
Submitted:
07 April 2025
Posted:
07 April 2025
You are already at the latest version
Abstract
Objectives. Biofilm infections caused by S. aureus present a significant threat to human health, leading to nosocomial infections on a global scale. To address this critical challenge and explore alternative antibiotic treatments, we investigated the potential of Solanum trilobatum (S. trilobatum), an edible and medicinally important herb plant in Ayurvedic medicine.
Keywords:
Staphylococcus aureus
; biofilm
; infectious disease
; Solanum trilobatum
; serine protease
1. Introduction
A bacterial biofilm is a layer of bacterial colonization that forms beneath a protective wall made of the matrix. This protective wall consists of Extracellular Polymeric Substances (EPS), which include proteins, lipids, polysaccharides, and extracellular DNA. These components play a vital role in developing various microbial infections [1]. Microbial cells within biofilms are known to survive extreme conditions and show resistance to factors such as UV radiation, metal-induced toxicity, exposure to acids, and changes in pH [2]. Additionally, EPS helps these pathogens evade the immune system by inhibiting neutrophil-mediated phagocytosis [3]. Furthermore, extracellular DNA (eDNA) and intercellular adhesins within EPS serve as barriers to antimicrobials, as eDNA can bind to human antimicrobial peptides (AMPs), reducing their effectiveness [4]. Moreover, many bacterial species have shown a greater tendency to accumulate on a variety of surfaces, eventually forming sedentary communities. These surfaces vary from plant and animal tissues to pipelines that carry household and industrial waste and prolonged usage of medical devices such as catheters, implants, and contact lenses [5].
Their proximity in these environments facilitates the efficient exchange of substrates, distributes the required metabolic products, and removes the toxic end products. This accumulation of mono- or poly-microbial aggregates results in the formation of biofilm comprising diverse communities of both bacteria and fungi [6]. Though they impart both positive and negative impacts on human health, biofilms are known to exhibit a profound effect in inducing several pathogenic forms of human diseases and plant infections. The most common examples are cystic fibrosis and dental plaque. While individuals affected by cystic fibrosis are susceptible to chronic Pseudomonas aeruginosa infections resulting in the formation of mucoid biofilm [7,8], patients affected by dental plaque may experience increased biofilm acidification, which in turn leads to the demineralization of the enamel, forming dental caries [9].
Staphylococcus aureus (S. aureus) is widely recognized as a major human pathogen responsible for a variety of diseases, including skin infections, bacteraemia, and infective endocarditis [10]. Among the different staphylococcal strains, S. aureus is considered the most virulent due to its diverse array of secreted and cell surface-associated virulence factors, immune evasion mechanisms, and toxin production [11]. These strains exhibit significant genetic diversity, enabling them to form biofilms and acquire antibiotic resistance. Particularly concerning are the methicillin-resistant S. aureus (MRSA) strains, which often display multiple antibiotic resistances [12]. They are a leading cause of healthcare-associated infections, as they can attach to and accumulate on host tissues or medical devices, leading to complications such as surgical site infections, intravascular catheter (IVC) infections, and implant-related infections [11]. Research indicates that up to 80% of human bacterial infections are associated with biofilm-forming microorganisms like S. aureus [13]. Furthermore, S. aureus strains can also form biofilms in other sectors, such as the food and dairy industries. In these environments, they tend to adhere to milking machines and food-contact surfaces, showing high resistance to antibiotics and sanitization procedures [14]. These biofilms are particularly resistant to both innate and adaptive immune defence systems, as well as to antimicrobial agents, resulting in persistent infections and treatment failures. Owing to their widespread prevalence in diseases and prolonged resistance to antimicrobial treatments, exploring novel approaches to combat or prevent biofilm formation has become of paramount importance worldwide.
Proteases hydrolyze protein peptide bonds and can be classified as serine, cysteine, aspartic, metallo, glutamic, and threonine proteases based on their catalytic residue. Among all the known families, serine proteases are the most extensively characterized proteases [15]. They are involved in the metabolic processes of all organisms and regulate many physiological pathways. In general, proteases from leaves are few and particularly serine proteases from plant leaves are very rare [16,17]. Most of the characterized plant proteases are cysteine proteases and they are readily reduced by air oxidation and metal ions [18]. The search for novel plant proteases having high stability has become a global research theme.
The plant, S. trilobatum, is well known for its traditional use of mucus clearance. Incidentally, this plant is known to demonstrate the existence of hydrolase. S. trilobatum has been in-use for various ailments in different parts of Asian countries [19,20] and its active principle ‘sobatum’ was reported to possess anti-tumor activity [21,22], anti-inflammatory activity [23] and the tannin of S. trilobatum leaves reported to have anti-bacterial activity [24,25]. The aqueous extract, methanol extract of leaves and stem were also reported to have anti-microbial activity [26]. The leaf extract of S. trilobatum was reported with anti–oxidant potency [27] as well as possessed oviposition deterrent and repellent activities against the mosquito Anopheles stephensi [28]. The presence of Nucleotide Binding Site (NBS)-Leucine rich repeating(LRR) resistance gene analogs have also been reported in S. trilobatum plant species [29]. While plant extracts such as Piper betle L [30], Eucalyptus globulus Labill, and Juglans regia L [31] have been studied for their antibiofilm properties against S. aureus strains, no published reports exist on the antibiofilm activity of S. trilobatum extracts. Furthermore, previous studies have primarily attributed the antibiofilm properties of these plant extracts to the presence of phytochemicals rather than proteins or enzymes. Therefore, this study presents a novel investigation into the potential of S. trilobatum-derived serine protease in combating S. aureus biofilms. In this context, we describe the identification, purification, and biofilm-eradicating potency of a serine protease from the leaves of S. trilobatum, an edible plant known for its medicinal properties.
2. Materials and Methods
2.1. Extraction and Purification
As reported earlier [25] the fresh leaves of S. trilobatum were collected from the Vandalur forest area near Chennai, India. After removing the thorns, the leaves were thoroughly washed with distilled water and left to air dry. A total of 100 g of the dried leaves were weighed and then extracted using a 50 mL buffer containing 20 mM Tris (pH 7.8),150mM Nacl. The resulting mixture was filtered through muslin cloth to remove large particulates.
The filtered extract was then centrifuged at 12,000 rpm for 40 minutes at 4°C to separate the supernatant (crude extract) from the pellet. Enzymatic assays were performed on the supernatant using trypsin as positive and the extraction buffer as negative controls [15].
For further purification, the crude extract was subjected to ammonium sulfate precipitation. Pulverized ammonium sulfate was gradually added to the extract with constant stirring to achieve 30% saturation. After standing for 2 hours at 4°C, the mixture was centrifuged again at 12,000 rpm for 40 minutes at 4°C. The resulting pellet was re-suspended in 3 mL of the extraction buffer. Subsequently, the saturation was adjusted to 30-60% and 60-80%, with the same centrifugation and re-suspension procedures applied for each fraction. The three obtained fractions were then extensively dialyzed against 20 mM Tris (pH 7.8) using 10 kDa cut-off dialysis membranes at 4°C overnight, followed by an additional 4 hours of dialysis against fresh buffer to remove excess salts.
2.2. Purification of the Active Fraction via Cationic Exchange Chromatography
Ion exchange chromatography was performed using a strong anion-exchange column, Q-Sepharose HP-FF 1 mL (GE Life Sciences), to purify the active 60-80% fraction. The column was pre-equilibrated with 10 column volumes (CV) of Buffer A [20 mM Tris, pH 7.8, containing 150 mM NaCl] and Buffer B [20 mM Tris, pH 7.8, containing 1 M NaCl], at a flow rate of 1 mL/min as mentioned by [25]. Before loading the active fraction, it was centrifuged briefly at 12,000 rpm at 4°C to remove any particulate matter, and the clear supernatant was loaded onto the column. The unbound proteins were washed out with 8 CV of Buffer A, while the bound proteins were eluted using a linear salt gradient, collecting fractions in 20 CV of Buffer B at a flow rate of 1 mL/min. The eluted and unbound fractions were analyzed for enzymatic activity to identify the fraction with the highest activity. Additionally, all fractions were subjected to polyacrylamide gel electrophoresis (PAGE) to assess their purity and homogeneity. This allowed for identification of the fraction with the desired enzyme activity and ensured that the protein was adequately purified.
2.3. Biofilm Eradication Assay
S. aureus strain of Microbial Type Culture Collection and Gene Bank (MTCC) No: 96 from Institute of Microbial Technology, India were grown overnight at 37 °C in an autoclaved LB medium was plated on a LB agar plate. A well-grown single colony was picked and inoculated in LB medium and grown overnight at 37 °C with agitation at 180 rpm. These bacterial suspensions were used as inoculums for all these biofilm experiments. The biofilm was formed by adding 900 µl of LB media to 100 µl from the inoculum of S. aureus, mixed in two different 24-well polystyrene plates from top to bottom in their respective wells. Wells were sealed with transparent tape and then incubated at 37 °C for 2 days in static conditions. After 2 days, the contents of the well were discarded carefully and washed once with a 0.1M NaCl solution, followed by water three times. The wells of the first plate alone were stained with a crystal violet solution (15 minutes) and then washed with water. The biofilms that were formed were observed through an optical microscope. Whereas in the second plate (unstained), the preformed biofilm was treated with 160 µM of purified protein as well as with a combination of protein fraction and ampicillin and kept overnight at 37°C in a static environment. After 12 hours, the contents of the well were carefully discarded and washed with water three times. The wells were stained with a crystal violet solution, and then they were washed with water. The changes in the biofilms were analyzed with the help of an optical microscope and the experiment adopted with slight modification from Jayarajan, S. et.al., [32].
