
Submitted:
31 March 2025
Posted:
02 April 2025
You are already at the latest version
Abstract
Graphene-based materials (GBMs) have shown significant promise in cancer therapy due to their unique physicochemical properties, biocompatibility, and ease of functionalization. Their ability to target solid tumors, penetrate the tumor microenvironment (TME), and act as efficient drug delivery platforms highlights their potential in nanomedicine. However, the complex and dynamic nature of the TME, characterized by metabolic heterogeneity, immune suppression, and drug resistance, poses significant challenges to effective cancer treatment. GBMs offer innovative solutions by enhancing tumor targeting, facilitating deep tissue penetration, and modulating metabolic pathways that contribute to tumor progression and immune evasion. Their functionalization with targeting ligands and biocompatible polymers improves their biosafety and specificity, while their ability to modulate immune cell interactions within the TME presents new opportunities for immunotherapy. Given the role of metabolic reprogramming in tumor survival and resistance, GBMs could be further exploited in metabolism-targeted therapies by disrupting glycolysis, mitochondrial respiration, and lipid metabolism to counteract the immunosuppressive effects of the TME. This review focuses on discussing research studies that design GBM nanocomposites with enhanced biodegradability, minimized toxicity, and improved efficacy in delivering therapeutic agents with the intention to reprogram TME for effective anticancer therapy. Additionally, exploring their potential of GBM nanocomposites in combination with immunotherapies and metabolism-targeted treatments could lead to more effective and personalized cancer therapies. By addressing these challenges, GBMs could play a pivotal role in overcoming current limitations in cancer treatment and advancing precision oncology.
Keywords:
graphene
; nanomaterials
; graphene oxide
; cancer
; photothermal therapy
; photodynamic therapy
1. Introduction
Graphene (Gr) and graphene-based materials (GBMs), such as graphene oxide (GO) and reduced graphene oxide (rGO) have gained significant attention in biomedical research, particularly for their potential in targeting solid tumors. Their application in cancer therapy and diagnostics (theranostics) stems from their unique physical, chemical, and biological properties [1]. GBMs owing to their exceptionally high surface area support efficiently the attachment of various therapeutic agents, including chemotherapeutic drugs [2], RNA molecules (like siRNA and mRNA) [2,3], or anticancer proteins and peptides [4], enhancing their effectiveness and potentially reducing side effects [5]. A focal point of GBMs research has been the ease of their functionalization with various molecules and ligands to improve tumor targeting specificity [3]. Particularly, the surface functionalization of GBMs has gained significant interest in enhancing or tailoring their biocompatible properties and optimizing their biosafety, since their toxicity profile may strongly vary depending on size, surface charge, and hydrophobicity [6,7]. The covalent and non-covalent modification of GBMs with biocompatible polymers and co-polymers, such as polyethylene glycol (PEG) [8], dextran [4], physical or hemisynthetic polysaccharides [9], and poly(lactic acid-co-ethylene glycol) (PLA-co-PEG) [10], has substantially improved their biological behavior in terms of solubility and colloidal stability [4], and improved the cellular accumulation and toxicity in tumor cells and normal cells [11]. The surface functionalization of GBMs for exploiting their effectiveness in biomedical applications has been an issue of interest in extensive reviews recently [1,2,3,4,5,6,7,8,9,10,11]. Especially, the surface modification with targeting ligands such as antibodies, peptides, or small RNA molecules can allow GBMs to increase their selectivity and binding affinity to tumor cell receptors that play an intimate role in carcinogenesis and tumor progression [2,3,12]. Such tumor-specific therapies improve the accumulation of therapeutic agents at the tumor site, while minimizing off-target side effects.
GBMs have emerged as key driving nanomaterials for overcoming tumor microenvironment (TME) biological barriers to promote deep tumor penetration and anticancer effectiveness (Figure 1). Their small size and ultrathin 2D plane provided them the ability to intercalate into tumor cell membranes and navigate through TME extracellular matrix (ECM) enhancing their therapeutic efficacy [13]. TME is a highly differentiating network with altered metabolic status favoring immune metabolic reprogramming, thus contributing to tumor cells’ differentiation and immune cells’ suppression [14]. The protein-rich ECM of TME consists of cell-secreted factors, including growth factors, cytokines, chemokines, and tumor antigens supporting cancer cells differentiation and mutation. In this ECM, cancer cells continuously escape immune surveillance, eventually leading to tumor growth and progression. ECM, abnormal vasculature, and interstitial matrix represent the non-cellular components of tumor stroma that coexist alongside the cellular components that most commonly include the capillary and vascular cells, immune cells (innate and adaptive), mesenchymal stromal cells (MSC) and cancer-associated fibroblasts (CAFs) [13,14,15]. The tumor stroma ECM acts as a steppingstone for the evolution of the metabolic alterations of tumor cells by supplying nutrients and growth factors highly supporting the formation of metabolic networks between the stromal and the surrounding cancer cells [16]. Such metabolic networks offer support for the transfer of signaling molecules (metabolites) from stromal cells (immune and non-immune) to neighboring cancer cells further promoting their proliferation, survival, and resistance. The stroma derived metabolites serve as intermediates of paracrine signaling for the support of metabolite-responsive signaling pathways [17]. The metabolic support from stroma cells by paracrine signaling most commonly involves the i) CAFs that mainly secrete alanine and supply with glutamine to promote cancer cells’ differentiation, ii) peripheral axons that secrete serine to regulate mRNA translation and iii) macrophages that secrete pyrimidines promoting drug resistance [16,17]. Moreover, through the activation of glycolysis and glutamine metabolism tumor stroma cells (especially CAFs) manage to create a reservoir of abundant molecules supply to promote metabolic flexibility and allow cancer cells to adjust their metabolic phenotypes adapting in TME [18]. This metabolic plasticity in combination with the increased oncogenic signaling within TME supports tumor cells survival and drug resistance limiting the efficacy of therapeutic agents (chemo- and immuno-therapies).
The increased cellular heterogeneity of TME and especially of tumor stroma, highly promotes the metabolic heterogeneity and reprogramming of cancer and stromal cells resulting in the metabolic immune suppression of TME [19]. Within TME, cancer and stromal cells are susceptible to metabolic stress due to their increased energy and nutrients requirements. The factors contributing to metabolic stress mainly include the increased competition in areas with high metabolic activity and the limited oxygen and nutrient supply, owing to the abnormal vasculature [20]. Thus, cancer cells fuel their bioenergetics demands by alternative processes, such as aerobic glycolysis for the conversion of glucose to lactate, highly affecting the extracellular levels of both metabolites and resulting in the upregulation of glycolytic enzymes, pH gradient, and hypoxia signaling [21]. Moreover, cancer cells use glutaminolysis for glutamine conversion to glutamate that is further catalyzed to αKG (alpha-ketoglutarate) from glutamate dehydrogenase (GDH) to enable mitochondrial ATP production. For cancer cells bioenergetics, glutamine via the tricarboxylic acid (TCA) cycle represents the main substitute source of carbon supply for the biosynthesis of lipids and metabolites and serves as a nitrogen supplier for the biosynthesis of amino acids and nucleotides [22]. Lipogenesis and lipolysis represent another source contributing to the energetic supply of cancer cells through the de novo synthesis of mono-unsaturated fatty acids (FAs) and the lipoprotein lipase (LPL)-mediated extracellular lipolysis for the additional supply with FAs, being essential building blocks of lipids and cholesterol synthesis [23,24]. Lipid metabolic reprogramming has been characterized as an essential cascade for cancer cells’ supply of FAs to support their increased growth and proliferation demands [24], and for stromal cells’ (especially immune cells) adaptability in the TME [23]. The immune cells (innate and adaptive) exploit the same bioenergetics resources for their function, and intercellular and intracellular communications [25]. The metabolic heterogeneity and the extensive crosstalk among cancer and stromal cells determine the fate of nutrients and metabolites within TME, shaping cancer cells’ progression and immune cells’ suppression. A distinct example is the increased production of reactive oxygen species (ROS) by the cancer cells that regulate the activation of CAFs, further stimulating aerobic glycolysis in CAFs. This process promotes the release of lactate from CAFs via monocarboxylate transporter (MCT) MCT4 that is transported to cancer cells by MCT1, leading to metabolic recycling [26]. The competitiveness among cancer and immune cells for nutrients and metabolites supply, in combination with the excessive consumption by the cancer cells, regulates the suppression of immune cells’ function [27]. In a study by Chang et al. [28], it was presented that glucose competition in the TME of mouse sarcoma model resulted in the increased glucose consumption by the cancer cells and the metabolic restriction of tumor-infiltrating CD8+ T lymphocytes (TILs) downregulating T cells responsiveness. Specifically, the mTOR activity, glycolytic capacity and IFN-γ expression of T-cells were impaired, promoting tumor growth and progression. In a recent study by Kim et al. [29] in lung adenocarcinoma (ADC) tissues from patients, cancer cells’ glycolysis was associated with increased hexokinase-2 (HK2) gene expression that promoted an immunosuppressive and pro-tumorigenic state downregulating CD8+ T-cells infiltration and increasing regulatory T-cells (Treg) infiltration. Fewer CD8+ T-cells and a low ratio of CD8+ T-cells to Tregs were associated with poor patients’ survival, signifying the importance of glycose metabolism in immune system suppression. Moreover, in a recent review by Qian et al. [30] it highlighted that tumor-associated macrophages (TAMs) heterogeneity and M2-polarization further promoted TME immunosuppressive effects and immune evasion of cancer cells, supporting drug resistance and metastasis. In this metabolically heterogeneous and immunosuppressive TME, immune evasion and drug resistance highly influence the effectiveness of immunotherapies and targeted therapies. By targeting metabolic pathways, such as aerobic glycolysis (Warburg effect), mitochondrial respiration, hypoxia and oxidative stress, novel strategies are offered in metabolism-regulated therapies for targeting TME [31].
2. Graphene-Based Nanocomposites in Reprogramming of TME
The therapeutic potential upon targeting TME has been extensively reviewed in terms of responsive targeted nanomedicines [15,32] that explore the metabolic immune reprogramming [33], immunotherapies [34], the inflammatory microenvironment [36], and the microvasculature [36]. Within TME, the cancer cells dispose of the efficiently healthy metabolic pathways and follow alternative routes that through metabolic reprogramming meet their biosynthesis and survival requirements [21]. The metabolic reprogramming of the TME affects the phenotype of cancer and stromal cells, including CAFs and immune cells, regulating cancer cells’ growth and immune cells’ suppression. The cellular crosstalk within the metabolic network of TME is supported by signaling molecules and oncogenes playing a vital role in cellular communications. Recently, there has been a great effort in targeting the metabolic pathways of TME by exploiting the advantages of nanomedicine [31]. The main obstacles in targeting TME metabolic pathways, to impair and exhaust cancer cells bioenergetics, emerged from the complexity of the metabolic network that require the activity and regulation of multiple metabolites, enzymes, and proteins. Moreover, metabolic reprogramming can be highly affected by the communications among cancer and noncancerous cells (such as non-cancerous epithelial cells, immune cells) of the TME, providing extra difficulties in metabolic targeting. The metabolic flexibility of TME offered another hallmark in metabolic targeting, highlighting the necessity of tumor metabolome analysis in patients [37,38].
However, the understanding of the role of oncogenes and oncogenic proteins in regulating cancer cells’ metabolism and TMEs’ metabolic reprogramming provided a profound alternative in targeting cancer metabolism [38]. The role of oncogenes is of vital importance in the proliferation of cancer cells for the activation of nutrients uptake, energy supply (through ATP biosynthesis) and the biosynthesis of proteins and lipids. In contrast to normal cells that are mainly based on mitochondrial oxidative phosphorylation (OXPHOS) for their energy supply during differentiation processes, cancer cells accumulate energy for their differentiation through aerobic glycolysis to avoid ROS formation (Warburg Effect). The switch to excessive glucose metabolism accompanied by glutamine and glycine influx by cancer cells has participated in the establishment of a hypoxic metabolic milieu [31,38,39]. The understanding of the underlying mechanisms of cancer metabolism and the complex metabolic reprogramming is still to be conquered, however the metabolites, enzymes, proteins, and transporters that take part in metabolic pathways represent useful therapeutic targets for nanomedicine therapeutics. By recognizing the vulnerabilities of TME metabolic milieu it becomes possible to lead cancer cells to energy deprivation by restraining their biochemical resources, to enhance the effectiveness of anticancer therapies and immunotherapies [31]. The application of nanotechnology in modulating TME metabolic reprogramming has been a revolution in view of the combinational therapies arising from exploiting the effects of metabolic inhibitors for cancer cells energy deprivation and anticancer therapies (Figure 2) [40,41].
The physicochemical properties of GBMs extend beyond the densely packed pristine hexagonal lattice and the π-π stacking and hydrophobic interactions. The covalent and non-covalent functionalization of GBMs’ surface has been extensively studied for increasing their biocompatibility, bioavailability, hydrophilicity, and facilitating the transportation of hydrophobic drug molecules, genes, and antibodies for targeted anticancer drug delivery [2,3]. Among GBMs, the oxidized forms of graphene such as GO, CGO, and rGO have presented excellent adhesion properties upon interaction with ECM proteins, genes, and growth factors of TME [4,5]. The amplified cell adhesion ability of GBMs is orchestrated by electrostatic and hydrophobic interactions, accompanied by covalent and hydrogen bonding. Such properties of GBMs have attracted considerable interest in their application on diagnostic biosensors and imaging devices for targeting crucial cancer biomarkers and tumor imaging, respectively [42,43]. Moreover, the significant applications of GBMs in photothermal (PTT) and photodynamic (PDT) combinational anticancer therapies are the main research interest of extended reviews [9]. GBMs, owing to their ability to absorb near infrared radiation (NIR) light and transmit heat, manage to induce local heat generation (hyperthermia) and promote thermal ablation to cancer cells. Also, GBMs have been extensively studied for PDT acting as photosensitizers due to their light absorption ability. Within the cellular TME, the GBMs transmit the absorbed energy generating ROS (such as singlet oxygen, superoxide anion radicals, hydroxyl radicals, and hydrogen peroxide) at cytotoxic levels promoting oxidative stress to cancer cells [44]. In the photothermal and photodynamic effects of GBMs, mitochondrial respiration, pH and hypoxic milieu, and the graphene-cell interactions highly affect the ROS production [45]. Such interactions have also been crucial for the development of metabolic heterogeneity within TME. However, little is known for the underlying mechanism of oxidative stress and thermal ablation of cancer cells by GBMs in combination with NIR light. In a recent review, the ability of GBMs to induce and modulate autophagy and the molecular mechanisms of graphene-cells interactions are highlighted [40]. Autophagy is a critical process for cancer cells to promote their survival within the heterogenic TME to overcome nutrients and metabolites starvation, by using autophagy-mediated recycling to maintain mitochondrial function and energy homeostasis [46,47]. Thus, autophagy inhibition by GBMs may be a beneficial target for metabolic reprogramming of TME. In a recent study by Zhang et al. [48], the effects of thermal ablation in plasma samples from patients with non-small cell lung cancer (NSCLC) revealed the significant impact on the regulation of metabolites associated with angiogenesis and inflammatory responses. CT-guided percutaneous microwave ablation was performed to patients after administration of local anesthesia with a needle reaching the tumor lesion and ablation being performed for 6-9 min at a 40-60 W power depending on the lesion size. Thermal ablation reduced the expression levels of pro-tumor associated proteins (angiopoietin 1, ANGPT1, tyrosine kinase, TIE2) and metabolic pathways by promoting a decreased ratio of metabolites (fatty acids, palmitic acid, and eicosapentaenoic acid) and steroid-related metabolites (corticosterone, cortisone, and cortisol) that participate in tumor angiogenesis and inflammatory responses. According to this study, after thermal ablation, pro-inflammatory proteins and metabolites induced elevated systemic inflammatory responses.
