
Submitted:
31 March 2025
Posted:
01 April 2025
You are already at the latest version
Abstract
Objective: Lung cancer remains the leading cause of cancer-related mortality worldwide, with significant resistance to conventional therapies, highlighting the urgent need for novel therapeutic strategies. Moringa oleifera (M. oleifera), a medicinal plant rich in diverse bioactive compounds, has shown promising potential for anti-lung carcinoma activity. This study investigates the molecular mechanisms underlying the therapeutic effects of M. oleifera bioactive compounds through an integrated systems biology and molecular docking approach. By constructing comprehensive compound-target-pathway networks, we aim to elucidate the multitarget pharmacology of M. oleifera compounds, providing valuable insights into their potential as therapeutic candidates.
Methods: Bioinformatics tools were used to identify 180 phytochemicals from M. oleifera, filtered using Lipinski’s Rule of Five and ADMET properties, yielding 14 lead compounds. Protein-protein interaction (PPI) analysis identified 89 overlapping lung cancer targets, with EGFR being the most enriched in pathway enrichment analysis.
Results: In the analysis, Caffeic acid showed the highest binding affinity (-28.97 kcal/mol) with EGFR through molecular docking and maintained stability during molecular dynamics simulations. This interaction also modulates 12 pathways critical to lung cancer, including MAPK, JAK-STAT, and PI3K/AKT pathways. The overall result suggests that Caffeic acid is an EGFR-mediated oncogenic signalling inhibitor.
Conclusion: Caffeic acid is a potential candidate for lung cancer therapy, warranting further experimental validation to translate these findings into clinical applications.
Keywords:
Lung cancer
; Moringa oleifera
; Network System Biology
; Molecular docking
; Molecular dynamics
1. Introduction
Lung cancer poses a substantial global public health challenge, representing the foremost cause of cancer-related fatalities worldwide, as reported by the World Health Organization (WHO). In 2022 alone, lung cancer was accountable for an estimated 1.8 million deaths, underscoring its status as the primary contributor to cancer mortality globally (source: https://gco.iarc.fr/). The incidence of lung cancer exhibits regional disparities, with North America, Europe, and Asia reporting the highest rates [1]. Tobacco smoking is the primary contributor to lung cancer, accounting for roughly 85% of all case. However, other factors such as exposure to environmental pollutants, genetic susceptibility, and other risk factors also contribute. Despite strides in treatment modalities, lung cancer prognosis remains poor, with a 5-year survival rate of less than 20% [2]. Lung cancer treatment approaches may differ based on cancer type and stage due to its complexity. Currently, lung cancer treatment options include surgery, radiation therapy, chemotherapy, targeted therapy, and immunotherapy. Chemotherapy, which employs drugs to eliminate cancer cells, is commonly administered to patients with advanced-stage lung cancer. Targeted therapy designed to pinpoint specific genes crucial for cancer cell proliferation has emerged as a promising approach. Various targeted therapies have demonstrated encouraging outcomes in the treatment of diverse forms of lung cancer [3,4]. Despite advancements in lung cancer treatment techniques, the issue of drug resistance and the growing number of lung cancer cases in the future highlights the pressing need for the development of new medications to combat this life-threatening disease [5]. Although existing treatment modalities have made progress, their efficacy varies among patients, and the emergence of drug resistance can compromise their effectiveness. Hence, the ongoing advancement of new and potent drugs for lung cancer treatment is imperative to address the escalating incidence of this condition. Natural compounds sourced from plants are increasingly recognized as a promising avenue for discovering novel alternative anticancer drugs owing to their enhanced structural diversity, low toxicity upon metabolism in the body, and complex nature. Secondary metabolites derived from plants are particularly important primary reservoirs of medicinal agents [6,7,8,9,10,11]. M. oleifera commonly referred as the drumstick tree and a staple vegetable in numerous countries since ancient times, originates from South Asia, primarily the foothills of the Himalayas, India, although it has also been cultivated and naturalized elsewhere. This tree boasts remarkable nutritional and medicinal properties [12,13,14]. M. oleifera has been traditionally employed to treat a myriad of ailments, including arthritis, asthma, hypertension, pain, coughs, diabetes, diarrhoea, dropsy, epilepsy, fever, headaches, hysteria, irritations, paralysis, skin infections, sores, tumors, weakness, wounds, among others, across various countries [15,16,17]. The bioactive constituents present in almost all parts of M. oleifera have been associated with their efficacy against various illnesses, including cancer [18]. Recent studies have revealed that natural compounds from M. oleifera can exert anticancer effects [19,20]. M. oleifera extracts offer substantial health benefits, particularly through alcoholic and aqueous extraction of leaf compounds emerging as the optimal source. Their effects vary with dosage, displaying cytotoxicity towards cancer cell lines while preserving normal cell lines [21,22]. M. oleifera roots have demonstrated inhibitory activity against various cancer cell lines by reducing cell proliferation and inducing apoptosis [23]. Leaf extracts have been shown to significantly induce apoptosis and reduce cell proliferation in a lung cancer cell line [24]. Studies on the alkaloid extract of M. oleifera indicate that inhibition of JAK2/STAT3 pathway activation is the mechanism by which the extract suppresses cell migration and proliferation, induces apoptosis, and halts cell cycle progression in lung cancer [25]. This study pioneered an innovative approach employing a network system biology-based strategy to identify active natural compounds from M. oleifera with potential therapeutic applications in lung cancer treatment. Molecular docking and simulation techniques were used to assess the anti-lung cancer properties of these M. oleifera active compounds, initiating the exploration of promising drug candidates derived from this plant. From a broader perspective, network system biology enables the examination of the intricate interplay between identified phytochemicals and various targets within the pathways associated with lung cancer [26,27,28]. Molecular docking was employed to elucidate the binding patterns of selected phytochemicals from M. oleifera to molecular targets linked to lung cancer. These analyses shed light on how these phytochemicals may influence cancer-related biological processes. Nevertheless, further validation of these findings through in vitro and in vivo studies is required. The detailed workflow of our study is shown in Figure 1.
2. Material and Methods
2.1. Data Mining of M. oleifera Natural Constituents
The phytochemical data of M. oleifera were obtained from publicly available databases such as Indian Medicinal Plants, Phytochemistry and Therapeutics 2.0 (IMPPAT 2.0) (https://cb.imsc.res.in/imppat/), Dr. Duke’s Phytochemical and Ethnobotanical Databases (https://phytochem.nal.usda.gov/), and Phytochemical Interactions DB (PCIDB) (https://www.genome.jp/db/pcidb/). The search outcomes were consolidated, and duplicate entries were eliminated.
2.2. 3D Structure Retrieval and Molecular Property Identification
The 3D and 2D structures of the compounds were obtained from PubChem and prepared using Discovery Studio 2022 (DS2022) software. The compound dataset was optimized using the CHARMm-based smart minimizer, which performs 1500 steps of steepest descent followed by conjugate gradient algorithms, achieving a convergence gradient of 0.001 kcal/mol [29]. The retrieved natural compounds were screened for drug-likeness based on Lipinski’s Rule of Five [30]. This rule is a guideline in drug discovery for evaluating the potential of compounds as effective orally administered drugs. Subsequently, we assessed the absorption, distribution, metabolism, excretion, and toxicity (ADMET) properties, which are critical for drug discovery and development, using the ADMET protocol in DS2022. These standard descriptors used in pharmaceutical characterization and drug selection indicate suitability for human administration [31].
2.3. Retrieval of Targets Related to Phytochemicals and Lung Cancer
The study utilized the Swiss Target Prediction to identify the target genes of the phytochemicals present in the M. oleifera plant (http://www.swisstargetprediction.ch/). Swiss-Target Prediction is a web-based tool for predicting the targets of bioactive molecules [32]. The selection criteria for genes contributing to lung cancer included a two-parameter threshold: a Gene-Disease Association (GDA) score of ≤ 0.1 from DisGeNET (https://www.disgenet.org/) [33] and Simple Somatic Mutation (SSM) affected cases with a mutation frequency of ≤ 1.06% from TCGA (https://portal.gdc.cancer.gov/). These parameters were used to identify genes for further analysis, and the target genes identified from these databases were combined to remove duplicates and to obtain a refined list of lung cancer genes. The final overlapping target genes between M. oleifera and lung cancer were identified and visualized using Venny 2.1(http://www.interactivenn.net/).
2.4. Protein-Protein Interaction (PPI) Network and Core Targets Identification
PPI networks are effective tools for discovering biological interactions among proteins. First, the intersected targets were considered potential therapeutic targets of the natural compounds of M. oleifera against Lung Cancer. Using the STRING database [34], the protein-protein interactions (PPI) network of shared genes was obtained. This database provides both experimental and predicted relationship information for network contraction. We investigated the functional associations between these targets using the STRING database. A high confidence threshold (>0.70) was established for the minimum interaction score. Network analysis visualization was conducted using Cytoscape software (version 3.9.0). In the PPI network, numerous nodes and edges represent proteins and their interactions. Nodes with a higher number of significant connections to other nodes were considered top-ranked. Three topological analyses (Degree, Closeness, and Betweenness) were employed in the PPI network to identify core targets [35].
2.5. Pathway Enrichment Analysis
Gene enrichment analysis was performed using g: profiler (https://biit.cs.ut.ee/gprofiler/gost) to investigate the Kyoto Encyclopedia of Genes and Genomes (KEGG) to identify lung cancer-associated pathways. From the PPI and network analysis, identified the core proteins that were uploaded to the g: profiler tool using Homo sapiens as the species. We considered those pathways that played crucial roles in lung cancer and were selected from the KEGG pathways using default cutoff values (P<0.05) that were considered statistically significant [36].
2.6. Compound-Targets–Pathways Network Construction
After enrichment analysis, the compound-target-pathway interaction network was constructed using Cytoscape 3.9.0. This network contained compounds, proteins, and pathways related to lung cancer enriched in KEGG pathway analysis and showed the hub proteins that contributed to pathway enrichment [37].
2.7. Molecular Docking
Based on the Compound–Targets-Pathways interaction network analysis, we identified the top interacting proteins with different pathways, and after that, we also selected the top interacting compounds with proteins. The top proteins and compounds were subjected to molecular docking using DS2022. We extracted the 3D structure of the EGFR (epidermal growth factor receptor) protein using the RCSB PDB (https://www.rcsb.org/) (PDB ID: 1M17), and the ‘Prepare Protein’ protocol was used to prepare the EGFR protein with DS2022. The active site was selected using docked ligand into the active cavity of EGFR by creating a sphere of 5Å radius using “Define and Editing” Protocol of DS2022 [38]. For molecular docking, the ‘CDOCKER’ protocol was utilized to extract the CDOCKER energy and interaction pattern of the compounds involved in docking [39].
2.8. Molecular Dynamics Simulation
A molecular dynamics simulation was conducted to analyze the stability of the protein-ligand complex. The best-docked complex was subjected to molecular dynamics (MD) simulation. The MD simulation conducted in an implicit solvent environment to study the stability, conformational changes, and dynamic behavior of the inhibitor within the binding cavity using the DS2022 protocol ‘Standard Dynamics Cascade’. [40]. The steepest descent minimization algorithm was used with a maximum of 5000 steps, to minimized the energy of docked complex. Position restraints were applied to the receptor and ligand of each system for 5000 steps throughout heating (300 K) using NVT (No. of atoms, Volume, Temperature) and a production run of 50ns. For the analysis of the MD simulation extracted, the trajectory files to explore the root mean square deviation (RMSD), root mean square fluctuation (RMSF), and radius of gyration (Rg).
