
Submitted:
01 April 2025
Posted:
02 April 2025
You are already at the latest version
Abstract
Beyond its immunological role, colostrum has emerged as a promising, non-invasive source of bioactive factors, including Mesenchymal Stromal Cells (MSCs). This study represents the first attempt to isolate and characterize MSCs from equine colostrum (C-MSCs) and assess their potential use in veterinary regenerative medicine. Colostrum (n=6) was collected immediately after delivery, centrifuged, and recovered cells cultured under standard conditions. C-MSCs displayed plastic adherence and heterogeneous morphology, including spindle-shaped and epithelial-like cells. PDT values varied among samples, and 4/6 showed rapid proliferation (< 2 days). CFU assays confirmed clonogenic potential, though significant inter-sample variability was observed (p< 0.05). Spheroid formation assays revealed differences in cell-cell adhesion: 4/6 samples formed stable spheroids within four days. Migration assay showed significant variability (p< 0.05): 1/6 achieved complete wound closure within 72 hours, whereas 5/6 reached ~30% at 96 hours. Differentiation assays confirmed trilineage potential, with all samples staining positive for adipogenic, chondrogenic, and osteogenic differentiation. RT-PCR confirmed MSC marker expression, while hematopoietic markers were absent. MHC-I expression was weak in 5/6 samples, whereas MHC-II was consistently negative. These findings support equine colostrum as a viable MSC source, though variability requires further validation with larger samples. Additional research is needed to investigate C-MSCs' immunomodulatory properties and therapeutic potential.
Keywords:
Equine
; Mesenchymal Stromal Cells
; Colostrum
; Cell characterization
1. Introduction
Colostrum is the first, thick, nutrient-rich secretion produced by the female mammary gland during the final 1-2 weeks of pregnancy, as a response to hormonal changes [1,2]. In equine species, due to the epitheliochorial nature of the placenta, colostrum is vital for foal survival, serving as the newborn's sole natural source of immunoglobulins, fulfilling essential nutritional needs and supporting gastrointestinal tract maturation [2,3,4].
The mammary gland is a highly metabolically active organ that undergoes cyclic phases of proliferation and hypertrophy during pregnancy and lactation, followed by post-lactation involution [5,6,7]. These processes are thought to be supported by mammary stromal cells (MaSCs), which possess the potential to differentiate into the various cellular components essential for lactation [8,9].
The functional unit of the mammary gland is the epithelium, composed of luminal and myoepithelial cells [10,11], both of which are thought to originate from a common progenitor derived from MaSCs [8,9,12] These MaSCs are likely of epithelial origin and, probably, they are present in milk, having been shed from the ductal and luminal epithelial layers. This shedding may result from the increased turnover of secretory tissue or mechanical shear forces induced by the continuous cycles of milk filling and emptying during lactation [7,13]. In 2007, Cregan et al. [14] provided the first evidence that breast milk contains cells with stem cell/progenitor properties. They showed that cell colonies established in culture from breast milk contained cells positive for the mammary stem cell marker CK5 and the general stem cell marker Nestin [14]. The ex vivo presence of CK5- and Nestin-positive cells in breast milk was subsequently confirmed by other investigators [7,15] and further expanded with the identification of breastmilk stem cells (BSCs) [16]. BSCs not only exhibit self-renewal capabilities in culture but also express pluripotency-associated genes, including the core transcription factors OCT4, SOX2 and NANOG, along with downstream targets KLF4, REX1, and GDF3 [16]. Remarkably, these BSC subpopulations appear highly plastic, capable of differentiating into cell types derived from all three germ layers, including neurons, glia, hepatocytes, pancreatic cells, cardiomyocytes, osteoblasts, chondrocytes, and adipocytes [10,16,17,18].
In addition to BSCs, various types of stem cells, including MSC-like (Mesenchymal stromal cells)-like cells, have been identified in colostrum and milk across different species [7,10,12,16,19,20,21,22,23,24,25,26]. Patki et al. [10] confirmed the mesenchymal characteristics of human breast milk-derived stem cells, which exhibit dual properties by being positive for both the epithelial marker E-Cadherin and the mesenchymal marker SMA (Smooth Muscle Actin), supporting the hypothesis that these cells may be of myoepithelial origin [10]. Moreover, these human breast milk-derived MSCs have been shown to possess the ability to differentiate into various human cell types, underscoring their potential applications in regenerative medicine [10].
These findings pave the way for further research into the use of milk and colostrum-derived stem cells as a non-invasive, accessible source for therapeutic applications in regenerative medicine, including in veterinary medicine. In vivo, these stem cells contribute not only to the structural remodeling of the mammary gland and the development of neonatal tissues but also play a role in epigenetic regulation in the offspring [26]. Studies in rats and mice demonstrated that maternal stem cells can survive in the gastrointestinal tract, enter the bloodstream, and integrate into various neonatal organs, where they differentiate into functional cells in a process known as microchimerism [27,28]. This process, which begins in utero and continues through breastfeeding, suggests that milk- and colostrum-derived stem cells not only support neonatal development but may provide long-term immune benefits and foster unique biological tolerance between mother and offspring [12,29].
Among stem cell types, MSCs are widely used for cell therapy due to their advantages, including self-renewal, multi-lineage differentiation, low risk of teratoma formation, and low immunogenicity [30,31]. MSCs can be derived from various tissues, such as bone marrow [32], adipose tissue [33], Wharton’s jelly [34], placenta [35], amniotic membrane [36], and amniotic fluid [37]. In 2006, the International Society for Cellular Therapy (ISCT) established the "golden criteria" for MSCs characterization. These criteria include fibroblast-like plastic-adherent morphology, high expression of mesenchymal biomarkers (CD73, CD90, CD105), minimal expression of hematopoietic or immune biomarkers (e.g., CD31, CD34, CD45, HLA-DR), and the ability to differentiate into adipocytes, osteoblasts, and chondrocytes [38]. Moreover, in response to stimuli, MSCs secrete a plethora of bioactive molecules with anti-inflammatory [39,40], immunomodulatory [39,41,42,43], and other therapeutic effects, such as inhibiting cell death, reducing fibrosis, promoting vascularization, and exhibiting antimicrobial properties [42,44,45].
Over the past two decades, studies have extensively characterized human milk-derived stem cells [5,6,7,10,14,16,18,22,23,46,47,48,49,50,51,52,53] and their therapeutic potential across various pathological conditions [54,55,56,57,58,59].
In veterinary species, studies have primarily focused on isolating and characterizing MSC-like cells from bovine, swine, and rabbit milk [19,60,61], highlighting their regenerative, pro-angiogenic [62], and antimicrobial [61] properties with potential applications in neonatal health and veterinary medicine.
However, research on equine mammary and milk-derived stem cells remains limited. To date, only one study has successfully isolated and characterized mammary stem cells (MaSCs) from equine mammary gland tissue [63]. Subsequent investigations have explored the role of extracellular vesicles [64] and microRNAs [65] in MaSC regulation, the development of equine mammary organoids for comparative research [66], and species-specific mechanisms of mammary cancer resistance [67].
Nevertheless, the presence and therapeutic potential of MSCs in equine colostrum remain largely unexplored. This study aims to isolate and characterize MSCs for the first time from equine colostrum collected immediately after delivery, laying the groundwork for future applications in veterinary regenerative medicine.
2. Materials and Methods
2.1. Materials
Chemicals were obtained from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). FBS (Fetal Bovine Serum) is branded GIBCO™ (Thermo Fisher Scientific, Waltham, MA, USA). Plastics were from Falcon™ (Corning Inc., NY, USA) and SarstedtTM (Sarstedt AG & Co. KG, Nümbrecht, Germany) unless otherwise stated.
2.2. Samples
Colostrum samples (n=6) were collected from mares hospitalized at the Equine Perinatology and Reproduction Unit of the Equine Clinical Service, Department of Veterinary Medical Sciences, University of Bologna (Italy), for delivery management. Samples were aseptically collected into sterile 15 mL tubes immediately after foaling. Colostrum quality, indicative of its IgG concentration [68], was assessed using a refractometer equipped with a Brix scale [68,69]. The portion of each sample not utilized for quality assessment was allocated for cell isolation. It was stored at +4°C for a maximum of 12 hours before being transported to the Laboratory of Animal Reproduction and Biotechnologies (Equine Clinical Service, Department of Veterinary Medical Sciences, University of Bologna, Italy). All procedures were conducted in compliance with the guidelines approved by the Animal Welfare Committee (CoBA) of the University of Bologna (protocol no. 4630/23).
2.3. Equine Colostrum-Derived MSCs Isolation and Culture
Equine colostrum-derived MSCs (C-MSCs) were isolated as previously described by Widjaja et al. [70] with some modifications. Briefly, colostrum was mixed and diluted 1:1 with Dulbecco’s polyphosphate-buffered saline (DPBS) plus antibiotics (100 IU/mL penicillin, 100 g/mL streptomycin) and antimycotics (2.5 mg/L Amphotericin B) in 50 mL conical tube and centrifuged at 1881×g for 20 min at 20°C. After centrifugation, the supernatant and the fat layer were discarded and the cell pellet was re-suspended in 2 mL DPBS, then transferred into a sterile 15 mL conical tube. The cell pellet was re-suspended and centrifuged at 1881×g for 10 min at 20°C. This operation was repeated twice. Finally, the cellular pellet was re-suspended in complete culture medium (DMEM F-12 + 10% FBS + 100 IU/mL penicillin + 100 g/mL streptomycin). Cells were plated into 25 cm2 culture flasks and incubated in 5% CO2 at 38.5°C, in a humidified atmosphere (Passage 0). After 24 h, the culture medium was completely replaced, and non-adherent cells were removed. The culture medium was changed every 3 days until reaching a confluence of 80-90%. At this time, cells were dissociated using a 0.25% Trypsin-EDTA (Ethylenediaminetetraacetic acid) solution, counted using a Neubauer improved chamber, and cryopreserved as described by Merlo et al. [71]. Briefly, cells in 0.5 mL of FBS were put in a 1.5 mL cryogenic tube at 4°C. After 10 min, the cell suspension was diluted 1:1 with FBS + 16% DMSO (Dimethyl sulfoxide, final concentration 8% DMSO) and maintained for a further 10 min at 4°C. Then the cryogenic tube was set to −80°C for 24 h in Mr. Frosty (Nalgene) and finally stored in liquid nitrogen.
Equine C-MSCs were thawed at 37°C in 20 mL DMEM + 10% FBS, then centrifuged at 470×g at 25°C for 10 min. The pellet was re-suspended in 1 mL of complete culture medium and cell concentration and viability were evaluated by staining cells with Trypan blue (0.4%) solution. Cells were seeded in a 25 cm2 flask (5000 cells/cm2) as “Passage 1” (P1). Third-passage (P3) cells were used for in vitro tests.
2.4. Characterization of Colostrum-Derived MSCs
When the confluence of 80–90% at P2 was reached, cells from all six colostrum samples were detached from the flask, and viability and concentrations were determined as described above. The cells were subsequently seeded to evaluate their growth rate through Population Doubling Time (PDT) analysis and for characterization via the colony-forming unit (CFU) assay, adhesion and migration assay, and tri-lineage in vitro differentiation. Furthermore, CD marker expression was evaluated by RT-PCR. For all samples, each test was carried out in three replicates.
2.4.1. Population Doubling Time (PDT) Analysis
To determine growth rate, C-MSCs were seeded in 6-well plate at a cell density of 5000 cells/cm2. Cell number was assessed every day for 7 days in triplicate. The mean number of cells was calculated and plotted on a semi-log curve against culture time to generate a growth curve. The population doubling time (PDT) was calculated from the log phase of the growth curve using the equation described by Bezerra et al. (2022) [72]:
where t represents the period of cultivation in days, N0 represents the initial number of cells, and Nt represents the number of cells in a specific period of culture.
