF-κB binding sites in its promoter regions, an advantage of Vpr's activation of NF-κB signaling via ATM is the potential direct enhancement of HIV-1 gene expression. Furthermore, the increased activation of NF-kB leads to the production of pro-inflammatory factors such as IFNα, IFNλ1, IRF1, IRF7, TNFα, IL-6, and IL-8, which play a key role in determining the state of chronic inflammation in HIV-1-infected individuals [100].
Of interest is that Vpr promotes the degradation of several DDR proteins, inhibiting double-strand DNA break repair by recruiting the CRL4ADCAF1 ubiquitin ligase complex [9]. CRL4ADCAF1 (also known as the CRL4 E3 ubiquitin ligase complex) is a specific variant of the CRL4 (Cullin-RING Ligase 4) complex, a type of E3 ubiquitin ligase. These complexes tag proteins with ubiquitin, signaling their degradation by the proteasome. For example, Vpr-expressing cells cannot repair DSBs induced by etoposide or other DSB-inducing compounds [9]. Li et al. observed that cells transfected with HIV-1 and HIV-2 Vpr decreased DNA repair by homologous recombination by 66% and 49%, respectively [9]. Homologous recombination repairs DNA before the cell enters mitosis (M phase), during the end of the S and G2 phases [101]. Cells often utilize non-homologous end joining (NHEJ) to repair DSBs when homologous recombination is repressed [102]. Likewise, to homologous recombination, HIV-1 Vpr expression also decreased NHEJ efficiency by 51%. Differently from HIV-1, HIV-2 Vpr did not significantly decrease NHEJ, indicating functional differences between HIV-1 and HIV-2 Vpr [9]. DDR is also inhibited by the HIV-1 accessory protein Vif (virion infectivity factor) [10]. Vif is an HIV-1 accessory protein required to counteract the antiviral activity of host APOBEC3 DNA cytosine deaminases. APOBEC3 (apolipoprotein B mRNA editing enzyme, catalytic subunit 3G) is a family of proteins that can bind to nascent HIV-1 particles and catalyze cytidine deamination to uridine in the single-stranded DNA substrate. This results in nonsense mutations, missense mutations, and abortive integration. The interaction of APOBEC3G with Vif results in the recruitment of APOBEC3 into a complex constituted by ubiquitin ligase E3, scaffold protein cullin5 (CUL5) and the substrate adaptors elongin B (ELOB) and elongin C (ELOC). This promotes polyubiquitination and degradation of APOBEC3 and inhibits the APOBEC3-mediated antiviral defense [103]. In addition, Vif antagonizes PP2A (protein phosphatase 2A) activity [10]. PP2A is an intracellular protein phosphatase that regulates cell growth and metabolism, DNA replication, DNA damage checkpoints, and DNA repair processes. PP2A governs the activation of kinases involved in DDR, such as ATM, ATR, DNA-PK, CHK1, and CHK2 [104,105,106,107]. PP2A antagonism mediated by Vif causes the accumulation of DSBs in the host DNA through the inactivation of kinases involved in DDR. However, pharmacologic and functional studies suggest that Vif blocks Vpr-directed ATM activation but not ATR [10]. In addition, Vif blocks ATM-directed induction of NF-κB signaling and phosphorylation [103]. The inhibition of cellular DNA repair mechanisms has essential implications in CD4+ T-cell depletion, inflammaging, and several comorbidities observed in HIV-1-infected patients (Figure 2).
3.2. DNA Damage and CD4+ T Cell Depletion
HIV-1 causes CD4+ T cell depletion not only by direct cytopathic effect (which generally occurs in the acute phase of infection) but also by direct and indirect mechanisms that damage or alter DNA (Figure 2). Being long-lived, CD4+ T cells are exposed for a long time to genomic insults that may affect different areas of DNA. As previously described, HIV-1 can inhibit several DNA damage repair mechanisms so that infected CD4+ T cells have a limited ability to repair genomic insults. This condition can induce premature senescence and apoptosis of infected cells [108]. Furthermore, in HIV-1 infection, the replicative pressure to replace dead CD4+ T cells causes progressive telomere shortening (a phenomenon defined as telomere attrition), which is a hallmark of cellular senescence [109,110].
Telomeres are specialized structures at the ends of linear chromosomes in eukaryotic cells. They consist of repetitive DNA sequences (in humans, the sequence is typically TTAGGG) and associated proteins. During DNA replication, the enzyme DNA polymerase cannot fully replicate the ends of linear DNA, leading to the progressive shortening of chromosomes. Telomeres buffer this loss by providing expendable DNA that can be shortened without affecting essential genes [111]. When they become critically short, cells enter a state called senescence (a non-dividing state) or undergo apoptosis, contributing to aging.
T lymphocytes can undergo a finite number of replications before entering a state of senescence and cell death. This limit to cellular replication depends on the erosion of telomeres that occurs after each cell division. Upon activation, T cells can proliferate by activating the enzyme telomerase, which can add telomeres to the ends of chromosomes, thus preventing the development of replicative senescence [112]. However, after repeated stimulation, highly differentiated T cells lose the ability to induce this enzyme [113]. To understand the role of telomeres in HIV-1 infection, Zhao et al. studied the homeostasis of T cells, telomeric DNA repair, and DNA damage mechanisms in HIV-1-infected individuals on ART with undetectable viremia. They observed that CD4+ T cells of these patients were susceptible to DNA damage that extended to the ends of chromosomes, leading to accelerated telomere erosion, mainly due to a suppression of the expression and activity of ATM and its downstream CHK2. In contrast, the DNA damage sensor MRN complex remained intact. Interestingly, ectopic ATM expression by transfection of HIV-1-derived CD4+ T cells with a Flag-His-ATM construct reduced DNA damage, apoptosis, and cellular dysfunction [13]. Considering that telomere integrity is crucial for cell survival, Khanal et al. conducted a series of in vitro studies to determine whether HIV-1-induced T-cell apoptosis is related to telomere damage [14]. They determined the sites of DNA damage by confocal microscopy by assessing the presence of 53BP1-induced nuclear foci. This analysis is based on the observation that following a genotoxic insult, 53BP1 functions as a docking site for other DNA repair factors to form nuclear foci visible under the microscope [113]. Dysfunctional telomere-induced 53BP1 was higher in primary CD4+ T cells after HIV-1 infection than in uninfected CD4+ T cells. Infected cells showed a marked shortening of telomeres compared to uninfected cells. Of note, the protease inhibitor raltegravir partially blocked or restored telomere lengths. Finally, they observed the PI3K/ATM pathway suppression in infected cells correlated with telomeric DNA damage and premature CD4+ T cell aging and apoptosis. Based on these findings, the authors suggested a model of CD4+ T-cell depletion in HIV-1 infection in which suppression of PI3K/ATM promotes telomeric DNA damage and erosion. This results in premature cell aging and apoptosis. ART cannot fully restore the PI3K/ATM pathway in HIV-1 infected cells, so infected cells would live in a state of high apoptotic susceptibility. In a univariate analysis, Alejos et al. observed that a shorter telomere length was associated with older age, HIV-1 RNA ≥ 100 000 copies/mL, CD4 count < 200 cells/μL, lower CD4:CD8 ratio, statin treatment, and current alcohol consumption [114]. In another study, higher levels of immune activation increased soluble CD14 levels, and a high percentage of CD38(+) HLA-DR(+) cells among CD4+ T cells correlated with shorter telomeres [115].
Among telomere shortening causes, accessory HIV-1 Tat protein plays a relevant role [116]. Tat protein can decrease the telomerase level in infected cells, and this reduction occurs in uninfected T cells when exposed to Tat protein (Figure 2) [116]. The inhibition induced by the Tat protein appears to be related to the ability of this protein to inhibit nuclear levels of hTERT (human telomerase reverse transcriptase), which is the catalytic subunit of telomerase with reverse transcriptase activity [116]. In addition, this protein alters the AKT pathway and the molecular interaction with chaperones required for hTERT phosphorylation, nuclear import, and activation [117]. ART initiated in the early stages of HIV-1 infection can slow telomere shortening in CD4+ T cells [117]. In contrast, ART administered at a late stage of infection is associated with significant and sustained telomere length shortening [118]. In HIV-1-infected adults with prolonged virological suppression, NRTIs-containing ART was associated with smaller gains in blood telomere length after 2 years of follow-up [119]. Tenofovir (tenofovir disoproxil fumarate and tenofovir alafenamide fumarate) and abacavir are potent inhibitors of human telomerase in vitro, likely due to homology between HIV-1 RT and hTERT [118,119,120,121,122]. In cross-sectional studies of patients treated with ART, longer exposure to tenofovir was associated with inhibition of telomerase activity and significantly shorter telomere length in CD4+ T cells [123]. A longitudinal study of long-term virologically suppressed patients showed that treatment with tenofovir or abacavir was associated with a smaller gain in telomere length than regimens with a sparing of NRTIs [119]. Furthermore, an increase in blood telomere length was observed in HIV-1-infected patients who switched to a therapeutic simplification with dolutegravir plus lamivudine compared to patients who continued on a triple regimen containing NRTIs [124]. Finally, a work on European children with HIV-1 acquired in the perinatal period and treated early showed that the size of the HIV-1 reservoir is associated with telomere shortening and immunosenescence [125].
3.3. DNA Damage and Its Role in Inflammaging in HIV-1 Infected Individuals
Inflammaging (a term that combines "inflammation" and "aging") is a chronic, low-grade inflammation that develops with age. Inflammaging of the immune system is a typical event in HIV-1 infection [126]. The relationship between aging and inflammation is complex and bidirectional [127]. A chronic, low-grade inflammatory state develops as individuals age. This persistent inflammation contributes to aging and the onset of age-related diseases [127]. Many of the alterations affecting the immune system in HIV-1-infected individuals resemble the progression of inflammation in the elderly [126]. HIV-1-mediated inflammation could result from different insults, such as incomplete viral proteins or particles released from the reservoirs, pro-inflammatory cytokines secreted by the cells, HIV-1-increased intestinal permeability, altered gut microbiota, and co-infections [128]. The accumulation of unrepaired DNA damage due to DDR inefficiency, the replicative pressure to compensate for CD4+ T cell depletion, and telomere attrition lead to T cell exhaustion and senescence, similar to that observed in the elderly [127]. Furthermore, it has been observed that cells with persistent DDRs may trigger the secretion of inflammatory cytokines associated with senescence that may induce a senescent state in neighboring uninfected cells [128]. Although cART effectively controls viral replication, residual inflammaging may persist even in individuals with normal CD4+ T cell counts (but often with an altered T cell CD4+/T cell CD8+ ratio) [129]. Such inflammaging is characterized by short telomeres leading to CD4+ T-cell depletion and senescence, similar to those observed in the elderly [110,130].
4. HIV-1 Integration and CD4+ T Cell Clonal Expansion
Although at the beginning of an HIV-1 infection, the viruses have identical genomes, these can diversify over time due to the appearance of spontaneous mutations [71,131]. However, clusters of identical viral sequences originating from clonally expanded infected cells are often observed in patients undergoing long-term antiretroviral therapy [132,133]. In patients treated with an efficient ART, plasma viremia can be reduced by more than ten thousand times, while the level of intracellular viral DNA decreases much less, on average, by only fifteen times [134]. This phenomenon has been related to a clonal expansion of infected cells hosting latently integrated provirus [134]. Upon suspension of ART, there is a rapid increase in plasma viremia. This indicates the presence of some cells in the latent reservoir capable of re-expanding and activating HIV-1 transcription [135,136,137]. Evidence that the provirus often integrates into a cancer-associated gene has suggested that this type of integration potentially can confer a proliferative advantage to clonal populations of infected cells [11]. The viral promoter LTR could grant the proliferative advantage, activating oncogenic gene expression [11]. However, other authors have demonstrated that integrating HIV-1 in cancer-related genes is a minor factor in the clonal expansion of infected CD4+ T cells [138]. Furthermore, HIV-1 proviruses integrated into or close to cancer-related genes are often defective and unable to determine the increase in viremia upon interruption of ART [138]. There is another category of genes, the KRAB-ZNF genes (KRAB = Krüppel associated box; ZNF = Zinc finger), where the insertion of HIV-1 proviruses is observed, especially in patients treated with long-term ART [12,83,84,85,139,140,141]. KRAB-ZNF proteins produced by these genes are the most abundant family of epigenetic repressors found only in tetrapod vertebrates. Upon binding to DNA, KRAB-ZNF proteins trigger transcriptional repression via interaction with TRIM28, which acts as a scaffold for heterochromatin protein 1 (HP1), H3K9me3-specific histone methyltransferase 1 (SETDB1), and the NuRD histone deacetylase-containing complex. Together, these proteins silence transcription by triggering H3K9 trimethylation and heterochromatin formation [142,143]. Huang et al. analyzed integration sites of near full-length HIV-1 genomes from individuals on long-term ART. They observed that, in clonally expanded cells of these individuals, proviruses were preferentially integrated within KRAB-ZNF genes. Unlike oncogenic genes, KRAB-ZNF genes do not promote cell proliferation, so the clonal expansion of CD4+ T-cells containing provirus integrations in KRAB-ZNF genes can be explained by the lack of production of the virus that is latently integrated, and thus the lack of cytolytic effect. KRAB-ZNF genes are associated with heterochromatin in memory CD4+ T cells, so they are under-expressed. [85]. Other studies indicate that HIV-1 proviruses integrated into KRAB-ZNF genes are intact and not defective and are only partially refractory to induction, suggesting that these genes might represent the region of host DNA where the provirus can integrate latently but can also be reactivated, for example, by ART withdrawal [12,144].
5. Capsid and Its Role in Driving HIV-1 Integration into Gene-Dense Regions or in Lamina-Associated Domains (LADs)
Although HIV-1 integration is primarily mediated by the interaction between integrase and LEDGF/p75 [55], recent studies suggest that capsid interactions with host factors CPSF6, Nup358 and Nup153 are critical for the selective targeting of integration sites [46,47,145,146]. Experimental depletion of these host factors reduces HIV-1 integration in gene-dense regions [147,148,149]. The primary capsid-binding host factor involved in HIV-1 integration targeting is CPSF6, and CPSF6 knockdown dramatically reduces the targeting of HIV-1 integration in gene-dense regions [39,150]. Tandem knockout experiments of CPSF6 and LEDGF/p75 have shown that CPSF6 plays a more dominant role than LEDGF/p75 in directing HIV-1 integration into gene-dense regions [39]. Furthermore, knockout or knockdown of CPSF6 significantly increases the preference for HIV-1 provirus integration into LADs [150]. LADs are regions of the genome that physically interact with the nuclear lamina. They are characterised by a repressive chromatin environment, low gene density, and limited transcriptional activity. LADs are rich in di- and trimethylated histone H3 lysine 9 (H3K9me2 and H3K9me3)—epigenetic modifications of the DNA-packaging protein histone H3 that promote heterochromatin formation. As a result, genes within LADs are prevalently repressed [151]. LADs are also enriched in GAGA repeats, which serve as binding sites for KRAB-ZNF proteins [152].
Interactions between the HIV-1 capsid and the host factors CPSF6 and Nup153 can be inhibited by capsid inhibitors, an emerging class of anti-HIV-1 drugs [153]. Since CPSF6 regulates integration site selectivity, treatment of HIV-1 infected cells with capsid inhibitors can influence the sites of HIV-1 integration. Bester et al. demonstrated that the capsid inhibitor GS-6207 (Lenacapavir) reduces HIV-1 integration in gene-dense regions while conversely enhancing integration within lamina-associated domains (LADs) [153]. This pattern parallels the effects observed following CPSF6 knockout or knockdown (Figure 4) [150].