2.4. Proteolytic Activity Measurement
Protease activity was measured using a modified UV spectroscopy method based on Benyon [33]. A 160 µM aliquot of the protease fraction was mixed with an equal volume of 1% casein (w/v) and incubated at 37°C for 30 minutes. The reaction was terminated by adding 200 µL of 10% ice-cold trichloroacetic acid (TCA), and the mixture was incubated for an additional 30 minutes at 37°C. After incubation, the sample was centrifuged at 10,000 rpm for 30 minutes at 4°C to separate the precipitate. The clear supernatant, containing the lytic fragments of casein, was collected and analyzed for absorbance at 280 nm using a UV spectrophotometer[34]. Proteolytic activity was assessed for both the aqueous crude extract and ammonium sulfate fractions, with positive (trypsin) and negative (lysis buffer used in protein preparation) controls include for comparison.
2.5. Effect of Standard Inhibitors on Protease Activity
The purified protease was assessed for its proteolytic activity using the casein agar radial diffusion method [15]. To determine the protease's characteristic nature, specific inhibitors were used: Ethylenediaminetetraacetic acid (EDTA) for metalloproteases, Iodoacetic acid (IAA) for cysteine proteases, and Phenylmethylsulfonyl fluoride (PMSF) for serine proteases. EDTA and IAA were dissolved in water, while PMSF was dissolved in 10% ethanol. The purified protease was pre-incubated with 5 mM EDTA, 2 mM IAA, and 2 mM PMSF in separate vials and the reaction mixture added into the respective wells of casein agar plate kept at 37°C overnight,the proteolytic activity was then measured from the digestion of casein zone.
2.6. Effect of Detergents, Organic Solvents and Metal Ions on Proteolytic Activity
The stability of the protease was evaluated in the presence of various organic solvents such as toluene, benzene, acetone, and isopropanol and surfactants and an oxidizing agent. The reaction mixture, consisting of purified protease and 0.6% casein solution (pH 7.8), was pre-incubated with 0.1 mM of various metal ions, including Ca²⁺, Zn²⁺, Cu²⁺, Mg²⁺, Mn²⁺, Fe²⁺, and Ba²⁺, at 37°C for 30 minutes. The reaction was then stopped by adding 10% Trichloroacetic acid (TCA) and incubated at room temperature for 30 minutes with slighter modification [34,35]. Afterward, the mixture was centrifuged at 10,000 rpm for 5 minutes to separate the precipitate. The supernatant was mixed with 0.5 M sodium carbonate and Folin-Ciocalteu reagent and incubated at room temperature for 30 minutes. The amount of tyrosine released from casein was quantified by measuring the absorbance at 660 nm. A control assay was performed without metal ions, and the resulting enzyme activity was considered as 100%. The effect of each metal ion on protease activity was determined by comparing the enzyme activity in the presence of the metal ions to the control. The same methodology followed to determine the effect of SDS, 0.2% Triton X-100, and Tween 80, along with 0.2% H₂O₂ as the oxidizing agent[36].
2.7. Effect of pH and Temperature on Proteolytic Activity
To assess the effect of pH on the purified protease, the enzyme was exchanged into buffers with varying pH levels: 20 mM sodium acetate buffer (pH 4.0 and 5.0), 20 mM sodium phosphate buffer (pH 6.0 and 7.0), and 20 mM Tris buffer (pH 8.0 and 9.0). The exchange was carried out for 12 hours at 4°C, after which the enzymatic activity was measured using the previously described method with some modifications [15]. The effect of temperature on enzymatic activity was determined by incubating the purified protease at different temperatures (20°C, 40°C, 60°C, and 80°C) for 30 minutes in 20 mM Tris buffer (pH 7.8). After incubation, proteolytic activity was assessed by measuring the zone of casein digestion as mentioned earlier.
2.8. Zymography
Zymography was performed at 4°C using 1% casein as a substrate, which was copolymerized with polyacrylamide gel. After electrophoresis under non-reducing conditions at 50 mV, the gel was washed twice with 50 mL of renaturation buffer (2.5% Triton X-100) for 2 hours to remove SDS. The gel was then incubated overnight at 37°C in a buffer containing 20 mM Tris (pH 7.8), 50 mM NaCl, and 10 mM CaCl₂. After incubation, the gel was stained with Coomassie Brilliant Blue G-250. The proteolytic activity of the enzyme was visualized as a clear, colorless zone against substrates background, indicating gelatin and casein digestion [37].
2.9. Determination of Vmax and km
The spectroscopic method was employed to determine the kinetic parameters, Vmax and Km, of S. trilobatum serine protease using casein as the substrate as described witha a slight modification [38]. A fixed concentration of 5 mM serine protease, along with a fixed reaction time, was used to measure the rate of proteolysis as the substrate concentration increased. To assess residual protease activity, varying concentrations of casein (ranging from 7 to 80 µM) were added to purified serine protease and incubated for 30 minutes at 37 °C in a 50 mM Tris buffer at pH 7.8. The reaction was stopped by adding 10% ice-cold TCA solution after the 30-minute mark. After incubation at 4 °C for an additional 30 min, the mixture was centrifuged at 12,000 rpm for 30 min, and the supernatant was used for the spectroscopic analysis. The progression of the proteolysis reaction was observed by measuring UV absorbance at 280 nm. The Km and Vmax value of serine protease was determined by the Substrate – Velocity curve. Assays were carried out in triplicates and the kinetic parameters were determined using Graph pad Prism 6.0 (San Diego, CA).
3. Results and Discussion
3.1. Isolation, and Purification of Protease from Solanum trilobatum
The S. trilobatum leaves were extracted in 20 mM Tris and 150 mM NaCl buffer, following which the protease activity of the crude was tested by the radial diffusion method (Figure 1a). By using casein as the substrate, the dialyzed ammonium sulfate fractions (0-30%, 30-60%, and 60-80%) along with the aqueous crude extract were evaluated by the UV spectroscopy method. The proteolytic activity was measured from the cleaved and liberated casein fragments, which were subsequently quantified by UV 280nm absorbance. Among the various fractions, the 60-80% fraction exhibited maximum protease activity similar to that of bovine trypsin (Figure 1b). Additionally, substantial protease activity was observed in the crude extract as well.
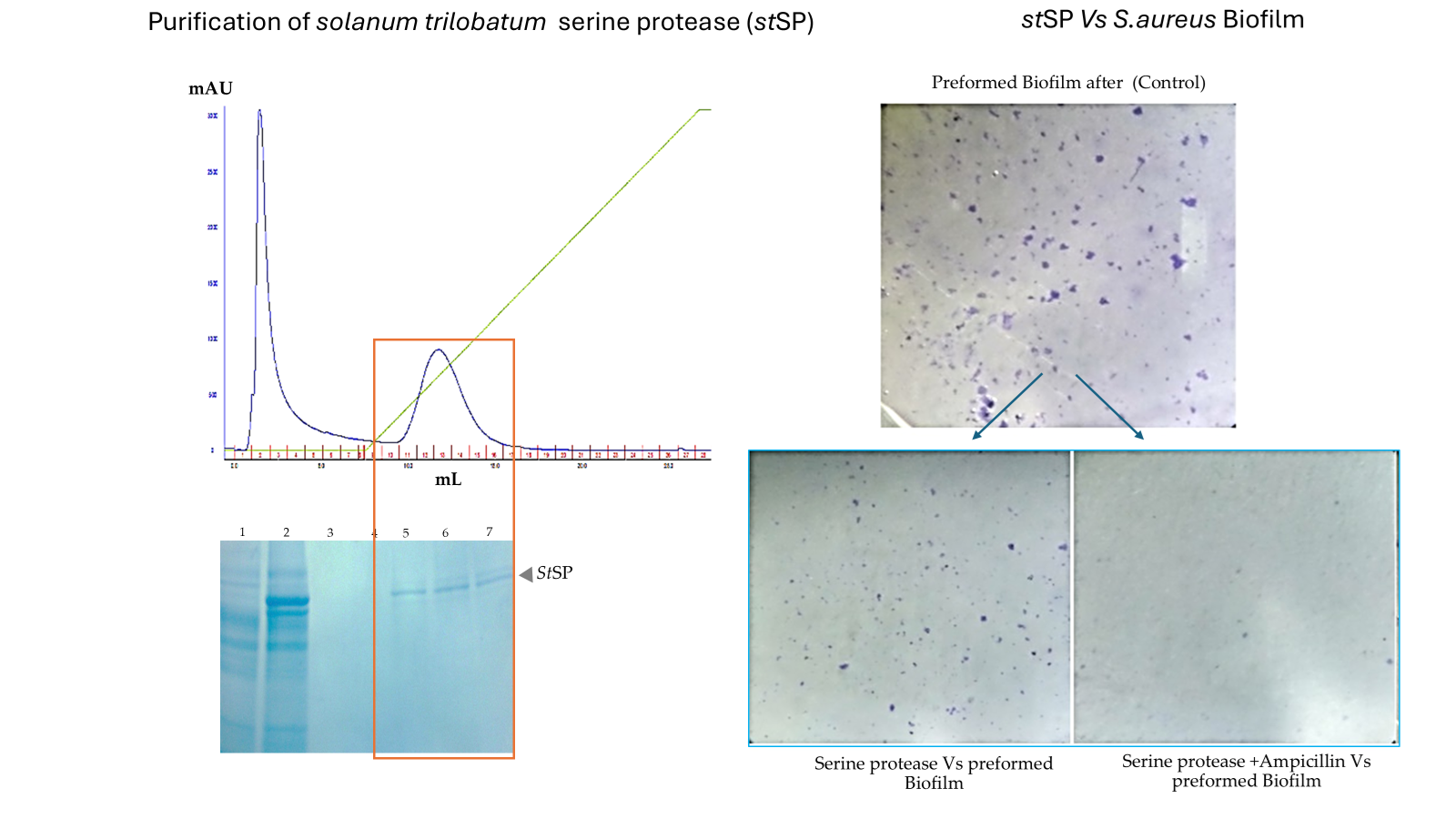
Based on this observation, the active fraction (60-80%) was loaded onto the Q- Sepharose Fast Flow anionic exchange column. While the unbound proteins were collected as an unabsorbed fraction with the same buffer, the bound proteins were eluted using an increasing concentration of salt. The elution profile of both the flow through and active fractions are depicted in Figure 2a. Following the chromatographic run, both the unbound and elution peak fractions were analyzed for their proteolysis activity on a casein agar plate. The activity study revealed that only the eluted peak (bound) fractions exhibited reliable protease activity when compared to the unbound fraction (Figure 2b). Using 10% SDS-PAGE, the homogeneity of those bound fractions displaying the caseinolytic activity was determined, confirming the protease's purity (Figure 2c). Interestingly, the molecular weight (~60 kDa) of this purified S. trilobatum Serine Protease (STSP) was found to be in accordance with serine proteases found in fruits of Cucumis trigonaus [39], leaves of wheat seeds [40] and seeds Solanum dubium [41].
