
Submitted:
08 April 2025
Posted:
09 April 2025
You are already at the latest version
Abstract
Mangifera indica L. by-products obtained by three extraction methods from three cultivars (Keitt, Sensation and Gomera-3) were tested for their antibacterial properties against 20 bacterial species. These species were selected based on their relevance to winemaking processes (Acetobacter, Gluconobacter and Gluconacetobacter), fermented meat products (Staphylococcus) and human diseases (Pseudomonas, Escherichia, Shigella and Klebsiella). All mango by-product extracts showed antimicrobial activity in agar diffusion and broth microdilution experiments. However, differences in antimicrobial activity against acetic acid bacteria were detected between the peel extracts obtained from the two extraction processes. Furthermore, a wide range of minimum inhibitory concentration (MIC) data was found; Staphylococcus spp. (10 species) showed MICs between 1.0-240 mgGAE/mL and Acetobacter spp. (4 species) showed MICs between 1.7-200 mgGAE/mL. The most sensitive bacteria belonged to staphylococcal species (MICs: 1 mgGAE/mL) and the most resistant was Gluconacetobacter saccharivorans (MIC >400 mgGAE/mL). In general, there was no significant correlation between the phenolic compounds identified and the MICs values. The minimum bactericidal concentration (MBC) revealed that the mango extracts have bacteriostatic effect. A simple and reliable method for the determination of MIC and MBC in microdilution assays with acetic acid bacteria was described. These results highlight the antibacterial properties of the mango by-products against species associated with food spoilage microorganisms and human deseases.
Keywords:
Mangifera indica
; antibacterial
; acetic acid bacteria
; Acetobacter
; Staphylococcus
; fermented cured sausages
1. Introduction
A variety of microorganisms play an important role in the production of many foods and beverages. In many cases, these microorganisms can improve the quality and organoleptic properties of the final product. However, in other cases, some microorganisms are frequently associated with the poisoning and spoilage of foods. To overcome these problems, classical chemical preservatives have been routinely added to foods and beverages. In the case of wines, the addition of sulphites has been widely used as a strategy to prevent the growth of acetic acid bacteria (AAB) and spoilage yeasts [1]. Despite of the apparent efficacy of these preservatives, their overuse has been associated with several drawbacks, such as the induction of drug resistance and the occurrence of adverse effects after their consumption. Recently, Alaiya and Odeniy [2] reviewed aspects of the antimicrobial resistance and the environmental threat of some synthetic antimicrobial agents, along with the potential use of mango (Mangifera indica L.) extracts in antimicrobial formulations. In this context, the use of plant-derived excipients is increasing and opening up new ways to solve current drug delivery challenges in the pharmaceutical and food industries. These novel and biocompatible alternatives have the potential to minimize microbial spoilage and ensure food safety for consumers. In addition, the use of natural compounds confers additional benefits to the bioeconomy by adding value to agricultural by-products. These natural antioxidants and antibacterial agents are also in the line with the preferences of health-conscious consumers [3,4]. However, despite significant progress in recent years, the search for natural antimicrobial compounds remains a challenging area of research; fruit-containing polyphenols represent a particularly promising class of compounds. In this context, several studies have shown the antimicrobial activity of mango by-products extracts, which are rich in polyphenols [5,6,7,8]. Nevertheless, most of these studies have focused on a limited number of pathogenic or food safety-associated bacterial species, therefore the diversity of microorganisms analyzed is generally scarce [9,10,11,12,13]. In the present work, we sought to address this knowledge gap by investigating the antimicrobial activity of mango by-products against 20 bacterial species, including: i) Six AAB species (13 strains) related to the winemaking processes, ii) Ten Staphylococcus species related to fermented meat products, food safety or human and animal diseases, and iii) Four species commonly associated with human infections. The in vitro antibacterial activity was evaluated using agar diffusion and broth microdilution assays, and parameters such as the minimum inhibitory concentration (MIC) and the minimum bactericidal concentration (MBC) were determined. In addition, the correlation of the antibacterial activity of the extracts with polyphenolic compounds (flavonoids, gallates and gallotannins, ellagic acid, etc.) was analyzed. Additionally, a reliable and simple method for the determination of MIC and MBC values in microdilution assays with AAB was described. Furthermore, the possible relationship between bacterial genotype and susceptibility was also investigated.
2. Materials and Methods
2.1. Plant Collection and Preparation of Mango By-Products Extracts
Three mango cultivars (Keitt, Sensation and Gomera-3) were collected from the fields of the mango collection (UTM 28R 364451.00 m E3133070.93 m N; 114 m.a.s.l.) of the Instituto Canario de Investigaciones Agrarias (Tenerife, Canary Islands, Spain) at the physiological stage of ripeness (mature-green) and allowed to ripen (full-ripeness or consumption stage) at 18 °C and 80-90 % relative humidity. The plant material was identified, processed and characterized. The extracts were obtained using optimized methods to achieve the maximum extraction of polyphenols and a high antioxidant capacity, as described in previous works [3,13,14,15]. Briefly, the peel and seed were manually separated (31±1 % and 4.2 ±2.1 % of the total fruit weight, respectively), cut into small pieces (0.5 x 1 cm) and freeze-dried at –40 °C in a vacuum (50 mPa) for 5 days. The dried mango peels and seeds were then ground to a fine powder (particle size between 355-500 µm) and stored at –20 °C until the extractions were performed. Microwave assisted extraction (MAE) (ETHOS 1, Milestone SRL, Sorisole, Italy) was used to obtain the extracts. Two types of peel extractions were performed: ethanol:water (1:1, v:v) at 75 °C and a weight-to-solvent volume of 1:50 (w:v) (extraction A), or 1:10 (w:v) (extraction B). On the other hand, the mango seed extraction was performed with acetone:water (1:1, v:v) at 50 °C and a weight-to-solvent volume of 1:30 (w:v). The resulting nine by-product extracts included three extracts (peel: extraction A and B, and seed) for each of the three mango cultivars (Keitt, Sensation and Gomera-3) [10,11,12,13].
The compounds present in the nine by-product extracts were identified by the following acronyms: KPA (Keitt Peel, extraction A); KPB (Keitt Peel, extraction B); SPA (Sensation Peel, extraction A); SPB (Sensation Peel, extraction B); GPA (Gomera Peel, extraction A); GPB (Gomera Peel, extraction B); KSC (Keitt Seed, extraction C); SSC (Sensation Seed, extraction C); GSC (Gomera Seed, extraction C).
The characterization of the polyphenolic composition of the mango by-product extracts was previously conducted [13].
2.2. Bacterial Species and Culture Conditions
The antibacterial activity of the extracts was tested against 20 bacterial species; i) six AAB species: Acetobacter cerevisiae, A. malorum, A. pasteurianus, A. tropicalis, Gluconacetobacter saccharivorans and Gluconobacter japonicus; ii) ten Staphylococcus species: Staphylococcus arlettae, S. aureus, S. carnosus subsp. carnosus, S. chromogenes, S. cohnii subsp. cohnii, S. equorum subsp. equorum, S. gallinarum, S. lugdunensis, S. sciuri subsp. sciuri and S. xylosus; iii) four species frequently associated with human infections: Escherichia coli, Klebsiella pneumoniae, Pseudomonas aeruginosa and Shigella dysenteriae.
The AAB strains were obtained from grapes from the Canary Islands (Spain); identified by the phylogenetic analysis of the 16S-23S rRNA gene ITS sequence and typed by the analysis of highly conserved repetitive DNA elements [(GTG)5-PCR and Enterobacterial Repetitive Intergenic Consensus (ERIC)-PCR]16. Staphylococcal strains were obtained from the Agricultural Research Service Culture Collection (NRRL), except for the clinically isolated S. aureus 11923-76 and S. lugdunensis 99705-65 obtained from the microbiology service of the Nuestra Señora de Candelaria University Hospital (Santa Cruz de Tenerife, Spain). E. coli, K. pneumonia, P. aeruginosa and S. dysenteriae strains were obtained from the American Type Culture Collection (ATCC) (Manassas, USA). All the strains were maintained in 30 % w/v glycerol at -80 °C until use. For experiments, all the bacteria strains (except the AAB) were grown separately on Mueller Hinton (MH) agar at 37 °C for 24 h, whereas the AAB strains were grown in GYC (5 % glucose, 1 % yeast extract, 1.5 % CaCO3, 1.5 % agar w/v) medium at 28 °C under aerobic conditions for 48 h.
2.3. Antibacterial Screening
The antibacterial activity of the mango peel and seed extracts was determined using two different bioassays: agar diffusion and micro-dilution.
2.3.1. Agar Diffusion Assay
The agar diffusion assay was used only in a preliminary and exploratory way to determine the antibacterial potential of the extracts. Each strain (except the AAB) was grown at an optical density at wavelength 600 nm (OD600) of 1 in MH broth with agitation (250 rpm) at 37 °C. The cell suspension was mixed with melted warm MH 1 % agar medium at a final OD600 of 0.033 and poured into sterile Petri plates. A similar procedure was used for AAB; YPD broth (0.5 % yeast extract, 0.3 % peptone, 2 % dextrose) with agitation at 28 °C was used to obtain the cell suspension, and melted warm GYC 1 % agar to perform the final mixture to be poured into the Petri plates. Finally, sixty microlitres of each extract was added to 8-mm-diameter wells dug in MH 1 % agar or GYC 1 % agar medium plates. After 2 h at 4 °C, the plates were incubated at 37 °C (MH agar) or 28 °C (GYC agar) for 24 h, and then the inhibition zones (mm) were measured.
