
Submitted:
15 April 2025
Posted:
15 April 2025
You are already at the latest version
Abstract
Biofilm formation is important for microbial survival, adaptation, and persistence within mutualistic and pathogenic systems in the Vibironaceae. Biofilms offer protection against environmental stressors, immune responses, and antimicrobial treatments by increasing host colonization and resilience. This review examines the mechanisms of biofilm formation in Vibrio species focusing on quorum sensing, cyclic-di-GMP signaling and host specific adaptations that influence biofilm structure and function. We discuss how biofilms differ between mutualistic and pathogenic species based on environmental and host signals. Recent advances in omics technologies such as transcriptomics and metabolomics have enhanced research in biofilm regulation under different conditions. Horizontal gene transfer and phase variation promotes greater fitness of bacterial biofilms due to the diversity of environmental isolates that utilize biofilms to colonize host species. Despite progress, questions remain regarding the long-term effects of biofilm formation and persistence on host physiology and biofilm community dynamics. Research integrating multidisciplinary approaches will help advance our understanding of biofilms and their implications for influencing microbial adaptation, symbiosis, and disease. These findings have applications in biotechnology and medicine, where genetic manipulation of biofilm regulation can enhance or disrupt microbiome stability and pathogen resistance, eventually leading to targeted therapeutic strategies.
Keywords:
symbiosis
; Vibrionaceae
; bioluminescence
; cephalopoda
1. Introduction
A biofilm is a structured community of microorganisms that adhere to surfaces and are embedded within a self-produced matrix of extracellular polymeric substances, or EPS. These microbial communities are highly organized and can be found in a wide range of environments. Biofilms play a pivotal role in the survival and persistence of many microorganisms, including both beneficial and pathogenic species. They are crucial for the establishment of symbiotic relationships between microbes and their hosts. In marine ecosystems, biofilms are integral to the establishment of host-microbe interactions, from nutrient exchange within the host to protection against environmental stressors. This review explores the significance of biofilms in a variety of symbiotic species within the Vibrionaceae. By examining biofilm-forming mechanisms in V. fischeri and its relatives, we will discuss the ecological and evolutionary significance of biofilms and broaden our understanding of biofilm formation across distinct species of Vibrio and Photobacterium in host-associated contexts.
One example of how biofilms provide a beneficial asset to the host are those found in sepiolid squids (Cephalopoda: Sepiolidae) and their bioluminescent symbionts in the genera Vibrio and Photobacterium. This association represents a tractable model system for studying the physiological, biochemical, and molecular mechanisms of animal host-microbe interactions. V. fischeri utilizes biofilm formation as a mechanism for host colonization, providing the squid with bioluminescence via quorum sensing, while benefiting from the nutrient-rich environment of the host light organ (Figure 1 [1]). Upon hatching, juvenile squids acquire V. fischeri from the surrounding seawater through a selective and targeted process. The bacteria then migrate toward and colonize the crypts of the squid’s light organ [1] In the nutrient-rich light organ, squids use V. fischeri for counterillumination during the night, a behavior that allows them to match downwelling moonlight and starlight, rendering it less visible to predators and prey below [2]. Biofilm formation enables V. fischeri to adhere to host epithelial surfaces in the light organ, grow rapidly, and establish a stable, self-sustaining population that replenishes itself each morning through a venting process with the onset of dawn. During this diurnal cycling, V. fischeri biofilm production is tightly regulated by a variety of genetic and environmental factors, most notably the quorum-sensing system LuxIR, which coordinates bacterial behavior in response to population cell density when cells reach late logrithimic phase right before dusk [3,4]. Additionally, the bacterium modulates its behavior in response to host-derived signals, such as nitric oxide and other immune factors, allowing it to adapt to the host light organ’s ever-changing internal environment [3,5,6] These biofilms not only enable attachment and colonization but also play crucial roles in protecting V. fischeri from host immune responses and environmental challenges, fostering a stable and persistent symbiosis [3,4,5,7,8,9].
However, biofilm-forming capabilities of V. fischeri are not unique in the microbial world. Other Vibrio species, such as V. cholerae and V. vulnificus, as well as Photobacterium species, exhibit diverse biofilm-forming strategies that enable them to thrive in both pathogenic and symbiotic states [10,11]. In the case of V. cholerae, biofilm formation is essential for its ecological persistence and successful colonization of the human intestine, with biofilm structures enhancing resistance to environmental stresses and immune responses [12,13]. Similarly, V. vulnificus, a known marine pathogen, forms biofilms on both biotic and abiotic surfaces, having a critical role in its pathogenicity and persistence in marine and clinical environments. In contrast, Photobacterium species, which form beneficial relationships with marine hosts, produce biofilms that enhance stability and facilitate bioluminescence, contributing to the fitness of both the host and the symbiont in deep-sea ecosystems [13,14]. Such studies of biofilm formation in Vibrio and Photobacterium species have far-reaching implications for understanding the microbial ecology, evolution, and persistence throughout various habitats [15,16,17].
Biofilms are a critical component of microbial life in diverse habitats, especially in marine ecosystems, where they influence ecosystem health, resilience, and function. Marine biofilms have an essential role in nutrient cycling, contributing to the regeneration of organic matter, nitrogen fixation, and other biogeochemical processes. For instance, biofilms in coral reefs, seagrasses, and oyster beds mediate cycling of nitrogen and other nutrients, thus supporting primary productivity and maintaining the stability of these vital ecosystems [18]. In these habitats, biofilms also help protect microbial communities from external stressors such as ocean acidification and warming, which threaten the integrity of their hosts and subsequently their environment. Biofilms are also integral to microbial survival, contributing to colonization, immune evasion, and nutrient exchange in many animal mutualisms. For example, biofilms formed by coral-associated Vibrio species play a complex role, simultaneously supporting beneficial associations when the host is healthy, while contributing to pathogenesis and coral disease under stressful conditions such prolonged exposure to high temperatures [19]. Understanding the delicate balance between mutualistic and pathogenic life histories of biofilms is critical for addressing global challenges such as coral bleaching, which are impacted by climate change [20].
Furthermore, biofilms in other host systems, such as those formed by V. vulnificus on shellfish or medical devices, highlight their importance in both public health concerns and their undetectable persistence within the environment.
Biofilm research also offers valuable insights into microbial adaptation and evolutionary dynamics of pathogenesis. The protective and structured environment of a biofilm allows microorganisms to endure fluctuating environmental conditions, develop resistance to antimicrobial agents, and acquire novel traits through genetic exchange, such as horizontal gene transfer [21,22,23]. For example, the close proximity of bacteria in biofilms facilitate the exchange of genes involved in antibiotic resistance or virulence factors between bacterial cells, which can have profound implications for both the emergence and evolution of pathogens [24,25]. The ability of biofilms to facilitate the adaptation of microbes to changing environments underscores their significance in applied microbiology [26,27]. Beyond fundamental ecological and biological insights, biofilm research holds great promise for various practical applications. In biotechnology, biofilms can be engineered for a range of purposes, such as in bioremediation, where they help degrade environmental pollutants [28,29], or in synthetic biology, where biofilm-forming bacteria are harnessed for the development of bio-based materials and devices [30]. Medical applications are dependent upon understanding biofilm formation for addressing chronic infections and microbial resistance, particularly in the context of biofilm-associated infections on medical devices and implants [31]. Furthermore, by studying biofilm dynamics in natural and clinical settings, we can better understand the persistence and spread of such pathogens, helping to develop public health strategies and policies [32].
