
Submitted:
07 April 2025
Posted:
08 April 2025
You are already at the latest version
Abstract
Beneficial interactions between nitrogen-fixing soil bacteria and legumes offer a solution to increase crop yield on Earth and potentially in future Martian colonies. In this study, we assessed the growth of the beta rhizobium Paraburkholderia phymatum in Martian simulant soil using Enhanced Mojave Mars Simulant 2 (MMS-2) that contains a high amount of iron (18.37 percent by weight) and aluminium (13.07 percent by weight). We observed that while P. phymatum wild-type’s growth was not affected by exposure to MMS-2, a mutant strain impaired in siderophore biosynthesis (ΔphmJK) grew less than P. phymatum wild-type on gradient plates prepared with increasing proportions of MMS-2 or aluminium concentration. This result suggests that the P. phymatum siderophore phymabactin alleviates aluminium-induced heavy metal stress. Using Ultra-high performance liquid chromatography-mass spectrometry (UHPLC MS), we showed that phymabactin can bind to aluminium more efficiently than iron. These results not only deepen our understanding of the behaviour of rhizobia in simulated extraterrestrial environments but also provide new insights into the potential use of P. phymatum for bioremediation and the multiple roles of the siderophore phymabactin.
Keywords:
siderophore
; rhizobium
; bioremediation
; metal
; space
; agriculture
1. Introduction
The prospect of extraterrestrial colonization requires the implementation and optimization of space farming to ensure that human settlements are as self-reliant as possible [1,2]. Indeed, supplying essential nutrients or goods to space settlers is increasingly difficult and expensive because of the planet’s great distance from Earth [1]. In addition, access to fresh and vitamin-rich food is crucial to human physical and psychological health [3,4]. Single-cell-based nutrition is critical in the long term due to its high nucleic acid content, which leads to uric acid formation and health problems such as gout or kidney stone formation in humans [5,6]. These concerns, therefore, motivate the need to develop and professionalize space farming. Yet, crops typically require the supplementation of nitrogen-based synthetic fertilizers to produce sufficient yields for human consumption [7,8]. One way to circumvent this issue and provide plants with soluble nitrogen is to take advantage of the beneficial relationship between legumes and nitrogen-fixing rhizobia [9]. Rhizobia are soil bacteria that can intracellularly colonize specialized legume root structures called nodules, where they convert atmospheric nitrogen into ammonia that can be used by the plant [9,10]. Nitrogen is the most limiting factor for plant growth and development since it is essential for synthesizing nucleic and amino acids, proteins, and chlorophyll [11,12]. Certain rhizobia also produce phytohormones like auxins and brassinosteroids that stimulate plant growth and root development [13,14,15,16,17]. Additionally, rhizobia can also have a health-protective effect on crops, either by directly stimulating their immune system or indirectly by preventing phytopathogen growth [18,19,20]. For example, some rhizobia produce siderophores that sequester iron from their microenvironment and prevent phytopathogens from obtaining it, thus inhibiting their growth [21]. Moreover, siderophores-producing soil bacteria were shown to protect plants against heavy metals-induced oxidative stress [22]. Indeed, certain types of siderophores can bind to heavy metals like aluminium, cadmium, copper, lead, and zinc, hence alleviating the stress induced by heavy-metal contamination in soil and thereby improving plant growth [23,24].
Paraburkholderia phymatum STM815T is a good model to study legume symbiosis, as this beta-rhizobium can nodulate more than 50 different legume species, including crops of human interest such as common bean and cowpea [15,25,26,27]. P. phymatum is also highly competitive against other soil bacteria in nodulating legume roots and shows remarkable abilities to survive abiotic stresses like those induced by salt or drought [15,26,28]. Importantly, we recently showed that P. phymatum can grow well in simulated microgravity and identified the phm gene cluster responsible for producing the hydroxamate-type siderophore phymabactin [29].
To determine the survival of P. phymatum under conditions that mimic extraterrestrial life, we tested its growth on a Martian soil simulant using the Enhanced Mojave Mars Simulant 2 (MMS-2). Indeed, data collected by different Mars exploration programs (Viking, Pathfinder, Spirit, and Opportunities) showed that heavy metals are present in high amounts on the surface of Mars [30,31]. It is estimated that there is between 18.5 and 21.7 percent by weight (wt%) iron on the Martian crust, while aluminium accounts for between 7.3 and 12.3 wt% [30]. By comparison, iron accounts for roughly 7 wt% and aluminium for 8 wt% of Earth’s crustal composition [32,33]. This study shows that a previously constructed P. phymatum strain, unable to produce the siderophore phymabactin, grew less than the wild-type strain when exposed to iron and aluminium-rich MMS-2. However, while the phm mutant was also growth-impaired in a medium supplemented with aluminium (102.4 mM), in a medium supplemented with 92 mM iron, the phm mutant was not affected in growth, suggesting that phymabactin is mainly neutralizing the toxic effects of aluminium. Finally, mass spectrometry analyses revealed that phymabactin extracted from the supernatant of P. phymatum can bind to aluminium even stronger than to iron. These results suggest that phymabactin production is not only beneficial for iron scavenging but also for binding to other heavy metals, thereby making P. phymatum a suitable candidate for space farming in Martian soil and bioremediation of aluminium-rich soils.
