
Submitted:
27 March 2025
Posted:
29 March 2025
You are already at the latest version
Abstract
Glioblastoma (GBM) is considered the most aggressive primary brain tumor with a high recurrence rate after treatment. Despite recent advances in molecular biology, the exact genetic and epigenetic pathways involved in gliomagenesis are not fully understood. Additionally, physicians currently lack robust non-invasive methods to diagnose and predict glioma with high accuracy. Cancer biomarkers, such as DNA, RNA, transcription factors, cell surface receptors, enzymes, and metabolites, can be found in tumor tissue, cerebrospinal fluid (CSF), and/or blood, and are currently being investigated to overcome these challenges. Non-coding RNAs (ncRNAs) are new classes of regulatory RNA that recently attracted attention due to their role in cancer and other diseases. ncRNAs are involved in tumor formation, invasion, and progression stages. They also regulate the cell cycle, apoptosis, autophagy, stemness, angiogenesis, blood-tumor-brain barrier integrity, and tumor metabolism. Expression levels of ncRNAs can be related to tumor grade, survival, treatment response (chemotherapeutic drugs or radiotherapy), and could determine overall prognosis. Therefore, the circulatory or local levels of these molecules could serve as real-time biomarkers. Here, we highlight the possible different non-coding RNAs that could be detected in the tissues of patients with glioma and their possible clinical applications as diagnostic and prognostic biomarkers.
Keywords:
Glioma
; ncRNAS
; Prognosis
; microRNAs
1. Introduction
Glioblastoma (GBM) is considered the most aggressive primary malignant brain tumor, even with advanced treatment options [1]. Despite significant advancements in our understanding of GBM, median survival has remained less than two years [2]. In 2005, the European Organization for Research and Treatment of Cancer (EORTC) and National Cancer Institute of Canada (NCIC) conducted a phase III study that showed that combined treatment with temozolomide and radiotherapy improved the outcome compared with radiotherapy alone by 5.2 to 6.7 months. [3].
Several factors contribute to treatment failure, such as the heterogeneity of the GBM microenvironment, the presence of regenerative cancer stem cells and the development of resistance to common therapies [4]. Therefore, designing new techniques that could aid in early diagnosis and monitoring of treatment response and tumor progression is crucial. A biomarker is described as a substance that can be objectively assessed as a parameter for a particular biological or pathological process or can measure responses to a specific therapy [4]. Cancer biomarkers can be obtained from tumor tissues or sera, and include DNA, RNA, enzymes, metabolites, transcription factors, and cell surface receptors [4].
Non-coding RNAs (ncRNAs) are new classes of regulatory RNA that control different physiological processes, such as gene regulation, chromatin arrangement, and cellular signaling [5]. Only less than 2% of the genome accounts for protein-coding genes, while the remaining represents non-coding elements that for a long time were considered as “junk” [6].
ncRNAs are classified into short and long types based on the nucleotide sequence length. Small ncRNAs (which include 20–200 nucleotides) include microRNAs (miRNAs), small nuclear RNAs (snRNAs), small interfering RNAs (siRNAs), small nucleolar RNAs (snoRNAs), and Piwi-associated small RNAs (piRNAs) [7]. Scientists thought that long non-coding RNAs (lncRNAs) are transcription nonfunctioning byproducts; however, certain functions have been recently uncovered with the help of modern sequencing technologies [8]. LncRNAs are divided into different groups on the basis of their locations in the genome. They include intronic lncRNAs, divergent lncRNAs, antisense lncRNAs, promoter-associated lncRNAs, intergenic lncRNAs, enhancer RNAs (eRNAs), and transcription-start-site-associated lncRNAs [9].
2. Results
2.2. MicroRNAs
MicroRNAs (miRNAs) are non-coding RNA molecules that comprise 19 to 25 nucleotides that control post-transcriptional gene silencing and serve as gene regulatory switches through targeting hundreds of intermediary mRNAs [10].
Two processes are involved in miRNA biogenesis. The first exists in the nucleus, where a nuclear RNase III converts the primary transcript (pri-miRNA) into a precursor (pre-miRNA) Figure 1. The pre-miRNA is transferred outside the nucleus and cleaved into a short-lived dsRNA (19–25 nucleotides) by Dicer. Then, the double-strand is untwisted, and the mature miRNA is formed from one strand that is included in an Argonaut (Ago)-protein-containing complex, known as the RNA-induced silencing complex (RISC) [11,12].
2.3. MicroRNAs as Biomarkers of GBM
Some tumor types affect the expression of miRNAs [16]. Minor genetic alterations involving the miRNA sequence can lead to major phenotypic changes.Their functional affiliation with cancer and the propensity to trigger several genetic changes suggest miRNAs as potential parameters for conferring cancer susceptibility. Several studies have recently reported that certain serum- or cerebrospinal fluid (CSF)-circulating miRNAs are differentially produced in patients with GBM and could serve as biomarkers with prognostic purposes [17] (Table 1). However, not all these microRNAs have been involved in clinical trials. Few microRNAs were used in clinical trials for use as potential biomarkers Table 2.
In 2013, miRNA expression was first identified in the sera of untreated patients diagnosed with high-grade glioma (WHO grades III-IV) and healthy controls. The serum miRNA levels were different among the high-grade astrocytomas patients and healthy controls. The findings revealed that 50 miRNAs were differentially expressed in astrocytoma, with approximately 15 being upregulated and the other 35 being downregulated. Seven miRNAs, including miR-15b, miR-23a, miR-133a, miR-150, miR-19, miR-497, and miR-548b-5p, were downregulated in grades II-IV astrocytoma patients and they showed high specificity and sensitivity for predicting malignant behavior. Additionally, after surgery, serum levels of these miRNAs were markedly elevated [18]. Dong et al. identified that 115 miRNAs were increased and 24 miRNAs were decreased in GBM tissues [19].
Manterola et al. studied the diagnostic utility of miRNAs isolated from the sera of patients diagnosed with GBM. They found that the expression levels of RNU6-1, miR-320, and miR-574-3p were highly associated with the diagnosis of GBM. RNU6-1 was also reported to be an independent marker of GBM diagnosis [19]. Wu et al. evaluated the level of miR-29 in different glial tumor patients’ serum at various stages. The level of miR-29 was significantly different in grade I–II patients compared to grade III-IV patients. Generally, the serum miR-29 level predicted progression to high-grade glioma [20]. Wei et al. revealed that miR-125b could be a possible biomarker for high grade gliomas with a high degree of accuracy. Their findings revealed that the serum levels of miR-125b were lower in GBM patients and the degree of reduction matched higher grade features [21]. Lai et al. measured blood miR-210 levels in patients with glioma grades I–IV. They found a seven-fold increase in miR-210 levels in serum samples from patients with high grade glioma compared with the healthy controls. Moreover, a high serum miR-210 level was linked to a high tumor grade and poor outcome [22]. Sun et al. discovered that preoperative serum from patients diagnosed with glioma had lower levels of miR-128 than patients harboring meningiomas [23]. In the glioma group, Yue et al. found that the serum expression of miR-205 was slightly lower than in other brain tumors. Compared with the low serum levels of miR-205, the higher serum levels of miR-205 indicated a longer survival [24]. The circulating miR-137 in GBM patients was screened in both early and advanced stages. Le et al. found that patients with GBM have lower levels of miR-137 in their blood. It was much lower in advanced GBM stages. Low serum miR-137 levels correlated strongly with higher histological grades and lower Karnofsky Performance Scale (KPS) [25]. Low blood levels of miR-137 correlated with poor survival in GBM patients [25]. Regazzo et al. measured serum and tissue miRNAs in 30 patients with brain cancers and 15 healthy controls. They noticed that patients with a high-grade glioma had lower concentration than low-grade tumors. GBM patients exhibited lower miR-497 and -125b levels than the low-grade group and this conclusion was confirmed by the final systematic review that included 19 studies [26].
2.4. MicroRNA-100 as a Tumor Biomarker of GBM
Zhang et al. [18] found that the relative serum miR-100 levels in GBM patients were significantly lower than those in the low risk factors group or healthy controls . Furthermore, the miR-100 levels in post-treatment blood samples were dramatically increased compared with the pre-treatment levels (P < 0.01) [18]. They observed that low serum miR-100 expression was significantly correlated with KPS (P = 0.0207), IDH1/2 mutation status (P = 0.0071), and MGMT methylation status (P = 0.0239). Therefore, miRNA-1oo could predict certain clinicopathological features of GBM. However, no correlation was noticed between its level and other clinical characteristics including age, gender, and tumor size [18].
2.5. How miRNA Expression May Be Associated with Carcinogenesis
Liu et al. discovered that the 3′-UTR of the PLK1 gene included binding sites for miR-100, making PLK1 a possible miR-100 target. In several tumors, PLK1 is considered a functional target for miR-100 [27]. Lee et al. found that PLK1 inhibition impacted cellular growth, induced apoptosis, and downregulated SOX2 expression in SF188 cells. Consistent with SOX2 inhibition, absent PLK1 effect allowed the cells to differentiate and express elevated levels of glial fibrillary acidic protein [28]. Therefore, we could suggest that the lower levels of microRNA-100 may disinhibit the PLK1 activity, enhancing cellular proliferation and growth.
Endothelial and vascular smooth muscle cells express miRNAs that could have inhibitory effects on neovascularization by regulating the mammalian target of rapamycin (mTOR) [29,30]. The overactivation of mTOR signaling is linked to tumor initiation and progression, and mTOR expression has been disrupted in certain cancers, including breast, kidney, lung, melanoma, bladder, brain, and renal carcinomas [29]. Hence, there may be a relation between the decreased level of miRNA-100 and the tumor vascularity.
2.6. MicroRNA-504
Nearly 25% of cancer patients will present with brain metastasis, which can be challenging to differentiate radiographically from other tumors such as GBM [31]. However, Jin et al. found that non-small cell lung cancer (NSCLC) and GBMs differ in their serum miRNA expression profile, potentially enabling a possible differentiation [32]. The serum levels of miRs 221, 608, and 504 were measured in patients with GBM, with NSCLC brain metastasis, and in the normal population. Jin et al. indicated that serum levels of miR-221 were upregulated in GBM and patients with metastatic NSCLC. Although both miR-608 and 504 were explicitly downregulated only in the GBM patient group, only miR-504 serum level can be potentially used as a reliable marker to differentiate between NSCLC metastasis and GBM [32].
2.7. MicroRNA-203
Chen et al. investigated the in vitro expression of miR-203 in GBM cells and found that it was significantly reduced compared to low grade tumors. In a human cohort, serum miR-203 expression was significantly lower in GBM patients compared with low-grade glioma (LGG) patients and normal controls. Furthermore, serum miR-203 distinguished GBM patients from LGG patients and healthy people [33]. A low serum miR-203 correlated well with larger tumor size, as well as a low KPS. Patients with lower blood levels of miR-203 have shorter overall survival (OS) and progression-free survival. Accordingly, miR-203 expression is an individualized prognostic factor for short OS in GBM patients [33].