2. Biofilms: Essential Structures in Symbiosis
Biofilms play a pivotal role in facilitating host-microbe interactions during symbiosis. By forming biofilms, mutualistic bacteria like V. fischeri can adhere to host tissues, resist abiotic stresses, and establish long-term associations with their host [33]. Biofilms are crucial in the colonization of the Hawaiian bobtail squid Euprymna scolopes, as it enables V. fischeri to populate the squid’s light organ and maintain bioluminescence as well as reduce immune responses from the host [1,34]. Other species, such as V. vulnificus, form biofilms on marine detritus, aiding in decomposition, nutrient acquisition, and exchange of important metabolites and possibly DNA [12,35]. Similarly, Photobacterium species form biofilms on fish skin (broad host range) and within fish light organs (species specific association), and only those associated with light organ containing hosts contribute to host protection against predators or communication through bioluminescence [36]. Within the light organ, beneficial bacteria recycle host-derived nutrients such as amino acids and peptides, while producing bioluminescence that benefits the host [37]. These interactions are also influenced by a variety of environmental factors and host-derived signaling that regulates biofilm development.
Environmental conditions, including light exposure [25,26], and host-derived signals such as nitric oxide and antimicrobial peptides, regulate biofilm formation in V. fischeri, influencing bacterial stress responses, gene expression, and biofilm architecture [16,38]. These dynamic interactions create a feedback loop between V. fischeri and its host by fine-tuning biofilm growth to optimize V. fischeri under such varying environmental conditions. For example, light regulated LuxIR signal integrates with the AinS quorum sensing pathway to modulate biofilm structure and density, allowing the bacteria to adjust their attachment and dispersal in response to population density and host signals [39]. Host secreted nutrients regulate bacterial metabolism, exopolysaccharides, and the adhesion necessary for biofilm stability [40,41]. Signaling via H-NOX regulators control bacterial gene expression, influencing stress responses and contributing to the regulation of colonization in symbiotic V. fischeri [42]. Together, these regulatory systems impose an adaptive response that allows V. fischeri to establish and maintain a stable and dynamic symbiotic association with bobtail squids.
3. Vibrio Fischeri as a Model for the Study of Host Associated Biofilms
Additionally, symbiosis between V. fischeri and the Hawaiian bobtail squid E. scolopes (Cephalopoda: Sepioliadae) serves as a hallmark model for studying the molecular mechanisms of biofilm regulation and production [3]. Quorum sensing is central to biofilm formation and symbiosis in V. fischeri. The LuxIR system mediates cell-density-dependent regulation of bioluminescence and biofilm maturation [43]. As bacterial populations grow, the autoinducer N-acyl-homoserine lactone (AHL) accumulates, triggering the activation of genes necessary for biofilm, light production, and other group behaviors such as motility and virulence [44,45]. Additionally, the syp symbiosis polysaccharide locus regulates the synthesis of extracellular matrix components essential for biofilm stability and structural integrity [46]. Mutations in genes such as rscS and sypG disrupt syp locus activation, impairing biofilm production and reducing the bacterium’s ability to establish stable biofilms with the host [46,47].
Beyond quorum sensing, colonization is also influenced by additional molecular regulators such as genes controlling chemotaxis and adherence [48]. For example, Clp has elevated expression which is often linked to better biofilm formers and more successful colonizers in V. fischeri biofilms [16]. sypF, a hybrid sensor kinase, coordinates biofilm formation via upstream response regulators that control pili expression and function that are essential for effective colonization [49]. Symbiotic loci such as pilA, flaA, and mshA are important for V. fischeri to locate, colonize, and attach to the host light organ. For example, flaA mutations result in impaired motility and reduction in colonization efficiency which highlights the importance of motility related genes for a successful symbiosis [50] V. fischeri uses flagellar motility and pili to navigate chemical gradients in the host, such as those produced by chitin oligosaccharides, and help them attach firmly to host tissues [51,52]. Surface adhesins, including those encoded by type IV pili and msh genes, further mediate bacterial attachment and biofilm stabilization [9]. For example, the pil operon in V. fischeri (pilABCD) encodes for components of the type IV pili, which produce a protein essential for attachment and persistence. Molecular differences in pilB and pilD loci found in different V. fischeri strains suggest these genes are critical for host recognition and specificity. Swapping pili genes from different V. fischeri strains demonstrate how such subtle differences in these loci are responsible for native versus non-native strain specificity [7,16,33].
4. Expanding the Lens: Other Vibrio and Photobacterium Species
While V. fischeri serves as an exemplary model for biofilms during symbiosis, other Vibrio species and related bacteria exhibit distinct biofilm strategies that highlight the versatility of biofilm-mediated interactions. Comparative studies on V. cholerae, V. vulnificus, and Photobacterium species reveal a wide array of biofilm mechanisms that contribute to their ecological success [53]. For instance, V. cholerae biofilms are crucial for environmental persistence and host pathogenicity, utilizing exopolysaccharides and extracellular DNA to enhance surface attachment and microbial resilience [54,55]. These biofilms facilitate V. cholerae survival in aquatic environments and transition to human hosts, providing an important role in its transmission and virulence [56,57,58]. Similarly, V. vulnificus forms robust biofilms on biotic surfaces (exoskeletons of arthropods) and abiotic surfaces (sand), creating a stable habitat as well as a springboard for opportunistic infections [59,60]. The ability of V. vulnificus to form biofilms on or in shellfish and medical devices contributes to its persistence in both natural and clinical settings, making it a significant concern for public health [61,62]. Photobacterium species, particularly those associated with deep-sea fish, form structured biofilms that support bioluminescence, indicating a functional convergence in biofilm-mediated interactions across Vibrio bacteria [31,36,63]. The biofilm matrix in these symbiotic interactions enhances the stability of luminescent bacteria, optimizing light production for host communication (high cell density), predator avoidance, and prey attraction [32,64].
Symbiotic versatility is evident in biofilms across different host environments. While V. fischeri establishes biofilms within the squid light organ, other Vibrio species colonize diverse hosts, including fish and many types of invertebrates. The variability in host-derived signals, including immune factors, microbial competitors, and nutrient availability, influences bacterial biofilm architecture and persistence differentially [3,65,66]. For example, biofilms in fish-associated Vibrio strains often exhibit high resistance to host immune responses, whereas coral-associated Vibrio biofilms play a dual role in both mutualistic and pathogenic associations under shifting environmental conditions [67,68,69]. Vibrio biofilms aid in nutrient cycling and organic matter decomposition in marine ecosystems. This enhances primary productivity and facilitates microbial interactions that regulate carbon and nitrogen fluxes. [19,43,70]. Additionally, interspecies biofilm interactions, such as those observed in coral-associated Vibrio communities, can influence ecosystem health and resilience against environmental stressors such as ocean acidification and warming [12,69,70]. For example, Vibrio biofilms participate in nitrogen cycling by interacting with other microbial communities that regulate denitrification processes, impacting water quality and ecosystem stability in oyster beds [71]
5. Adaptive and Functional Roles of Biofilms in Symbiosis
Biofilms are tightly regulated by genetic and molecular pathways that orchestrate surface attachment, matrix production, and dispersal [72]. Quorum sensing, a cell-density-dependent communication system, plays a central role in regulating bioluminescence and biofilm architecture across several Vibrio species [39,54,73,74]. V. cholerae utilizes the HapR regulator to modulate biofilm dispersal, balancing attachment with planktonic growth [40]. Additionally, the c-di-GMP signaling pathway integrates environmental cues such as nutrient availability and stress signals that regulate key processes in biofilm formation including matrix production as well as cell adhesion [34,75,76]. Variations in c-di-GMP levels are differentiated amongst Vibrio and Photobacterium species, contributing to species-specific biofilm strategies and resilience to environmental stressors [34,75]
Biofilms also provide a protective niche that enhances bacterial persistence in different environments by protecting bacteria from environmental stressors, antibiotics, and immune effectors. [77,78] In host associated biofilms, immune evasion occurs through mechanisms such as the suppression of innate immune response (inflammation, phagocytosis, and other immune cells), and modulating host signaling pathways [79,80,81]. For example, V. fischeri biofilms in the squid light organ protect the bacteria from oxidative stress while it simultaneously promotes symbiotic colonization [3,66]. However, V vulnificus biofilms are adept to resisting phagocytosis and antimicrobial peptides contributing to their virulence [82]. The V. vulnificus biofilm matrix contains polysaccharides and other extracellular components that act as a physical barrier to the environment [83]. The dense structure of biofilms protects against antimicrobial peptides, allowing their persistence despite the host immune response [84].