As a result, the fractions exhibiting proteolytic activity were pooled, concentrated, and used for further kinetic as well as other functional characterization studies. The proteolytic activity of this STSP was then evaluated again by UV spectroscopy using casein as the substrate (Figure 2d).
3.2. Inhibition of Serine Protease by Serine Protease Inhibitor (PMSF)
As outlined in the methodology section, the enzyme activity of STSP was tested in the presence of cysteine protease inhibitor Iodoacetamide (IAA), metalloprotease inhibitor (Ethylenediaminetetraacetic acid-EDTA), and serine protease inhibitor (Phenylmethylsulfonyl fluoride-PMSF). The results indicated that protease activity remained unaffected in the presence of either EDTA or IAA, as evidenced by distinct zones of digestion observed in wells containing these inhibitors (Figure 3a). Conversely, the incorporation of serine protease inhibitor PMSF to the purified protease completely inhibited its enzymatic activity (Figure 3b), confirming that STSP is a member of the serine protease family.
3.3. Staphylococcus aureus Pre-Formed Biofilm Eradicated by StSP
Biofilm eradication experiments were employed to understand the effect of these enzymes on preformed biofilm surfaces. The experiments reveal the destruction of preformed biofilm by the treatment of these enzymes. In Figure 4, the upper panels (a,b,c,d) shows the preformed biofilm on poly styrene surface of the micro-titre plate. The lower panel corresponds to respective sample treatment mentioned in the figure. The lower lane of the figure, StSP treated biofilm shows the reduced growth compared control. When preformed biofilm was incubated with media and grown static, profound propagation is witnessed (referred as “untreated”). The robustness of the biofilm could be witnessed the fact that ampicillin has no effect on biofilm and the propagation is as good as the untreated surface (Figure 4.h). Though differ by their specificities, the degree of disturbance exhibited by both enzyme and the combination of the enzyme and the antibiotic (StSP and Ampicillin) is comparable (Figure 4.e,g,h). From visual comparison StSP and the combination of enzyme and the antibiotic, the combined usage found to have better biofilm eradication potency than the simple enzyme (StSP) treatment (Figure 4.f). The presence of biofilm even after treatment with enzymes may be due to the spread of biofilm fragments which are cleaved off by enzymes. The reduced biofilm growth suggest the influence of antibiotic on the fragment or the planktonic bacteria excreted from biofilm after digestion of EPS.
3.4. Effect of Metal Ions
The enzymatic activity of serine protease isolated from S. trilobatum leaves has observed a significant enhancement with the addition of 0.1 mM Mn2+ and Fe2+ metal ions while experiencing a moderate activation with Cu2+ and Ba2+ (Figure 5a). Notably, the residual activity showed a remarkable increase of more than 2-fold, reaching up to 257% and 219% when incubated with Mn2+ and Fe2+ metal ions, respectively. However, the enzymatic activity of StSP remained unaffected by the presence of Zn2+ and Ca2+ ions among the tested metal ions (Figure 5a). Interestingly, similar activation of residual activity by a 60 kDa serine protease from pigeon pea leaves was observed with Mn2+, Fe2+, and Cu2+. In summary, none of the tested metal ions inhibited or altered the proteolytic activity of StSP, aligning with previously published reports that emphasize the role of metal ions in enhancing the activity of plant proteases [42].
3.5. Effect of Surfactants
The presence of 5 mM SDS and 0.2% Triton X-100 minimally impacted the activity of StSP, while the addition of 0.2% Tween resulted in a notable 20% enrichment in activity (Figure 5b). This finding aligns with previous studies on a 30 kDa serine protease from Scorzonera hispanica [43], where the proteolytic activity of STSP exhibited noteworthy resistance to these detergents. Particularly interesting is the fact that 80% of the activity was retained when incubated with 0.2% of the oxidizing agent H2O2. The resistance of S. trilobatum protease to these detergents and oxidizing agents opens new possibilities for its application in detergent formulations.
3.6. Effect of Organic Solvents
Enzymes often experience instability in the presence of various organic solvents, leading to denaturation. The stability of the StSP was assessed in the presence of different organic solvents, including toluene, benzene, acetone, and isopropanol (Figure 5c). However, when treated with acetone, the activity was retained at more than 100%, and it approached 100% when exposed to isopropanol and toluene. Consequently, the enzyme exhibited greater stability in acetone. The enzyme's stability in these solvents is comparable to that reported for serine proteases from Scorzonera hispanica [43].
3.7. Determination of Kinetic Parameters
UV-based enzyme-substrate kinetic parameters Vmax and Km for the purified StSP were determined by using casein as the substrate. The Michaelis-Menten substrate-velocity curve yielded Vmax and Km values of 48.63 µM/min and 14.63 µM respectively (Figure 6).
The determined Km value reflects the StSP's affinity towards the serine protease substrate, while Vmax signifies the enzyme's maximum catalytic ability, aiming for optimal efficiency. The study involved varying substrate concentrations (7–80 µM), revealing that at higher concentrations of the substrate, the proteolytic activity reached the enzyme's saturation point. This was evident in the linear regression plot and the Km of this enzyme, at 14.63 µM. The STSP enzyme has a Vmax of approximately 48.63 µM and a Km value of 14.08 µM/min. This suggests that its catalytic function is more efficient than that of a 69.9 kDa serine protease found in the seeds of Cyamopsis tetragonoloba, which has a Vmax of about 102 µM and a Km of approximately 56.56 µM/min [44]. Additionally, there is a stable 57.9 kDa serine protease derived from the latex of Wrightia tinctoria [45], which has a Km value of around 50 µM. Furthermore, a subtilisin-like serine protease, weighing about 59 kDa and reported from wheat leaves, exhibits a Vmax of approximately 2.27 mM and a Km of 1.18 mM [40].
3.8. Sustainability of Various pH and Temperature
The pH and temperature stability of the purified StSP were assessed across various ranges (pH 4.0–9.0) and temperatures (20 °C to 80 °C) using casein as the substrate, employing the radial diffusion method. As depicted in Figure 7a, the protease activity exhibited a gradual increase from an acidic pH of 4.0 to a maximum at a neutral pH. The optimal pH for enzymatic activity was determined to be 7.0, with a functional range identified between pH 6 and 8. The caseinolytic activity of the protease remained robust within a narrow temperature range of 40 °C to 50 °C, retaining full functionality. Up to 50 °C, the activity was nearly constant, but a sudden decline occurred at 60 °C and 70 °C, as illustrated in Figure 7b, and at 80 °C, the caseinolytic activity was completely lost. This loss may be attributed to a slight conformational change in the active or binding site, leading to the loss of proteolytic activity.
The moderate thermal stability and optimal activity at pH 7.0 observed in this study are consistent with other serine proteases reported from Kesinai leaves [46].
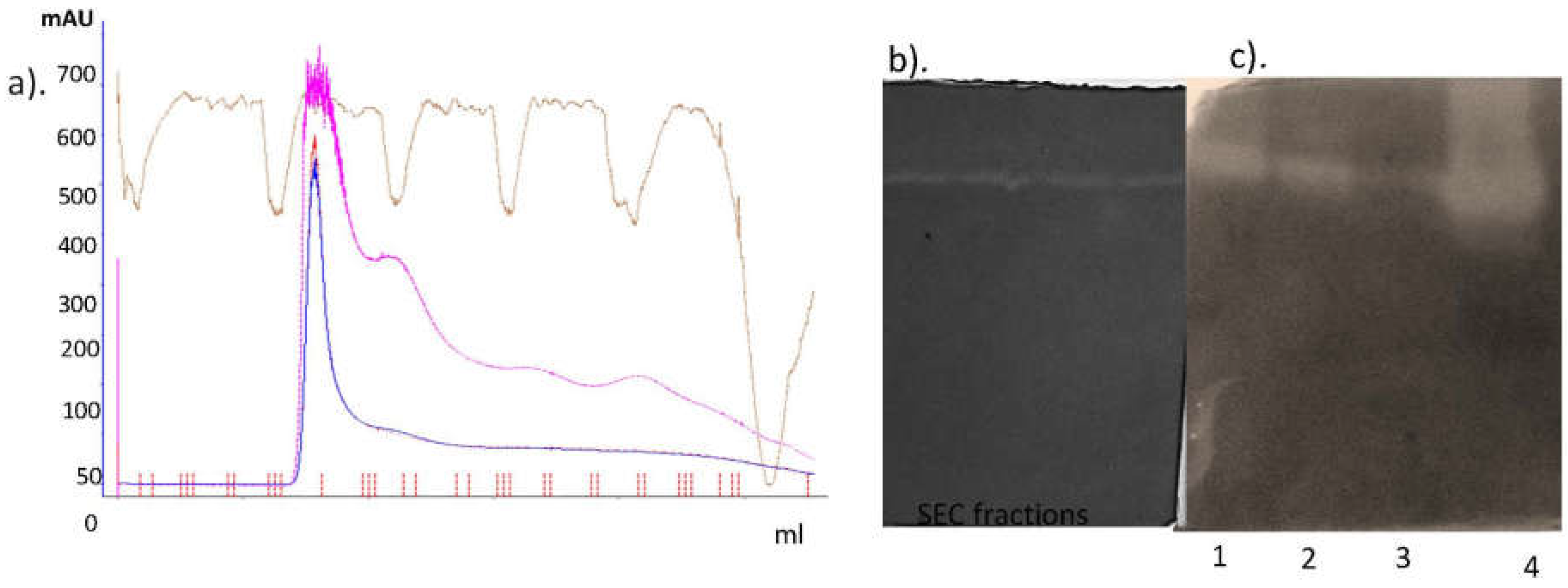
The proteolytic activity of the purified protein was confirmed through zymography on the background of using both gelatin (Figure 8.b) and casein (Figure 8.c) as substrates. Clear zones of digestion with those substrates were observed, as witnessed under the non-denaturing condition as a single bond of digestion. In Figure 8.b, the size exclusion fractions of Figure 8.a. corelated with SEC peak as the enzyme exhibited as a monomer in the native, non-denaturing and denaturing system.