2. Materials and Methods
2.1. Bacterial Strains, Media, and Cultivation
The bacterial strains and the antibiotics used in this study are listed in Supplementary Table S1. P. phymatum STM815 strains were grown in Luria-Bertani (LB) rich medium prepared without salt (LB-NaCl) or in AB minimal medium [34] with 15 mM of succinate (Sigma-Aldrich, St. Louis, MO, USA) as a carbon source. LB-NaCl was supplemented with 40 g/L of enhanced Mojave Mars Simulant 2 (MMS-2; The Martian Garden, Austin, Texas) to grow P. phymatum in artificial Martian soil. Trimethoprim was used to select the P. phymatum phmJK mutant (100 µg/mL).
2.2. Preparation of Linear Gradient Plates
P. phymatum’s growth was tested on MMS-2 using the linear gradient plate technique [35]. For this, two agar layers were poured into 12 cm square plates. First, 20 mL of LB-NaCl supplemented with 40 g/ L of MMS-2 or with 102,4 mM of AlCl3 (Sigma-Aldrich, St. Louis, MO, USA) or 92 mM FeCl3 (Sigma-Aldrich, St. Louis, MO, USA) was poured into tilted squared plates to form the bottom layer. After roughly 20 minutes, the second layer containing 20 mL of LB-NaCl was poured on the plate horizontally to cover the bottom layer. In this way, the proportion of MMS-2 or the concentration of AlCl3 or FeCl3 gradually increased along the horizontal axis. Pre-cultures of tested strains were grown in LB-NaCl media, washed twice, and set to an OD600 of 0.5. Sterile cotton swabs were used to draw lines on the gradient plates, alternating the strains to avoid variations due to plate irregularities. The plates were incubated for 48 hours at 28 °C.
2.3. Sample Preparation for Siderophore Screening Analysis
Bacterial supernatants of P. phymatum wild-type and ΔphmJK were prepared in triplicates and subjected to a siderophore screening analysis by UHPLC-MS. Therefore, 200 µL of bacterial supernatant was frozen using liquid nitrogen and lyophilized overnight in a vacuum concentrator at 8 °C. The dry lyophilizate was reconstituted in 300 µL of H2O/CH3OH (3:2) and centrifuged for 10 min at 5 °C and 14’000 rpm. Three LC-MS vials were filled with 100 µL of supernatant. One vial was spiked with 1 µL of an aqueous 100 mM FeCl3 solution, another vial was spiked with 1 µL of an aqueous 100 mM AlCl3 solution, and the last vial was not spiked. These samples were analysed by UHPLC-MS to investigate the formation of iron-phymabactin and aluminium-phymabactin complexes.
2.4. UHPLC-MS Method
Samples were analysed using a Vanquish Horizon UHPLC system (Thermo Fisher, Waltham, MA, USA) connected to a timsTOF Pro HR-QTOF mass spectrometer (Bruker, Bremen, Germany). The Vanquish Horizon UHPLC system was built from a binary pump H, a split sampler HT, and a temperature-controlled column compartment. Chromatographic separation was performed at 40 °C with an ACQUITY HSS T3 UPLC column (100 Å, 1.8 µm particle size, 2.1×100 mm, Waters, Milford, USA). The injection volume was 1 µL. The mobile phase consisted of A: Ultrapure H2O + 0.1% HCOOH and B: CH3CN + 0.1% HCOOH. A constant flow rate of 0.5 mL/min was applied using the following gradient: (I) 5% B isocratic from 0.0 to 0.5 min; (II) linear increase to 45% B from 0.5 to 6.0 min; (III) linear increase to 100% B from 6.0 to 6.1 min; (IV) 100% B isocratic from 6.1 to 9.0 min; (V) linear decrease to 5% B from 9.0 to 9.1 min; (VI) 5% B isocratic from 9.1 to 12.0 min. The mass spectrometer was operated in the positive ESI mode at 4500 V capillary voltage and 500 V endplate offset with an N2 nebulizer pressure of 2.2 bar and a dry gas flow of 10 L/min at 220 °C. Mass spectra were acquired in a mass range from m/z 20 to 1300 at a resolution of 40’000 (m/z 431 full width at half maximum) and a 12 Hz acquisition rate. Mass measurements were externally calibrated between m/z 118 and 1222 using an ESI-L Low Concentration tuning mix (Agilent, Santa Clara, USA). Internal mass calibration was performed at the beginning of each LC run between m/z 91 and 1247 using a 10 mM solution of sodium formate that was injected using a 6-port valve with a 20 µL loop giving a mass accuracy below 2 ppm.