Host-specific adaptations select specific biofilm traits through targeted specialization. Different hosts exert various selective pressures on their symbiotic bacteria that influences the genetic makeup of associated Vibrio species. With the development of whole genome sequencing and genome wide associated studies (GWAS), it has been revealed that Vibrio species associated with different hosts exhibit variations in biofilm-associated gene clusters, which are involved in production of exopolysaccharide and structural components essential for biofilm formation [84,85]. For example, pathogenic Vibrio species may acquire additional genes or operons that enhance exopolysaccharide production, allowing the formation of biofilms that are more robust and resistant to their host immune response [86]. Mutualistic strains may lose certain genes involved in biofilm production and regulation, relying instead on certain host derived factors such as nutrients or signaling molecules to stabilize biofilms [87]. These genetic adaptations are driven by host specific immune factors and competition amongst other microbes in the surrounding environment [88,89].
In the E. scolopes- Vibrio mutualism, the molecular mechanisms of biofilm regulation has been well-studied, but there are still areas that are not well understood. Since biofilm formation is essential for colonization and persistence, any subtle changes can be either beneficial or detrimental to the fitness of those symbionts [3,89]. Advancements in omics-based technologies facilitate the study of biofilm regulation at multiple levels. Transcriptomics and proteomics have revealed gene expression patterns associated with biofilm development, such as the upregulation of genes involved in surface attachment, matrix production, stress response, and quorum sensing [16,17,90], while metabolomics identifies key metabolic pathways that sustain biofilm communities [16,90]. CRISPR-based functional genomics approaches allow targeted studies of biofilm-related genes, providing mechanistic insights into their roles in host specificity and host switching [91,92]. While V. fischeri uses biofilms for mutualistic benefits, V. vulnificus biofilms enable the bacterium to evade immune responses. The different functions of biofilms highlight how bacteria can utilize and optimize biofilm phenotypes for survival, whether through cooperation with the host or to resist host defense mechanisms during pathogenesis.
6. Advances in Understanding Marine Biofilms
Recent research has shown the adaptability of marine biofilms, with microbial communities’ ability to thrive under fluctuating environmental conditions. Biofilm structure and composition exhibit dynamic alterations based on environmental factors such as temperature, salinity, and nutrient availability [8,26,66,93,94]. Environmental shifts such as climate change and human induced habitat alterations drive biofilm evolution. These changes highlight the plasticity and ability of biofilm-forming bacteria to adapt quickly, offering us insights into microbial evolution in real time.
In marine symbiotic systems, novel regulators of biofilms continue to be identified, such as hybrid two-component systems that enable bacteria to sense and respond to host-secreted metabolites [49,95,96]. Additionally, transcriptomic analyses have uncovered species-specific regulatory networks that differentiate commensal from pathogenic biofilms, shedding light on the fine-scale genetic adaptations that dictate host interactions [17,97,98]. Mechanisms of biofilm evolution serve as incubators for microbial diversification within the biofilm community as well as the planktonic bacteria that are released from these structures. The protective and close-knit environment of biofilms enhances horizontal gene transfer, facilitating the spread of adaptive traits such as antibiotic resistance, metabolic versatility, and host colonization factors [22,99]. Studies have demonstrated that biofilm-mediated selection pressures drive the emergence of phenotypic variants with enhanced fitness, contributing to microbial evolution in both natural and host-associated settings [5,27,100]. Vibrio biofilms promote phase variation and genetic heterogeneity, enabling populations to withstand environmental fluctuations and host immune challenges [7]. Understanding these evolutionary mechanisms offers insights into the persistence and resilience of V. fischeri within its mutualistic relationship with the squid host, revealing fundamental processes that govern microbial community dynamics and host-microbe interactions. This knowledge not only contributes to our understanding of microbial evolution and ecology at the species level, but provides insight on how these systems influence broader ecosystem functions, biodiversity, and the adaptability of organisms in the face of environmental pressures.
As we continue to investigate biofilm formation, structure, and function in host-associated systems, it is imperative to include the molecular mechanisms of biofilm dispersal in beneficial versus pathogenic Vibrio species, the role of interspecies interactions in biofilm architecture [5,101], and how these interactions shape stability within host-associated microbial communities. Additionally, metabolic exchange between biofilm-forming bacteria and their hosts, particularly in marine symbiotic systems, is being explored with a focus on nutrient exchange and chemical signaling [66,70]. To address these gaps, integrative approaches combining genomics, metabolomics, and live-imaging techniques are needed to better understand the dynamics of biofilm development in symbiotic relationships. The study of biofilms has significant biotechnological implications. Engineered biofilms hold promise for applications in bioremediation, wastewater treatment, and industrial fermentation processes [102,103]. For example, disrupting quorum sensing pathways or biofilm matrix synthesis has emerged as a promising strategy for controlling biofilm-associated pathogens [104]. By leveraging biofilm research, we can harness microbial communities for sustainable solutions while mitigating biofilm-associated challenges in both environmental and clinical settings.
7. Conclusion and Future Directions
The study of biofilms in V. fischeri and other symbiotic members of the Vibrionaceae provides critical insights into microbial ecology, host-microbe interactions, and evolutionary dynamics of how these intricate structures evolve and persist in our environment. In marine habitats, biofilms contribute to microbial succession, and the structuring of microbial communities on surfaces ranging from host tissues to substrates like sediments and ship hulls [12]. Biofilms are fundamental to the persistence of many different types of symbioses, facilitating colonization, enhancing microbial resilience, and shaping ecosystem processes [3,5,12]. Their formation enhances bacterial stability and host colonization, as observed in the structured biofilms of V. fischeri that mediate light organ colonization in sepiolid squid species [3,105]. Similarly, biofilms in pathogenic Vibrio species play a role in disease progression, demonstrating the dual nature of biofilm-mediated persistence [68].
