
Submitted:
17 March 2025
Posted:
24 March 2025
You are already at the latest version
Abstract
Background Antibiotic tolerance is associated with failure of antibiotic treatment and accelerates the development of antimicrobial resistance. The molecular mechanisms underlying antimicrobial tolerance remain, poorly understood. In some instances, tolerant bacteria can slow metabolism by extending the lag phase without altering antimicrobial susceptibility. We recently induced ceftriaxone (CRO) tolerance in the N. gonorrhoeae reference strain WHO P. In the current study, we characterized the transcriptomic profiles of these CRO-tolerant phenotypes. Methods To induce tolerance, WHO P strains were grown under three-hour intermittent CRO exposure (10× the MIC), followed by overnight growth in GC broth for seven consecutive days, with cultures maintained in sextuplicate. Two control cultures were maintained without CRO exposure. The tolerance and CRO susceptibility of the isolates were assessed using a modified Tolerance Disc (TD) test. Total RNA was isolated from tolerant isolates (n=12) and control (n = 3) strains, followed by Ribo depletion. Library preparation was carried out using the Stranded TruSeq RNA library preparation kit. Sequencing was performed on a NextSeq6000, v2. 1 × 150 bp (Illumina Inc., San Diego, CA, USA), and data analysis was performed using the CLC Genomics Workbench v22 (clcbio, Denmark). Results Transcriptomic analysis revealed no differentially expressed genes at the first time point that was assessed (after 1 day of CRO exposure). However, after 3 days of CRO exposure, 13 genes were found to be significantly downregulated, including tRNA-Ser (C7S06_RS03100) and tRNA-Leu (C7S06_RS04945) and ribosomal RNA genes (16S and 23S rRNA). Following 7 days of exposure, 51 genes were differentially expressed with most downregulated, such as SecB (Protein-export chaperone SecB) and tRNA-Ser (C7S06_RS01850) and the 16S and 23S ribosomal RNA genes. Conclusions The development of CRO-tolerance in N. gonorrhoeae was associated with the downregulation of various ribosomal genes and associated genes, reflecting a potential mechanism for bacterial survival under antibiotic stress.
Keywords:
tolerant
; RNA-Seq
; ceftriaxone
; WHO P
; Neisseria gonorrhoeae
; transcriptome
Introduction
Antibiotic tolerance poses a significant challenge in clinical settings and is increasingly recognized as a significant factor associated with treatment failure and recurrent infections [1]. Unlike resistance, where there is an increase in the minimum inhibitory concentration (MIC) of antibiotics [2,3,4], tolerance enables bacteria to acquire the ability to survive transient exposure to high antibiotic concentrations through adaptation without an increase in the MIC. One way they do this is by slowing down their metabolism by extending the lag phase [5,6,7,8,9,10,11]. The emergence of tolerance has been shown to facilitate the emergence of antimicrobial resistance (AMR) [11]. Understanding the mechanisms responsible for tolerance could thus assist with the development of not only novel treatment approaches to prevent treatment failures but also the emergence of AMR [11,12].
Antibiotic tolerance has been observed in numerous bacterial species. Although the mechanisms underpinning tolerance vary between different species they have frequently been found to include alterations in gene expression, changes in cell envelope properties, metabolic slowdown, activation of stress response pathways, and the formation of persister cells [13,14]. Antibiotic tolerance has been reported in clinically relevant pathogens such as Staphylococcus aureus, Escherichia coli, Pseudomonas aeruginosa, Streptococcus pneumoniae, and Mycobacterium spp. [13,15,16,17]. For instance, S. aureus can survive antibiotic exposure by forming persister cells that regrow, leading to recurrent infections [18], while M. tuberculosis enters a dormant state characterized by changes such as a dramatic downregulation of rRNA and ribosomal protein synthesis [19,20]. Upregulation of relA gene was seen in ciprofloxacin-tolerant S. aureus [21], and deletion of the sodium-proton antiporter gene nhaA, resulted in downregulated metabolism and upregulated stress responses in E.coli [22]. In Mycobacterium abscessus, the cytochrome bdtype quinol oxidase contributes to clofazimine tolerance [23]. These diverse mechanisms highlight the complexity of antibiotic tolerance across bacterial species and underscore the need for species-specific approaches in addressing this clinical challenge.
Resistance to ceftriaxone, the first-line treatment for gonorrhea - a sexually transmitted infection (STIs) caused by the Gram-negative bacterium Neisseria gonorrhoeae - has become a significant concern. Recent studies have shown that tolerance to ceftriaxone, can be induced in N. gonorrhoeae through cyclic antibiotic exposure [24]. Mutations in four genes that may contribute to ceftriaxone tolerance in N. gonorrhoeae, including mutation in the enolase (eno) gene, have been identified [25]. Of interest, the prevalence of antibiotic tolerance in N. gonorrhoeae varies by anatomical site, with tolerance to azithromycin being more common in anorectal than urogenital infections [26]. Gene expression studies have revealed several transcriptional regulators involved in bacterial antibiotic tolerance. For instance, marA, soxS, or robA systems in Escherichia coli have been implicated in mediating the induction of multi-drug tolerance [27,28]. No transcriptomic studies are available that provide insights into the mechanisms underlying ceftriaxone tolerance in N. gonorrhoeae.
This study aimed to provide the transcriptomic profile of in vitro-generated ceftriaxone tolerance strains of N. gonorrhoeae using RNA-Seq. We aimed to identify gene expression associated with the ceftriaxone-tolerant phenotype.
Materials and Methods
Induction of CRO Tolerance, Assessment of Tolerance and CRO Susceptibility
Previously, we induced ceftriaxone (CRO) tolerance in a highly susceptible N. gonorrhoeae CRO reference strain, WHO P, with a CRO MIC of 0.004 µg/ml [24]. Briefly, the overnight culture of WHO P reference colonies was suspended in Gonococcal (GC) broth and the turbidity was adjusted to 0.5–1.0 McFarland. These suspensions were then exposed to a high concentration of CRO at 10 times the MIC (0.04 µg/mL) for 3 hours in a cyclic manner. After each cycle, the cultures were resuspended in GC broth containing 0.04 µg/mL CRO, followed by washing the bacterial cells to remove antibiotics, and were incubated overnight at 36 oC in a 6% CO2 incubator. The experiment was conducted over seven consecutive days with six biological replicates and two control replicates that did not receive antibiotic exposure. The CRO MIC was determined after each exposure cycle using a gradient E-test, and samples were stored at -80 °C in skim milk with 30% glycerol.