3. Results
3.1. The Siderophore Phymabactin Is Important for the Growth of P. phymatum in Martian Soil and in Aluminium-Rich Medium
We previously showed that P. phymatum STM815T grows in simulated microgravity. In this study, we explore i) the ability of this strain to grow on an iron and aluminium-rich Martian soil simulant (MMS-2) and ii) the importance of P. phymatum siderophore phymabactin under these conditions. For this purpose, wild-type and the siderophore mutant ΔphmJK were grown on LB-NaCl gradient plates containing an increasing proportion of the Martian soil simulant MMS-2. In contrast to P. phymatum wild-type, a strain unable to produce the siderophore phymabactin (ΔphmJK) was not able to grow when cultivated with a high proportion of MMS-2 (Figure 1a), as well as in the presence of high concentrations of aluminium (Figure 1b). However, P. phymatum wild-type and ΔphmJK showed a similar growth on gradient plates prepared with 92 mM of FeCl3 (Figure 1c). As a control, both wild-type and mutant strains were inoculated on LB-NaCl plates and showed no difference in growth (data not shown).
3.2. Phymabactin Chelates Fe(III) and Al(III)
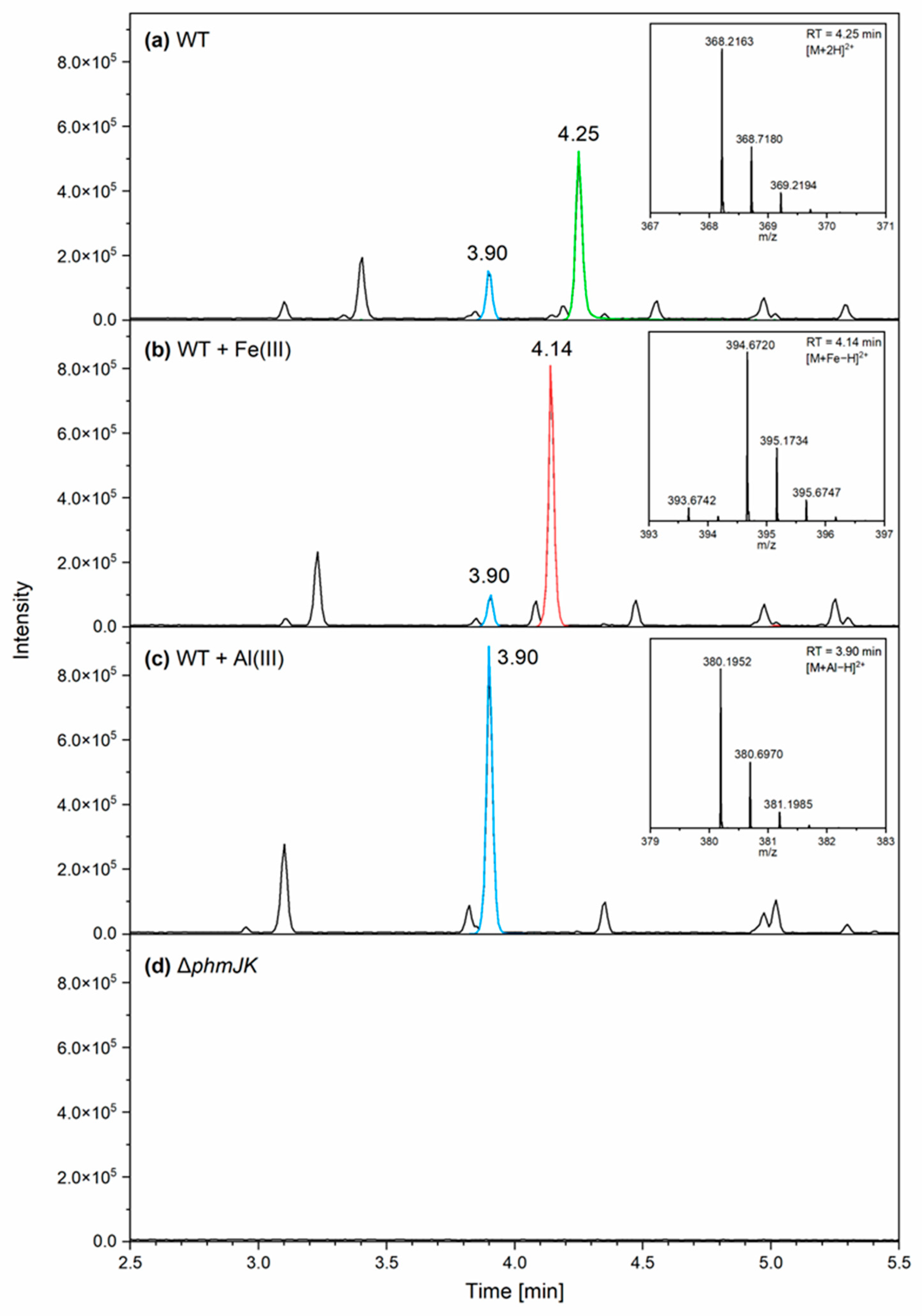
Supernatants of P. phymatum wild-type and ΔphmJK cells grown in minimal medium ABS prepared without iron were subjected to a siderophore screening analysis by UHPLC-MS. As shown in Figure 2, specific retention times, mass-to-charge ratio (m/z) shifts, and characteristic isotopic pattern distributions were observed for the complexation of phymabactin (Figure 2a) with iron (Figure 2b) and aluminium (Figure 2c). While the unbound siderophore had a retention time of 4.25 minutes, the Fe(III)-phymabactin complex displayed a retention time of 4.14 min, and the aluminium-phymabactin complex had a retention time of 3.90 min. The Al-phymabactin complex was detected in the P. phymatum sample spiked with FeCl3 or AlCl3, whereas the iron-phymabactin complex was only observed in the P. phymatum sample spiked with FeCl3. No siderophores or siderophore-metal complexes were detected in the supernatant of ΔphmJK cells (Figure 2d). These results suggest that phymabactin is binding to both metals with a stronger affinity to aluminium.
4. Discussion
P. phymatum is a soil bacterium that displays important traits such as reducing atmospheric nitrogen into ammonium in symbiosis with plants and in free-living conditions. Moreover, this strain can enter symbiosis with an unusually large number of plants, produces plant-like hormones, and is very resistant to abiotic and biotic stresses. We have recently shown that P. phymatum can grow well even under simulated microgravity conditions, making it an ideal candidate for space farming. In this study, we tested the growth of P. phymatum strains in Martian simulant soil (MMS-2), which is rich in iron (18.4 wt%) and aluminium (13.1 wt%). Indeed, Mars contains a high percentage of metals such as iron (Fe2O3; 19.2 wt%), aluminium (Al2O3; 9.4 wt%), chromium (Cr2O3; 0.5 wt%), and magnesium (MgO; 8.7 wt%) [36,37,38]. In comparison, on Earth, there are 4.7 wt% of Fe, 8.1 wt% of Al, 0.008 wt% of Cr, and 1.9 wt% of Mg [39]. In our study, we observed that the siderophore produced by P. phymatum (phymabactin) contributed to the growth of this strain in the Martian soil (Figure 1a). To test if the difference in growth between P. phymatum wild-type and a phymabactin mutant was due to the high concentration of heavy metal present in MMS-2, we grew the strains on linear gradient plates containing increasing concentrations of iron and aluminium, up to the same final concentration present in MMS-2. To our surprise, the siderophore phymabactin was found to be important for growth with high aluminium concentrations but did not play a role in P. phymatum fitness in an iron-rich environment (Figure 1c). This result may be explained by an inhibitory effect of high iron concentrations on P. phymatum siderophore production. Indeed, a potential ferric uptake regulation (Fur) binding sequence is localized upstream of the gene cluster responsible for phymabactin production [29], suggesting that the transcriptional repressor Fur binds to the phm promoter and regulates intracellular iron homeostasis. A similar effect has been observed in Burkholderia cenocepacia 715J, where the production of the siderophore ornibactin is negatively affected by iron supplementation [40]. The fact that ΔphmJK grows less than the wild-type strain in aluminium-rich media (Figure 1b) suggests that the metal does not hinder siderophore production and that aluminium is the component that prevents the growth of ΔphmJK in Martian simulant soil. Furthermore, we also observed that ΔphmJK grew less than P. phymatum wild-type when cultured in an aerated liquid medium and exposed to other metals such as zinc, lead, and copper, showing a similar behaviour after the addition of aluminium (Supplementary figure 1a-1d). Previous studies showed that a Burkholderia cenocepacia H111 siderophore mutant (not producing pyochelin and ornibactin) was affected in growth in the presence of these divalent cations [41]. P. phymatum wild-type grew even in the highest concentration of iron- and aluminium-rich Martian soil (Figure 1a-1c). This result can be explained by the fact that, in MMS-2, iron and aluminium are present as ferric oxide (Fe2O3) and aluminium oxide (Al2O3). In contrast, in the gradient plates assay, iron and aluminium were added as FeCl3 and AlCl3, respectively. Indeed, ferric oxide and aluminium oxide are less soluble than their chloride counterpart at neutral pH and, therefore, release fewer ions in their microenvironment, reducing their toxicity to microorganisms [42,43]. We showed here that phymabactin binds to aluminium and iron with retention times of 3.90 minutes and 4.14 minutes, respectively (Figure 2b and 2c), suggesting that -in addition to the iron scavenging function. P. phymatum phymabactin relieves the cell of metal-induced stress.