4. Conclusions
To the best of our knowledge, this is the first purification and characterization report on a 60 kDa serine protease from the leaves of S. trilobatum. The consumable nature and medicinal value of these leaves, coupled with their therapeutic properties against respiratory ailments, suggest a potential biological relevance in human health. While these leaves are already consumed as fresh juice and as a food additive, their newfound potential presents an innovative direction for the industry. The purified enzyme showcases proteolytic action, with an optimum pH of 7.0 and moderate thermal stability up to 50 °C. The purified StSP has displayed the capability to eradicate biofilms produced by S. aureus supports the traditional use of the leaves for alleviating throat infections,gastrointestinal and reducing mucus congestion.
Author Contributions
Conceptualization, K.G. and M.R.; methodology, N.R.; software, N.E.K; validation K.G. and M.R.; formal analysis and investigation, K.G.; resources, R.V.; data curation, K.E.; writing—original draft preparation, L.K.; writing—review and editing, M.R.; visualization, K.G.; supervision, K.G.; project administration, M.R.; funding acquisition, K.G. All authors have read and agreed to the published version of the manuscript.
Funding
The author thanks UGC, India for financial support.
Conflicts of Interest
All the authors declare no conflicts of interest.
References
- Ch’ng J.-H., Chong K. K., Lam L. N., Wong J. J., & Kline K. A. (2019). Biofilm-associated infection by Enterococci. Nat. Rev. Microbiol, 17, 82–94.
- Hall-Stoodley L., Costerton J. W., & Stoodley P. (2004). Bacterial biofilms: from the natural environment to infectious diseases. Nat. Rev. Microbiol, 2, 95–108. [CrossRef]
- Gunn J. S., Bakaletz L. O., & Wozniak D. J. (2016). What’s on the outside matters: the role of the extracellular polymeric substance of gram-negative biofilms in evading host immunity and as a target for therapeutic intervention. J. Biol. Chem, 291, 12538–12546. [CrossRef]
- Jones E. A., McGillivary G., & Bakaletz L. O. (2013). Extracellular DNA within a nontypeable Haemophilus influenzae-induced biofilm binds human defensin-3 and reduces its antimicrobial activity. J. Innate Immun, 5, 24–38. [CrossRef]
- Wu, H., Moser, C., Wang, H. Z., Høiby, N., & Song, Z. J. (2015). Strategies for combating bacterial biofilm infections. International Journal of Oral Science, 7(1), 1–7.
- Haaber, J., Cohn, M. T., Frees, D., Andersen, T. J., & Ingmer, H. (2012). Planktonic aggregates of Staphylococcus aureus protect against common antibiotics. PLoS One, 7(7), e41075.
- Ciofu, O., Mandsberg, L. F., Wang, H., & Hoiby, N. (2012). Phenotypes selected during chronic lung infection in cystic fibrosis patients: implications for the treatment of Pseudomonas aeruginosa biofilm infections. FEMS Immunology & Medical Microbiology, 65, 215–225. [CrossRef]
- Roy, R., Tiwari, M., Donelli, G., & Tiwari, V. (2018). Strategies for combating bacterial biofilms: A focus on anti-biofilm agents and their mechanisms of action. Virulence, 9(1), 522–554. [CrossRef]
- Salli, K. M., & Ouwehand, A. C. (2015). The use of in vitro model systems to study dental biofilms associated with caries: a short review. Journal of Oral Microbiology, 7, 26149. [CrossRef]
- Giordani, B., Costantini, P. E., Fedi, S., Cappelletti, M., Abruzzo, A., Parolin, C., ... & Vitali, B. (2019). Liposomes containing biosurfactants isolated from Lactobacillus gasseri exert antibiofilm activity against methicillin resistant Staphylococcus aureus strains. European Journal of Pharmaceutics and Biopharmaceutics, 139, 246-252. [CrossRef]
- Hogan, S., Zapotoczna, M., Stevens, N. T., Humphreys, H., O’Gara, J. P., & O’Neill, E. (2017). Potential use of targeted enzymatic agents in the treatment of Staphylococcus aureus biofilm-related infections. Journal of Hospital Infection, 96(2), 177–182.
- Gudiña, E. J., Rangarajan, V., Sen, R., & Rodrigues, L. R. (2013). Potential therapeutic applications of biosurfactants. Trends in pharmacological sciences, 34(12), 667-675.
- Römling, U. & C. Balsalobre. (2012). Biofilm infections, their resilience to therapy and innovative treatment strategies. J Intern Med, 272(6), 541–561. [CrossRef]
- Gharieb, R. M. A., Saad, M. F., Mohamed, A. S., & Tartor, Y. H. (2020). Characterization of two novel lytic bacteriophages for reducing biofilms of zoonotic multidrug-resistant Staphylococcus aureus and controlling their growth in milk. LWT, 124, 109145.
- Manohar, R., Kutumbarao, N. H. V., Krishna Nagampalli, R. S., Velmurugan, D., & Gunasekaran, K. (2018). Structural insights and binding of a natural ligand, succinic acid with serine and cysteine proteases. Biochemical and Biophysical Research Communications, 495(1), 679-685.
- Smith, A., Datta, S. P., Smith, G. H., Campbell, P. N., Bentley, R., & McKenzie, H. (2000). Oxford dictionary of biochemistry and molecular biology. Oxford University Press.
- Antão, C. M., & Malcata, F. X. (2005). Plant serine proteases: biochemical, physiological and molecular features. Plant Physiology and Biochemistry, 43(7), 637-650.
- Ahmed, I. A. M., Morishima, I., Babiker, E. E., & Mori, N. (2009). Characterisation of partially purified milk-clotting enzyme from Solanum dubium Fresen seeds. Food Chemistry, 116(2), 395-400.
- Govindan, S., Viswanathan, S., Vijayasekaran, V., & Alagappan, R. (1999). A pilot study on the clinical efficacy of Solanum xanthocarpum and S. trilobatum in bronchial asthma. Journal of Ethnopharmacology, 66(2), 205-210. [CrossRef]
- Madhavan, S., & Balu, S. (2000). Ethnobotanical studies on S. trilobatum L.-an Indian drug plant. In Ethnobotany and medicinal plants of Indian subcontinent (pp. 43-46). Jodhpur: Scientific Publishers.
- Mohanan, P., & Devi, K. (1996). Cytotoxic potential of the preparations from S. trilobatum and the effect of sobatum on tumour reduction in mice. Cancer Letters, 110(1), 71-76.
- Shahjahan, M., Sabitha, K., Jainu, M., & Devi, C. S. (2004). Effect of S. trilobatum against carbon tetra chloride induced hepatic damage in albino rats. Indian Journal of Medical Research, 120, 194-198.
- Emmanuel, S., Ignacimuthu, S., Perumalsamy, R., & Amalraj, T. (2006). Antiinflammatory activity of S. trilobatum. Fitoterapia, 77(7), 611-612.
- Doss, A., Mubarack, H. M., & Dhanabalan, R. (2009). Antibacterial activity of tannins from the leaves of S. trilobatum Linn. Indian Journal of Science and Technology, 2(2), 41-43.
- Radhakrishnan, M., Palayam, M., Altemimi, A. B., Karthik, L., Krishnasamy, G., Cacciola, F., ... Leucine-Rich, P. (2022). Anti-Bacterial Protein against Vibrio cholerae, Staphylococcus aureus from Solanum trilobatum Leaves. Molecules, 27(4), 1167.
- Kannabiran, K., & Thanigaiarassu, R. R. (2008). Antibacterial Activity of Saponin Isolated from the Leaves of S. trilobatum Linn. Journal of Applied Biological Sciences, 2(3).
- Shahjahan, M., Vani, G., & Shyamaladevi, C. (2005). Effect of S. trilobatum on the antioxidant status during diethyl nitrosamine induced and phenobarbital promoted hepatocarcinogenesis in rat. Chemico-biological Interactions, 156(2), 113-123.
- Rajkumar, S., & Jebanesan, A. (2005). Oviposition deterrent and skin repellent activities of S. trilobatum leaf extract against the malarial vector Anopheles stephensi. Journal of Insect Science, 5(1), 15.
- Thirumalaiandi, R., Selvaraj, M. G., Rajasekaran, R., & Subbarayalu, M. (2008). Cloning and characterization of resistance gene analogs from under-exploited plant species. Electronic Journal of Biotechnology, 11(4), 4-5.
- Jantorn, P., Tipmanee, V., Wanna, W., Prapasarakul, N., Visutthi, M., & Sotthibandhu, D. S. (2023). Potential natural antimicrobial and antibiofilm properties of Piper betle L. against Staphylococcus pseudintermedius and methicillin-resistant strains. Journal of ethnopharmacology, 317, 116820.
- Gomes, F., Martins, N., Ferreira, I. C., & Henriques, M. (2019). Anti-biofilm activity of hydromethanolic plant extracts against Staphylococcus aureus isolates from bovine mastitis. Heliyon, 5(5). [CrossRef]
- Jeyarajan, S.; Ranjith, S.; Veerapandian, R.; Natarajaseenivasan, K.; Chidambaram, P.; & Kumarasamy, A. (2024). Antibiofilm Activity of Epinecidin-1 and Its Variants Against Drug-Resistant Candida krusei and Candida tropicalis Isolates from Vaginal Candidiasis Patients. Infect. Dis. Rep, 16, 1214-1229.
- Beynon, R.J., & Bond, J.S. (1989). Proteolytic enzymes: A practical approach. IRL Press at Oxford University Press.