A possible explanation for this phenomenon may be inferred from the work of Emerson et al., which investigated the Gmr1-like family of Gypsy/Ty3-like retrotransposons in the ancestor of amniotes. [154]. Many C2H2–type ZFN proteins have a SCAN (SRE-ZBP, CtBP-Associated, and Novel Zinc finger) domain. This is a conserved motif of approximately 80 amino acids found at the N termini of ZFN proteins, involved in protein-protein interactions and capable of dimerization, leading to the creation of homo- and heterodimer protein complexes [155]. SCAN domains are often found alongside KRAB domains [156]. Evidence indicates that the SCAN domain was derived from the C-terminal portion of the gag capsid protein from the Gmr1-like family of Gypsy/Ty3-like retrotransposons in the ancestor of amniotes and was exploited or adaptively coopted for a novel function, by C2H2-type ZFN genes [154]. SCAN domain mediates capsid protein multimerization to form the core retroviral and retrotransposon capsid structure. In addition, protein structural similarity between the SCAN domain and the HIV C-terminal capsid has been observed [154]. There is evidence to suggest that the SCAN domain originally functioned to target ZFN proteins to retroelement capsids [154]. This interaction could influence viral assembly, replication, or restriction. A possible parallel is TRIM5α, a host protein that binds retroviral capsids and prevents infection; although TRIM5α utilises a RING and SPRY domain, a similar mechanism could apply to ZNF-SCAN proteins [157]. The potential interaction between capsid and ZFN proteins suggests that capsids unbound to CPSF6 and Nup153 may preferentially interact with ZNF proteins, which are more prevalent in LADs than in chromatin rich in active genes. This interaction may facilitate proviral integration into LADs [152]. In contrast, when the capsid is bound to host cell factors CPSF6 and Nup 153, the HIV-1 pre-integration complex is directed towards gene-dense regions. Although ZNF proteins with SCAN domains could potentially interact with retroviral capsid elements, direct experimental evidence is limited [154], and this remains an area of potential research in host-virus interactions.
Several outcomes are possible if a retrovirus inserts its DNA into a LAD. Because LADs are less transcriptionally active, an integrated provirus in these regions is more likely to be transcriptionally silent. As a result, the repressive chromatin environment of LADs can help maintain the provirus in a silent state. Insertional mutagenesis considerations can also be made. Retroviral integration in gene-rich and actively transcribed regions can sometimes activate proto-oncogenes or disrupt tumor suppressor genes. By contrast, LADs tend to be less gene-dense. Therefore, integration into LADs might be less likely to cause such overt pathogenic effects. Another question arises: can the proviruses integrated into the LAD form a latent reservoir capable of reactivating upon ART withdrawal? Long-term ART-treated individuals are characterized by large clones of intact proviruses preferentially integrated into heterochromatic regions [141]. Notably, in these individuals integration is enriched in KRAB-ZNF genes, which appear to play a key role in maintaining viral latency and mediating reactivation upon ART interruption [12,144]. Dragoni et al. reported that cells with proviruses integrated into ZNF genes (ZNF470 and ZNF721), in locations previously associated with deeper latency, proliferated extensively and produced virus upon stimulation with cognate Gag peptides [12]. Therefore, provirus integrated in ZNF genes represents a latent reservoir that can be reactivated. LADs are rich in di- and trimethylated histone 9 lysine and are enriched in GAGA repeats constituting binding sites for ZNF proteins, two conditions markedly repressing gene expression. Currently, no data indicate whether the reactivation of proviruses integrated into LADs is possible. However, it is important to note that LADs are infrequent targets for HIV-1 integration [141]. In contrast, in elite controllers, a significant enrichment of intact proviral sequences in LADs has been observed when clonal intact provirus sequences were counted as independent proviruses [158]. On the basis of these data, we propose a theoretical model in which long-term treatment with capsid inhibitors—over several years or even decades—could lead to the clonal expansion of CD4+ T cells harbouring proviruses integrated into LADs and silenced therein (Figure 5). Although this model remains entirely hypothetical at present, it could imply a greater resistance to viral rebound upon discontinuation of capsid inhibitor therapy. Testing this hypothesis would naturally require many years of treatment with this class of anti-HIV-1 drugs.
6. Comorbidities Caused by HIV-1-Induced DNA Damage
6.1. Cancer and Provirus Integration
Integration of HIV-1 provirus can alter the host genome, potentially contributing to tumor development. Several retroviruses can induce tumors in animals by inserting in or near oncogenes [159]. Retroviral provirus integration can cause DNA breaks or genomic rearrangements, increasing the risk of oncogenic mutations. If the provirus integrates near an oncogene, it can activate it through transcriptional promotion. Moreover, integration can disrupt the function of tumor suppressor genes, facilitating malignant transformation [160]. People with HIV-1 have a higher risk of developing certain types of cancers, known as AIDS-defining cancers (ADC) and non-AIDS-defining cancers (NADC) [161]. The pathogenesis of these malignancies is related to the HIV-1–induced immunodeficiency, which allows specific viruses that cause human cancers, such as Epstein-Barr virus, Kaposi sarcoma herpesvirus, and human papillomavirus, to replicate.
It is well-known that HIV-1 provirus frequently inserts near or into oncogenes. However, such insertions generally do not lead to neoplastic transformation but they could support clonal expansion of CD4+ T cells [11]. Moreover, if neoplastic transformation were induced by the insertion of the HIV-1 provirus into or near oncogenic genes, it would be expected to result in T-cell neoplasms. Although lymphomas in people affected by HIV-1 infection are mostly B-cell type, an increased risk of developing T-cell lymphomas has been reported [162]. Increased risk was observed for all subtypes of T-cell lymphoma, including mycosis fungoides, peripheral lymphomas, cutaneous lymphomas, and adult T-cell leukemia/lymphoma (ATLL) [162]. The first case of AIDS-associated non-Hodgkin's lymphoma in which HIV-1 infection may have played a central role in the lymphocyte transformation process was described by Herndier et al. in 1992. They observed monoclonal-integrated HIV-1 within the genome of T-cell lymphoma, and these cells produced the HIV-1 p24 antigen [163]. In 1994, Shiramizu et al. described four lymphomas in HIV-1 infected patients constituted by large immunoblastic cells. Two were T-cell lymphomas, one was B-cell lymphomas, and another was described as a Ki-1 positive lymphoma (CD3O) admixed with CD14/ CD68-reactive macrophages/histiocytes. HIV-1 p24 was detected in cells phenotypically similar to macrophages in cases 1, 3, and 4 and detected in T-cells in case 2. Monoclonal HIV-1 integration sites were observed in all four tumors. The provirus integration site was within the fur gene, upstream from the c-fes/fps protooncogene [164]. A rare case of B cell lymphoma in which a defective HIV-1 was integrated upstream of the first STAT3 coding exon was described in 2007. The provirus had only a 3' LTR sequence with more vigorous promoter activity than the STAT3 promoter, and lymphoma cells showed high levels of STAT3, suggesting that up-regulation of STAT3 caused by HIV-1 integration can induce B cell lymphoma [165]. STAT3 is implicated in B cell proliferation and terminal differentiation but can also play an essential role in numerous human malignancies [166]. The few cases of T-cell lymphomas described in the literature, in which the insertion of the HIV-1 provirus has played a role in their pathogenesis, may be attributed to the requirement for additional non-viral mutations to induce human T-cell lymphomas. This is likely the primary reason why proviral insertion in or near an oncogenic gene leads to the clonal expansion of non-neoplastic cells and only very rarely results in a T-cell lymphoma [167]. Some evidence suggests that, in certain individuals with HIV-1 infection, proviral activation of STAT3—and in some cases, both STAT3 and LCK—may compensate for one or more steps required for the development of T-cell lymphomas. HIV-1-infected individuals with proviral activation of these two genes may subsequently acquire additional relevant mutations, potentially leading to the emergence of T-cell lymphomas. [167].
6.2. Pulmonary Arterial Hypertension
Cardiopulmonary diseases such as pulmonary arterial hypertension have increased significantly in individuals who are HIV-infected despite the introduction of ART. Currently, the prevalence of pulmonary arterial hypertension in people with HIV-1 is 2000 times higher than in the general population [15]. HIV-1- pulmonary arterial hypertension is characterized by endothelial hyperplasia and hypertrophy of medial smooth muscle cells [168]. Cells of these tissues lack specific receptors and coreceptors for HIV-1, suggesting that indirect mechanisms allow HIV-1 to induce damage to pulmonary arterial vascular structures. Several reports indicate that two proteins of the virus, Tat and Nef, may play a key role in the pathogenesis of pulmonary hypertension by inhibiting apoptosis induced by components of the DNA repair machinery [15]. Tat is a regulatory protein that enhances the efficiency of viral transcription [169]. It can be secreted from infected cells into the extracellular environment and internalized by uninfected bystander cells [170]. After internalization into the cytoplasm, Tat moves to the nucleus, modulating cellular transcriptional and post-transcriptional processes. Tat interacts with the Tip60 protein, which is involved in DNA damage repair, by inhibiting its activity and facilitating proteasomal degradation [171,172,173]. Suppression of Tip60 blocks the activation of ATM kinase activity and prevents ATM-dependent phosphorylation of p53 and CHK2. The inhibition and degradation of Tip60 inhibits ATM activity, resulting in the non-repair of DNA and inhibition of p53-induced apoptosis [174]. Nef is an HIV-1 accessory protein that allows immune evasion and the establishment of viral persistence [175]. Nef can be transferred from infected CD4+ T cells into coronary endothelial cells and detected in the endothelial layer of vascular lesions associated with the development of pulmonary arterial hypertension [176]. Nef can activate AKT through a PI3K-dependent mechanism, which induces anti-apoptotic and pro-proliferative signals through Bad phosphorylation/activation [177].
6.3. DNA Damage in Renal Tubule Epithelial Cells
HIV-1-associated nephropathy is characterized by focal glomerulosclerosis that typically occurs in the advanced stages of HIV-1 infection, especially in persons of African ancestry [178]. HIV-1-associated nephropathy is characterized by severe proteinuria and rapid progression to end-stage renal disease. In patients treated with ART, progression to severe renal disease is much slower. HIV-1 can infect renal tubular epithelial cells, podocytes, and parietal epithelial cells, causing cell cycle dysregulation, inflammation, and cell death [179]. Because renal epithelial cells do not express HIV-1 receptors and coreceptors [180], the mechanisms by which renal cells are infected are likely different from those by which HIV-1 infects CD4+ T cells. Ray et al. observed that HIV-1-infected mononuclear cells could transfer the virus to renal tubular epithelial cells by direct cell-to-cell transfer [181]. Other authors have shown that this transfer was partly mediated by heparan sulfate proteoglycans [182]. It was also observed that cultured podocytes from children with HIV-1-associated nephropathy sustained low-level productive infection when exposed to cell-free virus [183]. The ability of cell-free HIV-1 to infect these podocytes depended on the presence of the HIV-1 env gene and cell surface proteoglycans [183] .
Although the pathogenesis of most renal diseases is still unclear, DNA damage and the related DDR have been observed in acute kidney injury and chronic kidney disease [16]. HIV-1 Vpr protein can induce DNA damage, block DNA replication, activation of ATR (ataxia telangiectasia and Rad3-related protein), ubiquitination of histone H2B, and phosphorylation of histone H2AX that causes chromatin remodeling and recruitment of DNA repair proteins [184,185]. In addition, Vpr can induce DSBs [7,18,99] and alter cellular transcription through modulation of Nf-Kb [7,18,99]. Rosenstiel et al. evaluated the levels of phosphorylated H2AX to determine whether activation of the DNA damage response pathway is involved in HIV-1-associated nephropathy [186]. These authors transduced HK2 cells, a cell line phenotypically similar to primary renal tubular epithelial cells, with lentivirus expressing either HIV-1 Vpr and a GFP reporter gene via an IRES intermediate or the GFP reporter alone. Four days after transduction, Vpr-expressing renal tubular epithelial cells were hypertrophic with enlarged nuclei and showed increased phosphorylated H2AX immunofluorescence compared to control cells. Similar results were also observed in vivo experiments in mice and human renal biopsies from HIV-1-infected patients, where an increase in phosphorylated H2AX was also observed in tubular cells.
6.4. Neurological Dysfunctions and DNA Damage
HIV-1 can infect the nervous system, causing central and peripheral nervous disorders. Moreover, the nervous system is an HIV-1 sanctuary in which infections can persist [187]. About 10–20% of HIV-1-infected patients receiving ART are affected by HIV-1-associated neurocognitive disorders that are characterized by impaired memory, anxiety, apathy, mania, and motor dysfunctions [188]. Infection of the nervous system by HIV-1 depends on the ability of macrophages and lymphocytes to cross the blood-brain barrier and infect perivascular macrophages, microglia, and astrocytes, causing damage in neurons, oligodendrocytes, brain microvascular endothelial cells, astrocytes, and microglial cells. Several HIV-1 proteins, including gp120, Tat, Nef and Vpr, can to directly induce neuronal cell death [189]. Numerous scientific reports indicate that impaired DNA repair mechanisms and the accumulation of DNA damage in nervous system cells can lead to various neurological syndromes [17,190,191,192]. Consequently, DNA damage is likely a pathogenic factor in the development of central and peripheral nervous system dysfunction, while the proper functioning of DNA repair mechanisms may play a crucial role in preventing pathological conditions. Increasing evidence suggests that Vpr contributes to neuropathogenesis in individuals infected with HIV-1 [193]. The neurotoxic effects of Vpr have been attributed to various factors, including alterations in cell cycle progression, dysregulation of cellular metabolism, and disrupted signalling [193]. However, given both the role of Vpr in inducing DNA damage and the growing evidence that DNA damage plays a significant role in the pathogenesis of numerous neurological syndromes, it is plausible to suggest that Vpr-induced DNA damage may be a central factor in the nervous system disorders observed in HIV-1-infected patients. Moreover, Vpr ability to inhibit DNA repair mechanisms could further contribute to the accumulation of DNA damage in nervous system cells. Interesting data have emerged from studying DNA damage caused by stabilizing DNA-topoisomerase complexes associated with defects in DNA repair systems. Topoisomerases are enzymes that catalyze changes in the topological state of DNA, interconverting relaxed and supercoiled forms through transient breaks of single-stranded (topoisomerase 1) or double-stranded (topoisomerase 2) DNA. During this activity, a cleavage intermediate complex is formed from the end of the DNA covalently bound with the tyrosine of the topoisomerase active site. Under certain conditions, this complex can be stabilized, preventing DNA binding and triggering a DDR. Neurological dysfunctions caused by the accumulation of topoisomerase-mediated DNA damage have been observed, leading to the conclusion that topoisomerase activity may play a relevant role in these pathologies [194,195]. There are several similarities between Vpr-induced DNA damage and the mechanisms of nervous system dysfunction caused by the stabilization of topoisomerase-DNA complexes. As reported by Iijima et al. [8]. Vpr-induced unwinding of DNA causes accumulation of supercoiling of DNA and formation of topoisomerase 1-DNA covalent complex. Therefore, Vpr may be involved in the development of HIV-1-associated neurocognitive disorder by the accumulation of supercoiling of DNA and formation of topoisomerase 1-DNA covalent complex that causes DSBs [8]. The inhibition of DNA repair systems by Vpr would prevent the repair of these DSBs, making the brain dysfunction model in HIV-1 very similar to other non-HIV-1-related models described in the scientific literature [196,197,198].