3. Results and Discussion
3.1. Screening of Active Components of M. oleifera
The phytochemical compounds of M. oleifera were retrieved from these databases (Indian Medicinal Plants, Phytochemistry and Therapeutics 2.0 (IMPPAT 2.0) [41], Dr Duke’s Phytochemical and Ethnobotanical Databases [42], and Phyto-Chemical Interactions DB [43]. The data retrieved from these databases were filtered after removing duplicate entries a total of 180 natural compounds were selected for further analysis. All the selected compounds were again subjected to Lipinski’s rule of 5 for drug-likeness. 167 natural compounds that fulfilled the cut-off criteria with the following rules: molar refractivity value of 40–130, Log P less than 5, number of hydrogen bond acceptors less than 10, number of hydrogen bond donors less than 10, and molecular mass less than 500 Da, as shown in Table S1. The compounds filtered from the Lipinski’s rule of 5. All (167 natural compounds) were subjected to another filtering process based on ADMET-based properties. In ADMET, we considered different properties for the filtration of compounds, such as absorption, water solubility, CYP2D6, Hepatotoxicity, and intestinal absorption. After the ADMET-based filtration, 14 natural compounds were identified as shown in Figure 2 and Table S2. Water solubility is a key factor in the drug bioavailability process; water solubility is categorized as the best if it is within the range of values less than 3 or 4. All selected compounds exhibited water solubility within the specified range. Concerning the next parameter, the intestinal absorption of all the compounds was 0, which is a good range, and shows that the intestine easily absorbs all the compounds and passes through the blood stream. The compounds showed no inhibition (false) of CYP2D6, which is an enzyme that plays a role in the digestive system and metabolic processes. Hepatotoxicity predictions revealed that all the predicted compounds were non-toxic.
3.2. The Potential Targets for Natural Compounds of M. oleifera and Lung Cancer
The Swiss Target Prediction database identified 508 unique genes targeted for the 14 natural compounds of M. oleifera obtained from the ADMET analysis (Table S3). Additionally, genes related to lung cancer were retrieved from DisGeNET (380) and TCGA (492) databases, which were given in Tables S4 and S5, respectively. After removing duplicates, 819 genes were considered from both databases (DisGeNET and TCGA). Using Venny to find the intersection of 508 genes from M. oleifera related targets and 819 lung cancer-related targets, a total of 89 overlapping targets were obtained, as shown in Figure 3, and provided in Table S6.
3.3. Protein-Protein Interaction and Network Analysis
A set of 89 common proteins was subjected to the STRING database with a confidence score cutoff of 0.90, and a PPI network was generated and visualized, as shown in Figure 4(i). The PPI network included 80 nodes(proteins), 314 edges, and 9 nodes that did not contribute to PPI interactions based on the parameters for the PPI network and were excluded from further analysis.
These nodes represent target proteins, and the edges indicate interactions between the proteins. The finalized PPI network was exported to Cytoscape v3.10 for further analysis. The Cytoscape Network Analyzer tool was used to evaluate the topological parameters of proteins within the network, includes protein degree, betweenness centrality (BC), and closeness centrality (CC). The degree represents the number of direct connections with the nodes in the network. Similarly, BC quantifies how often a node lies on the shortest paths between other nodes, and CC evaluates the importance of a node based on its efficiency in interacting with other nodes. The Hub proteins in the PPI network were identified using median cutoff criteria: a degree value of ≤6, a CC value of ≤0.0295, and a BC value of ≤0.4093. For more detail on the median-based cutoff criteria, we used the methodologies of Jie et al. and Ying et al. to identify significant proteins for our analysis [44,45]. Using all these parameters, selected 11 key proteins, namely STAT3, ESR1, EGFR, MAPK8, IGF1R, CTNNB1, CREBBP, HSP90AA1, HRAS, JAK2, and EP300 were identified as hub proteins, and the complete selection process were shown in the Figure 4. (ii-iv), and the information is given in Table S7.
3.4. KEGG Pathway Enrichment Analysis
A total of 56 KEGG pathways were enriched using 11 hub proteins obtained from PPI interactions and network analysis, of which 14 pathways were associated with lung cancers. A list of 11 proteins involved in the enrichment of these 14 pathways is presented in Figure 5. These proteins enrich significant pathways that play an essential role in lung cancer progression. JAK-STAT (Janus kinase-signal transducer and activator of transcription) signalling pathways are critical communication routes cells use to transmit extracellular signals to the nucleus. The gene participates in cell proliferation, differentiation, and apoptosis in lung cancer and can be used as a target for treating lung cancer [46]. The FoxO (Forkhead box O) signaling pathway is involved in various cellular processes, including apoptosis, cell cycle regulation, oxidative stress resistance, and metabolism. In lung cancer, deregulation of this pathway can contribute to the development and progression of cancer [47]. The mitogen-activated protein kinase (MAPK) signaling pathway, often due to mutations in upstream receptors like EGFR or in downstream components like KRAS, leads to continuous activation of MAPK signaling. This persistent activation drives uncontrolled cell division and tumor growth and enhances metastatic potential [48]. Ras signaling pathway stimulates angiogenesis by Ras proteins, which signal through the Ras/Raf/MAPK regulate various cellular functions. Deregulation of this pathway is a common event in cancer, as Ras is the most frequently mutated oncogene in lung cancer [49]. The GnRH (Gonadotropin-Releasing Hormone) signaling pathway, primarily known for its role in reproductive hormone regulation, has been implicated in lung cancer development through its influence on cellular proliferation and apoptosis [50]. The PI3K/AKT signaling pathway plays a significant role in the development and progression of lung cancer. It also plays a role in the tumor environment, such as in angiogenesis and inflammatory factor recruitment [51]. ErbB signaling pathway (erythroblastic oncogene B), often through mutations and overexpression, leads to the stimulation of downstream signaling cascades like the MAPK/ERK, PI3K/AKT and JAK/STAT pathways. In turn, they collectively contribute to uncontrolled cell growth, inhibition of apoptosis, angiogenesis, and metastasis in lung cancer [52]. The HIF-1 signaling pathway plays a crucial role in lung cancer progression by acting as a master regulator of cellular responses to hypoxia, enabling low oxygen conditions within the tumor microenvironment, promoting tumor growth, angiogenesis, invasion, and metastasis through the activation of various downstream genes [53]. The Rap1 signaling pathway impacts lung cancer progression by regulating cell adhesion, migration, and invasion, mainly through integrin-mediated interactions [54]. In lung cancer, PD-L1 expression helps tumors evade the immune system by activating the PD-1 checkpoint, effectively "putting the brakes" on immune responses and allowing cancer cells to grow unchecked [55]. Th17 cell differentiation pathways play a significant role in lung cancer progression by promoting tumor growth, angiogenesis, and inflammation [56]. The Wnt signaling pathway plays a significant role in lung cancer development and progression by promoting tumor cell proliferation, invasion and resistance to therapy [57]. Pathways in cancer and non-small-cell lung cancer pathways were also enriched by these proteins, which are important pathways for enhancing lung cancer.
From the protein-pathways interaction the most prominent protein identified is EGFR, which significantly impacts disease pathways. It is a key player, interacting with 12 pathways out of 14 pathways which is more than any other proteins, as indicated in Table 1. Inhibiting EGFR can significantly disrupt key pathways involved in lung cancer, including MAPK, JAK-STAT, PI3K/AKT, and HIF-1 signaling.
3.5. Compounds-Targets–Pathways Network Construction
We developed a Compounds-Targets–Pathways network comprising 14 natural compounds, of which two Furfural and Benzylamine did not form any interactions within the network. Then the network included 12 natural compounds, 11 target proteins and 14 lung cancer-related pathways. Our analysis revealed the interaction between the natural compounds and the target proteins, with some compounds demonstrating more extensive interactions than others. Notably, Gibberellin A29 and Caffeic Acid were the most highly connected compounds, interacting with five proteins (EGFR, MAPK8, CTNNB1, IGF1R, CREBBP) and four proteins (EGFR, EP300, STAT3, ESRI), respectively as shown in Figure 6. For further analysis, we focused on Gibberellin A29 and Caffeic Acid due to their association with the highest number of proteins. Additionally, we selected EGFR, as it plays a pivotal role in regulating the largest number of pathways.
3.6. Molecular Docking
To validate our findings from the compounds-proteins-pathways interactions, we conducted molecular docking to assess the interaction between the natural compounds (Gibberellin A29 and Caffeic Acid) with EGFR. Using the CDOCKER protocol in DS2022, we analysed the docking interaction. The docking complexe of Gibberellin A29 had CDOCKER energy of 46.3641 kcal/mol, this positive value indicates a week interaction. Based on the energy parameter, dropping this target from the study. In contrast, Caffeic Acid exhibited a higher negative CDOCKER energy -28.972 kcal/mol, suggesting a strong interaction with EGFR. EGFR-Caffeic Acid complex formed seven hydrogen bonds and one hydrophobic bond with active site residues. Caffeic acid established conventional hydrogen bonds with LYS721, MET769, GLU738, GLN767, LEU768, and THR830, and a hydrophobic interaction with LYS721. Caffeic acid forms conventional hydrogen bonds with LYS721, MET769, GLU738, GLN767, LEU768, and THR830, along with a hydrophobic interaction with LYS721, as detailed in Table S8. Notably, MET769, a key residue involved in EGFR inhibition, interacts with Caffeic acid, while LYS721 forms a hydrogen bond crucial for competing with ATP and blocking kinase activity. Additionally, GLU738 helps stabilize the inactive conformation of EGFR, preventing the kinase from transitioning to its active state and thereby inhibiting its binding capability [38,58,59,60,61]. To cross-validate the docking experiments, the CDOCKER energy of Caffeic acid was compared with Erlotinib, which was a known ligand of EGFR. Erlotinib exhibited a CDOCKER energy of -22.641 kcal/mol, whereas Caffeic acid had a higher CDOCKER energy of -28.972 kcal/mol. This energy difference indicates a stronger binding affinity for Caffeic acid compared to Erlotinib. This comparative analysis was performed using the CDOCKER protocol of DS2022. This comparative analysis was performed using the CDOCKER protocol of DS2022, CDOCKER protocol of DS2022. The interactions of EGFR with Caffeic acid and Erlotinib are shown in Figure 7, and residue interaction (bond formed) was provided in S8.
3.7. Molecular Dynamics Simulation
To evaluate the flexibility and overall stability of the docked complex, we performed 50ns time-dependent MD simulation using DS2022, analysing the results through RMSD (Root Mean Square Deviation), RMSF (Root Mean Square Fluctuation), and Radius of Gyration (Rg). The Figure 8(i) demonstrates that after molecular dynamics (MD) simulation, key interacting residues remain in place, contributing to the inhibition of the EGFR. Specifically, LYS721 forms a hydrophobic bond that stabilizes the interaction. Additionally, LEU768 and MET769 establish hydrogen bonds, as observed in the earlier molecular docking phase [38,39]. Notably, new hydrogen bonds also form with THR766, which is crucial for protein stabilization [62] and ASP831,which were not observed in the initial docking. The retention of these interactions, along with the formation of new stabilizing bonds, underscores the compound potential inhibitory effectiveness and stability in binding to the protein. During the MD simulation, five bonds were formed, three of which were retained while two were newly established. Meanwhile, four bonds Glu738, Met742, Gln767, and Thr830 initially observed in molecular docking, were lost. Detail information of bonds formed post MD stimulation is given in Table S9 and the table contained comparative information of bonds formed and loss during the docking and simulation in Table S10. The structural deviation of the EGFR in a complex with Caffeic acid was assessed using RMSD to evaluate the stability of the complex throughout the production run. Initially, the RMSD values of the EGFR in a complex with Caffeic acid increase rapidly at the beginning of the simulation. Initial conformation shows the RMSD values fluctuating within a range of 1.6 to 2.0 Å up to 10 nanoseconds, after which the complex achieved convergence, as illustrated in Figure 8 (ii). The RMSF values indicate the flexibility of residues, but we focused on the active site residues of EGFR which forms the bonds in simulations, the residues which play a significant role in the stability of the docked compound shows minimal fluctuation. Specifically, residues MET769, LYS721, THR766, LEU768, and ASP831 exhibited minimal fluctuations within a narrow range of 0.2 to 0.3 Å This suggest that these residues remain stabilized within the active site cavity of EGFR, as shown in Figure 8 (iii). The low fluctuation values of these active site residues further indicate a favorable and stable interaction [63,64] with the Caffeic acid. The radius of gyration (Rg) values was used to evaluate the compactness of the protein during MD simulation. Initially, the Rg value was 22.12 Å, which subsequently decreased to 22.04 Å, as shown in Figure 8(iv). Although this reduction in compactness was small (0.08 Å), but it indicates that EGFR became slightly more compact upon Caffeic acid binding and achieved a stable conformation. Together, these analyses demonstrate that EGFR attained structural stability in the presence of the Caffeic acid, which may support its enhanced inhibitory potential.