PDT=(t×log2) / log (Nt / N0)
2.4.2. CFU (Colony-Forming Unit) Assay
For determining the ability of cells to form colonies, 1 × 102 cells were cultured for 4 weeks in 6-well plates. Colonies were then fixed in 4% paraformaldehyde at RT for 1 h and stained with Giemsa 0.1% stain (30 min). Using an inverted light microscope (Eclipse TE 2000-U, Nikon Instruments Inc., Tokyo, Japan), the operator counted colonies formed by at least 16–20 nucleate cells.
2.4.3. Adhesion and Migration Assays
To determine whether C-MSCs preserved their adhesion capability, a spheroid formation assay was performed. Differently from the cell-substratum adhesion, performed on monolayer cultures adherent to rigid substrates, this test gives information about the direct cell-cell adhesion architecture found in normal tissues. Cells were cultured through the ‘hanging-drop’ method in Corning spheroid microplates (96-well, black/clear bottom round, Ultra-Low Attachment surface, Corning Inc., NY, USA, 5000 cells/25 µL drop). Bright-field images were acquired by a CCD camera (DS-Fi2, Nikon, Tokyo, Japan) mounted on an inverted light microscope (Eclipse TE 2000-U, Nikon Instruments Inc., Tokyo, Japan) using 4× magnification every 24 h from the seeding to the complete spheroid formation after 96 h. Starting from the binary masks obtained by Fiji software (ImageJ, v. 1.8.0_345/1.54g), the volume of each spheroid was computed using ReViSP (sourceforge.net/projects/revisp/, v. 2.3) [73], a software specifically designed to accurately estimate the volume of spheroids and to render an image of their 3D surface.
To assess cell migration potential, a scratch assay (or wound-healing assay) was carried out, as previously described by Liang et al. [74]. Briefly, P3 cells were seeded (5000 cells/cm2) in 6-well plates. At 80 to 90% confluence the cell monolayer was scraped using a 200 μL pipette tip. The cells were then washed twice with DPBS to remove any detached cells, and the plate was incubated under standard culture conditions. Images were captured at the same two locations of each well, both immediately after the tip-scratch (time 0 = t0) and every 24 h over a 96 h period (0, 24, 48, 72, 96 h). The distances of each scratch closure were measured by Fiji software (ImageJ, v. 1.8.0_345/1.54g), and the migration percentages (Wound Closure %) were calculated using the following formula [75]:
where tn corresponds to the distance at each specific time point analyzed.
[(distance at t0−distance at tn *100]/distance at t0
2.4.4. Multi-Lineage In Vitro Differentiation
In vitro differentiation potential toward osteogenic, adipogenic, and chondrogenic lineages of C-MSCs was also evaluated. Cells were seeded in 24-well plates at a density of 5000 cells/cm2 and cultured until subconfluency. Then, the culture medium was replaced, and cells were cultured for 3 weeks under specific induction media (Table 1) [76]. As a negative control, an equal number of cells was cultured in expansion medium. The specific induction media and expansion medium were replaced twice a week. To assess differentiation, cells were fixed with 4% paraformaldehyde at room temperature (RT) for 1 h. Differentiation was visualized using Oil Red O for adipogenic vacuoles, Alcian Blue for sulfated proteoglycans, and Alizarin Red S for calcium deposits. Bright-field images were captured using a CCD camera (DS-Fi2, Nikon, Tokyo, Japan) mounted on an inverted light microscope (Eclipse TE 2000-U, Nikon Instruments Inc., Tokyo, Japan) at 10× magnification.
Quantification of positively stained areas was performed using Fiji software (ImageJ, v. 1.8.0_345/1.54g) following the methodology described by Heyman et al. (2022) [77]. The "ColourDeconvolution2" plugin was applied to separate the stain and background channels, which were then converted to 8-bit grayscale images and binarized using auto-thresholding. The positively stained areas were measured, and the differentiation ratio (DR) was calculated as the ratio between the areas of the stain-positive signal and the background.
To compare differentiation across samples, a normalized DR was determined for each sample by dividing the mean DR of the differentiated wells by the mean DR of the non-induced controls.
2.4.5. Molecular Characterization
For the molecular characterization of C-MSCs, total RNA was extracted from snap-frozen cells using the NucleoSpin® RNA Kit (Macherey-Nagel, Düren, Germany) following the manufacturer’s protocol. Residual genomic DNA was removed by treating the RNA with Amplification-Grade DNase I (Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA). Complementary DNA (cDNA) was synthesized from the purified RNA using the RevertAid Reverse Transcriptase Kit (Thermo Fisher Scientific, Waltham, MA, USA).
The resulting cDNA was directly subjected to PCR using the 2× EpiArt™ HS Taq Master Mix (Vazyme Biotech Co., Ltd., Nanjing, China). The expression of genes encoding mesenchymal stromal cell markers (CD90, CD73), hematopoietic markers (CD45, CD34), and major histocompatibility complex markers (MHC-I, MHC-II) was analyzed. To ensure the proper expression of samples, GAPDH was used as a housekeeping gene. The specific primers employed for amplification are listed in Table 2 [78,79,80].
PCR products were resolved on a 2% agarose gel stained with ethidium bromide. Electrophoresis was conducted to verify the expected band sizes, and gels were visualized and photographed under ultraviolet light using a UVP PhotoDoc-It Imaging System (UVP, LLC, Upland, CA, USA). The presence of discrete bands corresponding to the expected sizes confirmed positive expression, while the absence of bands indicated a negative result. A 50 bp DNA Ladder (Thermo Fisher Scientific, Waltham, MA, USA) was used as a molecular size marker.
2.4.6. Statistical Analysis
Data are reported as mean ± standard deviation (SD). Statistical analyses and graph generation were performed using GraphPad Prism software (version 9.0.0). Data were analyzed, for normal distribution, using a Shapiro–Wilk test. Population doubling times (PDTs) and colony numbers were analyzed using one-way ANOVA, followed by Tukey’s post-hoc test for pairwise comparisons. Proliferation rates and wound closure percentages were evaluated with two-way ANOVA, followed by Tukey’s post-hoc test for comparisons between groups. Volumes of 3D spheroids at 96 hours and differentiation ratio (DRs) were analyzed using the Kruskal-Wallis test, followed by Dunn's multiple comparisons test. Results were considered significant when the probability of their occurrence due to chance alone was less than 5% (p < 0.05).
3. Results
3.1. Isolation and Morphological Characterization of Equine Colostrum-Derived Cell
Equine colostrum cells (n=6; ~20 mL/sample) were successfully isolated following the protocol detailed in the Materials and Methods section. Isolation was based solely on the cells' ability to adhere to plastic. Phase-contrast microscopy (Figure 1) revealed a morphologically heterogeneous population, comprising cells with a round/polyhedral, epithelial-like appearance, alongside spindle-shaped cells characteristic of Mesenchymal Stromal cells (MSCs). Cell colonies became visible within the first 24 hours and reached confluence in 10.17 ± 1.17 days, with a cell yield ranging from 10.4 to 36.4 × 103 cells/mL of colostrum.
A detailed summary of the colostrum samples, including volume, Brix Index at collection, number of cells isolated and time required to reach confluence, is reported in Table 3.
Undifferentiated cells were cultured up to passage three (P3) across all samples, with no observable changes in morphology during the culture period or after cryopreservation and thawing. Post-thaw viability, determined by Trypan Blue stain, at P3 exceeded 70%, while the initial viability was over 95%.
3.2. Characterization of Colostrum-Derived MSCs
3.4.1. Population Doubling Time (PDT) Analysis
To assess the growth efficiency and proliferation rate of successfully established colostrum-derived MSC (C-MSC) cultures, population doubling times (PDTs) were calculated. The semi-logarithmic growth curves (Figure 2) revealed variability in the growth patterns across the samples. Three samples (4-5-6) exhibited a brief lag phase of 1-2 days, followed by a significant and linear increase in cell concentration, while three samples demonstrated prolonged lag phases (1-2) or a slower overall growth rate (3). The mean PDT values, which confirm the variability between samples, are summarized in Table 3 and illustrated in Figure 2. One sample (3) exhibited the longest PDT (6.21± 1.84 days), indicating reduced proliferative efficiency, which was significantly different from the mean PDT of the other sample (2 ± 0.7 days, p < 0.0001; Figure 2). In contrast, 2/6 samples (2-4) showed the shortest PDTs, reflecting a higher proliferation rate (1.42 ± 0.28 and 1.22 ± 0.08, respectively). Overall, the sample effect on both growth rate and PDT was highly significant (p < 0.0001).
3.4.2. CFU (Colony-Forming Unit) Assay
The Colony Forming Unit (CFU) assay was performed to evaluate the self-renewal capacity of C-MSCs. At P3, MSCs from all six samples were seeded in 6-well plates at a density of 1 × 10² cells/well and cultured for 4 weeks. The resulting colonies were categorized as "dispersed," characterized by a vague macroscopic appearance and scattered cells under the microscope, as opposed to dense, compact colonies with the typical "fingerprint" pattern (Figure 3) [81]. Colonies were counted in triplicate, and the average number of colonies formed is summarized in Table 3 and presented in Figure 3, revealing significant sample-related variability (p < 0.0001). Two samples (2-4) demonstrated the highest clonogenic potential, forming significantly more CFUs than the other samples (71 ± 16.7 and 59.3 ± 7.37, respectively; p < 0.0001). Conversely, one sample (6) exhibited the lowest CFU count (13 ± 3.6), showing a statistically significant difference compared to samples 2, 4, and 5 (p < 0.0001, Figure 3). No significant differences were detected among samples 1, 3, and 5 (21 ± 2, 34.7 ± 11, and 40.3 ± 5.5, respectively; Figure 3).
3.4.3. Adhesion and Migration Assays
To assess the cell-cell adhesion capacity of C-MSCs, at P3 of in vitro culture, cells were cultured using the hanging-drop method in ultra-low attachment microplates (5000 cells/25 µL drop). Except for samples 1 and 3, all tested MSC populations demonstrated the ability to form three-dimensional spheroid-like structures. Spheroid formation was evident after 24 hours, and by the fourth day of incubation, most C-MSCs had developed stable, compact, and rigid spheroids with a spherical morphology (Figure 4).
In contrast, samples 1 and 3 failed to generate spheroids, instead forming loosely compact cellular sheets that remained easily dissociable, suggesting a lack of spheroid-forming capacity (Figure 4). Bright-field images were acquired at multiple time points (24, 48, 72, and 96 hours) and analyzed using ReViSP, an open-source voxel-based 3D image analysis tool capable of estimating spheroid volume starting from binary masks and rendering a three-dimensional surface representation (Figure 4).
Quantitative analysis of spheroid volume revealed dynamic structural changes over time, characterized by progressive compaction and a corresponding reduction in volume between 48 and 96 hours. By the end of the incubation period (4 days), spheroid volumes had stabilized, with no significant differences detected between the samples.
Given the growing interest in the role of MSCs in tissue repair [82], we aimed to evaluate their migratory capacity using the scratch assay, a highly quantifiable, adaptable, and cost-effective method. The percentage of wound closure was assessed at different time points (24, 48, 72, and 96 hours) and plotted against time, as shown in Figure 5.