7. New Therapeutic Strategies to Treat HIV Infection
7.1. HIV-1 Treatment Based on Capsid Inhibitors
Currently, most ART regimens use one or two drugs belonging to the NRTI class associated to an integrase or protease inhibitor. NRTIs can induce significant side effects, including lactic acidosis, pancreatitis, and renal failure [199,200,201]. Furthermore, several works have shown that NRTIs cause telomere shortening, resulting in cellular senescence and inflammation [118,119,120,121,122,123,124,125]. Currently, integrase inhibitors are drugs of first choice for the treatment of HIV-1 but are often combined with NRTIs. Capsid inhibitors are an emerging class of anti-HIV-1 drugs. Several key HIV-1 capsid inhibitors are either approved or under investigation. GS-6207 (Lenacapavir) was approved in 2022 by the FDA for treatment-experienced individuals with multidrug-resistant HIV [202]. It binds to a conserved arginine-rich region within the capsid hexamer pocket at the interface of CA monomers, inhibiting capsid disassembly (uncoating) and assembly in a dose-dependent manner. This mechanism affects both the early (nuclear import, reverse transcription) and late (assembly, maturation) stages of HIV replication. The binding of Lenacapavir allosterically alters the conformation of the capsid, disrupting the recruitment of host factors, such as CPSF6 and Nup153. CPSF6 functions in nuclear import and directs PIC to the host cell genome's nuclear speckles and gene-rich regions [28]. Without capsid-CPSF6 interaction, PIC is precluded from reaching nuclear speckles and gene-rich regions of chromatin [203]. Current clinical trials investigate Lenacapavir as an oral or subcutaneous agent as part of a dual therapy for both already treated and untreated patients. In phase 2 studies, the combination of Lenacapavir and Bictegravir treatment was well tolerated and induced a rapid reduction of plasma viremia in HIV-1-infected patients and was highly effective in maintaining virologic suppression [204,205,206,207]. As reported in section 5, the complexes LEDGF/integrase and CPSF6/capsid have different effects on the integration sites of HIV-1; moreover, Lenacapavir inhibits provirus integration into active genes, promoting integration into LADs, a condition that may favor the silencing of integrated proviruses. Future studies will be needed to understand the therapeutic potential of this drug combination in long-term treated patients. GS-CA1 and PF74 are capsid inhibitors currently in preclinical or early clinical trials [208]. They share a similar mechanism of action to GS-6207; however, PF74 binds near the host factor interaction sites, directly competing with host factors CPSF6 and Nup153 [208].
7.2. The "Shock and Kill" Therapeutic Approach
The "Shock and Kill" strategy is an experimental approach aimed at curing HIV-1 by targeting and eliminating latent viral reservoirs that persist despite ART. This method involves two primary steps: i) Shock (latency reversal), utilizing latency-reversing agents to reactivate dormant HIV-1 within infected cells, making the virus detectable; ii) Kill (elimination), where reactivated infected cells are targeted and destroyed by the immune system or therapeutic interventions. Several clinical trials have been conducted to evaluate the efficacy of this strategy. The ROADMAP study assessed the combination of the latency reversal agent romidepsin with the broadly neutralizing antibody 3BNC117. Despite reactivating latent HIV-1, the approach did not significantly reduce the viral reservoir [209]. Vorinostat, an HDAC inhibitor, was studied for its potential to reverse HIV-1 latency. While it successfully reactivated latent HIV-1 in some studies, eliminating the reactivated cells remains a challenge [210]. Studies have explored combining latency-reversing agents with agents that promote apoptosis of infected cells, such as BCL-2 inhibitors. However, an optimal combination of a latency-reversing agent and a BCL-2 inhibitor that enables maximal reactivation and selective elimination of HIV has yet to be identified. [211].
7.3. HIV-1 Treatment Based on Silencing or Knocking out the Provirus
Advancements in understanding the epigenetic regulation of HIV-1 have led to the development of novel therapeutic strategies, currently in the experimental stage, for treating individuals infected with the virus. The "Block and Lock" approach is an experimental strategy for achieving a functional cure for HIV-1, aiming to silence the virus permanently rather than eliminate it. In contrast to the "Shock and Kill" method, which seeks to reactivate and eradicate latent HIV-1, "Block and Lock" focuses on maintaining the virus in a deeply latent state, preventing its reactivation even in the absence of antiretroviral therapy [212]. To date, the only successful example of the ‘Block and Lock’ strategy is the use of didehydro-cortistatin A, a specific inhibitor of the Tat protein, which can achieve long-term suppression of HIV-1 reactivation through the establishment of epigenetic silencing [213,214,215]. Didehydro-cortistatin A inhibits HIV-1 by specific transcriptional and DNA methylation changes in genes related to the cell cycle, histones, and interferon responses. While preclinical studies have yielded promising results, translating this strategy into clinical practice remains challenging. Ensuring the long-term safety and efficacy of latency-promoting agents is crucial. Ongoing research continues to refine these approaches and evaluate their potential in human clinical trials. CRISPR-based gene editing is a promising strategy for curing HIV-1 infection by directly targeting and removing the viral genome from infected cells. CRISPR-Cas systems, particularly CRISPR-Cas9, cut and edit the HIV-1 genome or host genes involved in viral infection. This system consists of three main strategies: i) HIV-1 provirus elimination ("Cut and Remove"), in which CRISPR is programmed to target and excise the HIV-1 provirus from the DNA of infected cells [216]; ii) Blocking HIV-1 entry ("Gene Knockout") through gene-editing approaches aim to mimic mutations that make cells resistant to HIV-1 infection [217]; iii) Disrupting HIV-1 replication ("Gene Disruption") by CRISPR that disable essential HIV-1 genes (e.g., tat, rev, gag), preventing the virus from replicating [218]. This approach can keep latent HIV-1 from reactivating, reducing viral reservoirs. A prominent strategy involves the experimental therapy EBT-101, developed by Excision BioTherapeutics, Inc. EBT-101 utilizes the CRISPR-Cas9 system to excise HIV-1 proviral DNA from infected cells. Preclinical studies demonstrated its potential to remove HIV-1 DNA from various cell lines and animal models. In 2021, the U.S. Food and Drug Administration (FDA) authorised initiating clinical trials for EBT-101. Later, in July 2023, the FDA granted EBT-101 Fast Track Designation, accelerating its development process. Although preclinical animal studies have demonstrated promising curative potential, recent data presented at the 27th ASGCT meeting in Baltimore revealed that EBT-101 failed to prevent viral recurrence in three participants who discontinued antiretroviral therapy during the phase 1/2 trial, requiring them to restart ART treatment [219].
8. Conclusions
The intricate interactions between HIV-1 and the host genome, explored in this review, offer valuable insights into the underlying reasons for the poor outcomes in achieving a definitive cure for HIV-1 infection despite the use of the most advanced therapeutic strategies. Factors such as DNA damage induced by HIV-1, inhibition of DNA repair pathways, telomere attrition, the orientation of the provirus within host genes, clonal expansion of CD4+ T cells harboring the provirus in a latent form, and integration into specific genes like oncogenes or KRAB-ZNF genes are likely among the key contributors that hinder progress toward a lasting cure. A more in-depth study of cases where HIV-1-infected individuals have achieved a prolonged aviremic state without ART could guide research toward more effective therapeutic strategies. Bone marrow transplantation (BMT) has shown potential in eradicating HIV-1 in some instances, although it remains an exceptionally rare occurrence. The most notable examples are the cases of Timothy Ray Brown, known as the 'Berlin Patient,' and the 'London Patient,' both of whom received stem cell transplants from donors carrying the CCR5-Δ32 genetic mutation, which confers resistance to HIV-1 infection [220,221]. These cases suggest a gene-editing approach to mimic this mutation, rendering cells resistant to HIV-1 infection, could represent a feasible therapeutic strategy. The study of mechanisms through which elite controllers maintain aviremia or low viremia without ART could guide research towards more effective strategies for treating HIV-1 infection [222]. The aviremic or low-viremia state in elite controllers is often attributed to a combination of host genetic, epigenetic, and viral factors, such as the CCR5-Δ32 mutation, highly efficient HIV-1-specific CD8+ T cells, enhanced natural killer cell activity, increased production of interferons and other antiviral cytokines, and HIV-1 proviruses subject to epigenetic silencing. However, replicating this unique interaction of factors as therapeutic strategies in HIV-1-infected patients by combining treatments such as ‘Cut and Remove’, ‘Gene Knockout’ and ‘Gene Disruption’ remains a challenge. Lenacapavir stands out as an innovative drug due to its unique mechanism of action, distinguishing it from NRTIs, NNRTIs, protease inhibitors, and integrase inhibitors. Its use in combination with other antiviral drugs, such as bictegravir or islatravir [223], appears both safe and effective. Furthermore, its long-acting properties have the potential to significantly improve the quality of life for HIV-1-infected patients. Future studies will be needed to understand the therapeutic potential of capsid inhibitors in combination with other classes of anti-HIV-1 drugs in long-term treated patients.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not available.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Krebs, A.S.; Mendonça, L.M.; Zhang, P. Structural Analysis of Retrovirus Assembly and Maturation. Viruses 2021, 14, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Maertens, G.N.; Engelman, A.N.; Cherepanov, P. Structure and function of retroviral integrase. Nat. Rev. Microbiol. 2022, 20, 20–34. [Google Scholar] [CrossRef] [PubMed]
- Grawenhoff, J.; Engelman, A.N. Retroviral integrase protein and intasome nucleoprotein complex structures. World J. Biol. Chem. 2017, 8, 32–44. [Google Scholar] [CrossRef]
- Dopkins, N.; O’Mara, M.M.; Lawrence, E.; Fei, T. : Sandoval-Motta, S.; Nixon, D.F.; Bendall, M.L. A field guide to endogenous retrovirus regulatory networks. Mol. Cell. 2022, 82, 3763–3768. [Google Scholar] [CrossRef]
- Zahn, J.; Kaplan, M.H.; Fischer, S.; Dai, M.; Meng, F.; Saha, A.K.; Cervantes, P.; Chan, S.M.; Dube, D.; Omenn, G.S.; et al. Expansion of a novel endogenous retrovirus throughout the pericentromeres of modern humans. Genome Biol. 2015, 16, 74. [Google Scholar] [CrossRef]
- Roshal, M.; Kim, B.; Zhu, Y.; Nghiem, P.; Planelles, V. Activation of the ATR-mediated DNA damage response by the HIV-1 viral protein R. J. Biol. Chem. 2003, 278, 25879–86. [Google Scholar] [CrossRef]
- Zimmerman, E.S.; Sherman, M.P.; Blackett, J.L.; Neidleman, J.A.; Kreis, C.; Mundt, P.; Williams, S.A.; Warmerdam, M.; Kahn, J.; Hecht, F.M.; et al. Human immunodeficiency virus type 1 Vpr induces DNA replication stress in vitro and in vivo. J. Virol. 2006, 80, 10407–18. [Google Scholar] [CrossRef]
- Iijima, K.; Kobayashi, J.; Ishizaka, Y. Structural alteration of DNA induced by viral protein R of HIV-1 triggers the DNA damage response. Retrovirology 2018, 15, 8. [Google Scholar] [CrossRef]
- Li, D.; Lopez, A.; Sandoval, C.; Nichols, D.R.; Fregoso, O.I. HIV Vpr modulates the host DNA damage response at two independent steps to damage DNA and repress DSB repair. MBio 2020, 11, e00940–20. [Google Scholar] [CrossRef]
- Wong, T.; Luperchio, A.M.; Riley, S.; Salamango, D.J. Inhibition of ATM-directed antiviral responses by HIV-1 Vif. PLoS Pathog. 2023, 19, e1011634. [Google Scholar] [CrossRef]
- Wagner, T.A.; McLaughlin, S.; Garg, K.; Cheung, C.Y.; Larsen, B.B.; Styrchak, S.; Huang, H.C.; Edlefsen, P.T.; Mullins, J.I.; Frenkel, L.M. HIV latency. Proliferation of cells with HIV integrated into cancer genes contributes to persistent infection. Science 2014, 345, 570–573. [Google Scholar] [CrossRef]