4. Discussion:
Lung cancer is the leading cause of cancer-related deaths worldwide, claiming the highest mortality rates among all. Only about 20% of patients survive beyond five years after diagnosis. While significant progress has been made in understanding its genetic evolution, improving diagnostic techniques, and developing advanced treatments such as surgery, radiotherapy, chemotherapy, immunotherapy, and targeted therapies, the overall prognosis remains low [62,65]. Nature has provided medicinal ingredients for centuries, with many modern treatments rooted in natural sources [66]. For example, Morphine, derived from Papaver somniferum is used as an analgesic [67], while Taxol from Taxus brevifolia and Vinblastine and Vincristine from Catharanthus roseus are widely used as anticancer drugs [68,69]. Natural compounds are often favoured for their lower toxicity and reduced side effects compared to synthetic drugs. For instance, natural compound Galangin, which inhibits gastric cancer, is less toxic than the commonly used chemotherapy drug Fluorouracil [70]. Similarly, Luteolin is an excellent natural active compound with minimal toxicity compared to synthetic drugs used for liver disease treatment [71]. In the present study, we employed network system biology approach to investigate the mechanisms underlying the therapeutic effects of M. oleifera in treating lung cancer. This approach enabled a systematic evaluation by identifying key bioactive compounds from M. oleifera and their potential targets in lung cancer [72]. The multi-component nature of M. oleifera’s bioactive compound indicates its potential multi-targets to modulate diverse downstream signaling pathways involved in lung cancer progression and metastasis. In our study compounds from M. oleifera were collected from various databases and subsequently evaluated for their drug-likeness and ADMET profiles. Lipinski’s Rule of Five and ADMET analysis are used in drug discovery to evaluate the potential oral bioavailability of a molecule by assessing its physicochemical properties, identify compounds with characteristics likely to be well absorbed, distributed, metabolized, excreted, and have low toxicity within the body, thus improving the chances of a successful drug candidate during early development stage [73]. The core active compound in our study was Caffeic acid, is a polyphenol known for its various positive effects on human health, including potential anticancer activity. Caffeic acid has been reported to reduce the viability of human non-small-cell lung cancer cells and inhibit cell proliferation, particularly when used in combination with paclitaxel [74,75]. Beyond its anticancer properties, Caffeic acid exhibits a range of biological and pharmacological activities, including antioxidant [76], anti-inflammatory [77], and neuroprotective effects [78]. Caffeic acid has been found to inhibit the PI3K/Akt and AMPK pathways, which play roles in cancer progression [79], and the ERK1/2 pathway, which is involved in cell proliferation and survival [80]. It has also been observed to enhance apoptotic and antiproliferative activities, effectively inhibiting cancer progression [81]. The pathway analysis identified 16 pathways associated with lung cancer, with 14 linked to EGFR, 13 to HRAS, and 9 to STAT3. Further investigation revealed that all 13 HRAS-associated pathways were also present in the EGFR, and 8 out of the 9 STAT3-associated pathways overlapped with EGFR, thus excluding these pathways from subsequent analyses. Given the substantial overlap, we focused our analysis on EGFR, representing the most comprehensive set of common pathways related to lung cancer. EGFR is a crucial target for lung cancer treatment, as various structural aberrations and mutations in EGFR can lead to uncontrolled tumour cell proliferation and survival [82,83]. EGFR is activated through dimerization and subsequent auto-phosphorylation of its tyrosine kinase domain. This phosphorylation triggers downstream pathways, including the MAPK, PI3K/AKT, and STAT signaling pathways, ultimately promoting cell growth and proliferation [84,85]. Molecular docking was employed to investigate the interactions between EGFR and Caffeic acid and to explore the binding affinity. The docking results showed that Caffeic acid forms hydrogen bonds with key residues that play an important role within the active site and enable effective interactions with essential functional groups critical for EGFR activity. Additionally, hydrophobic interactions were observed, further stabilizing the inhibitor within the binding site cavity. These hydrophobic interactions, primarily present in a docked complex, are submitted subject to MD simulations to investigate the stability and binding affinity of protein-ligand interactions, and the study suggests that Caffeic acid could serve as an effective inhibitor of EGFR. Molecular dynamics simulations assessed the EGFR-Caffeic acid complex for stability and fluctuations. Different MD simulation analyses were performed using RMSD, RMSF, and Rg [64,86]. The RMSD value of the EGFR indicates the structural stability of the complex during the simulation, and RMSF explores the residues behaviour during the production run time. The active site residues show minimal fluctuations, which means that the residues interact with the Caffeic acid. They also show minimal fluctuations and structural stability of the EGFR. The Rg explore the compactness of the protein during MD simulation. Our result indicates a decrease in the Rg value, suggesting that the EGFR folding stabilized and achieved compactness. This integrated computational approach underscores the potential of M. oleifera’s natural compound, Caffeic acid, as a therapeutic agent for lung cancer. However, further studies are needed to explore the potential impact of the Caffeic acid in lung cancer treatment. Unlike synthetic EGFR inhibitors, Caffeic acid demonstrates higher binding affinities while maintaining lower toxicity. This aligns with studies reporting natural compounds as adjuncts to conventional therapies, enhancing efficacy while mitigating side effects. The ability of Caffeic acid to modulate multiple pathways suggests its potential as a multitarget therapy, particularly against advanced-stage lung cancer, where single-target drugs often fail due to resistance mechanisms.
5. Conclusion
The research, utilizing system biology and bioinformatics processes, emphasizes the potential efficacy and molecular mechanisms of M. oleifera in treating lung cancer. Our findings provide preliminary evidence that compounds of M. oleifera interact with 89 protein targets, with compound such as Caffeic acid showing results in inhibiting EGFR and modulating critical signaling pathways associated with lung cancer. The experimental approach encompassed both in vitro assays using EGFR-expressing lung cancer cell lines and in vivo validation, providing a comprehensive understanding of their potential therapeutic implications.
Supplementary Materials
The following supporting information is present in the Preprints.org.
Author Contributions
A.S. designed the study, collected and analysed the data, and wrote the manuscript. K.P.S provided expertise in statistical analysis and contributed to the writing and editing of the manuscript. S.G assisted with study design and data collection and contributed to the analysis and interpretation of the results. D.O. provided critical feedback on the manuscript and contributed to the conceptualization of the study. O.W. participated in the study’s design and provided technical expertise. All authors read and approved the final version of the manuscript.
Funding
This research was funded by the German Federal Ministry of Education and Research (BMBF) e: Med-MelAutim 01ZX2205B to S.G. and K.P.S.
Data Availability
The original contributions presented in this study are included in the article/Supplementary Material, and further inquiries can be directed to the corresponding author.
Competing Interests
The authors declare that the research was conducted without any commercial or financial relationships that could be construed as potential conflicts of interest.
References
- Thandra, K.C.; Barsouk, A.; Saginala, K.; Aluru, J.S.; Barsouk, A. Epidemiology of lung cancer. Contemp. Oncol. (Poznan, Poland) 2021, 25, 45–52. [Google Scholar] [CrossRef]
- Bade, B.C.; Dela Cruz, C.S. Lung Cancer 2020: Epidemiology, Etiology, and Prevention. Clin. Chest Med. 2020, 41, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Pirker, R. Conquering lung cancer: current status and prospects for the future. Pulmonology 2020, 26, 283–290. [Google Scholar] [CrossRef]
- Kennedy, K.; Hulbert, A.; Pasquinelli, M.; Feldman, L.E. Impact of CT screening in lung cancer: Scientific evidence and literature review. Semin. Oncol. 2022, 49, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Li, Y.; Xiong, L.; Wang, W.; Wu, M.; Yuan, T.; Yang, W.; Tian, C.; Miao, Z.; Wang, T.; et al. Small molecules in targeted cancer therapy: advances, challenges, and future perspectives. Signal Transduct. Target. Ther. 2021, 6, 1–48. [Google Scholar] [CrossRef]
- Kooti, W.; Servatyari, K.; Behzadifar, M.; Asadi-Samani, M.; Sadeghi, F.; Nouri, B.; Zare Marzouni, H. Effective Medicinal Plant in Cancer Treatment, Part 2: Review Study. J. Evid. Based. Complementary Altern. Med. 2017, 22, 982–995. [Google Scholar] [CrossRef]
- Dehelean, C.A.; Marcovici, I.; Soica, C.; Mioc, M.; Coricovac, D.; Iurciuc, S.; Cretu, O.M.; Pinzaru, I. Plant-Derived Anticancer Compounds as New Perspectives in Drug Discovery and Alternative Therapy. Molecules 2021, 26. [Google Scholar] [CrossRef]
- Siddiqui, A.J.; Jahan, S.; Singh, R.; Saxena, J.; Ashraf, S.A.; Khan, A.; Choudhary, R.K.; Balakrishnan, S.; Badraoui, R.; Bardakci, F.; et al. Plants in Anticancer Drug Discovery: From Molecular Mechanism to Chemoprevention. Biomed Res. Int. 2022, 2022. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Orhan, I.E.; Banach, M.; Rollinger, J.M.; Barreca, D.; Weckwerth, W.; Bauer, R.; Bayer, E.A.; et al. Natural products in drug discovery: advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef]
- Sezer, F.; Deniz, S.; Sevim, D.; Chaachouay, N.; Zidane, L. Plant-Derived Natural Products: A Source for Drug Discovery and Development. Drugs Drug Candidates 2024, Vol. 3, Pages 184-207 2024, 3, 184–207. [Google Scholar] [CrossRef]