Comparative analyses of biofilm strategies across Vibrio species reveal conserved regulatory mechanisms while highlighting species-specific adaptations. The quorum sensing and c-di-GMP signaling pathways that regulate biofilms are shared among multiple Vibrio species, yet their functional outputs vary depending on ecological niche and host interactions [106]. For example, while V. cholerae biofilms enhance environmental survival and transmission via waterborne routes, V. fischeri biofilms are tailored for long-term mutualistic colonization within squid and fish light organs [3,59]. These findings emphasize the ecological plasticity of biofilm regulation and the evolutionary pressures that shape biofilm-associated lifestyles in marine bacteria.
Despite these advancements, several key questions remain unanswered. How do biofilm-associated genetic networks evolve in response to environmental pressures? What are the long-term consequences of biofilms on host physiology and microbial community dynamics? Recent studies suggest that biofilms may drive microbial diversification through horizontal gene transfer and phase variation, yet the extent to which these processes contribute to long-term microbial adaptation remains unclear [6,92,107] Additionally, the interplay between host immune responses and biofilm formation requires further investigation, particularly in symbiotic systems where biofilms may actively modulate host signaling to maintain stable interactions [3,6,39]. Addressing these questions will require interdisciplinary approaches that integrate molecular biology, ecology, and computational modeling to develop a holistic understanding of biofilm dynamics.
Future biofilm research has great promise for applied microbial sciences. Understanding biofilm regulation can inform strategies for modulating microbial interactions in aquaculture, biotechnology, and medicine. In aquaculture, promoting beneficial biofilms may enhance probiotic effectiveness and pathogen resistance in farmed marine species [81]. In medicine, unraveling biofilm-associated resistance mechanisms can aid in the development of novel antimicrobial strategies, including quorum sensing inhibitors and biofilm-targeting therapeutics [23,108]. Furthermore, exploring biofilm resilience in marine environments may provide insights into microbial responses to climate change and anthropogenic impacts, such as ocean acidification and pollution [109,110]. Given their fundamental role in microbial ecology, biofilms represent a key focus for advancing both basic and applied microbiological research. By expanding our knowledge of biofilm biology, we can harness microbial communities for innovative solutions while mitigating biofilm-associated challenges in health and industry. As novel technologies, such as single-cell imaging, real-time metabolomics, and CRISPR-based functional genomics continue to advance, they will enable deeper exploration of biofilm heterogeneity and its implications for microbial adaptation and survival [92,109,111,112,113]. Integrating these approaches across diverse marine systems will not only deepen our understanding of biofilm-mediated symbioses but also provide actionable strategies for managing biofilms in both beneficial and detrimental contexts.
References
- Nyholm, S. V; McFall-Ngai, M.J. A lasting symbiosis: how the Hawaiian bobtail squid finds and keeps its bioluminescent bacterial partner. Nat Rev Microbiol 2021, 19, 666–679. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.W.; Nishiguchi, M.K. Counterillumination in the hawaiian bobtail squid, Euprymna scolopes Berry (Mollusca: Cephalopoda). Mar Biol 2004, 144, 1151–1155. [Google Scholar] [CrossRef]
- Fung, B.L.; Esin, J.J.; Visick, K.L. Vibrio fischeri : a model for host-associated biofilm formation. J Bacteriol 2024, 206. [Google Scholar] [CrossRef] [PubMed]
- Nyholm, S. V.; McFall-Ngai, M. The winnowing: establishing the squid–Vibrio symbiosis. Nat Rev Microbiol 2004, 2, 632–642. [Google Scholar] [CrossRef]
- Soto, W.; Nishiguchi, M.K. Microbial experimental evolution as a novel research approach in the Vibrionaceae and squid-Vibrio symbiosis. Front Microbiol 2014, 5. [Google Scholar] [CrossRef]
- Soto, W.; Nishiguchi, M.K. Environmental stress selects for innovations that drive Vibrio symbiont diversity. Front Ecol Evol 2021, 9. [Google Scholar] [CrossRef]
- Chavez-Dozal, A.; Hogan, D.; Gorman, C.; Quintanal-Villalonga, A.; Nishiguchi, M.K. Multiple Vibrio fischeri genes are involved in biofilm formation and host colonization. FEMS Microbiol Ecol 2012, 81, 562–573. [Google Scholar] [CrossRef]
- Nourabadi, N.; Nishiguchi, M.K. Ph adaptation drives diverse phenotypes in a beneficial bacterium-host mutualism. Front Ecol Evol 2021, 9. [Google Scholar] [CrossRef]
- Chavez-Dozal, A.A.; Gorman, C.; Lostroh, C.P.; Nishiguchi, M.K. Gene-swapping mediates host specificity among symbiotic bacteria in a beneficial symbiosis. PLoS One 2014, 9, e101691. [Google Scholar] [CrossRef]
- Visick, K.L.; Schembri, M.A.; Yildiz, F.; Ghigo, J.-M. Biofilms 2015: multidisciplinary approaches shed light into microbial life on surfaces. J Bacteriol 2016, 198, 2553–2563. [Google Scholar] [CrossRef]
- Yildiz, F.H.; Visick, K.L. Vibrio biofilms: so much the same yet so different. Trends Microbiol 2009, 17, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Dang, H.; Lovell, C.R. Microbial surface colonization and biofilm development in marine environments. Microbiol. Mol Biol Rev 2016, 80, 91–138. [Google Scholar] [CrossRef]
- Takemura, A.F.; Chien, D.M.; Polz, M.F. Associations and dynamics of Vibrionaceae in the environment, from the genus to the population level. Front Microbiol 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Zamborsky, D.J.; Nishiguchi, M.K. Phylogeographical patterns among mediterranean Sepiolid squids and their Vibrio symbionts: environment drives specificity among sympatric species. Appl Environ Microbiol 2011, 77, 642–649. [Google Scholar] [CrossRef] [PubMed]
- Chavez-Dozal, A.; Gorman, C.; Erken, M.; Steinberg, P.D.; McDougald, D.; Nishiguchi, M.K. Predation response of Vibrio fischeri biofilms to bacterivorus protists. Appl Environ Microbiol 2013, 79, 553–558. [Google Scholar] [CrossRef]
- Chavez-Dozal, A.; Gorman, C.; Nishiguchi, M.K. Proteomic and metabolomic profiles demonstrate variation among free-living and symbiotic Vibrio fischeri biofilms. BMC Microbiol 2015, 15, 226. [Google Scholar] [CrossRef]
- Chavez-Dozal, A.; Soto, W.; Nishiguchi, M.K. Identification of a transcriptomic network underlying the wrinkly and smooth phenotypes of Vibrio fischeri. J Bacteriol 2021, 203. [Google Scholar] [CrossRef]
- Yanovski, R.; Barak, H.; Brickner, I.; Kushmaro, A.; Abelson, A. The microbial community of coral reefs: biofilm composition on artificial substrates under different environmental conditions. Mar Biol 2024, 171, 74. [Google Scholar] [CrossRef]
- de Carvalho, C.C.C.R. Marine Biofilms: A successful microbial strategy with economic implications. Front Mar Sci 2018, 5. [Google Scholar] [CrossRef]
- Mao-Jones, J.; Ritchie, K.B.; Jones, L.E.; Ellner, S.P. How microbial community composition regulates coral disease development. PLoS Biol 2010, 8, e1000345. [Google Scholar] [CrossRef]
- Broszat, M.; Grohmann, E. Horizontal gene transfer in planktonic and biofilm modes. In; 2014; pp. 67–95.