To detect tolerant phenotypes, a modified TD test was used, wherein the colonies were inoculated on BDTM Columbia Agar with 5% Sheep Blood, followed by incubation and sequential exposure to discs with CRO (antibiotic discs with CRO conc. of 0.04 µg/mL) and discs with GC broth (nutrient discs) [24]. Tolerant colonies that emerged after 48 hours were further inoculated on fresh blood agar plates, harvested in RNA Shield (Zymo Research, Netherlands), snap-freezed and outsourced to Eurofins Genomics (Germany) for RNA isolation and sequencing.
RNA-Sequencing and Bioinformatic Analyses
A total of 12 tolerant isolates from four lineages evolved with CRO antibiotic, and three control isolates evolved without CRO were subjected to RNA-Seq (Table 1). The tolerant isolates included 3 isolates each from 3 time points and 4 lineages (after days 1, 3 and 7 of CRO exposure) (Table 1). Total RNA was isolated from a total of 15 isolates, followed by ribodepletion to remove ribosomal RNA. The Stranded TruSeq RNA Library Preparation Kit was used for cDNA library preparation. The library was then sequenced on a NextSeq6000, v2, 1 × 150 bp platform (Illumina Inc). Data analysis was conducted using CLC Genomics Workbench v22 (Clcbio, Denmark), according to Liu and Di 2020. Differential gene expression (DGE) analysis was performed to identify genes that showed significant differences in expression using Benjamini-Hochberg multiple testing correction using a false discovery rate of 0.05. The analysis also included controlling for time points across all groups to assess the differential expression due to CRO exposure. The raw reads generated are deposited at NCBI accession PRJNA1223718.
Results
In this study, a comparative transcriptomic analysis was carried out to identify differentially expressed genes (DEGs) between control (n=3) and tolerant isolates (n=12) across 4 lineages and 3 time points. The experimental conditions were as follows (Table 1):
- i.
- Condition 1: 10.2 (control sample) vs 10.3, 10.4, 10.7, 10.8 (tolerant samples) isolates from day 1 CRO-exposure
- ii.
- Condition 2: 12.1 (control sample) vs. 12.3.3, 12.4.3, 12.7.3, and 12.8.3 (tolerant samples) isolates from day 3 CRO-exposure
- iii.
- Condition 3: 16.1 (control sample) vs. 16.3.-2.3, 16.4.3, 16.7-2.3, and 16.8-2.3. (tolerant samples) isolates from day 7 CRO-exposure
- iv.
- Condition 4: all the control samples (n=3) vs. all the tolerant samples (n=12)
In condition 1, no DEGs were identified.
In condition 2, we identified 13 DEGs, which were significantly downregulated in the tolerant isolates (Figure 1 and Table 2). Notable DEGs included C7S06_RS03100 (tRNA-Ser) and C7S06_RS04945 (tRNA-Leu), with log2 fold changes of -7.40 and -8.83, respectively. Several ribosomal RNA genes, including 4 copies each of 23S ribosomal RNA (C7S06_RS07095, C7S06_RS09345, C7S06_RS09850, C7S06_RS11315) and and 16S ribosomal RNA (C7S06_RS07110, C7S06_RS09360, C7S06_RS09865, C7S06_RS11300) and ssrS (6S RNA), showed significant downregulation (Figure 1 and Table 2).
In Condition 3, a total of 51 DEGs were identified, and most were predominantly downregulated. Significant downregulation was observed in genes such as secB (Protein-export chaperone SecB and C7S06_RS01850 (tRNA-Ser), with log2 fold changes of -1.85 and -7.99, respectively. The downregulated genes also included 4 copies of 23S rRNA, 6S RNA, tRNAs (n=7), hypothetical protein (n=5), and pilin (Figure 2 and Table 2). Out of the 51 DEGs, eight genes were upregulated with log2 fold changes ranging between 1.13 to 2.03. These included specific transporter genes such as cysT (sulfate ABC transporter permease subunit, glnD ([protein-PII] uridylyltransferase, C7S06_RS05355 (sulfate ABC transporter substrate-binding protein, C7S06_RS07485 (PepSY domain-containing protein, C7S06_RS09460 (chloride channel protein, two amino acid ABC transporter permeases (C7S06_RS10580 and C7S06_RS10585) and C7S06_RS12240 (MFS transporter).
Discussion
Previously, we reported that tolerance to CRO can be induced in a CRO-susceptible (<0.004 µg/mL) N. gonorrhoeae WHO P reference isolate [24]. In the current analysis, we describe the differentially expressed genes using RNA-Seq for CRO-tolerant isolates (n=12). The DEGs identified in the various conditions underscore the complexity of bacterial adaptation during ceftriaxone tolerance, and the results provide significant insights into the molecular mechanisms underlying ceftriaxone tolerance in N. gonorrhoeae.
No DEGs were identified in condition 1, when isolates had been only been exposed to CRO for one day, suggesting that early response to antibiotic exposure may not involve significant transcriptional changes. In condition 2, following three days of CRO exposure, 13 DEGs were identified, all of which were significantly downregulated. Downregulation of tRNA genes (e.g., tRNA-Ser, tRNA-Leu), 16S and 23S ribosomal RNA genes suggests a decrease in overall bacterial protein synthesis. The abundance of individual tRNA varies with growth rate and reflects the codon frequencies in the corresponding mRNA pools [30]. However, when translation demand decreases due to stress reponse—e.g. reduced mRNA availability—tRNAs become susceptible to degradation by ribonucleases, thus preventing the accumulation of uncharged tRNAs [31]. Thus, the downregulation of tRNAs in CRO-tolerant N. gonorrhoeae could be a survival strategy where the bacteria can optimize their metabolic activity to survive under high antibiotic pressure.