- Dragoni, F.; Kwaa, A.K.; Traut, C.C.; Veenhuis, R.T.; Woldemeskel, B.A.; Camilo-Contreras, A.; Raymond, H.E.; Dykema, A.G.; Scully, E.P.; Rosecrans, A.M.; et al. Proviral location affects cognate peptide-induced virus production and immune recognition of HIV-1-infected T cell clones. J. Clin. Investig. 2023, 133, e171097. [Google Scholar] [CrossRef]
- Zhao, J.; Nguyen, L.N.T.; Dang, X.; Cao, D.; Khanal, S.; Schank, M.; Thakuri, B.K.C.; Ogbu, S.C.; Morrison, Z.D.; Wu, X.Y. , et al. ATM deficiency accelerates DNA damage, telomere erosion, and premature T cell aging in HIV-infected individuals on antiretroviral therapy. Front. Immunol. 2019, 10, 2531. [Google Scholar] [CrossRef]
- Khanal, S.; Tang, Q.; Cao, D.; Zhao, J.; Nguyen, L.N.; Oyedeji, O.S.; Dang, X.; Nguyen, L.N.T.; Schank, M.; Thakuri, B.K.C.; et al. Telomere and ATM Dynamics in CD4 T-Cell Depletion in Active and Virus-Suppressed HIV Infections. J. Virol. 2020, 94, e01061–20. [Google Scholar] [CrossRef]
- Simenauer, A.; Nozik-Grayck, E.; Cota-Gomez, A. The DNA Damage Response and HIV-Associated Pulmonary Arterial Hypertension. Int. J. Mol. Sci. 2020, 21, 3305. [Google Scholar] [CrossRef]
- Wang, P.; Ouyang, J.; Jia, Z.; Zhang, A.; Yang, Y. Roles of DNA damage in renal tubular epithelial cells injury. Front. Physiol. 2023, 14, 1162546. [Google Scholar] [CrossRef]
- Delint-Ramirez, I.; Madabhushi, R. NPAS4 juggles neuronal activity-dependent transcription and DSB repair with NuA4. Mol. Cell. 2023, 83, 1208–1209. [Google Scholar] [CrossRef]
- Chen, B. Molecular mechanism of HIV-1 entry. Trends Microbiol. 2019, 27, 878–891. [Google Scholar] [CrossRef]
- Novikova, M.; Zhang, Y.; Freed, E.O.; Peng, K. Multiple roles of HIV-1 capsid during the virus replication cycle. Virol. Sin. 2019, 34, 119–134. [Google Scholar] [CrossRef]
- Arhel, N. Revisiting HIV-1 uncoating. Retrovirology 2010, 7, 96–105. [Google Scholar] [CrossRef]
- Bukrinsky, M.I.; Haggerty, S.; Dempsey, M.P.; Sharova, N.; Adzhubel, A.; Spitz, L.; Lewis, P.; Goldfarb, D.; Emerman, M.; Stevenson, M. A nuclear localization signal within HIV-1 matrix protein that governs infection of non-dividing cells. Nature 1993, 365, 666–9. [Google Scholar] [CrossRef] [PubMed]
- Depienne, C.; Mousnier, A.; Leh, H.; Le Rouzic, E.; Dormont, D.; Benichou, S.; Dargemont, C. Characterization of the nuclear import pathway for HIV-1 integrase. J. Biol. Chem. 2001, 276, 18102–18107. [Google Scholar] [CrossRef] [PubMed]
- Gallay, P.; Hope, T.; Chin, D.; Trono, D. HIV-1 infection of nondividing cells through the recognition of Integrase by the Importin/Karyopherin pathway. Proc. Natl. Acad. Sci. USA 1997, 94, 9825–9830. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.L.; Spearman, P.; Ratner, L. Human immunodeficiency virus type 1 viral protein R localization in infected cells and virions. J. Virol. 1993, 67, 542–6550. [Google Scholar] [CrossRef]
- Yamashita, M.; Emerman, M. Capsid is a dominant determinant of retrovirus infectivity in nondividing cells. J. Virol. 2004, 78, 5670–5678. [Google Scholar] [CrossRef]
- Yamashita, M.; Perez, O.; Hope, T.J.; Emerman, M. Evidence for direct involvement of the capsid protein in HIV infection of nondividing cells. PLoS Pathog. 2007, 3, 1502–1510. [Google Scholar] [CrossRef]
- Dwivedi, R.; Prakash, P.; Kumbhar, B.V.; Balasubramaniam, M.; Dash, C. HIV-1 capsid and viral DNA integration. mBio 2024, 15, e0021222. [Google Scholar] [CrossRef]
- Burdick, R.C.; Morse, M.; Rouzina, I.; Williams, M.C.; Hu, W.S.; Pathak, V.K. HIV-1 uncoating requires long double-stranded reverse transcription products. Sci. Adv. 2024, 10, eadn7033. [Google Scholar] [CrossRef]
- Lesbats, P.; Engelman, A. N.; Cherepanov, P. Retroviral DNA integration. Chem. Rev. 2016, 116, 12730–12757. [Google Scholar]
- Anisenko, A.; Nefedova, A.; Agapkina, Y.; Gottikh, M. Both ATM and DNA-PK Are the Main Regulators of HIV-1 Post-Integrational DNA Repair. Int. J. Mol. Sci. 2023, 24, 2797–2809. [Google Scholar] [CrossRef]
- Vincent, K. A.; York-Higgins, D.; Quiroga, M.; Brown, P. O. Host sequences flanking the HIV provirus. Nucleic Acids Res. 1990, 18, 6045–6047. [Google Scholar] [CrossRef] [PubMed]
- Vink, C.; Groenink, M.; Elgersma, Y.; Fouchier, R. A.; Tersmette, M.; Plasterk, R. H. Analysis of the junctions between human immunodeficiency virus type 1 proviral DNA and human DNA. J. Virol. 1990, 64, 5626–5627. [Google Scholar] [CrossRef] [PubMed]
- Anisenko, A.N.; Knyazhanskaya, E.S.; Isaguliants, M.G.; Gottikh, M.B. A qPCR assay for measuring the post-integrational DNA repair in HIV-1 replication. J. Virol. Methods. 2018, 262, 12–19. [Google Scholar] [CrossRef]
- Anisenko, A.N.; Nefedova, A.A.; Kireev, I.I.; Gottikh, M.B. Post-Integrational DNA Repair of HIV-1 Is Associated with Activation of the DNA-PK and ATM Cellular Protein Kinases and Phosphorylation of Their Targets. Biochemistry (Mosc) 2024, 89, 1122–1132. [Google Scholar] [CrossRef]
- Barnes, G.; Rio, D. DNA double-strand-break sensitivity, DNA replication, and cell cycle arrest phenotypes of Ku-deficient Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 1997, 94, 867–72. [Google Scholar] [CrossRef]
- Schroder, A.R.; Shinn, P.; Chen, H.; Berry, C.; Ecker, J.R.; Bushman, F. HIV-1 integration in the human genome favors active genes and local hotspots. Cell 2002, 110, 521–529. [Google Scholar] [CrossRef]
- Wang, G.P.; Ciuffi, A.; Leipzig, J.; Berry, C.C.; Bushman, F.D. HIV integration site selection: Analysis by massively parallel pyrosequencing reveals association with epigenetic modifications. Genome Res. 2007, 17, 1186–1194. [Google Scholar] [CrossRef]
- Roth, S.L.; Malani, N.; Bushman, F.D. Gammaretroviral integration into nucleosomal target DNA in vivo. J. Virol. 2011, 85, 7393–7401. [Google Scholar] [CrossRef]
- Sowd, G.A.; Serrao, E.; Wang, H.; Wang, W.; Fadel, H.J.; Poeschla, E.M.; Engelman, A.N. A critical role for alternative polyadenylation factor CPSF6 in targeting HIV-1 integration to transcriptionally active chromatin. Proc. Natl. Acad. Sci. USA 2016, 113, E1054–E1063. [Google Scholar] [CrossRef]
- Kvaratskhelia, M.; Sharma, A.; Larue, R.C.; Serrao, E.; Engelman, A. Molecular mechanisms of retroviral integration site selection. Nucleic Acids Res. 2014, 42, 10209–10225. [Google Scholar] [CrossRef]
- Cohn, L.B.; Silva, I.T.; Oliveira, T.Y.; Rosales, R.A.; Parrish, E.H.; Learn, G.H.; Hahn, B.H.; Czartoski, J.L.; McElrath, M.J.; Lehmann, C.; et al. HIV-1 integration landscape during latent and active infection. Cell 2015, 160, 420–432. [Google Scholar] [CrossRef] [PubMed]
- Maldarelli, F.; Wu, X.; Su, L.; Simonetti, F.R.; Shao, W.; Hill, S.; Spindler, J.; Ferris, A.L.; Mellors, J.W.; Kearney, M.F.; et al. HIV latency. Specific HIV integration sites are linked to clonal expansion and persistence of infected cells. Science 2014, 345, 179–183. [Google Scholar] [CrossRef]
- Saleh, S.; Lu, H.K.; Evans, V.; Harisson, D.; Zhou, J.; Jaworowski, A.; Sallmann, G.; Cheong, K.Y.; Mota, T.M.; Tennakoon, S.; et al. HIV integration and the establishment of latency in CCL19-treated resting CD4+ T cells require activation of NF-κB. Retrovirology 2016, 13, 49–60. [Google Scholar] [CrossRef]
- Brady, T.; Agosto, L.M.; Malani, N.; Berry, C.C.; O’Doherty, U.; Bushman, F. HIV integration site distributions in resting and activated CD4 + T cells infected in culture. AIDS 2009, 23, 1461–1471. [Google Scholar] [CrossRef]
- Engelman, A.N.; Maertens, G.N. Virus-host interactions in retrovirus integration. In Retrovirus-Cell Interactions; Parent, L.J., Ed.; Academic Press: San Diego, CA, USA, 2018; pp. 163–198. [Google Scholar]
- Bedwell, G.J.; Engelman, A.N. Factors that mold the nuclear landscape of HIV-1 integration. Nucleic Acids Res. 2021, 49, 621–635. [Google Scholar] [CrossRef]
- Singh, P.K.; Bedwell, G.J.; Engelman, A.N. Spatial and genomic correlates of HIV-1 integration site targeting. Cells 2022, 11, 655. [Google Scholar] [CrossRef]
- Yedavalli, V.R.K.; Jeang, K.T. Methylation: a regulator of HIV-1-1 replication? Retrovirology.
- Willemsen, N.M.; Hitchen, E.M.; Bodetti, T.J.; Apolloni, A.; Warrilow, D.; Piller, S.C.; Harrich, D. Protein methylation is required to maintain optimal HIV-1-1 infectivity. Retrovirology 2006, 3, 92. [Google Scholar] [CrossRef]
- Sapp, N.; Burge, N.; Cox, K.; Prakash, P.; Balasubramaniam, M.; Thapa, S.; Christensen, D.; Li, M.; Linderberger, J.; Kvaratskhelia, M.; et al. HIV-1 preintegration complex preferentially integrates the viral DNA into nucleosomes containing trimethylated histone 3-lysine 36 modification and flanking linker DNA. J. Virol. 2022, 96, e01011–22. [Google Scholar] [CrossRef]
- Vansant, G.; Chen, H.C.; Zorita, E.; Trejbalová, K.; Miklík, D.; Filion, G.; Debyser, Z. The chromatin landscape at the HIV-1 provirus integration site determines viral expression. Nucleic Acids Res. 2020, 48, 7801–7817. [Google Scholar] [CrossRef]
- Bussey-Sutton, C.R.; Ward, A.; Fox, J.A. Turner, A.W.; Peterson, J.J.; Emery, A.; Longoria, A.R.; Gomez-Martinez, I.; Jones, C.; Hepperla, A.; et al. The histone methyltransferase SETD2 regulates HIV expression and latency. PLoS Pathog. 2024, 20, e1012281. [Google Scholar]
- Nguyen, K.; Karn, J. The sounds of silencing: dynamic epigenetic control of HIV latency. Curr. Opin. HIV AIDS. 2024, 19, 102–109. [Google Scholar] [CrossRef]
- Nikolaitchik, O.A.; Islam, S.; Kitzrow, J.P.; Duchon, A.; Cheng, Z.; Liu, Y.; Rawson, J.M.O.; Shao, W.; Nikolaitchik, M.; Kearney, M.F.; et al. HIV-1 usurps transcription start site heterogeneity of host RNA polymerase II to maximize replication fitness. Proc. Natl. Acad. Sci. U S A 2023, 120, e2305103120. [Google Scholar] [CrossRef]
- Ciuffi, A.; Llano, M.; Poeschla, E.; Hoffmann, C.; Leipzig, J.; Shinn, P.; Ecker, J.R.; Bushman, F. A role for LEDGF/p75 in targeting HIV DNA integration. Nat. Med. 2005, 11, 1287–1289. [Google Scholar] [CrossRef] [PubMed]
- Shun, M.C.; Raghavendra, N.K.; Vandegraaff, N.; Daigle, J.E.; Hughes, S.; Kellam, P.; Cherepanov, P.; Engelman, A. LEDGF/p75 functions downstream from preintegration complex formation to effect gene-specific HIV-1 integration. Genes Dev. 2007, 21, 1767–1778. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.K.; Plumb, M.R.; Ferris, A.L.; Iben, J.R.; Wu, X.; Fadel, H.J.; Luke, B.T.; Esnault, C.; Poeschla, E.M.; Hughes, S.H.; et al. LEDGF/p75 interacts with mRNA splicing factors and targets HIV-1 integration to highly spliced genes. Genes Dev. 2015, 29, 2287–2297. [Google Scholar] [CrossRef]
- Busschots, K.; Vercammen, J.; Emiliani, S.; Benarous, R.; Engelborghs, Y.; Christ, F.; Debyser, Z. The interaction of LEDGF/p75 with integrase is lentivirus-specific and promotes DNA binding. J. Biol. Chem. 2005, 280, 17841–17847. [Google Scholar] [CrossRef]
- Llano, M.; Delgado, S.; Vanegas, M.; Poeschla, E.M. Lens epithelium-derived growth factor/p75 prevents proteasomal degradation of HIV-1 integrase. J. Biol. Chem. 2004, 279, 55570–55577. [Google Scholar] [CrossRef]
- Cherepanov, P. LEDGF/p75 interacts with divergent lentiviral integrases and modulates their enzymatic activity in vitro. Nucleic Acids Res. 2007, 35, 113–124. [Google Scholar] [CrossRef]
- Kok, Y.L.; Vongrad, V.; Chaudron, S.E.; Shilaih, M.; Leemann, C.; Neumann, K.; Kusejko, K.; Di Giallonardo, F.; Kuster, H.; Braun, D,L. HIV-1 integration sites in CD4+ T cells during primary, chronic, and late presentation of HIV-1 infection. J.C.I. Insight 2021, 6, e143940. [Google Scholar] [CrossRef]
- Miura, Y.; Morooka, M.; Sax, N.; Roychoudhuri, R.; Itoh-Nakadai, A.; Brydun, A.; Funayama, R.; Nakayama, K.; Satomi, S.; Matsumoto, M.; et al. Bach2 promotes B cell receptor-induced proliferation of B lymphocytes and represses cyclin-dependent kinase inhibitors. J. Immunol. 2018, 200, 2882–2893. [Google Scholar] [CrossRef]
- Weng, X.; Zheng, M.; Liu, Y.; Lou, G. The role of Bach2 in regulating CD8 + T cell development and function. Cell. Commun. Signal. 2024, 22, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Shaposhnikov, D.; Kuffer, C.; Storchova, Z.; & Posern, G.; Posern, G. Myocardin related transcription factors are required for coordinated cell cycle progression. Cell cycle (Georgetown, Tex.) 2013, 12, 1762–1772. [Google Scholar] [CrossRef]
- Lei, J.; Hu, D.; Xue, S.; Mao, F.; Obeng, E.; Quan, Y.; Yu, W. HN1L is essential for cell growth and survival during nucleopolyhedrovirus infection in silkworm, Bombyx mori. PLoS One 2019, 14, e0216719. [Google Scholar] [CrossRef]