- Madsen, J.S.; Burmølle, M.; Hansen, L.H.; Sørensen, S.J. The interconnection between biofilm formation and horizontal gene transfer. FEMS Immunol Med Microbiol 2012, 65, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.W.; Mah, T.-F. Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol Rev 2017, 41, 276–301. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, M.; Brooks, B.; Brooks, A. The complex relationship between virulence and antibiotic resistance. Genes (Basel) 2017, 8, 39. [Google Scholar] [CrossRef] [PubMed]
- Ghazvinian, M.; Asgharzadeh Marghmalek, S.; Gholami, M.; Amir Gholami, S.; Amiri, E.; Goli, H.R. Antimicrobial resistance patterns, virulence genes, and biofilm formation in enterococci strains collected from different sources. BMC Infect Dis 2024, 24, 274. [Google Scholar] [CrossRef]
- Soto, W.; Gutierrez, J.; Remmenga, M.D.; Nishiguchi, M.K. Salinity and temperature effects on physiological responses of Vibrio fischeri from diverse ecological niches. Microb Ecol 2009, 57, 140–150. [Google Scholar] [CrossRef]
- Soto, W.; Rivera, F.M.; Nishiguchi, M.K. Ecological diversification of Vibrio fischeri serially passaged for 500 generations in novel squid host Euprymna tasmanica. Microb Ecol 2014, 67, 700–721. [Google Scholar] [CrossRef]
- McFall, A.; Coughlin, S.A.; Hardiman, G.; Megaw, J. Strategies for biofilm optimization of plastic-degrading microorganisms and isolating biofilm formers from plastic-contaminated environments. Sustain Microbiol 2024, 1. [Google Scholar] [CrossRef]
- Mitra, A.; Mukhopadhyay, S. Biofilm mediated decontamination of pollutants from the environment. AIMS Bioeng 2016, 3, 44–59. [Google Scholar] [CrossRef]
- Shi, Y.; Chen, T.; Shaw, P.; Wang, P.-Y. Manipulating bacterial biofilms using materiobiology and synthetic biology approaches. Front Microbiol 2022, 13. [Google Scholar] [CrossRef]
- Mishra, A.; Aggarwal, A.; Khan, F. Medical device-associated infections caused by biofilm-forming microbial pathogens and controlling strategies. Antibiotics 2024, 13, 623. [Google Scholar] [CrossRef]
- Machineni, L. Effects of biotic and abiotic factors on biofilm growth dynamics and their heterogeneous response to antibiotic challenge. J Biosci 2020, 45, 25. [Google Scholar] [CrossRef]
- Chavez-Dozal, A.; Nishiguchi, M.K. Variation in biofilm formation among symbiotic and free-living strains of Vibrio fischeri. J Basic Microbiol 2011, 51, 452–458. [Google Scholar] [CrossRef] [PubMed]
- Waters, C.M.; Lu, W.; Rabinowitz, J.D.; Bassler, B.L. Quorum sensing controls biofilm formation in Vibrio cholerae through modulation of cyclic di-GMP levels and repression of VpsT. J Bacteriol 2008, 190, 2527–2536. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Park, S.; Lee, K. Role of ntrc-regulated exopolysaccharides in the biofilm formation and pathogenic interaction of Vibrio vulnificus. Mol Microbiol 2009, 74, 436–453. [Google Scholar] [CrossRef] [PubMed]
- Urbanczyk, H.; Ast, J.C.; Dunlap, P. V. Phylogeny, genomics, and symbiosis of Photobacterium. FEMS Microbiol Rev 2011, 35, 324–342. [Google Scholar] [CrossRef]
- Battin, T.J.; Kaplan, L.A.; Denis Newbold, J.; Hansen, C.M.E. Contributions of microbial biofilms to ecosystem processes in stream mesocosms. Nature 2003, 426, 439–442. [Google Scholar] [CrossRef]
- Koch, E.J.; Moriano-Gutierrez, S.; Ruby, E.G.; McFall-Ngai, M.; Liebeke, M. The impact of persistent colonization by Vibrio fischeri on the metabolome of the host squid Euprymna scolopes. J Exp Biol 2020, 223. [Google Scholar] [CrossRef]
- Fung, B.L.; Visick, K.L. LitR and its quorum-sensing regulators modulate biofilm formation by Vibrio fischeri. J Bacteriol 2025. [Google Scholar] [CrossRef]
- Septer, A.N.; Visick, K.L. Lighting the way: how the Vibrio fischeri model microbe reveals the complexity of earth’s “simplest” life forms. J Bacteriol 2024, 206. [Google Scholar] [CrossRef]
- Pan, M.; Schwartzman, J.A.; Dunn, A.K.; Lu, Z.; Ruby, E.G. A single host-derived glycan impacts key regulatory nodes of symbiont metabolism in a coevolved mutualism. mBio 2015, 6. [Google Scholar] [CrossRef]
- Wang, Y.; Dufour, Y.S.; Carlson, H.K.; Donohue, T.J.; Marletta, M.A.; Ruby, E.G. H-nox–mediated nitric oxide sensing modulates symbiotic colonization by Vibrio fischeri. Proc Nat Acad Sci 2010, 107, 8375–8380. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.C.; Miyashiro, T. Quorum sensing in the squid-Vibrio symbiosis. Int J Mol Sci 2013, 14, 16386–16401. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, S.T.; Bassler, B.L. Bacterial quorum sensing: its role in virulence and possibilities for its control. Cold Spring Harb Perspect Med 2012, 2. [Google Scholar] [CrossRef] [PubMed]
- Lade, H.; Paul, D.; Kweon, J.H. N-acyl homoserine lactone-mediated quorum sensing with special reference to use of quorum quenching bacteria in membrane biofouling control. Biomed Res Int 2014, 2014, 162584. [Google Scholar] [CrossRef]
- Shibata, S.; Yip, E.S.; Quirke, K.P.; Ondrey, J.M.; Visick, K.L. Roles of the structural symbiosis polysaccharide (syp) genes in host colonization, biofilm formation, and polysaccharide biosynthesis in Vibrio fischeri. J Bacteriol 2012, 194, 6736–6747. [Google Scholar] [CrossRef]