Highly active metabolic pathways create critical alteration in the supply of nutrients and small molecules within TME. For example, aerobic glycolysis is mainly used for the metabolism of glucose by mitochondria. The oxidation of glucose, fatty acids and amino acids by mitochondria is promoted by the TCA cycle and the electron transport chain (ETC). This way excessive amounts of energy are produced for cancer cells’ proliferation and differentiation. Aerobic glycolysis is favored by cancer cells due to the rapid glucose metabolism accompanied by carbon and NAD+ (Nicotinamide adenine dinucleotide) regeneration that is a critical metabolite for cellular homeostasis [49]. NAD+ is formed by the linkage of adenosine monophosphate (AMP) and nicotinamide mononucleotide (NMN). The phosphorylation of AMP results in the formation of NADP+ (nicotinamide adenine dinucleotide phosphate). In the nicotinamide group of NAD+ and NADP+ the carbon atom of the pyridine ring in the opposite position of the primary amide (meta position) can accept a hydride anion (H+, 2e-) and provide the reduced forms of NADH and NADPH. The reduced forms (NADH and NADPH) undergo oxidation through the ETC to promote the synthesis of ATP (adenosine triphosphate). The intermediates of aerobic glycolysis (such as 3-Phosphoglycerate, glucose-6-phosphate, fructose-6-phosphate) in combination with NAD+/NADP+ cofactor reduction are essential participants in the pentose phosphate pathway (glucose-6-phosphate), serine synthesis (3-Phosphoglycerate) and fatty acids synthesis (pyruvate) [50]. Another essential metabolic pathway regulated by mitochondria is the hexosamine biosynthesis pathway (HBP) being vital for the synthesis of nucleotide sugars that play a fundamental role on the glycosylation of proteins and lipids. The glucose-6-phosphate and fructose-6-phosphate can also be metabolized by the HBP pathway by glutamine as an amide donor. Glutamine represents the main reservoir of nitrogen for amino acids and nucleic acids biosynthesis. Thus, glucose and glutamine metabolism are highly associated with cancer cells proliferation. Apart from the TCA cycle, ETC is essential to ATP production and mitochondrial aspartate formation by catabolism of glutamine in cancer cells. Aspartate and malate are the two major metabolites deriving from mitochondrial glutamine oxidation within the Krebs cycle (KC) functioning in normal cells. These metabolites are used for NADPH production that is crucial in anabolic processes and redox homeostasis. For aspartate production, glutamine enters the KC as α-ketoglutarate (α-KG) to be enzymatically converted to glutamate by glutaminase (GA). Glutamate is then further converted to α-KG by glutamate dehydrogenase (GLUD1), glutamate-oxaloacetate transaminase (GOT2) and glutamate pyruvate transaminase (GPT2) further releasing ammonia, aspartate and alanine, respectively. In cancer cells metabolism, GPT1 (cytosolic) and GPT2 (mitochondrial) are essential for energy metabolism by providing alanine for protein biosynthesis and feeding TCA cycle intermediates. Moreover, GOT acts as a pool by providing cytosolic aspartate for protein and nucleotide synthesis. Additionally, glutamine is necessary for the de novo enzymatic synthesis of asparagine from aspartate, by the action of asparagine synthetase (ASNS) that activates mTORC1 (mammalian target of rapamycin mTOR complex 1), further contributing to the synthesis of purines and pyrimidines [51]. With the significant increase in the application of immunotherapies the association among infiltrating immune cells and cancer cells has emerged. In this aspect, the critical role of metabolic pathways in the regulation of immune responses has been established [52]. The dysregulation of aerobic glycolysis and HBP pathways in cancer cells has a great impact on the immune cells’ signaling, cytokine production and oncogenes regulation thus influencing immune suppression. To facilitate TME metabolic reprogramming, tumor metabolism has been encountered as a checkpoint promoting immune suppression. To this aspect, metabolic inhibitors have been studied for targeting tumor metabolism by the blockade of glucose and glutamine metabolism for cancer cells energy exhaustion [49,50,51,52].
2.1. Graphene and Graphene Quantum Dots
The generation of mitochondrial ROS by the activation of metabolic pathways (TCA cycle and ETC) in cancer cells plays an essential part in tumor progression and metastasis. The elevated ROS formation is regulated by the cytosolic antioxidant system, that includes enzymatic antioxidants such as SOD (superoxide dismutase), CAT (catalase), GPX (glutathione peroxidase), TXN-TXNRD (thioredoxin - thioredoxin reductase), and non-enzymatic antioxidants such as GSH (glutathione), vitamins, selenium and metabolites (bilirubin, melatonin). The enzymatic and non-enzymatic antioxidants create an antioxidant network that regulates the expression of critical oncogenes (mutant KRAS and MYC genes) leading to the suppression of the antioxidant defense mechanism. In cancer progression, the presence of increased ROS expression levels can promote genomic and metabolic instability by the inactivation of tumor suppressor genes and mitogenic signaling pathways. However, a critical threshold exists in cancer cells being regulated by the imbalance between ROS production and cellular antioxidants presence in the cytosol. This imbalance may cause increased oxidative stress damaging proteins, lipids and DNA eventually leading to oxidative cancer cell death [53,54]. Thus, the dysfunction of mitochondrial ROS formation is a critical determinant of cancer therapeutics being targeted by disrupting the OXPHOS mechanism, by scavenging the ROS production, and by promoting oxidative stress cell death (Figure 3). Table 1 presents characteristic research studies.
Among GBMs, graphene (Gr) and graphene quantum dots (GQDs) have been investigated for ROS-targeting due to their superior photothermal/photodynamic light conversion ability after UV/Vis and NIR light radiation. Especially, GQDs have been associated with significant mitochondria accumulation ability due to their size (< 10 nm) and their surface functionalization capacities. In a recent study by Perini et al. [55] the functionalization of GQDs had great impact on membranes’ fluidity upon their interaction with U87MG glioblastoma cancer cells and glioblastoma neurospheres. The GQDs and the carboxylated-GQDs significantly increased membrane fluidity thus influencing cell membrane stability. This effect was associated with the negative surface charge of the GQDs that destabilized cell membrane and reduced the formation of neurospheres. Membrane fluidity is a critical aspect for cancer cells’ progression and metastasis, since the altered lipid metabolism promotes lipid-protein heterogeneities in the cell membrane composition. Particularly interesting is the expression of phosphatidylcholine that regulates the expression levels of enzymes and energy metabolites, promoting cancer cell progression. Moreover, the composition of the cellular membrane in highly metastatic cancer cells is associated with reduced cholesterol content thus increasing, their plasticity and fluidity to promote blood vessel penetration [56].

Table 1.
Applications of graphene (Gr) based nanocomposites in cancer therapeutics.
| Carrier Type | Agent | Characteristics | Ref |
|---|---|---|---|
| GQDs Carboxylated-GQDs |
- | increased membrane fluidity reduced formation of neurospheres in U87MG glioblastoma cancer cells and glioblastoma neurospheres |
[55] |
| Carboxylated-GQDs | DOX, TMZ | synergistic PTT on 3D spheroid model of glioblastoma, elevated intracellular ROS production, increased membrane permeability, elevated presence of tumor-associated antigens | [57] |
| nitrogen-doped GQD with HA and Fc | HA, Fc | CD44 cancer cell receptor targeting in HeLa cells, Fc facilitated a redox-based toxicity by the redox cycle of iron, oxidative stress targeting | [59] |
| nitrogen-doped GQD | TPP, ruthenium nitrosyl | mitochondria targeting, PTT, regulation of oxygen consumption and ATP synthesis, an inhibitory effect in the ETC, inhibited the in vivo tumor growth | [64] |
| GQDs | TPP | in vitro mitochondria monitoring | [68] |
| Hybrid GQDs - UCNP | TRITC | promoted increased in situ cytotoxic ROS formation, mitochondria dysfunction, decrease in mitochondria membrane potential, activation of caspase 3 apoptotic pathway | [69] |
| Graphene | GA | promoted the depletion of MMP, decreased the intracellular level of lipid droplets, induced DNA fragmentation | [72] |
| Pristine graphene | - | promote apoptotic mitochondria pathway signaling in murine RAW 264.7 macrophages, triggered increased ROS levels, activated Bcl-2 family pro-apoptotic proteins, activated MAPK and TGF-β signaling, activated caspace-3 related apoptotic pathway | [76] |
Abbreviations: PTT: Photothermal Therapy, DOX:doxorubicin, TMZ: temozolomide, ROS: reactive oxygen species, HA: hyaluronic acid, Fc: ferrocene, TPP: triphenylphosphonium, ETC: electron transport chain, UCNP: upconversion nanoparticles, TRITC: Tetramethylrhodamine-5-isothiocyanate, GA: gambogic acid, MMP: mitochondria membrane potential.
In another study, Perini et al. [57] employed carboxylic acid functionalized GQDs in combination with doxorubicin (DOX) and temozolomide (TMZ) for the synergistic PTT effect on a 3D spheroid model of glioblastoma. TMZ is an alkylating agent applied as a primary therapy for glioblastoma patients. Glioblastoma cells are highly dependent on redox regulation for their survival, thus elevated intracellular ROS production promotes TMZ-resistance through impaired redox balance [58]. The upregulation of ROS levels was studied as an effective way to target redox homeostasis to induce oxidative stress that may further promote cell death and sensitize cancer cells to anticancer therapies [58]. In this aspect, the synergy of chemotherapeutic agents as DOX with the photosensitizing properties of GQDs resulted in increased membrane permeability further enhancing DOX intracellular accumulation and efficacy. Upon NIR light PTT conversion in the 3D spheroid models, the GQDs induced significantly increased permeability and tumor accumulation, thus enhancing the localized therapeutic efficacy of DOX and TMZ and further inhibiting tumor growth. Moreover, the elevated presence of tumor-associated antigens and ROS (due to GQDs-induced PTT) promoted the migration of immune cells at the tumor site, suggesting the reactivation of immune responses. Oxidative stress targeting has also been studied recently by Campbell et al. [59], in nitrogen-doped GQD formulations functionalized with hyaluronic acid, HA, and ferrocene, Fc, (Fc-GQDs-HA) for the combined PTT and image-guided cancer therapy of HeLa cervical cancer cells. The N-doped GQDs (N-GQDs) are pronounced fluorophores for cellular and tumor multicolor imaging. Moreover, N-GQDs were applied in the targeting of the acidic extracellular TME since they presented pH-dependent fluorescence response [60]. The functionalization of N-GQDs with HA served in the targeting of CD44 cancer cell receptor overexpressed in HeLa cells and playing a key part in the regulation of cell proliferation and survival. The conjugation of Fc facilitated a redox-based toxicity in the cancer cells thought the redox cycle of iron. Specifically, the anticancer properties of Fc and its derivatives (photochemical and biochemical organometallic compounds) originate from the Fenton pathway that promotes the oxidation of Fe(II) to Fe(III) leading to increased ROS production [61]. The central iron atom of Fc presents great redox capacities by its electron donor-acceptor ability catalyzing the Fenton reaction of endogenous superoxide molecules promoting the oxidation of Fc to ferricinium cation. The Fe(III) ions are used in lipid metabolism promoting the generation of increased levels of lipid hydroperoxides that further induce ferroptosis (iron-dependent cell death) in cancer cells [62,63]. N-doped GQDs were also studied by Guo et al. [64] in mitochondria targeting photo-responsive platform for the combined delivery of nitric oxide (NO) and triphenylphosphonium (TPP) for the in vivo PTT anticancer therapy. The N-GQDs were surface functionalized with ruthenium nitrosyl that served as the NO donor under NIR light irradiation. The essential role of NO in mitochondrial respiration is based on its ability to regulate oxygen consumption and ATP synthesis by serving as a redox signaling molecule and promoting an inhibitory effect in the electron transport chain (ETC) of the mitochondria inner membrane. ETC is a collection of protein complexes (complex I, complex II, coenzyme Q, complex III, cytochrome C, and complex IV) that are used in the transportation (pass through) of electrons in a series of redox reactions that release energy. The released energy further creates a proton gradient used in the ATP synthesis. NO mainly inhibits the action of complex IV (Cytochrome c oxidase (COX)) that is the terminal enzyme of ETC responsible for reducing molecular oxygen to H2O and driving ATP synthesis [65]. More details on the mechanisms of NO inhibitory effect on the mitochondrial respiratory chain and biogenesis are presented in a review by Tengan et al. [66]. Defective mitochondrial respiration can be promoted in cancer cells by the prolonged exposure to increased levels of NO further regulating elevated ROS and RNS (reactive nitrogen species) production. The ruthenium nitrosyl N-GQDs presented increased mitochondrial targeting due to the surface functionalization with TPP+ cations that effectively target the negatively charged mitochondrial membrane. TPP+ belongs to the family of delocalized lipophilic cations (DLCs) that are selectively transported through the mitochondrial phospholipid membrane. The hyperpolarized potential of the mitochondria membrane can significantly be targeted by DLCs that due to their lipophilicity can be easily transported though the outer and inner mitochondria membranes [67]. Thus, the mitochondria localized elevated release of NO (due to ruthenium nitrosyl) under NIR light radiation in combination with PTT effect significantly inhibited the in vivo tumor growth. The targeting of mitochondria membrane potential has been an ideal tool for increasing the selectivity and therapeutic efficacy of ROS-inducing GBMs. In a study by Fan et al. [68], GQDs functionalized with TPP were studied for their application in the in vitro monitoring of mitochondria. The GQDs were surface functionalized with PEI (polyethyleneimine) offering an amid linkage to TPP. The GQDs-PEI-TPP graphenes were efficiently accumulated in the mitochondria as observed by the comparative fluorescence imaging of GQDs (green fluorescence) and mitochondria (Mitotracker Red fluorescence). The application of GQDs in mitochondria targeting was also studied by Zhang et al. [69] in hybrid nanoparticles of GQDs covalently functionalized with rare earth doped UCNP (upconversion nanoparticles). The GQDs-UCNP showed increased mitochondria specificity due to the conjugation of TRITC (Tetramethylrhodamine-5-isothiocyanate). Isothiocyanate (ITC) molecules are promising anticancer agents that express elevated mitochondria targeting leading to ITC-induced apoptotic cell death by activating caspase-9 related apoptotic pathway and by regulating Bcl-2 protein family members related to pro-apoptotic signaling in cancer cells. For the activation of caspase-9, mitochondria release cytochrome c from their intermembrane space inducing the loss of mitochondria membrane integrity [70,71]. The TRITC functionalized GQDs-UCNP promoted the increased in situ cytotoxic ROS formation (mainly singlet oxygen species, 1O2) in mitochondria upon NIR light irradiation. Moreover, the mitochondria dysfunction was confirmed by the significant decrease in mitochondria membrane potential and the activation of caspase 3 apoptotic pathway. Mitochondria damage is the main signal promoting apoptotic cell death in cancer cells, thus the combined mitochondria targeting and PDT effect resulted in important tumor suppression upon NIR irradiation in 4T1 tumor models treated with GQDs-UCNP/TRITC.
Graphene has also been applied in mitochondria targeting effective anticancer therapy by Saeed et al. [72] in breast and pancreatic cancer cells. Graphene was used for the nanodelivery of gambogic acid (GA), significantly enhancing its in vitro bioavailability. GA is a natural caged xanthonoid compound that highly promotes calcium ion (Ca2+) intracellular influx efficiently promoting the disregulation of Ca2+ homeostasis in various cancer cells. The increased upregulation of Ca2+ promotes the dysfunction of mitochondria membrane and the subsequent activation of mitochondria apoptosis signaling [73]. The mitochondria damage is promoted by GA thought the disruption of lipid metabolism, and the swelling of mitochondria (mega-mitochondria) and the endoplasmic reticulum (ER vacuolization) leading to paraptosis (vacuolization-associated cell death) [74,75]. The graphene-GA highly promoted the depletion of mitochondria membrane potential (MMP) and decreased the intracellular level of lipid droplets serving in energy storage. Moreover, nuclear DNA fragmentation was induced by the graphene-GA being a proof of apoptotic cancer cell death. However, GA was not the only compound contributing to the disruption of MMP. In a study by Li et al. [76] commercial pristine graphene was able to be internalized into murine RAW 264.7 macrophages and promote mitochondria pathway signaling to induce apoptotic cell death. Specifically, graphene triggered increased ROS levels and activated pro-apoptotic proteins of the Bcl-2 family (Bim and Bax) through the activation of MAPK and TGF-β signaling that stimulated the activation of cascape-3 related apoptotic pathway. Thus, pristine graphene was able to induce cytotoxicity to murine macrophage-like cells by regulating the mitochondria-induced apoptotic cell death.
2.2. Graphene Oxide
Hypoxia, acidity, oncogenic mutations, and immunosuppression constitute a TME that hinders the efficacy of anticancer therapies and immunotherapies by accumulating toxic metabolites and promoting nutrients’ starvation that induce an aggressive cancer cells’ phenotype and mechanisms of multidrug resistance (MDR). Recent therapeutic strategies lay on the versatility and facility of GBMs functionalization, especially GO, to deliver targeted nanomedicines for the reprogramming and modulation of TME. The application of GO is based on the great advantages offered by its binding oxygen functional groups, and the hydrophobic and π-π stacking interactions of its pristine graphene lattice. Nano GO has presented significant ability of cellular transportation either by endocytosis and plasma membrane mechanisms (intracellular) or by diffusion (paracellular pathways). Through functionalization (Figure 4), GO enabled the binding of targeting ligands for cancer cells’ membrane receptors and mitochondria membrane, to selectively deliver anticancer and immunotherapeutic agent for TME reprogramming (Table 2 presents research examples).
In a recent study by Han et al. [77] tumor and mitochondria targeting were assessed by the conjugation of hypericin (HY) on PEG functionalized GO through disulfide bonding. HY is a natural pleiotropic product acting as an anticancer agent by inhibiting pro-inflammatory mediators and increasing the expression levels of caspase-3 and caspase-4 promoting apoptotic cell death [78]. In the acidic and hypoxic TME, HY selectively binds to the low-density lipoprotein (LDL) receptors of cancer cell membranes promoting its non-specific accumulation [79,80]. Mitochondria are the primary target of the photodynamic action of HY by producing singlet oxygen molecules (1O2) through a type II oxygen-dependent photosensitization mechanism in the ETC at complex III [81]. Then, the generation of free radicals and reactive intermediates is promoted leading to the induction of mitochondria membrane permeability transition [82]. The HY functionalized GO vehicle was studied for the combined PDT and anticancer therapy of breast cancer in vitro and in vivo by the synergistic action of doxorubicin (DOX). The GO vehicle was internalized via endocytosis promoting the glutathione-triggered release of HY and the pH-triggered release of DOX. Upon NIR laser irradiation, HY targeted in the mitochondria generating singlet oxygen that further promoted the release of cytochrome c into the cytosol. The DNA damage promoted by DOX action and cytochrome c release, further promoted caspases activation that stimulated the apoptotic cancer cell death. In another study by Wu et al. [83], GO nanocomposites with DSPE-PEG2000 were used as a nanographene platform for the combined PDT/PTT and immunostimulatory anticancer effect in vitro and in vivo in EMT-6 transplantable mouse mammary tumor cell line. The amphiphilic DSPE-PEG was attached to the graphitic lattice of GO via hydrophobic interactions. Mitochondrial-targeting was demonstrated by the GO nanocomposite through the conjugation of the lipophilic cation alkyl-triphenylphosphonium (TPP+) in DSPE-PEG chain. The alkylated TPP+ has been extensively used in mito-targeting due to the negative membrane potential of the mitochondrial inner membrane resulting in the increased uptake of positively charged small molecules [84]. The PDT/PTT effect of GO was boosted by the IR820 NIR responsive photosensitizer to promote ROS-induced mitochondrial damage due to NIR-light conversion. Furthermore, the GO nanocomposite exhibited significant immunostimulatory effects owing to the linkage of CpG ODN nucleotide acid drug (synthetic unmethylated cytosine-guanosine oligodeoxynucleotides). The CpG ODN are short single-stranded synthetic DNA molecules that contain unmethylated oligodeoxynucleotides. CpG ODN are recognized by the TLR9 (Toll-like receptor 9) highly expressed on human B cells and pDCs (plasmacytoid dendritic cells) to stimulate immune responses (innate and adaptive) [85]. The combinational effect of CpG ODN with PDT/PTT resulted in the increased ROS production that promoted the mitochondrial induced cell death and the significant upregulation of proinflammatory cytokines (such as IL-6, TNF-α, INF-γ) improving immunogenicity in the TME and inhibiting tumor growth by 88 %.