Bacterial ssrS (6S RNA), a small non-coding RNA that is conserved in all bacteria, inhibits transcription via direct binding to RNA polymerase holoenzymes [32,33]. 6S RNA has been shown to be a key player in stress responses; for example, in E.coli, it is involved in oxidative stress response [34,35]. Deletion of 6S RNA in Bacillus thuringiensis, results in impaired growth in the stationary phase [36]. Downregulation of ssrS in conditions 2 to 4 may thus be compatible with the hypothesis that N. gonorrhoeae reduces its overall transcriptional activity in response to intermittent antimicrobial exposure.
In condition 3, after seven days of CRO exposure, the transcriptomic landscape revealed 51 DEGs, indicating a more extensive bacterial response. The downregulation of additional genes, including secB (Protein-export chaperone SecB), suggests less need for these systems during stress as chaperones play important roles in withstanding stress associated with intracellular survival [37,38]. Interestingly, in condition 3, the upregulation of specific transporter genes, such as cysT (Sulfate ABC transporter permease subunit) and glnD ([protein-PII] uridylyltransferase), was observed. Microorganisms require sulfur for growth, and in E. coli and Salmonella typhimurium, the ABC-type sulfate transporter, which is responsible for the transport of inorganic sulfate into microbial cells, consists of the periplasmic sulfate binding protein (Sbp) which interacts with the permease components CyST and CysW [39]. The upregulation of cysT in N. gonorrhoeae suggests that it helps maintain sulfur homeostasis, which is vital for bacterial survival and adaptation. The primary sensor in many bacterial nitrogen signal transduction (NSR) systems is the GlnD protein, a large multidomain uridylyltransferase and uridylyl cleavage enzyme whose primary uridylylation targets are the PII proteins, GlnB and GlnK [40]. The uridylylation state of these proteins is regulated by α-ketoglutarate and glutamine so that when GlnD senses that there is not enough nitrogen available to synthesize adequate glutamine, it activates the NSR by uridylylating the PII proteins [41–43]. Thus, the upregulation of glnD may reflect GlnD’s uridylylation of GlnB and GlnK, which in turn regulates the expression of genes involved in nitrogen uptake and assimilation.
In condition 4, where all the control samples (n=3) were compared to all the tolerant samples (n=12) across multiple time points, 10 DEGs were consistently identified, including downregulated tRNA genes and ssrS (6S RNA). This consistent downregulation across different conditions and time points suggests that these genes are central to the bacterial response to CRO exposure and may represent core components of the tolerance mechanism. Of note, two hypothetical proteins with no assigned function were identified, illustrating how much remains to be learned about tolerance in Neisseria spp.
The caveats of the study include the small sample sizes and the fact that the transcriptomic data were derived from in vitro experiments; as such, the findings may not entirely reflect the gene expression patterns that occur in vivo [20]. Finally, the observed downregulation of ribosomal genes could be due to the expression bias introduced during the ribodepletion of rRNA. Future directions should involve using in vivo animal models or clinical isolates from patients to validate the relevance of the identified DEGs. This is particularly important given studies that have found that tolerance facilitates the emergence of AMR [11]and our recent finding that a higher proportion of anorectal than urethral N. gonorrhoeae infections were tolerant [26]. Tolerance may thus partially explain why a higher proportion of anorectal infections are asymptomatic [26]. Gene knockout experiments need to be conducted to validate the roles of the identified DEGs in ceftriaxone tolerance.
In conclusion, this study provides valuable insights into the molecular basis of antibiotic tolerance. In general, tolerance was associated with the downregulation of central protein synthesis and later upregulation of genes encoding transporter proteins. Further research could evaluate if similar changes occur in human infections. It will also be important to assess if the probability and timing of emergence of tolerance varies by anatomical site of infection and exposure to various antimicrobials. We have not assessed the prevalence of tolerance in pharyngeal infections. This will be useful to do as infections at this site are more difficult to eradicate and have been frequently implicated in the emergence of antimicrobial resistance [44,45].
Author Contributions
C.K. and S.S.M.-B. conceptualized the study. M.B, Z.G and S.A performed the experiments. S.S.M-B performed the data analysis and wrote the first draft of the manuscript. S.S.M.-B. and C.K. interpreted the data. All the authors reviewed the manuscript.
Funding
This research was funded by SOFI 2021 grant: “Preventing the Emergence of untreatable STIs via radical Prevention” (PRESTIP).