- Yip, E.S.; Geszvain, K.; DeLoney-Marino, C.R.; Visick, K.L. The symbiosis regulator RscS controls the syp gene locus, biofilm formation and symbiotic aggregation by Vibrio fischeri. Mol Microbiol 2006, 62, 1586–1600. [Google Scholar] [CrossRef]
- Browne-Silva, J.; Nishiguchi, M.K. Gene sequences of the pil operon reveal relationships between symbiotic strains of Vibrio fischeri. Int J Syst Evol Microbiol 2008, 58, 1292–1299. [Google Scholar] [CrossRef]
- Darnell, C.L.; Hussa, E.A.; Visick, K.L. The putative hybrid sensor kinase sypf coordinates biofilm formation in Vibrio fischeri by acting upstream of two response regulators, SypG and VpsR. J Bacteriol 2008, 190, 4941–4950. [Google Scholar] [CrossRef]
- Millikan, D.S.; Ruby, E.G. Vibrio fischeri flagellin a is essential for normal motility and for symbiotic competence during initial squid light organ colonization. J Bacteriol 2004, 186, 4315–4325. [Google Scholar] [CrossRef]
- Colin, R.; Ni, B.; Laganenka, L.; Sourjik, V. Multiple functions of flagellar motility and chemotaxis in bacterial physiology. FEMS Microbiol Rev 2021, 45. [Google Scholar] [CrossRef]
- Ariyakumar, D.S.; Nishiguchi, M.K. Characterization of two host-specific genes, mannose-sensitive hemagglutinin (MshA) and Uridyl phosphate dehydrogenase (UDPDH) that are involved in the Vibrio Fischeri-Euprymna tasmanica mutualism. FEMS Microbiol Lett 2009, 299, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Nishiguchi, M.K.; Hirsch, A.M.; Devinney, R.; Vedantam, G.; Riley, M.A.; Mansky, L.M. Deciphering evolutionary mechanisms between mutualistic and pathogenic symbioses. Vie et Milieu 2008, 58, 87–106. [Google Scholar] [PubMed]
- Bellissimo, K.A.; Septer, A.N.; Whistler, C.A.; Rodríguez, C.; Stabb, E. V. Deletion of luxI increases luminescence of Vibrio fischeri. mBio 2024, 15. [Google Scholar] [CrossRef] [PubMed]
- Plate, L.; Marletta, M.A. Nitric oxide modulates bacterial biofilm formation through a multicomponent cyclic-di-GMP signaling network. Mol Cell 2012, 46, 449–460. [Google Scholar] [CrossRef]
- Silva, A.J.; Benitez, J.A. Vibrio cholerae biofilms and cholera pathogenesis. PLoS Negl Trop Dis 2016, 10, e0004330. [Google Scholar] [CrossRef]
- He, H.; Cooper, J.N.; Mishra, A.; Raskin, D.M. Stringent response regulation of biofilm formation in Vibrio cholerae. J Bacteriol 2012, 194, 2962–2972. [Google Scholar] [CrossRef]
- Lutz, C.; Erken, M.; Noorian, P.; Sun, S.; McDougald, D. Environmental reservoirs and mechanisms of persistence of Vibrio cholerae. Front Microbiol 2013, 4. [Google Scholar] [CrossRef]
- Conner, J.G.; Teschler, J.K.; Jones, C.J.; Yildiz, F.H. Staying alive: Vibrio cholerae ’s cycle of environmental survival, transmission, and dissemination. Microbiol Spectr 2016, 4. [Google Scholar] [CrossRef]
- Leighton, R.E.; Xiong, L.; Anderson, G.K.; Astarita, G.M.; Cai, G.; Norman, R.S.; Decho, A.W. Vibrio parahaemolyticus and Vibrio vulnificus in vitro biofilm dispersal from microplastics influenced by simulated human environment. Front Microbiol 2023, 14. [Google Scholar] [CrossRef]
- Chen, T.; Pu, M.; Subramanian, S.; Kearns, D.; Rowe-Magnus, D. PlzD Modifies Vibrio vulnificus foraging behavior and virulence in response to elevated c-di-GMP. mBio 2023, 14. [Google Scholar] [CrossRef]
- Lydon, K.A.; Kinsey, T.; Le, C.; Gulig, P.A.; Jones, J.L. Biochemical and virulence characterization of Vibrio vulnificus isolates from clinical and environmental sources. Front Cell Infect Microbiol 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Haygood, M.G.; Distel, D.L. Bioluminescent symbionts of flashlight fishes and deep-sea anglerfishes form unique lineages related to the genus Vibrio. Nature 1993, 363, 154–156. [Google Scholar] [CrossRef] [PubMed]
- Tanet, L.; Tamburini, C.; Baumas, C.; Garel, M.; Simon, G.; Casalot, L. Bacterial bioluminescence: light emission in Photobacterium phosphoreum is not under quorum-sensing control. Front Microbiol 2019, 10. [Google Scholar] [CrossRef]
- Kitts, G.; Rogers, A.; Teschler, J.K.; Park, J.H.; Trebino, M.A.; Chaudry, I.; Erill, I.; Yildiz, F.H. The Rvv two-component regulatory system regulates biofilm formation and colonization in Vibrio cholerae. PLoS Pathog 2023, 19, e1011415. [Google Scholar] [CrossRef] [PubMed]
- Marsden, A.E.; Grudzinski, K.; Ondrey, J.M.; DeLoney-Marino, C.R.; Visick, K.L. Impact of salt and nutrient content on biofilm formation by Vibrio fischeri. PLoS One 2017, 12, e0169521. [Google Scholar] [CrossRef]
- Ramos-Vivas, J.; Acosta, F. Editorial: Host-bacteria interactions in fish pathogens. Front Cell Infect Microbiol 2024, 14. [Google Scholar] [CrossRef]
- Mass, S.; Cohen, H.; Podicheti, R.; Rusch, D.B.; Gerlic, M.; Ushijima, B.; van Kessel, J.C.; Bosis, E.; Salomon, D. The coral pathogen Vibrio coralliilyticus uses a T6SS to secrete a group of novel anti-eukaryotic effectors that contribute to virulence. PLoS Biol 2024, 22, e3002734. [Google Scholar] [CrossRef]
- Zhao, W.; Chen, X.; Liu, R.; Tian, P.; Niu, W.; Zhang, X.-H.; Liu, J.; Wang, X. Distinct coral environments shape the dynamic of planktonic Vibrio spp. Environ Microbiome 2023, 18, 77. [Google Scholar] [CrossRef]
- Mincer, T.J.; Zettler, E.R.; Amaral-Zettler, L.A. Biofilms on plastic debris and their influence on marine nutrient cycling, productivity, and hazardous chemical mobility. In Hazardous chemicals associated with plastics in the marine environment; Springer: Cham; vol 78, pp. 221–233.