- Smith, E. R.; Cayrou, C.; Huang, R.; Lane, W. S.; Cote, J.; Lucchesi, J. C. A human protein complex homologous to the Drosophila MSL complex is responsible for the majority of histone H4 acetylation at lysine 16. Molec. Cell. Biol. 2005, 25, 9175–9188. [Google Scholar] [CrossRef]
- Makarov, E.M.; Makarova, O.V.; Achsel, T.; Luhrmann, R. The human homologue of the yeast splicing factor prp6p contains multiple TPR elements and is stably associated with the U5 snRNP via protein-protein interactions. J. Mol. Biol. 2000, 298, 567–75. [Google Scholar] [CrossRef]
- Han, Y.; Lassen, K.; Monie, D.; Sedaghat, A.R.; Shimoji, S.; Liu, X.; Pierson, T.C.; Margolick, J.B.; Siliciano, R.F.; Siliciano, J.D. Resting CD4+ T cells from human immunodeficiency virus type 1 (HIV-1)-infected individuals carry integrated HIV-1 genomes within actively transcribed host genes. Journal of virology 2004, 78, 6122–6133. [Google Scholar] [CrossRef]
- Ikeda, T.; Shibata, J.; Yoshimura, K.; Koito, A.; Matsushita, S. Recurrent HIV-1 integration at the BACH2 locus in resting CD4+ T cell populations during effective highly active antiretroviral therapy. The Journal of infectious diseases 2007, 195, 716–725. [Google Scholar] [CrossRef]
- Mack, K.D.; Jin, X.; Yu, S.; Wei, R.; Kapp, L.; Green, C.; Herndier, B.; Abbey, N.W.; Elbaggari, A.; Liu, Y.; et al. HIV insertions within and proximal to host cell genes are a common finding in tissues containing high levels of HIV DNA and macrophage-associated p24 antigen expression. Journal of acquired immune deficiency syndromes 2003, 33, 308–320. [Google Scholar] [CrossRef]
- Maldarelli, F.; Kearney, A.M.; Palmer, S.; Stephens, R.; Mican, J.; Polis, M.A.; Davey, R.T.; Kovacs, J.; Shao, W.; Rock-Kress, D.; et al. HIV populations are large and accumulate high genetic diversity in a nonlinear fashion. Journal of virology 2013, 87, 10313–10323. [Google Scholar] [CrossRef]
- Flucke, U.; Tops, B.B.; de Saint Aubain Somerhausen, N.; Bras, J.; Creytens, D.H.; Kusters, B.; Groenen, P.J.; Verdijk, M.A.; Suurmeijer, A.J.; Mentzel, T. Presence of C11orf95-MKL2 fusion is a consistent finding in chondroid lipomas: a study of eight cases. Histopathology 2013, 62, 925–930. [Google Scholar] [CrossRef]
- Kobayashi, S.; Taki, T.; Chinen, Y.; Tsutsumi, Y.; Ohshiro, M.; Kobayashi, T.; Matsumoto, Y.; Kuroda, J.; Horiike, S.; Nishida, K.; et al. Identification of IGHCdelta-BACH2 fusion transcripts resulting from cryptic chromosomal rearrangements of 14q32 with 6q15 in aggressive B-cell lymphoma/leukemia. Genes, chromosomes & cancer 2011, 50, 207–216. [Google Scholar]
- Muehlich, S.; Hampl, V.; Khalid, S.; Singer, S.; Frank, N.; Breuhahn, K.; Gudermann, T.; Prywes, R. The transcriptional coactivators megakaryoblastic leukemia 1/2 mediate the effects of loss of the tumor suppressor deleted in liver cancer 1. Oncogene 2012, 31, 3913–3923. [Google Scholar]
- Liu, J.; Sørensen, A.B.; Wang, B.; Wabl, M.; Nielsen, A.L. Pedersen FS. Identification of novel Bach2 transcripts and protein isoforms through tagging analysis of retroviral integrations in B-cell lymphomas. BMC Mol. Biol. 2009, 10, 2199–2210. [Google Scholar] [CrossRef]
- Wang, H.; Bei, L.; Shah, C.A.; Huang, W.; Platanias, L.C.; Eklund, E.A. The E3 ubiquitin ligase Triad1 influences development of Mll-Ell-induced acute myeloid leukemia. Oncogene 2018, 37, 2532–2544. [Google Scholar] [CrossRef]
- Geng, S.; Peng, W.; Wang, X.; Hu, X.; Liang, H.; Hou, J.; Wang, F.; Zhao, G.; Lü, M.; Cui, H. ARIH2 regulates the proliferation, DNA damage and chemosensitivity of gastric cancer cells by reducing the stability of p21 via ubiquitination. Cell Death & Disease, 2022; 13, 1–12. [Google Scholar] [CrossRef]
- Thomason, P.A.; King, J.S.; Insall, R.H. Mroh1, a lysosomal regulator localized by WASH-generated actin. J. Cell. Sci. 2017, 130, 1785–1795. [Google Scholar] [CrossRef]
- Wu, Y.; Gou, Y.; Wang, T.; Li, P.; Li, Y.; Lu, X.; Li, W.; Liu, Z. Exportin XPO6 upregulation activates the TLR2/MyD88/NF-kappaB signaling by facilitating TLR2 mRNA nuclear export in COPD pulmonary monocytes. Int. Immunopharmacol. 2024, 135, 112310. [Google Scholar] [CrossRef]
- Jordan, A.; Defechereux, P.; Verdin, E. The site of HIV-1 integration in the human genome determines basal transcriptional activity and response to Tat transactivation. EMBO J. 2001, 20, 1726–1738. [Google Scholar] [CrossRef]
- Jordan, A.; Bisgrove, D.; Verdin, E. HIV reproducibly establishes a latent infection after acute infection of T cells in vitro. EMBO J. 2003, 22, 1868–1877. [Google Scholar] [CrossRef]
- Pearson, R.; Kim, Y.K.; Hokello, J.; Lassen, K.; Friedman, J.; Tyagi, M.; Karn, J. Epigenetic silencing of human immunodeficiency virus (HIV) transcription by formation of restrictive chromatin structures at the viral long terminal repeat drives the progressive entry of HIV into latency. J. Virol. 2008, 82, 12291–12303. [Google Scholar] [CrossRef]
- Einkauf, K.B.; Lee, G.Q.; Gao, C.; Sharaf, R.; Sun, X.; Hua, S.; Chen, S.M.Y.; Jiang, C.; Lian, X.; Chowdhury, F.Z.; et al. Intact HIV-1 proviruses accumulate at distinct chromosomal positions during prolonged antiretroviral therapy. J. Clin. Invest. 2019, 129, 988–998. [Google Scholar] [CrossRef]
- Jiang, C.; Lian, X.; Gao, C.; Sun, X.; Einkauf, K.B.; Chevalier, J.M.; Chen, S.M.Y.; Hua, S.; Rhee, B.; Chang, K.; et al. Distinct viral reservoirs in individuals with spontaneous control of HIV-1. Nature 2020, 585, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.S.; Ramos, V.; Oliveira, T.Y.; Gaebler, C.; Jankovic, M.; Nussenzweig, M.C.; and Cohn, L.B. Integration features of intact latent HIV-1 in CD4+ T cell clones contribute to viral persistence. J. Exp. Med. 2021, 218, e20211427. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, K.; Das, B.; Dobrowolski, C.; Karn, J. Multiple histone lysine methyltransferases are required for the establishment and maintenance of HIV-1 latency. MBio 2017, 8, e00133–17. [Google Scholar] [CrossRef]
- Uckelmann, M.; Davidovich, C. Chromatin compaction by Polycomb group proteins revisited. Curr. Opin. Struct. Biol. 2024, 86, 102806. [Google Scholar] [CrossRef]
- Williams, S.A.; Greene, W.C. Regulation of HIV-1 Latency by T-cell Activation. Cytokine 2007, 39, 63–74. [Google Scholar] [CrossRef]
- Bosque, A.; Planelles, V. Induction of HIV-1 latency and reactivation in primary memory CD4+ T cells. Blood 2009, 113, 58–65. [Google Scholar] [CrossRef]
- Tyagi, M.; Pearson, R.J.; Karn, J. Establishment of HIV latency in primary CD4+ cells is due to epigenetic transcriptional silencing and P-TEFb restriction. J. Virol. 2010, 84, 6425–6437. [Google Scholar] [CrossRef]
- Han, Y.; Lin, Y.B.; An, W.; Xu, J.; Yang, H.C.; O'Connell, K.; Dordai, D.; Boeke, J.D.; Siliciano, J.D.; Siliciano, R.F. Orientation-dependent regulation of integrated HIV-1 expression by host gene transcriptional readthrough. Cell Host Microbe 2008, 2, 134–46. [Google Scholar] [CrossRef]
- Shan, L.; Yang, H.C.; Rabi, S.A.; Bravo, H.C.; Shroff, N.S.; Irizarry, R.A.; Zhang, H.; Margolick, J.B.; Siliciano, J.D.; Siliciano, R.F. Influence of host gene transcription level and orientation on HIV-1 latency in a primary-cell model. J. Virol. 2011, 11, 5384–5393. [Google Scholar] [CrossRef]
- Li, G.; Piampongsant, S.; Faria, N.R.; Voet, A.; Pineda-Peña, A.C.; Khouri, R.; Lemey, P.; Vandamme, A.M.; Theys, K. An integrated map of HIV genome-wide variation from a population perspective. Retrovirology 2015, 12, 18. [Google Scholar] [CrossRef]
- Jasinska, A.J. , Apetrei, C., Pandrea, I. Walk on the wild side: SIV infection in African non-human primate hosts-from the field to the laboratory. Front Immunol. 2023, 13, 1060985. [Google Scholar] [CrossRef] [PubMed]
- Casey, L.; Wen, X.; de Noronha, C.M. The functions of the HIV1 protein Vpr and its action through the DCAF1.DDB1.Cullin4 ubiquitin ligase. Cytokine 2010, 51, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, C.C.; Spies, M. Dynamic elements of replication protein A at the crossroads of DNA replication, recombination, and repair. Critical Reviews in Biochemistry and Molecular Biology 2020, 55, 482–507. [Google Scholar] [CrossRef] [PubMed]
- Dueva, R.; Iliakis, G. Replication protein A: a multifunctional protein with roles in DNA replication, repair and beyond. NAR Cancer 2020, 2, zcaa022. [Google Scholar] [CrossRef]
- Sandoval, C.; Nisson, K.; Fregoso, O.I. HIV-1 Vpr induces cell cycle arrest by activating the ATR DNA damage response pathway. PLoS Pathogens 2012, 8, e1002680. [Google Scholar]
- Andersen, J.L.; Planelles, V. The role of Vpr in HIV-1 pathogenesis. Curr. HIV Res. 2005, 3, 43–51. [Google Scholar] [CrossRef]
- Sandoval, C.; Nisson, K.; Fregoso, O.I. HIV-1 Vpr-induced DNA damage activates NF-κB through ATM-NEMO independent of cell cycle arrest. mBio. 2024, 15, e0024024. [Google Scholar] [CrossRef]
- Li, X.; Heyer, W.D. Homologous recombination in DNA repair and DNA damage tolerance. Cell Res. 2008, 18, 99–113. [Google Scholar] [CrossRef]
- Brandsma, I.; Gent, D.C. Pathway choice in DNA double strand break repair: observations of a balancing act. Genome Integr 2012, 3, 9–18. [Google Scholar] [CrossRef]
- Jäger, S.; Kim, D.Y.; Hultquist, J.F.; Shindo, K.; LaRue, R.S.; Kwon, E.; Li, M.; Anderson, B.D.; Yen, L.; Stanley, D.; et al. Vif hijacks CBF-β to degrade APOBEC3G and promote HIV-1 infection. Nature 2011, 481, 371–5. [Google Scholar] [CrossRef]
- Ramos, F.; Villoria, M.T.; Alonso-Rodriguez, E.; Clemente-Blanco, A. Role of protein phosphatases PP1, PP2A, PP4 and Cdc14 in the DNA damage response. Cell Stress 2019, 3, 70–85. [Google Scholar] [CrossRef] [PubMed]
- Campos, A.; Clemente-Blanco, A. Cell cycle and DNA repair regulation in the damage response: protein phosphatases take over the reins. Int. J. Mol. Sci. 2020, 21, 446–472. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, E.; Bruhn, C.; Peretti, M.; Cassani, C.; Carotenuto, W.V.; Elgendy, M.; Hubassi, G.; Lucca, C.; Bermejo, R.; Varasi, M.; et al. PP2A controls genome integrity by integrating nutrient-sensing and metabolic pathways with the DNA damage response. Mol. Cell. 2017, 67, 266–81. [Google Scholar] [CrossRef]
- Ambjorn, S.M.; Duxin, J.P.; Hertz, E.P.T.; Nasa, I.; Duro, J.; Kruse, T.; Lopez-Mendez, B.; Rymarczyk, B.; Cressey, L.E.; van Overeem Hansen, T.; et al. A complex of BRCA2 and PP2A-B56 is required for DNA repair by homologous recombination. Nat. Commun. 2021, 12, 5748–58. [Google Scholar] [CrossRef] [PubMed]
- Piekna-Przybylska, D.; Sharma, G. .; Maggirwar, S.B.; Bambara, R.A. Deficiency in DNA damage response, a new characteristic of cells infected with latent HIV. Cell Cycle 2017, 16, 968–978. [Google Scholar] [CrossRef]
- Bellon, M.; Nicot, C. Telomere dynamics in immune senescence and exhaustion triggered by chronic viral infection. Viruses 2017, 9, E289. [Google Scholar] [CrossRef]
- Fülöp, T.; Herbein, G.; Cossarizza, A.; Witkowski, J.M.; Frost, E.; Dupuis, G.; et al. Cellular senescence, immunosenescence and HIV. Interdiscip. Top. Gerontol. Geriatr. 2017, 42, 28–46. [Google Scholar]
- Blackburn, E.H. Switching and signaling at the telomere. Cell 2001, 106, 661–73. [Google Scholar]
- Lin, Y.; Damjanovic, A.; Metter, E.J.; Nguyen, H.; Truong, T.; Najarro, K.; Morris, C.; Longo, D.L.; Zhan, M.; Ferrucci, L.; et al. Age-associated telomere attrition of lymphocytes in vivo is co-ordinated with changes in telomerase activity, composition of lymphocyte subsets and health conditions. Clin. Sci. (Lond.) 2015, 128, 367–77. [Google Scholar]
- Rothkamm, K.; Barnard, S.; Moquet, J.; Ellender, M.; Rana, Z.; Burdak-Rothkamm, S. DNA damage foci: meaning and significance. Environ. Mol. Mutagen. 2015, 56, 491–504. [Google Scholar] [CrossRef]
- Alejos, B.; Stella-Ascariz, N.; Montejano, R.; Rodriguez-Centeno, J.; Schwimmer, C.; Bernardino, J.I.; Rodes, B.; Esser, S.; Goujard, C.; Sarmento-Castro, R.; et al. Determinants of blood telomere length in antiretroviral treatment-naive HIV-positive participants enrolled in the NEAT 001/ANRS 143 clinical trial. HIV Med. 2019, 20, 691–8. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, V.C.; Wit, F.W.; Joerink, M.; Maurer, I.; Harskamp, A.M.; Schouten, J.; Prins, M.; van Leeuwen, E.M.; Booiman, T.; Deeks, S.G.; et al. T-cell activation independently associates with immune senescence in HIV infected recipients of long-term antiretroviral treatment. J. Infect. Dis. 2016, 214, 216–225. [Google Scholar] [CrossRef]
- Comandini, A.; Naro, C.; Adamo, R.; Akbar, A.N.; Lanna, A.; Bonmassar, E.; Franzese, O. Molecular mechanisms involved in HIV-1-Tat mediated inhibition of telomerase activity in human CD4(+) T lymphocytes. Mol. Immunol. 2013, 54, 181–92. [Google Scholar] [CrossRef]