Conflicts of Interest
None
References
- Westblade, L.F.; Errington, J.; Dörr, T. Antibiotic tolerance. PLoS Pathog 2020, 16, e1008892. [Google Scholar] [CrossRef] [PubMed]
- Handwerger, S.; Tomasz, A. Antibiotic tolerance among clinical isolates of bacteria. Rev Infect Dis 1985, 7, 368–386. [Google Scholar] [CrossRef] [PubMed]
- Eric, M.; Pratt, W.B. The antimicrobial drugs; Eric M. Scholar, William B. Pratt., Second edi.; Oxford University Press: New York, 2000. [Google Scholar]
- Andrews, J.M. Determination of minimum inhibitory concentrations. J Antimicrob Chemother 2001, 48 (Suppl 1), 5–16. [Google Scholar] [CrossRef]
- Kotková, H.; Cabrnochová, M.; Lichá, I.; et al. Evaluation of TD test for analysis of persistence or tolerance in clinical isolates of Staphylococcus aureus. J Microbiol Methods 2019, 167, 105705. [Google Scholar] [CrossRef]
- McDermott, W. Microbial persistence. Yale J Biol Med 1958, 30, 257–291. [Google Scholar] [PubMed]
- Brauner, A.; Fridman, O.; Gefen, O.; Balaban, N.Q. Distinguishing between resistance, tolerance and persistence to antibiotic treatment. Nat. Rev. Microbiol. 2016, 14, 320–330. [Google Scholar] [CrossRef]
- Trastoy, R.; Manso, T.; Fernández-García, L.; et al. Mechanisms of Bacterial Tolerance and Persistence in the Gastrointestinal and Respiratory Environments. Clin Microbiol Rev 2018, 31. [Google Scholar] [CrossRef]
- Levin, B.R.; Rozen, D.E. Non-inherited antibiotic resistance. Nat Rev Microbiol 2006, 4, 556–562. [Google Scholar] [CrossRef]
- Levin-Reisman, I.; Ronin, I.; Gefen, O.; et al. Antibiotic tolerance facilitates the evolution of resistance. Science (80- ) 2017, 355, 826–830. [Google Scholar] [CrossRef]
- Levin-Reisman, I.; Brauner, A.; Ronin, I.; Balaban, N.Q. Epistasis between antibiotic tolerance, persistence, and resistance mutations. Proc Natl Acad Sci USA 2019, 116, 14734–14739. [Google Scholar] [CrossRef]
- Huemer, M.; Mairpady Shambat, S.; Brugger, S.D.; Zinkernagel, A.S. Antibiotic resistance and persistence-Implications for human health and treatment perspectives. EMBO Rep 2020, 21, e51034. [Google Scholar] [CrossRef]
- Kussell, E.; Kishony, R.; Balaban, N.Q.; Leibler, S. Bacterial persistence: a model of survival in changing environments. Genetics 2005, 169, 1807–1814. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K. Multidrug tolerance of biofilms and persister cells. Bact biofilms 2008, 107–131. [Google Scholar]
- Lazarovits, G.; Gefen, O.; Cahanian, N.; et al. Prevalence of Antibiotic Tolerance and Risk for Reinfection Among Escherichia coli Bloodstream Isolates: A Prospective Cohort Study. Clin Infect Dis an Off Publ Infect Dis Soc Am 2022, 75, 1706–1713. [Google Scholar] [CrossRef] [PubMed]
- Santi, I.; Manfredi, P.; Maffei, E.; et al. Evolution of antibiotic tolerance shapes resistance development in chronic Pseudomonas aeruginosa infections. MBio 2021, 12, e03482–20. [Google Scholar] [CrossRef]
- Walter, N.D.; Born, S.E.M.; Robertson, G.T.; et al. Mycobacterium tuberculosis precursor rRNA as a measure of treatment-shortening activity of drugs and regimens. Nat Commun 2021, 12, 1–11. [Google Scholar] [CrossRef]
- Conlon, B.P. Staphylococcus aureus chronic and relapsing infections: Evidence of a role for persister cells: An investigation of persister cells, their formation and their role in S. aureus disease. Bioessays 2014, 36, 991–996. [Google Scholar] [CrossRef]
- Salina, E.G.; Makarov, V. Mycobacterium tuberculosis Dormancy: How to Fight a Hidden Danger. Microorganisms 2022, 10, 2334. [Google Scholar] [CrossRef]
- Ignatov, D.V.; Salina, E.G.; Fursov, M.V.; et al. Dormant non-culturable Mycobacterium tuberculosis retains stable low-abundant mRNA. BMC Genomics 2015, 16, 954. [Google Scholar] [CrossRef]
- Miki, M.; Miyu, H.; Madhuri, S.; et al. Genetic and Transcriptomic Analyses of Ciprofloxacin-Tolerant Staphylococcus aureus Isolated by the Replica Plating Tolerance Isolation System (REPTIS). Antimicrob Agents Chemother 2019, 63, 10.1128–aac.02019. [Google Scholar] [CrossRef]
- Zheng, E.J.; Andrews, I.W.; Grote, A.T.; et al. Modulating the evolutionary trajectory of tolerance using antibiotics with different metabolic dependencies. Nat Commun 2022, 13, 2525. [Google Scholar] [CrossRef] [PubMed]
- Schildkraut, J.A.; Coolen, J.P.M.; Burbaud, S.; et al. RNA Sequencing Elucidates Drug-Specific Mechanisms of Antibiotic Tolerance and Resistance in Mycobacterium abscessus. Antimicrob Agents Chemother 2022, 66, e0150921. [Google Scholar] [CrossRef] [PubMed]
- Balduck, M.; Laumen, J.G.E.; Abdellati, S.; et al. Tolerance to Ceftriaxone in Neisseria gonorrhoeae: Rapid Induction in WHO P Reference Strain and Detection in Clinical Isolates. Antibiot 2022, 11, 1480. [Google Scholar] [CrossRef]