2.8. MicroRNA-301a
Lan et al. assessed the serum exosomal miR-301a levels in GBM patients and healthy individuals. They found a significant difference in miR-301a serum levels between those patients and non-glioma patients (P < 0.01). Moreover, the miR-301a serum levels were markedly elevated in both HGG and LGG patients. Furthermore, they discovered that the high miR-301a expression level correlated with the pathological grade of the tumor (P = 0.01) [34]. Interestingly, they also found that the miR-301a levels dropped significantly but were still detectable after surgical removal of these tumors (P < 0.01) [34]. The serum exosomal miR-301a expression levels were significantly higher in the glioma patients than those in all other patients diagnosed with meningioma, pituitary adenoma, and PCNSL [34]. Therefore, serum exosomal miR-301 can serve as a potential marker for distinguishing patients with gliomas from patients with other intracranial tumors. [34].
2.9. MicroRNA-145-5b
Zhang et al. found that serum miR-145-5p levels were significantly decreased in GBM patients compared with grade I/II glioma patients and the healthy population. They also discovered that low blood levels of miR-145-5p had a strong connection to aggressive clinicopathological parameters. According to the survival analysis, patients in the high serum miR-145-5p group lived considerably longer than people with low serum levels [35].
2.10. miR-29b
Zhong et al. determined the expression levels of serum miR-29b patients diagnosed with GBM, anaplastic astrocytoma (AA), and healthy volunteers. They found that patients with GBM had significantly lower serum miR-29b levels than anaplastic astrocytoma patients (P = 0.019) or healthy controls (P = 0.001). Interestingly, the serum levels of miR-29b levels were higher postoperatively. [36].
2.11. miR-451a, miR-485-3p, and miR-4298
The serum levels of miR-451a, miR-485-3p, and miR-4298 miRNAs were found to be different between 24 patients diagnosed with GBM and 12 healthy individuals (P < 0.05), according to Wang et al. The prognostic value of these miRNAs was evaluated among patients with GBM. A low serum miR-485-3p level was associated with a poor PFS and overall survival. Combining different miRNA could increase the diagnostic sensitivity of these markers rather than using a single molecule.
2.12. miR-223 and miR-320e
Preoperative and postoperative serum miRNA analysis was performed on a prospective group of GBM patients and healthy volunteers. Candidate miRNAs in 11 patients were tested with several follow-up samples, and the results were compared with MRI measured tumor volumes. Morokoff et al. found nine-gene miRNA signatures that could differentiate GBM patients from healthy controls with 99.8% accuracy. In LGG and GBM, miR-223 and miR-320e showed the most significant changes associated with the tumor volume. In two cases of pseudo-progression, miRNA levels did not rise, reflecting the test’s potential utility in directing care decisions in challenging cases of pseudoprogression [37]. With robust longitudinal studies, these miRNAs could possibly be used to differentiate between true progression and pseudoprogression.
2.13. miR-4763-3p, miR-1915-3p, and miR-3679-5p
The glioma Index involves three miRNAs: miR-4763-3p, miR-1915-3p, and miR-3679-5p. Patients with diffuse GBM had higher serum levels of miR-4763-3p and miR-1915-3p than the controls, indicating a potential oncogenic role. On the other hand, the levels of miR-3679-5p were lower in patients with diffuse GBM, indicating the possible role of miRNA as a tumor suppressor [38]. About 100 patients with diffuse glioma were compared with 200 healthy individuals and, the area under the curve (AUC) was 0.99 (95% CI = 0.99–1.00), with a measured sensitivity of 0.95 (95% CI = 0.89–1.00) and specificity of 0.97 (95% CI = 0.93–1.00). Using the Glioma Index, researchers were able to identify 39 positive samples out of 42 primary central nervous system lymphoma (PCNSL) samples, 25 positive samples out of 28 metastatic brain tumors, and 2 of 2 spinal tumors (100%) [38]. Combining the diagnostic values of different biomarkers using glioma index produces more robust results than using a single marker and increases accuracy..
2.14. Long Non-coding RNAs
Since the coding genome comprises only ~2% of DNA and several clinically significant mutations have been identified in the non-coding sequences, the dysregulation of the non-coding genome may have a role in tumorigenesis, as discussed earlier for miRNA [39]. Long non-coding RNAs (lncRNAs) are defined as non-protein-coding structures with a length of more than 200 nucleotides. While the exact functions of the vast majority of the ~ 40,000 lncRNAs are still uncovered, some of these transcripts are potential regulators of cellular growth and proliferation and may have oncogenic potential [40,41].
There are four potential regulatory pathways that lncRNAs exert on gene expression: epigenetic control, transcriptional (molecule decoy), post-transcriptional (splicing and mRNA decay), and translational regulation [42]. LncRNAs play essential roles in tumorigenesis via controlling the expression of oncogenes, tumor suppressor genes, and other cancer-associated genes [42]. The possible use of circulating lncRNAs as biomarkers is attracting attention. LncRNAs are used as predictors of survival in patients with heart failure [43], a mortality predictor in patients with acute kidney insult, and a new biomarker for diagnosis of coronary artery disease [44], esophageal squamous cell carcinoma [45], gastric cancer [46], and GBM [47,48,49]. Table 3 shows the various lncRNAs studied in GBM.
2.15. HOTAIR
HOTAIR (HOX Transcript Antisense Intergenic RNA) is a long non-coding RNA that relates to the homeobox superfamily and is transcribed on chromosome 12q13.13 from the HOXC locus [45,50]. HOTAIR is a lncRNA measuring approximately 2,015 bp long, located between the HOXC11 and HOXC12 genes [45,50]. HOTAIR-PRC2-LSD1 complex binds to the promoters of genes coding for metastatic inhibitors (such as PCDH10, PCDHB5, and JAM2) Figure 2, silencing them through the modification of H3K27me3 (tri-methylation) and H3K4me2 (demethylation) [51].
Figure 2.
Chromatin-modifying complex as polycomb-repressive complex 2 (PRC2) targets to which HOTAIR can bind and induce changes. PRC2 complex induces the methylation of lysine on histone H3. H3K27-methylation is a form of gene silencing influenced by histone methyltransferase EZH2 (Enhancer of Zeste homolog 2), a component (PRC2). LSD1 is known to form a complex with REST (RE1-Silencing Transcription factor) and CoREST, which have vital roles in gene silencing. Histone methylation and demethylation are dynamic processes of epigenetic modification participating in the regulation of gene transcription, chromatin packaging, and cellular differentiation.
Figure 2.
Chromatin-modifying complex as polycomb-repressive complex 2 (PRC2) targets to which HOTAIR can bind and induce changes. PRC2 complex induces the methylation of lysine on histone H3. H3K27-methylation is a form of gene silencing influenced by histone methyltransferase EZH2 (Enhancer of Zeste homolog 2), a component (PRC2). LSD1 is known to form a complex with REST (RE1-Silencing Transcription factor) and CoREST, which have vital roles in gene silencing. Histone methylation and demethylation are dynamic processes of epigenetic modification participating in the regulation of gene transcription, chromatin packaging, and cellular differentiation.
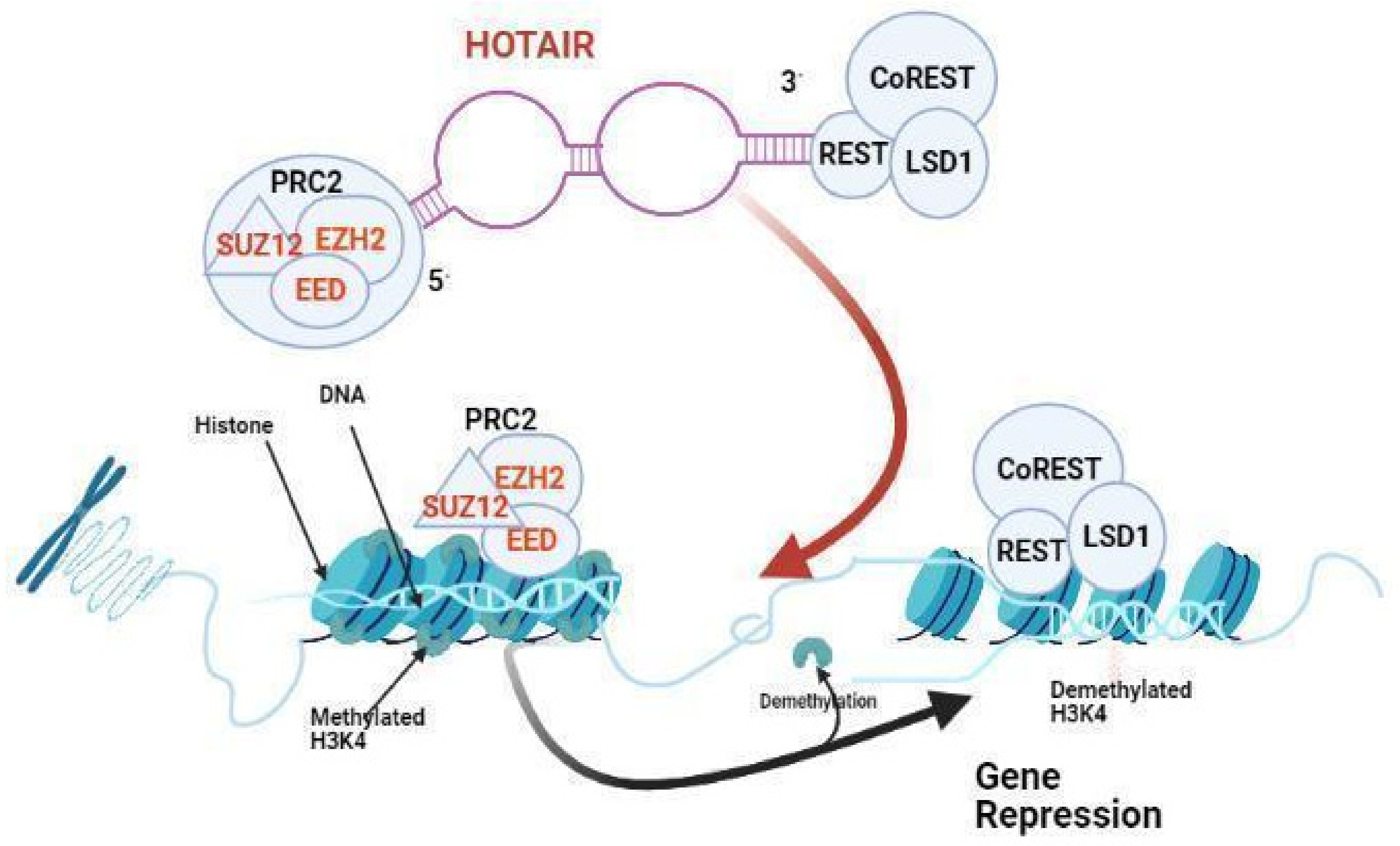
Figure 3.
A summary showing the involvement of different non-coding RNAs in the pathogenesis of GBM.
Figure 3.
A summary showing the involvement of different non-coding RNAs in the pathogenesis of GBM.