- Stella-Ascariz, N.; Montejano, R.; Rodriguez-Centeno, J.; Alejos, B.; Schwimmer, C.; Bernardino, J.I.; Rodes, B.; Allavena, C.; Hoffmann, C.; Gisslén, M.; et al. Blood telomere length changes after ritonavir-boosted darunavir combined with raltegravir or tenofovir-emtricitabine in antiretroviral-naïve adults infected with HIV-1. J. Infect. Dis. 2018, 218, 1523–30. [Google Scholar] [CrossRef]
- Raffenberg, M.; Engel, T.; Schoepf, I.C.; Kootstra, N.A.; Reiss, P.; Braun, D.L.; Thorball, C.W.; Fellay, J.; Kouyos, R.D.; Ledergerber, B.; et al. Impact of delaying antiretroviral treatment during primary human immunodeficiency virus infection on telomere length. J. Infect. Dis. 2021, 224, 1775–84. [Google Scholar] [CrossRef]
- Montejano, R.; Stella-Ascariz, N.; Monge, S.; Bernardino, J.I.; Pérez-Valero, I.; Montes, M.L.; Valencia, E.; Martín-Carbonero, L.; Moreno, V.; González-Garcia, J.; et al. Impact of nucleos(t)ide reverse transcriptase inhibitors on blood telomere length changes in a prospective cohort of aviremic HIV-infected adults. J. Infect. Dis. 2018, 218, 1531–40. [Google Scholar] [CrossRef]
- Kaushal, S.; Landay, A.L.; Lederman, M.M.; Connick, E.; Spritzler, J.; Kuritzkes, D.R.; Kessler, H.; Levine, B.L.; St Louis, D.C.; June, C. H. Increases in T cell telomere length in HIV infection after antiretroviral combination therapy for HIV-1 infection implicate distinct population dynamics in CD4+ and CD8+ T cells. Clin. Immunol. 1999, 92, 14–24. [Google Scholar] [CrossRef]
- Hukezalie, K.R.; Thumati, N.R.; Côté, H.C.; Wong, J.M. In vitro and ex vivo inhibition of human telomerase by anti-HIV nucleoside reverse transcriptase inhibitors (NRTIs) but not by non-NRTIs. PLoS One 2012, 7. [Google Scholar] [CrossRef]
- Stella-Ascariz, N.; Montejano, R.; Pintado-Berninches, L.; Monge, S.; Bernardino, J.I.; Pérez-Valero, I.; Montes, M.L.; Mingorance, J.; Perona, R.; Arribas, J.R. Differential effects of tenofovir, abacavir, emtricitabine, and darunavir on telomerase activity in vitro. J. Acquir. Immune Defic. Syndr. 2017, 74, 91–4. [Google Scholar] [CrossRef]
- Leeansyah, E.; Cameron, P.U.; Solomon, A.; Tennakoon, S.; Velayudham, P.; Gouillou, M.; Spelman, T.; Hearps, A.; Fairley, C.; Smit, deV. ; et al. Inhibition of telomerase activity by human immunodeficiency virus (HIV) nucleos(t)ide reverse transcriptase inhibitors: a potential factor contributing to HIV-associated accelerated aging. J. Infect. Dis. 2013, 207, 1157–65. [Google Scholar] [CrossRef]
- Lombardi, F.; Sanfilippo, A.; Fabbiani, M.; Borghetti, A.; Ciccullo, A.; Tamburrini, E.; Di Giambenedetto, S. Blood telomere length gain in people living with HIV switching to dolutegravir plus lamivudine versus continuing triple regimen: a longitudinal, prospective, matched, controlled study. J. Antimicrob. Chemother. 2023, 9, 2315–2322. [Google Scholar] [CrossRef] [PubMed]
- Dalzini, A.; Ballin, G.; Dominguez-Rodriguez, S.; Rojo, P.; Petrara, M. R.; Foster, C.; Cotugno, N.; Ruggiero, A.; Nastouli, E.; Klein, N.; et al. Size of HIV-1 reservoir is associated with telomere shortening and immunosenescence in early-treated European children with perinatally acquired HIV-1. J. Int. AIDS Soc. 2021, 11, e25847. [Google Scholar] [CrossRef] [PubMed]
- Appay, V.; Sauce, D. Assessing immune aging in HIV-infected patients. Virulence 2017, 8, 529–38. [Google Scholar] [CrossRef]
- Jose, S.S.; Bendickova, K.; Kepak, T.; Krenova, Z.; Fric, J. Chronic inflammation in immune aging: role of pattern recognition receptor crosstalk with the telomere complex? Front. Immunol. 2017, 8, 1078. [Google Scholar] [CrossRef]
- Rodier, F.; Coppé, J.P.; Patil, C.K.; Hoeijmakers, W.A.; Muñoz, D.P.; Raza, S.R.; Freund, A.; Campeau, E.; Davalos, A.R.; Campisi, J. Persistent DNA damage signaling triggers senescence-associated inflammatory cytokine secretion. Nat. Cell Biol. 2009, 11, 973–9. [Google Scholar] [CrossRef]
- Van Epps, P.; Kalayjian, R.C. Human immunodeficiency virus and aging in the era of effective antiretroviral therapy. Infect. Dis. Clin. North Am. 2017, 31, 791–810. [Google Scholar] [CrossRef]
- Blanco, J.R.; Jarrin, I.; Martinez, A.; Siles, E.; Larrayoz, I.M.; Cañuelo, A.; Gutierrez, F.; Gonzalez-Garcia, J.; Vidal, F.; Moreno, S.; et al. Shorter telomere length predicts poorer immunological recovery in virologically suppressed HIV-1-infected patients treated with combined antiretroviral therapy. J. Acquir. Immune Defic. Syndr. 2015, 68, 21–9. [Google Scholar] [CrossRef]
- Shankarappa, R.; Gupta, P.; Learn Jr, G.H.; Rodrigo, A.G.; Rinaldo Jr, C.R.; Gorry, M.C.; Mullins, J.I.; Nara, P.L.; Ehrlich, G.D. Evolution of human immunodeficiency virus type 1 envelope sequences in infected individuals with differing disease progression profiles. Virology 1998, 241, 251–259. [Google Scholar] [CrossRef]
- Bailey, J.R.; Sedaghat, A.R.; Kieffer, T.; Brennan, T.; Lee, P.K.; Wind-Rotolo, M.; Haggerty, C.M.; Kamireddi, A.R.; Liu, Y.; Lee, J.; et al. Residual human immunodeficiency virus type 1 viremia in some patients on antiretroviral therapy is dominated by a small number of invariant clones rarely found in circulating CD4+ T cells. Journal of virology 2006, 80, 6441–6457. [Google Scholar] [CrossRef]
- Kearney, M.F.; Spindler, J.; Shao, W.; Yu, S.; Anderson, E.M.; O'Shea, A.; Rehm, C.; Poethke, C.; Kovacs, N.; Mellors, J.W.; et al. Lack of Detectable HIV-1 Molecular Evolution during Suppressive Antiretroviral Therapy. PLoS pathogens 2014, 10, e1004010. [Google Scholar] [CrossRef]
- Besson, G.J.; Lalama, C.M.; Bosch, R.J.; Gandhi, R.T.; Bedison, M.A.; Aga, E.; Riddler, S.A.; McMahon, D.K.; Hong, F.; Mellors, J.W. HIV-1 DNA decay dynamics in blood during more than a decade of suppressive antiretroviral therapy. Clin. Infect. Dis. 2014, 59, 1312–1321. [Google Scholar] [CrossRef] [PubMed]
- Pollack, R.A.; Jones, R.B.; Pertea, M.; Bruner, K.M.; Martin, A.R.; Thomas, A.S.; Capoferri, A.A.; Beg, S.A.; Huang, S.H.; Karandish, S.; et al. Defective HIV-1 Proviruses Are Expressed and Can Be Recognized by Cytotoxic T Lymphocytes, which Shape the Proviral Landscape. Cell Host Microbe, 2017, 21, 494–506. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.C.; Shan, L.; Hosmane, N.N.; Wang, J.; Laskey, S.B.; Rosenbloom, D.I.; Lai, J.; Blankson, J.N.; Siliciano, J.D.; Siliciano, R.F. Replication-competent noninduced proviruses in the latent reservoir increase barrier to HIV-1 cure. Cell 2013, 155, 540–551. [Google Scholar] [CrossRef] [PubMed]
- Musick, A.; Spindler, J.; Boritz, E.; Perez, L.; Crespo-Velez, D.; Patro, S.C.; Sobolewski, M.D.; Bale, M.J.; Reid, C.; Keele, B.F.; et al. HIV Infected T Cells Can Proliferate in vivo Without Inducing Expression of the Integrated Provirus. Front. Microbiol. 2019, 10, 2204. [Google Scholar] [CrossRef]
- Coffin, J.M.; Bale, M.J.; Wells, D.; Guo, S.; Luke, B.; Zerbato, J.M.; Sobolewski, M.D.; Sia, T.; Shao, W.; Wu, X.; et al. Integration in oncogenes plays only a minor role in determining the in vivo distribution of HIV integration sites before or during suppressive antiretroviral therapy. PLoS Pathog. 2021, 17, e1009141. [Google Scholar] [CrossRef]
- Einkauf, K.B.; Osborn, M.R.; Gao, C.; Sun, W.; Sun, X.; Lian, X.; Parsons, E.M.; Gladkov, G.T.; Seiger, K.W.; Blackmer, J.E.; et al. Parallel analysis of transcription, integration, and sequence of single HIV-1 proviruses. Cell 2022, 185, 266–282.e15. [Google Scholar] [CrossRef]
- Cole, B.; Lambrechts, L.; Gantner, P.; Noppe, Y.; Bonine, N.; Witkowski, W.; Chen, L.; Palmer, S.; Mullins, J.I.; Chomont, N.; et al. In-depth single-cell analysis of translation-competent HIV-1 reservoirs identifies cellular sources of plasma viremia. Nat. Commun. 2021, 12, 3727. [Google Scholar] [CrossRef]
- Lian, X.; Seiger, K.W.; Parsons, E.M.; Gao, C.; Sun, W.; Gladkov, G.T.; Roseto, I.C.; Einkauf, K.B.; Osborn, M.R.; Chevalier, J.M.; et al. Progressive transformation of the HIV-1 reservoir cell profile over two decades of antiviral therapy. Cell Host Microbe 2023, 31, 83–96.e5. [Google Scholar] [CrossRef]
- Matsui, T.; Leung, D.; Miyashita, H.; Maksakova, I.A.; Miyachi, H.; Kimura, H.; Tachibana, M.; Lorincz, M.C.; Shinkai, Y. Proviral silencing in embryonic stem cells requires the histone methyltransferase ESET. Nature 2010, 464, 927–931. [Google Scholar] [CrossRef]
- Schultz, D.C.; Ayyanathan, K.; Negorev, D.; Maul, G.G.; Rauscher, F.J. , 3rd. SETDB1: A novel KAP-1-associated histone H3, lysine 9-specific methyltransferase that contributes to HP1-mediated silencing of euchromatic genes by KRAB zinc-finger proteins. Genes Dev. 2002, 16, 919–932. [Google Scholar] [CrossRef]
- Halvas, E.K.; Joseph, K.W.; Brandt, L.D.; Guo, S.; Sobolewski, M.D.; Jacobs, J.L.; Tumiotto, C.; Bui, J.K.; Cyktor, J.C.; Keele, B.F.; et al. HIV-1 viremia not suppressible by antiretroviral therapy can originate from large T cell clones producing infectious virus. J. Clin. Invest. 2020, 11, 5847–5857. [Google Scholar] [CrossRef] [PubMed]
- Marini, B.; Kertesz-Farkas, A.; Ali, H.; Lucic, B.; Lisek, K.; Manganaro, L.; Pongor, S.; Luzzati, R.; Recchia, A.; Mavilio, F.; et al. Nuclear architecture dictates HIV-1 integration site selection. Nature, 2015; 521, 227–231. [Google Scholar] [CrossRef]
- Matreyek, K.A.; Yücel, S.S.; Li, X.; Engelman, A. Nucleoporin Nup153 phenylalanine-glycine motifs engage a common binding pocket within the HIV-1 capsid protein to mediate lentiviral infectivity. PLoS Pathog 2013, 9, e1003693. [Google Scholar] [CrossRef]
- Ocwieja, K.E.; Brady, T.L.; Ronen, K.; Huegel, A.; Roth, S.L.; Schaller, T.; James, L.C.; Towers, G.J.; Young, J.A.; Chanda, S.K.; et al. HIV integration targeting: a pathway involving transportin-3 and the nuclear pore protein RanBP2. PLoS Pathog 2011, 7, e1001313. [Google Scholar] [CrossRef]
- Koh, Y.; Wu, X.; Ferris, A.L.; Matreyek, K.A.; Smith, S.J.; Lee, K.; KewalRamani, V.N.; Hughes, S.H.; Engelman, A. Differential effects of human immunodeficiency virus type 1 capsid and cellular factors nucleoporin 153 and LEDGF/p75 on the efficiency and specificity of viral DNA integration. J. Virol. 2013, 87, 648–658. [Google Scholar] [CrossRef]
- Di Nunzio, F.; Fricke, T.; Miccio, A.; Valle-Casuso, J.C.; Perez, P.; Souque, P.; Rizzi, E.; Severgnini, M.; Mavilio, F.; Charneau, P.; et al. Nup153 and Nup98 bind the HIV-1 core and contribute to the early steps of HIV-1 replication. Virology 2013, 440, 8–18. [Google Scholar] [CrossRef]
- Achuthan, V.; Perreira, J.M.; Sowd, G.A.; Puray-Chavez, M.; McDougall, W.M.; Paulucci-Holthauzen, A.; Wu, X.; Fadel, H.J.; Poeschla, E.M.; Multani, A.S.; et al. Capsid-CPSF6 interaction licenses nuclear HIV-1 trafficking to sites of viral DNA integration. Cell Host Microbe 2018, 24, 392–404. [Google Scholar] [CrossRef]
- Briand, N.; Collas, P. Lamina-associated domains: peripheral matters and internal affairs. Genome Biol. 2020, 1, 85. [Google Scholar] [CrossRef]
- Alagna, N.S.; Thomas, T.I.; Wilson, K.L.; Reddy, K.L. Choreography of lamina-associated domains: structure meets dynamics. FEBS Lett. 2023 Nov;597(22):2806-2822. [CrossRef]
- Bester, S.M.; Wei, G.; Zhao, H.; Adu-Ampratwum, D.; Iqbal, N.; Courouble, V.V.; Francis, A.C.; Annamalai, A.S.; Singh, P.K.; Shkriabai, N.; et al. Structural and mechanistic bases for a potent HIV-1 capsid inhibitor. Science 2020, 6514, 360–364. [Google Scholar] [CrossRef]
- Emerson, R.O.; Thomas, J.H. Gypsy and the birth of the SCAN domain. J. Virol. 2011, 22, 12043–52. [Google Scholar] [CrossRef]
- Williams, A.J.; Blacklow, S.C.; Collins, T. The zinc finger-associated SCAN box is a conserved oligomerization domain. Mol. Cell. Biol. 1999, 19, 8526–8535. [Google Scholar] [CrossRef]
- Bellefroid, E.J.; Poncelet, D.A.; Lecocq, P.J.; Revelant, O.; Martial, J.A. The evolutionarily conserved Krüppel-associated box domain defined a subfamily of eu-karyotic multifingered proteins. Proc. Natl. Acad. Sci. U. S. A. 1991, 88, 3608–3612. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.; Skorupka, K.A.; Pak, A.J.; Ganser-Pornillos, B.K.; Pornillos, O.; Voth, G.A. TRIM5α self-assembly and compartmentalization of the HIV-1 viral capsid. Nat. Commun. 2020, 11, 1307–1316. [Google Scholar] [CrossRef]
- Jiang, C.; Lian, X.; Gao, C.; Sun, X.; Einkauf, K.B.; Chevalier, J.M.; Chen, S.M.Y.; Hua, S.; Rhee, B.; Chang, K.; et al. Distinct viral reservoirs in individuals with spontaneous control of HIV-1. Nature 2020, 585, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, N.; Jolicoeur, P. Retroviral pathogenesis. in Retrovirus, JM Coffin, SH Hughes, HE Varmus, Eds. (Cold Spring Harbor Laboratory Press, 1997), pp. 475–585.