- Manoharan-Basil, S.S.; Balduck, M.; Abdellati, S.; et al. Enolase is implicated in the emergence of gonococcal tolerance to ceftriaxone. Antibiotics 2023, 12, 534. [Google Scholar] [CrossRef]
- Balduck, M.; Strikker, A.; Gestels, Z.; et al. The Prevalence of Antibiotic Tolerance in Neisseria gonorrhoeae Varies by Anatomical Site. Pathogens 2024, 13, 538. [Google Scholar] [CrossRef]
- White, D.G.; Goldman, J.D.; Demple, B.; Levy, S.B. Role of the acrAB locus in organic solvent tolerance mediated by expression of marA, soxS, or robA in Escherichia coli. J Bacteriol 1997, 179, 6122–6126. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Haycocks, J.R.J.; Middlemiss, A.D.; et al. The multiple antibiotic resistance operon of enteric bacteria controls DNA repair and outer membrane integrity. Nat Commun 2017, 8, 1444. [Google Scholar] [CrossRef]
- Liu, C.-H.; Di, Y.P. Analysis of RNA Sequencing Data Using CLC Genomics Workbench. Methods Mol Biol 2020, 2102, 61–113. [Google Scholar] [PubMed]
- Dong, H.; Nilsson, L.; Kurland, C.G. Co-variation of tRNA abundance and codon usage in Escherichia coli at different growth rates. J Mol Biol 1996, 260, 649–663. [Google Scholar] [CrossRef]
- Sørensen, M.A.; Fehler, A.O.; Lo Svenningsen, S. Transfer RNA instability as a stress response in Escherichia coli: Rapid dynamics of the tRNA pool as a function of demand. RNA Biol 2018, 15, 586–593. [Google Scholar] [CrossRef]
- Wassarman, K.M. 6S RNA, a Global Regulator of Transcription. Microbiol Spectr 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- Storz, G.; Vogel, J.; Wassarman, K.M. Regulation by small RNAs in bacteria: expanding frontiers. Mol Cell 2011, 43, 880–891. [Google Scholar] [CrossRef] [PubMed]
- Holmqvist, E.; Wagner, E.G.H. Impact of bacterial sRNAs in stress responses. Biochem Soc Trans 2017, 45, 1203–1212. [Google Scholar] [CrossRef]
- Burenina, O.Y.; Elkina, D.A.; Ovcharenko, A.; et al. Involvement of E. coli 6S RNA in Oxidative Stress Response. Int. J. Mol. Sci. 2022, 23, 3653. [Google Scholar] [CrossRef]
- Li, Z.; Zhu, L.; Yu, Z.; et al. 6S-1 RNA Contributes to Sporulation and Parasporal Crystal Formation in Bacillus thuringiensis. Front Microbiol 2020, 11, 604458. [Google Scholar] [CrossRef]
- Beare, P.A.; Unsworth, N.; Andoh, M.; et al. Comparative genomics reveal extensive transposon-mediated genomic plasticity and diversity among potential effector proteins within the genus Coxiella. Infect Immun 2009, 77, 642–656. [Google Scholar] [CrossRef]
- Yadav, A.; Brewer, M.N.; Elshahed, M.S.; Shaw, E.I. Comparative Transcriptomics and Genomics from Continuous Axenic Media Growth Identifies Coxiella burnetii Intracellular Survival Strategies. bioRxiv 2023. [Google Scholar]
- Kertesz, M.A. Bacterial transporters for sulfate and organosulfur compounds. Res Microbiol 2001, 152, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Yurgel, S.N.; Rice, J.; Mulder, M.; Kahn, M.L. GlnB/GlnK PII proteins and regulation of the Sinorhizobium meliloti Rm1021 nitrogen stress response and symbiotic function. J Bacteriol 2010, 192, 2473–2481. [Google Scholar] [CrossRef]
- Ikeda, T.P.; Shauger, A.E.; Kustu, S. Salmonella typhimurium apparently perceives external nitrogen limitation as internal glutamine limitation. J Mol Biol 1996, 259, 589–607. [Google Scholar] [CrossRef]
- Javelle, A.; Severi, E.; Thornton, J.; Merrick, M. Ammonium sensing in Escherichia coli. Role of the ammonium transporter AmtB and AmtB-GlnK complex formation. J Biol Chem 2004, 279, 8530–8538. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Peliska, J.A.; Ninfa, A.J. Enzymological characterization of the signal-transducing uridylyltransferase/uridylyl-removing enzyme (EC 2.7.7.59) of Escherichia coli and its interaction with the PII protein. Biochemistry 1998, 37, 12782–12794. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.Y.S.; Hatzis, C.L.; Lau, A.; et al. Treatment efficacy for pharyngeal Neisseria gonorrhoeae: a systematic review and meta-analysis of randomized controlled trials. J Antimicrob Chemother 2020, 75, 3109–3119. [Google Scholar] [CrossRef]
- Kenyon, C.; Laumen, J.; Manoharan-Basil, S. Choosing New Therapies for Gonorrhoea: We Need to Consider the Impact on the Pan-Neisseria Genome. A Viewpoint. Antibiot 2021, 10, 515. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Heatmap of differentially expressed genes in condition 2 showing the expression levels of all 13 DEGs across the CRO-tolerant and control isolates.
Figure 1.
Heatmap of differentially expressed genes in condition 2 showing the expression levels of all 13 DEGs across the CRO-tolerant and control isolates.
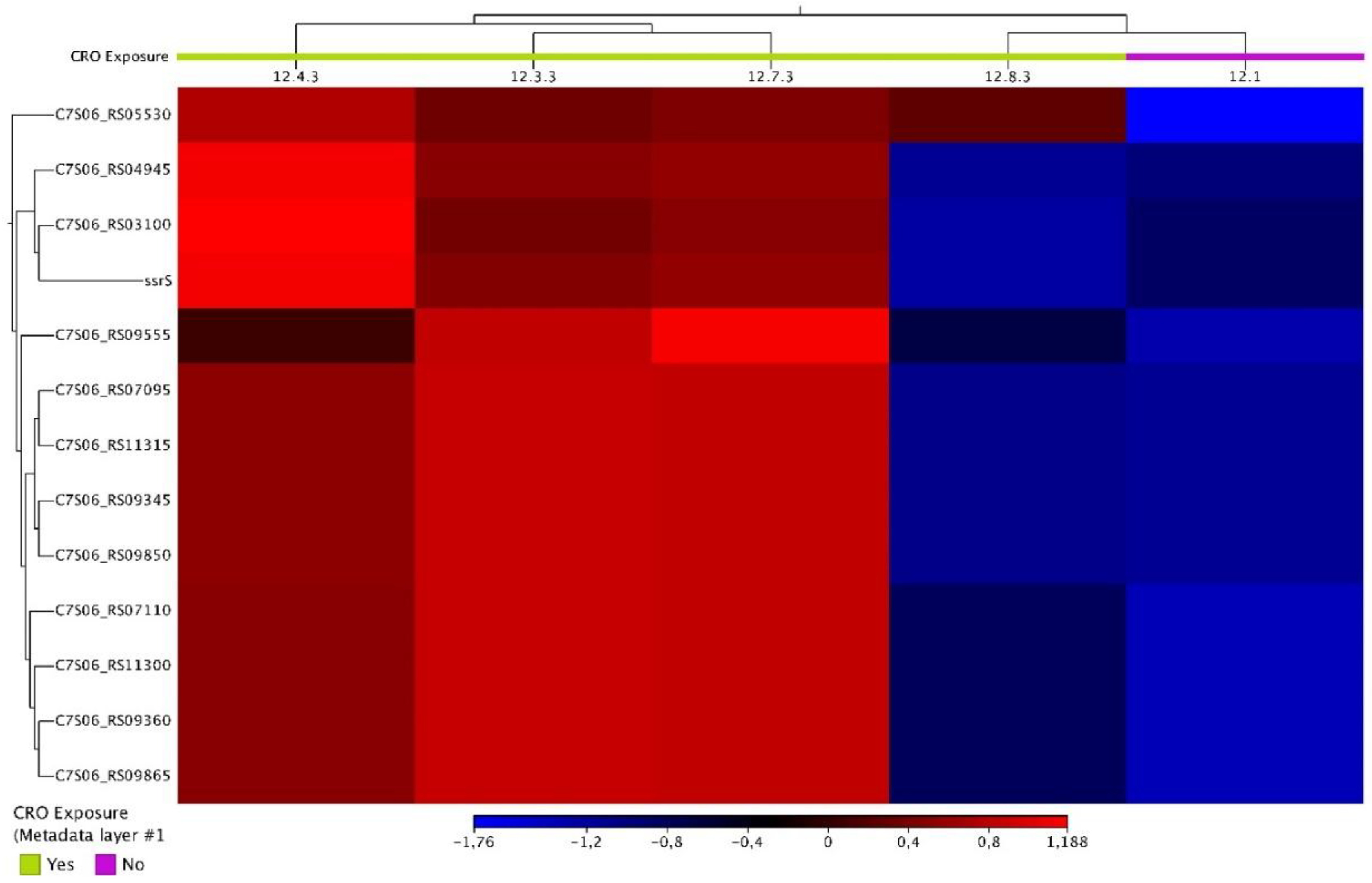
Figure 2.