Table 2.
Applications for graphene oxide (GO) based nanocomposites in cancer therapeutics.
| Carrier Type | Agent | Characteristics | Ref |
|---|---|---|---|
| GO-PEG | HY, DOΧ | Combined PDT and anticancer therapy, internalization via endocytosis, glutathione-triggered HY release, pH-triggered DOX release, HY-triggered generation of singlet oxygen in mitochondria, cytochrome c release into the cytosol, caspases activation | [77] |
| GO- DSPE-PEG2000 | TPP+, IR820 NIR Photosensitizer, CpG ODN |
combined PDT/PTT and immunostimulatory anticancer effect, mitochondrial targeting, increased ROS production, mitochondrial induced cell death, upregulation of proinflammatory cytokines (IL-6, TNF-α, INF-γ) | [83] |
| GO-ICG | TPPB | increased mitochondrial accumulation, synergistic PDT/PTT, elevated ROS (1O2, O2–) production, GO hyperthermia effect, inhibition of ATP synthesis | [86] |
| GO-PEG | integrin αvβ3 mAb, PPa | on/off phototoxicity switches, FRET mechanism, mitochondria accumulation, localized ROS formation, promoting cell apoptosis | [88] |
| GO-MitP | MTX | Increased mitochondria localization, promoting mitochondria dysfunction upon AFM, decrease in MMP, downregulation in ATP expression levels, cytochrome c release, activation of caspase 3 apoptotic pathway | [89] |
| GO- β-CD/Plys/PEG/MitP | TPM-Azo, TF | Increased mitochondria accumulation, stimulating mitochondrial aggregation, reducing ATP levels, disruption of cancer cells cycle, arrest at the G2 phase, decreased cell viability, increased cytochrome c expression into the cytosol | [92] |
| GO-GA | DOX | mitochondria targeting, selective pH-dependent DOX release, decreased the MMP, opening of the mitochondrial permeability transition pores, increase in the ratio of Bax/Bcl-2 proteins, activation of caspase-mediated apoptotic pathway | [94] |
| GO-GE11 | Ori | promoted mitochondrial dysfunction, decreased the MMP, induced cell cycle arrest increasing the G2/M phase, downregulation of the Ras/Raf/MEK/ERK pathways | [99] |
Abbreviations: PEG: polyethylene glycol, HY: hypericin, PDT: photodynamic therapy, DOX, doxorubicin, DSPE-PEG2000: 1,2-distearoyl-sn-glycero-3-phosphoethanolamine-poly(ethylene glycol)-2000, PTT: photothermal therapy, TPP+: alkyl-triphenylphosphonium, NIR: near-infrared region, CpG ODN: synthetic unmethylated cytosine-guanosine oligodeoxynucleotides, ROS: reactive oxygen species, ICG: indocyanine green, TPPB: (4-carboxybutyl) triphenyl phosphonium bromide, mAb: monoclonal antibody, PPa: pyropheophorbide-a photosensitizer, FRET: Fluorescence Resonance Energy Transfer, MitP: mitochondria targeting peptide, MTX: mitoxantrone, AFM: alternating magnetic field, MMP: mitochondrial membrane potential, ATP: Adenosine triphosphate, β-CD: β-cyclodextrin, TPM-Azo: TPM-Azobenzene, Plys: polylysine, TF: transferrin, GA: glycyrrhetinic acid, GE11: EGFR targeting peptide, Ori: Oridonin.
The synergistic effect of mitochondria-targeting and PDT/PTT for enhanced anticancer treatment against drug–resistant osteosarcoma (OS) was studied in vitro and in vivo by Zeng et al. [86] in PEI (polyethylenimine)-modified PEG-GO nanosheets. The GO nanocomposites were conjugated with indocyanine green (ICG) and with the mitochondria-targeting ligand (4-carboxybutyl) triphenyl phosphonium bromide (TPPB). ICG is an FDA-approved NIR fluorescent and contrast agent for imaging and image-guided surgery. Owing to its ability to convert the absorbed light to energy, ICG has been used in cancer theranostic applications as a photosensitizer to induce hyperthermia and increase ROS expression levels, as singlet oxygen (1O2) and superoxide (O2–) [87]. The incorporation of ICG into the GO nanocomposites considerably increased the mitochondrial accumulation through mito-specific TPPB targeting. After NIR light irradiation, the increased intracellular localization of the nanocomposites enabled the synergistic PDT/PTT therapy of doxorubicin-resistant MG63/DOX OS cancer cells in vitro and in vivo in OS tumor-bearing mice. The elevated production of intracellular ROS (1O2, O2–), due to ICG, in combination with GO hyperthermia effect resulted in the irreversible apoptotic damage of cancer cells through mitochondria dysfunction. The mito-damage was associated with the inhibition of ATP synthesis that further suspended the energy supply of MG63/DOX cancer cells. The suppressed ATP production in vivo upon NIR light, resulted in the inhibition of tumor growth and the increased presence of apoptotic or necrotic cancer cells in tumor tissues. The GO nanocomposites expressed elevated biosafety as it was assessed by blood biochemistry markers (alanine aminotransferase ALT, aspartate aminotransferase AST, alkaline phosphatase ALP). In another study by Wei et al. [88] GO nanocomposites modified with an integrin αvβ3 monoclonal antibody (mAb) and a pyropheophorbide-a (PPa) photosensitizer that was conjugated with PEG polymer were evaluated as on/off phototoxicity switches for tumor and mitochondria targeted delivery. The switch on/off mechanism was based on the hydrophobic interactions between PPa and the graphitic plane of GO in aqueous environment, such as in the blood circulation and the cytoplasm of cancer cells. Within aqueous environment, intense π-π stacking interactions are generated between PPa and GO promoting the formation of a stable structure. Upon laser light irradiation, the excited PPa transferred its energy to GO through FRET mechanism (Fluorescence Resonance Energy Transfer) occupying an off phototoxicity state. The GO nanocomposites exhibited a preferential accumulation into the mitochondria in vitro in U87-MG glioblastoma and MCF-7 breast cancer cells, due to the mAb providing an effective distribution of PPa and translocation from the lysosome to the mitochondria. The underlying mechanism for effective mito-translocation was associated with the electrostatic interaction between the mitochondria membrane and the polarized GO nanocomposites that promoted their physical attraction to the mitochondria. Within the lipid environment of the mitochondrial membrane, the π-π stacking interactions among PPa and GO were destabilized promoting the on phototoxicity state. Upon light irradiation, the switched-on state of the PPa photosensitizer permitted the localized ROS formation into the mitochondria promoting cell apoptosis.
The targeting of mitochondria is an effective approached to reprogram TME promoting apoptotic cancer cell death. Apart from targeting ligands, peptides have also been used in mitochondria targeting applications. Zhu et al. [89] studied the in vitro application of magnetic GO nanocomposites conjugated with mitochondria targeting peptide (MitP) for the selective delivery of mitoxantrone (MTX) and the promotion of mitochondria dysfunction. MTX is a DNA topoisomerase II inhibitor that forms a DNA-topoisomerase II/MTX complex to inhibit the DNA transcription and replication processes [90]. MTX cardiotoxicity arises from the induced collapse of mitochondrial membrane potential that promotes the formation of abnormal mitochondria, ATP depletion and mitochondriopathy [91]. The MitP-GO nanocomposites presented increased localization in the mitochondria of A549 lung adenocarcinoma cancer cells, promoting the selective delivery of mitoxantrone. Upon AMF (alternating magnetic field) application, the effect of MTX on promoting mitochondria dysfunction was assessed by the downregulation in the expression levels of ATP and the decrease in mitochondrial membrane potential. Moreover, the mitochondria damage was further confirmed by the release of cytochrome c and the subsequent activation of caspase 3 apoptotic pathway. The application of MitP was also exploited by Zhang et al. [92] in β-cyclodextrin (β-CD)- grafted GO nanoassemblies with TPM-Azobenzene (TPM-Azo). The TPM-Azo was prepared by covalent functionalization of polylysine (Plys) with the tumor targeting protein transferrin (TF), the MitP, and PEG to improve biodispersibility and biocompatibility. Azo, under hypoxia conditions, is a photoactive molecule activated in the absence of oxygen that can act as a nitrogen mustard deactivator in mitochondrial targeting. Nitrogen mustards can generate intra- and inter-strand crosslinking with DNA inducing DNA damage. However, the DNA repair mechanisms of cancer cells may promote drug resistance and mutagenic effects. The reapplication of Azo into the mitochondria is based on the presence of mitochondria targeting ligands or peptides, to increase mito-localization, bypass cellular drug resistance mechanisms, and off-target liver activity [93]. In the GO nanoassemblies, the MitP highly promoted the in vitro accumulation of the system in the mitochondria of A549 lung cancer cells, further inducing mitochondrial aggregation and decreasing ATP expression levels. The application of UV and NIR light irradiation promoted the dissociation of the TMP-Azo from the GO nanoassemblies further stimulating mitochondrial aggregation and lowering ATP levels. The mitochondrial dysfunction promoted by the GO nanoassemblies was associated with disruption of cancer cells’ cycle, inducing arrest at the G2 phase thus preventing cancer cells from entering mitosis, and inhibiting cancer cells proliferation. Thus, the G2 cell cycle arrest was related with decreased A549 cell viability. Under NIR light, the photothermal transition activity of the nanoassemblies significantly decreased cell viability and increased cytochrome c expression levels from the mitochondria into the cytosol. The in vivo application of the GO nanoassemblies in S180 tumor-bearing mice resulted in elevated antitumor effect significantly inhibiting tumor growth.
The mitochondria-triggered apoptosis is an effective pathway for anticancer therapy that was also studied by Zhang et al. [94] in glycyrrhetinic acid (GA) functionalized GO nanocomposites for the selective delivery of doxorubicin in mitochondria. GA is a biologically active metabolite that derives from glycyrrhizin, a natural product of Glycyrrhiza glabra (or licorice) with broad anticancer properties and mitochondria targeting ability [95]. The anticancer properties of GA are related with the inhibition of protein kinase C (PKC) α/βII and activation of c-Jun NH2-terminal kinase of JNK pathway promoting apoptosis of non-small cell lung cancer cells [96]. In the mitochondria, GA can significantly decrease mitochondrial membrane potential and promote caspase-3 mediated apoptosis, [97] and restrict energy metabolism by binding to and inhibiting the activity of the enzyme serine hydroxymethyltransferase 2 (SHMT2) [95]. GA by interacting with complex I of the mitochondrial respiratory chain promotes oxidative stress facilitating the generation of ROS, such as hydrogen peroxide (H2O2), superoxide radicals and the highly reactive hydroxyl radicals that further lead to thiol oxidation. This mechanism promotes the opening of the mitochondrial permeability transition pores in the presence of calcium ions (Ca2+) inducing mitochondria targeting effects to GA [98]. Thus, the GA functionalized GO effectively promoted mitochondria targeting for the selective pH-dependent release of DOX and the synergistic induction of mitochondria apoptosis and programmed cancer cell death. The nanocomposites decreased the mitochondrial membrane potential in vitro and in vivo in HepG2 hepatocellular carcinoma nude mice. The opening of the mitochondrial permeability transition pores was verified by the increase in the ratio of Bax/Bcl-2 proteins facilitating the release of cytochrome c into the cytosol and further activating the caspase-mediated apoptotic pathway (especially caspase-3). The activation of mitochondria apoptotic signaling pathways was studied by Jiang et al. [99] in GO nanodrugs functionalized with GE11 EGFR targeting peptide for the delivery of Oridonin (Ori). Ori is a natural terpenoid interfering in the apoptotic signaling pathways by modulating the Bcl-2/Bax protein expression. The Bcl-2 and Bcl-xl belong to the anti-apoptotic protein family, while the Bax and Bak to the pro-apoptotic protein family. Thus, the downregulation of Bcl-2, in combination with upregulation of Bax expression promoted the release of cytochrome c to the cytoplasm leading to caspase activation and caspase-triggered apoptosis [100]. The GO-Ori nanodrug was evaluated in vitro in KYSE-30 and EC-109 human oesophageal squamous cell carcinoma cells that overexpress the EGFR receptor. GO-Ori nanodrug effectively promoted mitochondrial dysfunction by decreasing the mitochondrial membrane potential and increasing the Bax/Bcl-2 ratio. Moreover, the increased lysosomal accumulation of the GO-Ori nanodrug significantly promoted toxicity and induced cell cycle arrest increasing the G2/M phase. Thus, nanodrug triggered cancer cells’ apoptosis by cell cycle arrest, and mitochondria mediated apoptosis. Moreover, the effective targeting of EGFR resulted in the downregulation of the Ras/Raf/MEK/ERK pathways that are linked with biological responses including growth, proliferation, migration and apoptosis.
2.3. Reduced Graphene Oxide
Since the metabolic reprogramming of TME in solid tumors is mainly based on the targeting of mitochondrial function to induce mito-mediated apoptosis and energy deprivation, the implementation of rGO has received increasing attention. The ROS formation plays a crucial role in the regulation of mitochondria function and reprogramming with elevated ROS levels acting as signaling molecules that modulate cellular pathways involved in cancer cell survival and proliferation. ROS formation plays a dual role in the mitochondria of cancer cells, since it contributes to cellular adaptation, survival, and progression while also driving DNA damage and genomic instability creating a mutagenic environment that drives the evolution of aggressiveness and metastasis [101]. Targeting the mechanisms that control ROS generation in mitochondria holds therapeutic promise in cancer treatment (Figure 5). The driving force behind using rGO in targeting ROS generation in the TME in cancer cells stems from several of its unique properties, which make it highly suitable for modulating ROS production and improving cancer treatment [102]. rGO has a lower ratio of oxygen groups than GO, offering less hydrophilicity but has a higher electrical conductivity that in combination with its small size, sharp edges and surface provides the ability to be transported through cellular membranes promoting the damage of cellular membranes and increased cytotoxicity (Table 3 presents characteristic research examples).
In a recent study by Kretowski et al. [103], rGO stimulated autophagy and cell cycle arrest in MDA-MB-231 and ZR-75-1 breast cancer cells in vitro, further promoting apoptosis signaling pathway by decreasing the mitochondrial membrane potential and activating caspase-9 and caspase-3 cytosolic expression. In another study by Zhang et al. [104], rGO in combination with gamma irradiation was evaluated in H9C2 rat myocardial cells in vitro and in in vivo experiments with C57BL/6 male mice. rGO could promote oxidative stress damage with elevated levels of ROS expression, while reducing the mitochondria membrane potential. Apart from mitochondrial dysfunction, rGO in synergy with increasing radiation dose promoted the damage of myocardial tissue by reducing the expression levels of enzymes, including aspartate aminotransferase and lactate dehydrogenase.
Table 3.
Applications for reduced graphene oxide (rGO) based nanocomposites in cancer therapeutics.
| Carrier Type | Agent | Characteristics | Ref |
|---|---|---|---|
| rGO | - | stimulated autophagy and cell cycle arrest in cancer cells, promoted apoptosis signaling pathway, decreased the MMP, activated caspase-9 and caspase-3 cytosolic expression | [103] |
| rGO | Gamma irradiation | oxidative stress damage, elevated levels of ROS expression, damage of myocardial tissue, reduced expression levels of enzymes | [104] |
| rGO-MNPs | Coum, CMP | Combined chemophotodynamic effect, elevated ROS production, regulation of apoptotic cell death, cell shrinkage and nuclear fragmentation, increased expression of p53 and Bax proteins | [105] |
| rGO | MG-132 | increased apoptosis and necrosis of breast cancer cells, increasing ROS formation, caspase-8 and caspase-9 apoptotic pathways activation, reduced enzymatic antioxidants activity (SOD, GPx) | [106] |
| rGO-AgNPs | - | Decreased viability and cell growth, leakage of intracellular LDH, damage of cellular membrane integrity, ROS generation, increased MDA pro-oxidant levels, decreased antioxidant GSH production, activation of caspase-3 apoptotic death | [111] |
| rGO-AgNPs | Cis | Inhibition of viability and proliferation, increased Cis-induced ROS expression levels, increased oxidative stress, elevated MDA expression levels, loss of MMP, upregulation of apoptotic genes, downregulation of anti-apoptotic genes | [112] |
| rGO-Arg/Pro | - | increased cellular membranes adhesion, reduced cell proliferation, downregulation of VEGF expression, caspsase-3 activation, inhibition of FGF2, regulation of apoptosis | [115] |
Abbreviations: MMP: mitochondrial membrane potential, ROS: reactive oxygen species, MNPs: magnetic nanoparticles, CMP: camptothecin, ROS: reactive oxygen species, MG-132: aldehyde proteasome inhibitor peptide, SOD: superoxide dismutase, GPx: glutathione peroxidase, AgNPs: silver nanoparticles, LDH: lactate dehydrogenase, MDA: malonaldehyde, GSH: glutathione, Cis: cisplatin, MMP: mitochondria membrane potential, Arg: Arginine, Pro: Proline, FGF2: fibroblast growth factor-2.