2.3.2. Micro-Dilution Antibacterial Assay
The MICs of the extracts were determined by a serial dilution method in 96-well sterile microtitre plates. The antimicrobial activity bioassays were optimized according to the requirements and characteristics of each bacterial species. All strains (except the AAB) were grown up to the exponential phase (OD600=0.5) in MH broth with agitation at 37 ºC. After that the cells were centrifuged (2500 rcf for 10 min.) and resuspended in sterile MH broth concentrate (2.5X) at an OD600 of 0.1 (K. pneumoniae, S. dysenteriae and Staphylococcus species) or 0.25 (E. coli and P. aeruginosa) and mixed with the different extracts at a concentration of between 1.0 and 240 mg mg gallic acid equivalents per mL (GAE/mL). Briefly, 44 µL of cell suspension (OD600=0.1 or 0.25) and 66 µL of the extract were added to each well. The final inoculum contained approximately 2 x 105 CFU/mL in a total volume of 110 µL/well of MH (1X). The microtitre plates were mixed and incubated under aerobic conditions at 37 ºC for 24 h and then examined using a binocular microscope and a spectrophotometer.
The AAB strains were grown in YPD broth (up to OD600=1.0) with agitation at 28 °C. The cells were concentrated by centrifugation at 2500 rcf for 10 min. and resuspended in GYC broth concentrate (2.5X) at an OD600 of 1.25 and mixed with the extracts at a concentration of between 1.7 and 400 mg GAE/mL. Briefly, 44 µL of the cell suspension in GYC and 66 µL of extract were added in each well. The microtitre plates were mixed and incubated under aerobic conditions at 28 °C for 24 h, and the analysis was performed using a bromo cresol purple (BCP) alkaline solution (0.1% w/v BCP + OHK 0.05M). For that, 8 µL/well of the BCP solution was added and after one minute the color of the reaction was observed [positive reaction (acid)=yellow, negative reaction (alkaline)=violet].
Positive controls (containing bacteria without extracts) and negative controls (containing extracts without bacteria) were included in each microplate of each experiment. The MIC was determined as the lowest concentration of extracts that inhibited the visual growth of the bacterial culture on the microplate. The spectrophotometric analysis (except the AAB) was determined using a microdilution automatic reader at a wavelength of 600 nm, and each test sample was considered negative when the bacteria growth displayed 90 % inhibition compared with the positive control. In addition, the MBC was determined via serial subcultivation of 10 µL aliquots of each well in microtitre plates containing 90 µL of fresh MH broth per well and further incubation for 24 to 48 h at 37 °C. The lowest concentration with no visible growth (at the binocular microscope) and the lowest determined by spectrophotometric analysis was defined as the MBC. In the case of AAB, the MBC was determined with fresh GYC broth at 28 °C and the evaluation was performed with BCP alkaline solution. Each sample was tested in triplicate in separate experiments. Figure 1 illustrates the whole procedure.
2.4. Statistical Analysis
All results are expressed as mean ± SD (n = 3). Spearman’s correlation analysis was used to determine the correlation between antibacterial capacity and phenolic compounds of the extracts. Differences were considered significant at p < 0.05. All statistical analyses were performed with the Statgraphics Centurion 18 V18.1.14 software.
3. Results
3.1. Bioactive Compound in Mango By-Products Extracts
A total of 30 phenolic compounds were identified in a previous work [13] from nine mango by-products extracts obtained from peels and seeds of the three cultivars (Keitt, Sensation and Gomera 3) (Figure 2). Among them, five main groups of bioactive compounds were found: Flavoinds (10 compounds), Xanhones (1), Gallates and gallotannins (10), Ellagic acid and derivatives (2) and Benzophenone derivatives (7).
The flavonoid concentration (g/100 g dw) was significantly different (p < 0.05) among the peel of the three mango cultivars and in the extracts obtained with the two extraction conditions evaluated (A or B) (Figure 2). In general, extracts obtained with process A showed 1.4 times more (on average) flavonoid concentration than extracts B. Moreover, cultivar Gomera-3 showed 2.5 and 6.5 times more flavonoids than Sensation and Keitt cultivars, respectively. Furthermore, the concentration of flavonoids in the seeds was significantly lower than in the peel (between 4.5 and 40 times on average, independently of extraction). Only four of the ten compounds identified in the peel were detected.
Gallates and gallotannins in mango peel did not show significant differences (p < 0.05) among cultivars (within the same extraction process). However, the peel of the Sensation and Gomera-3 cultivars showed higher extraction yield in extract B (31.5 ± 1.75 g/100 g dw) than in extract A (25.0 ± 4.06 g/100 g dw). In addition, seed extracts showed a significantly lower yield compared to peel extracts (13.8 times on average), however the number of identified compounds was similar.
The content of ellagic acid derivatives in the extracts A showed no significant differences between cultivars, however, in the extracts B, the concentration in Keitt cultivar was significantly lower (p < 0.05) than in the Sensation and Gomera-3 cultivars (1.7 and 1.3 times, respectively). In seed extracts, the Sensation cultivar showed a significantly higher ellagic acid concentration in B extracts (24.9 ± 4.9 g/ 100 g dw) compared to the other cultivars (16.7 ± 3.4 g/ 100 g dw). Likewise, the mean concentration in Sensation seed extracts exceeded that of the peel by 1.65 times.
Finally, the highest concentration of xanthones was observed in the seed extracts of cultivar Keitt (4.15 ± 0.18 g/100 g dw), followed by peel extracts A and B of the Gomera-3 cultivar (1.41 and 1.29 g/100 g dw, respectively). For this group of compounds, the only compound identified and quantified was mangiferin.
3.2. Antibacterial Activity of Mango By-Products
3.2.1. Acetic Acid Bacteria
A preliminary and exploratory screening of the antibacterial activity of mango by-products against three AAB species (A. malorum, A. tropicalis and G. japonicus) was performed using agar diffusion assay. All extracts (from peel and seed) had an inhibition effect against tested bacteria. Likewise, the extraction process and the cultivar affected the bioactive compounds extracted and therefore their antibacterial activity. Peel extracts obtained by process B showed higher antibacterial activity in agar diffusion assay (1.5 times on average) than extracts obtained by process A (data not shown). On the other hand, the peel and seed extracts from Gomera-3 and Sensation cultivars showed higher antibacterial activity than extracts obtained from Keitt cultivar.
For a quantitative analysis of the antibacterial activity, the MIC and the MBC were determined against six species of AAB (thirteen genotypes typified by ERIC-PCR and GTG5-PCR). In this case, all the extracts were diluted to a final concentration between 1.7 and 400 mg GAE/mL. The serial dilution assays confirm the antibacterial properties of the extracts, with the exception of G. saccharivorans that was resistant at the maximum concentration tested (400 mg GAE/mL). A wide range of MIC and MBC values was observed among the different bacterial species, with MICs from 1.7 mg GAE/mL in A. cerevisiae to 200 mg GAE/mL in A. tropicalis and G. japonicus, and MBCs from 17 mg GAE/mL to 400 mg GAE/mL in the same species mentioned above (Table 1).
Taking into account the MICs values in peel extracts, two arbitrary groups can be defined; Group I, species with MICs ≤ 50 mg GAE/mL (A. cerevisiae, A. pasteurianus and A. malorum), and Group II, species with MICs ≥ 50 mg GAE/mL (A. tropicalis, G. japonicus and G. saccharivorans) (Table 1). In no case, extracts B had MICs greater than extracts A. Peel extracts obtained by extraction A showed higher MICs values than extracts obtained by extraction B in each bacterial species tested in at least in one of the cultivars. In Gomera-3 cultivar MICs from extracts A were higher than extracts B in the 66.6 % of the genotypes (Ac2, Ac3, Ac5, Am17, Am25, Am26, Ap16 and Gj2), while in Keitt cultivar MICs from extracts A were higher than extracts B in the 58.3 % of the genotypes (Ac2, Ac3, At1, At2, Gj1, Gj2 and Gj3). In the extracts obtained by process A, A. cerevisiae and A. malorum showed no differences between cultivars, however, A. pasteurianus showed a lower MIC value with the extract obtained from cultivar Keitt, and A. tropicalis with those from Gomera-3. Elsewhere, the highest MIC values in G. japonicus were observed in the Keitt cultivar. In the case of extracts obtained by process B, Sensation cultivar showed higher MICs than the other cultivars with A. cerevisiae (Ac3), A. pasteurianus (Ap16), and A. tropicalis (At1, At2) (Table 1). While Gomera-3 cultivar showed highest MICs with G. japonicus (Gj1, Gj3) and lowest MICs with A. cerevisiae (Ac5), A. pasteurianus (Ap16) and A. malorum (Am17, Am25, Am26).
In seed extracts, the results were variable, being able to highlight the greater MICs values in A. malorum and A. tropicalis with Keitt cultivar, and the equality of the results among cultivars in G. japonicus assays (Table 1). In addition, it is important to note the differences between the MIC data of the peel and seed extracts on the different bacterial species and cultivars. For example, in A. cerevisiae the MICs of seed extracts of Sensation and Keitt cultivars showed higher values than in peel extracts, however, in A. pasteurianus (Ap16) the relation was the opposite (MICs of peel extracts were highest than seed extracts). Likewise, in the Sensation cultivar, the MIC of the seed extracts were lower than the peel extracts.