Heatmap of differentially expressed genes in condition 3 showing the expression levels of all 51 DEGs across the CRO-tolerant and control isolates.
Figure 2.
Heatmap of differentially expressed genes in condition 3 showing the expression levels of all 51 DEGs across the CRO-tolerant and control isolates.

Figure 3.
Heatmap of differentially expressed genes in condition 4 showing the expression levels of all 10 DEGs across the CRO-tolerant and control isolate.
Figure 3.
Heatmap of differentially expressed genes in condition 4 showing the expression levels of all 10 DEGs across the CRO-tolerant and control isolate.
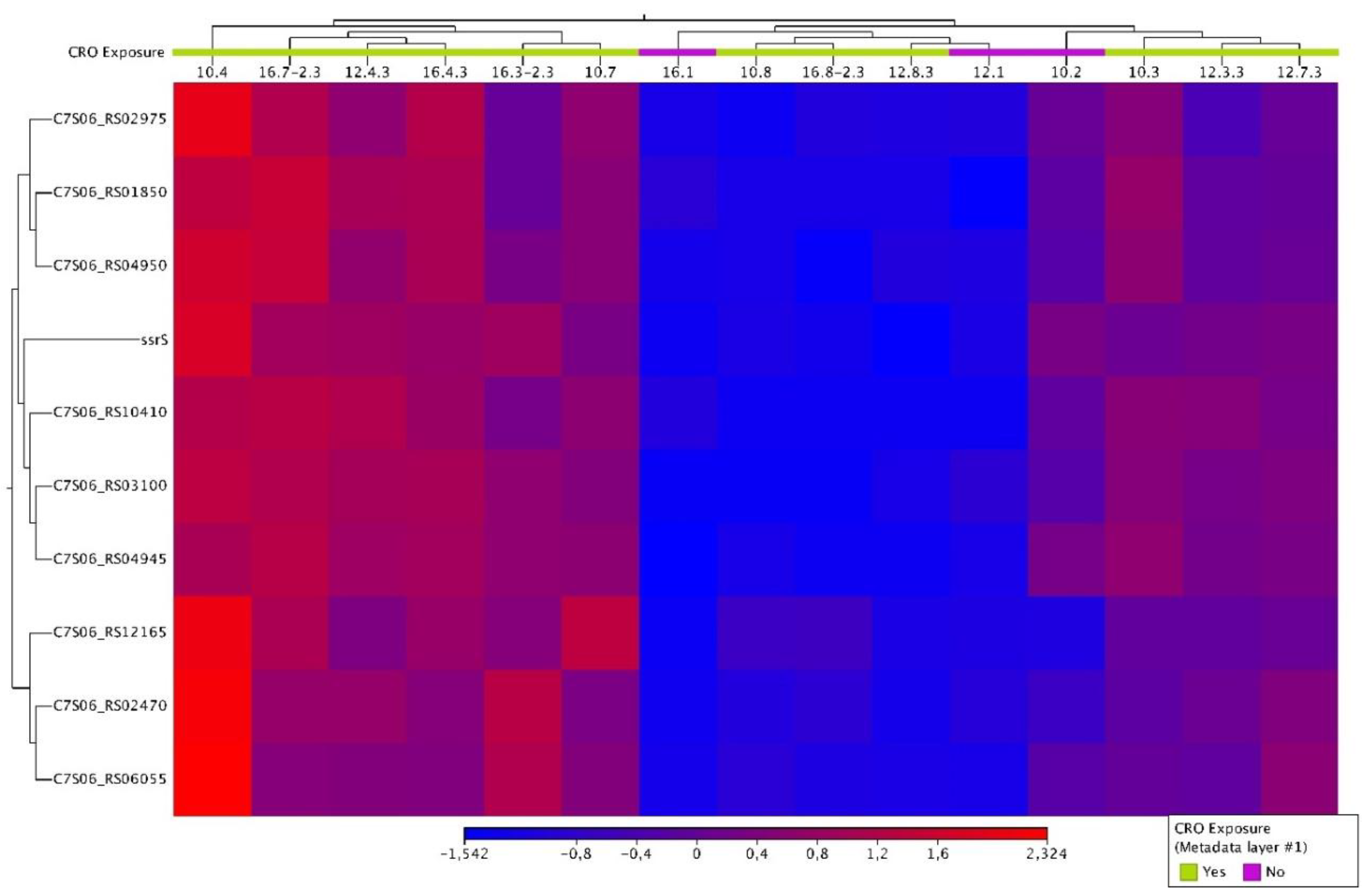

Table 1.