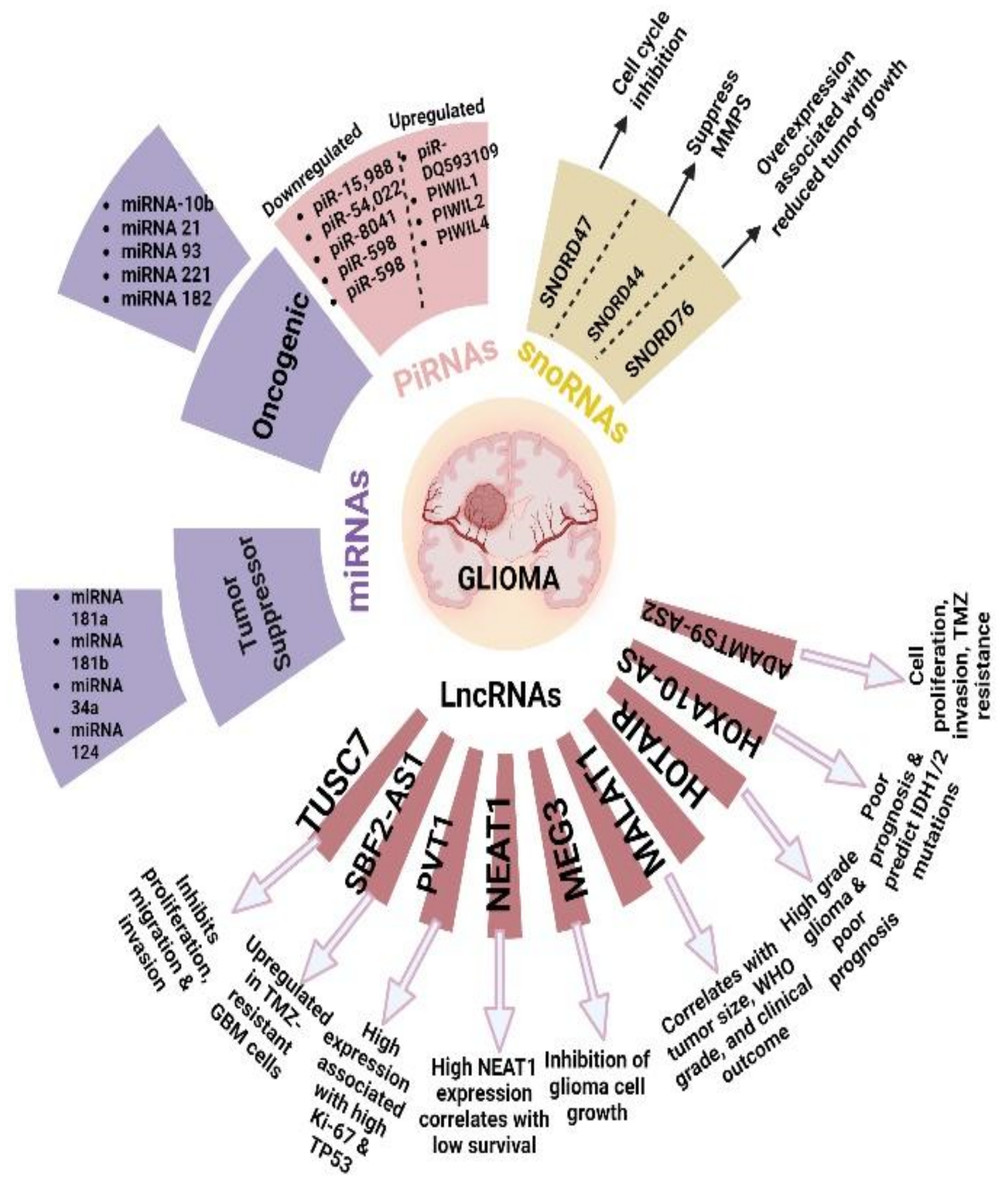
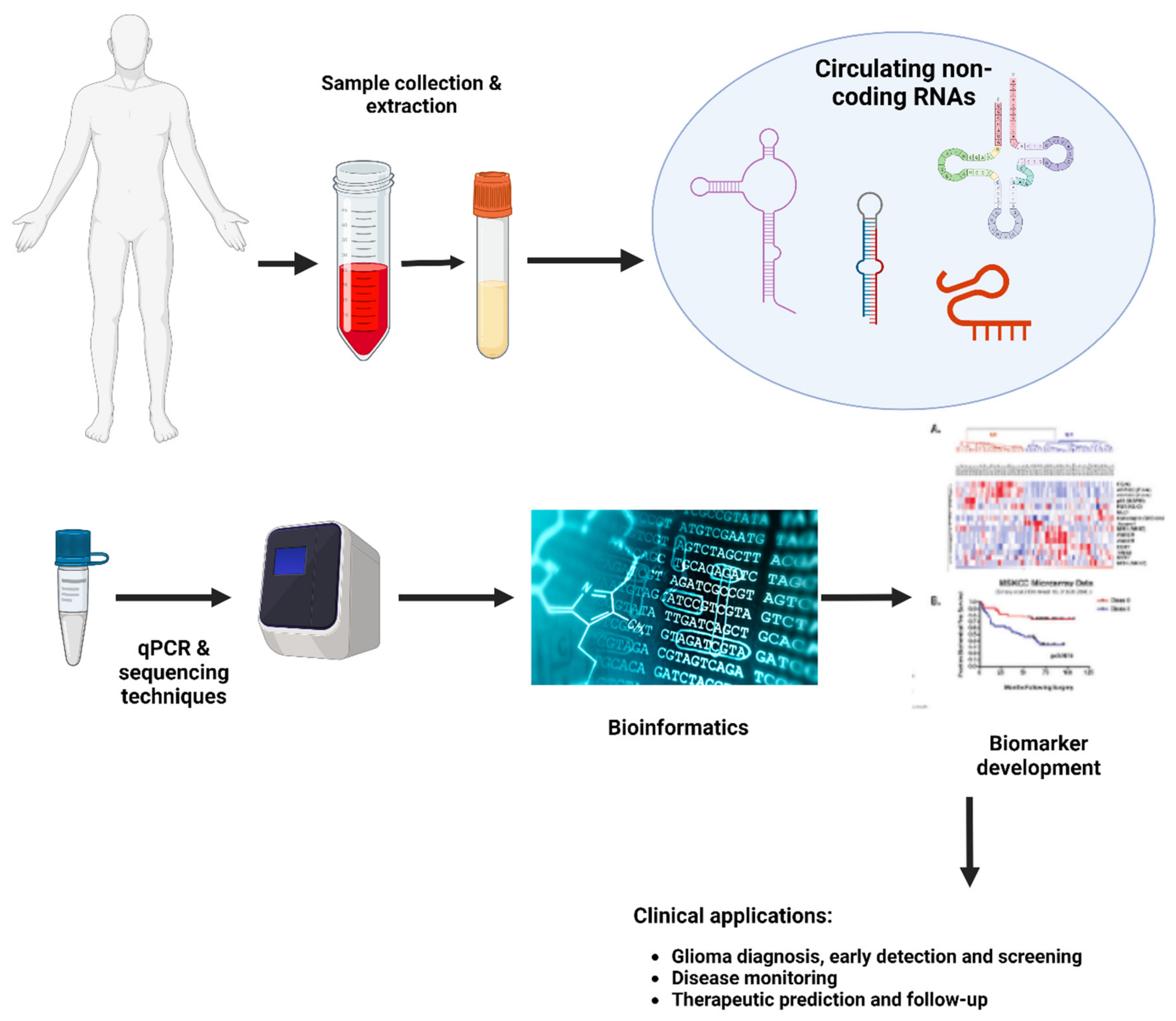
Figure 4.
Main stages for developing a non-coding biomarker.
HOTAIR was an important research focus in multiple tumors. It was found to be a potential diagnostic biomarker for breast cancer [52], and it can predict disease progression in patients with bladder cancer [53]. It is considered a novel diagnostic biomarker for esophageal squamous cell carcinoma [45], and, in general, dysregulation of HOTAIR expression is associated with the aggressive malignancies [53]. Its role in GBM has only more recently been elucidated, however.
2.16. HOTAIR Expression Correlates with Glioma Grade
Zhou et al. estimated the level of HOTAIR expression by whole gene profiling of different gliomas and normal brain samples, and its expression level in GBM was significantly higher than its level in other low-grade tumors and normal brain samples [54]. This implies the possible role of HOTAIR as a diagnostic marker for patients diagnosed with gliomas.
Zhang et al. studied approximately 295 glioma samples from the Chinese Glioma Genome Atlas, including low- and high-grade gliomas [55]. The expression of HOTAIR was observed in HGG more than in low-grade glioma (LGG) (P < 0.001). Furthermore, GBM showed higher HOTAIR transcript levels, compared with those observed in normal tissues (P = 0.093), LGGs (P < 0.001), or anaplastic gliomas (AGs) (P = 0.011). [47].
Tan et al. compared HOTAIR levels in the serum from GBM patients and healthy controls. Serum HOTAIR levels in patients diagnosed with GBM were significantly higher than in the healthy controls (P < 0.0001). High-grade gliomas express higher levels of HOTAIR in their tissues [48].
2.17. The Prognostic Significance of HOTAIR Expression in Glioma
Zhang et al. studied the association between HOTAIR expression and overall survival. They found that HGG samples that showed upregulated HOTAIR expression have shortened survival [47]. Furthermore, HOTAIR expression was inversely correlated with overall survival in anaplastic astrocytoma and GBM [47]. The microarray analysis showed that HGG expressed more HOTAIR compared with LGG (P < 0.001), and GBM samples that were highly positive for HOTAIR had an unfavorable outcome (P = 0.0088) [47].
HOTAIR is identified as an important marker for tumor grade, outcome, and molecular subtype in glioma [47]. The Cancer Genome Atlas (TCGA) network classified GBM into classical, mesenchymal, neural, and proneural subtypes based on molecular features [56]. Zhang et al. classified GBM into two main categories: classical and mesenchymal subtypes (mesenchymal-like) and the neural and proneural subtypes (proneural-like) [47] according to the response to temozolomide therapy [56]. They found that the mesenchymal-like subtype has higher HOTAIR expression than the other subtypes [47].
2.18. HOTAIR as a Biomarker for Responsiveness of GBM Cells to BET Inhibitors
Bromodomain-containing 4 (BRD4), a member of the Bromodomain and Extraterminal (BET) protein family, can control HOTAIR levels through binding to the HOTAIR promoter. Inhibition of BRD4 activity with small molecule Bromodomain and Extraterminal (BET) inhibitors decreased BRD4 binding at the HOTAIR promoter and downregulated HOTAIR levels [57]. Since HOTAIR is overexpressed in GBM, and it is essential for supporting tumor cell proliferation, inhibition of HOTAIR by I-BET151 could induce cell cycle arrest in GBM cells [57]. Pastori et al. suggested that HOTAIR can be used as a biomarker, measuring the response of GBM cells to BET inhibitors [57].
2.19. GBM Evolved Drug-Resistant Mechanisms by lncRNA SBF2-AS1
Zhang et al. found that long non-coding RNA SBF2-AS1 was upregulated in TMZ-resistant GBM cells and its overexpression enhanced the TMZ resistance. In contrast, its inhibition sensitized resistant GBM cells to TMZ [58]. GBM cells could modify the tumor microenvironment to promote chemotherapy resistance by secreting oncogenic lncSBF2-AS1-enriched exosomes. Therefore, serum exosomal lncSBF2-AS1 may serve as a possible diagnostic biomarker for TMZ-resistant GBM [58].