- Nagampalli, R. S. K., Gunasekaran, K., Narayanan, R. B., Peters, A., & Bhaskaran, R. (2014). A structural biology approach to understand human lymphatic filarial infection. PLoS Neglected Tropical Diseases, 8(2), e2662. [CrossRef]
- Zanphorlin, L. M., Cabral, H., Arantes, E., Assis, D., Juliano, L., Juliano, M. A., ... & Bonilla-Rodriguez, G. O. (2011). Purification and characterization of a new alkaline serine protease from the thermophilic fungus Myceliophthora sp. Process Biochemistry, 46(11), 2137-2143.
- Damare S, Mishra A, D'Souza-Ticlo-Diniz D, Krishnaswamy A, & Raghukumar C. (2020). A deep-sea hydrogen peroxide-stable alkaline serine protease from Aspergillus flavus. 3 Biotech, 10(12), 528.
- Alagarasan, C., Ramya, K. S., Manohar, R., & Siva, G. V. (2018). PARTIAL PURIFICATION OF CYSTEINE PROTEASE FROM NIGELLA SATIVA L SEEDS.
- Karthik, L., Manohar, R., Elamparithi, K., & Gunasekaran, K. (2019). Purification, characterization and functional analysis of a serine protease inhibitor from the pulps of Cicer arietinum L.(Chick Pea). Indian Journal of Biochemistry and Biophysics (IJBB), 56(2), 117-124..
- Asif-Ullah, M., Kim, K.-S., & Yu, Y.G. (2006). Purification and characterization of a serine protease from Cucumis trigonus Roxburghi. Phytochemistry, 67(9), 870-875.
- Fan, T., Bykova, N.V., Rampitsch, C., & Xing, T. (2016). Identification and characterization of a serine protease from wheat leaves. European Journal of Plant Pathology, 1-12.
- Ahmed, I.A.M., Morishima, I., Babiker, E.E., & Mori, N. (2009). Dubiumin, a chymotrypsin-like serine protease from the seeds of Solanum dubium Fresen. Phytochemistry, 70(4), 483-491. [CrossRef]
- Alici, E. H., & Arabaci, G. (2018). A novel serine protease from strawberry (Fragaria ananassa): Purification and biochemical characterization. International journal of biological macromolecules, 114, 1295-1304.
- Nadaroglu, H., & Demır, N. (2012). Purification and characterization of a novel serine protease compositain from compositae (Scorzonera hispanica L.). European Food Research and Technology, 234, 945-953.48.
- Rawaliya, R. K., Patidar, P., Sharma, S., & Hajela, K. (2022). Purification and biochemical characterization of protease from the seeds of Cyamopsis tetragonoloba. J Appl Biol Biotechnol, 10, 172-180.
- Tomar, R., Kumar, R., & Jagannadham, M. V. (2008). A stable serine protease, wrightin, from the latex of the plant Wrightia tinctoria (Roxb.) R. Br.: purification and biochemical properties. Journal of agricultural and food chemistry, 56(4), 1479-1487. [CrossRef]
- Mehrnoush, A., Mustafa, S., Sarker, M.Z.I., & Yazid, A.M.M. (2011). Optimization of the conditions for extraction of serine protease from Kesinai Plant (Streblus asper) leaves using response surface methodology. Molecules, 16(11), 9245-9260. [CrossRef]
Figure 1.
a). Caseinolytic activity of the aqueous crude extract. (1. Positive control, 2-6, Negative controls), Well No. 1. 2mM Trypsin, 2. 20 mM Tris (7.8) + 150 mM NaCl (Extraction buffer), 3. 150 mM NaCl, 4. 20 mM Tris (7.8), 5. Water, 6. 1M NaCl, 7. Crude extract S. trilobatum 250 μM. b). Serine protease activity of the dialyzed fractions was obtained from UV Spectroscopic assay where casein was used as the substrate.
Figure 1.
a). Caseinolytic activity of the aqueous crude extract. (1. Positive control, 2-6, Negative controls), Well No. 1. 2mM Trypsin, 2. 20 mM Tris (7.8) + 150 mM NaCl (Extraction buffer), 3. 150 mM NaCl, 4. 20 mM Tris (7.8), 5. Water, 6. 1M NaCl, 7. Crude extract S. trilobatum 250 μM. b). Serine protease activity of the dialyzed fractions was obtained from UV Spectroscopic assay where casein was used as the substrate.
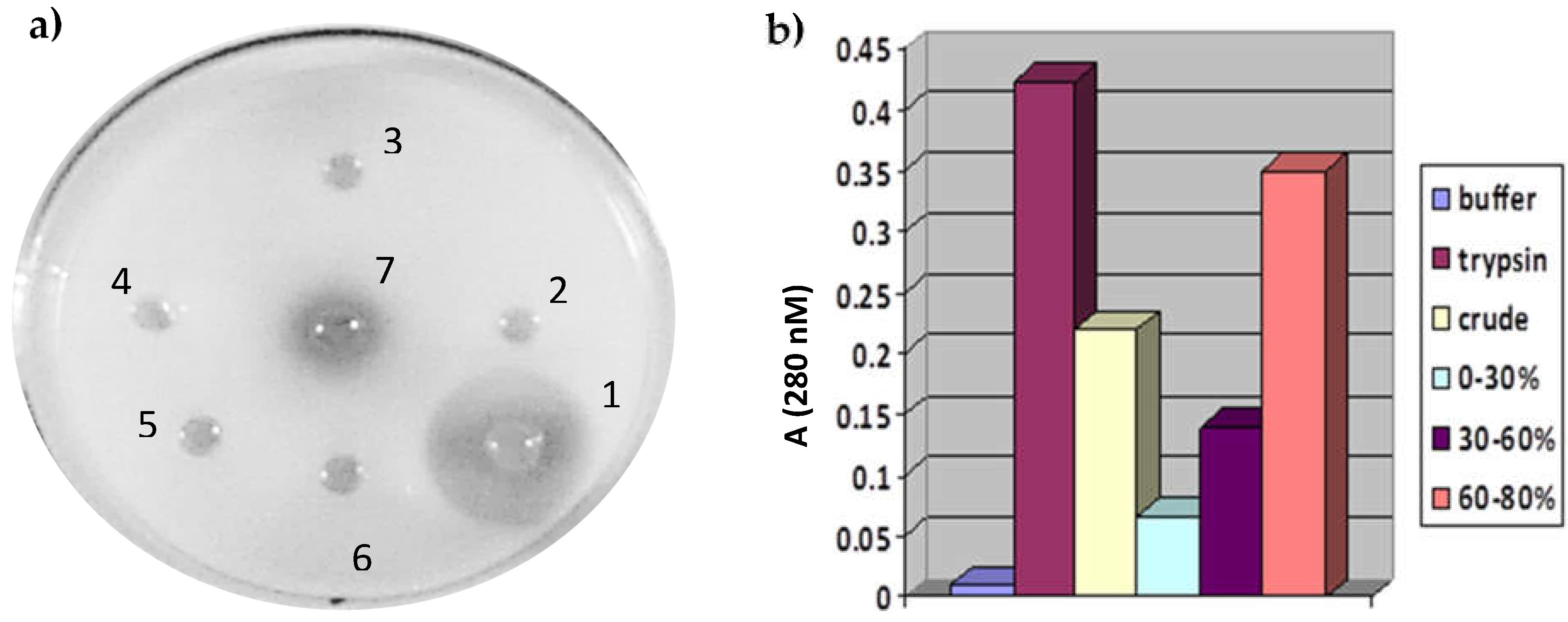
Figure 2.
a). Ion Exchange Chromatography (IEC) of the 60-80% Fraction: The green line represents the rise of conductivity by a salt linear gradient from 0 M to 1 M NaCl. (X-axis: 1ml/min, Y-axis: UV 280nm). b). Caseinolytic activity of IEC fractions: Well No. 1. IEC elution tube number 12, 2. IEC flow-through fraction, 3. 2mM Trypsin (Positive Control), 4. IEC elution 13, 5.IEC Elution Buffer 20mM tris+1M NaCl (Negative Control). c). IEC fractions on 10% SDS PAGE: Lane No. 1. Flow through, 2. 60-80% ammonium sulfate fraction, Elution 10, 4. Elution 11, 5. Elution 12, 6. Elution 13, 7. Elution 14. d). Purified StSP with standard protein ladder.
Figure 2.
a). Ion Exchange Chromatography (IEC) of the 60-80% Fraction: The green line represents the rise of conductivity by a salt linear gradient from 0 M to 1 M NaCl. (X-axis: 1ml/min, Y-axis: UV 280nm). b). Caseinolytic activity of IEC fractions: Well No. 1. IEC elution tube number 12, 2. IEC flow-through fraction, 3. 2mM Trypsin (Positive Control), 4. IEC elution 13, 5.IEC Elution Buffer 20mM tris+1M NaCl (Negative Control). c). IEC fractions on 10% SDS PAGE: Lane No. 1. Flow through, 2. 60-80% ammonium sulfate fraction, Elution 10, 4. Elution 11, 5. Elution 12, 6. Elution 13, 7. Elution 14. d). Purified StSP with standard protein ladder.