Quantitative analysis revealed significant differences in wound closure percentages among the groups (Two-way ANOVA, p < 0.0001). Notably, sample 2 demonstrated the highest migratory capacity, achieving complete wound closure (100%) at 72 hours, significantly outperforming all other groups. In contrast, samples 1, 4, 5, and 6 exhibited a more gradual increase in wound closure over time, without exceeding approximately 30% at 96 h (Figure 5).
Inter-group comparisons at individual time points revealed intermediate wound closure dynamics for sample 3. Although its performance showed modest but non-significant improvements over the slower-migrating groups (1, 4, 5, and 6), it remained significantly lower than sample 2 (p < 0.001, Figure 5).
3.4.4. Multi-Lineage In Vitro Differentiation
According to the criteria established by the International Society for Cell & Gene Therapy (ISCT) [38], at P3 of in vitro culture, C-MSCs were evaluated for their potential to differentiate into osteogenic, chondrogenic and adipogenic lineages under specific induction conditions.
After 3 weeks of adipogenic induction, all isolated cells exhibited adipogenic differentiation, as demonstrated by a morphological transition from spindle-shaped to round or oval-shaped cells. This was accompanied by the presence of intracytoplasmic lipid droplets, confirmed by positive staining with Oil Red O (Figure 6). In contrast, control MSCs maintained their spindle-shaped morphology, forming a monolayer without staining with Oil Red O, confirming the absence of adipogenic differentiation (Figure 6).
Chondrogenic differentiation was observed in C-MSCs from all samples after 21 days of culture in a chondrogenic medium. This was marked by a morphological change from spindle-shaped cells to larger, round cell aggregates, along with the secretion of an extracellular matrix enriched in sulfated proteoglycans, a defining trait of cartilage tissue. Positive Alcian Blue staining (Figure 6) confirmed the presence of sulfated proteoglycans, while no staining was detected in the control wells (Figure 6), indicating a lack of chondrogenic differentiation.
Following osteogenic induction, C-MSCs from all samples exhibited matrix calcium deposition, as demonstrated by positive Alizarin Red S staining, a key marker of osteogenic differentiation (Figure 6). No calcium deposition was observed in the control groups (Figure 6).
Quantification of positively stained areas using ImageJ software showed no statistically significant differences in adipogenic, chondrogenic, or osteogenic differentiation capacities between the groups (p > 0.005).
3.4.5. Molecular Characterization
Following the guidelines of the International Society for Cell & Gene Therapy (ISCT) [38], the molecular characterization of C-MSCs was performed by analyzing the expression of genes encoding MSC markers (CD90, CD73), hematopoietic markers (CD45, CD34), and major histocompatibility complex markers (MHC-I, MHC-II). To ensure the proper expression of samples, GAPDH was used as a housekeeping gene (Figure 7). The PCR results are summarized in Table 4. Qualitative PCR analysis demonstrated that MSCs isolated from all six colostrum samples expressed the MSC markers CD90 and CD73, while hematopoietic markers CD34 and CD45 were absent at P3 of in vitro culture (Figure 7). Additionally, MHC-I expression was weakly positive in all samples except for sample 1, where it was absent, while MHC-II expression was consistently negative across all samples (Figure 7).
4. Discussion
The expanding field of regenerative medicine has intensified the search for innovative and advantageous stem cell sources. Recent studies have demonstrated that milk, including colostrum, harbors a heterogeneous population of cells, including stem and progenitor cells with significant regenerative potential [5,6,7,10,14,16,18,22,23,46,47,48,49,50,51,52,53].
While interest in the therapeutic applications of milk- and colostrum-derived stem cells in veterinary medicine is increasing, little is known about the presence and characteristics of mesenchymal stem/stromal cells in equine colostrum.
This study aimed to isolate and characterize equine colostrum-derived MSCs (C-MSCs) to validate colostrum as a promising, non-invasive stem cell source. Building on our previous findings on fetal- and adipose-derived MSCs [83,84], we sought to compare C-MSCs with these well-characterized sources, assessing their potential for veterinary and translational medicine.
Our findings confirmed the successful isolation of C-MSCs, with a cell yield ranging from 10.4 to 36.4 × 10³ cells/mL. These cells exhibited plastic adherence and a fibroblast-like morphology, consistent with prior reports on MSCs isolated from human breast milk [47,85]. The presence of a heterogeneous cell population in colostrum, including both fibroblast-like and round-shaped cells, aligns with studies on bovine and human milk-derived MSCs, where an epithelial-to-mesenchymal transition (EMT) has been observed during culture [10,47,60].
As reported in studies on human breast milk MSCs, the lactation stage significantly influences stem cell quantity and viability, with a higher concentration found in early lactation[6,47,48]. This variation may explain discrepancies in isolation success across studies. Like findings by Fan et al. [7], who encountered difficulties in culturing breast milk-derived MSCs from later lactation stages, our results suggest that the early postpartum period is optimal for MSC isolation.
MSC characterization was performed at passage three, when the culture reached a homogeneous population, in accordance with previous studies [86,87,88]. Population doubling time (PDT) is a key parameter for assessing MSC proliferative potential in therapeutic applications [89,90], exhibited notable inter-sample variability. While some C-MSC samples exhibited a rapid expansion, others displayed a slower proliferation rate, with an average PDT of 2.7 ± 0.6 days. As previously reported, PDT is highly dependent on the tissue of origin and culture conditions [91,92]. Our findings align with those observed before for Wharton’s jelly MSCs (WJ-MSCs, 2.7 ± 0.9 days) [83], while C-MSCs proliferated faster than bone marrow MSCs (BM-MSCs, 3.6 ± 1.4 days) [83] but slower than amniotic fluid MSCs (AF-MSCs, 27.13 ± 2.6 hours) [91], adipose tissue MSCs (AT-MSCs, 2.2 ± 1.1 days), and umbilical cord blood MSCs (UCB-MSCs, 2.4 ± 1.3 days) [83] . These results emphasize the variability in MSC proliferation dynamics based on tissue source and culture conditions, even when carried out in the same laboratory.
The colony-forming unit (CFU) assay confirmed the self-renewal capacity of C-MSCs. Our results contrast with those reported by Fan et al.; indeed, authors failed to obtain CFU formation from human breast milk MSCs, possibly due to differences in lactation stage, culture conditions, or intrinsic biological differences between species [7]. The ability of C-MSCs to form colonies suggests a robust self-renewal potential, comparable to that of MSCs derived from equine fetal annexes and other postnatal sources [93]. Further comparative studies are required to elucidate the specific factors influencing CFU efficiency across different species and MSC sources.
Spheroid formation was assessed using the hanging-drop method, with most samples forming compact spheroids indicative of strong cell-cell adhesion properties. Notably, smaller spheroids correlated with increased adhesion ability, a characteristic previously described in fetal-derived MSCs, such as WJ-MSCs, compared to adult tissue-derived MSCs [83]. This suggests that C-MSCs share adhesion properties similar to those observed in fetal MSCs.
The migratory potential of MSCs is a key characteristic that influences their capacity for systemic application and integration into host tissues during therapeutic interventions [30]. The scratch assay demonstrated efficient C-MSC migration, achieving approximately 30% wound closure at 96 hours. Comparative studies in our laboratory showed similar migration efficiencies among MSCs from various sources, including amniotic membrane (AM-MSCs, 34.14 ± 4.51%) and Wharton’s jelly (WJ-MSCs, 38.20 ± 2.88%) [84]. Comparable migration rates were observed among MSCs from WJ, UCB, BM, and AT, though AT-MSCs exhibited notable inter-sample variability [83]. These findings indicate that C-MSCs possess migration capabilities like fetal annex-derived MSCs, reinforcing their regenerative potential.
C-MSCs exhibited a multipotent differentiation capacity, successfully differentiating into osteogenic, chondrogenic, and adipogenic lineages, as confirmed by specific staining performed after 21 days of culture in the appropriate differentiation medium. These results align with prior studies on MSCs from human breast milk [10,16] and bovine milk [60], which demonstrated similar tri-lineage differentiation potential. The mesodermal differentiation ability of C-MSCs further validates their identity as MSCs, in accordance with the criteria established by the International Society for Cellular Therapy (ISCT) [38]. Notably, unlike MSCs from adult equine tissues [94] and our previous findings on equine AF-MSCs, UCB-MSCs, and WJ-MSCs [87], C-MSCs displayed minimal donor-to-donor variability.
To confirm the mesenchymal identity of equine C-MSCs, we evaluated MSC surface marker expression. These cells expressed CD90 and CD73 while lacking hematopoietic markers CD34 and CD45, consistent with findings on MSCs from AF, CB, and WJ [87]. Similar expression patterns have been reported in MSCs from milk across species, including rabbits, bovines, and humans, with high levels of CD29, CD166, CD44, and CD105, and an absence of CD45 and CD34 [10,19,22,47,59,60,95].
A key feature of MSCs is their immunomodulatory potential, which is largely influenced by the expression of major histocompatibility complex (MHC) molecules. In our study, MHC-I expression varied among samples, with some exhibiting weak expression and one lacking it entirely. Previous studies on equine BM-MSCs have shown MHC-I expression with heterogeneous MHC-II expression [96,97]. In horses, MSCs that express MHC-I but not MHC-II have been shown to avoid triggering T-cell proliferation [97]. The absence of MHC-II expression is therefore a crucial factor for potential allogeneic applications, as it enables MSCs to evade immune rejection [97,98]. Our findings confirm that equine C-MSCs did not express MHC-II, in agreement with our previous studies on equine WJ- and AT-MSCs [84] and recent findings on AF-MSCs [91].
The successful isolation and characterization of equine C-MSCs highlight their potential as a novel, non-invasive source of regenerative cells for veterinary medicine. Their ease of collection, multipotency, and immunomodulatory properties position them as strong candidates for equine regenerative therapies, with possible translational implications for human medicine. However, the variability observed among samples from different donors necessitates validation with a larger sample size. Future studies should also investigate their in vivo therapeutic potential and interactions with host tissues.
5. Conclusions
This study provides the first comprehensive characterization of mesenchymal stromal cells isolated from equine colostrum (C-MSCs), confirming their mesenchymal phenotype, self-renewal ability, migration potential, and multipotency. These findings pave the way for further investigations into their applications in veterinary regenerative medicine, with promising translational implications for human therapies.
Author Contributions
Conceptualization, E.I. and B.M.; methodology, E.I.; validation, E.I. and A.C.; formal analysis, A.C.; investigation, A.C. and F.B.; resources, E.I. and B.M.; data curation, A.C.; writing—original draft preparation, A.C.; writing—review and editing, E.I.; supervision, E.I and B.M.; project administration, E.I.; funding acquisition, E.I. All authors have read and agreed to the published version of the manuscript.
Funding
Angelita Capone PhD fellowship is funded by the Italian Ministry of University and Research (MUR) under the National Recovery and Resilience Plan (NRRP, ex D.M. 118/23).
Institutional Review Board Statement
All sample collection procedures were conducted in compliance with the guidelines approved by the University of Bologna's Animal Welfare Committee (CoBA) (protocol no. 4630/23).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are contained within this article.