Certainly, one of the primary advantages of rGO is its ability to generate ROS, especially singlet oxygen and free radicals upon light activation (NIR or UV-vis light), which is particularly useful in PDT [45]. By targeting mitochondria with rGO and functionalized rGO, ROS accumulation in these organelles can lead to oxidative damage, mitochondrial dysfunction, and apoptotic cell death. rGO can interact with mitochondria directly, either by localizing on their surface or internalizing into the organelle, facilitating ROS-induced damage. The effect of rGO in the production of ROS can be greatly improved by its surface functionalization to provide engineered rGO enabling better targeting of cancer cells and mitochondria with targeting molecules like peptides or antibodies that bind specifically to cancer cell markers or mitochondria-associated receptors [102]. In a recent study by Vinothini et al. [105], rGO was functionalized with coumarin (Coum) photosensitizer and magnetic nanoparticles (MNPs) for the controlled release of camptothecin (CMP), in combination with PDT under UV light irradiation. The synergistic chemophotodynamic effect of the rGO nanodrug highly promoted cytotoxicity of MCF-7 human breast cancer cells through elevated production of ROS and regulation of apoptotic cell death. Cellular apoptosis was accompanied by cell shrinkage and nuclear fragmentation, with increased expression levels of p53 and Bax proteins. The intracellular ROS levels significantly increased at 24 h upon UV irradiation, exhibiting a short lifetime due to their participation in redox reaction that promoted cell membrane damage. The rGO nanodrug exhibited synergistic antitumor effect in vivo in DMBA-induced mammary tumor-bearing rats, with decreased tumor volume/weight, due to the increased apoptotic cell death and tumor tissue damage. Characteristic was the upregulation of the in vivo expression levels of p53 gene being a crucial biomarker of apoptosis and DNA damage, and of Bax protein indicating the regulation of apoptotic signaling pathways. Moreover, the rGO nanodrug promoted the regulation of mitochondria respiration in vivo by significantly decreasing the enzymatic expression levels of acid phosphatase, β-D-glucuronidase, cathepsin and LDH. In another study by Kretowski et al. [106], the synergistic effect of rGO and enzymatic inhibition was studied in the induction of apoptosis through oxidative stress. The peptide aldehyde proteasome inhibitor MG-132 was mixed with rGO to evaluate the anticancer effect of their nanocomposite against ZR-75-1 and MDA-MB-231 breast cancer cells. The proteasome is a large multi-subunit protein complex present in the nucleus and cytoplasm of cells with the main function to degrade damaged, misfolded, or superfluous proteins by proteolysis. To be recognized, the proteins are tagged with ubiquitin (a small protein) and translocated into the catalytic core of proteasome allowing them to be degraded into smaller peptides. This process is essential for maintaining protein homeostasis (proteostasis) and regulating various cellular processes, such as cell cycle progression, signal transduction, and stress responses [107]. The MG-132 inhibitor can selectively induce apoptosis in glioblastoma cells by inhibiting the PI3K/Akt and NFκB signaling pathways involved in cancer initiation and progression. By promoting cell cycle arrest at the G2/M phase, the MG-132 prevented cancer cells from entering mitosis thus inhibiting their proliferation. Moreover, the MG-132 inhibitor promoted mitochondrial dysfunction by decreasing the mitochondrial membrane potential and opening the mitochondrial permeability transition pore resulting in the downregulation of the anti-apoptotic bcl-xl protein expression levels and further activation of caspase-3 apoptotic signaling [108]. The nanocomposite of rGO and MG-132 inhibitor, significantly increased apoptosis and necrosis of breast cancer cells and promoted the activation of caspase-8 and caspase-9 apoptotic pathways. The rGO with MG-132 highly supported intracellular oxidative stress by increasing ROS formation and decreasing thiol groups (Glutatione, GSH) expression levels because of reduced enzymatic antioxidants activity such as SOD (superoxide dismutase) and GPx (glutathione peroxidase).
The importance of rGO nanocomposites with silver nanoparticles (AgNPs) in anticancer therapy has attracted attention specifically regarding ROS formation by mitochondria due to their ability to target and disrupt mitochondrial function, a key pathway for cancer cell apoptosis. The rGO/Ag nanocomposites significantly increase oxidative stress due to their ability to be internalized by cells allowing them to interact directly with the mitochondria, increasing ROS levels in a more localized manner [109]. Moreover, AgNPs can facilitate the production of ROS by acting as a catalyst in Fenton-like reactions, where silver nanoparticles can react with cellular components (like hydrogen peroxide, H2O2) to generate highly toxic hydroxyl radicals. rGO, being a good electron donor, can enhance the transfer of electrons during redox reactions, leading to a higher concentration of ROS in the mitochondria. The combination with AgNPs can act synergistically to catalyze ROS production significantly increasing ROS accumulation leading to oxidative stress and further promoting mitochondria dysfunction [110]. The anticancer properties of rGO-AgNPs nanocomposites were evaluated on their effect on TME by Gurunathan et al. [111] in A2780 epithelial ovarian carcinoma cells. The rGO-AgNPs nanocomposites expressed increased inhibitory effect on cell viability accompanied with pronounced decrease of cell growth. Moreover, the rGO-AgNPs significantly promoted the leakage of intracellular LDH (lactate dehydrogenase) as a response to the damage of cellular membrane integrity. ROS generation was significantly promoted inducing oxidative stress and increasing the intracellular expression level of pro-oxidant MDA (malonaldehyde) while decreasing antioxidant GSH (glutathione) production. The strong effect of rGO-AgNPs nanocomposites on inducing oxidative stress highly promoted caspase-3 activation and induced apoptotic cell death. In a recent study by Yuan et al. [112], rGO-AgNPs nanocomposites were studied in combination with cisplatin (Cis) to evaluate their in vitro synergistic effect on apoptosis and autophagy of HeLa human cervical cancer cells. The synergy of Cis with rGO-AgNPs highly inhibited the viability and proliferation of cervical cancer cells promoting increased LDH leakage due to cellular membrane damage, in comparison to Cis, pristine rGO and AgNPs. The rGO-AgNPs acted synergistically with Cis in supporting the oxidation-reduction status (redox) of cancer cells, promoting a pronounced increase on the Cis-induced ROS expression levels. The increased oxidative stress of the cancer cells was verified by the elevated MDA expression levels. Moreover, the combination of Cis with rGO-AgNPs had a strong influence on the loss of mitochondria membrane potential and on gene expression levels related to mitochondrial dysfunction. The activation of apoptosis as a response to oxidative stress and DNA damage was verified by the upregulation of apoptotic genes (P53, P21, BAX, BAK, CASP3, and CASP9) and downregulation of anti-apoptotic genes (BCL2 and BCL2L1). The activation of CASP3 and CASP9 gene expression was associated with mitochondrial induced apoptosis pathway. Cisplatin exerts its cytotoxic effects primarily through DNA damage, leading to the activation of signaling pathways, including pathways for the regulation of autophagy. By disrupting DNA replication and transcription, Cis promotes the activation of DNA damage cellular responses, including the upregulation of p53 protein expression that is related to the promotion of BECN1 (Beclin-1) autophagy-related gene. Cis-induced DNA damage promotes oxidative stress and mitochondrial dysfunction leading to a decrease in cellular ATP levels and AMPK protein kinase activation. In turn, the activation of AMPK can inhibit mTOR pathway promoting the induction of autophagy in response to Cis-induced stress. Collectively, DNA damage, oxidative stress, p53 activation, and AMPK/mTOR inhibition contribute to the cellular responses to Cis that can influence the activation of autophagy [113]. Interestingly, the combinational treatment of Cis with rGO-AgNPs significantly increased the induced autophagic cell death through the generation of autophagosomes and upregulation of autophagy-related (ATG) genes expression. The pronounced presence of autophagic vacuoles was combined with increased generation of autophagolysosomes. Under the elevated ROS production and oxidative stress promoted by Cis and rGO-AgNPs combination, the extended presence of autophagy endorsed cancer cell death.
The surface protein adsorption ability of rGO was evaluated on the cellular responses and induced toxicological profile on MDA-MB-231 mammary epithelial cancer cells. It was demonstrated that the lower the surfactants’ concentration the higher the protein adsorption (biocorona formation). The rGO with increased protein corona promoted pronounced increase on ROS generation, lipid membrane peroxidation, oxidative stress, and inhibited mitochondrial ATP synthesis [114]. The coating and surface functionalization of rGO has been used to shape the cellular membrane interactions of rGO, including the mitochondrial membrane. In a study by Sawosz et al. [115], the functionalization of rGO with Arginine (Arg) or Proline (Pro) was evaluated on their anticancer effect in glioblastoma multiforme U87 cells in vitro and in tumors in vivo. The Arg and Pro amino acids were conjugated on rGO by reduction process to improve stability and protect rGO from agglomeration. Both Pro and Arg play important roles in cellular metabolism, signaling, and stress responses. Pro metabolism and degradation pathways participate in regulating the function of p53 protein inducing stress signal responses, including DNA damage, oxidative stress and nutrient deprivation. Such signals act through the mitochondrial function by proline dehydrogenase (PRODH) enzymatic action that converts proline to glutamate while generating ROS byproducts and activating p53 expression through oxidative stress mechanisms [116]. Arg metabolism is important in cancer biology, as it can influence cell survival and apoptosis by regulating NO (nitric oxide) production through the action of nitric oxide synthase (NOS) further activating p53 expression. NO can activate the mitochondrial apoptotic pathway, leading to mitochondrial dysfunction and the release of pro-apoptotic factors like cytochrome c, which subsequently activates p53-mediated cell death pathways [117]. The amino acids functionalization of rGO increased its adhesion to cellular membranes thus increasing toxicity. Moreover, the rGO Pro and Arg nanocomposites reduced the tumor volume by inhibiting cell proliferation. The reduced cell proliferation rate was accompanied by the inhibition of FGF2 (fibroblast growth factor-2) and downregulation of VEGF expression. Moreover, rGO nanocomposites promoted caspsase-3 activation and mRNA genes expression, such as MDM2 (mouse double minute 2), COX6 (cytochrome c oxidase 6) indicating the regulation of apoptosis. Among the rGO nanocomposites, the Arg functionalized sufficiently induced the intrinsic apoptotic pathways and COX6 activation, while proline inhibited the proapoptotic rGO activity.
3. Discussion
Reprogramming the heterogenic TME is a complex and challenging task, primarily due to its diversity and dynamic nature. TME is composed of various cell types, including cancer cells, stromal cells, immune cells, endothelial cells, and extracellular matrix components, all of which can vary significantly between different regions within the same tumor. The various cellular and molecular components of TME contribute to its immunosuppressive nature. Immune cells, such as regulatory T cells (Tregs), myeloid-derived suppressor cells (MDSCs), and tumor-associated macrophages (TAMs), often dominate TME, creating an immunosuppressive environment that prevents the immune system from effectively targeting, reprogramming, and eliminating tumor cells. The use of GBM nanocomposites offers a promising strategy in reprogramming the TME for overcoming the challenges of heterogeneity and immunosuppression in cancer therapy, potentially improving the effectiveness of both immune-based and chemo-based nanomedicine therapies [118]. The advancements in nanotechnology have led to the development of personalized nanomedicine that can more effectively deliver drugs and therapeutic agents into the TME, bypassing its physical barriers. Personalized nanomedicine has become a promising approach for enhancing the precision and effectiveness of cancer therapies since they can combine different treatment modalities, including chemotherapy, radiotherapy, gene therapy, and immunotherapy, into a single nanoparticle platform. This multi-functional approach can overcome resistance mechanisms within the TME and achieve more comprehensive treatment outcomes [119]. Moreover, the combination of advanced nanomaterials with GBM in nanomedicine design has led to the development of biodegradable and biocompatible nanocomposites that reduce potential toxicities and facilitate the targeted release of therapeutic and diagnostic agents based on the patient’s specific tumor characteristics [120]. GBM nanocomposites are promising candidates for personalized anticancer nanomedicine owing to their unique advancements. Surface modification of GBMs enhances biocompatibility and colloidal stability behavior in biological environments. Covalent functionalization with polymers like polyethylene glycol (PEG), poly(vinyl alcohol) (PVA), and chitosan improves pharmacokinetics, reduces non-specific interactions, and facilitates targeted drug delivery. Additionally, conjugating graphene-based nanocarriers with specific ligands (e.g., peptides, folic acid, aptamers, and monoclonal antibodies) enables targeted drug delivery, increasing intratumor drug concentration. Beyond drug delivery, functionalized graphene materials show potential for multifunctional combinational applications, including PTT and PDT, strengthening their NIR light absorption capabilities [120,121,122,123].
Personalized GBM nanomedicines can utilize the patient's unique molecular profile to selectively target specific biomarkers. By leveraging patient-specific tumor characteristics, GBM nanocomposites enhance precision medicine approaches, improving treatment efficacy and reducing side effects. This innovation in nanomedicine design paves the way for safer and more efficient cancer therapies. For these applications, GBM nanocomposites can be engineered to recognize and bind to specific biomarkers expressed by tumor cells or the surrounding TME, such as overexpressed receptors or abnormal extracellular matrix (ECM) components [124,125,126]. Incorporating biomarkers into GBMs for personalized therapy involves identifying unique molecular features of a patient's tumor to tailor treatment. Tumors exhibit distinct genetic, proteomic, and metabolic profiles that influence their behavior and response to therapies. By functionalizing GBM nanomedicines with specific ligands, antibodies, or peptides that target these biomarkers, it is possible to precisely deliver therapeutic agents to tumor cells within the microenvironment. This approach enables the reprogramming of the TME by selectively targeting pro-tumorigenic cells (such as tumor-associated macrophages or fibroblasts) and reprogramming them to an anti-tumor phenotype. Moreover, biomarkers can also guide the design of nanomedicines that overcome resistance mechanisms, such as hypoxia or immune evasion. This personalized approach enhances the efficiency of treatment, reduces off-target effects, and allows for more effective modulation of the TME, ultimately improving therapeutic outcomes.
GBM nanocomposites can also be engineered to trigger or enhance immune responses, such as targeting immune checkpoint inhibitors or activating dendritic cells, that have shown promising results in personalized cancer treatment. Such therapies can target both tumor cells and immune cells within the TME, potentially enhancing immunotherapies by stimulating both innate and adaptive immune responses [126]. One of the primary challenges in immunotherapy is immune evasion by tumors, which limits the effectiveness of current treatments. Among GBMs, GO and rGO offer a novel approach to overcoming such limitations by facilitating immune activation to suppress tumorigenesis. GBMs can trigger immunomodulatory effects by interacting with TLRs (Toll-like receptors) stimulating the expression of tumor necrosis factor (TNF)-α, and interleukin (IL)-10, IL-1α and IL-6. Moreover, GBMs can be engaged in macrophage autophagy through the TLR pathways further inducing immunogenic cell death (ICD), especially through the TLR4 and TLR3 signaling, being essential parts of ICD-stimulated immunogenicity and cytotoxic effects of both CD8+ T cells and NK cells [126,127,128]. GBM nanocomposites, GO and rGO, have been studied as multifunctional and biodegradable vaccine carriers for personalized cancer immunotherapy. Traditional vaccines face challenges in sustaining antigen presentation and efficiently stimulating both CD4+ and CD8+ T cell responses, especially with neoantigens derived from tumor mutations. In a recent study by Xu et al. [129] pegylated rGO was functionalized with neoantigens and CpG oligodeoxynucleotide (CpG ODN being a TLR-9 agonist) for the development of personalized rGO cancer vaccines for neoantigen-specific CD8+ T cell responses. PET imaging with 64Cu demonstrated that rGO-PEG significantly enhanced vaccine delivery to lymph nodes (>100-fold compared to soluble vaccines). Additionally, rGO-PEG induced ROS generation in DCs (dendritic cells), leading to endolysosomal alkalization and promoting sustained antigen presentation to T cells. In vivo, a single dose of rGO-PEG vaccination generated strong CD8+ T cell responses and eliminated tumors, making it a promising nanoplatform for personalized cancer vaccination. While research has primarily focused on CD8+ T cell enhancement and antigen presentation [130], there are still many unexplored areas that could further improve GO-based cancer immunotherapy. One particularly promising direction is the modulation of immune evasion pathways, such as the PD-L1/PD-1 axis, which plays a critical role in suppressing immune responses in the TME [126].
4. Conclusions
The reprogramming of the TME plays a crucial role in cancer cell survival, immune evasion, and therapeutic resistance. Cancer cells shift from oxidative phosphorylation to aerobic glycolysis (Warburg effect) and exploit glucose and glutamine metabolism to sustain energy production and biosynthesis. These metabolic alterations influence stromal and immune cells, contributing to immune suppression and therapy resistance. Targeting these metabolic pathways using nanomedicine and other advanced therapeutic strategies has shown promising potential, but challenges persist due to the complexity and adaptability of TME metabolism. Recent advances in graphene-based materials (GBMs) have opened new opportunities for targeting TME metabolism. GBMs enhance TME targeting drug delivery, promote photothermal and photodynamic therapies, and induce oxidative stress in cancer cells. Additionally, metabolic inhibitors targeting glucose and glutamine metabolism offer potential strategies to starve cancer cells and improve immunotherapy outcomes. However, the complexity and adaptability of cancer metabolism pose challenges, necessitating further research into metabolic pathways, oxidative stress mechanisms, and tumor metabolome analysis. Overcoming these challenges could enhance the efficacy of combinational therapies and pave the way for more effective TME modulating cancer treatments.