The MBC assay was performed to confirm cell death in the MIC test. Under the tested concentrations, most of the AAB species displayed difference between the MIC and MBC values, which indicates that the mango extracts have a bacteriostatic effect. The MBCs range between 17 and 400 mg GAE/mL. In general terms, the differences between bacterial species or genotypes and between cultivars were similar to those described in the case of MICs; Group I include species with MBCs ≤ 100 mg GAE/mL, and Group II species with MBCs ≥ 100 mg GAE/mL (Table 1). Likewise, peel extracts obtained by process A showed higher MBCs values than extracts obtained by process B in each bacterial genotype tested in at least in one of the cultivars.
Finally, the MICs values were correlated with the polyphenolic content of the extracts to examine if any of the phenolic compounds were associated with the antibacterial activity. As shown in Table 4 the statistical correlation analysis shows that the phenolic compounds quantified in our study have no significant correlation with the MICs of AAB tested, except A. cerevisiae (Ac5). In this case, a significant (p < 0.05) negative correlation was observed between MICs values and the concentration (g/100 g dw) of flavonoids (r = -0,8385), gallates and gallotannins (r = -0,7267), and total phenolic compounds (r = -0,8385) (Table 4).
3.2.2. Staphylococcus Species
The agar diffusion assay indicated that all extracts (from peel and seed) had an inhibition effect against all the tested species. Nevertheless, the antimicrobial activity of the extracts (inhibition halo) on the different bacterial species tested was diverse. Furthermore, the extraction process and the cultivar affected the antibacterial activity of the obtained extracts. The peel extracts of process B showed higher antibacterial activity (1.5 times on average) than extracts obtained by process A, and Sensation cultivar (peel and seed extracts) showed (in most cases) the higher antibacterial activity (data not shown).
The quantitative analysis of the antibacterial activity (MIC and MBC) was determined against 10 species of Staphylococcus (Table 2). The MIC values ranged from 1 to 240 mg GAE/mL and the MBCs from 1 to 240 mg GAE/mL, except for S. lugdunensis (more resistant to some extracts). Taking into account the MIC values in peel extracts, two arbitrary groups can be defined; Group I, species with MICs ≤ 5 mg GAE/mL, and Group II, species with MICs ≥ 5 mg GAE/mL (Table 2).
In peel extracts, no differences were observed between MICs of both extraction processes (A and B) in Gomera-3 and Sensation cultivars, however, in cultivar Keitt the results were different: five bacteria species of group I presented lower MICs with the extraction process B, and three species of group II presented lower MICs with process A. On the other hand, the extracts obtained with process A showed no differences between cultivars in the bacterial species of group I, while in group II the cultivar affected the MIC values (except S. aureus NRRL B-767). In extracts obtained with process B, Keit cultivar presented the lowest MICs in the bacteria of group I, while in group II, three bacterial species (S. aureus, S. cohnii and S. lugdunensis) showed the opposite effect. In seed extracts, the MICs of eight bacterial species showed no differences between cultivars, however, in S. arlettae, S. equorum and S. gallinarum the MICs of the extracts of cultivar Keitt were lower. Furthermore, no significant correlation was observed between the phenolic compounds and the MICs values of the Staphylococcus species tested (Table 4).
3.2.3. Human Pathogenic Bacteria
To compare the antibacterial activity of our extracts with other published studies, we used potentially human pathogenic bacteria generally tested in in vitro assays: E. coli, K. pneumoniae, P. aeruginosa and S. dysenteriae. The agar diffusion assay showed the antibacterial properties of the extracts (from peel and seed) on all the species tested. P. aeruginosa was the most susceptible species and, in general terms, the extracts obtained by process B showed higher inhibitory capacity than those obtained by process A. Also, no differences were observed between cultivars (data not shown).
The serial dilution assays confirm that all extracts had antibacterial properties, with MICs between 10 and 240 mg GAE/mL (Table 3). In the same way as in agar diffusion assays, differences between species were detected; P. aeruginosa was the most sensitive (MIC 10 mg GAE/mL) and K. pneumoniae was the most resistant species (MIC 240 mg GAE/mL).
No differences in MICs values of peel extracts were observed among extraction processes of the four bacterial species tested, except for S. dysenteriae which showed a higher susceptibility to the extracts obtained with process B. Likewise, no differences in MIC values among cultivars (in peel and seed extracts) were observed, except in P. aeruginosa where Keitt seed extracts have a lower MIC (10 mg GAE/ml) than Gomera-3 and Sensation (30 mg GAE/ml). The MBC results indicated that mango extracts have a bacteriostatic effect on the species tested (Table 3). Finally, Spearman’s analysis with significant negative correlation coefficient was found between the MICs of P. aeruginosa and the concentration (g/100 g dw) of flavonoids, gallates and gallotannins and total phenolic compounds (r = -0,8216, p < 0.05). Likewise, significant negative correlation was found between the MICs of S. dysenteriae and the concentration of gallates and gallotannins (r = -0,7303, p < 0.05) (Table 4).
4. Discussion
Pathogenic bacteria and antibiotic resistance represent a major global public health problem [14,15]. Therefore, the search of novel antibacterial products environmentally friendly for the treatment of infections caused by drug-resistant bacteria is essential. In this context, several studies describe different antioxidant and antimicrobial compounds obtained from mango extracts (mainly from leaves, fruit peel, root, stem bark and seed). Furthermore, several review articles have focused on analyzing the pharmacological properties of these extracts, which are generally attributed to the presence of phytochemical substances such as tannins, alkaloids, flavonoids, mangiferin, phenols, etc. [2,16,17,18]. Nevertheless, the antimicrobial activity of the extracts is influenced by several factors, including the phenological stage and part of the plant (roots, leaves, bark, fruit peel and seed, etc.), the cultivar, the solvent and extraction method applied, the season of the year [13,18,19,20,21] and many other variables that are still unknown.
As mentioned above, some studies describe the antibacterial potential of mango extracts, however, most studies test these properties mainly on a small number of bacterial species of human clinical interest, while only a few studies include different bacterial strains or species related to food spoilage [22]. This last aspect is important, as many foods currently incorporate different synthetic preservatives that inhibit the microbial proliferation and increase the shelf life of the product [23]. However, in some cases, these compounds may cause asthma, allergies, nausea, diarrhea, lung irritation, and can even be carcinogenic and harmful to the circulatory, hepatic and urinary systems [24,25,26,27]. Therefore, the use of natural plant extracts as food preservatives is considered a healthier alternative to successfully increase the shelf life of processed foods without exposing consumers to the harmful effects of synthetic preservatives. In this context, many pharmaceutical companies are focusing on the search for alternative preservatives of natural origin to eliminate synthetic preservatives from food due to their undesirable effects on humans [28,29,30].
An example of this can be seen when wines are spoiled by AAB during maturation or storage. The proliferation of AAB in wine results in the production of metabolites such as acetic acid, acetaldehyde and ethyl acetate, which give the wine undesirable sensory characteristics and are a serious commercial problem [31,32]. To avoid this, sulfur dioxide has been traditionally used to control spoilage microorganisms during winemaking. Nevertheless, several natural products can be used for the control of bacteria in wine, such as lysozyme and bacteriocins [31]. Additionally, it has been demonstrated that phenolic compounds possess antibacterial and antifungal properties and some types of these compounds can affect the wine bacterial growth [35,36]. However, a scarce number of studies evaluates the effect of polyphenols on the AAB growth. In this sense, Alañón et al. [32] used oenological woods extracts against A. acetii and G. oxydans, and García-Ruiz et al. [34] and Pastorkova et al. [33] used commercial phenolic compounds against A. aceti and G. oxydans, and A. aceti, A. oeni and A. pasteurianus, respectively. In the same way, Sabel et al., [1] analyzed five commercial phenolic compounds (gallic acid, ferulic acid, sinapic acid, resveratrol and syringaldehyde) against two AAB (G. cerinus 9533 and A. acetii 3508) obtained from the German Collection of Microorganisms and Cell Cultures (DSMZ, Braunschweig).
In our work, we report the ability of mango by-products to inhibit the growth of six AAB species (13 genotypes) of enological origin (microvinifications of red and white grapes) and demonstrate the wide range of sensitivity between the different AAB species; ranging from species with high sensitivity, such as A. cerevisiae, to species resistant to high concentrations of phenols, such as G. saccharivorans. Several works support the antimicrobial activity of ellagic acid and their derivates. This compound can be present in free form or as derivatives, principally as complex polymers called ellagitannins [37,38]. The mechanism of action described for ellagic acid is related to their ability coupled with protein in bacteria walls and, in other cases, their capacity to inhibit gyrase activity which is associated with the cleavage of DNA strand during the replication process [37] Likewise, others authors describe a strong influence of the extraction process to obtain extracts rich in antibacterial compounds [8]. In our work, the extracts with high content in gallates and gallotannins showed highest antibacterial activity against A. cerevisiae (Ac5) (Table 4). Probably, this result is due to the iron-chelating ability of the gallotannins [4]. However, the rest of the AAB species tested did not show a significant correlation with any of the phenolic compounds quantified.