Acknowledgments
The authors thank all the technical staff and veterinarians of the Equine Perinatology and Reproduction Unit of the Equine Clinical Service, Department of Veterinary Medical Sciences, University of Bologna, for their assistance in providing access to the samples.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- McCue, P.M. Colostrum Banking. Equine Reproductive Procedures 2014, 299–301. [CrossRef]
- de Lima, T.C.; de Sobral, G.G.; de França Queiroz, A.E.S.; Chinelate, G.C.B.; Porto, T.S.; Oliveira, J.T.C.; Carneiro, G.F. Characterization of Lyophilized Equine Colostrum. J Equine Vet Sci 2024, 132, 104975. [CrossRef]
- Perkins, G.A.; Goodman, L.B.; Wimer, C.; Freer, H.; Babasyan, S.; Wagner, B. Maternal T-Lymphocytes in Equine Colostrum Express a Primarily Inflammatory Phenotype. Vet Immunol Immunopathol 2014, 161, 141–150. [CrossRef]
- Dukes, H.H.; Reece, W.O. Fisiologia Dos Animais Domésticos; Guanabara Koogan: Rio de Janeiro, 2019;
- Pichiri, G.; Lanzano, D.; Piras, M.; Dessì, A.; Reali, A.; Puddu, M.; Noto, A.; Fanos, V.; Coni, C.; Faa, G.; et al. Human Breast Milk Stem Cells: A New Challenge for Perinatologists. JOURNAL OF PEDIATRIC AND NEONATAL INDIVIDUALIZED MEDICINE 2016, 5, e050120-1. [CrossRef]
- Indumathi, S.; Dhanasekaran, M.; Rajkumar, J.S.; Sudarsanam, D. Exploring the Stem Cell and Non-Stem Cell Constituents of Human Breast Milk. Cytotechnology 2013, 65, 385–393. [CrossRef]
- Fan, Y.; Chong, Y.S.; Choolani, M.A.; Cregan, M.D.; Chan, J.K.Y. Unravelling the Mystery of Stem/Progenitor Cells in Human Breast Milk. PLoS One 2010, 5, e14421. [CrossRef]
- Visvader, J.E. Keeping Abreast of the Mammary Epithelial Hierarchy and Breast Tumorigenesis. Genes Dev 2009, 23, 2563–2577. [CrossRef]
- Visvader, J.E.; Lindeman, G.J. Mammary Stem Cells and Mammopoiesis. Cancer Res 2006, 66, 9798–9801. [CrossRef]
- Patki, S.; Kadam, S.; Chandra, V.; Bhonde, R. Human Breast Milk Is a Rich Source of Multipotent Mesenchymal Stem Cells. Hum Cell 2010, 23, 35–40. [CrossRef]
- Lochter, A. Plasticity of Mammary Epithelia during Normal Development and Neoplastic Progression. 2011, 76, 997–1008. [CrossRef]
- Hassiotou, F.; Geddes, D.T.; Hartmann, P.E. Cells in Human Milk: State of the Science. Journal of Human Lactation 2013, 29, 171–182. [CrossRef]
- Somasundaram, I.; Kaingade, P.; Bhonde, R. Breast Milk-Derived Mesenchymal Stem-Like Cells: History and Mystery. Stem cell and Non-stem Cell Components of Breast Milk 2023, 45–53. [CrossRef]
- Cregan, M.D.; Fan, Y.; Appelbee, A.; Brown, M.L.; Klopcic, B.; Koppen, J.; Mitoulas, L.R.; Piper, K.M.E.; Choolani, M.A.; Chong, Y.S.; et al. Identification of Nestin-Positive Putative Mammary Stem Cells in Human Breastmilk. Cell Tissue Res 2007, 329, 129–136. [CrossRef]
- Thomas, E.; Zeps, N.; Rigby, P.; Hartmann, P. Reactive Oxygen Species Initiate Luminal but Not Basal Cell Death in Cultured Human Mammary Alveolar Structures: A Potential Regulator of Involution. Cell Death & Disease 2011 2:8 2011, 2, e189–e189. [CrossRef]
- Hassiotou, F.; Beltran, A.; Chetwynd, E.; Stuebe, A.M.; Twigger, A.J.; Metzger, P.; Trengove, N.; Lai, C.L.; Filgueira, L.; Blancafort, P.; et al. Breastmilk Is a Novel Source of Stem Cells with Multilineage Differentiation Potential. Stem Cells 2012, 30, 2164–2174. [CrossRef]
- Twigger, A.J.; Hepworth, A.R.; Tat Lai, C.; Chetwynd, E.; Stuebe, A.M.; Blancafort, P.; Hartmann, P.E.; Geddes, D.T.; Kakulas, F. Gene Expression in Breastmilk Cells Is Associated with Maternal and Infant Characteristics. Scientific Reports 2015 5:1 2015, 5, 1–14. [CrossRef]
- Hosseini, S.M.; Talaei-Khozani, T.; Sani, M.; Owrangi, B. Differentiation of Human Breast-Milk Stem Cells to Neural Stem Cells and Neurons. Neurol Res Int 2014, 2014, 807896. [CrossRef]
- Abd Allah, S.H.; Shalaby, S.M.; El-Shal, A.S.; El Nabtety, S.M.; Khamis, T.; Abd El Rhman, S.A.; Ghareb, M.A.; Kelani, H.M. Breast Milk MSCs: An Explanation of Tissue Growth and Maturation of Offspring. IUBMB Life 2016, 68, 935–942. [CrossRef]
- Briere, C.E.; McGrath, J.M.; Jensen, T.; Matson, A.; Finck, C. Breast Milk Stem Cells. Advances in Neonatal Care 2016, 16, 410–419. [CrossRef]
- Hassiotou, F.; Hartmann, P.E. At the Dawn of a New Discovery: The Potential of Breast Milk Stem Cells. Advances in Nutrition 2014, 5, 770–778. [CrossRef]
- Kaingade, P.; Somasundaram, I.; Sharma, A.; Patel, D.; Marappagounder, D. Cellular Components, Including Stem-Like Cells, of Preterm Mother’s Mature Milk as Compared with Those in Her Colostrum: A Pilot Study. https://home.liebertpub.com/bfm 2017, 12, 446–449. [CrossRef]
- Kaingade, P.M.; Somasundaram, I.; Nikam, A.B.; Sarang, S.A.; Patel, J.S. Assessment of Growth Factors Secreted by Human Breastmilk Mesenchymal Stem Cells. https://home.liebertpub.com/bfm 2016, 11, 26–31. [CrossRef]
- Sam Vijay Kumar, J.; Rajkumar, H.; Nagasamy, S.; Ghose, S.; Subramanian, B.; Adithan, C. Human Colostrum Is a Rich Source of Cells with Stem Cell-Like Properties. SBV Journal of Basic, Clinical and Applied Health Science 2017, 1, 26–31. [CrossRef]
- Sani, M.; Hosseini, S.M.; Salmannejad, M.; Aleahmad, F.; Ebrahimi, S.; Jahanshahi, S.; Talaei-Khozani, T. Origins of the Breast Milk-Derived Cells; an Endeavor to Find the Cell Sources. Cell Biol Int 2015, 39, 611–618. [CrossRef]
- Witkowska-Zimny, M.; Kaminska-El-Hassan, E. Cells of Human Breast Milk. Cell Mol Biol Lett 2017, 22, 1–11. [CrossRef]
- Hassiotou, F.; Heath, B.; Ocal, O.; Filgueira, L.; Geddes, D.; Hartmann, P.; Wilkie, T. Breastmilk Stem Cell Transfer from Mother to Neonatal Organs (216.4). The FASEB Journal 2014, 28, 216.4. [CrossRef]
- Aydın, M.Ş.; Yiğit, E.N.; Vatandaşlar, E.; Erdoğan, E.; Öztürk, G. Transfer and Integration of Breast Milk Stem Cells to the Brain of Suckling Pups. Scientific Reports 2018 8:1 2018, 8, 1–9. [CrossRef]
- Barinaga, M. Cells Exchanged during Pregnancy Live On. Science (1979) 2002, 296, 2169–2172. [CrossRef]
- Hanson, L.Å.; Silfverdal, S.A.; Strömbäck, L.; Erling, V.; Zaman, S.; Olcén, P.; Telemo, E. The Immunological Role of Breast Feeding. Pediatr Allergy Immunol 2001, 12 Suppl 14, 15–19. [CrossRef]
- Hamosh, M. Bioactive Factors in Human Milk. Pediatr Clin North Am 2001, 48, 69–86. [CrossRef]
- Ranera, B.; Lyahyai, J.; Romero, A.; Vázquez, F.J.; Remacha, A.R.; Bernal, M.L.; Zaragoza, P.; Rodellar, C.; Martín-Burriel, I. Immunophenotype and Gene Expression Profiles of Cell Surface Markers of Mesenchymal Stem Cells Derived from Equine Bone Marrow and Adipose Tissue. Vet Immunol Immunopathol 2011, 144, 147–154. [CrossRef]
- Alipour, F.; Parham, A.; Mehrjerdi, H.K.; Dehghani, H. Equine Adipose-Derived Mesenchymal Stem Cells: Phenotype and Growth Characteristics, Gene Expression Profile and Differentiation Potentials. Cell Journal (Yakhteh) 2015, 16, 456. [CrossRef]
- Venugopal, P.; Balasubramanian, S.; Majumdar, A. Sen; Ta, M. Isolation, Characterization, and Gene Expression Analysis of Wharton’s Jelly-Derived Mesenchymal Stem Cells under Xeno-Free Culture Conditions. Stem Cells Cloning 2011, 4, 39–50. [CrossRef]
- In ’t Anker, P.S.; Scherjon, S.A.; Kleijburg-van der Keur, C.; de Groot-Swings, G.M.J.S.; Claas, F.H.J.; Fibbe, W.E.; Kanhai, H.H.H. Isolation of Mesenchymal Stem Cells of Fetal or Maternal Origin from Human Placenta. Stem Cells 2004, 22, 1338–1345. [CrossRef]
- Mihu, C.M.; Rus Ciucă, D.; Soritău, O.; Suşman, S.; Mihu, D. Isolation and Characterization of Mesenchymal Stem Cells from the Amniotic Membrane. Rom J Morphol Embryol 2009, 50, 73–77.