5. Future Directions
To enhance the therapeutic effectiveness of GBMs in TME targeting cancer treatment, it is essential to gain deeper insights into the mechanisms of oxidative stress, autophagy, and metabolic heterogeneity within cancer cells (Figure 6). Oxidative stress refers to the imbalance between the production of ROS and the cell's ability to neutralize them with the production of antioxidants. These elevated ROS levels can damage cellular components such as DNA, proteins, and lipids, potentially leading to mutations and further promoting cancer cell survival and proliferation. However, cancer cells are also equipped with defense mechanisms to adapt to oxidative stress, such as the upregulation of antioxidant enzymes. Understanding how oxidative stress contributes to tumor survival and resistance to treatments is essential for developing strategies that can either enhance the oxidative stress within tumors or block the cellular defense mechanisms. GBMs have excellent electron transfer capabilities, which can potentially interact or even modulate ROS levels. Thus, the development of GBM nanodrugs can help in the regulation of tumor oxidative stress by either increasing ROS levels to induce cancer cell death or reducing ROS to protect normal cells. For GBMs to inhibit the antioxidant defense mechanism in cancer cells the selective targeting of antioxidant enzymes can be induced blocking their protective mechanism and rendering tumor cells to be more susceptible to treatments like chemotherapy or radiation therapy. The combination of GBMs with traditional cancer therapies (chemotherapy, radiation) can create synergistic effects, thus by increasing oxidative stress, GBM nanodrugs could potentially enhance the efficacy of chemotherapy and radiation therapies. In addition to ROS modulation, GBMs can be functionalized to target multiple pathways involved in tumor survival, such as growth factors, signaling pathways, or drug efflux pumps. Tumors adapt to chronic oxidative stress by activating pathways like NRF2 (Nuclear Factor Erythroid 2-related Factor 2), which upregulates antioxidant proteins. By using GBMs to probe these pathways or to interfere with their activation, the understanding of tumors resistance to oxidative damage can be promoted and new strategies can be developed to reverse this adaptation [131].
Autophagy is another crucial cellular process involved in cancer progression and therapy resistance. It is a double-edged sword mechanism by which cancer cells degrade and recycle damaged or unnecessary components to survive under nutrient-deprived or stressful conditions providing them a survival advantage to resist to therapeutics. But can also serve as a tumor suppressor mechanism through the disruption of autophagy processes or excessive autophagic flux further promoting apoptotic cell death. Thus, the role of autophagy in cancer therapy is complex, as it can either support tumor survival or, if manipulated correctly, lead to cell death. Research into autophagy inhibitors or activators is rapidly growing, and future therapeutic approaches may seek to fine-tune this process in cancer cells. GBMs, due to their large surface area and the ability to be functionalized with various molecules, can be used as a delivery platforms for autophagy-modulating drugs. These drugs could either enhance or inhibit autophagy in cancer cells to induce cell death or prevent cancer cell survival. For example, autophagy-inducing agents can be delivered by GBMs to activate autophagy potentially forcing cancer cells to undergo autophagic cell death. By specifically functionalizing GBMs to target autophagic pathways, they could assist either in inhibiting cancer cells’ survival or in promoting cell death. Such functionalization could be followed by peptides or agonists binding to autophagy proteins such as Beclin-1 agonist, mTOR inhibitors such as rapamycin, ligands for targeting autophagy-related receptors such as LC3 (microtubule-associated light chain 3 involved in autophagosome formation), monoclonal antibodies specific to autophagy-regulating proteins or receptors like Beclin-1, LC3, or PI3K (phosphoinositide 3 kinase) that could improve the selectivity of the GBMs. The interaction of GBMs with autophagy signaling pathways could affect key autophagy-regulating proteins, such as Beclin-1, LC3, mTOR, and AMPK (AMP-activated protein kinase) either enhancing or suppressing autophagy thus modulating the survival or death of cancer cells. In cancer cells, autophagy often serves as a survival mechanism under oxidative stress. By using GBMs to stimulate oxidative stress, it may be possible to overwhelm cancer cells, promoting a shift from autophagic survival to autophagic cell death. By either inhibiting or activating autophagy, GBMs could enhance the efficacy of chemotherapy and radiation therapy by preventing cancer cells from repairing the damage induced by the treatments. Therefore, autophagy modulation could be integrated with GBM as a valuable strategy in overcoming therapy resistance [132].
These factors play a critical role in tumor progression and drug resistance, making them key areas of focus for improving GBM-based therapeutic strategies that are known for their ability to induce localized oxidative stress. Future research should aim to overcome the metabolic flexibility of cancer cells, which allows them to adapt and survive under stress conditions. By exploring the ways in which these mechanisms influence cancer cell behavior, researchers can develop more targeted and effective treatment strategies that harness the unique properties of GBMs. Integrating multi-modal therapeutic approaches, which combine GBMs with other treatments including PTT/PDT, ultrasound therapy and magnetic targeting, could provide a more direct strategy for tackling cancers metabolic plasticity. Moreover, GBMs can be used to deliver drugs directly to tumors, disrupt the tumor microenvironment, or even induce localized oxidative stress. When combined with other treatments, such as targeted therapies, chemotherapy, or immunotherapy, these materials could enhance the overall therapeutic efficacy by attacking multiple vulnerabilities in cancer cells. A multi-pronged approach would not only help tackle the tumor's metabolic plasticity but also prevent resistance from developing, offering a more comprehensive and potent strategy against cancer. Moreover, therapies designed to inhibit key metabolic enzymes or pathways could potentially limit the ability of cancer cells to adapt to this environment. Future research on identifying specific metabolic vulnerabilities and designing therapeutic approaches to deprive the cellular energy sources rendering them more susceptible to treatment could open the way for more effective therapies [133].
A crucial direction for future research will be the development of personalized treatment based on the metabolic profile of individual tumors. Refining the analysis of tumor metabolomes will offer a direct understanding of the metabolic profiles specific to different cancer types, thereby enhancing the precision and effectiveness of metabolic-targeted therapies. The complexity of cancer metabolism has long been an obstacle to developing highly effective therapies. The unique metabolic profiles exhibited by distinct cancer types, in combination with metabolic alterations within the same type, demonstrate significant metabolic heterogeneity. By identifying unique metabolic signatures, a more precise understanding could unlock new potentials for metabolic-targeted therapies. The integration of advanced technologies like metabolomics and proteomics, combined with GBMs, could facilitate the identification of key metabolic targets and enable the design of highly personalized therapeutic regimens. The properties of GBMs align perfectly with the increased level of specificity and effectiveness required for metabolic-targeted therapies. Moreover, GBMs can interact with the TME inducing oxidative stress and modulating the metabolic pathways of cancer cells enhancing the therapeutic effect, as presented in this review. The combination of tumor metabolomics with GBM targeted therapies could be crucial for the development of precise and more effective personalized therapies improving cancer treatment outcomes [134].
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Alhazmi, H.A.; Ahsan, W.; Mangla, B.; Javed, S.; Hassan, M.Z.; et al. Graphene-based biosensors for disease theranostics: Development, applications, and recent advancements. Nanotechnol. Rev. 2022, 11, 96. [Google Scholar] [CrossRef]
- Jampilek, J.; Kralova, K. Advances in Drug Delivery Nanosystems Using Graphene-Based Materials and Carbon Nanotubes. Materials 2021, 14, 1059. [Google Scholar] [CrossRef]
- Shafiee, A.; Iravani, S.; Varma, R.S. Graphene and graphene oxide with anticancer applications: Challenges and future perspectives. Med. Comm. 2022, 3, e118. [Google Scholar] [CrossRef] [PubMed]
- Itoo, A.M.; Vemula, S.L.; Gupta, M.T.; Giram, M.V.; Kumar, S.A.; et al. Multifunctional graphene oxide nanoparticles for drug delivery in cancer. J. Contr. Rel. 2022, 350, 26. [Google Scholar] [CrossRef]
- Hosseini, S.M.; Mohammadnejad, J.; Najafi-Taher, R.; Zadeh, Z.B.; Tanhaei, M.; et al. Multifunctional Carbon-Based Nanoparticles: Theranostic Applications in Cancer Therapy and Diagnosis. ACS Appl. Bio Mater. 2023, 6, 1323. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Chakraborty, S.; Monikh, F.A.; Varsou, D.D.; Chetwynd, A.J.; et al. Functionalization of Graphene-Based Materials: Biological Behavior, Toxicology, and Safe-By-Design Aspects. Adv Biol (Weinh). 2021, 5, e2100637. [Google Scholar] [CrossRef]
- Fadeel, B.; Busy, C.; Merino, S.; Vazquez, E.; Flahaut, E.; et al. Safety Assessment of Graphene-Based Materials: Focus on Human Health and the Environment. ACS Nano. 2018, 12, 10582. [Google Scholar] [CrossRef]
- Angelopoulou, A.; Papachristodoulou, M.; Voulgari, E.; Mouikis, A.; Zygouri, P.; et al. Paclitaxel-Loaded, Pegylated Carboxylic Graphene Oxide with High Colloidal Stability, Sustained, pH-Responsive Release and Strong Anticancer Effects on Lung Cancer A549 Cell Line. Pharmaceutics 2024, 16, 1452. [Google Scholar] [CrossRef]
- Sattari, S.; Adeli, M.; Beyranvand, S.; Nemati, M. Functionalized Graphene Platforms for Anticancer Drug Delivery. I.J.Naanomed. 2021, 16, 5955. [Google Scholar] [CrossRef]
- Angelopoulou, A.; Voulgari, E.; Diamanti, E.K.; Gournis, D.; Avgoustakis, K. Graphene oxide stabilized by PLA–PEG copolymers for the controlled delivery of paclitaxel. Eur. J. Pharm. Biopharm. 2015, 93, 18. [Google Scholar] [CrossRef]
- Rahimi, S.; Chen, Y.; Zareian, M.; Pandit, S.; Mijakovic, I. Cellular and subcellular interactions of graphene-based materials with cancerous and non-cancerous cells. Adv. Drug Deliv. Rev. 2022, 189, 114467. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yang, Y.; Li, K.; Li, J. Application of graphene oxide in tumor targeting and tumor therapy. J Biomater Sci Polym Ed. 2023, 34, 2551. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, X.; Yue, H. Two-dimensional nanomaterials induced nano-bio interfacial effects and biomedical applications in cancer treatment. J. Nanobiotechnol. 2024, 22, 67. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.; Tang, Y.Q.; Miao, H. Metabolism in tumor microenvironment: Implications for cancer immunotherapy. MedComm 2020, 1, 47. [Google Scholar] [CrossRef]
- Avgoustakis, K.; Angelopoulou, A. Biomaterial-Based Responsive Nanomedicines for Targeting Solid Tumor Microenvironments. Pharmaceutics 2024, 16, 179. [Google Scholar] [CrossRef]
- Kimmelman, A.C.; Sherman, M.H. The Role of stroma in Cancer Metabolism. Cold Spring Harb Perspect Med. 2024, 14, a041540. [Google Scholar] [CrossRef] [PubMed]
- Schworer, S.; Vardhana, S.A.; Thompson, C.B. Cancer metabolism drives a stromal regenerative response. Cell Metab. 2019, 29, 576. [Google Scholar] [CrossRef]
- Ahmed, N.; Escalona, R.; Leung, D.; Chan, E.; Kannourakis, G. Tumour microenvironment and metabolic plasticity in cancer and cancer stem cells: Perspectives on metabolic and immune regulatory signatures in chemoresistant ovarian cancer stem cells. Semin. Cancer Biol. 2018, 53, 265. [Google Scholar] [CrossRef]
- Arner, E.N.; Rathmell, J.C. Metabolic Programming and Immune Suppression in the Tumor Microenvironment. Cancer Cell. 2023, 41, 421. [Google Scholar] [CrossRef]
- Zhao, Y.; Ye, X.; Xiong, Z.; Ihsan, A.; Ares, I.; et al. Cancer Metabolism: The Role of ROS in DNA Damage and Induction of Apoptosis in Cancer Cells. Metabolites 2023, 13, 796. [Google Scholar] [CrossRef]
- Schiliro, C.; Firestein, B.L. Mechanisms of Metabolic Reprogramming in Cancer Cells Supporting Enhanced Growth and Proliferation. Cells 2021, 10, 1056. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Alesi, G.N.; Kang, S. Glutaminolysis as a target for cancer therapy. Oncogene. 2016, 35, 3619. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Zou, T.; Shen, X.; Nelson, P.J.; Li, J.; et al. Lipid metabolism in cancer progression and therapeutic strategies. MedComm, 2020, 2, 27. [Google Scholar] [CrossRef]
- Zaidi, N.; Lupien, L.; Kuemmerle, N.B.; Kinlaw, W.B.; Swinnen, J.V.; et al. Lipogenesis and lipolysis: the pathways exploited by the cancer cells to acquire fatty acids. Prog Lipid Res. 2013, 52, 585. [Google Scholar] [CrossRef]