- Tzvetkov, N.T.; Kirilov, K.; Matin, M.; Atanasov, A.G. Natural product drug discovery and drug design: two approaches shaping new pharmaceutical development. Nephrol. Dial. Transplant. 2024, 39, 375–378. [Google Scholar] [CrossRef]
- Anwar, F.; Latif, S.; Ashraf, M.; Gilani, A.H. Moringa oleifera: A food plant with multiple medicinal uses. Phyther. Res. 2007, 21, 17–25. [Google Scholar] [CrossRef]
- Pandey, A.; Pradheep, K.; Gupta, R.; Nayar, E.R.; Bhandari, D.C. “Drumstick tree” (Moringa oleifera Lam.): A multipurpose potential species in India. Genet. Resour. Crop Evol. 2011, 58, 453–460. [Google Scholar] [CrossRef]
- Islam, Z.; Islam, S.M.R.; Hossen, F.; Mahtab-Ul-Islam, K.; Hasan, M.R.; Karim, R. Moringa oleifera is a Prominent Source of Nutrients with Potential Health Benefits. Int. J. Food Sci. 2021, 2021. [Google Scholar] [CrossRef]
- Alhakmani, F.; Kumar, S.; Khan, S.A. Estimation of total phenolic content, in–vitro antioxidant and anti–inflammatory activity of flowers of Moringa oleifera. Asian Pac. J. Trop. Biomed. 2013, 3, 623–627. [Google Scholar] [CrossRef] [PubMed]
- Nkukwana, T.T.; Muchenje, V.; Pieterse, E.; Masika, P.J.; Mabusela, T.P.; Hoffman, L.C.; Dzama, K. Effect of Moringa oleifera leaf meal on growth performance, apparent digestibility, digestive organ size and carcass yield in broiler chickens. Livest. Sci. 2014, 161, 139–146. [Google Scholar] [CrossRef]
- Kashyap, P.; Kumar, S.; Riar, C.S.; Jindal, N.; Baniwal, P.; Guiné, R.P.F.; Correia, P.M.R.; Mehra, R.; Kumar, H. Recent Advances in Drumstick (Moringa oleifera) Leaves Bioactive Compounds: Composition, Health Benefits, Bioaccessibility, and Dietary Applications. Antioxidants 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Tiloke, C.; Anand, K.; Gengan, R.M.; Chuturgoon, A.A. Moringa oleifera and their phytonanoparticles: Potential antiproliferative agents against cancer. Biomed. Pharmacother. 2018, 108, 457–466. [Google Scholar] [CrossRef]
- Mumtaz, M.Z.; Kausar, F.; Hassan, M.; Javaid, S.; Malik, A. Anticancer activities of phenolic compounds from Moringa oleifera leaves: in vitro and in silico mechanistic study. Beni-Suef Univ. J. Basic Appl. Sci. 2021, 10, 1–11. [Google Scholar] [CrossRef]
- Wu, Y.-Y.; Xu, Y.-M.; Y Lau, A.T.; Ferreira de Oliveira, P.; Ribeiro, D.; Ascenso, A.; Santos, C. Anti-Cancer and Medicinal Potentials of Moringa Isothiocyanate. Mol. 2021, Vol. 26, Page 7512 2021, 26, 7512. [Google Scholar] [CrossRef]
- Ramamurthy, S.; Varghese, S.; Sudarsan, S.; Muruganandhan, J.; Mushtaq, S. Moringa oleifera: Antioxidant, Anticancer, Anti-inflammatory, and Related Properties of Extracts in Cell Lines: A Review of Medicinal Effects, Phytochemistry, and Applications. J. Contemp. Dent. Pract. 2021. [Google Scholar] [CrossRef]
- Singh, J.; Gautam, D.N.S.; Sourav, S.; Sharma, R. Role of Moringa oleifera Lam. in cancer: Phytochemistry and pharmacological insights. Food Front. 2023, 4, 164–206. [Google Scholar] [CrossRef]
- Abd-Rabou, A.A.; Abdalla, A.M.; Ali, N.A.; Zoheir, K.M.A. Moringa oleifera Root Induces Cancer Apoptosis more Effectively than Leave Nanocomposites and Its Free Counterpart. Asian Pac. J. Cancer Prev. 2017, 18, 2141. [Google Scholar] [CrossRef]
- Bhadresha, K.; Thakore, V.; Brahmbhatt, J.; Upadhyay, V.; Jain, N.; Rawal, R. Anticancer effect of Moringa oleifera leaves extract against lung cancer cell line via induction of apoptosis. Adv. Cancer Biol. - Metastasis 2022, 6, 100072. [Google Scholar] [CrossRef]
- Xie, J.; Peng, L.J.; Yang, M.R.; Jiang, W.W.; Mao, J.Y.; Shi, C.Y.; Tian, Y.; Sheng, J. Alkaloid Extract of Moringa oleifera Lam. Exerts Antitumor Activity in Human Non-Small-Cell Lung Cancer via Modulation of the JAK2/STAT3 Signaling Pathway. Evidence-based Complement. Altern. Med. 2021, 2021. [Google Scholar] [CrossRef]
- Ji, H.; Li, K.; Xu, W.; Li, R.; Xie, S.; Zhu, X. Prediction of the Mechanisms by Which Quercetin Enhances Cisplatin Action in Cervical Cancer: A Network Pharmacology Study and Experimental Validation. Front. Oncol. 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Lem, F.F.; Jiunn, D.; Lee, H.; Chee, F.T.; Chin, S.N.; Lin, K.M.; Yew, C.W. Network pharmacology approach to reveals therapeutic mech-anism of traditional plants formulation used by Malaysia in-digenous ethnics in coronaviruses infection. ChemRxiv 2022. [Google Scholar] [CrossRef]
- Li, L.; Yang, L.; Yang, L.; He, C.; He, Y.; Chen, L.; Dong, Q.; Zhang, H.; Chen, S.; Li, P. Network pharmacology: a bright guiding light on the way to explore the personalized precise medication of traditional Chinese medicine. Chin. Med. 2023, 18, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Gogoi, B.; Saikia, S.P. Virtual Screening and Network Pharmacology-Based Study to Explore the Pharmacological Mechanism of Clerodendrum Species for Anticancer Treatment. Evidence-based Complement. Altern. Med. 2022, 2022. [Google Scholar] [CrossRef]
- Lipinski, C.A. Lead- and drug-like compounds: The rule-of-five revolution. Drug Discov. Today Technol. 2004, 1, 337–341. [Google Scholar] [CrossRef]
- Guan, L.; Yang, H.; Cai, Y.; Sun, L.; Di, P.; Li, W.; Liu, G.; Tang, Y. ADMET-score – a comprehensive scoring function for evaluation of chemical drug-likeness. Medchemcomm 2019, 10, 148. [Google Scholar] [CrossRef]
- Gfeller, D.; Grosdidier, A.; Wirth, M.; Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: a web server for target prediction of bioactive small molecules. Nucleic Acids Res. 2014, 42. [Google Scholar] [CrossRef] [PubMed]
- Piñero, J.; Ramírez-Anguita, J.M.; Saüch-Pitarch, J.; Ronzano, F.; Centeno, E.; Sanz, F.; Furlong, L.I. The DisGeNET knowledge platform for disease genomics: 2019 update. Nucleic Acids Res. 2020, 48, D845–D855. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607. [Google Scholar] [CrossRef]
- Zhang, M.; Lu, P.; Zhao, F.; Sun, X.; Ma, W.; Tang, J.; Zhang, C.; Ji, H.; Wang, X. Uncovering the molecular mechanisms of Curcumae Rhizoma against myocardial fibrosis using network pharmacology and experimental validation. J. Ethnopharmacol. 2023, 300. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: new perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 2017, 45, D353–D361. [Google Scholar] [CrossRef]
- Liu, J.; Meng, J.; Li, R.; Jiang, H.; Fu, L.; Xu, T.; Zhu, G.Y.; Zhang, W.; Gao, J.; Jiang, Z.H.; et al. Integrated network pharmacology analysis, molecular docking, LC-MS analysis and bioassays revealed the potential active ingredients and underlying mechanism of Scutellariae radix for COVID-19. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef]
- Nasab, R.R.; Mansourian, M.; Hassanzadeh, F.; Shahlaei, M. Exploring the interaction between epidermal growth factor receptor tyrosine kinase and some of the synthesized inhibitors using combination of in-silico and in-vitro cytotoxicity methods. Res. Pharm. Sci. 2018, 13, 509. [Google Scholar] [CrossRef]
- Koska, J.; Spassov, V.Z.; Maynard, A.J.; Yan, L.; Austin, N.; Flook, P.K.; Venkatachalam, C.M. Fully automated molecular mechanics based induced fit protein - Ligand docking method. J. Chem. Inf. Model. 2008, 48, 1965–1973. [Google Scholar] [CrossRef]
- Yang, C.; Yang, Z.; Tong, K.; Wang, J.; Yang, W.; Yu, R.; Jiang, F.; Ji, Y. Homology modeling and molecular docking simulation of martentoxin as a specific inhibitor of the BK channel. Ann. Transl. Med. 2022, 10, 71–71. [Google Scholar] [CrossRef]
- Mohanraj, K.; Karthikeyan, B.S.; Vivek-Ananth, R.P.; Chand, R.P.B.; Aparna, S.R.; Mangalapandi, P.; Samal, A. IMPPAT: A curated database of Indian Medicinal Plants, Phytochemistry And Therapeutics. Sci. Rep. 2018, 8, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Lans, C.; van Asseldonk, T. Dr. Duke’s Phytochemical and Ethnobotanical Databases, a Cornerstone in the Validation of Ethnoveterinary Medicinal Plants, as Demonstrated by Data on Pets in British Columbia. 2020, 219–246. [Google Scholar] [CrossRef]
- Ali M 2019 PCIDB - PhytoChemical Interactions DataBase presentation by: DR. MOHAMMED ALI ABD EL-HAMMED ABD ALLAH Contact information Phone: Research gatea. [CrossRef]
- Zhao, J.; Mo, C.; Shi, W.; Meng, L.; Ai, J. Network Pharmacology Combined with Bioinformatics to Investigate the Mechanisms and Molecular Targets of Astragalus Radix-Panax notoginseng Herb Pair on Treating Diabetic Nephropathy. Evid. Based. Complement. Alternat. Med. 2021, 2021. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yuan, Y.; Wang, W.; He, Y.; Zhong, H.; Zhou, X.; Chen, Y.; Cai, X.J.; Liu, L. qin Mechanisms underlying the therapeutic effects of Qingfeiyin in treating acute lung injury based on GEO datasets, network pharmacology and molecular docking. Comput. Biol. Med. 2022, 145. [Google Scholar] [CrossRef]
- Rah, B.; Rather, R.A.; Bhat, G.R.; Baba, A.B.; Mushtaq, I.; Farooq, M.; Yousuf, T.; Dar, S.B.; Parveen, S.; Hassan, R.; et al. JAK/STAT Signaling: Molecular Targets, Therapeutic Opportunities, and Limitations of Targeted Inhibitions in Solid Malignancies. Front. Pharmacol. 2022, 13, 1–19. [Google Scholar] [CrossRef]
- Ebrahimnezhad, M.; Natami, M.; Bakhtiari, G.H.; Tabnak, P.; Ebrahimnezhad, N.; Yousefi, B.; Majidinia, M. FOXO1, a tiny protein with intricate interactions: Promising therapeutic candidate in lung cancer. Biomed. Pharmacother. 2023, 169, 115900. [Google Scholar] [CrossRef]
- Pradhan, R.; Singhvi, G.; Dubey, S.K.; Gupta, G.; Dua, K. MAPK Pathway: a Potential Target for the Treatment of Non-small-cell Lung Carcinoma. Future Med. Chem. 2019, 11, 793–795. [Google Scholar] [CrossRef]
- Bahar, M.E.; Kim, H.J.; Kim, D.R. Targeting the RAS/RAF/MAPK pathway for cancer therapy: from mechanism to clinical studies. Signal Transduct. Target. Ther. 2023 81 2023, 8, 1–38. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, Z.; Tang, D.; Liu, H.; Luo, S.; Stinchcombe, T.E.; Glass, C.; Su, L.; Lin, L.; Christiani, D.C.; et al. Potentially functional variants of HBEGF and ITPR3 in GnRH signaling pathway genes predict survival of non-small cell lung cancer patients. Transl. Res. 2021, 233, 92–103. [Google Scholar] [CrossRef]
- Sanaei, M.J.; Razi, S.; Pourbagheri-Sigaroodi, A.; Bashash, D. The PI3K/Akt/mTOR pathway in lung cancer; oncogenic alterations, therapeutic opportunities, challenges, and a glance at the application of nanoparticles. Transl. Oncol. 2022, 18, 101364. [Google Scholar] [CrossRef]
- Del Re, M.; Cucchiara, F.; Petrini, I.; Fogli, S.; Passaro, A.; Crucitta, S.; Attili, I.; De Marinis, F.; Chella, A.; Danesi, R. erbB in NSCLC as a molecular target: current evidences and future directions. ESMO Open 2020, 5, 724. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Xing, C.; Deng, Y.; Ye, C.; Peng, H. HIF-1α signaling: Essential roles in tumorigenesis and implications in targeted therapies. Genes Dis. 2023, 11, 234. [Google Scholar] [CrossRef]