- In ’t Anker, P.S.; Scherjon, S.A.; Kleijburg-van der Keur, C.; Noort, W.A.; Claas, F.H.J.; Willemze, R.; Fibbe, W.E.; Kanhai, H.H.H. Amniotic Fluid as a Novel Source of Mesenchymal Stem Cells for Therapeutic Transplantation. Blood 2003, 102, 1548–1549. [CrossRef]
- Horwitz, E.M.; Le Blanc, K.; Dominici, M.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Deans, R.J.; Krause, D.S.; Keating, A. Clarification of the Nomenclature for MSC: The International Society for Cellular Therapy Position Statement. Cytotherapy 2005, 7, 393–395. [CrossRef]
- Le Blanc, K.; Davies, L.C. Mesenchymal Stromal Cells and the Innate Immune Response. Immunol Lett 2015, 168, 140–146. [CrossRef]
- Caplan, A.I. MSCs: The Sentinel and Safe-Guards of Injury. J Cell Physiol 2016, 231, 1413–1416. [CrossRef]
- Bernardo, M.E.; Fibbe, W.E. Mesenchymal Stromal Cells: Sensors and Switchers of Inflammation. Cell Stem Cell 2013, 13, 392–402. [CrossRef]
- Ferreira, J.R.; Teixeira, G.Q.; Santos, S.G.; Barbosa, M.A.; Almeida-Porada, G.; Gonçalves, R.M. Mesenchymal Stromal Cell Secretome: Influencing Therapeutic Potential by Cellular Pre-Conditioning. Front Immunol 2018, 9, 425936. [CrossRef]
- Serra, M.; Cunha, B.; Peixoto, C.; Gomes-Alves, P.; Alves, P.M. Advancing Manufacture of Human Mesenchymal Stem Cells Therapies: Technological Challenges in Cell Bioprocessing and Characterization. Curr Opin Chem Eng 2018, 22, 226–235. [CrossRef]
- Sutton, M.T.; Fletcher, D.; Ghosh, S.K.; Weinberg, A.; Van Heeckeren, R.; Kaur, S.; Sadeghi, Z.; Hijaz, A.; Reese, J.; Lazarus, H.M.; et al. Antimicrobial Properties of Mesenchymal Stem Cells: Therapeutic Potential for Cystic Fibrosis Infection, and Treatment. Stem Cells Int 2016, 2016, 5303048. [CrossRef]
- Vizoso, F.J.; Eiro, N.; Cid, S.; Schneider, J.; Perez-Fernandez, R. Mesenchymal Stem Cell Secretome: Toward Cell-Free Therapeutic Strategies in Regenerative Medicine. International Journal of Molecular Sciences 2017, Vol. 18, Page 1852 2017, 18, 1852. [CrossRef]
- Coni, P.; Piras, M.; Piludu, M.; Lachowicz, J.I.; Matteddu, A.; Coni, S.; Reali, A.; Fanos, V.; Jaremko, M.; Faa, G.; et al. Exploring Cell Surface Markers and Cell-Cell Interactions of Human Breast Milk Stem Cells. J Public Health Res 2023, 12. [CrossRef]
- Goudarzi, N.; Shabani, R.; Ebrahimi, M.; Baghestani, A.; Dehdashtian, E.; Vahabzadeh, G.; Soleimani, M.; Moradi, F.; Katebi, M. Comparative Phenotypic Characterization of Human Colostrum and Breast Milk-Derived Stem Cells. Hum Cell 2020, 33, 308–317. [CrossRef]
- Li, S.; Zhang, L.; Zhou, Q.; Jiang, S.; Yang, Y.; Cao, Y. Characterization of Stem Cells and Immune Cells in Preterm and Term Mother’s Milk. Journal of Human Lactation 2019, 35, 528–534. [CrossRef]
- Vijay Tukaram Mali, S.M.P. Cytology of the Human Milk in the First Post Partum Week - A Clinical Perspective. J Cytol Histol 2014, s4. [CrossRef]
- Sani, M.; Hosseini, S.M.; Salmannejad, M.; Aleahmad, F.; Ebrahimi, S.; Jahanshahi, S.; Talaei-Khozani, T. Origins of the Breast Milk-Derived Cells; an Endeavor to Find the Cell Sources. Cell Biol Int 2015, 39, 611–618. [CrossRef]
- Zaitsev, K. V.; Mezheritskii, S.A.; Stepanenko, N.P.; Gostyukhina, A.A.; Zhukova, O.B.; Kondrat’eva, E.I.; Stepanov, I.A.; Dzyuman, A.N.; Nikolaevskaya, E.E.; Vorob’ev, V.A.; et al. Immunological and Phenotypic Characterization of Cell Constituents of Breast Milk. Cell and Tissue Biology 2016 10:5 2016, 10, 410–415. [CrossRef]
- Twigger, A.J.; Engelbrecht, L.K.; Bach, K.; Schultz-Pernice, I.; Pensa, S.; Stenning, J.; Petricca, S.; Scheel, C.H.; Khaled, W.T. Transcriptional Changes in the Mammary Gland during Lactation Revealed by Single Cell Sequencing of Cells from Human Milk. Nat Commun 2022, 13. [CrossRef]
- Thomas, E.; Zeps, N.; Cregan, M.; Hartmann, P.; Martin, T. 14-3-3σ (Sigma) Regulates Proliferation and Differentiation of Multipotent P63-Positive Cells Isolated from Human Breastmilk. Cell Cycle 2011, 10, 278–284. [CrossRef]
- Nageeb, M.M.; Saadawy, S.F.; Attia, S.H. Breast Milk Mesenchymal Stem Cells Abate Cisplatin-Induced Cardiotoxicity in Adult Male Albino Rats via Modulating the AMPK Pathway. Sci Rep 2022, 12. [CrossRef]
- Pisano, C.; Galley, J.; Elbahrawy, M.; Wang, Y.; Farrell, A.; Brigstock, D.; Besner, G.E. Human Breast Milk-Derived Extracellular Vesicles in the Protection Against Experimental Necrotizing Enterocolitis. J Pediatr Surg 2020, 55, 54–58. [CrossRef]
- Borhani-Haghighi, M.; Navid, S.; Mohamadi, Y. The Therapeutic Potential of Conditioned Medium from Human Breast Milk Stem Cells in Treating Spinal Cord Injury. Asian Spine J 2020, 14. [CrossRef]
- Khamis, T.; Abdelalim, A.F.; Abdallah, S.H.; Saeed, A.A.; Edress, N.M.; Arisha, A.H. Breast Milk MSCs Transplantation Attenuates Male Diabetic Infertility Via Immunomodulatory Mechanism in Rats. Adv Anim Vet Sci 2019, 7, 145–153. [CrossRef]
- Khamis, T.; Abdelalim, A.F.; Saeed, A.A.; Edress, N.M.; Nafea, A.; Ebian, H.F.; Algendy, R.; Hendawy, D.M.; Arisha, A.H.; Abdallah, S.H. Breast Milk MSCs Upregulated β-Cells PDX1, Ngn3, and PCNA Expression via Remodeling ER Stress /Inflammatory /Apoptotic Signaling Pathways in Type 1 Diabetic Rats. Eur J Pharmacol 2021, 905. [CrossRef]
- Khamis, T.; Abdelalim, A.F.; Abdallah, S.H.; Saeed, A.A.; Edress, N.M.; Arisha, A.H. Early Intervention with Breast Milk Mesenchymal Stem Cells Attenuates the Development of Diabetic-Induced Testicular Dysfunction via Hypothalamic Kisspeptin/Kiss1r-GnRH/GnIH System in Male Rats. Biochim Biophys Acta Mol Basis Dis 2020, 1866. [CrossRef]
- Pipino, C.; Mandatori, D.; Buccella, F.; Lanuti, P.; Preziuso, A.; Castellani, F.; Grotta, L.; Di Tomo, P.; Marchetti, S.; Di Pietro, N.; et al. Identification and Characterization of a Stem Cell-Like Population in Bovine Milk: A Potential New Source for Regenerative Medicine in Veterinary. https://home.liebertpub.com/scd 2018, 27, 1587–1597. [CrossRef]
- Marquez-Curtis, L.A.; Dorobantu, L.S.; Sauvageau, D.; Elliott, J.A.W. Cryopreservation of Swine Colostrum-Derived Cells. Cryobiology 2020, 97, 168–178. [CrossRef]
- Danev, N.; Harman, R.M.; Oliveira, L.; Huntimer, L.; Van de Walle, G.R. Bovine Milk-Derived Cells Express Transcriptome Markers of Pluripotency and Secrete Bioactive Factors with Regenerative and Antimicrobial Activity. Scientific Reports 2023 13:1 2023, 13, 1–12. [CrossRef]
- Spaas, J.H.; Chiers, K.; Bussche, L.; Burvenich, C.; Van De Walle, G.R. Stem/Progenitor Cells in Non-Lactating Versus Lactating Equine Mammary Gland. https://home.liebertpub.com/scd 2012, 21, 3055–3067. [CrossRef]
- Bussche, L.; Rauner, G.; Antonyak, M.; Syracuse, B.; McDowell, M.; Brown, A.M.C.; Cerione, R.A.; Van De Walle, G.R. Microvesicle-Mediated Wnt/β-Catenin Signaling Promotes Interspecies Mammary Stem/Progenitor Cell Growth. J Biol Chem 2016, 291, 24390–24405. [CrossRef]
- Miller, J.L.; Kanke, M.; Rauner, G.; Bakhle, K.M.; Sethupathy, P.; Van de Walle, G.R. Comparative Analysis of MicroRNAs That Stratify in Vitro Mammary Stem and Progenitor Activity Reveals Functionality of Human MiR-92b-3p. J Mammary Gland Biol Neoplasia 2022, 27, 253–269. [CrossRef]
- Bartlett, A.P.; Harman, R.M.; Weiss, J.R.; Van de Walle, G. Establishment and Characterization of Equine Mammary Organoids Using a Method Translatable to Other Non-Traditional Model Species. Development 2022, 149. [CrossRef]
- Harman, R.M.; Das, S.P.; Kanke, M.; Sethupathy, P.; Van de Walle, G.R. MiRNA-214-3p Stimulates Carcinogen-Induced Mammary Epithelial Cell Apoptosis in Mammary Cancer-Resistant Species. Commun Biol 2023, 6. [CrossRef]
- Cash, R.S.G. Colostral Quality Determined by Refractometry. Equine Vet Educ 1999, 11, 36–38. [CrossRef]
- Magalhaes, H.B.; Canisso, I.F. Colostrum Conductivity, PH and Brix Index as Predictors of Passive Immunity Transfer in Foals. Equine Vet J 2024. [CrossRef]
- Widjaja, S.L.; Salimo, H.; Yulianto, I.; Soetrisno Proteomic Analysis of Hypoxia and Non-Hypoxia Secretome Mesenchymal Stem-like Cells from Human Breastmilk. Saudi J Biol Sci 2021, 28, 4399–4407. [CrossRef]
- Merlo, B.; Pirondi, S.; Iacono, E.; Rossi, B.; Ricci, F.; Mari, G. Viability, in Vitro Differentiation and Molecular Characterization of Equine Adipose Tissue-Derived Mesenchymal Stem Cells Cryopreserved in Serum and Serum-Free Medium. Cryoletters 2016, 37, 243–252.