2.20. HOX-Related LncRNAs
Morgan and Bridges suggested that specific genes in fruit flies are responsible for the proper spatial orientation of body parts [59]. Scientists then identified responsible genes (homeotic or HOX genes). Mutations affecting HOX genes cause different body segments to look similar, so that they are called “homeotic” genes. Those genes are categorized in clusters; in humans, approximately four clusters (HOXA, HOXB, HOXC, and HOXD) and 39 HOX genes were identified [60]. HOX genes are involved in embryogenesis, but they can control different cellular processes in adults [61,62]. As HOX genes control cancer-related processes, such as cellular growth, proliferation, migration, invasion, and stemness, some cancers exhibit dysregulation of HOX gene expression [61,62].
HOXA cluster antisense RNA 2 (HOXA-AS2) is a 1048-bp LncRNA located between the HOXA3 and HOXA4 genes in the HOXA cluster [66]. In NB4 promyelocytic leukemia cells treated with all-trans-retinoic acid, HOXA-AS2 was identified as a repressor for apoptosis [67]. HOXA-AS2 is reported to be dysregulated in other tumors, such as liver and colorectal cancer [68]. It stimulates cell migration and enhances invasion in NSCLC through upregulation of IGF-2 [69]. HOXA-AS2 was reported to epigenetically inhibit P21, PLK3, and DDIT3 transcription, which could enhance the process of tumorigenesis in gastric cancer [70]. HOXA-AS2 was upregulated in glioma surgical samples and correlated with tumor size [49]. HOXA-AS2 may serve as a prognostic marker in patients with glioma; however, there is no established clinical evidence to use it for diagnosis.
HOXA10-AS is a LncRNA transcribed from the antisense strand of the homeobox A10 (HOXA10) gene locus at chromosome 7p15.3 [71]. Higher expression of HOXA10 was noticed in different cancers and correlated with poor prognosis [72,73,74]. HOXA10 was reported to be highly expressed in glioma tissue compared with the normal brain [41].
2.21. MALAT1
Metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) is located on chromosome 11q13 as an intergenic LncRNA [75]. GBM cells exhibited lower expression levels of MALAT1 than cancer stem cells [76]. The serum level of this LncRNA has been studied as a prognostic marker in breast and bladder cancers [77,78]. In GBM patients, higher expression of MALAT1 was associated with poor survival [79,80]. Cao et al. found that patients diagnosed with GBM have lower MALAT1 levels compared with healthy volunteers [81]. Li et al. did not find any significant association between the MALAT1 level and survival [80]. Therefore, further investigation is required to utilize MALAT1 as a prognostic marker.
2.22. H19
H19 is a LncRNA that localizes on 11p15.5, a location that is associated with some embryonic tumors [82]. The serum level of H19 has been evaluated as a possible diagnostic marker in gastric cancer [83]. However, its diagnostic value has not been widely investigated in GBM [84]. H19 exerts a role in angiogenesis and EMT through interactions with different miRNAs. Measuring the level of H19 and its interacting miRNAs might have a possible prognostic value in patients diagnosed with GBM.
2.23. PVT1
Plasmacytoma variant translocation 1 (PVT1) is an intergenic lncRNA that localizes to chromosome 8q24, an area that is associated with certain cancers [85]. Its role in lung, breast cancer, and glioma has been investigated [86]. GBM cells showed higher expression levels of PVT1 than normal cells [87]. Higher expression levels of PVT1 in GBM patients are associated with certain aggressive tumor characteristics, such as IDH-wildtype status and higher tumor grades [87,88,89,90]. Therefore, PVT1 could be used as a prognostic indicator for GBM patients, and future studies should be designed to confirm that.
2.24. NEAT1
A LncRNA located at chromosome 11q13.1 and has been considered as an oncogene [91,92,93]. He et al. found that high expression levels of NEAT1 were associated with poor OS [94]. Further investigations are required to evaluate the possible use of NEAT1 as an independent prognostic and diagnostic marker in GBM.
2.25. Circular RNAs
circRNAs were first described in plants in 1976 [95]. With the progressive advances in RNA detection techniques, circRNAs were identified to have intronic, exonic, intergenic and antisense sequences [96,97]. circRNAs are involved in different cellular processes as gene regulation, transcription regulation, interacting with different proteins and epigenetic regulation [98]. circRNAs can control the mRNA expression through complementary binding or adsorption regulating different downstream effects [99,100]. The role of circRNAs is evident in several cancers. circRNA_0005529 and circ-TGFBR2, interact with miR-527 and miR-107 to control human gastric cancer cell proliferation and spread [157]. In esophageal squamous cell cancer, circ-FOXO3 targeted miR-23a to downregulate cellular proliferation [101].
Different studies have demonstrated that circRNAs are dysregulated in glioma. Abnormal expression levels of these molecules were reported in glial tissue compared to normal brain tissues [102,103]. These circRNAs may play a role in migration, apoptosis, proliferation, invasion and angiogenesis of glioma. Table 4 lists glioma related dysregulated circRNAs and the possible functions of each type.
2.26. PIWI-Interacting RNAs (piRNAs)
piRNAs are non-coding molecules derived from single-stranded primary transcripts that are produced from genetic loci termed PIWI clusters [104]. The processing of piRNA involves three steps: pre-piRNAs synthesis: the modification of the 5 ′ and 3′ ends and methylation [105]. These non-coding elements can influence epigenetic reprogramming and can control transcription, translation, and mRNA degradation [106]. Few studies have evaluated the role of piRNAs in glioma. Wang et al. demonstrated that piRNAs play a role in the pathogenesis of GBM [107].
Interestingly, these non-coding RNAs have been detected in the serum of patients with colorectal cancer [108]. These piRNAs are stable for a long time promoting their use as biomarkers, however, their detection is more challenging than other non-coding RNAs [109]. The rs147061479 variant of piR-598 was associated with an estimated risk for gliomagenesis [110]. Moreover, Jacob et al. identified some piRNAs that are downregulated in GBM, such as piR-15,988, piR-20,249, piR-54,022, and piR-8041 Table 5 [111].
2.27. Small Nucleolar RNAs
Unlike the most studied microRNAs and LncRNAs, small nucleolar RNAs are not widely investigated in terms of GBM [112]. The nucleolus contains a large number of long molecules called small nucleolar RNAs (snoRNAs), which associate with certain proteins to form small nucleolar RNPs (snoRNPs). They are known to participate in processing of rRNAs, the synthesis of 2′-O-methylated nucleotides and pseudouridines for post-translational processing of rRNA [113].They are classified into two main categories, one group contains the box C (RUGAUGA) and D (CUGA) motifs, whereas the other group contains the box H (ANANNA) and ACA motifs [114]. Several snoRNAs are predominantly expressed in the central nervous system [115].
Previous research indicates that snoRNAs may be dysregulated in cancer and play a role in proliferation and cell invasion [116]. SNORD44 was down-regulated in breast cancer and has prognostic significance [117]. snoRNA U50 could suppress tumor progression and is frequently deleted in prostate and breast cancer [118]. snoRNAs could work as both oncogenes and tumor suppressor genes [118,119,120]. However, research involving snoRNAs in glioma is scarce. There is not much evidence in literature regarding the possible use of snoRNAs as diagnostic or prognostic markers in glioma. SNORD47 was down-regulated in glioma tissue and correlated negatively with tumor stage [121]. [121] Xia et al. found that SNORD44 expression was downregulated in glioma cells, and suppressed the expression of metalloproteinases [116]. Chen et al. reported that downregulation of SNORD76 was associated with a malignant phenotype of glioma and the overexpression of SNORD76 in tumor xenografts was associated with a reduced tumor growth [122].
3. Discussion
3.1. Different Challenges Facing Non-coding RNAs as Biomarkers for GBM
Challenges intrinsic to the RNA biological features
As ncRNAs circulate in body fluids away from the primary brain tumor, their use as non-invasive biomarkers is readily apparent. [123]. However, RNA has an inherent unstable molecular feature, which poses logistical and technical challenges to obtain and process the samples maintaining the integrity of the RNA molecule. For example, miRNAs show inherent stability due to their extensive secondary structures while LncRNAs may require further modification to keep their stability [124,125]. For appropriate markers to be approved for clinical practice, proper standardization techniques should be applied. Setting the cutoff values of these markers is challenging due to degradation vulnerability during freezing thawing cycles, transportation and technical differences between labs [126]. The pool of ncRNAs can be altered during sample preparation due to the possibility of red blood cells hemolysis or release from the platelets during the coagulation process [127]. Rapid processing of the sample, in an absolutely RNAse-free environment, and avoiding repeated freeze thaw cycles can ensure consistent results. The stability of these molecules during various disease stages is questionable as it may be affected by chemotherapy, radiotherapy or surgical manipulation. Longitudinal studies with characteristic long term follow up intervals should be designed to determine the oscillatory changes that could affect ncRNAs levels throughout the full disease and treatment course.
Due to the lipoid nature of the extracellular vesicles carrying ncRNAs, the postprandial lipemia may alter their levels [128]. Moreover, aggregation of the vesicles and contamination with other soluble plasma lipoproteins may undermine their accuracy [129]. Recent purification techniques using bind elute chromatography (BEC), in which vesicles are highly extracted from other protein contaminants could be helpful [130]. This method can be used in the future for more accurate purification of RNAs in patients with glioma.
ncRNAs are predominantly found intracellularly, but as they travel through bodily fluids, their concentration mostly changes, which may make clinical interpretation inaccurate. Further studies are needed to determine which source is accurate and compare solid tissue biopsies versus body fluid ncRNAs levels in matched cohort studies. ncRNA subtypes differ among vertebrates and other species making animal models technically challenging [131].
3.2. Sensitivity and Specificity
For considering a molecule as a novel biomarker, the study sample should be large enough to achieve a reliable result and adequate statistical significance. Pepe et al. designed specific criteria to increase the predictive value of a diagnostic biomarker based on prospective-specimen collection, retrospective-blinded-evaluation (ProBE) for selecting samples [132]. The sample size must be accurately considered depending on the stage of the disease and the type of study. In reference to Table 2, the largest sample size in the clinical trials involving miRNAs was 500. https://beta.clinicaltrials.gov/study/NCT03770468. The sample size could be variable depending on the phase of the biomarker testing. In the preclinical phases, the sample size can be small as the statistical significance of the study in this stage is highly dependent on the type of molecule being tested rather than the sample size [133]. Next generation sequencing is commonly utilized in this stage as it can identify the entire genomic sequence of the target molecule [134]. In the validation stage, the results of the discovery phase are usually replicated, requiring larger samples to determine the specificity and sensitivity of each marker. Generally, for a marker to be appropriate for clinical use, the lower acceptable values for sensitivity and specificity should be 0.8 (80%) and 0.6 (60%), respectively [135]. miRNA-10b (mir-10b) is being tested as a biomarker for GBM in a clinical trial involving only 200 participants. In a study by Tan SK et al., the HOTAIR level was measured in 32 GBM serum samples and 40 healthy controls which is considered a relatively small sample size [48]. Zhou et al. reviewed 28 articles investigating miRNA as a marker for glioma diagnosis and the overall sensitivity was 85%, specificity was 90%, and AUC was 93% [136].