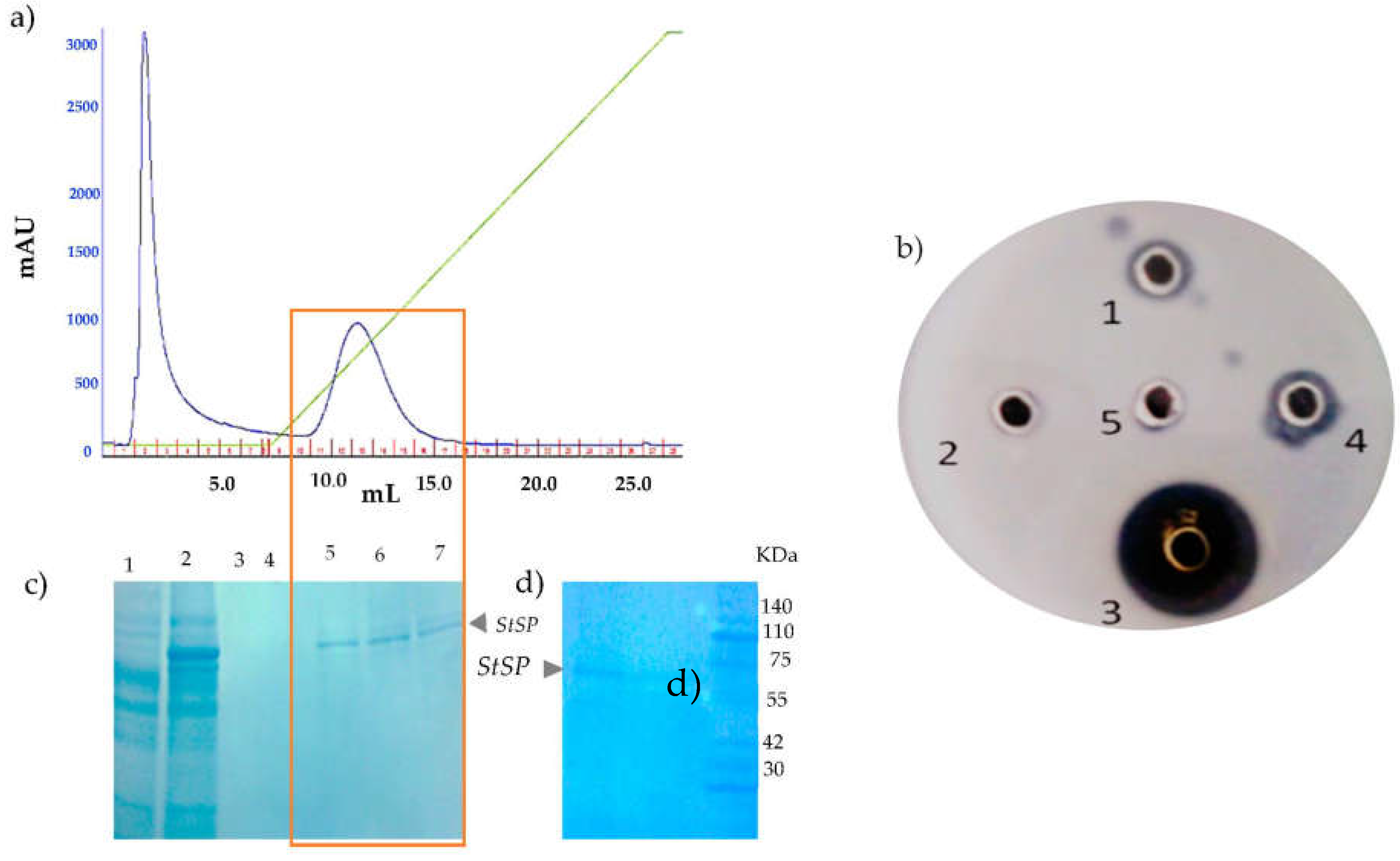
Figure 3.
a) Effect of metalloprotease inhibitor (EDTA) and cysteine protease inhibitor (IAA): Well No. 1. 100 µg purified protease incubated with 5 mM EDTA, 2. 100 µg Trypsin (Positive control), 3. 100 µg purified protease incubated with 2 mM IAA, 4. 100 µg purified protease, 5. Buffer (Negative control). b) Effect of Serine Protease Inhibitor (PMSF): Well No: 1.100 µg purified protease, 2. 100 µg purified protease preincubated with 2 mM PMSF, 3. 100 µg Trypsin (Positive control), 4. 100 µg purified protease, 5. 10 % Ethanol (Negative control), 6. Buffer (Negative control).
Figure 3.
a) Effect of metalloprotease inhibitor (EDTA) and cysteine protease inhibitor (IAA): Well No. 1. 100 µg purified protease incubated with 5 mM EDTA, 2. 100 µg Trypsin (Positive control), 3. 100 µg purified protease incubated with 2 mM IAA, 4. 100 µg purified protease, 5. Buffer (Negative control). b) Effect of Serine Protease Inhibitor (PMSF): Well No: 1.100 µg purified protease, 2. 100 µg purified protease preincubated with 2 mM PMSF, 3. 100 µg Trypsin (Positive control), 4. 100 µg purified protease, 5. 10 % Ethanol (Negative control), 6. Buffer (Negative control).
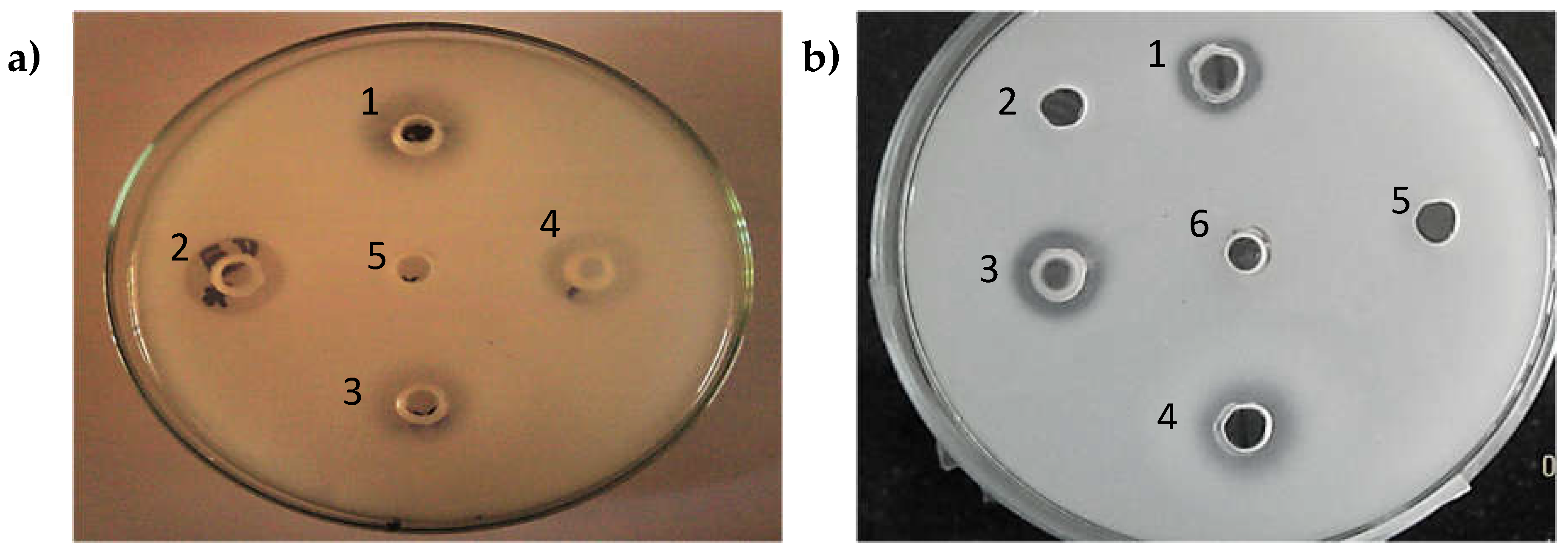
Figure 4.
The biofilm eradication potency of StSP. The control panels c) and g) shows the S.aureus-preformed biofilm on the surface of the microtiter plate at initial and after 12hr intervals of incubation. The samples (Serine protease, Serine protease+ampicillin, Ampicillin) treated panels showed in a), b) and d), respectively. The eradication potency of StSP samples after 12 hours of treatment were observed on panel e), f) and h).
Figure 4.
The biofilm eradication potency of StSP. The control panels c) and g) shows the S.aureus-preformed biofilm on the surface of the microtiter plate at initial and after 12hr intervals of incubation. The samples (Serine protease, Serine protease+ampicillin, Ampicillin) treated panels showed in a), b) and d), respectively. The eradication potency of StSP samples after 12 hours of treatment were observed on panel e), f) and h).
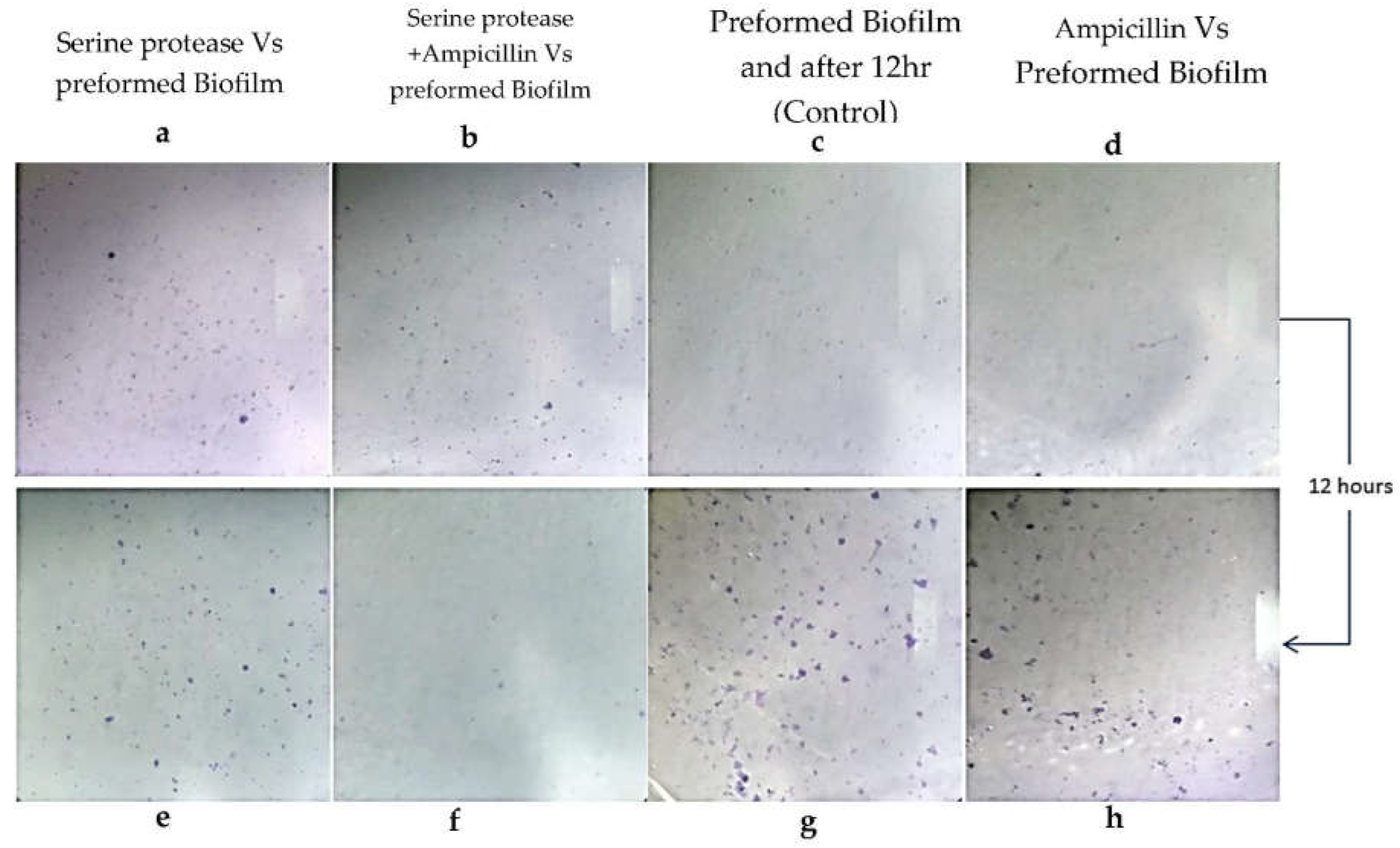
Figure 5.
a: Effect of Metal Ions towards StSP.