Siderophores-producing soil bacteria have been shown to reduce heavy metal toxicity by capturing the metal in soil or increasing plants’ systemic resistance, making them more resilient [22]. Heavy metals such as copper, cadmium, or lead are human activity’s common byproducts and phytotoxic compounds that cause plant growth inhibition and even death [44]. Yet, soybean grown in lead-supplemented growth media and inoculated with the siderophore-producing and plant growth-promoting rhizobacteria (PGPR) Pseudomonas putida K9P9 showed a 70.6% and 28.6% increase in shoot and root length, respectively, compared to the uninoculated control [45]. A similar effect was observed in Paraburkholderia fungorum BRRh-4 that alleviated the stress of rapeseed grown in cadmium-contaminated soil and induced a 40% increase in height compared to the uninoculated control [46].
In the future, studies looking at the symbiotic and nitrogen-fixation efficiency of P. phymatum with legumes cultivated in MMS-2 or growth media containing heavy metals will help to understand the role of PGPR in bioremediation. Finally, this study provided new insights into the potential of P. phymatum as an inoculant in extraterrestrial environments such as Martian soil. It also revealed the role of the siderophore phymabactin as a possible bioremediation tool to protect plants from heavy metals, both on Earth and in extraterrestrial environments.
5. Conclusions
Our study unveiled the significance of P. phymatum’s siderophore production in its ability to thrive in Martian-like soil. We used Martian soil simulant media and showed that P. phymatum strains capable of producing phymabactin had a growth advantage compared to a mutant strain lacking this ability. This growth advantage is attributed to the high aluminium content in the simulated Martian soil. Notably, phymabactin demonstrated a superior capacity to bind aluminium ions compared to iron, as confirmed through UHPLC-MS analysis. These findings reveal that beyond its nitrogen-fixing capabilities across diverse legume species, P. phymatum demonstrates remarkable potential for thriving in the metal-rich soils characteristic of Mars and in decontaminating soils with high metal content.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Supplementary Figure S1; Supplementary Table S1; Supplementary Table S2.
Author Contributions
The following author contributions were made. Conceptualization: D.G. and G.P. Methodology: D.G, L. Bu., L. Bi and G.P. Investigation: D.G. and L. Bu. Data analysis: D.G., L. Bu., M.E., L. Bi. and G.P. Visualization: D.G., L. Bu., M.E. L. Bi. and G.P. Funding acquisition: M.E., L. Bi. and G.P. Writing: D.G., L. Bu., M.E., L. Bi. and G.P.
Funding
This work was supported by the Swiss Space PRODEX proposal FISURI (ESA PRODEX ARRANGEMENT No 4000144510) to Marcel Egli and the Swiss National Science Foundation (SNFS) with project number 310030_215282 to Gabriella Pessi.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Bernd Rattenbacher for providing MMS soil. We acknowledge Yilei Liu and Julie Ahrens for their help in preparing microbiology material. We would like to thank Christian Schwander for his support in optimizing the chromatographic method as part of his bachelor’s thesis.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| MMS-2 | enhanced Mojave Mars Simulant 2 |
| UHPLC MS | Ultra-high performance liquid chromatography-mass spectrometry |
| wt% | Percent per weight |
References
- Nguyen, M.; Knowling, M.; Tran, N.N.; Burgess, A.; Fisk, I.; Watt, M.; Escribà-Gelonch, M.; This, H.; Culton, J.; Hessel, V. Space farming: Horticulture systems on spacecraft and outlook to planetary space exploration. Plant Physiol. Biochem. 2023, 194, 708–721. [Google Scholar] [CrossRef]