- Maffuid, K.; Cao, Y. Decoding the Complexity of Immune–Cancer Cell Interactions: Empowering the Future of Cancer Immunotherapy. Cancers 2023, 15, 4188. [Google Scholar] [CrossRef]
- Kim, J.; DeBerardinis, R.J. Mechanisms and Implications of Metabolic Heterogeneity in Cancer. Cell Metab. 2019, 30, 434. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Hua, G.; Hu, X. Lipid metabolism associated crosstalk: the bidirectional interaction between cancer cells and immune/stromal cells within the tumor microenvironment for prognostic insight. Cancer Cell Int. 2024, 24, 295. [Google Scholar] [CrossRef]
- Chang, C.H.; Qiu, J.; O'Sullivan, D.; Buck, M.D.; Noguchi, T.; et al. Metabolic Competition in the Tumor Microenvironment Is a Driver of Cancer Progression. Cell. 2015, 162, 1229. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Koh, J.; Song, S.G.; Yim, J.; Kim, M.; et al. High tumor hexokinase-2 expression promotes a pro-tumorigenic immune microenvironment by modulating CD8+/regulatory T-cell infiltration. BMC Cancer. 2022, 22, 1120. [Google Scholar] [CrossRef]
- Qian, Y.; Yin, Y.; Zheng, X.; Liu, Z.; Wang, X. Metabolic regulation of tumor-associated macrophage heterogeneity: insights into the tumor microenvironment and immunotherapeutic opportunities. Biomark Res, 2024, 12, 1. [Google Scholar] [CrossRef]
- Ren, M.; Zheng, X.; Gao, H.; Jiang, A.; Yao, Y.; et al. Nanomedicines Targeting Metabolism in the Tumor Microenvironment. Front. Bioeng. Biotechnol. 2022, 10, 943906. [Google Scholar] [CrossRef]
- Roma-Rodrigues, C.; Pombo, I.; Raposo, L.; Pedrosa, P.; Fernandes, A.R.; et al. Nanotheranostics Targeting the Tumor Microenvironment. Front. Bioeng. Biotechnol 2019, 7, 197. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Zhang, Y.; Zhao, Z.; Gao, M. Developing targeted therapies for neuroblastoma by dissecting the effects of metabolic reprogramming on tumor microenvironments and progression. Theranostics. 2024, 14, 3439. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, L.; Zhang, Z.; Wu, P.; Zhang, Y; et al. Advances in targeting tumor microenvironment for immunotherapy. Front. Immunol. 2024, 15, 1472772. [Google Scholar] [CrossRef]
- Bhat, A.A.; Nisar, S.; Singh, M.; Ashraf, B.; Masoodi. Et al. Cytokine- and chemokine-induced inflammatory colorectal tumor microenvironment: Emerging avenue for targeted therapy. Cancer Commun. 2022, 42, 689. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Guo, S.; Deng, J.; Shen, J.; Du, F.; et al. VEGF/VEGFR-Targeted Therapy and Immunotherapy in Non-small Cell Lung Cancer: Targeting the Tumor Microenvironment. Int J Biol Sci. 2022, 18, 3845. [Google Scholar] [CrossRef]
- Yamaguchi, R.; Perkins, G. Challenges in targeting cancer metabolism for cancer therapy. EMBO Rep. 2012, 13, 1034. [Google Scholar] [CrossRef]
- Teicher, B.A.; Marston Linehan, W.; Helman, L.J. Targeting Cancer Metabolism. Clin Cancer Res., 2012, 18, 5537. [Google Scholar] [CrossRef]
- Saadh, M.J.; Mustafa, M.A.; Qassem, L.Y.; Ghadir, G.K.; Alaraj, M.; et al. Targeting hypoxic and acidic tumor microenvironment by nanoparticles: A review. J. Drug Deliv. Sci. Technol., 2024, 96, 105660. [Google Scholar] [CrossRef]
- Zhou, Y.; Yuan, J.; Xu, K.; Li, S.; Liu, Y. Nanotechnology Reprogramming Metabolism for Enhanced Tumor Immunotherapy. ACS Nano. 2024, 18, 1846. [Google Scholar] [CrossRef]
- Zhang, X.; An, M.; Zhang, J.; Zhao, Y.; Liu, Y. Nano-medicine therapy reprogramming metabolic network of tumour microenvironment: new opportunity for cancer therapies. J. Drug Target 2024, 32, 241. [Google Scholar] [CrossRef] [PubMed]
- Iannazzo, D.; Espro, C.; Celesti, C.; Ferlazzo, A.; Neri, G. Smart Biosensors for Cancer Diagnosis Based on Graphene Quantum Dots. Cancers 2021, 13, 3194. [Google Scholar] [CrossRef] [PubMed]
- Pourmadadi, M.; Soleimani Dinani, H.; Saeidi Tabar, F.; Khassi, K.; Janfaza, S.; Tasnim, N.; Hoorfar, M. Properties and Applications of Graphene and Its Derivatives in Biosensors for Cancer Detection: A Comprehensive Review. Biosensors 2022, 12, 269. [Google Scholar] [CrossRef] [PubMed]
- Uzdrowska, K.; Knap, N.; Gulczynski, J.; Kuban-Jankowska, A.; Struck-Lewicka, W.; et al. Chasing Graphene-Based Anticancer Drugs: Where are We Now on the Biomedical Graphene Roadmap? Int J Nanomedicine. 2024, 19, 3973. [Google Scholar] [CrossRef]
- Tabish, T.A.; Zhang, S.; Winyard, P.G. Developing the next generation of graphene-based platforms for cancer therapeutics: The potential role of reactive oxygen species. Redox Biology, 2018, 15, 34. [Google Scholar] [CrossRef]
- White, E.; Mehnert, J.M.; Chan, C.S. Autophagy, Metabolism, and Cancer. Clin Cancer Res. 2015, 21, 5037. [Google Scholar] [CrossRef]
- Poillet-Perez, L.; White, E. Role of tumor and host autophagy in cancer metabolism. Genes Dev. 2019, 33, 610. [Google Scholar] [CrossRef]
- Zhang, X.; Shao, S.; Song, N.; Yang, B.; Liu, F.; et al. Integrated omics characterization reveals reduced cancer indicators and elevated inflammatory factors after thermal ablation in non-small cell lung cancer patients. Respir Res 2024, 25, 309. [Google Scholar] [CrossRef]
- Leone, R.D.; Powell, J.D. Metabolism of immune cells in cancer. Nat Rev Cancer 2020, 20, 516. [Google Scholar] [CrossRef]
- Navas, L.E.; Carnero, A. NAD+ metabolism, stemness, the immune response, and cancer. Sig Transduct Target Ther 2021, 6, 2. [Google Scholar] [CrossRef]
- Jin, J.; Byun, J.-K.; Choi, Y.-K.; Park, K.-G. Targeting glutamine metabolism as a therapeutic strategy for cancer. Exp Mol Med 2023, 55, 706. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Liu, Z.; Liu, W.; Liu, Z. Current knowledge and potential intervention of hexosamine biosynthesis pathway in lung cancer. World J Surg Onc 2023, 21, 334. [Google Scholar] [CrossRef]
- An, X.; Yu, W.; Liu, J.; Tang, D.; Yang, L.; et al. Oxidative cell death in cancer: mechanisms and therapeutic opportunities. Cell Death Dis 2024, 15, 556. [Google Scholar] [CrossRef] [PubMed]
- Arfin, S.; Jha, N.K.; Jha, S.K.; Kesari, K.K.; Ruokolainen, J.; et al. Oxidative Stress in Cancer Cell Metabolism. Antioxidants 2021, 10, 642. [Google Scholar] [CrossRef] [PubMed]
- Perini, G.; Palmieri, V.; Ciasca, G.; Primiano, A.; Gervasoni, J.; et al. Functionalized Graphene Quantum Dots Modulate Malignancy of Glioblastoma Multiforme by Downregulating Neurospheres Formation. C — J. Carbon Research 2021, 7, 4. [Google Scholar] [CrossRef]
- Szlasa, W.; Zendran, I.; Zalesinska, A.; Tarek, M.; Kulbacka, J. Lipid composition of the cancer cell membrane. J Bioenerg Biomembr. 2020, 52, 321. [Google Scholar] [CrossRef]
- Perini, G.; Palmieri, V.; Friggeri, G.; et al. Carboxylated graphene quantum dots-mediated photothermal therapy enhances drug-membrane permeability, ROS production, and the immune system recruitment on 3D glioblastoma models. Cancer Nano 2023, 14, 13. [Google Scholar] [CrossRef]
- Campos-Sandoval, J.A.; Gomez-Garcia, M.C.; de los Santos-Jimenez, J.; Mates, J.M.; Alonso, F.J.; et al. Antioxidant responses related to temozolomide resistance in glioblastoma. Neurocchem. Intern. 2021, 149, 105136. [Google Scholar] [CrossRef]
- Campbell, E.; Hasan, M.T.; Gonzalez-Rodriguez, R.; Truly, T.; Lee, B.H.; et al. Graphene quantum dot formulation for cancer imaging and redox-based drug delivery. Nanomed.: Nanotechnol. Biol. Med. 2021, 37, 102408. [Google Scholar] [CrossRef]
- Campbell, E.; Hasan, M.T.; Gonzalez-Rodriguez, R.; Akkaraju, G.; Naumov, A.V. Doped Graphene Quantum Dots for Intracellular Multicolor Imaging and Cancer Detection. ACS Biomater. Sci. Engineer. 2019, 5, 4671. [Google Scholar] [CrossRef]
- Tomar, V.; Kumar, P.; Sharma, D.; Joshi, R.K.; Nemiwal, M. Anticancer potential of ferrocene-containing derivatives: Current and future prospective. J. Mol. Struct. 2025, 1319, 139589. [Google Scholar] [CrossRef]
- Li, W.; Yu, J.; Wang, J.; Fan, X.; Xu, X.; et al. How does ferrocene correlate with ferroptosis? Multiple approaches to explore ferrocene-appended GPX4 inhibitors as anticancer agents. Chem Sci. 2024, 15, 10477. [Google Scholar] [CrossRef] [PubMed]
- Favaron, C.; Gabano, E.; Zanellato, I.; Gaiaschi, L.; Casali, C.; et al. Effects of Ferrocene and Ferrocenium on MCF-7 Breast Cancer Cells and Interconnection with Regulated Cell Death Pathways. Molecules 2023, 28, 6469. [Google Scholar] [CrossRef]
- Guo, M.; Xiang, H.-J.; Wang, Y.; Zhang, Q.-L.; An, L.; et al. Ruthenium nitrosyl functionalized graphene quantum dots as an efficient nanoplatform for NIR-light-controlled and mitochondria-targeted delivery of nitric oxide combined with photothermal therapy. Chem. Commun., 2017, 53, 3253. [Google Scholar] [CrossRef]
- Poderoso, J.J.; Helfenberger, K.; Poderoso, C. The effect of nitric oxide on mitochondrial respiration. Nitric Oxide. 2019, 88, 61. [Google Scholar] [CrossRef] [PubMed]
- Tengan, C.H.; Moraes, C.T. NO control of mitochondrial function in normal and transformed cells. Biochim Biophys Acta. 2018, 1858, 573. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Myers, C.R.; Wang, Y.; You, M. Mitochondria as a Novel Target for Cancer Chemoprevention: Emergence of Mitochondrial-targeting Agents. Cancer Prev Res (Phila). 2021, 14, 285. [Google Scholar] [CrossRef]
- Fan, Z.; Nie, Y.; Wei, Y.; Zhao, J.; Liao, X.; et al. Facile and large-scale synthesis of graphene quantum dots for selective targeting and imaging of cell nucleus and mitochondria. Mater. Sci. Eng. C. 2019, 103, 109824. [Google Scholar] [CrossRef]
- Zhang, D.; Wen, L.; Huang, R.; Wang, H.; Hu, X.; et al. Mitochondrial specific photodynamic therapy by rare-earth nanoparticles mediated near-infrared graphene quantum dots. Biomater. 2018, 153, 14. [Google Scholar] [CrossRef]
- Dong, X.; Yu, X.; Lu, M.; Xu, Y.; Zhou, L.; et al. Quantitative chemical proteomics reveals that phenethyl isothiocyanate covalently targets BID to promote apoptosis. Cell Death Discov. 2024, 10, 456. [Google Scholar] [CrossRef]
- Tang, L.; Zhang, Y. Mitochondria are the primary target in isothiocyanate-induced apoptosis in human bladder cancer cells Mol. Cancer Ther 2005, 4, 1250. [Google Scholar] [CrossRef] [PubMed]
- Saeed, L.M.; Mahmood, M.; Pyrek, S.J.; Fahmi, T.; Yang, X.; et al. Single-walled carbon nanotube and graphene nanodelivery of gambogic acid increases its cytotoxicity in breast and pancreatic cancer cells. J. Appl. Toxicol. 2014, 34, 1188. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Sun, Q.; Chen, S.; Ma, D.; Qi, Y.; et al. pH-responsive hydrogel with gambogic acid and calcium nanowires for promoting mitochondrial apoptosis in osteosarcoma. J. Control. Real. 2025, 377, 563. [Google Scholar] [CrossRef]
- Seo, M.J.; Lee, D.M.; Kim, I.Y.; Lee, D.; Choi, M.-K.; et al. Gambogic acid triggers vacuolization-associated cell death in cancer cells via disruption of thiol proteostasis. Cell Death Dis. 2019, 10, 187. [Google Scholar] [CrossRef]
- Hatami, E.; Jaggi, M.; Chauhan, S.C.; Yallapu, M.M. Gambogic acid: A shining natural compound to nanomedicine for cancer therapeutics. Biochim Biophys Acta Rev Cancer. 2020, 1874, 188381. [Google Scholar] [CrossRef]
- Li, Y.; Liu, Y.; Fu, Y.; Wei, T.; Le Guyader, L.; et al. The triggering of apoptosis in macrophages by pristine graphene through the MAPK and TGF-beta signaling pathways. Biomaterials 2012, 33, 402. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Zhang, C.; Ma, T.; Zhang, C.; Luo, J.; et al. Hypericin-functionalized graphene oxide for enhanced mitochondria-targeting and synergistic anticancer effect. Acta Biomater. 2018, 77, 268. [Google Scholar] [CrossRef]
- Choudhary, N.; Collignon, T.E.; Tewari, D.; Bishayee, A. Hypericin and its anticancer effects: From mechanism of action to potential therapeutic application. Phytomed. 2022, 105, 154356. [Google Scholar] [CrossRef]
- Lenkavska, L.; Blascakova, L.; Jurasekova, Z.; Macajova, M.; Bolcik, B.; et al. Benefits of hypericin transport and delivery by low- and high-density lipoproteins to cancer cells: From in vitro to ex ovo. Photodiagnosis Photodyn Ther. 2019, 25, 214. [Google Scholar] [CrossRef]
- Bulkova, V.; Vargova, J.; Babincak, M.; Jendzelovsky, R.; Zdrahal, Z.; et al. New findings on the action of hypericin in hypoxic cancer cells with a focus on the modulation of side population cells. Biomed Pharmacother. 2023, 163, 114829. [Google Scholar] [CrossRef]
- Miccoli, L.; Beurdeley-Thomas, A.; De Pinieux, G.; Sureau, S.; Oudard, S.; et al. Light-induced Photoactivation of Hypericin Affects the Energy Metabolism of Human Glioma Cells by Inhibiting Hexokinase Bound to Mitochondria. Cancer Res. 1998, 58, 5777. [Google Scholar] [PubMed]
- Theodossiou, T.A.; Hothersall, J.S.; De Witte, P.A.; Pantos, A.; Agostinis, P. The Multifaceted Photocytotoxic Profile of Hypericin. Mol. Pharmaceutics 2009, 6, 1775. [Google Scholar] [CrossRef]
- Wu, C.; Wang, L.; Tian, Y.; Guan, X.; Liu, Q.; et al. “Triple-Punch” Anticancer Strategy Mediated by Near-Infrared Photosensitizer/CpG Oligonucleotides Dual-Dressed and Mitochondria-Targeted Nanographene. ACS Appl. Mater. Interfaces 2018, 10, 6942. [Google Scholar] [CrossRef] [PubMed]
- Zielonka, J.; Joseph, J.; Sikora, A.; Hardy, M.; Ouari, O.; et al. Mitochondria-Targeted Triphenylphosphonium-Based Compounds: Syntheses, Mechanisms of Action, and Therapeutic and Diagnostic Applications. Chem Rev. 2017, 117, 10043. [Google Scholar] [CrossRef] [PubMed]