With regard to the genus Staphylococcus many species are considered by the European Food Safety Authority [39] as food-borne disease-causing agents due to the ability to produce enterotoxins. Coagulase negative staphylococci (CNS) are commonly present in fermented foods, cheeses, sausages and meat products and are often resistant to one or more antibiotics [40]. Although this group of microorganisms is part of the normal non-pathogenic commensal biota, many studies have shown that it can be one of the most common causes of nosocomial infections, especially in immunocompromised and hospitalized patients [41]. In fact, in particular S. lugdunensis, a component of the human skin microbiome mainly associated with moist areas of the body, such as the inguinal fold and perineum, and the nasal cavity (less frequently) [42,43], can cause a destructive form of infective endocarditis [44,45] and a wide range of nosocomial and community-acquired infections in the skin and soft tissues, bones and joints, prosthetic joint, vascular catheters and abscesses [46,47,48]. In addition, S. xylosus and S. sciuri have been isolated from cheese as potentially responsible for food poisoning, and in commercial salamis, they are commonly associated with two other species carrying genes involved in the biosynthesis of toxins: S. saprophyticus and S. carnosus [49]. All these evidences highlight the need to find and include natural compounds during the manufacture of sausages, to control the growth of potentially harmful staphylococcal species. In this sense, a large number of studies have demonstrated the ability of different natural compounds to control staphylococci. However, most of the published work focuses mainly on a few Staphylococcus species of medical interest, especially S. aureus [9,51,52,53]. Previous studies described the antibacterial activity of mango seed kernel extracts against methicillin resistant S. aureus (MRSA) [50,53]. In our work, we show the effectivity of mango polyphenolic extracts against ten Staphylococcus species associated with fermented meat products, food safety or human and animal diseases. In a general way and in contrast with the results obtained with the AAB, no differences were observed between the extraction process and cultivars (except for certain data of Keitt cultivar) and no significant correlation was observed between the phenolic compounds and the MICs values of the Staphylococcus species tested. Therefore, probably, the antibacterial activity of mango extracts is due to a synergistic effect between the different phenolic compounds. However, different studies highlight the antibacterial capacity of certain phenolic compounds. For example, mangiferin and its analogs have been reported for their moderate activity against S. aureus [54,55,56]. On the other hand, our results are in agreement with those of Gupta et al. [51] using ethanolic and methanolic mango extracts against S. aureus. Kabuki et al. [53] tested ethanolic extracts from mango seed against 18 bacterial species, including different strains of S. aureus, obtaining MICs between 0.05-1 mg GAE/mL. These values are several times lower than those observed in the present work (between 7.5-10 mg GAE/mL), probably because a different solvent (ethanol) was used to obtain the mango extracts. Torres-León et al. [8] investigated mango seed extracts obtained by MAE, Fermentation-Assisted Extraction (FAE) and Solvent-Assisted Extraction (SAE) and showed the antibacterial properties against six isolates of S. aureus. The MICs revealed a high antibacterial activity (7.31–750 μg/mL) compared to synthetic antibiotics (vancomycin and penicillin) and were influenced by the extraction technique: SAE had the highest biological activity. Finally, our work demonstrates the antibacterial capacity of mango extracts on a wide diversity of species of Staphylococcus (other than S. aureus) associated with food spoilage or pathogenic for humans. We also demonstrate the wide range of sensitivities between species; from high sensitivity (species of Group 1, Table 2) to highly resistant (such as S. lugdunensis). Consequently, this finding suggests that mango extracts are promising compounds for further development as natural antibacterial agents with pharmaceutical and food industry applications.
In relation with different human pathogenic bacteria, several studies describe the in vitro antibacterial activity of mango extracts. However, the high diversity of extraction processes (ethanol, methanol, etc.), cultivars, parts of the plant (stem bark, leaves, rind, seeds, dried pulp, etc.), fruit ripening conditions and the in vitro evaluation methods (disks, wells, diffusion agar, macro or micro dilution, etc.) used in the different works, strongly affect the total polyphenolic content as well as their antimicrobial activity potential of the extracts. Consequently, the MICs values for the same bacterial species are very diverse and are expressed in different ways (mm halo, mg, mg/ml, mg GAE/ml, etc.), which makes cross-data comparisons very difficult. For example, the MIC data for E. coli or P. aeruginosa may range from 1.25 to 100 mg/ml. Another aspect widely reported in different studies and in some cases duly argued [4,7] is the description of the greater susceptibility of the Gram positive bacteria concerning the Gram negative, likely as a result of the barrier effect caused by the outer membrane. Given the results of the present study we cannot confirm this concept since, from 10 Gram negative (AAB and human pathogenic bacteria) and 10 Gram positive (Staphylococcus spp) species tested, we have observed a wide range of MIC data: 1.7 to >400 mg GAE/ml and 1.0 to 120 mg GAE/ml, respectively. However, although the maximum MIC value recorded for Gram negative bacteria (>400 mg GAE/ml) was at least 3.3 times higher than for Gram positive bacteria (120 mg GAE/ml), the range of values in both types of bacteria is wide spread. For this reason, to confirm this general concept, the mode of action of the extracts should be studied in more detail and a greater number of bacterial species and strains should be tested.
5. Discussion
It has been well documented that mango fruits and their by-products are an important source of bioactive compounds such as phenolic acids, gallotannins, flavonoids, catechins, benzophenones, carotenoids, tocopherols and xanthonoids. This article describes the antibacterial properties of mango by-products (peel and seed) from three cultivars (Keitt, Sensation and Gomera-3) widely distributed in the Canary Islands, against AAB related to the winemaking processes, Staphylococcus species related to fermented meat products, food safety or human and animal diseases, and other bacterial species frequently associated with human infections. The bacterial species was the most important factor to consider in the evaluation of the extracts activity (MICs data), showing a high diversity of data between species within the same genus; MICs from 1.7 to 200 mg GAE/mL in four Acetobacter species, and 1.0 to 240 mg GAE/mL in ten Staphylococcus species. In general, no significant correlation between the phenolic compounds identified and the MICs was detected and the MBC revealed the bacteriostatic effect of the mango extracts. In addition, a simple and reliable method to assess MIC and MBC of AAB was described.
Most food products require protection against microbial spoilage during storage to maintain food quality and safety and prevent nutritional and organoleptic losses during the consumption period. In this context, natural compounds have a great interest in the industry, as a result of the potential to provide food quality and safety with a reduced impact on human health. In this sense, this work highlights the antibacterial properties of mango and demonstrates the potential of extracts as a natural and functional ingredient for the control of spoilage or potentially food-borne pathogenic microorganisms.
Author Contributions
“Conceptualization, methodology, investigation, formal analysis, data curation, writing—original draft preparation, writing—review and editing: Federico Laich and Eva Dorta; visualization, supervision, project administration, funding acquisition Federico Laich, Eva Dorta, Mónica González and Gloria Lobo. All authors have read and agreed to the published version of the manuscript.”
Institutional Review Board Statement
“Not applicable” for studies not involving humans or animals.
Data Availability Statement
We encourage all authors of articles published in MDPI journals to share their research data. In this section, please provide details regarding where data supporting reported results can be found, including links to publicly archived datasets analyzed or generated during the study. Where no new data were created, or where data is unavailable due to privacy or ethical restrictions, a statement is still required. Suggested Data Availability Statements are available in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Acknowledgments
Eva Dorta acknowledges the Spanish National Institute for Agricultural and Food Research and Technology (INIA). Proyects I+D+i RTA2006-00187, CAIA 2024-0010-05-02 and CAIA 2024-0010-05-01.
Conflicts of Interest
The authors declare no conflicts of interest.”
Abbreviations
| AAB | Acetic Acid Bacteria |
| FAE | Fermentation-Assisted Extraction |
| GAE | mg gallic acid equivalents per mL |
| GPA | Gomera Peel, extraction A |
| GPB | Gomera Peel, extraction B |
| GSC | Gomera Seed, extraction C |
| MBC | Minimum Bactericidal Concentration |
| MIC | Minimum Inhibitory Concentration |
| KPA | Keitt Peel, extraction A |
| KPB | Keitt Peel, extraction B |
| KSC | Keitt Seed, extraction C |
| SAE | Solvent-Assisted Extraction |
| SPA | Sensation Peel, extraction A |
| SPB | Sensation Peel, extraction B |
| SSC | Sensation Seed, extraction C |
References
- Sabel, A.; Bredefeld, S.; Schlander, M.; Claus, H. Wine Phenolic Compounds: Antimicrobial Properties against Yeasts, Lactic Acid and Acetic Acid Bacteria. Beverages 2017, 3, 29. [Google Scholar] [CrossRef]
- Alaiya, M.A.; Odeniyi, M.A. Utilisation of Mangifera indica plant extracts and parts in antimicrobial formulations and as a pharmaceutical excipient: a review. Futur. J. Pharm. Sci. 2023, 9, 29. [Google Scholar] [CrossRef] [PubMed]
- Barata, A.; Caldeira, J.; Botelheiro, R.; Pagliara, D.; Malfeito-Ferreira, M.; et al. Survival patterns of Dekkera bruxellensis in wines and inhibitory effect of sulphur dioxide. Int J Food Microbiol 2008, 121, 201–207. [Google Scholar]
- Widsten, P.; Cruz, C.D.; Fletcher, G.C.; Pajak, M.A.; McGhie, T.K. Tannins and Extracts of Fruit By products: Antibacterial Activity against Foodborne Bacteria and Antioxidant Capacity. J. Agric. Food. Chem. 2014, 62, 11146–11156. [Google Scholar]
- Gupta, A.K.; Gurjar, P. S.; Beer, K.; Pongener, A.; Ravia, S.C.; Singh, S.; Verma, A.; Singh, A.; Thakure, M.; Tripathyf, S.; Vermaf, D.K. A review on valorization of different byproducts of mango (Mangifera indica L. ) for functional food and human health. Food Biosc. 2022, 48, 101783. [Google Scholar]
- García-Mahecha, M. Bioactive compounds in extracts from the agro-industrial waste of mango. Molecules 2023, 28, 458. [Google Scholar] [CrossRef] [PubMed]
- Kucuk, N.; Primožiˇc, M.; Kotnik, P.; Knez, Ž.; Leitgeb, M. Mango peels as an industrial by-product: a sustainable source of compounds with antioxidant, enzymatic, and antimicrobial activity. Foods 2024, 13, 553. [Google Scholar] [CrossRef]
- Torres-León, C.; de Azevedo Ramos, B.; dos Santos Correi, M.T.; Carneiro-da-Cunha, M.G.; Ramirez-Guzman, N., Alves; Ascacio-Valdes, J.; Álvarez-Pérez, O.B.; Aguilar, C.N. Antioxidant and anti-staphylococcal activity of polyphenolic-rich extracts from Ataulfo mango seed. LWT – Food Sci Tech 2021, 148, 111653.