- Zhang, X.; Lin, H.; Wang, X.; Austin, B. Significance of Vibrio species in the marine organic carbon cycle—a review. Sci China Earth Sci 2018, 61, 1357–1368. [Google Scholar] [CrossRef]
- Alotaibi, G.F. Factors influencing bacterial biofilm formation and development. Am J Biomed Sci Res 2021, 12, 617–626. [Google Scholar] [CrossRef]
- Filippini, G.; Bugnot, A.B.; Varkey, D.R.; Siboni, N.; Ferguson, A.; Gribben, P.E.; Erickson, K.; Palmer, J.; Dafforn, K.A. Nitrogen-cycling genes in oyster reefs and surrounding sediments: relationships with environmental factors and respective nitrogen rates. Mar Pollut Bull 2023, 197, 115710. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.E.; O’toole, G.A. Microbial biofilms: from ecology to molecular genetics. Microbiol Mol Biol Rev 2000, 64, 847–867. [Google Scholar] [CrossRef] [PubMed]
- McCaughey, C.S.; Trebino, M.A.; McAtamney, A.; Isenberg, R.Y.; Mandel, M.J.; Yildiz, F.H.; Sanchez, L.M. A Label-free approach for relative spatial quantitation of c-di-GMP in microbial biofilms. Anal Chem 2024, 96, 8308–8316. [Google Scholar] [CrossRef] [PubMed]
- Teschler, J.K.; Nadell, C.D.; Drescher, K.; Yildiz, F.H. Mechanisms underlying Vibrio cholerae biofilm formation and dispersion. Annu Rev Microbiol 2022, 76, 503–532. [Google Scholar] [CrossRef]
- Shrestha, P.; Razvi, A.; Fung, B.L.; Eichinger, S.J.; Visick, K.L. Mutational analysis of Vibrio fischeri c-di-GMP-modulating genes reveals complex regulation of motility. J Bacteriol 2022, 204. [Google Scholar] [CrossRef]
- Ragupathi, H.; Pushparaj, M.M.; Gopi, S.M.; Govindarajan, D.K.; Kandaswamy, K. Biofilm matrix: a multifaceted layer of biomolecules and a defensive barrier against antimicrobials. Arch Microbiol 2024, 206, 432. [Google Scholar] [CrossRef]
- Almatroudi, A. Biofilm resilience: molecular mechanisms driving antibiotic resistance in clinical contexts. Biology (Basel) 2025, 14, 165. [Google Scholar] [CrossRef]
- Roilides, E.; Simitsopoulou, M.; Katragkou, A.; Walsh, T.J. How biofilms evade host defenses. Microbiol Spectr 2015, 3. [Google Scholar] [CrossRef]
- Hitzler, S.U.J.; Fernández-Fernández, C.; Montaño, D.E.; Dietschmann, A.; Gresnigt, M.S. Microbial adaptive pathogenicity strategies to the host inflammatory environment. FEMS Microbiol Rev 2025, 49. [Google Scholar] [CrossRef]
- Tanveer, M.; Ntakiyisumba, E.; Won, G. Revealing antimicrobial resistance profile and associated factors of Vibrio vulnificus isolated from clinical, environmental, and seafood samples across Asia: a systematic review and meta-analysis. Heliyon 2024, 10, e40334. [Google Scholar] [CrossRef]
- Azeem, K.; Fatima, S.; Ali, A.; Ubaid, A.; Husain, F.M.; Abid, M. Biochemistry of bacterial biofilm: insights into antibiotic resistance mechanisms and therapeutic intervention. Life 2025, 15, 49. [Google Scholar] [CrossRef] [PubMed]
- Mandel, M.J.; Dunn, A.K. Impact and influence of the natural Vibrio-squid symbiosis in understanding bacterial–animal interactions. Front Microbiol 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Muthulakshmi, T.; Mothadaka, M.P. Vibrio vulnificus and its antimicrobial resistance. In Handbook on antimicrobial resistance. In Handbook on antimicrobial resistance; Springer Nature Singapore: Singapore, 2023; pp. 1–18. [Google Scholar]
- Yang, Y.; Yan, J.; Olson, R.; Jiang, X. 2025. [CrossRef]
- Pang, R.; Xie, T.; Wu, Q.; Li, Y.; Lei, T.; Zhang, J.; Ding, Y.; Wang, J.; Xue, L.; Chen, M.; et al. Comparative genomic analysis reveals the potential risk of Vibrio parahaemolyticus isolated from ready-to-eat foods in china. Front Microbiol 2019, 10, 186. [Google Scholar] [CrossRef] [PubMed]
- Steenackers, H.P.; Parijs, I.; Foster, K.R.; Vanderleyden, J. Experimental evolution in biofilm populations. FEMS Microbiol Rev 2016, 40, 373–397. [Google Scholar] [CrossRef]
- Norsworthy, A.N.; Visick, K.L. Gimme Shelter: How Vibrio fischeri successfully navigates an animal’s multiple environments. Front Microbiol 2013, 4. [Google Scholar] [CrossRef]
- Kuper, T.J.; Islam, M.M.; Peirce-Cottler, S.M.; Papin, J.A.; Ford, R.M. Spatial transcriptome-guided multi-scale framework connects Psuedomonas aeruginosa metabolic states to oxidative stress biofilm microenvironment. PLoS Comput Biol 2024, 20, e1012031. [Google Scholar] [CrossRef]
- Lee, H.H.; Ostrov, N.; Wong, B.G.; Gold, M.A.; Khalil, A.S.; Church, G.M. Functional genomics of the rapidly replicating bacterium Vibrio natriegens by CRISPRi. Nat Microbiol 2019, 4, 1105–1113. [Google Scholar] [CrossRef]
- Pipes, B.L.; Nishiguchi, M.K. Nocturnal acidification: a coordinating cue in the Euprymna scolopes–Vibrio fischeri symbiosis. Int J Mol Sci 2022, 23, 3743. [Google Scholar] [CrossRef]
- Sampaio, A.; Silva, V.; Poeta, P.; Aonofriesei, F. Vibrio spp.: life strategies, ecology, and risks in a changing environment. Diversity (Basel) 2022, 14, 97. [Google Scholar] [CrossRef]
- Vezzulli, L.; Pezzati, E.; Brettar, I.; Höfle, M.; Pruzzo, C. Effects of global warming on Vibrio ecology. Microbiol Spectr 2015, 3. [Google Scholar] [CrossRef]
- Sauer, K. Cyclic di-GMP and the regulation of biofilm dispersion. In Microbial Cyclic Di-Nucleotide Signaling; Springer International Publishing: Cham, 2020; pp. 545–560. [Google Scholar]
- Hussa, E.A.; Darnell, C.L.; Visick, K.L. RscS functions upstream of SypG to control the syp locus and biofilm formation in Vibrio fischeri. J Bacteriol 2008, 190, 4576–4583. [Google Scholar] [CrossRef] [PubMed]