Evolution of ceftriaxone tolerance in the samples assessed. The experiment was conducted over seven consecutive days with six biological replicates and two control replicates of N. gonorrhoeae WHO-P. Whilst lineages 1-6 were exposed to daily ceftriaxone, the controls were exposed to identical conditions with the exception of not being exposed to ceftriaxone daily. The isolates that were subjected to RNA-Seq are colored in grey.
Table 1.
Evolution of ceftriaxone tolerance in the samples assessed. The experiment was conducted over seven consecutive days with six biological replicates and two control replicates of N. gonorrhoeae WHO-P. Whilst lineages 1-6 were exposed to daily ceftriaxone, the controls were exposed to identical conditions with the exception of not being exposed to ceftriaxone daily. The isolates that were subjected to RNA-Seq are colored in grey.
| CRO exposure [in days] | ||||||||
| Days (d) | d0 | d1 | d2 | d3 | d4 | d5 | d6 | d7 |
| Control 1 | - | 10.2 | - | 12.1 | - | - | - | 16.1 |
| Control 2 | - | - | - | - | - | - | - | - |
| Lineage 1 | - | 10.3 T | 11.3-2.3 T | 12.3.3 T | 13.3.3 T | 14.3.3 T | 15.3-2.3 T | 16.3-2.3 T |
| Lineage 2 | - | 10.4 T | 11.4-2.3 T | 12.4.3 T | 13.4.3 T | 14.4.3 T | 15.4-2.3 T | 16.4-2.3 T |
| Lineage 3 | - | - | 11.5-2.3 T | - | - | - | - | 16.5-2.3 T |
| Lineage 4 | - | - | - | 12.6.3 T | - | - | - | 16.6-2.3 T |
| Lineage 5 | - | 10.7 T | 11.7-2.3 T | 12.7.3 T | 13.7.3 T | 14.7.3 T | 15.7.3 T | 16.7-2.3 T |
| Lineage 6 | - | 10.8 T | 11.8-2.3 T | 12.8.3 T | - | - | - | 16.8-2.3 T |
- No tolerant samples stored/collected. T- Tolerant isolates.
Table 2.
List of differentially expressed genes for conditions 1 to 4.
| Condition | Samples | Name | Product | Log2 fold change | P-value | FDR p-value |
|---|---|---|---|---|---|---|
| Condition 1 | 10.2 vs 10.3, 10.4, 1.0.7, 10.8 | NA | NA | NA | NA | NA |
| Condition 2 | 12.1 vs 12.3.3, 12.4.3, 12.7.3, 12.8.3 | C7S06_RS03100 | tRNA-Ser | -7.39860353 | 1.38E-06 | 2.84E-04 |
| C7S06_RS04945 | tRNA-Leu | -8.83399392 | 2.96E-08 | 1.34E-05 | ||
| C7S06_RS05530 | helix-turn-helix domain-containing protein | -6.9900684 | 1.34E-06 | 2.84E-04 | ||
| C7S06_RS07095 | 23S ribosomal RNA | -7.36462585 | 6.27E-07 | 2.25E-04 | ||
| C7S06_RS07110 | 16S ribosomal RNA | -8.49118611 | 1.91E-09 | 1.13E-06 | ||
| C7S06_RS09345 | 23S ribosomal RNA | -7.36644228 | 6.99E-07 | 2.25E-04 | ||
| C7S06_RS09360 | 16S ribosomal RNA | -8.50896375 | 2.00E-09 | 1.13E-06 | ||
| C7S06_RS09555 | hypothetical protein | -3.17366339 | 3.48E-06 | 6.55E-04 | ||
| C7S06_RS09850 | 23S ribosomal RNA | -7.37838277 | 9.49E-07 | 2.38E-04 | ||
| C7S06_RS09865 | 16S ribosomal RNA | -8.55848201 | 1.93E-09 | 1.13E-06 | ||
| ssrS | 6S RNA | -4.0081717 | 1.04E-04 | 0.017997931 | ||
| C7S06_RS11300 | 16S ribosomal RNA | -8.57394491 | 1.10E-09 | 1.13E-06 | ||
| C7S06_RS11315 | 23S ribosomal RNA | -7.34451019 | 8.95E-07 | 2.38E-04 | ||
| Condition 3 | 16.1 vs 16.3.3, 16.4.3, 16.7-2.3, 16.8-2.3 | secB | protein-export chaperone SecB | -1.84506299 | 1.12E-04 | 9.76E-03 |
| C7S06_RS01160 | 2,3-diphosphoglycerate-dependent phosphoglycerate mutase | -1.28680155 | 7.56E-04 | 0.035587745 | ||
| C7S06_RS01850 | tRNA-Ser | -7.99931846 | 1.70E-06 | 4.27E-04 | ||
| C7S06_RS02470 | IS1595 family transposase | -3.02614034 | 3.15E-06 | 5.92E-04 | ||
| htpX | protease htpX | -1.91863904 | 4.27E-04 | 0.023548408 | ||
| C7S06_RS02975 | tRNA-Gln | -5.7211052 | 1.49E-05 | 1.98E-03 | ||
| cysT | sulfate ABC transporter permease subunit CysT | 1.46371918 | 1.68E-04 | 0.012145005 | ||
| C7S06_RS03025 | isoprenylcysteine carboxyl methyltransferase family protein | -1.30810263 | 2.18E-04 | 0.014056098 | ||
| C7S06_RS03100 | tRNA-Ser | -15.0233146 | 5.45E-04 | 0.026758275 | ||
| C7S06_RS03105 | tRNA-Ser | -7.48792977 | 2.47E-04 | 0.015104502 | ||
| C7S06_RS03165 | hypothetical protein | -6.48897383 | 2.84E-06 | 5.82E-04 | ||
| C7S06_RS03180 | hypothetical protein | -3.58579449 | 1.59E-05 | 1.99E-03 | ||
| C7S06_RS03675 | carbonic anhydrase family | -1.83404995 | 2.14E-04 | 0.014056098 | ||