- Bezerra, A.F.; Alves, J.P.M.; Fernandes, C.C.L.; Cavalcanti, C.M.; Silva, M.R.L.; Conde, A.J.H.; Tetaping, G.M.; Ferreira, A.C.A.; Melo, L.M.; Rodrigues, A.P.R.; et al. Dyslipidemia Induced by Lipid Diet in Late Gestation Donor Impact on Growth Kinetics and in Vitro Potential Differentiation of Umbilical Cord Wharton’s Jelly Mesenchymal Stem Cells in Goats. Vet Res Commun 2022, 46, 1259–1270. [CrossRef]
- Bellotti, C.; Duchi, S.; Bevilacqua, A.; Lucarelli, E.; Piccinini, F. Long Term Morphological Characterization of Mesenchymal Stromal Cells 3D Spheroids Built with a Rapid Method Based on Entry-Level Equipment. Cytotechnology 2016, 68, 2479–2490. [CrossRef]
- Liang, C.C.; Park, A.Y.; Guan, J.L. In Vitro Scratch Assay: A Convenient and Inexpensive Method for Analysis of Cell Migration in Vitro. Nature Protocols 2007 2:2 2007, 2, 329–333. [CrossRef]
- Rossi, B.; Merlo, B.; Colleoni, S.; Iacono, E.; Tazzari, P.L.; Ricci, F.; Lazzari, G.; Galli, C. Isolation and in Vitro Characterization of Bovine Amniotic Fluid Derived Stem Cells at Different Trimesters of Pregnancy. Stem Cell Rev Rep 2014, 10, 712–724. [CrossRef]
- Mizuno, H.; Hyakusoku, H. Mesengenic Potential and Future Clinical Perspective of Human Processed Lipoaspirate Cells. Journal of Nippon Medical School 2003, 70, 300–306. [CrossRef]
- Heyman, E.; Meeremans, M.; Devriendt, B.; Olenic, M.; Chiers, K.; De Schauwer, C. Validation of a Color Deconvolution Method to Quantify MSC Tri-Lineage Differentiation across Species. Front Vet Sci 2022, 9, 987045. [CrossRef]
- Mohanty, N.; Gulati, B.R.; Kumar, R.; Gera, S.; Kumar, P.; Somasundaram, R.K.; Kumar, S. Immunophenotypic Characterization and Tenogenic Differentiation of Mesenchymal Stromal Cells Isolated from Equine Umbilical Cord Blood. In Vitro Cell Dev Biol Anim 2014, 50, 538–548. [CrossRef]
- Corradetti, B.; Lange-Consiglio, A.; Barucca, M.; Cremonesi, F.; Bizzaro, D. Size-Sieved Subpopulations of Mesenchymal Stem Cells from Intervascular and Perivascular Equine Umbilical Cord Matrix. Cell Prolif 2011, 44, 330–342. [CrossRef]
- Desmarais, J.A.; Demers, S.P.; Suzuki, J.; Laflamme, S.; Vincent, P.; Laverty, S.; Smith, L.C. Trophoblast Stem Cell Marker Gene Expression in Inner Cell Mass-Derived Cells from Parthenogenetic Equine Embryos. Reproduction 2011, 141, 321–332. [CrossRef]
- Spaas, J.H.; Schauwer, C. De; Cornillie, P.; Meyer, E.; Soom, A. Van; Van de Walle, G.R. Culture and Characterisation of Equine Peripheral Blood Mesenchymal Stromal Cells. Vet J 2013, 195, 107–113. [CrossRef]
- Fu, X.; Liu, G.; Halim, A.; Ju, Y.; Luo, Q.; Song, G. Mesenchymal Stem Cell Migration and Tissue Repair. Cells 2019, 8. [CrossRef]
- Merlo, B.; Teti, G.; Lanci, A.; Burk, J.; Mazzotti, E.; Falconi, M.; Iacono, E. Comparison between Adult and Foetal Adnexa Derived Equine Post-Natal Mesenchymal Stem Cells. BMC Vet Res 2019, 15. [CrossRef]
- Iacono, E.; Pascucci, L.; Rossi, B.; Bazzucchi, C.; Lanci, A.; Ceccoli, M.; Merlo, B. Ultrastructural Characteristics and Immune Profile of Equine MSCs from Fetal Adnexa. Reproduction 2017, 154, 509–519. [CrossRef]
- Kumari, P.; Raval, A.; Rana, P.; Mahto, S.K. Regenerative Potential of Human Breast Milk: A Natural Reservoir of Nutrients, Bioactive Components and Stem Cells. Stem Cell Rev Rep 2023, 19, 1307–1327. [CrossRef]
- Ranera, B.; Ordovás, L.; Lyahyai, J.; Bernal, M.L.; Fernandes, F.; Remacha, A.R.; Romero, A.; Vázquez, F.J.; Osta, R.; Cons, C.; et al. Comparative Study of Equine Bone Marrow and Adipose Tissue-Derived Mesenchymal Stromal Cells. Equine Vet J 2012, 44, 33–42. [CrossRef]
- Iacono, E.; Brunori, L.; Pirrone, A.; Pagliaro, P.P.; Ricci, F.; Tazzari, P.L.; Merlo, B. Isolation, Characterization and Differentiation of Mesenchymal Stem Cells from Amniotic Fluid, Umbilical Cord Blood and Wharton’s Jelly in the Horse. Reproduction 2012, 143, 455–468. [CrossRef]
- Xie, L.; Zhang, N.; Marsano, A.; Vunjak-Novakovic, G.; Zhang, Y.; Lopez, M.J. In Vitro Mesenchymal Trilineage Differentiation and Extracellular Matrix Production by Adipose and Bone Marrow Derived Adult Equine Multipotent Stromal Cells on a Collagen Scaffold. Stem Cell Rev Rep 2013, 9, 858–872. [CrossRef]
- Bhat, S.; Viswanathan, P.; Chandanala, S.; Prasanna, S.J.; Seetharam, R.N. Expansion and Characterization of Bone Marrow Derived Human Mesenchymal Stromal Cells in Serum-Free Conditions. Scientific Reports 2021 11:1 2021, 11, 1–18. [CrossRef]
- Mehrian, M.; Lambrechts, T.; Marechal, M.; Luyten, F.P.; Papantoniou, I.; Geris, L. Predicting in Vitro Human Mesenchymal Stromal Cell Expansion Based on Individual Donor Characteristics Using Machine Learning. Cytotherapy 2020, 22, 82–90. [CrossRef]
- Kim, E.Y.; Lee, E.J.; Kim, R.E.; Kil, T.Y.; Kim, M.K. Evaluation of Stability and Safety of Equine Mesenchymal Stem Cells Derived from Amniotic Fluid for Clinical Application. Front Vet Sci 2024, 11, 1330009. [CrossRef]
- Schwarz, C.; Leicht, U.; Rothe, C.; Drosse, I.; Luibl, V.; Röcken, M.; Schieker, M. Effects of Different Media on Proliferation and Differentiation Capacity of Canine, Equine and Porcine Adipose Derived Stem Cells. Res Vet Sci 2012, 93, 457–462. [CrossRef]
- Lange-Consiglio, A.; Corradetti, B.; Meucci, A.; Perego, R.; Bizzaro, D.; Cremonesi, F. Characteristics of Equine Mesenchymal Stem Cells Derived from Amnion and Bone Marrow: In Vitro Proliferative and Multilineage Potential Assessment. Equine Vet J 2013, 45, 737–744. [CrossRef]
- Hillmann, A.; Ahrberg, A.B.; Brehm, W.; Heller, S.; Josten, C.; Paebst, F.; Burk, J. Comparative Characterization of Human and Equine Mesenchymal Stromal Cells: A Basis for Translational Studies in the Equine Model. Cell Transplant 2016, 25, 109–124. [CrossRef]
- Somasundaram, I.; Kaingade, P.; Bhonde, R. Applications of Breast Milk-Derived Cell Components: Present and Future Perspectives. Stem cell and Non-stem Cell Components of Breast Milk 2023, 71–77. [CrossRef]
- Kamm, J.L.; Parlane, N.A.; Riley, C.B.; Gee, E.K.; Dittmer, K.E.; McIlwraith, C.W. Blood Type and Breed-Associated Differences in Cell Marker Expression on Equine Bone Marrow-Derived Mesenchymal Stem Cells Including Major Histocompatibility Complex Class II Antigen Expression. PLoS One 2019, 14. [CrossRef]
- Schnabel, L. V.; Pezzanite, L.M.; Antczak, D.F.; Felippe, M.J.B.; Fortier, L.A. Equine Bone Marrow-Derived Mesenchymal Stromal Cells Are Heterogeneous in MHC Class II Expression and Capable of Inciting an Immune Response in Vitro. Stem Cell Res Ther 2014, 5. [CrossRef]
- Joel, M.D.M.; Yuan, J.; Wang, J.; Yan, Y.; Qian, H.; Zhang, X.; Xu, W.; Mao, F. MSC: Immunoregulatory Effects, Roles on Neutrophils and Evolving Clinical Potentials. Am J Transl Res 2019, 11, 3890.
Figure 1.
Monolayer of Equine Colostrum-derived MSCs; magnification 4×, 10×; Scale bar: 100 μm.

Figure 2.
Proliferative capacity of equine colostrum-derived MSCs, evaluated through semi-logarithmic growth curves (a) and Population Doubling Time (PDT) analysis (b). Data are presented as mean (a) and ad mean ± SD (b). Statistical analysis: One-way ANOVA with post-hoc Tukey’s test (b). Different letters indicate significant differences (p < 0.05) among samples.
Figure 2.
Proliferative capacity of equine colostrum-derived MSCs, evaluated through semi-logarithmic growth curves (a) and Population Doubling Time (PDT) analysis (b). Data are presented as mean (a) and ad mean ± SD (b). Statistical analysis: One-way ANOVA with post-hoc Tukey’s test (b). Different letters indicate significant differences (p < 0.05) among samples.
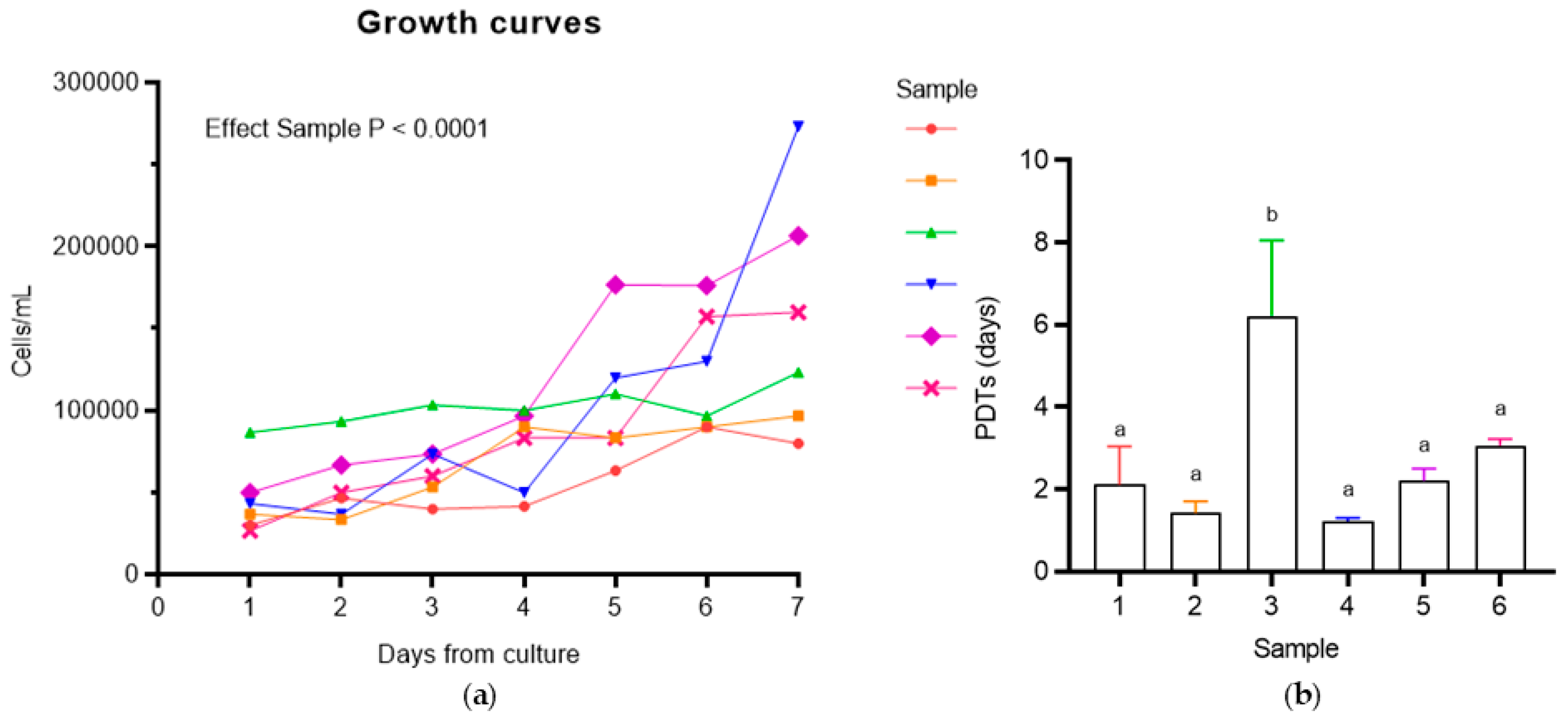
Figure 3.
Colony-forming unit (CFU) assay of colostrum-derived MSCs. (a) Representative image of dispersed CFUs after Giemsa staining. Magnification: 10×; Scale bar: 100 μm. (B) Average number of CFUs per sample. Data are expressed as mean ± SD. Statistical analysis: One-way ANOVA with post-hoc Tukey's test. Different letters indicate significant differences (p < 0.05) among samples.