- Iovane, M.; Izzo, L.G.; Romano, L.E.; Aronne, G. Simulated microgravity affects directional growth of pollen tubes in candidate space crops. Front. Plant Sci. 2023, 14. [Google Scholar] [CrossRef]
- Smith, S.M.; Lane, H.W.; Zwart, S.R. Spaceflight Metabolism and Nutritional Support, 413–439. [CrossRef]
- Odeh, R.; Guy, C.L. Gardening for Therapeutic People-Plant Interactions during Long-Duration Space Missions. Open Agriculture 2017, 2, 1–13. [Google Scholar] [CrossRef]
- Nasseri, A.T.; Rasoul-Ami, S.; Morowvat, M.H.; Ghasemi, Y. Single Cell Protein: Production and Process. American J. of Food Technology 2011, 6, 103–116. [Google Scholar] [CrossRef]
- Li, Y.P.; Ahmadi, F.; Kariman, K.; Lackner, M. Recent advances and challenges in single cell protein (SCP) technologies for food and feed production. NPJ Sci. Food 2024, 8, 66. [Google Scholar] [CrossRef]
- Tei, F.; Neve, S. de; Haan, J. de; Kristensen, H.L. Nitrogen management of vegetable crops. Agricultural Water Management 2020, 240, 106316. [Google Scholar] [CrossRef]
- Oyetunji, O.; Bolan, N.; Hancock, G. A comprehensive review on enhancing nutrient use efficiency and productivity of broadacre (arable) crops with the combined utilization of compost and fertilizers. J. Environ. Manage. 2022, 317, 115395. [Google Scholar] [CrossRef] [PubMed]
- Lindström, K.; Mousavi, S.A. Effectiveness of nitrogen fixation in rhizobia. Microb. Biotechnol. 2020, 13, 1314–1335. [Google Scholar] [CrossRef]
- Masson-Boivin, C.; Sachs, J.L. Symbiotic nitrogen fixation by rhizobia-the roots of a success story. Curr. Opin. Plant Biol. 2018, 44, 7–15. [Google Scholar] [CrossRef]
- Bellés-Sancho, P.; Beukes, C.; James, E.K.; Pessi, G. Nitrogen-Fixing Symbiotic Paraburkholderia Species: Current Knowledge and Future Perspectives. Nitrogen 2023, 4, 135–158. [Google Scholar] [CrossRef]
- Liu, X.; Hu, B.; Chu, C. Nitrogen assimilation in plants: current status and future prospects. J. Genet. Genomics 2022, 49, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Vargas, L.K.; Volpiano, C.G.; Lisboa, B.B.; Giongo, A.; Beneduzi, A.; Passaglia, L.M.P. Potential of Rhizobia as Plant Growth-Promoting Rhizobacteria, 153–174. [CrossRef]
- Bellés-Sancho, P.; Liu, Y.; Heiniger, B.; Salis, E. von; Eberl, L.; Ahrens, C.H.; Zamboni, N.; Bailly, A.; Pessi, G. A novel function of the key nitrogen-fixation activator NifA in beta-rhizobia: Repression of bacterial auxin synthesis during symbiosis. Front. Plant Sci. 2022, 13, 991548. [Google Scholar] [CrossRef]
- Bellés-Sancho, P.; Lardi, M.; Liu, Y.; Hug, S.; Pinto-Carbó, M.A.; Zamboni, N.; Pessi, G. Paraburkholderia phymatum Homocitrate Synthase NifV Plays a Key Role for Nitrogenase Activity during Symbiosis with Papilionoids and in Free-Living Growth Conditions. Cells 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, B.J.; Mathesius, U. Phytohormone regulation of legume-rhizobia interactions. J. Chem. Ecol. 2014, 40, 770–790. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.Z.; Huang, J.; Gyaneshwar, P.; Zhao, D. Rhizobium sp. IRBG74 Alters Arabidopsis Root Development by Affecting Auxin Signaling. Front. Microbiol. 2017, 8, 2556. [Google Scholar] [CrossRef]
- Li, B.; Dong, C.; Chu, Z.; Zhang, W.; Wang, M.; Liu, H.; Xie, B. Synthesis, characterization and application of ion exchange resin as a slow-release fertilizer for wheat cultivation in space. Acta Astronautica 2016, 127, 579–586. [Google Scholar] [CrossRef]