Research has been directed to increase the sensitivity and specificity of non-coding molecules for the diagnosis of cancer. To compensate for the moderate specificity/sensitivity of some circulating ncRNAs and promote their diagnostic accuracy, several studies have compiled the diagnostic values of more than a single circulating RNA candidate. For example, SPRY4-IT1, a lncRNA when combined with POU3F3 and HNF1AAS1, the sensitivity and specificity for detecting esophageal cell carcinoma significantly increased [45]. Yang et al. reported that the pool of these miRNAs was upregulated in glioma (miR-15b, miR-23a, miR-133a, miR-150, miR-197, miR-497, and miR-548b-5pl) collectively which support the use of multiple markers simultaneously [137].
Other technical methods were designed to increase the sensitivity of detection of these non coding molecules compared to the traditional RT-PCR. Morlion et al. designed a distinct lncRNA capture sequencing set that consists of 565,878 capture probes for 49,372 human lncRNA genes [138]. A new biosensor was developed for lncRNA detection called “SPCE Au NCs/MWCNT-NH2” in lung cancer [139]. Applicability of this biosensor in glioma should be investigated in future studies. A validated robust use of these molecules can be achieved by combining clinical, bioinformatic, and quantitative imaging data collectively involving different machine learning algorithms [140].
The consistent use of these ncRNAs as investigational biomarkers for glioma is still non-comprehensive. Clinical trials involving ncRNAs in glioma have traditionally had a small sample size compared with trials in other cancers, and have not focused on the correlation of these markers with different patient clinical findings. To date, there are only three clinical trials involving ncRNA as a biomarker in patients diagnosed with GBM.
4. Conclusions and Future Remarks
Timely, accurate diagnosis and monitoring treatment response in patients with high grade gliomas is vitally important to improve outcomes. Recently, ncRNAs have garnered interest as potential diagnostic and prognostic tools given their ability to be found distant to the tumor site, such as in CSF and blood. However, there are still significant hurdles before adoption of ncRNAs in clinical use. Future studies should be directed at ncRNAs in GBM as they showed promising potential to aid in GBM diagnosis, prognosis, and treatment.
Author Contributions
Dr Azab was responsible for conceptualization, study design , drafting and Dr Attallah was responsible for reviewing, editing and table design.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this article.
References
- Hazra R, Debnath R, Tuppad A. Glioblastoma stem cell long non-coding RNAs: therapeutic perspectives and opportunities. Front Genet. 2024 Jul 2;15:1416772.
- Razavi, S.M., et al., Immune Evasion Strategies of Glioblastoma. Front Surg, 2016. 3: p. 11.
- Stupp, R., et al., Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N Engl J Med, 2005. 352(10): p. 987-96.
- Wu, L. and X. Qu, Cancer biomarker detection: recent achievements and challenges. Chem Soc Rev, 2015. 44(10): p. 2963-97.
- Kapranov, P., et al., RNA maps reveal new RNA classes and a possible function for pervasive transcription. Science, 2007. 316(5830): p. 1484-8.
- Wahlestedt, C., Targeting long non-coding RNA to therapeutically upregulate gene expression. Nat Rev Drug Discov, 2013. 12(6): p. 433-46.
- Ma, L., V.B. Bajic, and Z. Zhang, On the classification of long non-coding RNAs. RNA Biol, 2013. 10(6): p. 925-33.
- Gomes, A.Q., S. Nolasco, and H. Soares, Non-coding RNAs: multi-tasking molecules in the cell. Int J Mol Sci, 2013. 14(8): p. 16010-39.
- Kondo, Y., K. Shinjo, and K. Katsushima, Long non-coding RNAs as an epigenetic regulator in human cancers. Cancer Sci, 2017. 108(10): p. 1927-1933.
- Mercer, T.R., M.E. Dinger, and J.S. Mattick, Long non-coding RNAs: insights into functions. Nat Rev Genet, 2009. 10(3): p. 155-9.
- Lu, T.X. and M.E. Rothenberg, MicroRNA. J Allergy Clin Immunol, 2018. 141(4): p. 1202-1207.
- Barciszewska, A.M., MicroRNAs as efficient biomarkers in high-grade gliomas. Folia Neuropathol, 2016. 54(4): p. 369-374.
- Ryan, B.M., A.I. Robles, and C.C. Harris, Genetic variation in microRNA networks: the implications for cancer research. Nat Rev Cancer, 2010. 10(6): p. 389-402.
- Stark, A., et al., Identification of Drosophila MicroRNA targets. PLoS Biol, 2003. 1(3): p. E60.
- Grimson, A., et al., MicroRNA targeting specificity in mammals: determinants beyond seed pairing. Mol Cell, 2007. 27(1): p. 91-105.
- Filipowicz, W., S.N. Bhattacharyya, and N. Sonenberg, Mechanisms of post-transcriptional regulation by microRNAs: are the answers in sight? Nat Rev Genet, 2008. 9(2): p. 102-14.
- Lu, J., et al., MicroRNA expression profiles classify human cancers. Nature, 2005. 435(7043): p. 834-8.
- Zhang, H., et al., Serum miR-100 is a potential biomarker for detection and outcome prediction of glioblastoma patients. Cancer Biomark, 2019. 24(1): p. 43-49.
- Dong, L., et al., miRNA microarray reveals specific expression in the peripheral blood of glioblastoma patients. Int J Oncol, 2014. 45(2): p. 746-56.
- Wu, J., L. Li, and C. Jiang, Identification and Evaluation of Serum MicroRNA-29 Family for Glioma Screening. Mol Neurobiol, 2015. 52(3): p. 1540-1546.
- Wei, X., et al., Serum MicroRNA-125b as a Potential Biomarker for Glioma Diagnosis. Mol Neurobiol, 2016. 53(1): p. 163-170.
- Lai, N.S., et al., Serum microRNA-210 as a potential noninvasive biomarker for the diagnosis and prognosis of glioma. Br J Cancer, 2015. 112(7): p. 1241-6.
- Sun, J., et al., Serum microRNA-128 as a biomarker for diagnosis of glioma. Int J Clin Exp Med, 2015. 8(1): p. 456-63.
- Yue, X., et al., Downregulation of serum microRNA-205 as a potential diagnostic and prognostic biomarker for human glioma. J Neurosurg, 2016. 124(1): p. 122-8.
- Li, H.Y., et al., Circulating microRNA-137 is a potential biomarker for human glioblastoma. Eur Rev Med Pharmacol Sci, 2016. 20(17): p. 3599-604.
- Regazzo, G., et al., A restricted signature of serum miRNAs distinguishes glioblastoma from lower grade gliomas. J Exp Clin Cancer Res, 2016. 35(1): p. 124.
- Liu, J., et al., MicroRNA-100 is a potential molecular marker of non-small cell lung cancer and functions as a tumor suppressor by targeting polo-like kinase 1. BMC Cancer, 2012. 12: p. 519.
- Lee, C., et al., Polo-like kinase 1 inhibition kills glioblastoma multiforme brain tumor cells in part through loss of SOX2 and delays tumor progression in mice. Stem Cells, 2012. 30(6): p. 1064-75.
- Xu, K., P. Liu, and W. Wei, mTOR signaling in tumorigenesis. Biochim Biophys Acta, 2014. 1846(2): p. 638-54.
- Grundmann, S., et al., MicroRNA-100 regulates neovascularization by suppression of mammalian target of rapamycin in endothelial and vascular smooth muscle cells. Circulation, 2011. 123(9): p. 999-1009.
- Tsuchiya, K., et al., Differentiation between solitary brain metastasis and high-grade glioma by diffusion tensor imaging. Br J Radiol, 2005. 78(930): p. 533-7.
- Jin, Z., et al., Serum expression level of miR-504 can differentiate between glioblastoma multiforme and solitary brain metastasis of non-small cell lung carcinoma. J buon, 2017. 22(2): p. 474-480.
- Chen, J., L. Yang, and X. Wang, Reduced circulating microRNA-203 predicts poor prognosis for glioblastoma. Cancer Biomark, 2017. 20(4): p. 521-526.
- Lan, F., et al., Serum exosomal miR-301a as a potential diagnostic and prognostic biomarker for human glioma. Cell Oncol (Dordr), 2018. 41(1): p. 25-33.
- Zhang, Y., et al., Diagnostic and prognostic significance of serum miR-145-5p expression in glioblastoma. Int J Clin Exp Pathol, 2019. 12(7): p. 2536-2543.
- Zhong, F., T. Huang, and J. Leng, Serum miR-29b as a novel biomarker for glioblastoma diagnosis and prognosis. Int J Clin Exp Pathol, 2019. 12(11): p. 4106-4112.
- Morokoff, A., et al., Serum microRNA is a biomarker for post-operative monitoring in glioma. J Neurooncol, 2020. 149(3): p. 391-400.
- Ohno, M., et al., Assessment of the Diagnostic Utility of Serum MicroRNA Classification in Patients With Diffuse Glioma. JAMA Netw Open, 2019. 2(12): p. e1916953.
- Fujimoto, A., et al., Erratum: Whole-genome mutational landscape and characterization of noncoding and structural mutations in liver cancer. Nat Genet, 2016. 48(6): p. 700.
- Schlosser, K., et al., Assessment of Circulating LncRNAs Under Physiologic and Pathologic Conditions in Humans Reveals Potential Limitations as Biomarkers. Sci Rep, 2016. 6: p. 36596.
- Isaev, K., et al., Pan-cancer analysis of non-coding transcripts reveals the prognostic onco-lncRNA HOXA10-AS in gliomas. Cell Rep, 2021. 37(3): p. 109873.
- Sun, W., et al., Regulatory mechanisms of long noncoding RNAs on gene expression in cancers. Cancer Genet, 2017. 216-217: p. 105-110.
- Kumarswamy, R., et al., Circulating long noncoding RNA, LIPCAR, predicts survival in patients with heart failure. Circ Res, 2014. 114(10): p. 1569-75.
- Yang, Y., et al., Plasma long non-coding RNA, CoroMarker, a novel biomarker for diagnosis of coronary artery disease. Clin Sci (Lond), 2015. 129(8): p. 675-85.
- Tong, Y.S., et al., Identification of the long non-coding RNA POU3F3 in plasma as a novel biomarker for diagnosis of esophageal squamous cell carcinoma. Mol Cancer, 2015. 14: p. 3.
- Siri, G., et al., A comprehensive review of the role of lncRNAs in gastric cancer (GC) pathogenesis, immune regulation, and their clinical applications. Pathol Res Pract, 2023. 241: p. 154221.
- Zhang, J.X., et al., HOTAIR, a cell cycle-associated long noncoding RNA and a strong predictor of survival, is preferentially expressed in classical and mesenchymal glioma. Neuro Oncol, 2013. 15(12): p. 1595-603.
- Tan, S.K., et al., Serum long noncoding RNA HOTAIR as a novel diagnostic and prognostic biomarker in glioblastoma multiforme. Mol Cancer, 2018. 17(1): p. 74.
- Gao, Y., et al., Long Non-Coding RNA HOXA-AS2 Regulates Malignant Glioma Behaviors and Vasculogenic Mimicry Formation via the MiR-373/EGFR Axis. Cell Physiol Biochem, 2018. 45(1): p. 131-147.
- Cantile, M., et al., Hyperexpression of HOXC13, located in the 12q13 chromosomal region, in well-differentiated and dedifferentiated human liposarcomas. Oncol Rep, 2013. 30(6): p. 2579-86.
- Croce, C.M., LINCing chromatin remodeling to metastasis. Nat Biotechnol, 2010. 28(9): p. 931-2.