Figure 3.
Colony-forming unit (CFU) assay of colostrum-derived MSCs. (a) Representative image of dispersed CFUs after Giemsa staining. Magnification: 10×; Scale bar: 100 μm. (B) Average number of CFUs per sample. Data are expressed as mean ± SD. Statistical analysis: One-way ANOVA with post-hoc Tukey's test. Different letters indicate significant differences (p < 0.05) among samples.
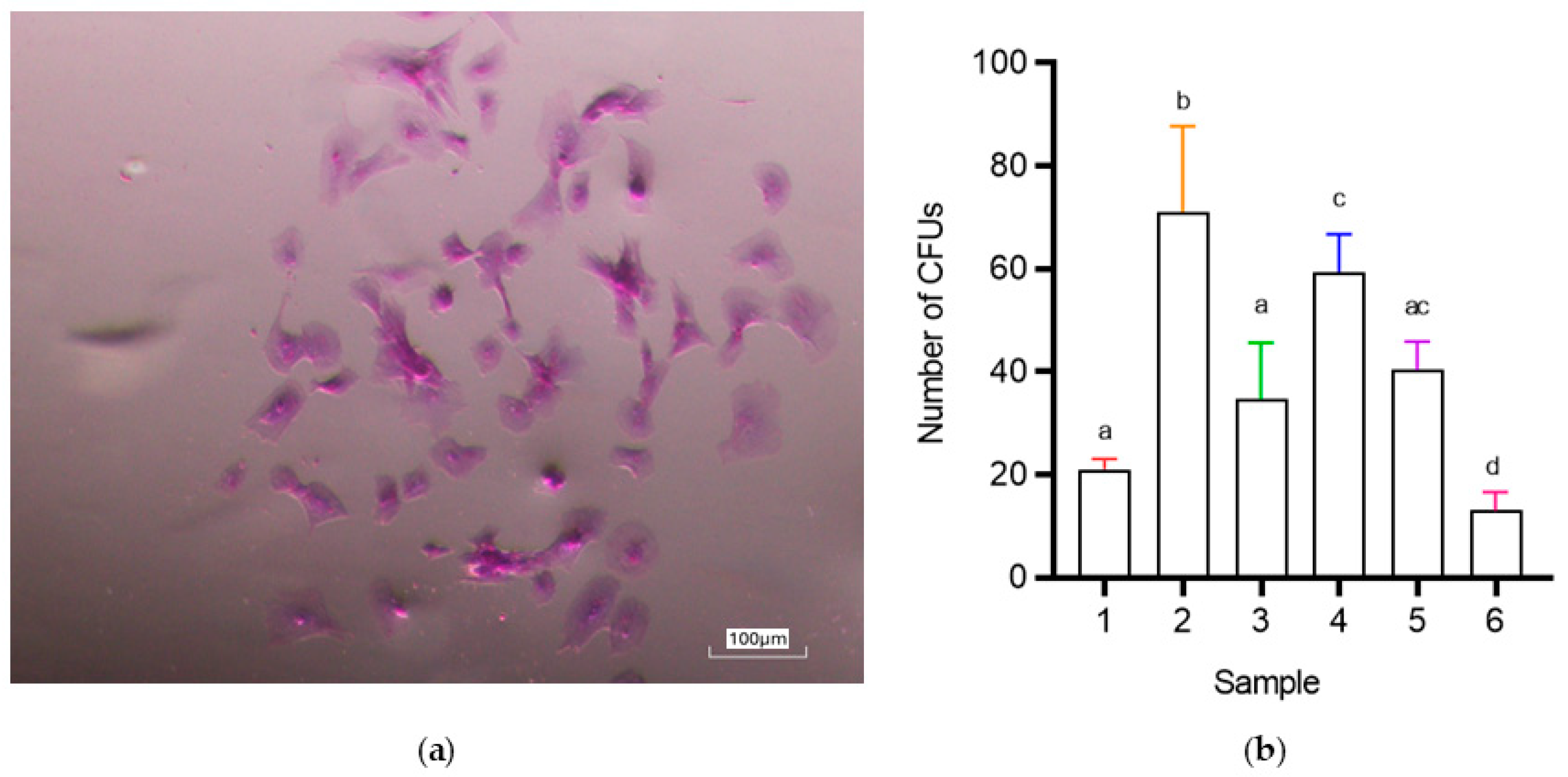
Figure 4.
Adhesion assay of colostrum-derived MSCs. Representative brightfield images showing spheroid-forming (a) and non-forming (b) colostrum-derived MSCs after 1 to 4 days of hanging drop culture (5000 cells/25 µL drop). Magnification: 10×; Scale bar: 100 μm. DIV, day in vitro. 3D spheroid volume reconstruction (c) was performed using ReViSP software, starting from the acquired 2D brightfield images. Measure unit: pixel.
Figure 4.
Adhesion assay of colostrum-derived MSCs. Representative brightfield images showing spheroid-forming (a) and non-forming (b) colostrum-derived MSCs after 1 to 4 days of hanging drop culture (5000 cells/25 µL drop). Magnification: 10×; Scale bar: 100 μm. DIV, day in vitro. 3D spheroid volume reconstruction (c) was performed using ReViSP software, starting from the acquired 2D brightfield images. Measure unit: pixel.
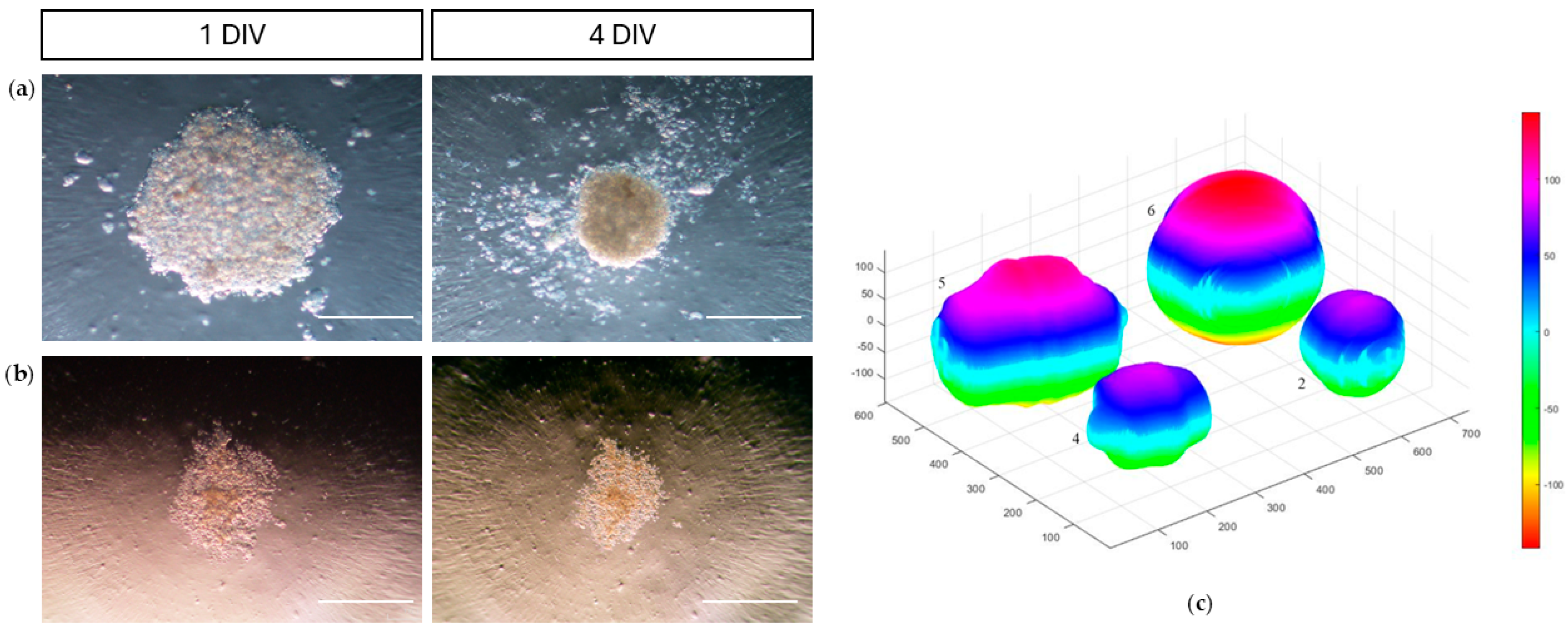
Figure 5.
Scratch assay on colostrum-derived MSCs. Wound closure percentages were calculated using ImageJ software at 24, 48, 72, and 96 hours using the formula: [(distance at t0−distance at tn *100]/distance at t0. Data, presented as mean values, were plotted over time.
Figure 5.
Scratch assay on colostrum-derived MSCs. Wound closure percentages were calculated using ImageJ software at 24, 48, 72, and 96 hours using the formula: [(distance at t0−distance at tn *100]/distance at t0. Data, presented as mean values, were plotted over time.
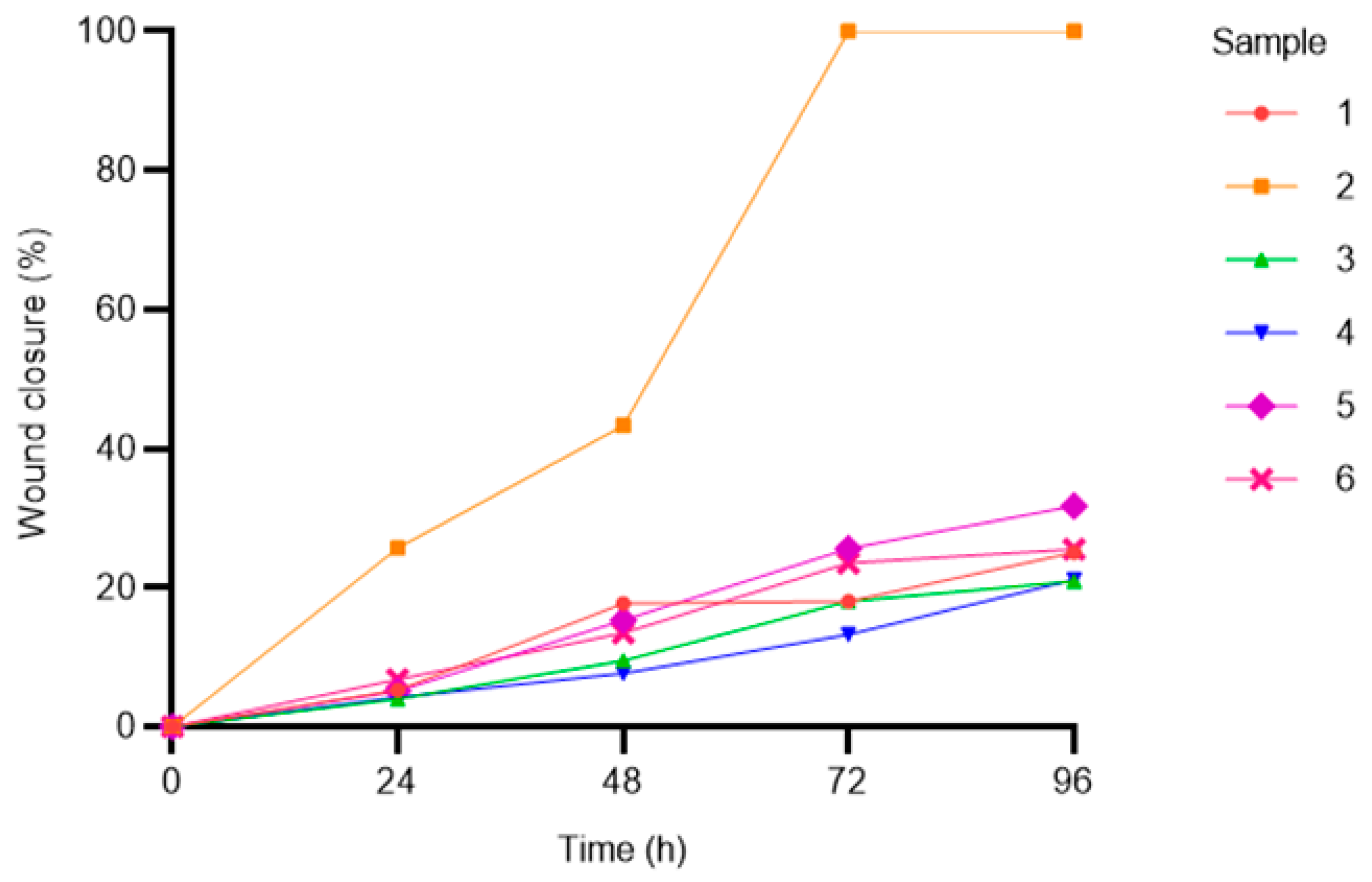
Figure 6.