- Zhang, Y.L.; Wang, R.C.; Cheng, K.; Ring, B.Z.; Su, L. Roles of Rap1 signaling in tumor cell migration and invasion. Cancer Biol. Med. 2017, 14, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.W.; Li, Y.; Yang, Y.; Yang, H.K.; Dong, J.M.; Xiao, Z.H.; He, X.; Guo, J.H.; Wang, R.Q.; Dai, B.; et al. Tumor immunotherapy resistance: Revealing the mechanism of PD-1 / PD-L1-mediated tumor immune escape. Biomed. Pharmacother. 2024, 171, 116203. [Google Scholar] [CrossRef] [PubMed]
- Anvar, M.T.; Rashidan, K.; Arsam, N.; Rasouli-Saravani, A.; Yadegari, H.; Ahmadi, A.; Asgari, Z.; Vanan, A.G.; Ghorbaninezhad, F.; Tahmasebi, S. Th17 cell function in cancers: immunosuppressive agents or anti-tumor allies? Cancer Cell Int. 2024, 24. [Google Scholar] [CrossRef]
- Zhu, W.; Wang, H.; Zhu, D. Wnt/β-catenin signaling pathway in lung cancer. Med. Drug Discov. 2022, 13, 100113. [Google Scholar] [CrossRef]
- Yun, C.H.; Mengwasser, K.E.; Toms, A.V.; Woo, M.S.; Greulich, H.; Wong, K.K.; Meyerson, M.; Eck, M.J. The T790M mutation in EGFR kinase causes drug resistance by increasing the affinity for ATP. Proc. Natl. Acad. Sci. USA 2008, 105, 2070–2075. [Google Scholar] [CrossRef]
- Palanivel, S.; Yli-Harja, O.; Kandhavelu, M. Molecular interaction study of novel indoline derivatives with EGFR-kinase domain using multiple computational analysis. J. Biomol. Struct. Dyn. 2022, 40, 7545–7554. [Google Scholar] [CrossRef]
- Jura, N.; Endres, N.F.; Engel, K.; Deindl, S.; Das, R.; Lamers, M.H.; Wemmer, D.E.; Zhang, X.; Kuriyan, J. Mechanism for activation of the EGF receptor catalytic domain by the juxtamembrane segment. Cell 2009, 137, 1293. [Google Scholar] [CrossRef]
- Martin-Fernandez, M.L.; Clarke, D.T.; Roberts, S.K.; Zanetti-Domingues, L.C.; Gervasio, F.L. Structure and Dynamics of the EGF Receptor as Revealed by Experiments and Simulations and Its Relevance to Non-Small Cell Lung Cancer. Cells 2019, 8, 316. [Google Scholar] [CrossRef]
- Vicidomini, G. Current Challenges and Future Advances in Lung Cancer: Genetics, Instrumental Diagnosis and Treatment. Cancers 2023, 15, 3710. [Google Scholar] [CrossRef] [PubMed]
- Oyewusi, H.A.; Wu, Y.S.; Safi, S.Z.; Wahab, R.A.; Hatta, M.H.M.; Batumalaie, K. Molecular dynamics simulations reveal the inhibitory mechanism of Withanolide A against α-glucosidase and α-amylase. J. Biomol. Struct. Dyn. 2023, 41, 6203–6218. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.P.; Singh, A.; Wolkenhauer, O.; Gupta, S.K. Regulatory Role of IL6 in Immune-Related Adverse Events during Checkpoint Inhibitor Treatment in Melanoma. Int. J. Mol. Sci. 2024, 25, 10600. [Google Scholar] [CrossRef]
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef] [PubMed]
- Nasim, N.; Sandeep, I.S.; Mohanty, S. Plant-derived natural products for drug discovery: current approaches and prospects. Nucl. 2022, 65, 399–411. [Google Scholar] [CrossRef]
- Stoker, D.G.; Reber, K.R.; Waltzman, L.S.; Ernst, C.; Hamilton, D.; Gawarecki, D.; Mermelstein, F.; McNicol, E.; Wright, C.; Carr, D.B. Analgesic efficacy and safety of morphine-chitosan nasal solution in patients with moderate to severe pain following orthopedic surgery. Pain Med. 2008, 9, 3–12. [Google Scholar] [CrossRef]
- Sze, D.M.Y.; Miller, K.; Neilan, B. Development of Taxol and Other Endophyte Produced Anti-Cancer Agents. Recent Pat. Anticancer. Drug Discov. 2008, 3, 14–19. [Google Scholar] [CrossRef]
- Rai, V.; Tandon, P.K.; Khatoon, S. Effect of Chromium on Antioxidant Potential of Catharanthus roseus Varieties and Production of Their Anticancer Alkaloids: Vincristine and Vinblastine. Biomed Res. Int. 2014, 2014, 934182. [Google Scholar] [CrossRef]
- Liang, X.; Wang, P.; Yang, C.; Huang, F.; Wu, H.; Shi, H.; Wu, X. Galangin Inhibits Gastric Cancer Growth Through Enhancing STAT3 Mediated ROS Production. Front. Pharmacol. 2021, 12, 646628. [Google Scholar] [CrossRef]
- Yao, C.; Dai, S.; Wang, C.; Fu, K.; Wu, R.; Zhao, X.; Yao, Y.; Li, Y. Luteolin as a potential hepatoprotective drug: Molecular mechanisms and treatment strategies. Biomed. Pharmacother. 2023, 167, 115464. [Google Scholar] [CrossRef]
- Li, W.; Yuan, G.; Pan, Y.; Wang, C.; Chen, H. Network Pharmacology Studies on the Bioactive Compounds and Action Mechanisms of Natural Products for the Treatment of Diabetes Mellitus: A Review. Front. Pharmacol. 2017, 8, 74. [Google Scholar] [CrossRef] [PubMed]
- Ya’u Ibrahim, Z.; Uzairu, A.; Shallangwa, G.; Abechi, S. Molecular docking studies, drug-likeness and in-silico ADMET prediction of some novel β-Amino alcohol grafted 1,4,5-trisubstituted 1,2,3-triazoles derivatives as elevators of p53 protein levels. Sci. African 2020, 10, e00570. [Google Scholar] [CrossRef]
- Lin, C.L.; Chen, R.F.; Chen, J.Y.F.; Chu, Y.C.; Wang, H.M.; Chou, H.L.; Chang, W.C.; Fong, Y.; Chang, W.T.; Wu, C.Y.; et al. Protective Effect of Caffeic Acid on Paclitaxel Induced Anti-Proliferation and Apoptosis of Lung Cancer Cells Involves NF-κB Pathway. Int. J. Mol. Sci. 2012, 13, 6236. [Google Scholar] [CrossRef]
- Cal, M.; Szakonyi, Z.; Pavlíková, N. Caffeic Acid and Diseases—Mechanisms of Action. Int. J. Mol. Sci. 2022, 24, 588. [Google Scholar] [CrossRef]
- Lopes, R.; Costa, M.; Ferreira, M.; Gameiro, P.; Fernandes, S.; Catarino, C.; Santos-Silva, A.; Paiva-Martins, F. Caffeic acid phenolipids in the protection of cell membranes from oxidative injuries. Interaction with the membrane phospholipid bilayer. Biochim. Biophys. Acta - Biomembr. 2021, 1863, 183727. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.X.; Wang, L.; Yang, J.L.; Li, L.Z.; Xu, W.M.; Li, T. Caffeic acid phenethyl ester attenuates pro-inflammatory and fibrogenic phenotypes of LPS-stimulated hepatic stellate cells through the inhibition of NF-κB signaling. Int. J. Mol. Med. 2014, 33, 687–694. [Google Scholar] [CrossRef]
- Mudgal, J.; Basu Mallik, S.; Nampoothiri, M.; Kinra, M.; Hall, S.; Grant, G.D.; Anoopkumar-Dukie, S.; Davey, A.K.; Rao, C.M.; Arora, D. Effect of coffee constituents, caffeine and caffeic acid on anxiety and lipopolysaccharide-induced sickness behavior in mice. J. Funct. Foods 2020, 64, 103638. [Google Scholar] [CrossRef]
- Mirzaei, S.; Gholami, M.H.; Zabolian, A.; Saleki, H.; Farahani, M.V.; Hamzehlou, S.; Far, F.B.; Sharifzadeh, S.O.; Samarghandian, S.; Khan, H.; et al. Caffeic acid and its derivatives as potential modulators of oncogenic molecular pathways: New hope in the fight against cancer. Pharmacol. Res. 2021, 171, 105759. [Google Scholar] [CrossRef]
- Alam, M.; Ahmed, S.; Elasbali, A.M.; Adnan, M.; Alam, S.; Hassan, M.I.; Pasupuleti, V.R. Therapeutic Implications of Caffeic Acid in Cancer and Neurological Diseases. Front. Oncol. 2022, 12, 860508. [Google Scholar] [CrossRef]
- Secme, M.; Mutlu, D.; Elmas, L.; Arslan, S. Assessing effects of caffeic acid on cytotoxicity, apoptosis, invasion, GST enzyme activity, oxidant, antioxidant status and micro-RNA expressions in HCT116 colorectal cancer cells. South African J. Bot. 2023, 157, 19–26. [Google Scholar] [CrossRef]
- Uribe, M.L.; Marrocco, I.; Yarden, Y. EGFR in Cancer: Signaling Mechanisms, Drugs, and Acquired Resistance. Cancers 2021, 13, 2748. [Google Scholar] [CrossRef] [PubMed]
- Harrison, P.T.; Vyse, S.; Huang, P.H. Rare epidermal growth factor receptor (EGFR) mutations in non-small cell lung cancer. Semin. Cancer Biol. 2020, 61, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Chi, S.A.; Yu, H.; Choi, Y. La; Park, S.; Sun, J.M.; Lee, S.H.; Ahn, J.S.; Ahn, M.J.; Choi, D.H.; Kim, K.; et al. Trends in Survival Rates of Non–Small Cell Lung Cancer With Use of Molecular Testing and Targeted Therapy in Korea, 2010-2020. JAMA Netw. Open 2023, 6, e232002–e232002. [Google Scholar] [CrossRef]
- Alam, M.; Hasan, G.M.; Eldin, S.M.; Adnan, M.; Riyaz, M.B.; Islam, A.; Khan, I.; Hassan, M.I. Investigating regulated signaling pathways in therapeutic targeting of non-small cell lung carcinoma. Biomed. Pharmacother. 2023, 161, 114452. [Google Scholar] [CrossRef] [PubMed]
- Hollingsworth, S.A.; Dror, R.O. Molecular dynamics simulation for all. Neuron 2018, 99, 1129. [Google Scholar] [CrossRef]
Figure 1.
This workflow identifies natural compounds from M. oleifera that target lung cancer cells. The methods were enhanced using various filters to refine the selection. Initially, natural compounds were retrieved from the databases and filtered based on parameters such as Lipinski’s Rule of 5 and ADMET. The compounds were then input into the SwissTarget database to identify related proteins. Lung cancer-related proteins were obtained from DisGeNET and TCGA databases. Proteins common between the two datasets were identified. Using the STRING database, these shared proteins were analysed through a protein-protein interaction (PPI) network. The PPI network was further examined in Cytoscape using a network analyzer tool. The most promising proteins were subjected to enrichment analysis and a network-based approach was used to identify potential targets. Compound-protein-pathway networks were employed to identify the most suitable natural compounds. Molecular docking was performed to determine the optimal compounds. Finally, the stability of the lead compound was assessed using molecular dynamics (MD) simulations.
Figure 1.
This workflow identifies natural compounds from M. oleifera that target lung cancer cells. The methods were enhanced using various filters to refine the selection. Initially, natural compounds were retrieved from the databases and filtered based on parameters such as Lipinski’s Rule of 5 and ADMET. The compounds were then input into the SwissTarget database to identify related proteins. Lung cancer-related proteins were obtained from DisGeNET and TCGA databases. Proteins common between the two datasets were identified. Using the STRING database, these shared proteins were analysed through a protein-protein interaction (PPI) network. The PPI network was further examined in Cytoscape using a network analyzer tool. The most promising proteins were subjected to enrichment analysis and a network-based approach was used to identify potential targets. Compound-protein-pathway networks were employed to identify the most suitable natural compounds. Molecular docking was performed to determine the optimal compounds. Finally, the stability of the lead compound was assessed using molecular dynamics (MD) simulations.