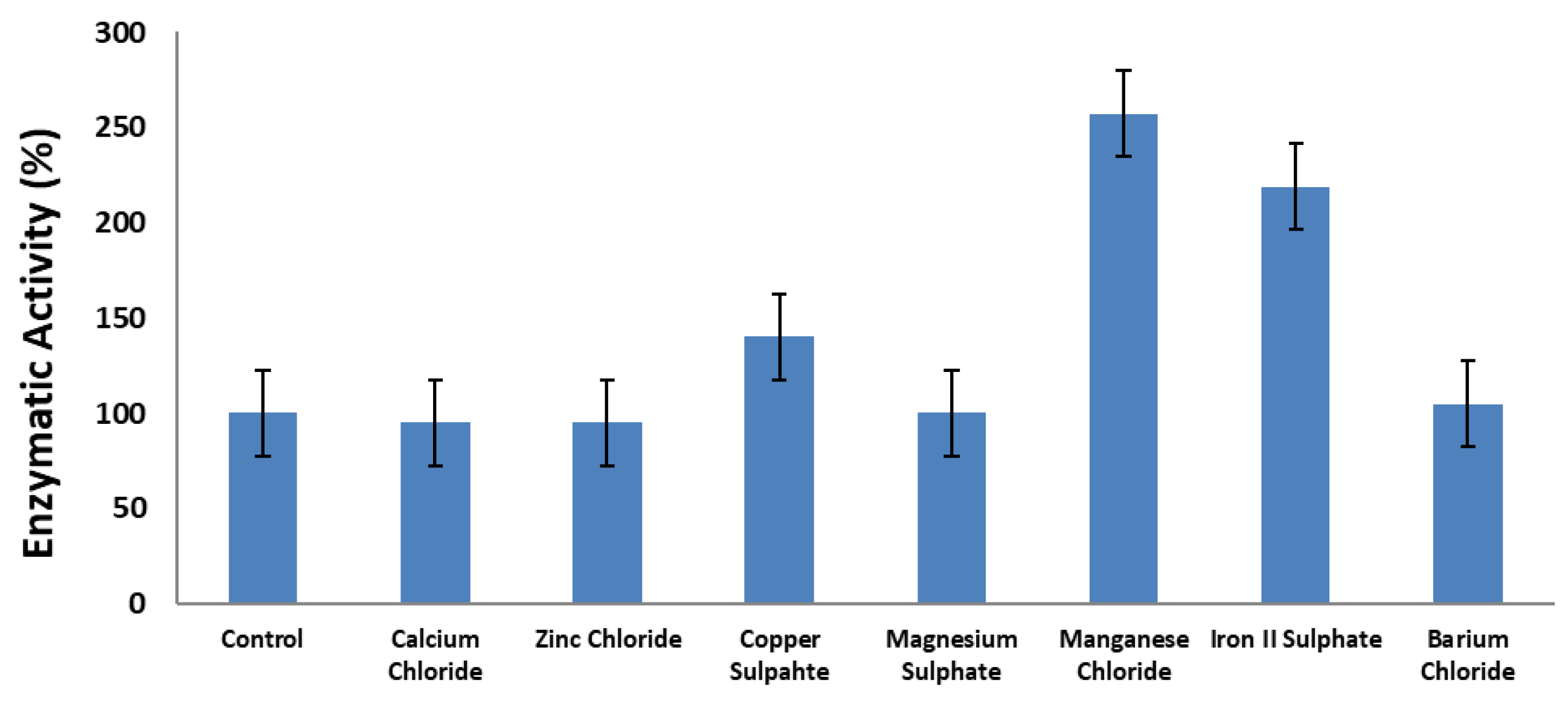
Figure 5.
b: Effect of Various Surfactants towards stSP.
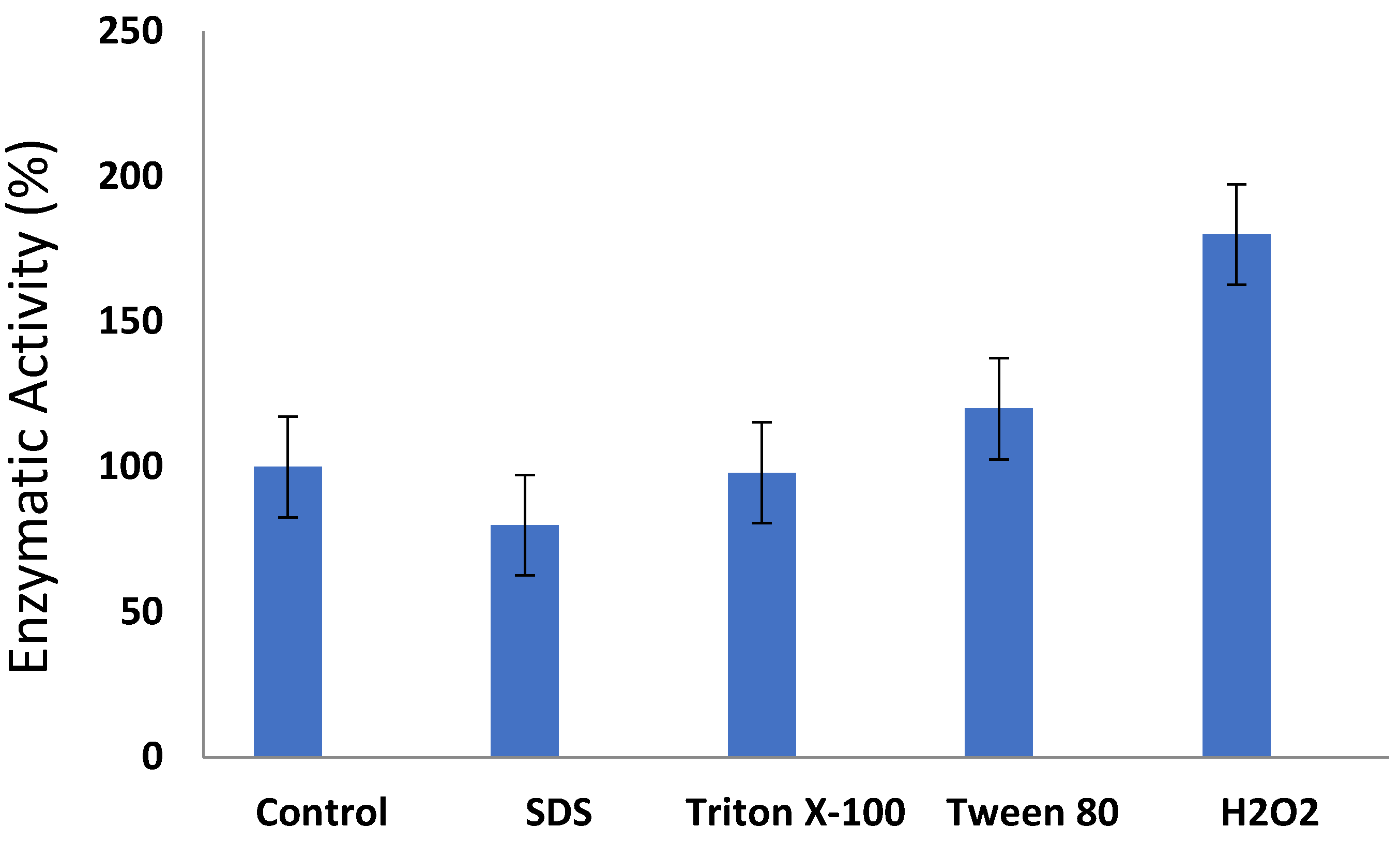
Figure 5.
c: Effect of Various Organic Solvents towards stSP.