- Neil, J.C.; Cameron, E.R. Retroviral insertion sites and cancer: fountain of all knowledge? Cancer Cell 2002, 4, 253–5. [Google Scholar] [CrossRef] [PubMed]
- McNally, G.A. HIV and Cancer: An Overview of AIDS-Defining and Non-AIDS-Defining Cancers in Patients With HIV. Clin. J. Oncol. Nurs. 2019, 3, 327–331. [Google Scholar] [CrossRef]
- Biggar, R.J.; Engels, E.A.; Frisch, M.; Goedert, J.J.; AIDS Cancer Match Registry Study Group. Risk of T-cell lymphomas in persons with AIDS. J. Acquir. Immune Defic. Syndr. 2001, 4, 371–6. [Google Scholar] [CrossRef]
- Herndier, B.G.; Shiramizu, B.T.; Jewett, N.E.; Aldape, K.D.; Reyes, G.R.; McGrath, M.S. ; Acquired immunodeficiency syndrome-associated T-cell lymphoma: Evidence for human immunodeficiency virus type 1-associated T-cell transformation. Blood 1992, 79, 1768–1774. [Google Scholar]
- Shiramizu, B.; Herndier, B.G.; McGrath, M.S. ; Identification of a common clonal human immunodeficiency virus integration site in human immunodeficiency virus-associated lymphomas. Cancer Res. 1994, 54, 2069–2072. [Google Scholar]
- Katano, H.; Sato, Y.; Hoshino, S.; Tachikawa, N.; Oka, S.; Morishita, Y.; Ishida, T.; Watanabe, T.; Rom, W.N.; Mori, S.; et al. Integration of HIV-1 caused STAT3-associated B cell lymphoma in an AIDS patient. Microbes Infect. 2007, 9, 1581–1589. [Google Scholar] [CrossRef]
- Tolomeo, M.; Cascio, A. The Multifaced Role of STAT3 in Cancer and Its Implication for Anticancer Therapy. Int. J. Mol. Sci. 2021, 2, 603. [Google Scholar] [CrossRef]
- Mellors, J.W.; Guo, S.; Naqvi, A.; Brandt, L.D.; Su, L.; Sun, Z.; Joseph, K.W.; Demirov, D.; Halvas, E.K.; Butcher, D.; Scott, B.; et al. Insertional activation of STAT3 and LCK by HIV-1 proviruses in T cell lymphomas. Sci. Adv. 2021, 42, eabi8795. [Google Scholar] [CrossRef]
- Almodovar, S.; Knight, R.; Allshouse, A.A.; Roemer, S.; Lozupone, C.; McDonald, D.; Widmann, J.; Voelkel, N.F.; Shelton, R.J.; Suarez, E.B.; et al. Human Immunodeficiency Virus nef signature sequences are associated with pulmonary hypertension. AIDS Res. Hum. Retrovir. 2012, 28, 607–618. [Google Scholar] [CrossRef] [PubMed]
- Musinova, Y.R.; Sheval, E.V.; Dib, C.; Germini, D.; Vassetzky, Y.S. Functional roles of HIV-1 Tat protein in the nucleus. Cell. Mol. Life Sci. 2016, 3, 589–601. [Google Scholar] [CrossRef]
- Manes, T.L.; Simenauer, A.; Geohring, J.L.; Flemming, J.; Brehm, M.; Cota-Gomez, A. The HIV-Tat protein interacts with Sp3 transcription factor and inhibits its binding to a distal site of the sod2 promoter in human pulmonary artery endothelial cells. Free Radic. Biol. Med. 2020, 147, 102–113. [Google Scholar] [CrossRef]
- Col, E.; Caron, C.; Chable-Bessia, C.; Legube, G.; Gazzeri, S.; Komatsu, Y.; Yoshida, M.; Benkirane, M.; Trouche, D.; Khochbin, S. HIV-1 Tat targets Tip60 to impair the apoptotic cell response to genotoxic stresses. EMBO J. 2005, 24, 2634–2645. [Google Scholar] [CrossRef]
- Zhang, S.M.; Song, M.; Yang, T.Y.; Fan, R.; Liu, X.D.; Zhou, P.K. HIV-1 tat impairs cell cycle control by targeting the Tip60, Plk1 and cyclin B1 ternary complex. Cell Cycle 2012, 11, 1217–1234. [Google Scholar] [CrossRef]
- Martina, C.; Fabienne, H.; Vesco, M.; Edwige, C.; Cécile, C.; Stefan, D.; Saadi, K. Control of the Histone-Acetyltransferase Activity of Tip60 by the HIV-1 Transactivator Protein, Tat. Biochemistry 1999, 38, 8826–8830. [Google Scholar]
- Sun, Y.; Jiang, X.; Chen, S.; Fernandes, N.; Price, B.D. A role for the Tip60 histone acetyltransferase in the acetylation and activation of ATM. Proc. Natl. Acad. Sci. USA 2005, 102, 13182–13187. [Google Scholar] [CrossRef]
- Baur, A. Functions of the HIV-1 Nef protein. Curr. Drug Targets Immune Endocr. Metabol. Disord. 2004, 4, 309–13. [Google Scholar] [CrossRef]
- Wang, T.; Green, L.A.; Gupta, S.K.; Kim, C.; Wang, L.; Almodovar, S.; Flores, S.C.; Prudovsky, I.A.; Jolicoeur, P.; Liu, Z.; et al. Transfer of intracellular HIV Nef to endothelium causes endothelial dysfunction. PLoS One. 2014, 3, e91063. [Google Scholar] [CrossRef]
- Wolf, D.; Witte, V.; Laffert, B.; Blume, K.; Stromer, E.; Trapp, S.; d'Aloja, P.; Schürmann, A.; Baur, A.S. HIV-1 Nef associated PAK and PI3-kinases stimulate Akt-independent Bad-phosphorylation to induce anti-apoptotic signals. Nat. Med. 2001, 11, 1217–24. [Google Scholar] [CrossRef] [PubMed]
- Rednor, S.J.; Ross, M.J. Molecular Mechanisms of Injury in HIV-Associated Nephropathy. Front. Med. (Lausanne). 2018, 7, 5–177. [Google Scholar] [CrossRef] [PubMed]
- Bruggeman, L.A.; Ross, M.D.; Tanji, N.; Cara, A.; Dikman, S.; Gordon, R.E.; Burns, G.C.; D'Agati, V.D.; Winston, J.A.; Klotman, M.E.; et al. Renal epithelium is a previously unrecognized site of HIV-1 infection. J. Am. Soc. Nephrol. 2000, 11, 2079–87. [Google Scholar] [CrossRef]
- Eitner, F.; Cui, Y.; Hudkins, K.L.; Stokes, M.B.; Segerer, S.; Mack, M.; Lewis, P.L.; Abraham, A.A.; Schlöndorff, D.; Gallo, G.; et al. Chemokine receptor CCR5 and CXCR4 expression in HIV-associated kidney disease. J. Am. Soc. Nephrol. 2000, 11, 856–867. [Google Scholar] [CrossRef]
- Ray, P.E.; Liu, X.H.; Henry, D.; Dye 3rd, L.; Xu, L.; Orenstein, J.M.; Schuztbank, T.E. Infection of human primary renal epithelial cells with HIV-1 from children with HIV-associated nephropathy. Kidney Int. 1998, 53, 1217–29. [Google Scholar]
- Chen, P.; Chen, B.K.; Mosoian, A.; Hays, T.; Ross, M.J.; Klotman, P.E.; Klotman, M.E. Virological synapses allow HIV-1 uptake and gene expression in renal tubular epithelial cells. J. Am. Soc. Nephrol. 2011, 22, 496–507. [Google Scholar] [CrossRef]
- Li, J.; Das, J.R.; Tang, P.; Han, Z.; Jaiswal, J.K.; Ray, P.E. Transmembrane TNF- facilitates HIV-1 infection of podocytes cultured from children with HIV-associated nephropathy. J. Am. Soc. Nephrol. 2016, 28, 862–75. [Google Scholar] [CrossRef]
- Ikura, T.; Tashiro, S.; Kakino, A.; Shima, H.; Jacob, N.; Amunugama, R.; Yoder, K.; Izumi, S.; Kuraoka, I.; Tanaka, K.; et al. DNA damage-dependent acetylation and ubiquitination of H2AX enhances chromatin dynamics. Mol. Cell. Biol. 2007, 20, 7028–40. [Google Scholar] [CrossRef]
- Prabhu, K.S.; Kuttikrishnan, S.; Ahmad, N.; Habeeba, U.; Mariyam, Z.; Suleman, M.; Bhat, A.A.; Uddin, S. H2AX: A key player in DNA damage response and a promising target for cancer therapy. Biomed. Pharmacother. 2024, 175, 116663. [Google Scholar] [CrossRef]
- Rosenstiel, P.E.; Chan, J.; Snyder, A.; Planelles, V.; D'Agati, V.D.; Klotman, P.E.; Klotman, M.E. HIV-1 Vpr activates the DNA damage response in renal tubule epithelial cells. AIDS 2009, 15, 2054–6. [Google Scholar] [CrossRef]
- Spudich, S.; González-Scarano, F. HIV-1-related central nervous system disease: current issues in pathogenesis, diagnosis, and treatment. Cold Spring Harb. Perspect. Med. 2012, 2, a007120. [Google Scholar] [CrossRef] [PubMed]
- Boisse, L.; Gill, M.J.; Power, C. HIV infection of the central nervous system: clinical features and neuropathogenesis. Neurol. Clin. 2008, 26, 799–819. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.; Power, C. Regulation of neural cell survival by HIV-1 infection. Neurobiol. Dis. 2012, 21, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Chopp, M.; Chan, P.H.; Hsu, C.Y.; Cheung, M.E.; Jacobs, T.P. DNA damage and repair in central nervous system injury: National Institute of Neurological Disorders and Stroke Workshop Summary. Stroke. 1996, 27, 363–9. [Google Scholar] [CrossRef]
- Madabhushi, R.; Pan, L.; Tsai, L.H. DNA damage and its links to neurodegeneration. Neuron 2014, 83, 266–282. [Google Scholar] [CrossRef]
- Madabhushi, R.; Gao, F.; Pfenning, A.R.; Pan, L.; Yamakawa, S.; Seo, J.; Rueda, R.; Phan, T.X.; Yamakawa, H.; Pao, P.C.; et al. Activity-induced DNA breaks govern the expression of neuronal early-response genes. Cell 2015, 161, 1592–605. [Google Scholar] [CrossRef]
- James, T.; Nonnemacher, M.R.; Wigdahl, B.; Krebs, F.C. Defining the roles for Vpr in HIV-1-associated neuropathogenesis. J. Neurovirol. 2016, 22, 403–15. [Google Scholar] [CrossRef]
- Crewe, M.; Madabhushi, R. Topoisomerase-mediated DNA damage in neurological disorders. Front Aging Neurosci. 2021, 13, 751742–59. [Google Scholar] [CrossRef]
- Madabhushi, R. The Roles of DNA Topoisomerase IIβ in Transcription. Int. J. Mol. Sci. 2018, 19, 1917. [Google Scholar] [CrossRef]
- McKinnon, P.J. DNA repair deficiency and neurological disease. Nat. Rev. Neurosci. 2009, 10, 100–112. [Google Scholar] [CrossRef]
- McKinnon, P.J. Maintaining genome stability in the nervous system. Nat. Neurosci. 2013, 16, 1523–1529. [Google Scholar] [CrossRef] [PubMed]
- Shull, E.R.; Lee, Y.; Nakane, H.; Stracker, T.H.; Zhao, J.; Russell, H.R.; Petrini, J.H.; McKinnon, P.J. Differential DNA damage signaling accounts for distinct neural apoptotic responses in ATLD and NBS. Genes Dev. 2009, 23, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Hollinger, A.; Cueni, N.; Marzolini, C.; Dickenmann, M.; Landmann, E.; Battegay, M.; Martinez, A. E,. Siegemund, M.; Leuppi-Taegtmeyer, A. Lactic acidosis and hyperlactatemia associated with lamivudine accumulation and sepsis in a kidney transplant recipient-a case report and review of the literature. AIDS Res. Ther. 2021, 1, 56. [Google Scholar]
- Oliveira, N.M.; Ferreira, F.A.; Yonamine, R.Y.; Chehter, E.Z. Antiretroviral drugs and acute pancreatitis in HIV/AIDS patients: is there any association? A literature review. Einstein (Sao Paulo). 2014, 1, 112–119. [Google Scholar] [CrossRef]
- Jao, J.; Wyatt, C.M. Antiretroviral medications: adverse effects on the kidney. Adv. Chronic Kidney Dis. 2010, 1, 72–82. [Google Scholar] [CrossRef]
- Hitchcock, A.M.; Kufel, W.D.; Dwyer, K.A.M.; Sidman, E.F. Lenacapavir: A novel injectable HIV-1 capsid inhibitor. Int. J. Antimicrob. Agents 2024, 1, 107009. [Google Scholar] [CrossRef]
- Engelman, A.N. HIV Capsid and Integration Targeting. Viruses 2021, 1, 125. [Google Scholar] [CrossRef]
- Mounzer, K.; Slim, J.; Ramgopal, M.; Hedgcock, M.; Bloch, M.; Santana, J.; Mendes, I.; Zhang, X.; Sklar, P.; Montezuma-Rusca, J.M.; et al. Efficacy and safety of bictegravir plus lenacapavir: 48-week outcomes in virologically suppressed people with HIV-1 on complex antiretroviral regimens at baseline. AIDS 2024, , Munich. Oral abstract OAB2602. 22–26 July.
- Mounzer, K. , Slim, J.; Ramgopal, M.; Hedgcock, M.; Bloch, M.; Santana, J.; Mendes, I.; Guo, Y.; Arora, P.; Montezuma-Rusca, J.M. et al. Phase 2 study of switch to daily BIC + LEN in individuals on a complex HIV treatment regimen. CROI 2024, Denver.
- Mounzer, K. , Slim, J.; Ramgopal, M., Hedgcock, M., Bloch, M., Santana, J., Mendes, I., Guo, Y., Arora, P., Eds.; Montezuma-Rusca, J.M. et al. Efficacy and Safety of Switching to Daily Bictegravir Plus Lenacapavir From a Complex Human Immunodeficiency Virus Treatment Regimen: A Randomized, Open-Label, Multicenter Phase 2 Study (ARTISTRY-1). Clin. Infect. Dis. 2024. [Google Scholar]
- Doan, J.; Brunzo-Hager, S.; Satterly, B.; Cory, T.J. Expanding therapeutic options: lenacapavir + bictegravir as a potential treatment for HIV. Expert Opin. Pharmacother. 2023, 18, 1949–1956. [Google Scholar] [CrossRef]
- Pak, A.J.; Grime, J.M.A.; Yu, A.; Voth, GA. Off-pathway assembly: a broad-spectrum mechanism of action for drugs that undermine controlled HIV-1 viral capsid formation. J. Am. Chem. Soc. 2019, 141, 10214–10224. [Google Scholar] [CrossRef]
- Gruell, H.; Gunst, J.D.; Cohen, Y.Z.; Pahus, M.H.; Malin, J.J.; Platten, M.; Millard, K.G.; Tolstrup, M.; Jones, R.B.; Conce Alberto, W.D.; et al. Effect of 3BNC117 and romidepsin on the HIV-1 reservoir in people taking suppressive antiretroviral therapy (ROADMAP): a randomised, open-label, phase 2A trial. Lancet Microbe 2022, 3, e203–e214. [Google Scholar] [CrossRef]
- Maxwell, J.W.; Falcinelli, S.D.; Nefedov, A.; Dorfmeier, C.; Wu, G.; Dewey, M.; Webber, A.L.; Archin, N.M.; Margolis, D.M.; Hazuda, D.J.; et al. Cellular Gene Modulation of HIV-Infected CD4 T Cells in Response to Serial Treatment with the Histone Deacetylase Inhibitor Vorinostat. J. Virol. 2020, 13, e00351–20. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekar, A.P.; Badley, A.D. Prime, shock and kill: BCL-2 inhibition for HIV cure. Front. Immunol. 2022, 13, 1033609. [Google Scholar] [CrossRef] [PubMed]
- Ahlenstiel, C.L.; Symonds, G.; Kent, S.J.; Kelleher, A.D. Block and Lock HIV Cure Strategies to Control the Latent Reservoir. Front. Cell. Infect. Microbiol. 2020, 10, 424. [Google Scholar] [CrossRef]
- Mousseau, G.; Kessing, C.F.; Fromentin, R.; Trautmann, L.; Chomont, N.; Valente, S.T. The Tat Inhibitor Didehydro-Cortistatin A Prevents HIV-1 Reactivation From Latency. mBio, 2015, 4, e00465. [Google Scholar] [CrossRef]
- Li, C.; Mousseau, G.; Valente, S.T. Tat Inhibition by Didehydro-Cortistatin A Promotes Heterochromatin Formation at the HIV-1 Long Terminal Repeat. Epigenet. Chromatin 2019, 1, 23. [Google Scholar] [CrossRef]
- Kessing, C.F.; Nixon, C.C.; Li, C.; Tsai, P.; Takata, H.; Mousseau, G.; Ho, P.T.; Honeycutt, J.B.; Fallahi, M.; Trautmann, L.; et al. In Vivo Suppression of HIV Rebound by Didehydro-Cortistatin A, a "Block-And-Lock" Strategy for HIV-1 Treatment. Cell Rep. 2017, 3, 600–11. [Google Scholar] [CrossRef]
- Bhowmik, R.; Chaubey, B. CRISPR/Cas9: a tool to eradicate HIV-1. AIDS Res. Ther. 2022, 1, 58. [Google Scholar] [CrossRef]
- Dash, P.K.; Chen, C.; Kaminski, R.; Su, H.; Mancuso, P.; Sillman, B.; Zhang, C.; Liao, S.; Sravanam, S.; Liu, H.; et al. CRISPR editing of CCR5 and HIV-1 facilitates viral elimination in antiretroviral drug-suppressed virus-infected humanized mice. Proc. Natl. Acad. Sci. U. S. A. 2023, 19, e2217887120. [Google Scholar] [CrossRef]
- Herskovitz, J.; Hasan, M.; Patel, M.; Blomberg, W.R.; Cohen, J.D.; Machhi, J.; Shahjin, F.; Mosley, R.L.; McMillan, J.; Kevadiya, B.D.; et al. CRISPR-Cas9 Mediated Exonic Disruption for HIV-1 Elimination. EBioMedicine 2021, 73, 103678. [Google Scholar] [CrossRef]
- Cohrt, K.O., News. Excision’s EBT-101 demonstrates safety in clinical trial but does not cure HIV - The 2nd CRISPR Medicine Conference 2025, -11, Copenhagen, Denmark. 7 April.
- Gupta, R.K.; Peppa, D.; Hill, A.L.; Gálvez, C.; Salgado, M.; Pace, M.; McCoy, L.E.; Griffith, S.A.; Thornhill, J.; Alrubayyi, A. Evidence for HIV-1 cure after CCR5Δ32/Δ32 allogeneic haemopoietic stem-cell transplantation 30 months post analytical treatment interruption: a case report. The Lancet HIV 2020, 7, 5–e340. [Google Scholar] [CrossRef]
- Hütter, G.; Nowak, D.; Mossner, M.; Ganepola, S.; Müßig, A.; Allers, K.; Schneider, T.; Homann, J.; Kücherer, C.; Blau, O. "Long-term remission and HIV cure in a patient with a CCR5-delta32/Delta32 mutation after stem cell transplantation. " New England Journal of Medicine 2009, 360, 692–698. [Google Scholar] [PubMed]
- Li, J.Z.; Blankson, J.N. How elite controllers and posttreatment controllers inform our search for an HIV-1 cure. J. Clin. Invest. 2021, 11, e149414. [Google Scholar]
- Derbalah, A.; Karpick, H.C.; Maize, H.; Skersick, P.; Cottrell, M.; Rao, G.G. Role of islatravir in HIV treatment and prevention: an update. Curr. Opin. HIV AIDS 2022, 4, 240–246. [Google Scholar] [CrossRef]
Figure 1.
The viral integrase and Ku70 complex recruits the Ku80, DNA-PK, and ATM at the integration site. ATM phosphorylates CHK2 and H2AX, two key proteins that initiate activation of the DNA damage checkpoint. CHK2 inhibits CDC25, preventing cell entry into the S and M phases. The phosphorylated form of H2AX is a scaffold that attracts repair factors to DNA damage sites and retains them until the damage is removed.