- Abdullah, A.; Saeed Mirghani, M.; Jamal, P. Antibacterial activity of malaysian mango kernel. Afr J Biotechnol 2011, 10, 18739–18748. [Google Scholar]
- Dorta, E.; González, M.; Lobo, M.G.; Laich, F. Antifungal activity of mango peel and seed extracts against clinically pathogenic and food spoilage yeast. Nat Prod Res 2015, 26, 1–7. [Google Scholar] [CrossRef]
- Dorta, E.; Lobo, M.G.; González, M. Improving the efficiency of antioxidant extraction from mango peel by using microwave-assisted extraction. Plant Food Hum Nutr 2013, 68, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Dorta, E.; Lobo, M.G.; González, M. Optimization of factors affecting extraction of antioxidants from mango seed. Food Bioproc Tech 2013, 6, 1067–1081. [Google Scholar] [CrossRef]
- Dorta, E.; Gonzalez, M.; Lobo, M.G.; Sanchez-Moreno, C.; de Ancos, B. Screening of phenolic compounds in by-product extracts from mangoes (Mangifera indica L. ) by HPLC-ESI-QTOF-MS and multivariate analysis for use as a food ingredient. Food Res Int 2014, 57, 51–60. [Google Scholar]
- WHO, World Health Organization. Antimicrobial resistance. Available online: https://www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance (accessed on 12 December 2024).
- Prestinaci, F.; Pezzotti, P.; Pantosti, A. Antimicrobial resistance: a global multifaceted phenomenon. Pathogens and Global Health 2015, 109, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Jahurul, M.H.A.; Jahurul, I.S.M.; Ghafoor, Z.K.; Al-Juhaimi, F.Y. Mango (Mangifera indica L. ) by-products and their valuable components: a review. Food Chem 2015, 183, 173–180. [Google Scholar]
- Parvez, G.M. Pharmacological activities of mango (Mangifera indica): a review. J Pharmacognosy Phytochem 2016, 5, 01–07. [Google Scholar]
- Ediriweera, M.K.; Tennekoon, K.H.; Samarakoon, S.R. A review on ethnopharmacological applications, pharmacological activities and bioactive compounds of Mangifera indica (mango). Evid-Based Complement Alternat Med. 2017. [CrossRef]
- Maharaj, A.; Naidoo, Y.; Dewir, Y.H.; Rihan, H. Phytochemical screening and antibacterial and antioxidant activities of Mangifera indica L. Leaves. Hortcf 2022, 8, 909. [Google Scholar] [CrossRef]
- Manzur, A. G.; Junior, S.M.; Morais-Costa, F.; Mariano, E.G.; Careli, R.T.; da Silva, L.M.; Duarte, E.R. Extract of Mangifera indica L. leaves may reduce biofilms of Staphylococcus spp. in stainless steel and teat cup rubbers. Food Sci Technol int 2019, 108201321985852. [Google Scholar] [CrossRef]
- Engels, C.; Gänzle, M. G.; Schieber, A. Fast LC–MS analysis of gallotannins from mango (Mangifera indica L. ) kernels and effects of methanolysis on their antibacterial activity and iron binding capacity. Food Res Int 2012, 45, 422–426. [Google Scholar]
- Kabuki, T.; Nakajima, H.; Arai, M.; Ueda, S.; Kuwabara, Y.; et al. Characterization of novel antimicrobial compounds from mango (Mangifera indica L. ) kernel seeds. Food Chem 2000, 71, 61–66. [Google Scholar] [CrossRef]
- Adams, M.R.; Moss, M.O. Food Microbiology, 2nd ed.; Royal Society of Chemistry: Cambridge, UK, 2000; pp. 101–109. [Google Scholar]
- Gunnison, A.F.; Jacobsen, D.W. Sulfite hypersensitivity. A critical review. Crit. Rev. Toxicol. 1987, 17, 185–214. [Google Scholar]
- Bomhard, E.M.; Brendler-Schwaab, S.Y.; Freyberger, A.; Herbold, B.A.; Leser, K.H.; Richter, M. Ophenylphenol and its sodium and potassium salts: A toxicological assessment. Crit. Rev. Toxicol. 2002, 32, 551–626. [Google Scholar] [CrossRef] [PubMed]
- Oms-Oliu, G.; Rojas-Graü, M.A.; González, L.A.; Varela, P.; Soliva-Fortuny, R.; Hernando, M.I.H.; Munuera, I.P.; Fiszman, S.; Martín-Belloso, O. Recent approaches using chemical treatments to preserve quality of fresh-cut fruit: A review. Posth. Biol. Technol. 2010, 57, 139–148. [Google Scholar] [CrossRef]
- Kim, S.J.; Cho, A. R.; Han, J. Antioxidant and antimicrobial activities of leafy green vegetable extracts and their application to meat product preservation. Food Control 2013, 29, 112e120. [Google Scholar] [CrossRef]
- Dillon, V. M.; Board, R. G. Future prospects for natural antimicrobial food preservation systems. In V. M. Dillon, & R. G. Board (Eds.), Natural antimicrobial systems and food preservation (pp. 297e303). 1994, Wallingford: CAB International.
- Parke, D. V.; Lewis, D. F. V. Safety aspects of food preservatives. Food Addit. Contam 1992, 9, 561e577. [Google Scholar] [CrossRef]
- Bañón, S.; Díaz, P.; Rodríguez, M.; Garrido, M. D.; Price, A. Ascorbate, green tea and grape seed extracts increase the shelf-life of low sulphite beef patties. Meat Scie 2007, 77, 626e633. [Google Scholar] [CrossRef]
- Bartowsky, E.J.; Henschke, P.A. Acetic acid bacteria spoilage of bottled red wine - A review. Int J Food Microbiol 2008, 125, 60–70. [Google Scholar] [CrossRef]
- Alañón, M.E.; García-Ruíz, A.; Díaz-Maroto, M.C.; Pérez-Coello, MS.; et al. Antimicrobial and antioxidant activity of pressurized liquid extracts from oenological woods. Food Control, 2015, 50, 581–588. [Google Scholar] [CrossRef]
- Pastorkova, E.; Zakova, T.; Landa, P.; Novakova, J.; Vadlejch, J.; et al. Growth inhibitory effect of grape phenolics against wine spoilage yeasts and acetic acid bacteria. Int J Food Microbiol 2013, 161, 209–213. [Google Scholar] [CrossRef]
- García-Ruiz, A.; Cueva, C.; González-Rompinelli, E.M.; Yuste, M.; Torres Martín-Álvarez, J.; et al. Antimicrobial phenolic extracts able to inhibit lactic acid bacteria growth and wine malolactic fermentation. Food Control 2012, 28, 212–219. [Google Scholar] [CrossRef]
- Reguant, C.; Bordons, A.; Arola, L.; Rozès, N. Influence of phenolic compounds on the physiology of Oenococcusoeni from wine. J Appl Microbiol 2000, 88, 1065–1071. [Google Scholar] [CrossRef]
- Vivas, N.; Lonvaud-Funel, A.; Glories, Y. Effect of phenolic acids and anthocyanins on growth, viability and malolactic activity of a lactic acid bacterium. Food Microbiol 1997, 14, 291–300. [Google Scholar] [CrossRef]
- Sepúlveda, L.; Ascacio, L.; Rodríguez-Herrera, R.; Aguilera-Carbó, A.; Aguilar, C.N. Ellagic acid: Biological properties and biotechnological development for production processes. Afr. J. Biotechnol 2011, 10, 4518–4523. [Google Scholar]
- Ríos, J.L.; Giner, R.M.; Marín, M.; Recio, M.C. A Pharmacological Update of Ellagic Acid. Planta Med 2018, 84, 1068–1093. [Google Scholar] [CrossRef] [PubMed]
- EFSA – European Food Safety Authority, The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-borne Outbreaks in 2012. The EFSA Journal 2014, 12, 3547.