- Zhang, Y., et al., Circulating long non-coding HOX transcript antisense intergenic ribonucleic acid in plasma as a potential biomarker for diagnosis of breast cancer. Thorac Cancer, 2016. 7(6): p. 627-632.
- Berrondo, C., et al., Expression of the Long Non-Coding RNA HOTAIR Correlates with Disease Progression in Bladder Cancer and Is Contained in Bladder Cancer Patient Urinary Exosomes. PLoS One, 2016. 11(1): p. e0147236.
- Zhou, X., et al., HOTAIR is a therapeutic target in glioblastoma. Oncotarget, 2015. 6(10): p. 8353-65.
- Zhang, J.X., et al., Unique genome-wide map of TCF4 and STAT3 targets using ChIP-seq reveals their association with new molecular subtypes of glioblastoma. Neuro Oncol, 2013. 15(3): p. 279-89.
- Verhaak, R.G., et al., Integrated genomic analysis identifies clinically relevant subtypes of glioblastoma characterized by abnormalities in PDGFRA, IDH1, EGFR, and NF1. Cancer Cell, 2010. 17(1): p. 98-110.
- Pastori, C., et al., The Bromodomain protein BRD4 controls HOTAIR, a long noncoding RNA essential for glioblastoma proliferation. Proc Natl Acad Sci U S A, 2015. 112(27): p. 8326-31.
- Zhang, Z., et al., Exosomal transfer of long non-coding RNA SBF2-AS1 enhances chemoresistance to temozolomide in glioblastoma. J Exp Clin Cancer Res, 2019. 38(1): p. 166.
- Bridges, C.B., Current Maps of the Location of the Mutant Genes of Drosophila Melanogaster. Proc Natl Acad Sci U S A, 1921. 7(4): p. 127-32.
- Quinonez, S.C. and J.W. Innis, Human HOX gene disorders. Mol Genet Metab, 2014. 111(1): p. 4-15.
- Tsuboi, M., et al., The transcription factor HOXB7 regulates ERK kinase activity and thereby stimulates the motility and invasiveness of pancreatic cancer cells. J Biol Chem, 2017. 292(43): p. 17681-17702.
- Wu, S.Y., et al., A miR-192-EGR1-HOXB9 regulatory network controls the angiogenic switch in cancer. Nat Commun, 2016. 7: p. 11169.
- Nagata, H., et al., Genome-wide screening of DNA methylation associated with lymph node metastasis in esophageal squamous cell carcinoma. Oncotarget, 2017. 8(23): p. 37740-37750.
- Sui, B.Q., et al., HOXB13 expression and promoter methylation as a candidate biomarker in gastric cancer. Oncol Lett, 2018. 15(6): p. 8833-8840.
- Li, B., Q. Huang, and G.H. Wei, The Role of HOX Transcription Factors in Cancer Predisposition and Progression. Cancers (Basel), 2019. 11(4).
- Wang, J., et al., LncRNA HOXA-AS2 and its molecular mechanisms in human cancer. Clin Chim Acta, 2018. 485: p. 229-233.
- Zhao, H., et al., HOX antisense lincRNA HOXA-AS2 is an apoptosis repressor in all trans retinoic acid treated NB4 promyelocytic leukemia cells. J Cell Biochem, 2013. 114(10): p. 2375-83.
- Wang, F., et al., HOX Antisense lincRNA HOXA-AS2 Promotes Tumorigenesis of Hepatocellular Carcinoma. Cell Physiol Biochem, 2016. 40(1-2): p. 287-296.
- Zheng, F.X., et al., Long noncoding RNA HOXA-AS2 promotes cell migration and invasion via upregulating IGF-2 in non-small cell lung cancer as an oncogene. Eur Rev Med Pharmacol Sci, 2019. 23(11): p. 4793-4799.
- Xie, M., et al., Long noncoding RNA HOXA-AS2 promotes gastric cancer proliferation by epigenetically silencing P21/PLK3/DDIT3 expression. Oncotarget, 2015. 6(32): p. 33587-601.
- Shen, Z.H., K.M. Zhao, and T. Du, HOXA10 promotes nasopharyngeal carcinoma cell proliferation and invasion via inducing the expression of ZIC2. Eur Rev Med Pharmacol Sci, 2017. 21(5): p. 945-952.
- Eoh, K.J., et al., Dysregulated expression of homeobox family genes may influence survival outcomes of patients with epithelial ovarian cancer: analysis of data from The Cancer Genome Atlas. Oncotarget, 2017. 8(41): p. 70579-70585.
- Lim, J.Y., et al., Overexpression of miR-196b and HOXA10 characterize a poor-prognosis gastric cancer subtype. World J Gastroenterol, 2013. 19(41): p. 7078-88.
- Kurscheid, S., et al., Chromosome 7 gain and DNA hypermethylation at the HOXA10 locus are associated with expression of a stem cell related HOX-signature in glioblastoma. Genome Biol, 2015. 16(1): p. 16.
- Sun, Y. and L. Ma, New Insights into Long Non-Coding RNA MALAT1 in Cancer and Metastasis. Cancers (Basel), 2019. 11(2).
- Kim, S.S., et al., Targeted nanocomplex carrying siRNA against MALAT1 sensitizes glioblastoma to temozolomide. Nucleic Acids Res, 2018. 46(3): p. 1424-1440.
- Huang, S.K., et al., A Panel of Serum Noncoding RNAs for the Diagnosis and Monitoring of Response to Therapy in Patients with Breast Cancer. Med Sci Monit, 2018. 24: p. 2476-2488.
- Duan, W., et al., Identification of a serum circulating lncRNA panel for the diagnosis and recurrence prediction of bladder cancer. Oncotarget, 2016. 7(48): p. 78850-78858.
- Zhou, Q., et al., lncRNAs as potential molecular biomarkers for the clinicopathology and prognosis of glioma: A systematic review and meta-analysis. Gene, 2018. 668: p. 77-86.
- Li, J., et al., Clinicopathological and prognostic significance of long noncoding RNA MALAT1 in human cancers: a review and meta-analysis. Cancer Cell Int, 2018. 18: p. 109.
- Cao, S., et al., Tumor-suppressive function of long noncoding RNA MALAT1 in glioma cells by suppressing miR-155 expression and activating FBXW7 function. Am J Cancer Res, 2016. 6(11): p. 2561-2574.
- Chung, W.Y., et al., Chromosome 11p15.5 regional imprinting: comparative analysis of KIP2 and H19 in human tissues and Wilms’ tumors. Hum Mol Genet, 1996. 5(8): p. 1101-8.
- Zhou, X., et al., Identification of the long non-coding RNA H19 in plasma as a novel biomarker for diagnosis of gastric cancer. Sci Rep, 2015. 5: p. 11516.
- Shen, L., et al., Prognostic evaluation of serum long non-coding RNA H19 for endoscopic keyhole surgery or craniotomy in glioma. Ann Clin Biochem, 2020. 57(5): p. 365-372.
- Wang, W., et al., PVT1 Promotes Cancer Progression via MicroRNAs. Front Oncol, 2019. 9: p. 609.
- Han, Y., et al., Knockdown of lncRNA PVT1 Inhibits Glioma Progression by Regulating miR-424 Expression. Oncol Res, 2019. 27(6): p. 681-690.
- Fang, J. and J. Huang, Clinical significance of the expression of long non-coding RNA PVT1 in glioma. Cancer Biomark, 2019. 24(4): p. 509-513.
- Zou, H., et al., lncRNAs PVT1 and HAR1A are prognosis biomarkers and indicate therapy outcome for diffuse glioma patients. Oncotarget, 2017. 8(45): p. 78767-78780.
- Xue, W., et al., PVT1 regulates the malignant behaviors of human glioma cells by targeting miR-190a-5p and miR-488-3p. Biochim Biophys Acta Mol Basis Dis, 2018. 1864(5 Pt A): p. 1783-1794.
- Chen, Y., et al., Long Non-coding RNA Expression Profiling Identifies a Four-Long Non-coding RNA Prognostic Signature for Isocitrate Dehydrogenase Mutant Glioma. Front Neurol, 2020. 11: p. 573264.
- You, J., et al., MicroRNA-449a inhibits cell growth in lung cancer and regulates long noncoding RNA nuclear enriched abundant transcript 1. Indian J Cancer, 2014. 51 Suppl 3: p. e77-81.
- Chakravarty, D., et al., The oestrogen receptor alpha-regulated lncRNA NEAT1 is a critical modulator of prostate cancer. Nat Commun, 2014. 5: p. 5383.
- Choudhry, H., et al., Tumor hypoxia induces nuclear paraspeckle formation through HIF-2α dependent transcriptional activation of NEAT1 leading to cancer cell survival. Oncogene, 2015. 34(34): p. 4546.
- He, C., et al., Aberrant NEAT1 expression is associated with clinical outcome in high grade glioma patients. Apmis, 2016. 124(3): p. 169-74.
- Sanger, H.L., et al., Viroids are single-stranded covalently closed circular RNA molecules existing as highly base-paired rod-like structures. Proc Natl Acad Sci U S A, 1976. 73(11): p. 3852-6.
- Goodall, G.J. and V.O. Wickramasinghe, RNA in cancer. Nat Rev Cancer, 2021. 21(1): p. 22-36.
- Wang, Z. and X. Lei, Identifying the sequence specificities of circRNA-binding proteins based on a capsule network architecture. BMC Bioinformatics, 2021. 22(1): p. 19.
- Razavi, Z.S., et al., Gynecologic cancers and non-coding RNAs: Epigenetic regulators with emerging roles. Crit Rev Oncol Hematol, 2021. 157: p. 103192.
- Khanipouyani, F., H. Akrami, and M.R. Fattahi, Circular RNAs as important players in human gastric cancer. Clin Transl Oncol, 2021. 23(1): p. 10-21.
- Zhang, X., et al., circRNA_0005529 facilitates growth and metastasis of gastric cancer via regulating miR-527/Sp1 axis. BMC Mol Cell Biol, 2021. 22(1): p. 6.
- Xing, Y., et al., Circular RNA circ-Foxo3 inhibits esophageal squamous cell cancer progression via the miR-23a/PTEN axis. J Cell Biochem, 2020. 121(3): p. 2595-2605.
- Liu, J., et al., Overexpression of circular RNA circ-CDC45 facilitates glioma cell progression by sponging miR-516b and miR-527 and predicts an adverse prognosis. J Cell Biochem, 2020. 121(1): p. 690-697.
- Cao, Q., et al., Circular METRN RNA hsa_circ_0037251 Promotes Glioma Progression by Sponging miR-1229-3p and Regulating mTOR Expression. Sci Rep, 2019. 9(1): p. 19791.
- Vagin, V.V., et al., A distinct small RNA pathway silences selfish genetic elements in the germline. Science, 2006. 313(5785): p. 320-4.
- Gainetdinov, I., et al., A Single Mechanism of Biogenesis, Initiated and Directed by PIWI Proteins, Explains piRNA Production in Most Animals. Mol Cell, 2018. 71(5): p. 775-790.e5.
- Grivna, S.T., et al., A novel class of small RNAs in mouse spermatogenic cells. Genes Dev, 2006. 20(13): p. 1709-14.