- Hug, S.; Liu, Y.; Heiniger, B.; Bailly, A.; Ahrens, C.H.; Eberl, L.; Pessi, G. Differential Expression of Paraburkholderia phymatum Type VI Secretion Systems (T6SS) Suggests a Role of T6SS-b in Early Symbiotic Interaction. Front. Plant Sci. 2021, 12, 699590. [Google Scholar] [CrossRef]
- Hug, S.; Heiniger, B.; Bolli, K.; Paszti, S.; Eberl, L.; Ahrens, C.H.; Pessi, G. Paraburkholderia sabiae Uses One Type VI Secretion System (T6SS-1) as a Powerful Weapon against Notorious Plant Pathogens. Microbiol. Spectr. 2023, 11, e0162223. [Google Scholar] [CrossRef]
- Geetha, S.J.; Joshi, S.J. Engineering rhizobial bioinoculants: a strategy to improve iron nutrition. ScientificWorldJournal. 2013, 2013, 315890. [Google Scholar] [CrossRef]
- Huo, Y.; Kang, J.P.; Ahn, J.C.; Kim, Y.J.; Piao, C.H.; Yang, D.U.; Yang, D.C. Siderophore-producing rhizobacteria reduce heavy metal-induced oxidative stress in Panax ginseng Meyer. J. Ginseng Res. 2021, 45, 218–227. [Google Scholar] [CrossRef]
- Fahde, S.; Boughribil, S.; Sijilmassi, B.; Amri, A. Rhizobia: A Promising Source of Plant Growth-Promoting Molecules and Their Non-Legume Interactions: Examining Applications and Mechanisms. Agriculture 2023, 13, 1279. [Google Scholar] [CrossRef]
- Schalk, I.J.; Hannauer, M.; Braud, A. New roles for bacterial siderophores in metal transport and tolerance. Environ. Microbiol. 2011, 13, 2844–2854. [Google Scholar] [CrossRef] [PubMed]
- Bellés-Sancho, P.; Lardi, M.; Liu, Y.; Eberl, L.; Zamboni, N.; Bailly, A.; Pessi, G. Metabolomics and Dual RNA-Sequencing on Root Nodules Revealed New Cellular Functions Controlled by Paraburkholderia phymatum NifA. Metabolites 2021, 11. [Google Scholar] [CrossRef]
- Campos, S.B. de; Lardi, M.; Gandolfi, A.; Eberl, L.; Pessi, G. Mutations in Two Paraburkholderia phymatum Type VI Secretion Systems Cause Reduced Fitness in Interbacterial Competition. Front. Microbiol. 2017, 8, 2473. [Google Scholar] [CrossRef]
- Lardi, M.; Campos, S.B. de; Purtschert, G.; Eberl, L.; Pessi, G. Competition Experiments for Legume Infection Identify Burkholderia phymatum as a Highly Competitive β-Rhizobium. Front. Microbiol. 2017, 8, 1527. [Google Scholar] [CrossRef]
- Lardi, M.; Pessi, G. Functional Genomics Approaches to Studying Symbioses between Legumes and Nitrogen-Fixing Rhizobia. High Throughput 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Golaz, D.; Papenfuhs, C.K.; Bellés-Sancho, P.; Eberl, L.; Egli, M.; Pessi, G. RNA-seq analysis in simulated microgravity unveils down-regulation of the beta-rhizobial siderophore phymabactin. NPJ Microgravity 2024, 10, 44. [Google Scholar] [CrossRef]
- Peters, G.H.; Abbey, W.; Bearman, G.H.; Mungas, G.S.; Smith, J.A.; Anderson, R.C.; Douglas, S.; Beegle, L.W. Mojave Mars simulant—Characterization of a new geologic Mars analog. Icarus 2008, 197, 470–479. [Google Scholar] [CrossRef]
- B A E Lehner; C N Haenggi; J Schleppi; S J J Brouns; A Cowley. Bacterial modification of lunar and Martian regolith for plant growth in life support systems 2018.
- Yan, L.; Riaz, M.; Li, S.; Cheng, J.; Jiang, C. Harnessing the power of exogenous factors to enhance plant resistance to aluminum toxicity; a critical review. Plant Physiol. Biochem. 2023, 203, 108064. [Google Scholar] [CrossRef]
- Peys, A.; Isteri, V.; Yliniemi, J.; Yorkshire, A.S.; Lemougna, P.N.; Utton, C.; Provis, J.L.; Snellings, R.; Hanein, T. Sustainable iron-rich cements: Raw material sources and binder types. Cement and Concrete Research 2022, 157, 106834. [Google Scholar] [CrossRef]
- Clark, D.J.; Maaløe, O. DNA replication and the division cycle in Escherichia coli. Journal of Molecular Biology 1967, 23, 99–112. [Google Scholar] [CrossRef]
- Szybalski, W. Gradient plates for the study of microbial resistance to antibiotics. Bacteriological Proceedings 1952.