- Hanagata, N. Structure-dependent immunostimulatory effect of CpG oligodeoxynucleotides and their delivery system. Int J Nanomedicine. 2012, 7, 2181. [Google Scholar] [CrossRef]
- Zeng, W.-N.; Yu, Q.-P.; Wang, D.; Liu, J.-L.; Yang, Q.-J.; et al. Mitochondria-targeting graphene oxide nanocomposites for fluorescence imaging-guided synergistic phototherapy of drug-resistant osteosarcoma. J Nanobiotechnol. 2021, 19, 79. [Google Scholar] [CrossRef]
- Wang, H.; Li, X.; Tse, B.W.-C.; Yang, H.; Thorling, C.A.; et al. Indocyanine green-incorporating nanoparticles for cancer theranostics. Theranostics. 2018, 8, 1227. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Zhou, F.; Zhang, D.; Chen, Q.; Xing, D. A graphene oxide based smart drug delivery system for tumor mitochondria-targeting photodynamic therapy. Nanoscale, 2016, 8, 3530. [Google Scholar] [CrossRef]
- Zhu, H.; Zhang, B.; Zhu, N.; Li, M.; Yu, Q. Mitochondrion targeting peptide-modified magnetic graphene oxide delivering mitoxantrone for impairment of tumor mitochondrial functions. Chin. Chem. Lett. 2021, 32, 1220. [Google Scholar] [CrossRef]
- Evison, B.J.; Sleebs, B.E.; Watson, K.G.; Phillips, D.R.; Cutts, S.M. Mitoxantrone, More than Just Another Topoisomerase II Poison. Med. Res. Rev. 2015, 36, 248. [Google Scholar] [CrossRef]
- Kim, C.-W.; Choi, K.-C. Effects of anticancer drugs on the cardiac mitochondrial toxicity and their underlying mechanisms for novel cardiac protective strategies. Life Sci. 2021, 277, 119607. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Yu, Q.; Zhang, Y.-M.; Liu, Y. Two-dimensional supramolecular assemblies based on β-cyclodextrin-grafted graphene oxide for mitochondrial dysfunction and photothermal therapy. Chem. Commun., 2019, 55, 12200. [Google Scholar] [CrossRef] [PubMed]
- Verwilst, P.; Han, J.; Lee, J.; Mun, S.; Kang, H.-G.; et al. Reconsidering azobenzene as a component of small-molecule hypoxia-mediated cancer drugs: A theranostic case study. Biomater. 2017, 115, 104. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, Z.; Zheng, Y.; Geng, Y.; Han, C.; et al. Glycyrrhetinic Acid Functionalized Graphene Oxide for Mitochondria Targeting and Cancer Treatment In Vivo. Small. 2017, 14, 1703306. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Li, L.; Peng, Q.; Gan, C.; Gao, L.; et al. Glycyrrhetinic acid restricts mitochondrial energy metabolism by targeting SHMT2. iScience 2022, 25, 104349. [Google Scholar] [CrossRef]
- Song, J.; Ko, H.-S.; Sohn, E.J.; Kim, B.; Kim, J.H.; et al. Inhibition of protein kinase C α/βII and activation of c-Jun NH2-terminal kinase mediate glycyrrhetinic acid induced apoptosis in non-small cell lung cancer NCI-H460 cells. Bioorg. Med. Chem. Lett. 2014, 4, 1188. [Google Scholar] [CrossRef]
- Guo, Q.; Zhao, M.; Wang, Q.; Lu, T.; Luo, P.; et al. Glycyrrhetinic acid inhibits non-small cell lung cancer via promotion of Prdx6- and caspase-3-mediated mitochondrial apoptosis. Biomed. Pharmacother. 2024, 173, 116304. [Google Scholar] [CrossRef]
- Fiore, C.; Salvi, M.; Palermo, M.; Sinigaglia, G.; Armanini, D.; et al. On the mechanism of mitochondrial permeability transition induction by glycyrrhetinic acid. Biochim. Biophys. Acta – Bioenerg. 2004, 1658, 195. [Google Scholar] [CrossRef]
- Jiang, J.-H.; Pi, J.; Jin, H.; Cai, J.-Y. Functional graphene oxide as cancer-targeted drug delivery system toselectively induce oesophageal cancer cell apoptosis. Artif Cells Nanomed Biotechnol. 2018, 46, 5297. [Google Scholar] [CrossRef]
- Chauhan, A.; Joshi, H.; Kandari, D.; Aggarwal, D.; Chauhan, R.; et al. Oridonin: A natural terpenoid having the potential to modulate apoptosis and survival signaling in cancer. Phytomed. Plus. 2025, 5, 100721. [Google Scholar] [CrossRef]
- Haque, S.; Tripathy, S.; Patra, C.R. (2022). Graphene Based Nanomaterials for ROS-Mediated Cancer Therapeutics. In: Chakraborti, S. (eds) Handbook of Oxidative Stress in Cancer: Therapeutic Aspects. Springer, Singapore. pp 1-26. [CrossRef]
- Dash, B.S.; Jose, G.; Lu, Y.-J.; Chen, J.-P. Functionalized Reduced Graphene Oxide as a Versatile Tool for Cancer Therapy. Int. J. Mol. Sci. 2021, 22, 2989. [Google Scholar] [CrossRef] [PubMed]
- Kretowski, R.; Cechowska-Pasko, M. The Reduced Graphene Oxide (rGO) Induces Apoptosis, Autophagy and Cell Cycle Arrest in Breast Cancer Cells. Int. J. Mol. Sci. 2022, 23, 9285. [Google Scholar] [CrossRef]
- Zhang, J.; Cao, H.-Y.; Wang, J.-Q.; Wu, G.-D.; Wang, L. Graphene Oxide and Reduced Graphene Oxide Exhibit Cardiotoxicity Through the Regulation of Lipid Peroxidation, Oxidative Stress, and Mitochondrial Dysfunction. Front. Cell Dev. Biol. 2021, 9, 616888. [Google Scholar] [CrossRef]
- Vinothini, K.; Rajendran, N.K.; Rajan, M.; Ramu, A.; Marraiki, N.; et al. A magnetic nanoparticle functionalized reduced graphene oxide-based drug carrier system for a chemo-photodynamic cancer therapy. New J. Chem., 2020, 44, 5265. [Google Scholar] [CrossRef]
- Kretowski, R.; Szynaka, B.; Jablonska-Trypuc, A.; Kiełtyka-Dadasiewicz, A.; Cechowska-Pasko, M. The Synergistic Effect of Reduced Graphene Oxide and Proteasome Inhibitor in the Induction of Apoptosis through Oxidative Stress in Breast Cancer Cell Lines. Int. J. Mol. Sci. 2024, 25, 5436. [Google Scholar] [CrossRef]
- Tarjanyi, O.; Haerer, J.; Vecsernyes, M.; Berta, G.; Stayer-Harci, A. et a. Prolonged treatment with the proteasome inhibitor MG-132 induces apoptosis in PC12 rat pheochromocytoma cells. Sci Rep 2022, 12, 5808. [Google Scholar] [CrossRef]
- Zanotto-Filho, A.; Braganhol, E.; Battastini, A.M.O. Moreire, J.C.F. Proteasome inhibitor MG132 induces selective apoptosis in glioblastoma cells through inhibition of PI3K/Akt and NFkappaB pathways, mitochondrial dysfunction, and activation of p38-JNK1/2 signaling. Invest New Drugs 2012, 30, 2252. [Google Scholar] [CrossRef] [PubMed]
- Kavinkumar, T.; Varunkumar, K.; Ravikumar, V.; Manivannan, S. Anticancer activity of graphene oxide-reduced graphene oxide-silver nanoparticle composites. J. Colloid Interface Sci. 2017, 505, 1125. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.; Zhang, J.; Gu, Z.; Chen, Y. Nanocatalysts-augmented Fenton chemical reaction for nanocatalytic tumor therapy. Biomaterials. 2019, 211, 1. [Google Scholar] [CrossRef]
- Gurunathan, S.; Han, J.W.; Park, J.H.; Kim, E.; Choi, Y.; et al. Reduced graphene oxide–silver nanoparticle nanocomposite: a potential anticancer nanotherapy. I.J. Nanomed. 2015, 10, 6257. [Google Scholar] [CrossRef]
- Yuan, Y.-G.; Gurunathan, S. Combination of graphene oxide–silver nanoparticle nanocomposites and cisplatin enhances apoptosis and autophagy in human cervical cancer cells. Int J Nanomedicine. 2017, 12, 6537. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, L.; Tian, Y.; Gu, M.; Wang, Y.; et al. Autophagy-driven regulation of cisplatin response in human cancers: Exploring molecular and cell death dynamics. Cancer Let. 2024, 587. [Google Scholar] [CrossRef]
- Coreas, R.; Castillo, C.; Li, Z.; Yan, D.; Gao, Z.; et al. Biological impacts of reduced graphene oxide affected by protein corona formation. Chem Res Toxicol. 2022, 35, 1244. [Google Scholar] [CrossRef] [PubMed]
- Sawosz, E.; Jaworski, S.; Kutwin, M.; Vadalasetty, K.P.; Grodzik, M.; Wierzbicki, M.; Kurantowicz, N.; Strojny, B.; Hotowy, A.; Lipińska, L.; et al. Graphene Functionalized with Arginine Decreases the Development of Glioblastoma Multiforme Tumor in a Gene-Dependent Manner. Int. J. Mol. Sci. 2015, 16, 25214–25233. [Google Scholar] [CrossRef]
- Liu, Y.; Borchert, G.L.; Donald, S.; Diwan, B.; Anver, M.; et al. Proline oxidase functions as a mitochondrial tumor suppressor in human cancers. Cancer Res. 2009, 69, 6414. [Google Scholar] [CrossRef]
- Keshet, R.; Erez, A. Arginine and the metabolic regulation of nitric oxide synthesis in cancer. Dis. Model Mech. 2018, 11, dmm033332. [Google Scholar] [CrossRef] [PubMed]
- Hajipour Keyvani, A.; Mohammadnejad, P.; Pazoki-Toroudi, H.; Perez Gilabert, I.; Chu, T.; et al. Advancements in Cancer Treatment: Harnessing the Synergistic Potential of Graphene-Based Nanomaterials in Combination Therapy. ACS Appl. Mater. Interfaces 2025, 17, 2, 2756. [Google Scholar] [CrossRef] [PubMed]
- Rasool, M.; Malik, A.; Waquar, S.; Arooj, M.; Zahid, S.; et al. New challenges in the use of nanomedicine in cancer therapy. Bioengineered. 2022, 13, 759. [Google Scholar] [CrossRef]
- Asadi, M.; Ghorbani, S.H.; Mahdavian, L.; Aghamohammadi, M. Graphene-based hybrid composites for cancer diagnostic and therapy. J. Transl. Med. 2024, 22, 611. [Google Scholar] [CrossRef]
- Boddu, A.; Obireddy, S.R.; Zhang, D.; Krishna Rao, K.S.V.; Lai, W.-F. ROS-generating, pH-responsive and highly tunable reduced graphene oxide-embedded microbeads showing intrinsic anticancer properties and multi-drug co-delivery capacity for combination cancer therapy. Drug Deliv. 2022, 29, 2481. [Google Scholar] [CrossRef]
- Khakpour, E.; Salehi, S.; Naghib, S.M.; Ghorbanzadeh, S.; Zhang, W. Graphene-based nanomaterials for stimuli-sensitive controlled delivery of therapeutic molecules. Front Bioeng Biotechnol. 2023, 11, 1129768. [Google Scholar] [CrossRef]
- Semenov, K.N.; Shemchuk, O.S.; Ageev, S.V.; Andoskin, P.A.; Iurev, G.O.; et al. Development of Graphene-Based Materials with the Targeted Action for Cancer Theranostics. Biochemistry Moscow. 2024, 89, 1362. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Mukherjee, A.; Bytesnikova, Z.; Ashrafi, A.M.; Richtera, L.; et al. 2D graphene-based advanced nanoarchitectonics for electrochemical biosensors: Applications in cancer biomarker detection. Biosens. Bioelectron. 2024, 250, 116050. [Google Scholar] [CrossRef]
- Iannazzo, D.; Espro, C.; Celesti, C.; Ferlazzo, A.; Neri, G. Smart Biosensors for Cancer Diagnosis Based on Graphene Quantum Dots. Cancers 2021, 13, 3194. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, M.S.; Sangrizeh, F.H.; Jahani, N.; Abedin, M.S.; Chaleshgari, S.; et al. Graphene oxide nanoarchitectures in cancer therapy: Drug and gene delivery, phototherapy, immunotherapy, and vaccine development. Environ. Res. 2023, 237, 117027. [Google Scholar] [CrossRef]
- Fucikova, J.; Keep, O.; Kasikova, L.; Petroni, G.; Yamazaki, T.; et al. Detection of immunogenic cell death and its relevance for cancer therapy. Cell Death Dis 2020, 11, 1013. [Google Scholar] [CrossRef]
- Yunus, M.A; Ramli, M.M; Osman, N.H.; Mohamed, R. Stimulation of Innate and Adaptive Immune Cells with Graphene Oxide and Reduced Graphene Oxide Affect Cancer Progression. Arch. Immunol. Ther. Exp. 2021, 69, 20. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Hong, H.; Lee, Y.; Park, K.S.; Sun, M.; et al. Efficient Lymph Node-Targeted Delivery of Personalized Cancer Vaccines with Reactive Oxygen Species-Inducing Reduced Graphene Oxide Nanosheets. ACS Nano 2020, 14, 10–13268. [Google Scholar] [CrossRef]
- Yue, H.; Wei, W.; Gu, Z.; Ni, D.; Luo, N.; et al. Exploration of graphene oxide as an intelligent platform for cancer vaccines. Nanoscale, 2015, 7, 19949. [Google Scholar] [CrossRef]
- Jomova, K.; Alomar, S.Y.; Alwasel, S.H.; Nepovimova, E.; Kuca, K.; et al. Several lines of antioxidant defense against oxidative stress: antioxidant enzymes, nanomaterials with multiple enzyme-mimicking activities, and low-molecular-weight antioxidants. Arch Toxicol 2024, 98, 1323. [Google Scholar] [CrossRef]
- Ristic, B.; Harhaji-Trajkovic, L.; Bosnjak, M.; Dakic, I.; Mijatovic, S.; et al. Modulation of Cancer Cell Autophagic Responses by Graphene-Based Nanomaterials: Molecular Mechanisms and Therapeutic Implications. Cancers 2021, 13, 4145. [Google Scholar] [CrossRef] [PubMed]
- Sandbhor, P.; Palkar, P.; Bhat, S.; John, G.; Goda, J.S. Nanomedicine as a multimodal therapeutic paradigm against cancer: on the way forward in advancing precision therapy. Nanoscale, 2024, 16, 6330. [Google Scholar] [CrossRef] [PubMed]
- Tabish, T.A.; Narayan, R.J. Mitochondria-targeted graphene for advanced cancer therapeutics. Acta Biomaterial 2021, 129, 43. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Representation of the applications of GBMs in TME reprogramming through mitochondria targeting. (Created with the assistance of BioRender.com “https://app.biorender.com/user/signin?illustrationId=6156d45891063d00af8af51d (accessed on 18/03/2025 up to 23/03/2025) and Microsoft ppt).
Figure 1.
Representation of the applications of GBMs in TME reprogramming through mitochondria targeting. (Created with the assistance of BioRender.com “https://app.biorender.com/user/signin?illustrationId=6156d45891063d00af8af51d (accessed on 18/03/2025 up to 23/03/2025) and Microsoft ppt).
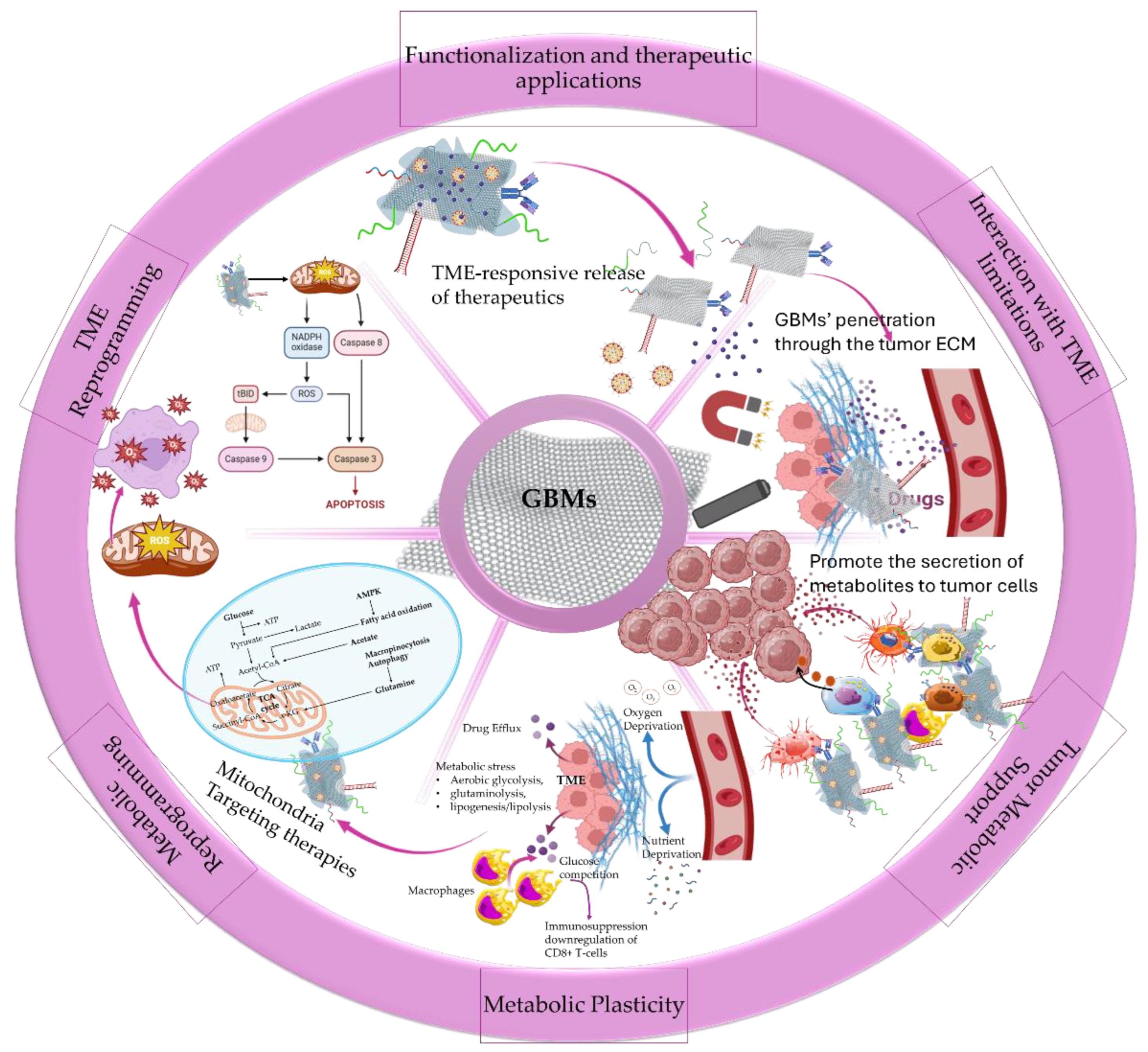
Figure 2.
Schematic representation of the application of GBMs nanocomposites in the metabolic reprogramming in TME and the advantages of nanomedicine in targeting tumor metabolism. (Created with the assistance of BioRender.com “https://app.biorender.com/user/signin?illustrationId=6156d45891063d00af8af51d (accessed on 18/03/2025 up to 23/03/2025) and Microsoft ppt).
Figure 2.
Schematic representation of the application of GBMs nanocomposites in the metabolic reprogramming in TME and the advantages of nanomedicine in targeting tumor metabolism. (Created with the assistance of BioRender.com “https://app.biorender.com/user/signin?illustrationId=6156d45891063d00af8af51d (accessed on 18/03/2025 up to 23/03/2025) and Microsoft ppt).
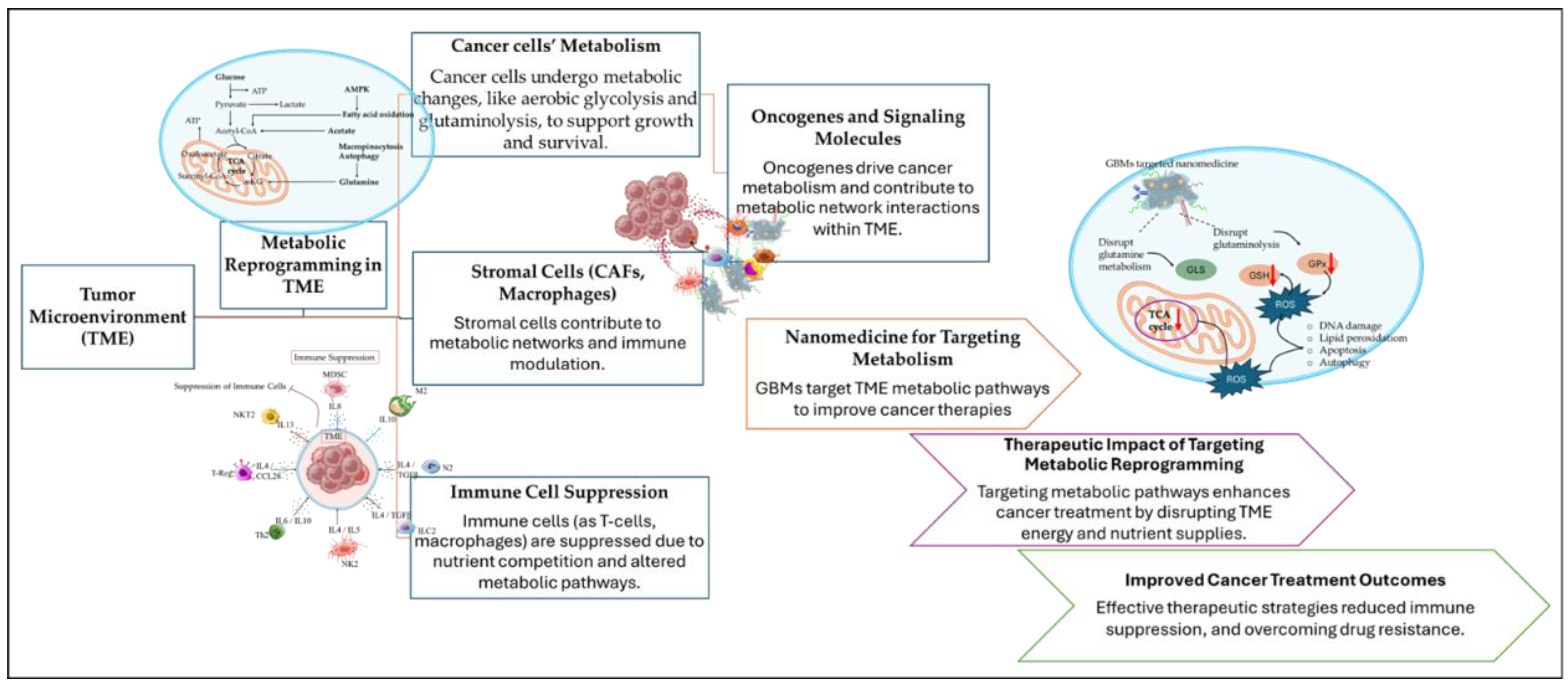
Figure 3.