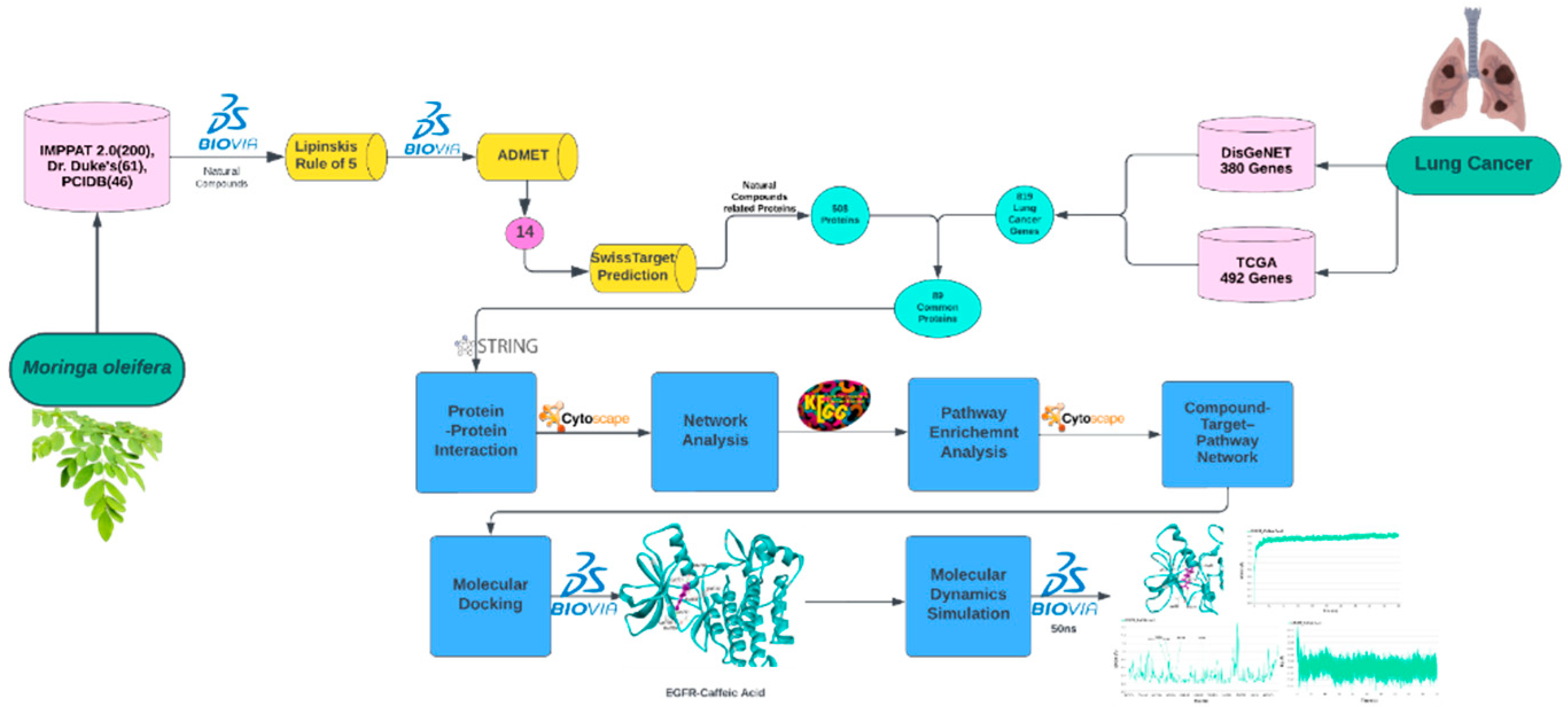
Figure 2.
The 14 natural compounds were selected after ADMET filtration using DS 2022. The 2D images of these compounds and their names and PubChem IDs were shown.
Figure 2.
The 14 natural compounds were selected after ADMET filtration using DS 2022. The 2D images of these compounds and their names and PubChem IDs were shown.
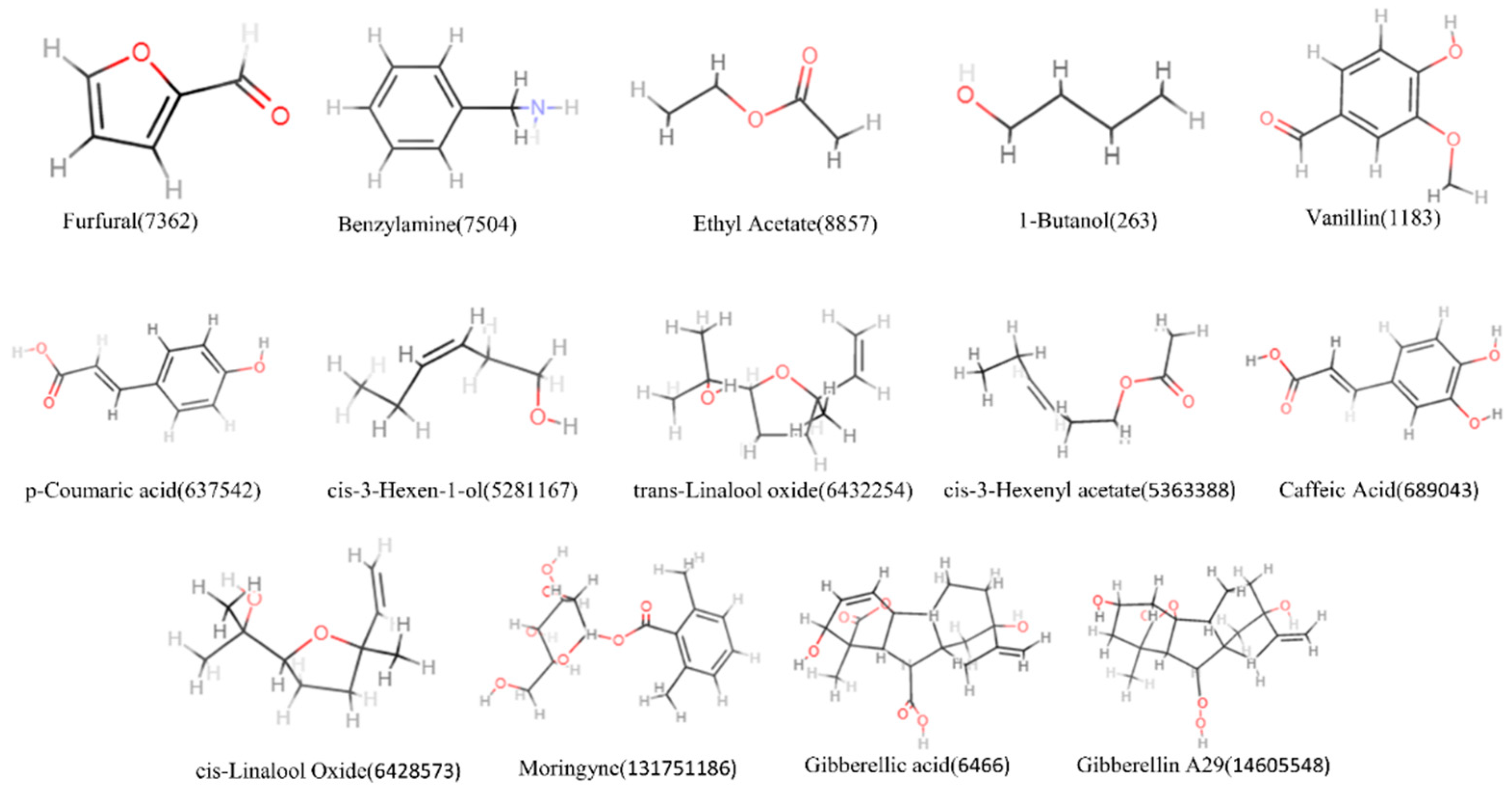
Figure 3.
Venn diagram of lung cancer-associated genes and M. oleifera Associated Genes; 89 common genes were present in both domains.
Figure 3.
Venn diagram of lung cancer-associated genes and M. oleifera Associated Genes; 89 common genes were present in both domains.
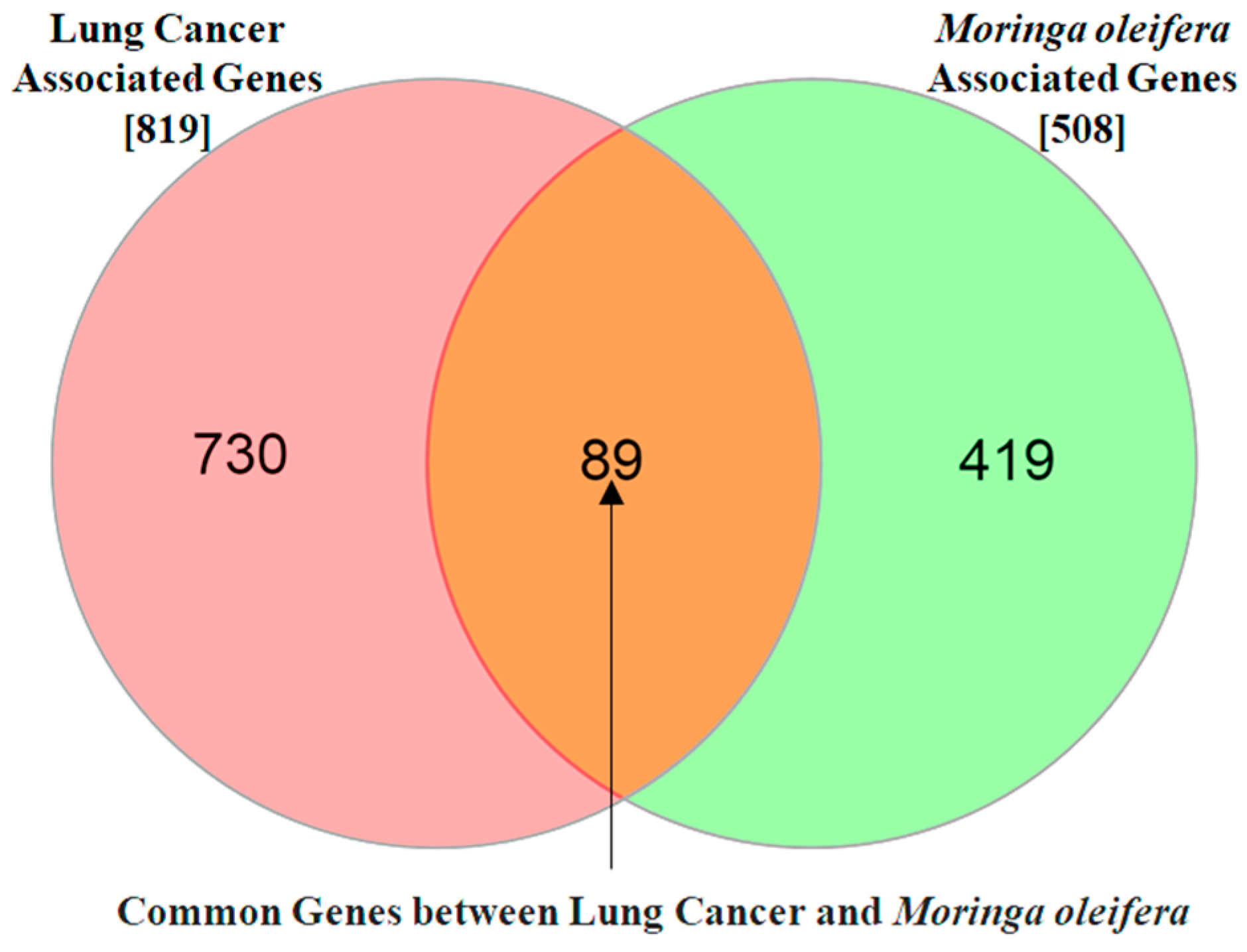
Figure 4.
The protein-protein interaction (PPI) analysis of 89 common targets between lung cancer and Moringa oleifera resulted in a network of 80 nodes and 314 edges. The screening process for the PPI network was conducted as follows: (i) The initial PPI network included 80 lung cancer targets.(ii) A refined PPI network was generated based on significant proteins with a degree median of ≤6.(iii) Another subset was created for targets with a closeness centrality median of ≤0.0295.(iv) Finally, a PPI network was constructed for targets with a betweenness centrality median of ≤0.4093.This networks highlight the most relevant targets for further analysis.
Figure 4.
The protein-protein interaction (PPI) analysis of 89 common targets between lung cancer and Moringa oleifera resulted in a network of 80 nodes and 314 edges. The screening process for the PPI network was conducted as follows: (i) The initial PPI network included 80 lung cancer targets.(ii) A refined PPI network was generated based on significant proteins with a degree median of ≤6.(iii) Another subset was created for targets with a closeness centrality median of ≤0.0295.(iv) Finally, a PPI network was constructed for targets with a betweenness centrality median of ≤0.4093.This networks highlight the most relevant targets for further analysis.