Figure 1.
The viral integrase and Ku70 complex recruits the Ku80, DNA-PK, and ATM at the integration site. ATM phosphorylates CHK2 and H2AX, two key proteins that initiate activation of the DNA damage checkpoint. CHK2 inhibits CDC25, preventing cell entry into the S and M phases. The phosphorylated form of H2AX is a scaffold that attracts repair factors to DNA damage sites and retains them until the damage is removed.
Figure 2.
Schematic representation of the complex interactions between HIV-1 and the host cell genome. The factors that promote virus production are: i) integration into nuclear speckles; ii) integration in the same orientation as host genes; iii) integration within genes marked by H3K36me3, H3K9ac, or H3K4me3 epigenetic histone modifications. Integration into heterochromatin, in the opposite orientation to host genes, or in genes marked by H3K27 or H3K9 histone epigenetic modifications, promotes viral latency. Integration and Vpr can cause DNA damage, which is not repaired due to PI3K/ATM suppression by Vif. Replication pressure caused by HIV-1-induced CD4+ T cell depletion, PI3K/ATM suppression, DNA damage, reduced telomerase levels, and treatment with NRTIs leads to progressive telomere shortening. Telomere shortening is a cause of CD4+ T cell depletion and, together with DNA damage, contributes to inflammaging.
Figure 2.
Schematic representation of the complex interactions between HIV-1 and the host cell genome. The factors that promote virus production are: i) integration into nuclear speckles; ii) integration in the same orientation as host genes; iii) integration within genes marked by H3K36me3, H3K9ac, or H3K4me3 epigenetic histone modifications. Integration into heterochromatin, in the opposite orientation to host genes, or in genes marked by H3K27 or H3K9 histone epigenetic modifications, promotes viral latency. Integration and Vpr can cause DNA damage, which is not repaired due to PI3K/ATM suppression by Vif. Replication pressure caused by HIV-1-induced CD4+ T cell depletion, PI3K/ATM suppression, DNA damage, reduced telomerase levels, and treatment with NRTIs leads to progressive telomere shortening. Telomere shortening is a cause of CD4+ T cell depletion and, together with DNA damage, contributes to inflammaging.
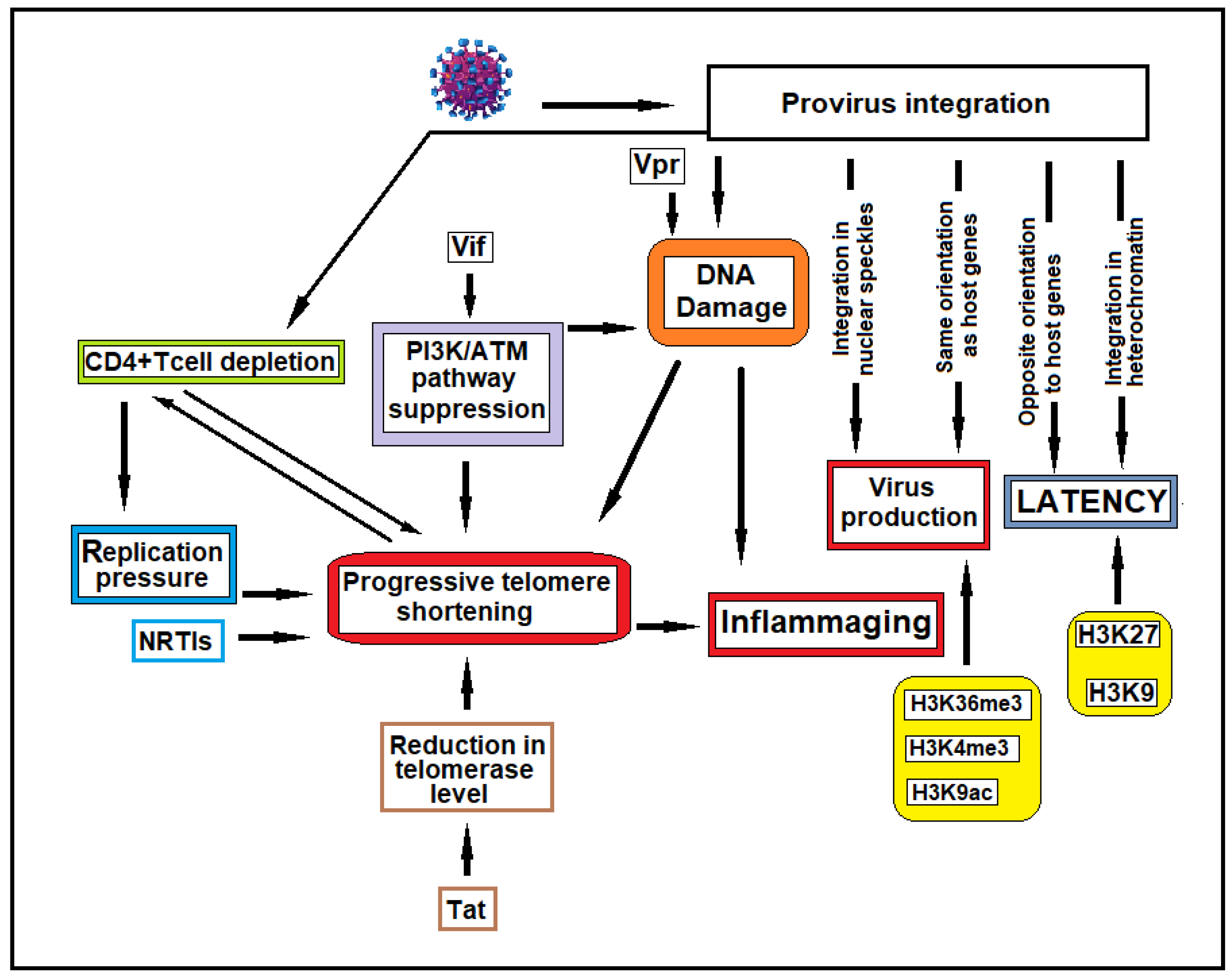
Figure 3.
Vpr unwinds double-stranded DNA by converting it into a relaxed form and induces the ubiquitination of histone H2B. This chromatin remodeling promotes efficient loading of RPA70 that activates the ATR-dependent DDR. In addition, Vpr-induced unwinding of double-stranded DNA results in the accumulation of negatively supercoiled DNA and covalent complexes of topoisomerase 1 and DNA, which cause DSBs. Vpr promotes the degradation of several DDR proteins, inhibiting double-strand DNA break repair by recruiting the CRL4ADCAF1 ubiquitin ligase complex. Vif blocks Vpr-directed activation of ATM but not ATR.
Figure 3.
Vpr unwinds double-stranded DNA by converting it into a relaxed form and induces the ubiquitination of histone H2B. This chromatin remodeling promotes efficient loading of RPA70 that activates the ATR-dependent DDR. In addition, Vpr-induced unwinding of double-stranded DNA results in the accumulation of negatively supercoiled DNA and covalent complexes of topoisomerase 1 and DNA, which cause DSBs. Vpr promotes the degradation of several DDR proteins, inhibiting double-strand DNA break repair by recruiting the CRL4ADCAF1 ubiquitin ligase complex. Vif blocks Vpr-directed activation of ATM but not ATR.
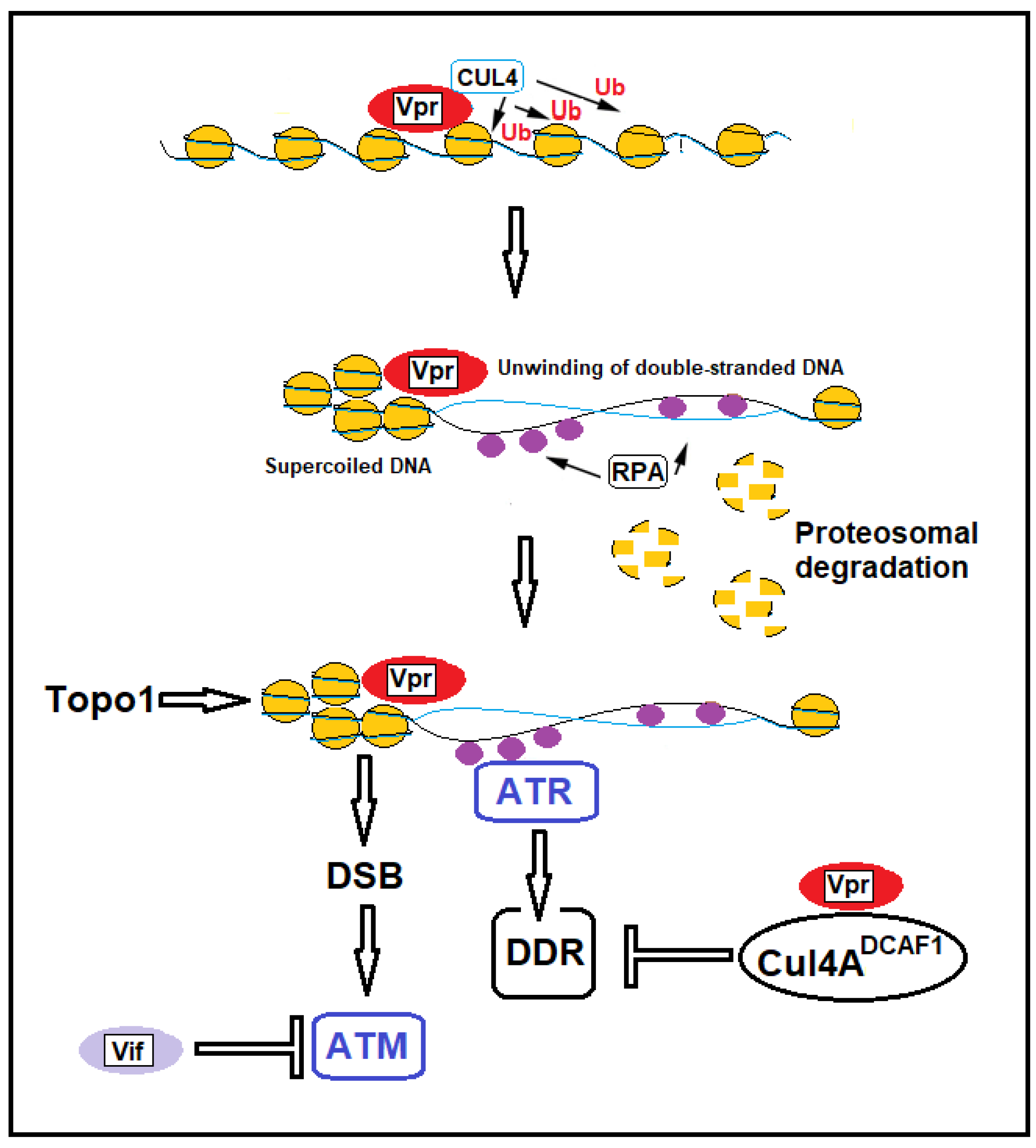
Figure 4.
CPSF6 plays a predominant role in directing HIV-1 integration into gene-dense regions of euchromatin. Knockout or knockdown of the host factor CPSF6, or treatment with Lenacapavir, results in a significant preference for HIV-1 provirus integration into LADs. The repressive chromatin environment of LADs may help maintain the provirus in a silent state.
Figure 4.
CPSF6 plays a predominant role in directing HIV-1 integration into gene-dense regions of euchromatin. Knockout or knockdown of the host factor CPSF6, or treatment with Lenacapavir, results in a significant preference for HIV-1 provirus integration into LADs. The repressive chromatin environment of LADs may help maintain the provirus in a silent state.

Figure 5.
A) Long-term ART-treated individuals are characterized by large clones of intact proviruses preferentially integrated in heterochromatin locations. B) A theoretical model in which long-term treatment with ART including capsid inhibitors could lead to the clonal expansion of CD4+ T cells harbouring proviruses integrated into LADs and silenced therein.
Figure 5.
A) Long-term ART-treated individuals are characterized by large clones of intact proviruses preferentially integrated in heterochromatin locations. B) A theoretical model in which long-term treatment with ART including capsid inhibitors could lead to the clonal expansion of CD4+ T cells harbouring proviruses integrated into LADs and silenced therein.
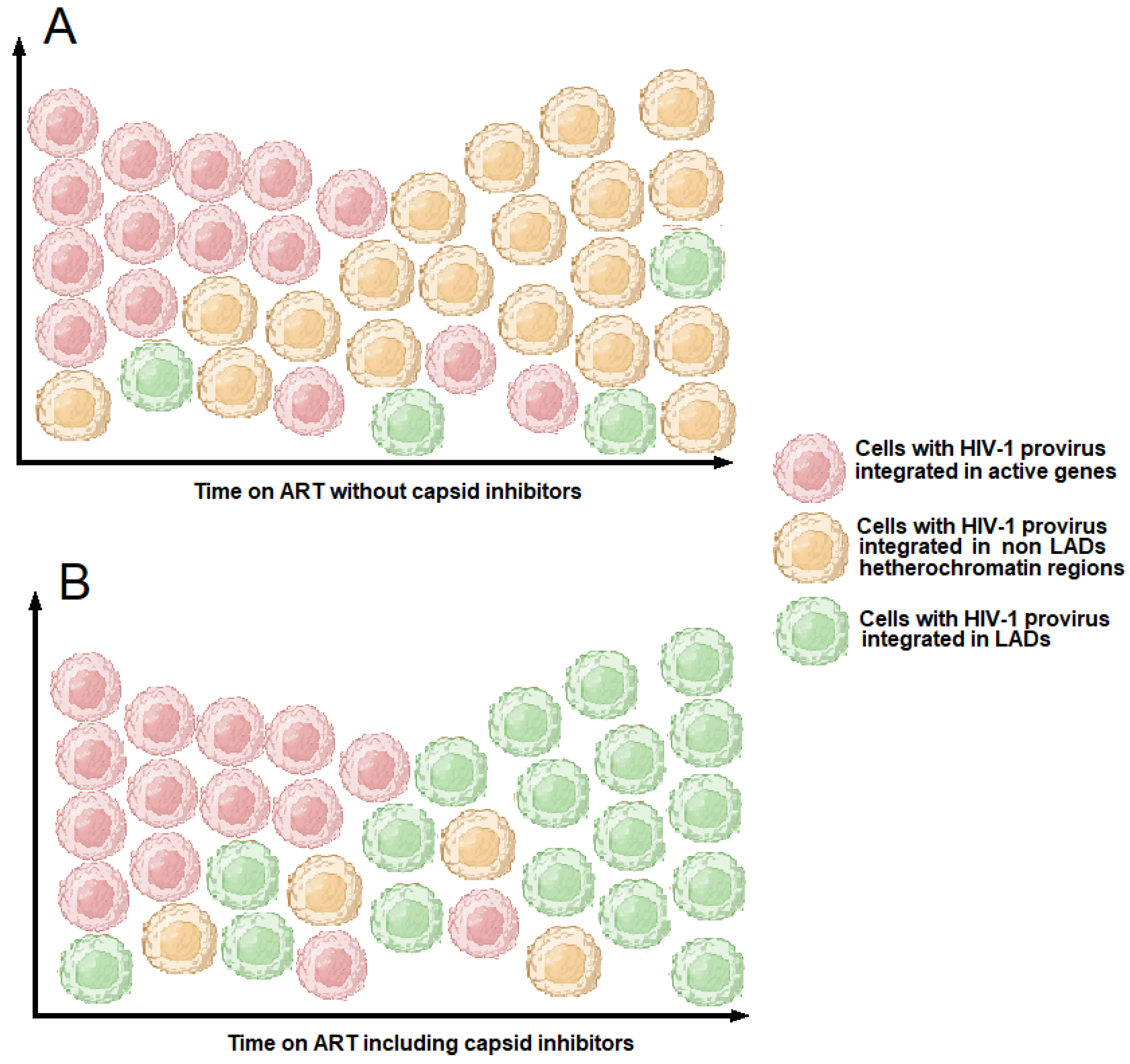
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