- Götz, F.; Bannerman, T.; Schleifer, K.H. The Genera Staphylococcus and Macrococcus. In: M Dworkin, S Falkow, E Rosenberg, K-H Schleifer, E Stackebrandt. (Eds.). The Prokaryotes (New York: Springer) 2006, 4, 5–75. [Google Scholar]
- Becker, K.; Heilmann, C.; Peters, G. Coagulase-Negative Staphylococci. Clin Microbiol Rev 2014, 27, 870–926. [Google Scholar] [CrossRef] [PubMed]
- Bieber, L.; Kahlmeter, G. Staphylococcus lugdunensis in several niches of the normal skin flora. Clin Microbiol Infect 2010, 16, 385–388. [Google Scholar] [CrossRef]
- Liu, C.M.; Price, L.B.; Hungate, B.A.; Abraham, A.G.; Larsen, L.A.; Christensen, K.; Stegger, M.; Skov, R. ; Andersen, Staphylococcus aureus and the ecology of the nasal microbiome. Sci Adv 2015, 1, e1400216. [Google Scholar] [CrossRef]
- Kyaw, H.; Raju, F.; Shaikh, A.Z.; Lin, A.N.; Lin, A.T.; Abboud, J.; Reddy, S. Staphylococcus lugdunensis endocarditis and cerebrovascular accident: a systemic review of risk factors and clinical outcome. Cureus 2018, 10, e2469. [Google Scholar] [CrossRef]
- Liu, P.Y.; Huang, Y.F.; Tang, C.W.; Chen, Y.Y.; Hsieh, K.S.; Ger, L.P.; Chen, Y.S.; Liu, Y.C. Staphylococcus lugdunensis infective endocarditis: a literature review and analysis of risk factors. J Microbiol Immunol Infect 2010, 43, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Frank, K.L.; Del Pozo, J.L.; Patel, R. From clinical microbiology to infection pathogenesis: how daring to be different works for Staphylococcus lugdunensis. Clin Microbiol Rev 2008, 21, 111–133. [Google Scholar] [CrossRef] [PubMed]
- Douiri, N.; Hansmann, Y.; Lefebvre, N.; Riegel, P.; Martin, M.; Baldeyrou, M.; Christmann, D.; Prevost, G.; Argemi, X. Staphylococcus lugdunensis: a virulent pathogen causing bone and joint infections. Clin Microbiol Infect 2016, 22, 747–748. [Google Scholar] [CrossRef]
- Argemi, X.; Prevost, G.; Riegel, P.; Keller, D.; Meyer, N.; Baldeyrou, M.; Douiri, N.; Lefebvre, N.; Meghit, K.; Ronde Oustau, C.; Christmann, D.; Cianferani, S.; Strub, J.M.; Hansmann, Y. VISLISI trial, a prospective clinical study allowing identification of a new metalloprotease and putative virulence factor from Staphylococcus lugdunensis. Clin Microbiol Infect 2017, 23, 334.e1–334. [Google Scholar] [CrossRef]
- Casaes Nunes, R.S.; Pires de Souza, C.; Pereira, K.S.; Del Aguila, E.M.; Flosi Paschoalin, V.M. Identification and molecular phylogeny of coagulase-negative staphylococci isolates from Minas Frescal cheese in southeastern Brazil: Superantigenic toxin production and antibiotic resistance. J Dairy Sci 2016, 99, 2641–2653. [Google Scholar] [CrossRef]
- Jiamboonsri, P.; Pithayanukul, P.; Bavovada, R.; Chomnawang, M.T. The inhibitory potential of Thai mango seed kernel extract against methicillin-resistant Staphylococcus aureus. Molec 2011, 16, 6255–6270. [Google Scholar] [CrossRef]
- Gupta, P.; Deshmukh, P.; Ravishankerv. Antimicrobial and phytochemical screening of Mangifera indica against skin aliments. J Pure ApplMicrobio 2010, 4, 387–392. [Google Scholar]
- Mirghani, M.E.S.; Yosuf, F.; Kabbashi, N.A.; Vejayan, J.; Yosuf, Z.B.M. Antibacterial activity of mango Kernel extracts. J Appl Sci 2009, 9, 3013–3019. [Google Scholar] [CrossRef]
- Kabuki, T.; Nakajima, H.; Arai, M.; Ueda, S.; Kuwabara, Y.; Dosako, S. Characterization of novel antimicrobial compounds from mango (Mangifera indica L.) kernel seeds. Food Chem. 2001, 71, 61–66. [Google Scholar] [CrossRef]
- Luo, F. L.v.Q.; Zhao, Y.; Hu, G.; Huang, G.; et al. Quantification and purification of mangiferin from chinese mango (Mangifera indica L.) cultivars and its protective effect on human umbilical vein endothelial cells under H2O2 induced stress. Int J Mol Sci 2012, 13, 11260–11274. [Google Scholar] [CrossRef]
- Salomon, S.; Sevilla, I.; Betancourt, R.; Romero, A.; Nuevas-Paz, L.; et al. Extraction of mangiferin from Mangifera indica L. leaves using microwave assisted technique. Emir J Food Agr 2014, 26, 616–622. [Google Scholar] [CrossRef]
- Singh, S.K.; Kumar, Y.; Sadish, K.S.; Sharma, V.K.; Dua, K.; et al. Antimicrobial evaluation of mangiferin analogues. Indian J Pharma Sci 2009, 71, 328–331. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Graphical illustration of the procedure to obtain mango extracts and to evaluate the in vitro antimicrobial potential against 20 bacterial species of food and human health relevance. Minimum inhibitory and bactericidal concentrations (MIC and MBC).
Figure 1.
Graphical illustration of the procedure to obtain mango extracts and to evaluate the in vitro antimicrobial potential against 20 bacterial species of food and human health relevance. Minimum inhibitory and bactericidal concentrations (MIC and MBC).
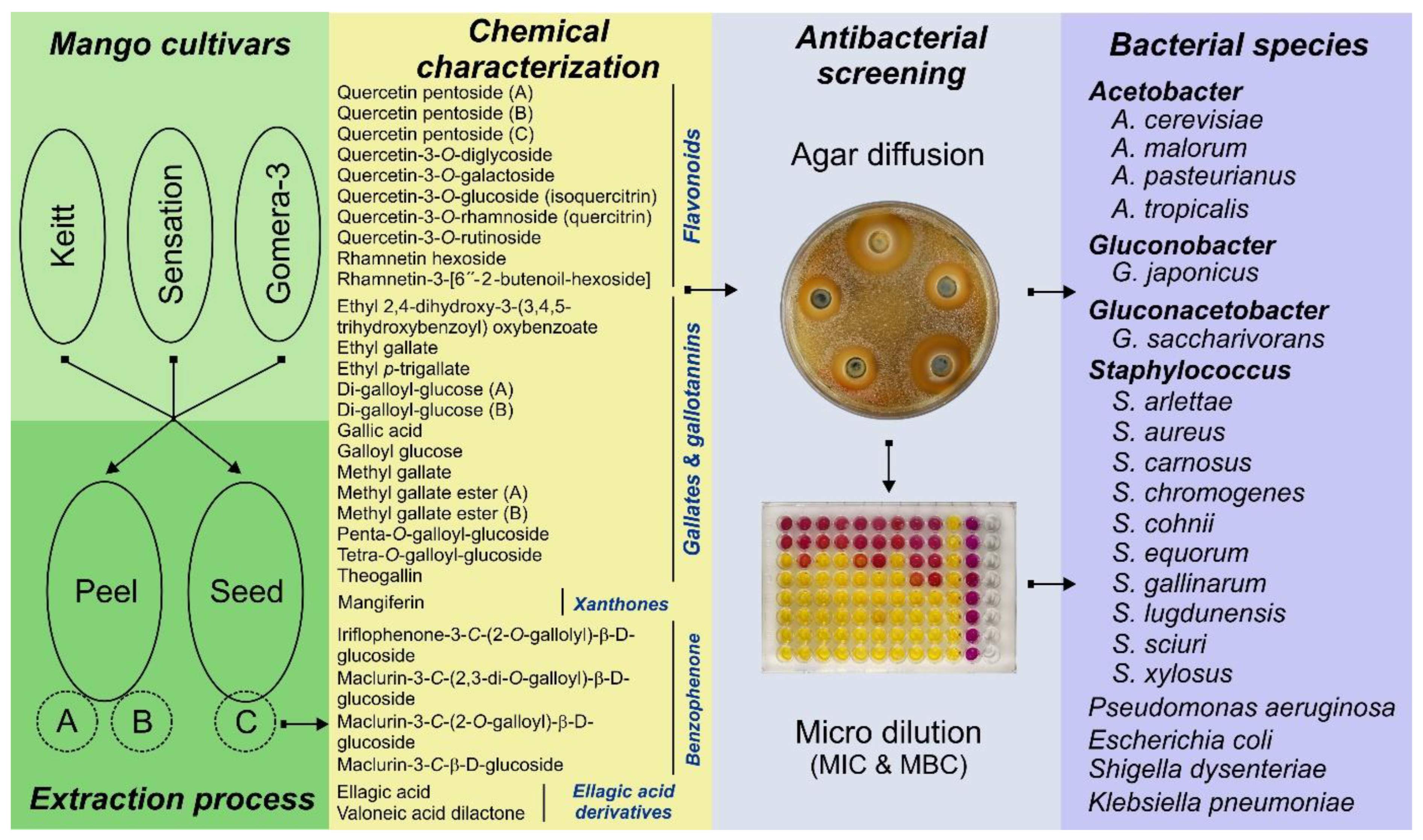
Figure 2.