Schematic representation of the effect of Gr and GQDs nanocomposites on the antioxidant defense of mitochondria to promote ROS imbalance and oxidative stress triggered cell death. (Created with the assistance of BioRender.com “https://app.biorender.com/user/signin?illustrationId=6156d45891063d00af8af51d (accessed on 18/03/2025 up to 23/03/2025 and Microsoft ppt).
Figure 3.
Schematic representation of the effect of Gr and GQDs nanocomposites on the antioxidant defense of mitochondria to promote ROS imbalance and oxidative stress triggered cell death. (Created with the assistance of BioRender.com “https://app.biorender.com/user/signin?illustrationId=6156d45891063d00af8af51d (accessed on 18/03/2025 up to 23/03/2025 and Microsoft ppt).
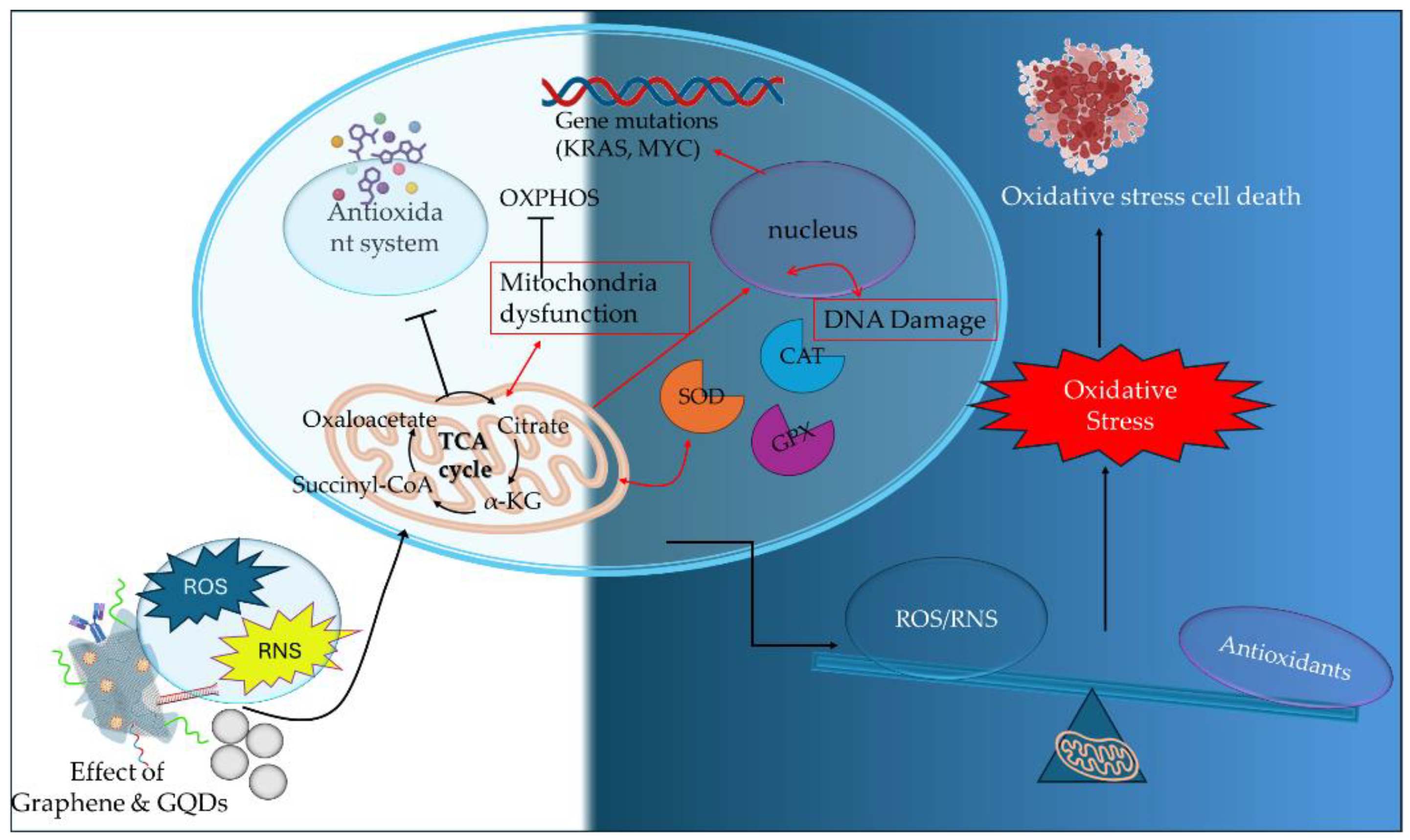
Figure 4.
Visual representation of the role of GO nanocomposites in targeting cancer cells and mitochondria for enhanced drug delivery and effective stimulation of autophagy and apoptotic cell death through ROS production. Created with the support of BioRender.com “https://app.biorender.com/user/signin?illustrationId=6156d45891063d00af8af51d (accessed on 18/03/2025 up to 23/03/2025) and Microsoft ppt).
Figure 4.
Visual representation of the role of GO nanocomposites in targeting cancer cells and mitochondria for enhanced drug delivery and effective stimulation of autophagy and apoptotic cell death through ROS production. Created with the support of BioRender.com “https://app.biorender.com/user/signin?illustrationId=6156d45891063d00af8af51d (accessed on 18/03/2025 up to 23/03/2025) and Microsoft ppt).
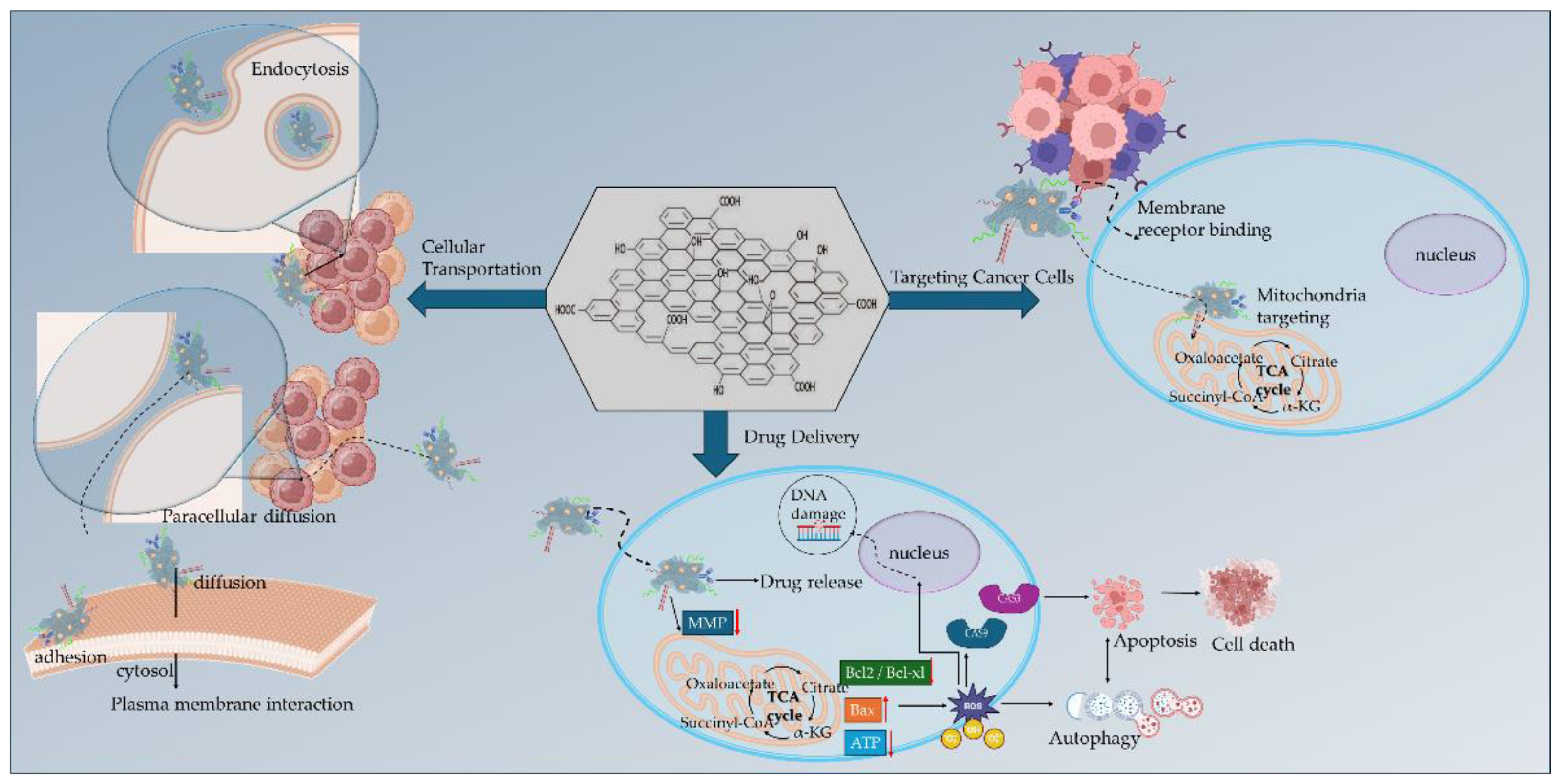
Figure 5.
Schematic of the role of rGO nanocomposites in targeting cancer cells and mitochondria for enhanced drug delivery to promote apoptotic cell death and synergistic effect with AgNPs to further promote oxidative stress and trigger cancer cell death. Created with the support of BioRender.com “https://app.biorender.com/user/signin?illustrationId=6156d45891063d00af8af51d (accessed on 18/03/2025 up to 23/03/2025) and Microsoft ppt).
Figure 5.
Schematic of the role of rGO nanocomposites in targeting cancer cells and mitochondria for enhanced drug delivery to promote apoptotic cell death and synergistic effect with AgNPs to further promote oxidative stress and trigger cancer cell death. Created with the support of BioRender.com “https://app.biorender.com/user/signin?illustrationId=6156d45891063d00af8af51d (accessed on 18/03/2025 up to 23/03/2025) and Microsoft ppt).
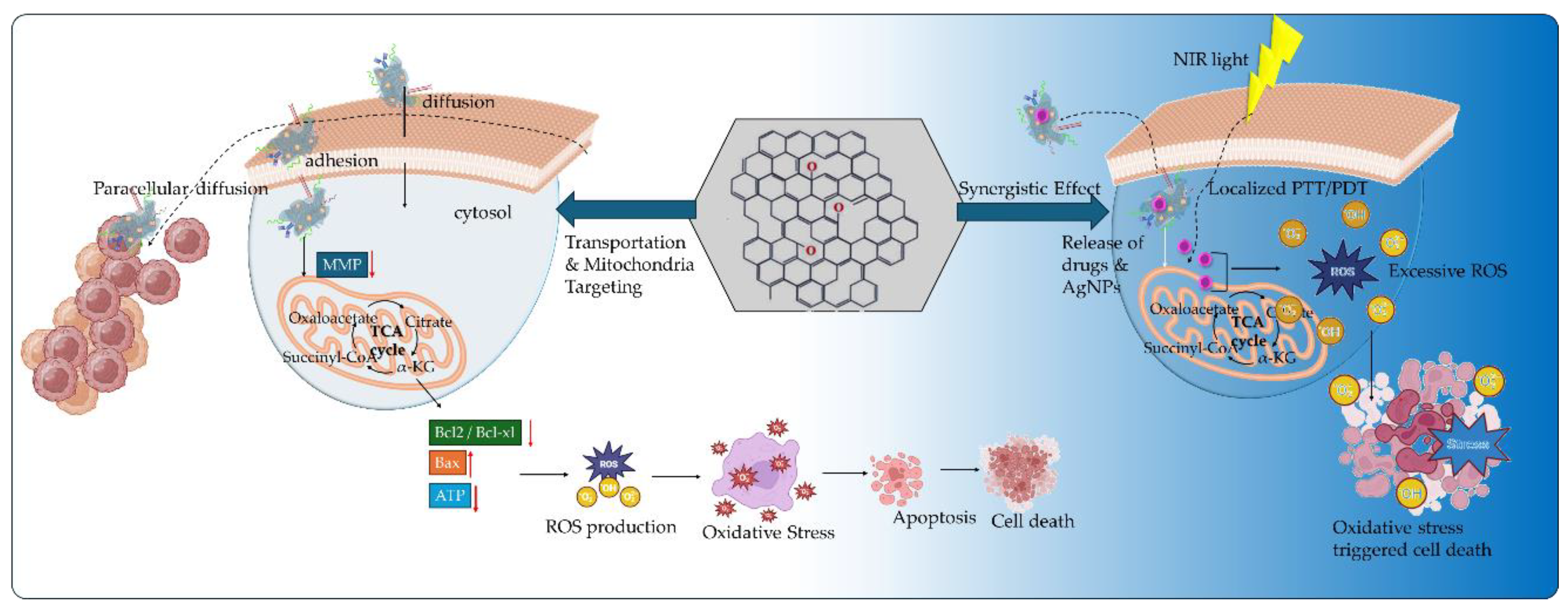
Figure 6.
Schematic illustration of the future directions on GBMs.
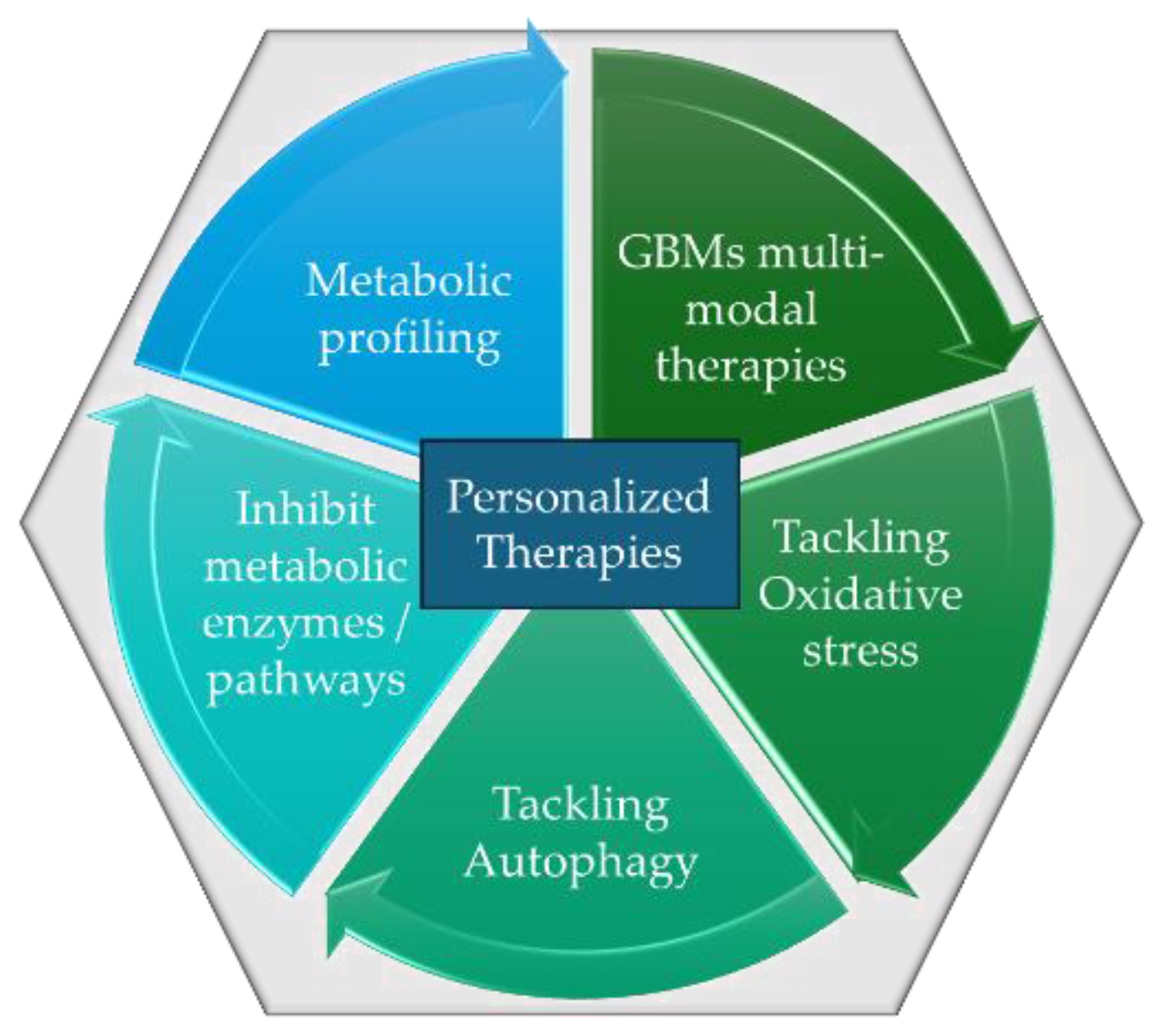
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