- Griend, J.A. Vander; Isenberg, R.Y.; Kotla, K.R.; Mandel, M.J. Transcriptional pathways across colony biofilm models in the symbiont. Vibrio fischeri, 2023. [Google Scholar]
- Youngblom, M.A.; Smith, T.M.; Murray, H.J.; Pepperell, C.S. Adaptation of the mycobacterium tuberculosis transcriptome to biofilm growth. PLoS Pathog 2024, 20, e1012124. [Google Scholar] [CrossRef] [PubMed]
- León, M.; Kokkari, C.; García, K.; Castillo, D.; Katharios, P.; Bastías, R. Diversification of Vibrio anguillarum driven by the bacteriophage CHOED. Front Microbiol 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.; Hölscher, T.; Dragoš, A.; Cooper, V.S.; Kovács, Á.T. Laboratory evolution of microbial interactions in bacterial biofilms. J Bacteriol 2016, 198, 2564–2571. [Google Scholar] [CrossRef]
- Barraud, N.; Kjelleberg, S.; Rice, S.A. Dispersal from microbial biofilms. Microbiol Spectr 2015, 3. [Google Scholar] [CrossRef]
- Edel, M.; Horn, H.; Gescher, J. Biofilm systems as tools in biotechnological production. Appl Microbiol Biotechnol 2019, 103, 5095–5103. [Google Scholar] [CrossRef]
- de Oliveira, N.S.; da Silva Ramos, R.C.P.; de Paula, R.C.; da Costa Pereira, M.G.; Rosa, R.T.; Bianchini, L.F.; Rosa, E.A.R. Advantages of using biofilms to obtain high-value molecules by microbial biotransformations. Explor Drug Sci 2025. [Google Scholar] [CrossRef]
- Koo, H.; Allan, R.N.; Howlin, R.P.; Stoodley, P.; Hall-Stoodley, L. Targeting microbial biofilms: current and prospective therapeutic strategies. Nat Rev Microbiol 2017, 15, 740–755. [Google Scholar] [CrossRef]
- Ruby, E.G.; Lee, K.-H. The Vibrio fischeri-Euprymna scolopes light organ association: current ecological paradigms. Appl Environ Microbiol 1998, 64, 805–812. [Google Scholar] [CrossRef]
- Prentice, J.A.; Bridges, A.A.; Bassler, B.L. Synergy between c-di-GMP and quorum-sensing signaling in Vibrio cholerae biofilm morphogenesis. J Bacteriol 2022, 204. [Google Scholar] [CrossRef]
- Kilic, T. Factors affecting biofilm formation and the effects of these factors on bacteria. In Exploring Bacterial Biofilms; IntechOpen, 2025. [Google Scholar]
- Naga, N.G.; El-Badan, D.E.; Ghanem, K.M.; Shaaban, M.I. It is the time for quorum sensing inhibition as alternative strategy of antimicrobial therapy. Cell Communication and Signaling 2023, 21, 133. [Google Scholar] [CrossRef] [PubMed]
- Yun, M.S.; Sun, J.; Lovejoy, C.; Lee, S.H. Editorial: Microbial response to a rapidly changing marine environment: global warming and ocean acidification. Front Microbiol 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Portas, A.; Carriot, N.; Barry-Martinet, R.; Ortalo-Magné, A.; Hajjoul, H.; Dormoy, B.; Culioli, G.; Quillien, N.; Briand, J.-. François. Shear stress controls prokaryotic and eukaryotic biofilm communities together with eps and metabolomic expression in a semi-controlled coastal environment in the NW Mediterranean sea. Environ Microbiome 2024, 19, 109. [Google Scholar] [CrossRef]
- Chan, W.Y.; Rudd, D.; van Oppen, M.J. Spatial metabolomics for symbiotic marine invertebrates. Life Sci Alliance 2023, 6, e202301900. [Google Scholar] [CrossRef] [PubMed]
- Yeor-Davidi, E.; Zverzhinetsky, M.; Krivitsky, V.; Patolsky, F. Real-time monitoring of bacterial biofilms metabolic activity by a redox-reactive nanosensors array. J Nanobiotechnology 2020, 18, 81. [Google Scholar] [CrossRef]
- Yan, X.; Liao, H.; Wang, C.; Huang, C.; Zhang, W.; Guo, C.; Pu, Y. An improved bacterial single-cell RNA-seq reveals biofilm heterogeneity. Elife 2024, 13. [Google Scholar] [CrossRef]
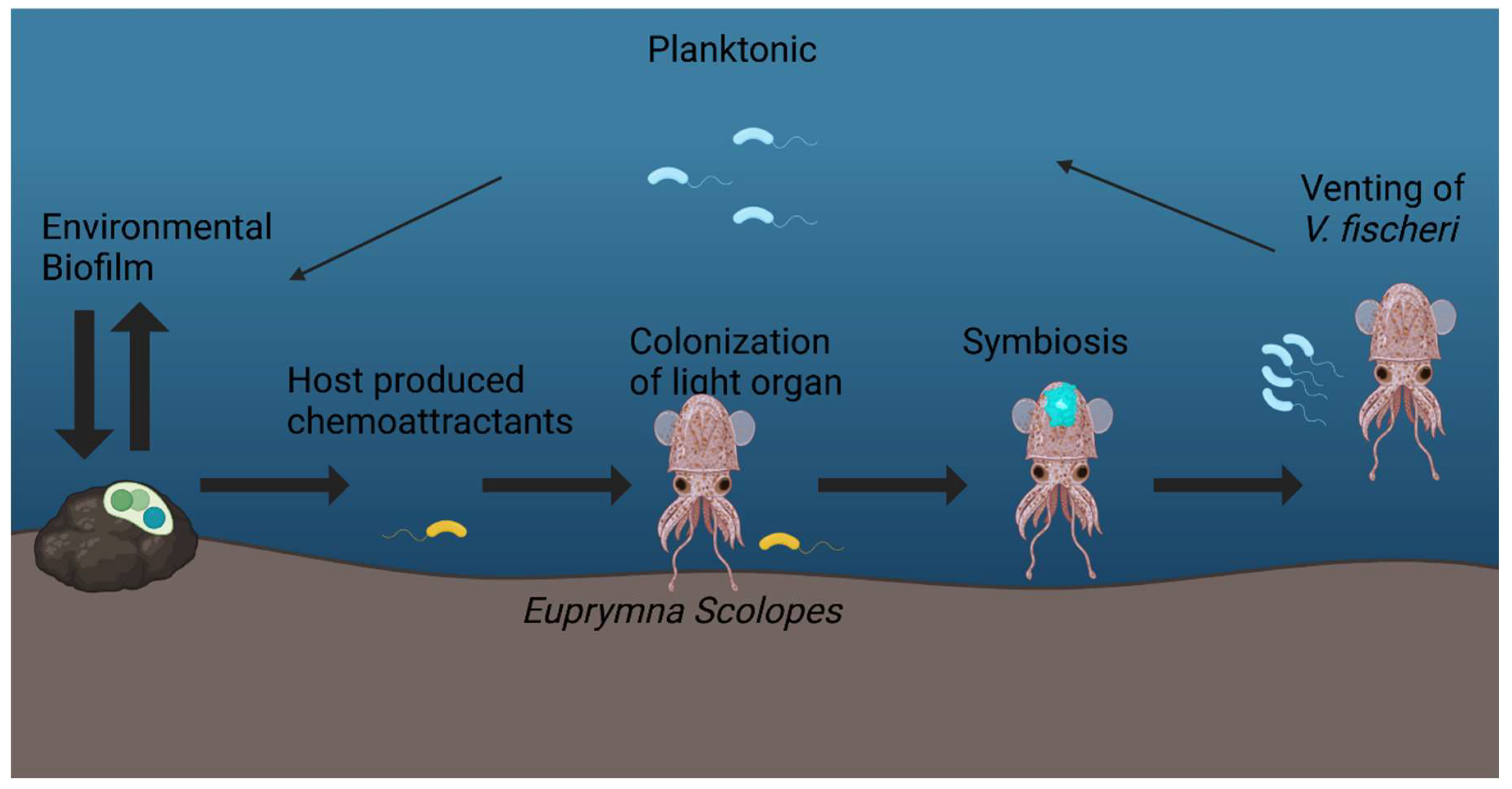
Figure 1.
Transition between environmental and squid host-associated Vibrio fischeri. V. fischeri exists in both environmental biofilms and a planktonic (free-living) state. Host-produced chemo-attractants recruit planktonic V. fischeri to the squid’s light organ where a series of specific biochemical filters only allow symbiotic competent V. fischeri to colonize. After colonization, V. fischeri can then establish a dense community or biofilm, allowing the production of bioluminescence through quorum sensing. Once bioluminescence is no longer needed for counterillumination and with the onset of daylight, the squid vents 95% of its symbiont population at dawn, releasing competent V. fischeri back into the environment, continuing the cycle between free-living and host associated life histories.
Figure 1.
Transition between environmental and squid host-associated Vibrio fischeri. V. fischeri exists in both environmental biofilms and a planktonic (free-living) state. Host-produced chemo-attractants recruit planktonic V. fischeri to the squid’s light organ where a series of specific biochemical filters only allow symbiotic competent V. fischeri to colonize. After colonization, V. fischeri can then establish a dense community or biofilm, allowing the production of bioluminescence through quorum sensing. Once bioluminescence is no longer needed for counterillumination and with the onset of daylight, the squid vents 95% of its symbiont population at dawn, releasing competent V. fischeri back into the environment, continuing the cycle between free-living and host associated life histories.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