Compounds identified and quantified in mango peel and seed extracts from three mango varieties produced in the Canary Island by HPLC-ESI-QTOF-MS. (a) Bioactive compounds identified in peel and seed extracts. (b) Quantified principal phenolic compounds (g/100 g dry weight) in peel and seed extracts by HPLC. KPA (Keitt Peel, extraction A); KPB (Keitt Peel, extraction B); SPA (identified in Sensation Peel, extraction A); SPB (Sensation Peel, extraction B); GPA (Gomera 3 Peel, extraction A); GPB (Gomera 3 Peel, extraction B); KSC (Keitt Seed, extraction C); SSC (Sensation Seed, extraction C); GSC (Gomera 3 Seed, extraction C). Each color represents a specific family of compounds. In compounds where no color is present, it indicates that only trace amounts of these compounds were detected.
Figure 2.
Compounds identified and quantified in mango peel and seed extracts from three mango varieties produced in the Canary Island by HPLC-ESI-QTOF-MS. (a) Bioactive compounds identified in peel and seed extracts. (b) Quantified principal phenolic compounds (g/100 g dry weight) in peel and seed extracts by HPLC. KPA (Keitt Peel, extraction A); KPB (Keitt Peel, extraction B); SPA (identified in Sensation Peel, extraction A); SPB (Sensation Peel, extraction B); GPA (Gomera 3 Peel, extraction A); GPB (Gomera 3 Peel, extraction B); KSC (Keitt Seed, extraction C); SSC (Sensation Seed, extraction C); GSC (Gomera 3 Seed, extraction C). Each color represents a specific family of compounds. In compounds where no color is present, it indicates that only trace amounts of these compounds were detected.
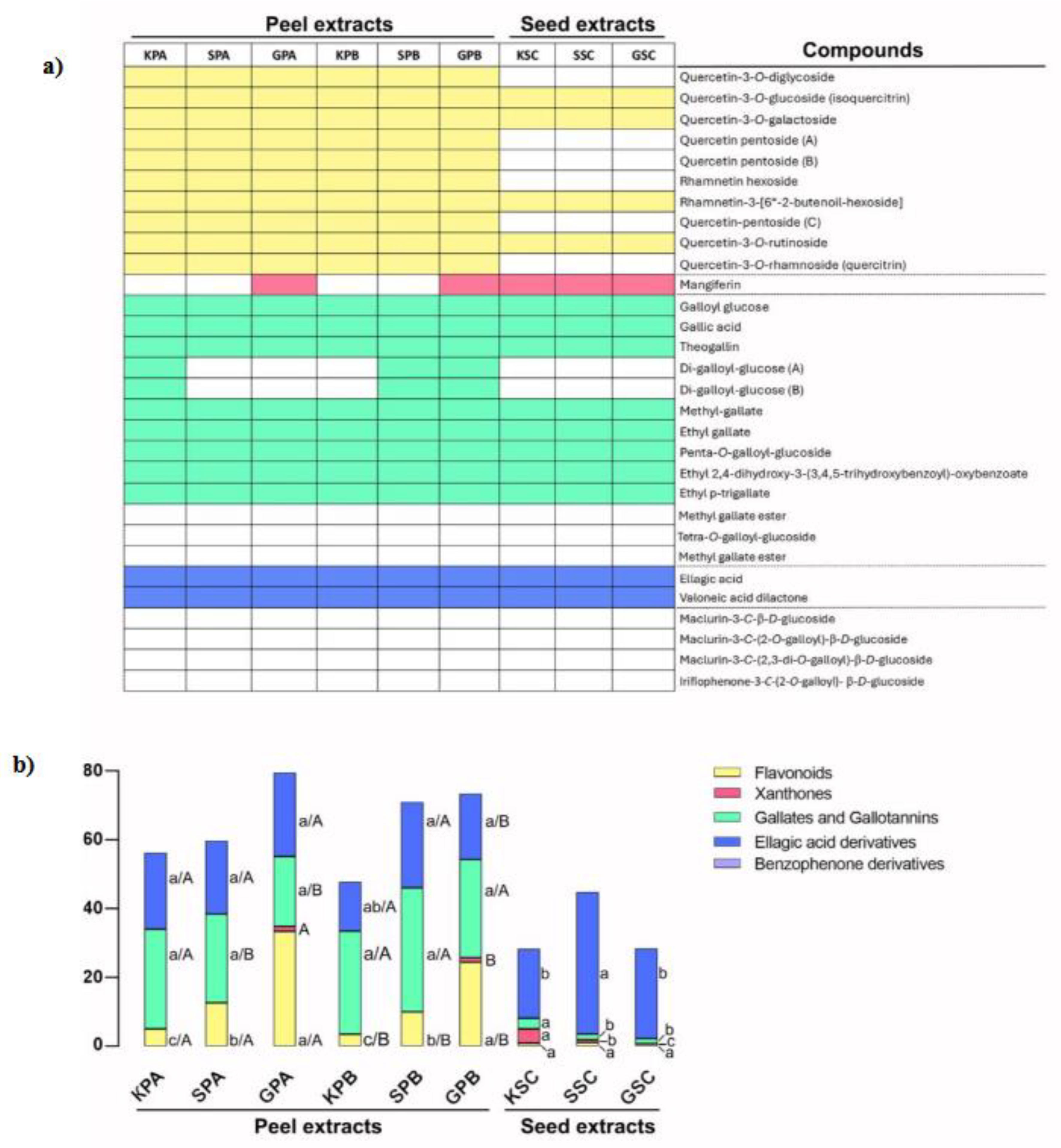

Table 1.
The minimum inhibitory and bactericidal concentrations (MIC and MBC) of mango peel and seed extracts against different acetic acid bacteria species.
Table 1.
The minimum inhibitory and bactericidal concentrations (MIC and MBC) of mango peel and seed extracts against different acetic acid bacteria species.
| AceticAcid Bacteria | Peel extracts (mg GAE/mL) | Seed extracts (mg GAE/mL) |
|||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GPA | SPA | KPA | GPB | SPB | KPB | GSC | SSC | KSC | |||||
| Group I | Acetobacter A. cerevisiae(Ac3-6A2) |
MIC | 8.3 | 8.3 | 8.3 | 1.7 | 8.3 | 1.7 | 8.3 | 17 | 17 | ||
| MBC | 25 | 25 | 25 | 17 | 25 | 17 | 50 | 50 | 50 | ||||
| A. cerevisiae(Ac5- T5) | MIC | 8.3 | 8.3 | 8.3 | 1.7 | 8.3 | 8.3 | 17 | 17 | 17 | |||
| MBC | 25 | 25 | 25 | 17 | 25 | 25 | 50 | 50 | 50 | ||||
| A. cerevisiae (Ac2-6A1) | MIC | 8.3 | 8.3 | 8.3 | 1.7 | 1.7 | 1.7 | 8.3 | 25 | 17 | |||
| MBC | 25 | 25 | 25 | 17 | 25 | 17 | 25 | 50 | 50 | ||||
| A. pasteurianus(Ap16-Lz75) | MIC | 50 | 50 | 25 | 8.3 | 50 | 25 | 17 | 8.3 | 8.3 | |||
| MBC | 100 | 100 | 100 | 50 | 100 | 100 | 50 | 50 | 50 | ||||
| A. malorum(Am17-T33) | MIC | 50 | 50 | 50 | 25 | 50 | 50 | 25 | 25 | 100 | |||
| MBC | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 200 | ||||
| A. malorum(Am25-P21) | MIC | 50 | 50 | 50 | 25 | 50 | 50 | 25 | 25 | 100 | |||
| MBC | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 200 | ||||
| A. malorum(Am26-Lz67) | MIC | 50 | 50 | 50 | 25 | 50 | 50 | 25 | 25 | 100 | |||
| MBC | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 50 | 200 | ||||
| Group II | A. tropicalis(At1-T191) | MIC | 100 | 200 | 200 | 100 | 200 | 100 | 50 | 50 | 200 | ||
| MBC | 200 | 400 | 400 | 200 | 400 | 200 | 100 | 100 | 400 | ||||
| A. tropicalis(At2-T59) | MIC | 100 | 200 | 200 | 100 | 200 | 100 | 50 | 50 | 200 | |||
| MBC | 200 | 400 | 400 | 200 | 400 | 200 | 100 | 100 | 400 | ||||
| Gluconobacter G. japonicus(Gj1-P37) |
MIC | 100 | 50 | 200 | 100 | 50 | 50 | 50 | 50 | 50 | |||
| MBC | 200 | 100 | 400 | 200 | 100 | 100 | 100 | 100 | 100 | ||||
| G. japonicus(Gj3-Lz59) | MIC | 100 | 50 | 200 | 100 | 50 | 50 | 50 | 50 | 50 | |||
| MBC | 200 | 100 | 400 | 200 | 100 | 100 | 100 | 100 | 100 | ||||
| G. japonicus(Gj2-P92) | MIC | 100 | 100 | 200 | 50 | 50 | 50 | 50 | 50 | 50 | |||
| MBC | 200 | 200 | 400 | 200 | 100 | 100 | 100 | 200 | 100 | ||||
| Gluconacetobacter G. saccharivorans(Gs1-T80) |
MIC | R* | R | R | R | R | R | R | R | R | |||
| MBC | R* | R | R | R | R | R | R | R | R | ||||
GPA, Gomera-3 Extracción A; SPA, Sensation Extracción A; KPA, Keitt Extracción A; GPB, Gomera-3 Extracción B; SPB, Sensation Extracción B; KPB, Keitt Extracción B; GSC, Gomera-3 Extracción C; SSC, Sensation Extracción C; KPC, Keitt Extracción C.*Resistant up to 400 mg gallic acid equivalents per mL (GAE/mL) (highest dose tested).