- Wang, X., et al., MiRNA-154-5p inhibits cell proliferation and metastasis by targeting PIWIL1 in glioblastoma. Brain Res, 2017. 1676: p. 69-76.
- Qu, A., et al., A serum piRNA signature as promising non-invasive diagnostic and prognostic biomarkers for colorectal cancer. Cancer Manag Res, 2019. 11: p. 3703-3720.
- Krishnan, P. and S. Damaraju, The Challenges and Opportunities in the Clinical Application of Noncoding RNAs: The Road Map for miRNAs and piRNAs in Cancer Diagnostics and Prognostics. Int J Genomics, 2018. 2018: p. 5848046.
- Jacobs, D.I., et al., PIWI-Interacting RNAs in Gliomagenesis: Evidence from Post-GWAS and Functional Analyses. Cancer Epidemiol Biomarkers Prev, 2016. 25(7): p. 1073-80.
- Jacobs, D.I., et al., piRNA-8041 is downregulated in human glioblastoma and suppresses tumor growth in vitro and in vivo. Oncotarget, 2018. 9(102): p. 37616-37626.
- Williams, G.T. and F. Farzaneh, Are snoRNAs and snoRNA host genes new players in cancer? Nat Rev Cancer, 2012. 12(2): p. 84-8.
- Hoang, C. and A.R. Ferré-D’Amaré, Cocrystal structure of a tRNA Psi55 pseudouridine synthase: nucleotide flipping by an RNA-modifying enzyme. Cell, 2001. 107(7): p. 929-39.
- Kiss, T., Small nucleolar RNA-guided post-transcriptional modification of cellular RNAs. Embo j, 2001. 20(14): p. 3617-22.
- Runte, M., et al., The IC-SNURF-SNRPN transcript serves as a host for multiple small nucleolar RNA species and as an antisense RNA for UBE3A. Hum Mol Genet, 2001. 10(23): p. 2687-700.
- Xia, X.R., et al., Effects of small nucleolar RNA SNORD44 on the proliferation, apoptosis and invasion of glioma cells. Histochem Cell Biol, 2020. 153(4): p. 257-269.
- Appaiah, H.N., et al., Persistent upregulation of U6:SNORD44 small RNA ratio in the serum of breast cancer patients. Breast Cancer Res, 2011. 13(5): p. R86.
- Dong, X.Y., et al., SnoRNA U50 is a candidate tumor-suppressor gene at 6q14.3 with a mutation associated with clinically significant prostate cancer. Hum Mol Genet, 2008. 17(7): p. 1031-42.
- Dong, X.Y., et al., Implication of snoRNA U50 in human breast cancer. J Genet Genomics, 2009. 36(8): p. 447-54.
- Okugawa, Y., et al., Clinical significance of SNORA42 as an oncogene and a prognostic biomarker in colorectal cancer. Gut, 2017. 66(1): p. 107-117.
- Xu, B., et al., SNORD47, a box C/D snoRNA, suppresses tumorigenesis in glioblastoma. Oncotarget, 2017. 8(27): p. 43953-43966.
- Chen, L., et al., SNORD76, a box C/D snoRNA, acts as a tumor suppressor in glioblastoma. Sci Rep, 2015. 5: p. 8588.
- Fanelli, G.N., et al., LONG-NONCODING RNAs in gastroesophageal cancers. Noncoding RNA Res, 2018. 3(4): p. 195-212.
- Statello, L., et al., Gene regulation by long non-coding RNAs and its biological functions. Nat Rev Mol Cell Biol, 2021. 22(2): p. 96-118.
- Hombach, S. and M. Kretz, Non-coding RNAs: Classification, Biology and Functioning. Adv Exp Med Biol, 2016. 937: p. 3-17.
- Baldassarre, A., et al., Circulating microRNAs and Bioinformatics Tools to Discover Novel Diagnostic Biomarkers of Pediatric Diseases. Genes (Basel), 2017. 8(9).
- Iempridee, T., et al., Identification of reference genes for circulating long noncoding RNA analysis in serum of cervical cancer patients. FEBS Open Bio, 2018. 8(11): p. 1844-1854.
- Onódi, Z., et al., Isolation of High-Purity Extracellular Vesicles by the Combination of Iodixanol Density Gradient Ultracentrifugation and Bind-Elute Chromatography From Blood Plasma. Front Physiol, 2018. 9: p. 1479.
- Linares, R., et al., High-speed centrifugation induces aggregation of extracellular vesicles. J Extracell Vesicles, 2015. 4: p. 29509.
- Corso, G., et al., Reproducible and scalable purification of extracellular vesicles using combined bind-elute and size exclusion chromatography. Sci Rep, 2017. 7(1): p. 11561.
- Necsulea, A., et al., The evolution of lncRNA repertoires and expression patterns in tetrapods. Nature, 2014. 505(7485): p. 635-40.
- Pepe, M.S., et al., Pivotal evaluation of the accuracy of a biomarker used for classification or prediction: standards for study design. J Natl Cancer Inst, 2008. 100(20): p. 1432-8.
- Corchete, L.A., et al., Systematic comparison and assessment of RNA-seq procedures for gene expression quantitative analysis. Sci Rep, 2020. 10(1): p. 19737.
- Auer, P.L. and R.W. Doerge, Statistical design and analysis of RNA sequencing data. Genetics, 2010. 185(2): p. 405-16.
- Ray, P., et al., Statistical evaluation of a biomarker. Anesthesiology, 2010. 112(4): p. 1023-40.
- Zhou, Q., et al., MicroRNAs as potential biomarkers for the diagnosis of glioma: A systematic review and meta-analysis. Cancer Sci, 2018. 109(9): p. 2651-2659.
- Yang, C., et al., Identification of seven serum microRNAs from a genome-wide serum microRNA expression profile as potential noninvasive biomarkers for malignant astrocytomas. Int J Cancer, 2013. 132(1): p. 116-27.
- Morlion, A., et al., Custom long non-coding RNA capture enhances detection sensitivity in different human sample types. RNA Biol, 2021. 18(sup1): p. 215-222.
- Chen, M., et al., A novel biosensor for the ultrasensitive detection of the lncRNA biomarker MALAT1 in non-small cell lung cancer. Sci Rep, 2021. 11(1): p. 3666.
- Lobato-Delgado, B., B. Priego-Torres, and D. Sanchez-Morillo, Combining Molecular, Imaging, and Clinical Data Analysis for Predicting Cancer Prognosis. Cancers (Basel), 2022. 14(13).
- Manterola, L., et al., A small noncoding RNA signature found in exosomes of GBM patient serum as a diagnostic tool. Neuro Oncol, 2014. 16(4): p. 520-7.
- Wang, Z.Q., et al., Low serum level of miR-485-3p predicts poor survival in patients with glioblastoma. PLoS One, 2017. 12(9): p. e0184969.
- Yan, Y., et al., Novel Function of lncRNA ADAMTS9-AS2 in Promoting Temozolomide Resistance in Glioblastoma via Upregulating the FUS/MDM2 Ubiquitination Axis. Front Cell Dev Biol, 2019. 7: p. 217.
- Xavier-Magalhães, A., et al., Effects of the functional HOTAIR rs920778 and rs12826786 genetic variants in glioma susceptibility and patient prognosis. J Neurooncol, 2017. 132(1): p. 27-34.
- Shen, J., et al., Serum HOTAIR and GAS5 levels as predictors of survival in patients with glioblastoma. Mol Carcinog, 2018. 57(1): p. 137-141.
- Li, J., et al., Epigenetic repression of long non-coding RNA MEG3 mediated by DNMT1 represses the p53 pathway in gliomas. Int J Oncol, 2016. 48(2): p. 723-33.
- Cai, H., et al., Long non-coding RNA taurine upregulated 1 enhances tumor-induced angiogenesis through inhibiting microRNA-299 in human glioblastoma. Oncogene, 2017. 36(3): p. 318-331.
- Cai, H., et al., The long noncoding RNA TUG1 regulates blood-tumor barrier permeability by targeting miR-144. Oncotarget, 2015. 6(23): p. 19759-79.
- Shang, C., et al., Long Non-coding RNA TUSC7, a Target of miR-23b, Plays Tumor-Suppressing Roles in Human Gliomas. Front Cell Neurosci, 2016. 10: p. 235.
- He, Z., et al., FUS/circ_002136/miR-138-5p/SOX13 feedback loop regulates angiogenesis in Glioma. J Exp Clin Cancer Res, 2019. 38(1): p. 65.
- Liu, X., et al., SRSF10 inhibits biogenesis of circ-ATXN1 to regulate glioma angiogenesis via miR-526b-3p/MMP2 pathway. J Exp Clin Cancer Res, 2020. 39(1): p. 121.
- Qu, Y., et al., Circular RNA circ_0079593 indicates a poor prognosis and facilitates cell growth and invasion by sponging miR-182 and miR-433 in glioma. J Cell Biochem, 2019. 120(10): p. 18005-18013.
- Chen, C., et al., Circular RNA Pleiotrophin promotes carcinogenesis in glioma via regulation of microRNA-122/SRY-box transcription factor 6 axis. Eur J Cancer Prev, 2020. 29(2): p. 165-173.
- Shen, S., et al., PIWIL1/piRNA-DQ593109 Regulates the Permeability of the Blood-Tumor Barrier via the MEG3/miR-330-5p/RUNX3 Axis. Mol Ther Nucleic Acids, 2018. 10: p. 412-425.
- Li, J., et al., High expression of PIWIL2 promotes tumor cell proliferation, migration and predicts a poor prognosis in glioma. Oncol Rep, 2017. 38(1): p. 183-192.
Figure 1.
miRNA synthesis. Different types of dysregulated miRNAs in GBM.