Multilineage differentiation of colostrum-derived MSCs. (a) Adipogenic differentiation: Representative Oil Red O staining shows intracellular lipid droplets after 3 weeks of induction. (b) Adipogenic control: Representative image of cells cultured in standard medium for 21 days, showing normal morphology and no staining. (c) Chondrogenic differentiation: Representative Alcian Blue staining highlights glycosaminoglycans in the cartilage matrix after 3 weeks of induction. (d) Chondrogenic control: Representative image of cells in standard medium for 21 days, showing normal morphology and no staining. (e) Osteogenic differentiation: Representative Alizarin Red S staining indicates extracellular calcium deposition after 3 weeks of induction. (f) Osteogenic control: Representative image of cells in standard medium for 21 days, showing normal morphology and no staining. Magnifications: Induction panels: 10×, controls: 4×. Scale bar: 100 μm.
Figure 6.
Multilineage differentiation of colostrum-derived MSCs. (a) Adipogenic differentiation: Representative Oil Red O staining shows intracellular lipid droplets after 3 weeks of induction. (b) Adipogenic control: Representative image of cells cultured in standard medium for 21 days, showing normal morphology and no staining. (c) Chondrogenic differentiation: Representative Alcian Blue staining highlights glycosaminoglycans in the cartilage matrix after 3 weeks of induction. (d) Chondrogenic control: Representative image of cells in standard medium for 21 days, showing normal morphology and no staining. (e) Osteogenic differentiation: Representative Alizarin Red S staining indicates extracellular calcium deposition after 3 weeks of induction. (f) Osteogenic control: Representative image of cells in standard medium for 21 days, showing normal morphology and no staining. Magnifications: Induction panels: 10×, controls: 4×. Scale bar: 100 μm.

Figure 7.
RT-PCR analysis of MSC marker expression in colostrum-derived MSCs. (a) Expression of the housekeeping gene GAPDH, MSC markers CD90 and CD73 (b), hematopoietic markers CD34 and CD45 (c), and major histocompatibility complex markers MHC-I and MHC-II (d). Lane M in each panel corresponds to the molecular weight marker (50 bp DNA ladder).
Figure 7.
RT-PCR analysis of MSC marker expression in colostrum-derived MSCs. (a) Expression of the housekeeping gene GAPDH, MSC markers CD90 and CD73 (b), hematopoietic markers CD34 and CD45 (c), and major histocompatibility complex markers MHC-I and MHC-II (d). Lane M in each panel corresponds to the molecular weight marker (50 bp DNA ladder).
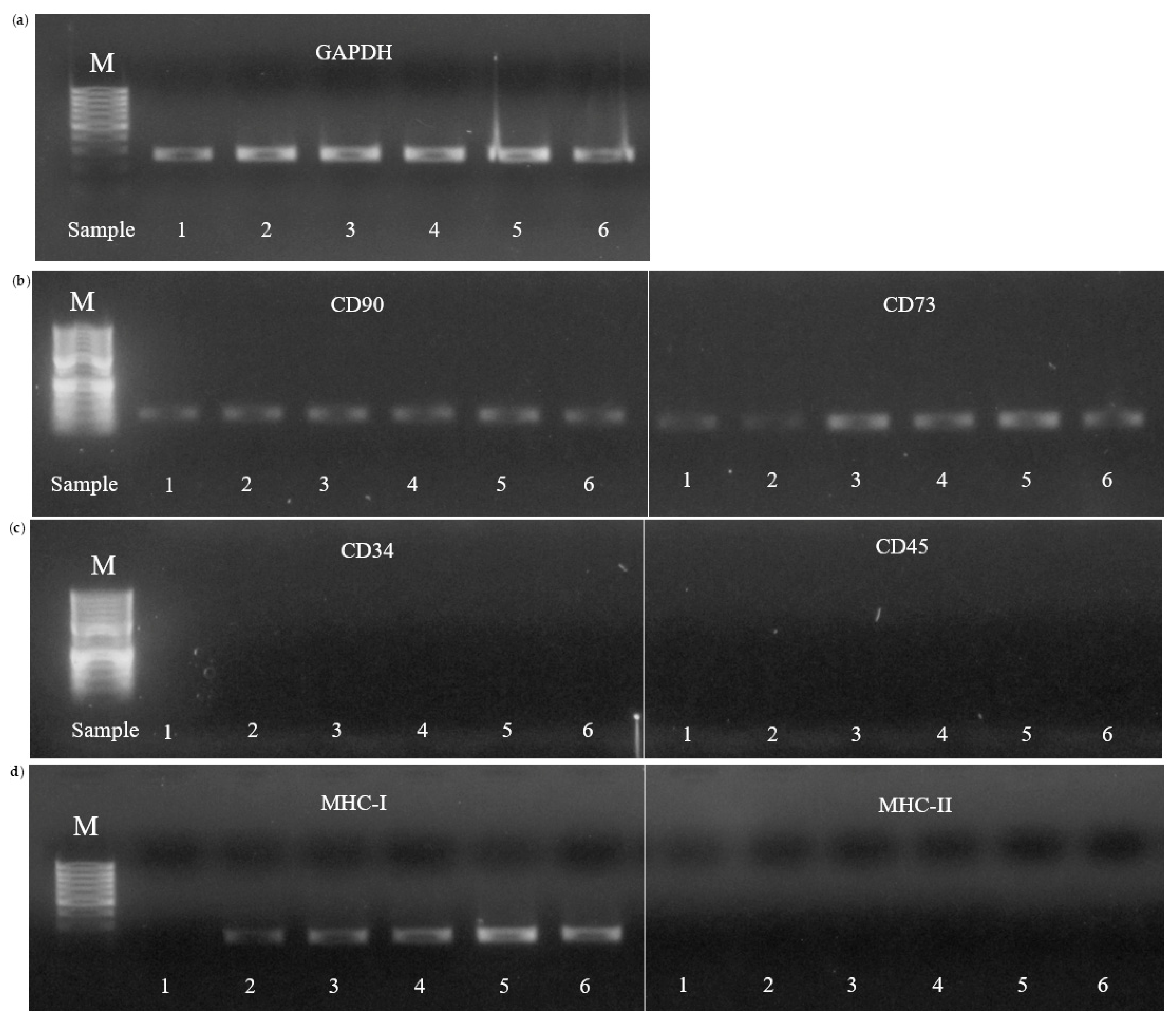

Table 1.
Specific induction media compositions.
| Adipogenic | Chondrogenic | Osteogenic |
| DMEM | DMEM | DMEM |
| 10% FBS | 1% FBS | 10% FBS |
| 1 µM DXM 2 (removed after 6 days) |
0.1 µM DXM 2 | 0.1 µM DXM 2 |
| 0.5 mM IBMX 1 (removed after 3 days) |
50 nM AA2P 4 | 50 µM AA2P 4 |
| 10 µg/mL insulin | 6.25 µg/mL insulin | 10 mM BGP 5 |
| 0.1 mM indomethacin | 10 ng/mL hTGF-β1 3 |
1 IBMX: isobutylmethylxanthine, 2 DXM: dexamethasone, 3 hTGF-β1: human transforming growth factor-β1, 4 AA2P: ascorbic acid 2-phosphate, 5 BGP: beta-glycerophosphate.
Table 2.
Primers sequences for PCR analysis.
| Primers | References | Sequences (5’ →3’) | bp |
| MSC marker | |||
| CD90 | [78] | F: TGCGAACTCCGCCTCTCT R: GCTTATGCCCTCGCACTTG |
93 |
| CD73 | [78] | F: GGGATTGTTGGATACACTTCAAAAG R: GCTGCAACGCAGTGATTTCA |
90 |
| Hematopoietic markers | |||
| CD34 | [78] | F: CACTAAACCCTCTACATCATTTTCTCCTA R: GGCAGATACCTTGAGTCAATTTCA |
101 |
| CD45 | [78] | F: TGATTCCCAGAAATGACCATGTA R: ACATTTTGGGCTTGTCCTGTAAC |
101 |
| MHC markers | |||
| MHC-I | [79] | F: GGAGAGGAGCAGAGATACA R: CTGTCACTGTTTGCAGTCT |
218 |
| MHC-II | [79] | F: TCTACACCTGCCAAGTG R: CCACCATGCCCTTTCTG |
178 |
| Housekeeping | |||
| GAPDH | [80] | F: GTCCATGCCATCACTGCCAC R: CCTGCTTCACCACCTTCTTG |
262 |
Table 3.
Characteristics of Colostrum Samples (Volume, Brix Index at collection, Cell Yield, and Days to Confluence) and Colostrum-Derived MSCs (Mean Population Doubling Times and Number of Colonies).
Table 3.
Characteristics of Colostrum Samples (Volume, Brix Index at collection, Cell Yield, and Days to Confluence) and Colostrum-Derived MSCs (Mean Population Doubling Times and Number of Colonies).
| Sample | Volume (mL) | Brix Index (%) |
Cell Yeld (×10³ cells/mL) |
Days to Confluence |
Mean PDTs (days) |
Number of CFU |
| 1 | 25 | 25 | 260 | 10 | 2.1 ± 0.9 | 21 ± 2 |
| 2 | 23 | 29.5 | 30 | 10 | 1.4 ± 0.3 | 71 ± 16.7 |
| 3 | 17 | 27 | 300 | 11 | 6.2 ± 1.8 | 34.7 ± 11 |
| 4 | 20 | 26 | 500 | 8 | 1.2 ± 0.1 | 59.3 ± 7.4 |
| 5 | 21 | 35 | 480 | 11 | 2.2 ± 0.3 | 40.3 ± 5.5 |
| 6 | 11 | 28 | 400 | 11 | 3.1 ± 0.2 | 13 ± 3.6 |
| Mean | 19.5 | 28.4 | 328.3 | 10.2 | 2.7 ± 0.6 | 39.9 ± 7.7 |
Table 4.
Results of PCR analysis of MSCs derived from six colostrum samples at P3.
| Sample | MSC marker |
Hematopoietic markers |
MHC markers | |||
| CD 90 | CD73 | CD34 | CD 45 | MHC-I | MHC-II | |
| 1 | + | + | - | - | . | - |
| 2 | + | + | - | - | +/– | - |
| 3 | + | + | - | - | +/– | - |
| 4 | + | + | - | - | +/– | - |
| 5 | + | + | - | - | +/– | - |
| 6 | + | + | - | - | +/– | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