Table 4.
Spearman correlations between the phenolic compounds and the minimum inhibitory concentration (MIC) of each bacterial species tested.
Table 4.
Spearman correlations between the phenolic compounds and the minimum inhibitory concentration (MIC) of each bacterial species tested.
| Species | Isolate code | Flavonoids | Xanthones | Gallates gallotannins |
Ellagic acid derivatives |
Total phenol compounds |
|---|---|---|---|---|---|---|
| Acetobacter | ||||||
| A. cerevisiae | Ac3-6A2 | -0,4518 | 0,3708 | -0,5809 | 0,5809 | -0,5164 |
| A. cerevisiae | Ac5-T5 | -0,8385* | 0,2725 | -0,7267* | 0,5404 | -0,8385* |
| A. cerevisiae | Ac2-6A1 | -0,4433 | 0,4445 | -0,7714* | 0,4699 | -0,5497 |
| A. pasteurianus | Ap16-Lz75 | 0,5285 | -0,5339 | 0,4766 | 0,0693 | 0,5632 |
| A. malorum | Am17-T33 | 0,0559 | 0 | 0,3354 | -0,4099 | -0,0745 |
| A. malorum | Am25-P21 | 0,0559 | 0 | 0,3354 | -0,4099 | -0,0745 |
| A. malorum | Am26-Lz67 | 0,0559 | 0 | 0,3354 | -0,4099 | -0,0745 |
| A. tropicalis | At1-T191 | 0,2673 | -0,2745 | 0,5612 | -0,3742 | 0,1604 |
| A. tropicalis | At2-T59 | 0,2673 | -0,2745 | 0,5612 | -0,3742 | 0,1604 |
| Gluconobacter | ||||||
| G. japonicus | Gj1-P37 | 0,5578 | 0,13 | 0,2191 | -0,1594 | 0,5578 |
| G. japonicus | Gj3-Lz59 | 0,5578 | 0,13 | 0,2191 | -0,1594 | 0,5578 |
| G. japonicus | Gj2-P92 | 0,4781 | -0,2445 | 0,1394 | 0 | 0,3984 |
| Staphylococcus | ||||||
| S. arlettae | NRRL B-14764 | 0,414 | -0,1622 | -0,1035 | 0,6211 | 0,5175 |
| S. equorum subsp. equorum | NRRL B-14765 | 0,414 | -0,1622 | -0,1035 | 0,6211 | 0,5175 |
| S. carnosus subsp. carnosus | NRRL B-14760 | 0,1369 | 0,3575 | -0,4108 | 0,5477 | 0,1369 |
| S. xylosus | NRRL B-14776 | 0,1369 | 0,3575 | -0,4108 | 0,5477 | 0,1369 |
| S. sciuri subsp. sciuri | NRRL B-14767 | 0,1369 | 0,3575 | -0,4108 | 0,5477 | 0,1369 |
| S. gallinarum | NRRL B-14763 | 0,3651 | 0,0953 | -0,2739 | 0,5477 | 0,4564 |
| S. cohnii subsp. cohnii | NRRL B-14756 | 0,6739 | -0,4167 | 0,7625* | -0,5763 | 0,6739 |
| S. lugdunensis | 99705-65 | -0,0791 | -0,2615 | 0,4480 | -0,4216 | -0,0264 |
| S. aureus | 11923-76 | 0,4099 | -0,0973 | 0,4845 | -0,5963 | 0,4099 |
| Pseudomonas aeruginosa | ATCC 27853 | -0,8216* | 0,4767 | -0,8216* | 0,4564 | -0,8216* |
| Klebsiella pneumoniae | ATCC 13883 | -0,2739 | 0,286 | -0,7303* | 0,4564 | -0,3651 |
* Indicates significant correlation at p < 0.05.
Table 2.
The minimum inhibitory and bactericidal concentrations (MIC and MBC) of mango peel and seed extracts against different Staphylococcus species.
Table 2.
The minimum inhibitory and bactericidal concentrations (MIC and MBC) of mango peel and seed extracts against different Staphylococcus species.
| Staphylococcus species | Peel extracts (mg GAE/mL) |
Seed extracts (mg GAE/mL) |
|||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GPA | SPA | KPA | GPA | SPA | KPA | GSC | SSC | KSC | |||||
| Group I |
S. arlettae NRRL B-14764 |
MIC | 5 | 5 | 5 | 5 | 5 | 1 | 5 | 5 | 1 | ||
| MBC | 5 | 5 | 5 | 5 | 5 | 1 | 5 | 5 | 5 | ||||
|
S. equorum subsp. equorum NRRL B-14765 |
MIC | 5 | 5 | 5 | 5 | 5 | 1 | 5 | 5 | 1 | |||
| MBC | 5 | 5 | 5 | 5 | 5 | 1 | 5 | 5 | 5 | ||||
|
S. carnosus subsp. carnosus NRRL B-14760 |
MIC | 5 | 5 | 5 | 5 | 5 | 1 | 5 | 5 | 5 | |||
| MBC | 10 | 10 | 10 | 10 | 10 | 5 | 10 | 10 | 5 | ||||
|
S. xylosus NRRL B-14776 |
MIC | 5 | 5 | 5 | 5 | 5 | 1 | 5 | 5 | 5 | |||
| MBC | nd | nd | nd | nd | nd | nd | nd | nd | nd | ||||
|
S. sciuri subsp. sciuri NRRL B-14767 |
MIC | 5 | 5 | 5 | 5 | 5 | 1 | 5 | 5 | 5 | |||
| MBC | 30 | 15 | 15 | 30 | 15 | 15 | 120 | 120 | 120 | ||||
|
S. chromogenes NRRL B-14759 |
MIC | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | |||
| MBC | 5 | 10 | 10 | 5 | 10 | 5 | 10 | 10 | 5 | ||||
| Group II |
S. gallinarum NRRL B-14763 |
MIC | 10 | 10 | 5 | 10 | 10 | 5 | 10 | 10 | 5 | ||
| MBC | 30 | 15 | 10 | 15 | 15 | 10 | 240 | 240 | 240 | ||||
|
S. aureus subsp. aureus NRRL B-767 |
MIC | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 | |||
| MBC | nd | nd | nd | nd | nd | nd | nd | nd | nd | ||||
|
S. aureus 11923-76 |
MIC | 30 | 10 | 30 | 30 | 10 | 120 | 10 | 10 | 10 | |||
| MBC | 60 | 15 | 60 | 60 | 15 | 240 | 30 | 15 | 15 | ||||
|
S. cohnii subsp. cohnii NRRL B-14756 |
MIC | 15 | 15 | 10 | 15 | 15 | 60 | 5 | 5 | 5 | |||
| MBC | nd | nd | nd | nd | nd | nd | nd | nd | nd | ||||
|
S. lugdunensis 99705-65 |
MIC | 30 | 60 | 30 | 120 | 120 | 240 | 60 | 60 | 60 | |||
| MBC | 120 | >240 | 120 | 240 | 240 | >240 | 120 | 120 | 120 | ||||
GPA, Gomera-3 Extracción A; SPA, Sensation Extracción A; KPA, Keitt Extracción A; GPB, Gomera-3 Extracción B; SPB, Sensation Extracción B; KPB, Keitt Extracción B; GSC, Gomera-3 Extracción C; SSC, Sensation Extracción C; KPC, Keitt Extracción C; nd, not determined; mg gallic acid equivalents per mL (GAE/mL).
Table 3.
The minimum inhibitory and bactericidal concentrations (MIC and MBC) of mango peel and seed extracts against different bacterial species frequently associated with human infections.
Table 3.
The minimum inhibitory and bactericidal concentrations (MIC and MBC) of mango peel and seed extracts against different bacterial species frequently associated with human infections.
| Species | Peel extracts (mg GAE/mL) |
Seed extracts (mg GAE/mL) |
|||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GPA | SPA | KPA | GPB | SPB | KPB | GSC | SSC | KSC | |||||
|
Pseudomonas aeruginosa ATCC 27853 |
MIC | 10 | 10 | 10 | 10 | 10 | 10 | 30 | 30 | 30 | |||
| MBC | 30 | 30 | 30 | 30 | 30 | 30 | 60 | 60 | 30 | ||||
|
Escherichia coli ATCC 25922 |
MIC | 30 | 30 | 30 | 30 | 30 | 30 | 30 | 30 | 30 | |||
| MBC | 60 | 60 | 60 | 60 | 60 | 60 | 60 | 60 | 60 | ||||
|
Shigella dysenteriae ATCC 13313 |
MIC | 120 | 120 | 120 | 60 | 60 | 60 | 120 | 120 | 120 | |||
| MBC | >240 | >240 | >240 | >240 | >240 | >240 | >240 | >240 | >240 | ||||
|
Klebsiella pneumoniae ATCC 13883 |
MIC | 240 | 240 | 240 | 240 | 240 | 240 | 240 | 240 | 240 | |||
| MBC | >240 | >240 | >240 | >240 | >240 | >240 | >240 | >240 | >240 | ||||
GPA, Gomera-3 Extracción A; SPA, Sensation Extracción A; KPA, Keitt Extracción A; GPB, Gomera-3 Extracción B; SPB, Sensation Extracción B; KPB, Keitt Extracción B; GSC, Gomera-3 Extracción C; SSC, Sensation Extracción C; KPC, Keitt Extracción C; nd, not determined; mg gallic acid equivalents per mL (GAE/mL).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