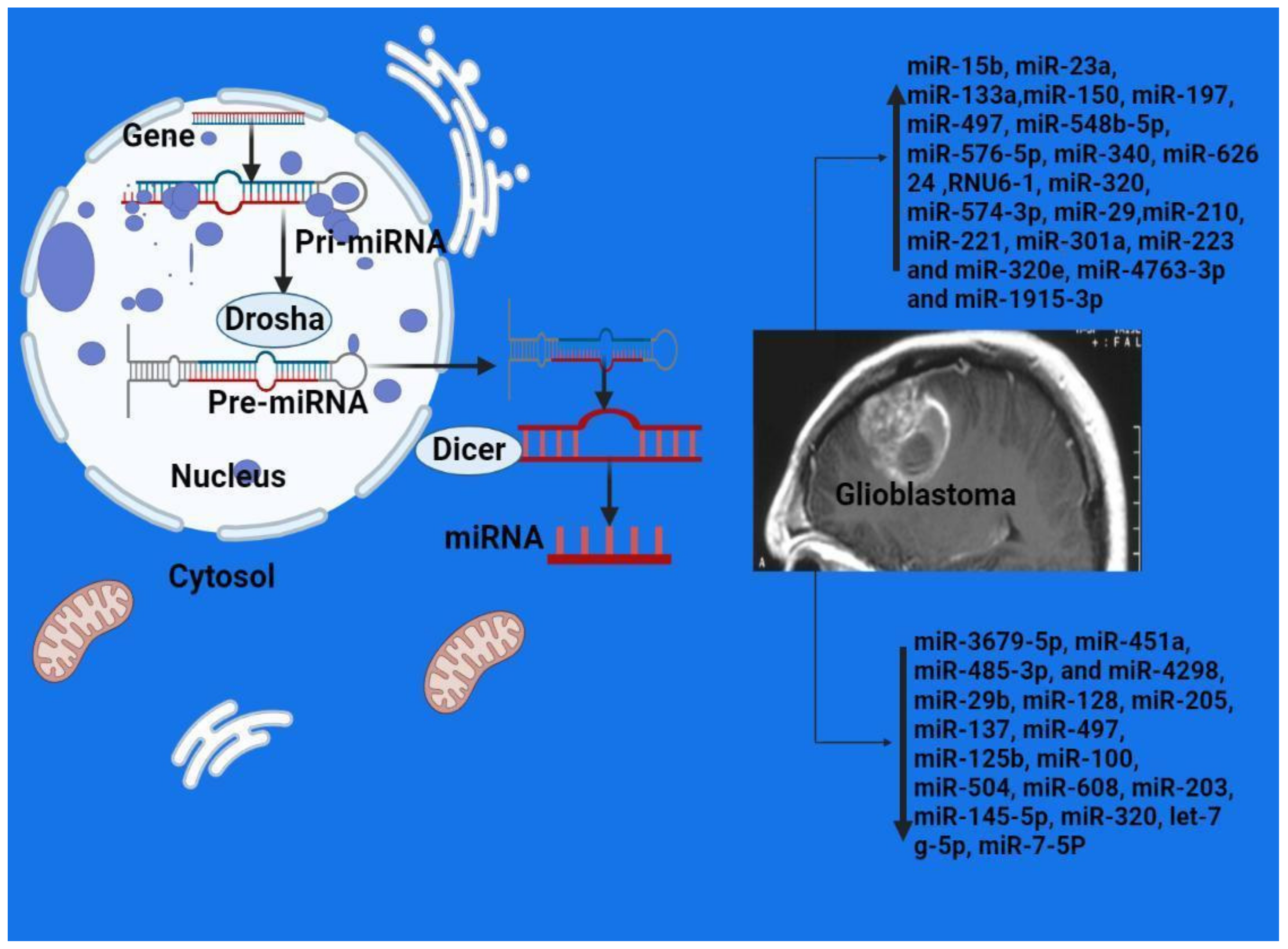

Table 1.
MicroRNAs as biomarkers for diagnostic and prognostic purposes in GBM.
| Reference | Sample | Method | Overexpressed | Downexpressed |
|---|---|---|---|---|
| [18] 2013 |
GBM+ normal control | Microarray RT-PCR | miR-15b, miR-23a, miR-133a, miR-150, miR-197, miR-497, and miR-548b-5p | |
| [137] 2014 |
GBM+ normal control | Microarray RT-PCR | miR-576-5p, miR-340, and miR-626 24 | miR-320, let-7 g-5p, and miR-7-5P |
| [19] 2014 |
Glioma and normal controls | Microarray RT-PCR | RNU6-1, miR-320, and miR-574-3p | |
| [141] 2015 |
Glioma | RT-PCR | miR-29 | |
| [20] 2014 |
Glioma | RT-PCR | miR-125b | |
| [21] 2015 |
Glioma and normal controls | RT-PCR | miR-210 | |
| [22] 2015 |
Glioma and normal controls | RT-PCR | miR-128 | |
| [23] 2015 |
Glioma | RT-PCR | miR-205 | |
| [24] 2016 |
Glioma and normal controls | RT-PCR | miR-137 | |
| [25] 2016 |
Glioma and normal controls | RT-PCR | miR-497 and miR-125b | |
| [17] 2019 |
Glioma and normal controls | RT-PCR | miR-100 | |
| [26] 2017 |
Glioma and normal controls | RT-PCR | miR-221 | miR-504 and miR-608 |
| [32] 2017 |
Glioma and normal controls | RT-PCR | miR-203 | |
| [33] 2018 |
Glioma and normal controls | RT-PCR | miR-301a | |
| [34] 2019 |
Glioma and normal controls | RT-PCR | miR-145-5p | |
| [35] 2019 |
Glioma and normal controls | RT-PCR | miR-29b | |
| [36] 2017 |
Glioma and normal controls | RT-PCR | miR-451a, miR-485-3p, and miR-4298 | |
| [142] 2020 |
Glioma and normal controls | dd-PCR | miR-223 and miR-320e | |
| [37] 2020 |
Glioma and normal controls | RT-PCR | miR-4763-3p and miR-1915-3p | miR-3679-5p |
Table 2.
Examples of clinical trials involving microRNAs as possible diagnostic and prognostic tools.
Table 2.
Examples of clinical trials involving microRNAs as possible diagnostic and prognostic tools.
| Micro-RNA or/Targeted Micro-RNA | Official Title | Start Date | Study Type | Number Enrolled | Recruitment Status | Clinical Trial Number | Reference |
|---|---|---|---|---|---|---|---|
| Micro-RNAs level | Molecular Genetic, Host-derived and Clinical Determinants of Long-term Survival in Glioblastoma | July 5, 2015 | Observational | 599 | Active Non-recruiting | NCT03770468 | https://beta.clinicaltrials.gov/study/NCT03770468. accessed on 10 December 2022. |
| miRNA-10b | Evaluating the Expression Levels of MicroRNA-10b in Patients with Gliomas | May 2013 | Observational | 200 | Recruiting | NCT01849952 | https://www.clinicaltrials.gov/ct2/show/NCT01849952. accessed on 10 December 2022 |
| MicroRNAs level | Clinical Evaluation of a New Cancer Diagnosis Center at Kristianstad General Hospital, Sweden (CPF-DC) | October 2012 | Interventional (Clinical Trial) | 388 | Completed | NCT01709539 |
https://beta.clinicaltrials.gov/study/NCT01709539?tab=results# publications. accessed on 10 December 2022 |
Table 3.
The most studied LncRNAs in GBM.
| LncRNA Type | Cell Line | Method | Target Pathway |
Suggested Prognostic/Diagnostic Value | Reference |
|---|---|---|---|---|---|
| ADAMTS9-AS2 | T98G and U118 human glioma cell lines | qRT-PCR | mTOR pathway | Promotion of cell proliferation, migration and invasion Role in TMZ resistance | [143] |
| HOTAIR | U87MG and human glioma samples | qRT-PCR | Cell cycle and proliferation pathway | HOTAIR is overexpressed in GBM, it sustains cell proliferation. | [57] |
| HOTAIR | Serum exosomes | qRT-PCR | Cell cycle and proliferation pathway | Its expression was significantly correlated with high grade brain tumors. Its serum level from GBM patients was significantly higher than in the healthy volunteers. | [48] |
| HOTAIR | Human glioma tissue | qRT-PCR and RFLP | Cell cycle and proliferation pathway | HOTAIR SNPs rs920778 and rs12826786 do not play a significant role in glioma susceptibility and were associated with better survival of patients diagnosed with WHO grade III anaplastic oligodendroglioma. | [144] |
| HOTAIR | Patient serum | qRT-PCR | Cell cycle and proliferation pathway | High levels of HOTAIR were associated with higher mortality rates. | [145] |
| HOTAIR | Human glioma tissue and LN229 and U87 |
Gene set enrichment analysis |
Cell cycle and proliferation pathway | HOTAIR expression was related to higher glioma grade and poor prognosis. | [47] |
| HOXA10-AS | G432 and G797 and human glioma tissue | Machine learning | Unknown | HOXA10-AS activation is a robust marker of poor prognosis and can predict IDH1/2 mutations. | [41] |
| SBF2-AS1 | U87, LN229, A172, T98, U251, and (HEK) 293 T cells | qRT-PCR and FISH assays | Unknown | SBF2-AS1 was upregulated in TMZ-resistant GBM cells, and overexpression of SBF2-AS1 promoted resistance to TMZ. | [58] |
| MALAT1 | U87 and SHG139 and human glioma tissue | RT-qPCR | MAPK pathway | Expression of MALAT1 was significantly decreased in glioma specimens. MALAT1 expression was correlated with tumor size, WHO grade, and clinical outcome. | [80,81] |
| MEG3 | U251, U87 and A172 human glioma cell lines | RT-qPCR | p53 pathway PTEN pathway | Inhibition of glioma cell growth | [146] |
| PVT1 | Human glioma tissue | qRT-PCR | Cell cycle pathway BMP pathway NOTCH pathway | The expression levels of PVT1 were significantly higher in GBM tissues than in normal tissues, and PVT1 level was closely related to tumor grade. | [87] |
| PVT1 | Human glioma tissue | qRT-PCR | Cell cycle pathway, BMP pathway NOTCH pathway | High expression of PVT1 was associated with high Ki-67 level and more TP53 mutation and indicates poor survival. | [88] |
| PVT1 | U87 and U251 and human embryonic kidney cell line (HEK-293T) | qRT-PCR | Cell cycle pathway BMP pathway NOTCH pathway | PVT1 was upregulated in glioma specimens and cell lines. | [89] |
| NEAT1 | Human glioma tissue | qRT-PCR | WNT/β-Catenin pathway PI3K/AKT/mTOR pathway MEK/ERK pathway | Patients with high NEAT1 expression showed unfavorable overall survival. | [94] |
| TUG1 | Human glioma tissue and U251 MG, U87MG | qRT-PCR | WNT/β-Catenin pathway EMT pathway Neuronal differentiation pathway | Promotion of cell proliferation, migration and invasion regulation of glioma cell stemness enhances VEGF expression and angiogenesis | [147,148] |
| TUSC7 | Human glioma tissues and U251 , U87 | qRT-PCR | PI3K/Akt pathway | Inhibition of cell proliferation, migration and invasion | [149] |
Table 4.
Dysregulated circRNAs in GBM and its possible functions.
| Authors | Type | Expression Level | Cell Line | Mechanism | Target Molecule | Reference |
|---|---|---|---|---|---|---|
| Cao et al. | hsa_circ_0037251 | Upregulated | U373, U251 (HEK) 293T cells | Inhibits apoptosis | miR-1229-3 mTORp | [103] |
| He et al., | circ_002136 | Upregulated | U87 | Promotes angiogenesis | miR-138-5p/SOX13 | [150] |
| Liu et a, | circ-ATXN1 | Upregulated | U87 | Promotes angiogenesis | miR-526b-3p/VEGFA | [151] |
| Qu et al., | circ_0079593 | Upregulated | U118, U251, U87MG, and LN229 | Prognostic marker | miR-182 | [152] |
| Chen et al., | circ_PTN | Upregulated | U251 and U87 | Promotes cellular proliferation | miR-122/SOX6 | [153] |
Table 5.
Examples of piRNAs that have variable expression levels in GBM.
| piRNA type | Cell Line | Expression Level | Reference |
|---|---|---|---|
| piR-15,988 | Human glioma tissue | Downregulated | [111] |
| piR-54,022 | Human glioma tissue | Downregulated | [111] |
| piR-8041 | Human glioma tissue | Downregulated | [111] |
| piR-DQ593109 | U87 cell line and HEK293T | Upregulated | [154] |
| PIWIL1 | U87 cell line and HEK293T | Upregulated | [154] |
| piR-598 | U87 and A172 and human glioma tissue | Downregulated | [110] |
| PIWIL2 | H4, A172, U251, U87, and U118 and human glioma tissue | Upregulated | [155] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