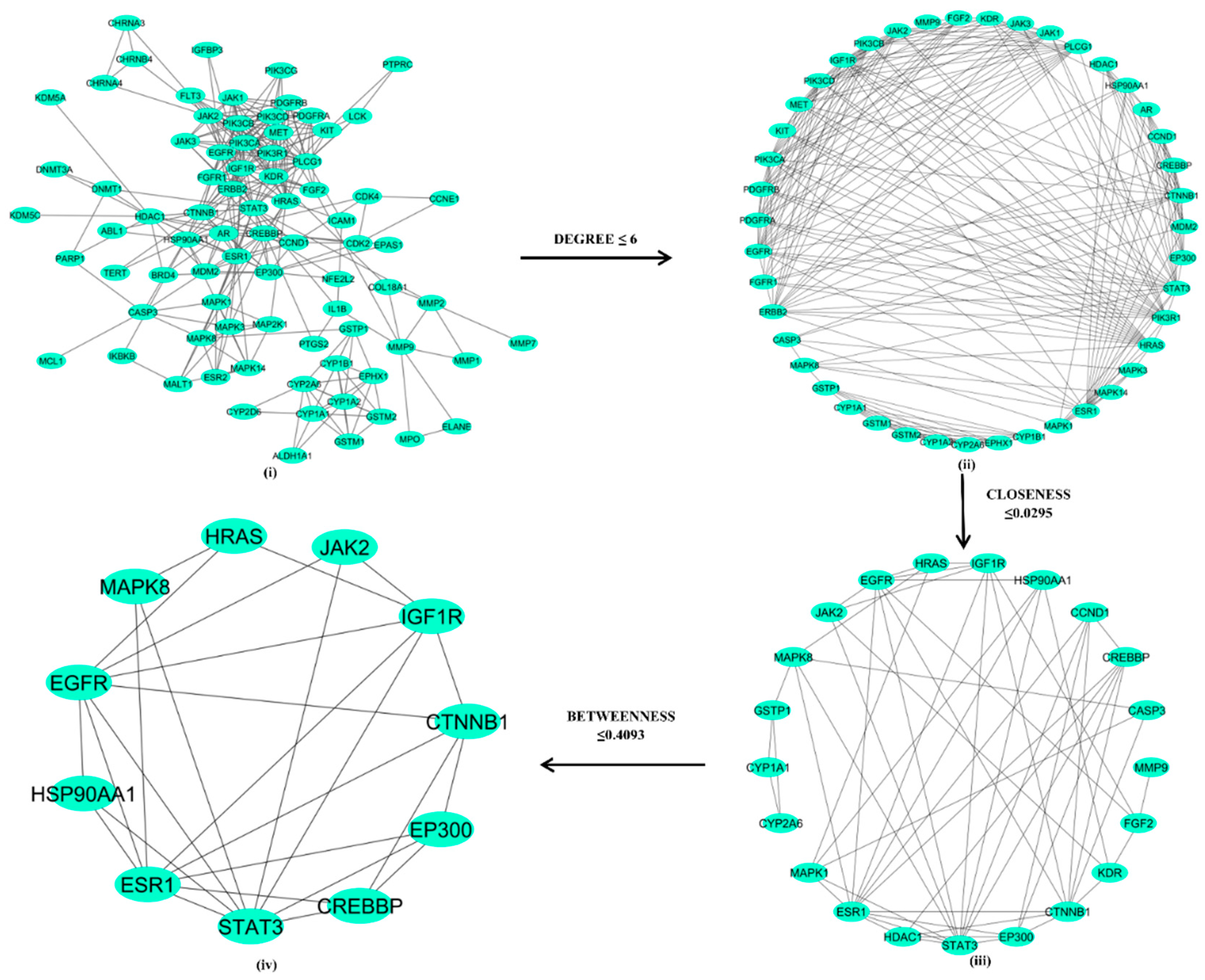
Figure 5.
Diagram showing Pathway-Protein interaction from the 11 key proteins. Within the network, all proteins underwent an analysis of enrichment pathways. Pathways are represented by green triangles, whereas proteins are represented by round nodes. A total of 14 pathways were enriched using different proteins in the network. From the analysis of this interactions, we observed that EGFR has the maximum interaction with the pathways, indicating that it has a greater impact than the other proteins.
Figure 5.
Diagram showing Pathway-Protein interaction from the 11 key proteins. Within the network, all proteins underwent an analysis of enrichment pathways. Pathways are represented by green triangles, whereas proteins are represented by round nodes. A total of 14 pathways were enriched using different proteins in the network. From the analysis of this interactions, we observed that EGFR has the maximum interaction with the pathways, indicating that it has a greater impact than the other proteins.
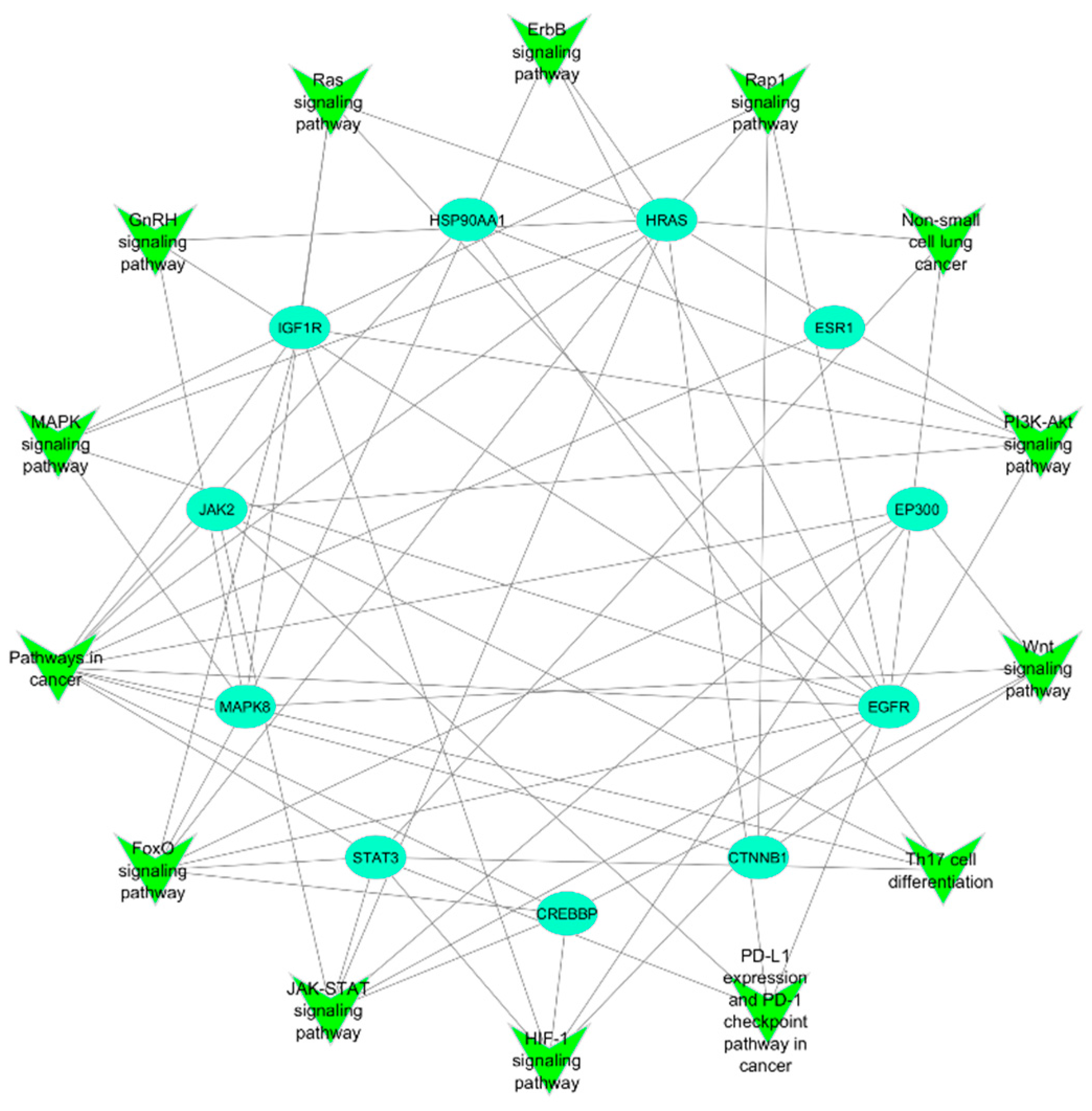
Figure 6.
Figure illustrates the network between natural compounds-target proteins-pathways interacting among themselves. Natural compounds are represented by the rectangle’s boxes, target proteins are represented by the circle and pathways are represented by the triangle nodes, respectively. Out of 14 natural compounds only 12 can interact with 11 target proteins. Eleven protein targets are making the interaction with the 14 pathways which plays vital role in the lung cancer progression. We also highlight the connection showing maximum compounds with the EGFR with dotted pink colour arrow and connection of EGFR targeting maximum pathways with dotted orange colour arrow.
Figure 6.
Figure illustrates the network between natural compounds-target proteins-pathways interacting among themselves. Natural compounds are represented by the rectangle’s boxes, target proteins are represented by the circle and pathways are represented by the triangle nodes, respectively. Out of 14 natural compounds only 12 can interact with 11 target proteins. Eleven protein targets are making the interaction with the 14 pathways which plays vital role in the lung cancer progression. We also highlight the connection showing maximum compounds with the EGFR with dotted pink colour arrow and connection of EGFR targeting maximum pathways with dotted orange colour arrow.
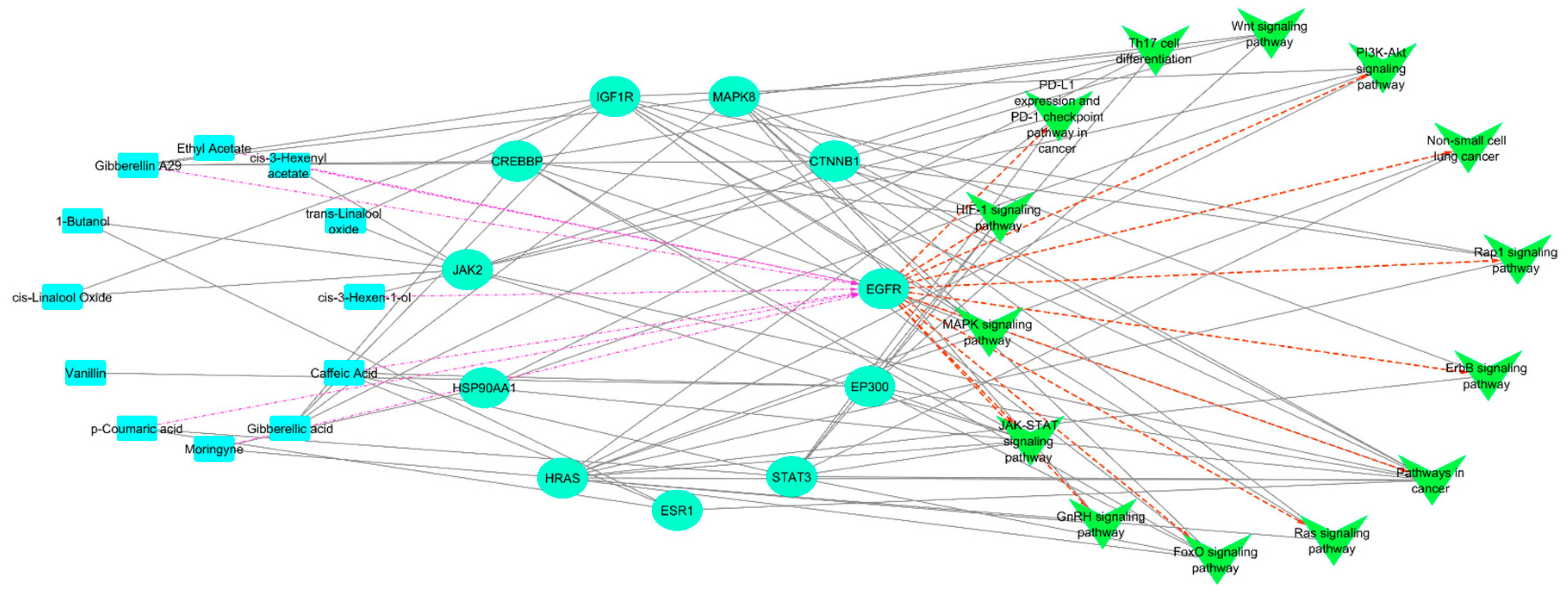
Figure 7.
The figure illustrates the comparative molecular docking analysis interactions of the control ligand (Erlotinib) with the EGFR active site shown in Figure 7 (i) and the natural compound (Caffeic acid) with the EGFR active site in Figure 7 (ii). In both the figures EGFR is representing cyan colour and ligands are purple colour.