- Costa, N.; Bonetto, A.; Ferretti, P.; Casarotto, B.; Massironi, M.; Altieri, F.; Nava, J.; Favero, M. Analytical data on three Martian simulants. Data Brief 2024, 57, 111099. [Google Scholar] [CrossRef]
- Cannon, K.M.; Britt, D.T.; Smith, T.M.; Fritsche, R.F.; Batcheldor, D. Mars global simulant MGS-1: A Rocknest-based open standard for basaltic martian regolith simulants. Icarus 2019, 317, 470–478. [Google Scholar] [CrossRef]
- Achilles, C.N.; Downs, R.T.; Ming, D.W.; Rampe, E.B.; Morris, R.V.; Treiman, A.H.; Morrison, S.M.; Blake, D.F.; Vaniman, D.T.; Ewing, R.C.; et al. Mineralogy of an active eolian sediment from the Namib dune, Gale crater, Mars. JGR Planets 2017, 122, 2344–2361. [Google Scholar] [CrossRef]
- Yaroshevsky, A.A. Abundances of chemical elements in the Earth’s crust. Geochem. Int. 2006, 44, 48–55. [Google Scholar] [CrossRef]
- Agnoli, K.; Lowe, C.A.; Farmer, K.L.; Husnain, S.I.; Thomas, M.S. The ornibactin biosynthesis and transport genes of Burkholderia cenocepacia are regulated by an extracytoplasmic function sigma factor which is a part of the Fur regulon. J. Bacteriol. 2006, 188, 3631–3644. [Google Scholar] [CrossRef]
- Mathew, A.; Jenul, C.; Carlier, A.L.; Eberl, L. The role of siderophores in metal homeostasis of members of the genus Burkholderia. Environ. Microbiol. Rep. 2016, 8, 103–109. [Google Scholar] [CrossRef]
- Bhuvaneshwari, M.; Bairoliya, S.; Parashar, A.; Chandrasekaran, N.; Mukherjee, A. Differential toxicity of Al2O3 particles on Gram-positive and Gram-negative sediment bacterial isolates from freshwater. Environ. Sci. Pollut. Res. Int. 2016, 23, 12095–12106. [Google Scholar] [CrossRef]
- Botté, A.; Zaidi, M.; Guery, J.; Fichet, D.; Leignel, V. Aluminium in aquatic environments: abundance and ecotoxicological impacts. Aquat Ecol 2022, 56, 751–773. [Google Scholar] [CrossRef]
- Tripathi, M.; Munot, H.P.; Shouche, Y.; Meyer, J.M.; Goel, R. Isolation and functional characterization of siderophore-producing lead- and cadmium-resistant Pseudomonas putida KNP9. Curr. Microbiol. 2005, 50, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, M.; Kumar, S.; Makarana, G.; Goel, R. Metal-Tolerant Bioinoculant Pseudomonas putida KNP9 Mediated Enhancement of Soybean Growth under Heavy Metal Stress Suitable for Biofuel Production at the Metal-Contaminated Site. Energies 2023, 16, 4508. [Google Scholar] [CrossRef]
- Raihan, M.R.H.; Rahman, M.; Mahmud, N.U.; Adak, M.K.; Islam, T.; Fujita, M.; Hasanuzzaman, M. Application of Rhizobacteria, Paraburkholderia fungorum and Delftia sp. Confer Cadmium Tolerance in Rapeseed (Brassica campestris) through Modulating Antioxidant Defense and Glyoxalase Systems. Plants (Basel) 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Moulin, L.; Munive, A.; Dreyfus, B.; Boivin-Masson, C. Nodulation of legumes by members of the beta-subclass of Proteobacteria. Nature 2001, 411, 948–950. [Google Scholar] [CrossRef]
Figure 1.
Representative figure of P. phymatum wild-type (WT) and a siderophore mutant (ΔphmJK) grown on linear gradient plates prepared with LB-NaCl agar medium supplemented with (a) 40 g/L of MMS-2, (b) 102.4 mM of AlCl3, and (c) 92 mM FeCl3 for the bottom layer. LB-NaCl agar medium was used for the upper layer. The plates were incubated for 48 hours at 28 °C. Three (n=3) biological replicates were performed. The black line shows where the cells stop growing, referring to a specific concentration on a steadily increasing gradient from the top to the bottom.
Figure 1.
Representative figure of P. phymatum wild-type (WT) and a siderophore mutant (ΔphmJK) grown on linear gradient plates prepared with LB-NaCl agar medium supplemented with (a) 40 g/L of MMS-2, (b) 102.4 mM of AlCl3, and (c) 92 mM FeCl3 for the bottom layer. LB-NaCl agar medium was used for the upper layer. The plates were incubated for 48 hours at 28 °C. Three (n=3) biological replicates were performed. The black line shows where the cells stop growing, referring to a specific concentration on a steadily increasing gradient from the top to the bottom.

Figure 2.
UHPLC-MS base peak chromatograms of (a) P. phymatum wild-type (WT), (b) WT spiked with FeCl3, (c) WT spiked with AlCl3, and (d) ΔphmJK. Phymabactin was eluted at a retention time (RT) of 4.25 min, the Fe(III)-phymabactin complex at an RT of 4.14 min, and the Al(III)-phymabactin complex at an RT of 3.90 min. The respective mass spectra of the double-charged molecular ions are shown in the boxes on the right side.
Figure 2.
UHPLC-MS base peak chromatograms of (a) P. phymatum wild-type (WT), (b) WT spiked with FeCl3, (c) WT spiked with AlCl3, and (d) ΔphmJK. Phymabactin was eluted at a retention time (RT) of 4.25 min, the Fe(III)-phymabactin complex at an RT of 4.14 min, and the Al(III)-phymabactin complex at an RT of 3.90 min. The respective mass spectra of the double-charged molecular ions are shown in the boxes on the right side.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