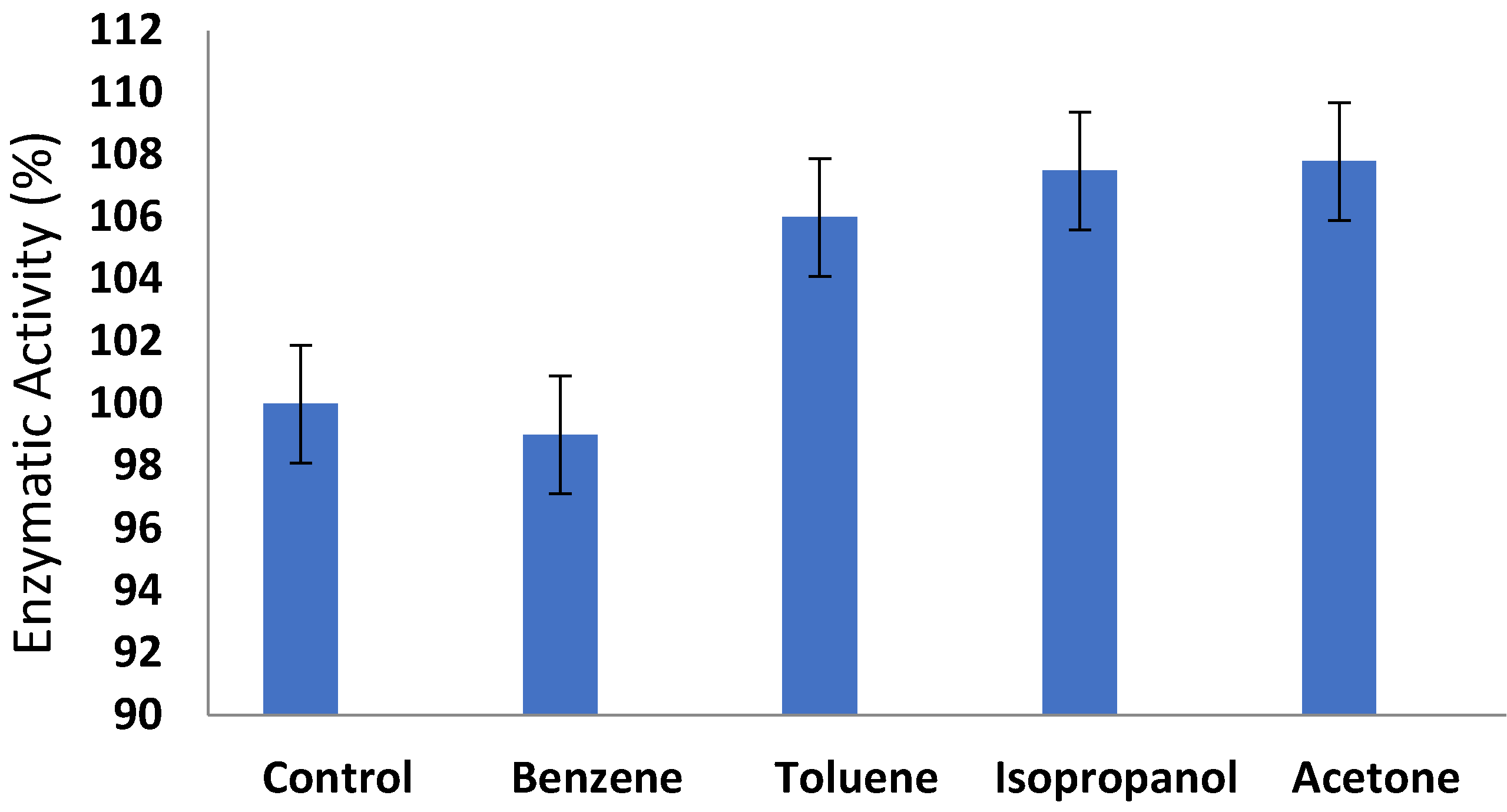
Figure 6.
Determination of kinetic parameters, Vmax & Km of the purified stSP.
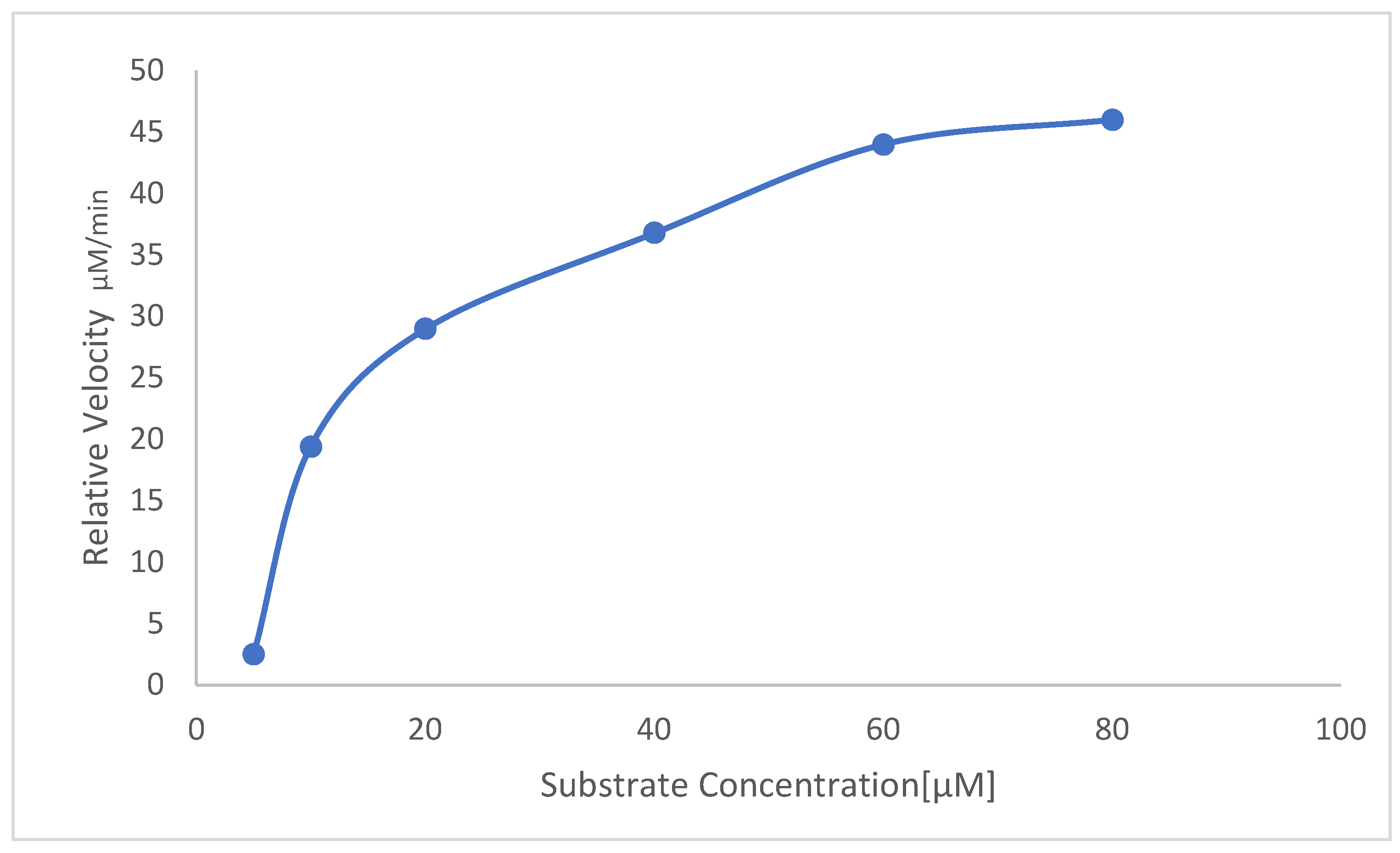
Figure 7.
a, b: Effect of pH and Temperature.
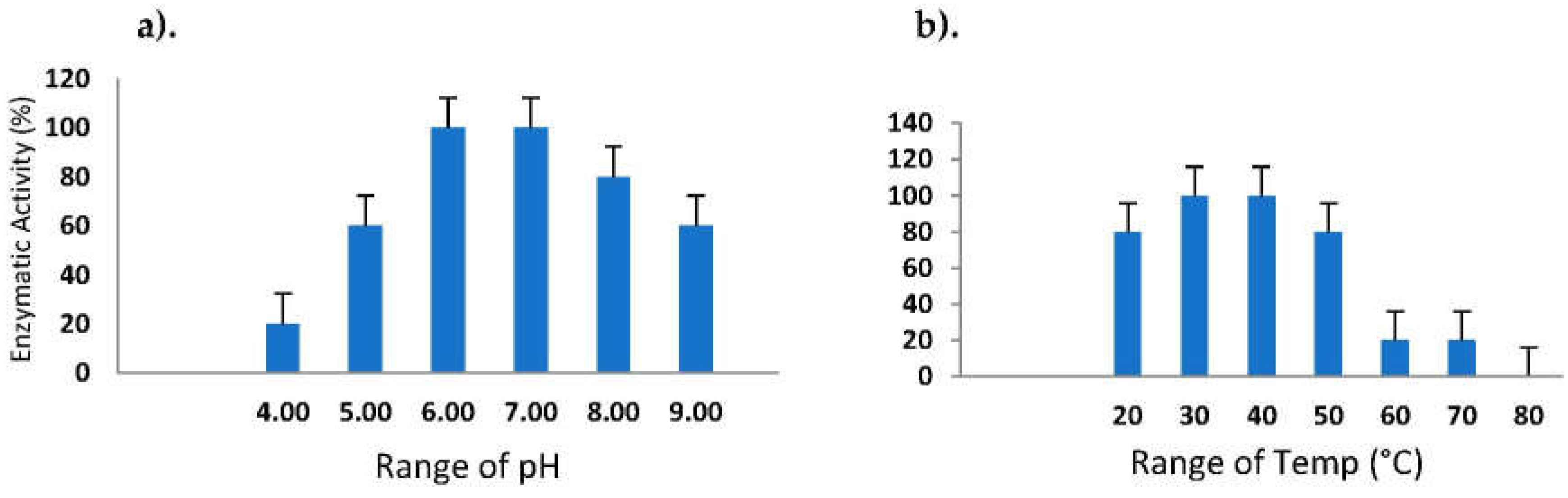
Figure 8.
a). Size exclusion chromatography of Serine Protease on G- 100 Sephadex resin. b). Caseinolytic activity of size exclusion fractions. c). Zymography: Lane 1. Purified serine protease, 2. 60-80% Ammonium Sulfate Fraction, 3. Aqueous extract of S. trilobatum leaves, 4. Purified and concentrated StSP.
Figure 8.
a). Size exclusion chromatography of Serine Protease on G- 100 Sephadex resin. b). Caseinolytic activity of size exclusion fractions. c). Zymography: Lane 1. Purified serine protease, 2. 60-80% Ammonium Sulfate Fraction, 3. Aqueous extract of S. trilobatum leaves, 4. Purified and concentrated StSP.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