Figure 7.
The figure illustrates the comparative molecular docking analysis interactions of the control ligand (Erlotinib) with the EGFR active site shown in Figure 7 (i) and the natural compound (Caffeic acid) with the EGFR active site in Figure 7 (ii). In both the figures EGFR is representing cyan colour and ligands are purple colour.
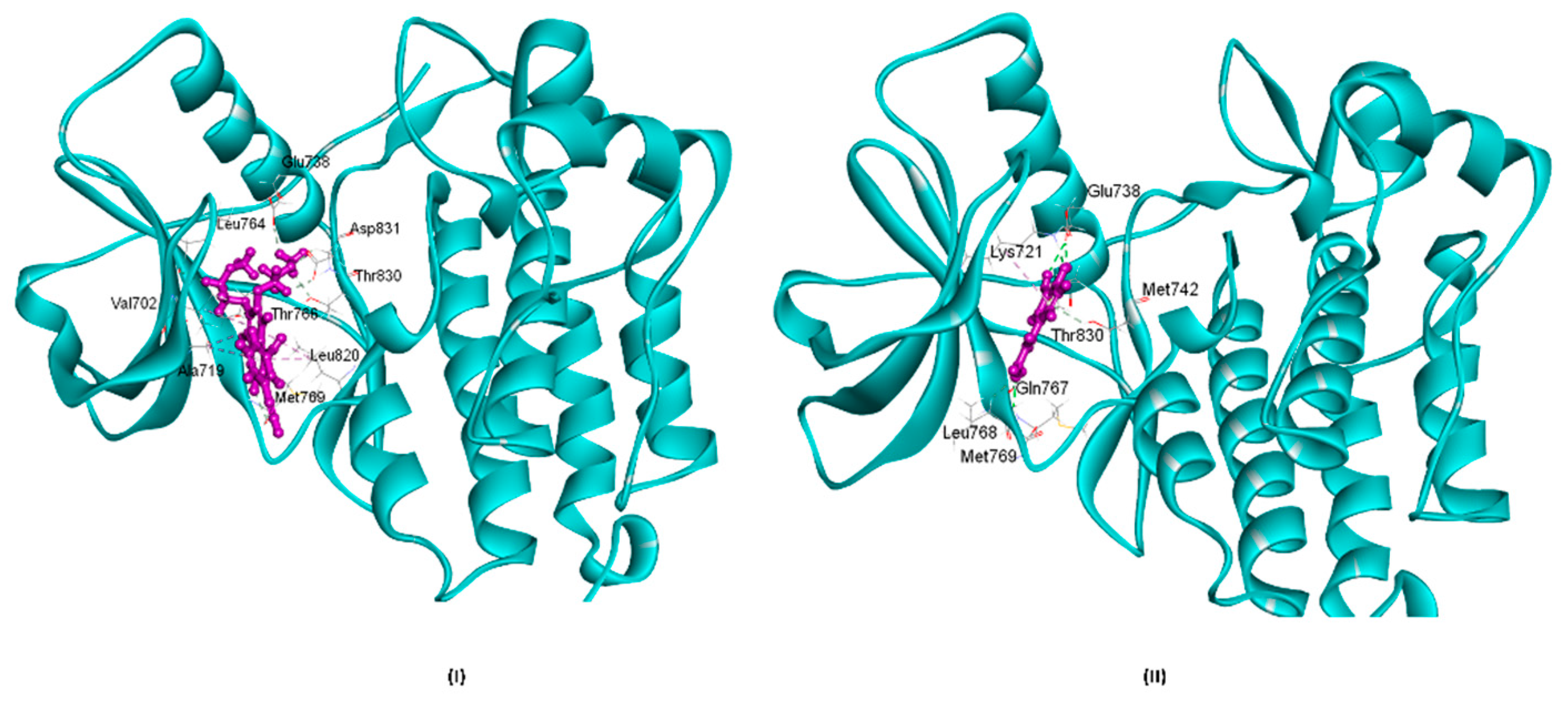
Figure 8.
This illustration depicts data regarding the bonds established by Caffeic acid within the active site of the EGFR protein throughout MD simulation as shown in figure 8 (i). Figure 8 (ii) Illustrate the root mean square deviation (RMSD) of the docked complex of Caffeic Acid over a simulation period of 50 nanoseconds (ns). In the production run, the starting frame up to 10ns shows the increase in the trend of RMSD values ranging from 1.6 to 2.0 Å in Caffeic Acid. After the equilibrium condition, the EGFR-Caffeic Acid complex exhibited less deviation. The overall MD result showed that Caffeic Acid complex had converged, and no deviations were observed in the protein. In Figure 8 (iii) RMSF, the graph illustrates the fluctuations of EGFR residues involved in bond formation during the MD simulation. It was observed that all these residues exhibited minimal fluctuations within the active site cavity of EGFR. The graph indicates that most fluctuations remained within a range of 0.8 Å. Figure 8 (iv) Illustrates the radius of gyration (Rg) of a docked complex of Caffeic Acid, which became more compact during the MD simulation. Initially, it measured around 22.12 Å, but after achieving a stable conformation during the MD simulation, it decreased to 22.04 Å.
Figure 8.
This illustration depicts data regarding the bonds established by Caffeic acid within the active site of the EGFR protein throughout MD simulation as shown in figure 8 (i). Figure 8 (ii) Illustrate the root mean square deviation (RMSD) of the docked complex of Caffeic Acid over a simulation period of 50 nanoseconds (ns). In the production run, the starting frame up to 10ns shows the increase in the trend of RMSD values ranging from 1.6 to 2.0 Å in Caffeic Acid. After the equilibrium condition, the EGFR-Caffeic Acid complex exhibited less deviation. The overall MD result showed that Caffeic Acid complex had converged, and no deviations were observed in the protein. In Figure 8 (iii) RMSF, the graph illustrates the fluctuations of EGFR residues involved in bond formation during the MD simulation. It was observed that all these residues exhibited minimal fluctuations within the active site cavity of EGFR. The graph indicates that most fluctuations remained within a range of 0.8 Å. Figure 8 (iv) Illustrates the radius of gyration (Rg) of a docked complex of Caffeic Acid, which became more compact during the MD simulation. Initially, it measured around 22.12 Å, but after achieving a stable conformation during the MD simulation, it decreased to 22.04 Å.
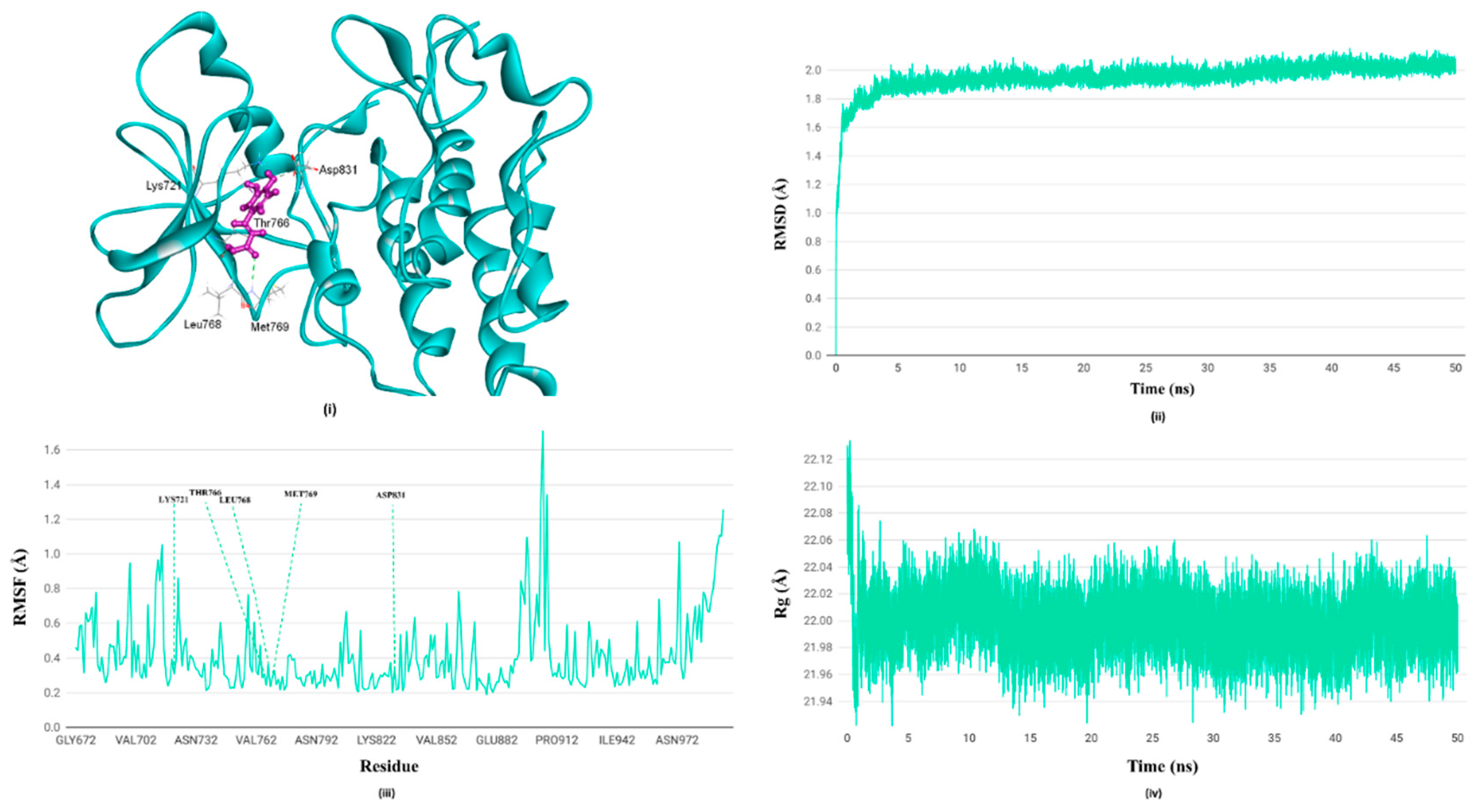

Table 1.
This table highlights 11 proteins identified through network analysis and their interconnection to 14 pathways. The proteins present in the pathways are represented using tick marks, while blanks indicate no interaction with pathways. Among these, EGFR stands out as a key player, interacting with 12 pathways more than any other protein. Notably, these include major pathways associated with lung cancer, such as non-small cell lung cancer, Ras signaling, PI3K/AKT signaling, MAPK signaling, and JAK-STAT signaling. This suggests that inhibiting EGFR could potentially have a significant impact on reducing the activity of pathways linked to lung cancer.
Table 1.
This table highlights 11 proteins identified through network analysis and their interconnection to 14 pathways. The proteins present in the pathways are represented using tick marks, while blanks indicate no interaction with pathways. Among these, EGFR stands out as a key player, interacting with 12 pathways more than any other protein. Notably, these include major pathways associated with lung cancer, such as non-small cell lung cancer, Ras signaling, PI3K/AKT signaling, MAPK signaling, and JAK-STAT signaling. This suggests that inhibiting EGFR could potentially have a significant impact on reducing the activity of pathways linked to lung cancer.
| Pathways in cancer | FoxO signaling pathway | JAK-STAT signaling pathway | HIF-1 signaling pathway | PD-L1 expression and PD-1 checkpoint pathway in cancer | Th17 cell differentiation | Wnt signaling pathway | PI3K-Akt signaling pathway | Non-small cell lung cancer | Rap1 signaling pathway | ErbB signaling pathway | Ras signaling pathway | GnRH signaling pathway | MAPK signaling pathway | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EGFR | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ||
| STAT3 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | |||||||
| ESR1 | ✓ | |||||||||||||
| MAPK8 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ||||||
| IGF1R | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | |||||||
| CTNNB1 | ✓ | ✓ | ✓ | |||||||||||
| CREBBP | ✓ | ✓ | ✓ | ✓ | ✓ | |||||||||
| HSP90AA1 | ✓ | ✓ | ✓ | |||||||||||
| HRAS | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ||||
| JAK2 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ||||||||
| EP300 | ✓ | ✓ | ✓ | ✓ | ✓ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