| C7S06_RS04010 | alpha-hydroxy-acid oxidizing protein | -2.16189144 | 5.07E-05 | 4.98E-03 | ||
| glnD | [protein-PII] uridylyltransferase | 1.13672092 | 4.82E-04 | 0.024762705 | ||
| C7S06_RS04945 | tRNA-Leu | -16.2019008 | 1.93E-04 | 0.013209519 | ||
| C7S06_RS04950 | tRNA-Ser | -8.19485807 | 1.55E-07 | 1.17E-04 | ||
| C7S06_RS05295 | serine hydroxymethyltransferase | -1.44687599 | 1.30E-04 | 0.010872726 | ||
| C7S06_RS05355 | sulfate ABC transporter substrate-binding protein | 1.20861114 | 8.61E-04 | 0.03813733 | ||
| C7S06_RS05905 | protein-disulfide reductase DsbD | -1.94221647 | 8.44E-04 | 0.03813733 | ||
| C7S06_RS06055 | hypothetical protein | -2.92315591 | 2.87E-05 | 3.24E-03 | ||
| C7S06_RS06120 | nucleoid-associated protein | -1.05820139 | 5.62E-04 | 0.027017799 | ||
| C7S06_RS06180 | nitronate monooxygenase family protein | -1.47848462 | 4.65E-04 | 0.024439693 | ||
| C7S06_RS13545 | glutamate dehydrogenase | -2.85215757 | 2.35E-04 | 0.014715194 | ||
| C7S06_RS06985 | hemolysin III family protein | -2.22464042 | 2.37E-05 | 2.82E-03 | ||
| C7S06_RS07095 | 23S ribosomal RNA | -8.91263195 | 1.07E-05 | 1.85E-03 | ||
| C7S06_RS07110 | 16S ribosomal RNA | -9.86029166 | 4.90E-07 | 1.61E-04 | ||
| C7S06_RS07485 | PepSY domain-containing protein | 1.81879489 | 4.40E-04 | 0.023678349 | ||
| C7S06_RS07550 | GrxB family glutaredoxin | -1.79669437 | 1.67E-04 | 0.012145005 | ||
| grpE | nucleotide exchange factor GrpE | -1.53580284 | 8.15E-05 | 7.67E-03 | ||
| C7S06_RS08065 | NAD(P)H-dependent oxidoreductase | -3.44183551 | 1.08E-04 | 9.75E-03 | ||
| xth | exodeoxyribonuclease III | -1.15381669 | 5.19E-04 | 0.026030321 | ||
| C7S06_RS08615 | metalloregulator ArsR/SmtB family transcription factor | -1.88716147 | 1.99E-06 | 4.50E-04 | ||
| C7S06_RS08785 | hypothetical protein | -2.31744916 | 3.43E-05 | 3.69E-03 | ||
| C7S06_RS08795 | immunity 41 family protein | -4.78449117 | 0 | 0 | ||
| C7S06_RS09345 | 23S ribosomal RNA | -8.94549528 | 1.28E-05 | 1.92E-03 | ||
| C7S06_RS09360 | 16S ribosomal RNA | -9.91623085 | 5.20E-07 | 1.61E-04 | ||
| C7S06_RS09460 | chloride channel protein | 1.18464463 | 8.43E-04 | 0.03813733 | ||
| C7S06_RS09850 | 23S ribosomal RNA | -8.90732152 | 1.21E-05 | 1.92E-03 | ||
| C7S06_RS09865 | 16S ribosomal RNA | -9.93544646 | 5.64E-07 | 1.61E-04 | ||
| C7S06_RS10210 | pilin | -6.01251507 | 3.06E-04 | 0.017737818 | ||
| C7S06_RS10410 | tRNA-Met | -6.9457042 | 1.63E-04 | 0.012145005 | ||
| ssrS | 6S RNA | -5.31517647 | 1.04E-07 | 1.17E-04 | ||
| C7S06_RS10580 | amino acid ABC transporter permease | 2.03399688 | 1.72E-04 | 0.012145005 | ||
| C7S06_RS10585 | amino acid ABC transporter permease | 1.76436842 | 2.55E-04 | 0.015174939 | ||
| C7S06_RS10775 | hypothetical protein | -1.76534988 | 1.65E-04 | 0.012145005 | ||
| C7S06_RS11300 | 16S ribosomal RNA | -9.8672322 | 5.21E-07 | 1.61E-04 | ||
| C7S06_RS11315 | 23S ribosomal RNA | -8.94465167 | 1.41E-05 | 1.98E-03 | ||
| C7S06_RS11330 | helix-hairpin-helix domain-containing protein | -1.28389104 | 3.92E-04 | 0.022138925 | ||
| C7S06_RS12160 | tRNA-Asp | -9.4247375 | 5.70E-07 | 1.61E-04 | ||
| C7S06_RS12240 | MFS transporter | 1.82688438 | 4.79E-05 | 4.91E-03 | ||
| Condition 4 | All Control (n=3) vs All Tolerant (n=12) isolates | C7S06_RS01850 | tRNA-Ser | -6.10107845 | 1.91E-07 | 1.45E-04 |
| C7S06_RS02470 | IS1595 family transposase | -2.46234082 | 5.57E-05 | 0.015579126 | ||
| C7S06_RS02975 | tRNA-Gln | -3.88761923 | 3.93E-05 | 0.015579126 | ||
| C7S06_RS03100 | tRNA-Ser | -7.28200512 | 5.49E-09 | 1.25E-05 | ||
| C7S06_RS04945 | tRNA-Leu | -4.95129854 | 6.05E-05 | 0.015579126 | ||
| C7S06_RS04950 | tRNA-Ser | -5.37973119 | 1.22E-07 | 1.39E-04 | ||
| C7S06_RS06055 | hypothetical protein | -2.54434011 | 6.16E-05 | 0.015579126 | ||
| C7S06_RS10410 | tRNA-Met | -5.07346986 | 6.95E-05 | 0.015835072 | ||
| ssrS | 6S RNA | -3.37642264 | 5.21E-05 | 0.015579126 | ||
| C7S06_RS12165 | tRNA-Val | -3.7538874 | 3.59E-05 | 0.015579126 |
NA- not available.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



