
Submitted:
22 April 2025
Posted:
22 April 2025
You are already at the latest version
Abstract
Purinergic P2X receptors (P2X) are ATP-gated ion channels that are broadly expressed in the brain, particularly in the hypothalamus. As ionic channels with high permeability to calcium, P2X play an important and active role in neural functions. The hypothalamus contains a number of small nuclei with many molecularly defined types of peptidergic neurons that affect a wide range of physiological functions, including water balance, blood pressure, metabolism, food intake, circadian rhythm, childbirth and breastfeeding, growth, stress, body temperature and multiple behaviors. This review focuses on the possible role of P2X and extracellular ATP in specific hypothalamic functions and the cellular expression of P2X in the most important hypothalamic nuclei, such as the supraoptic nucleus (SON), paraventricular nucleus (PVN), suprachiasmatic nucleus (SCN), anteroventral periventricular nucleus (AVPV), arcuate nucleus (ARC), ventromedial hypothalamic nucleus (VMH), dorsomedial hypothalamic nucleus (DMH), tuberomammillary nucleus (TMN) and lateral hypothalamic area (LHA).
Keywords:
hypothalamus
; supraoptic nucleus (SON)
; paraventricular nucleus (PVN)
; arcuare nucleus (ARC)
; suprachiasmatic nucleus (SCN)
; P2X
; extracellular ATP
1. Introduction
Hypothalamus is the part of the brain that is functionally connected to the limbic system and pituitary gland. This relatively small area of the brain contains a number of small nuclei, which play a key role in coordinating fundamental body functions. Particularly, hypothalamus has important functions in maintaining the homeostasis of body fluids, regulating blood flow, lactation, parturition, body temperature, appetite, systemic metabolism, growth, sleep and waking cycle, and social behavior [1,2]. Anatomically, the hypothalamus can be arranged into three main regions: the anterior (Figure 1), middle and posterior hypothalamus. Each major region contains nuclei that serve certain physiological functions. The anterior region contains at least five important nuclei: supraoptic (SON), paraventricular (PVN), suprachiasmatic (SCN), anteroventral periventricular nucleus (AVPV) and anterior hypothalamic nucleus (AHN). The middle region contains three nuclei: the arcuate nucleus (ARC), the ventromedial hypothalamic (VMH) and dorsomedial hypothalamic (DMH) nucleus. The posterior region contains the tuberomammillary nucleus (TMN) and the lateral hypothalamic region (LHA), which serves for the integration of autonomic and limbic information. Hypothalamus contains many molecularly defined neuronal populations that synthesize neuropeptides alongside classical neurotransmitters such as γ-aminobutyric acid (GABA) and glutamate. Molecular characterization of neuronal subtypes is presented in an integrated reference atlas, “HypoMap“, which provides a comprehensive single-cell transcriptomic atlas of the mouse hypothalamus and reveals transcriptomic diversity of cell types in different hypothalamic nuclei [3].
The hypothalamus is known to abundantly express purinergic P2X receptors (P2X) activated by extracellular adenosine-5'-triphosphate (ATP). Hypothalamic neurons express several subtypes of P2X [4], which are also expressed in hypothalamic astrocytes [5,6], tanycytes [7,8] and microvessels [9]. Although purinergic signaling has a well-established role in the regulation of energy homeostasis and feeding behavior [10], and extracellular ATP is known to be involved in regulating the sleep-wake cycle [11], the role of P2X in hypothalamic functions is not yet fully understood. Several reviews described the expression and function of P2X and purinergic signaling pathways in the hypothalamic-neurohypophysial system [5,12], the hypothalamic-adenohypophysial system [13,14,15], endocrine glands [16,17] and circadian system [18,19,20]. This review focuses on the distribution of P2X in the most important hypothalamic nuclei, and the possible involvement of extracellular ATP and P2X in endocrine and non-endocrine hypothalamic functions.
2. Nuclei and Neuron Types in the Hypothalamus
2.1. Supraoptic Nucleus (SON)
Supraoptic nucleus (SON) is located at the bottom of the anterior hypothalamus (Figure 1), adjacent to the chiasma opticum and in the vicinity of hypothalamic blood vessels. Its shape and size depend on the species, it can be oval shaped or elongated, and in humans it stretches through most of the ventral side of the hypothalamus [21]. SON contains two types of magnocellular neurons that synthesize and secrete two hormones: oxytocin and vasopressin, also called arginin-vasopressin (AVP). The axons of magnocellular neurons transport oxytocin and vasopressin stored in large dense-cored vesicles from the soma to the posterior pituitary, where both hormones are secreted into the systemic circulation. The main function of oxytocin and vasopressin is to regulate childbirth and breastfeeding, and to maintain the homeostasis of body fluid. Oxytocin stimulates the contraction of myoepithelial cells in the lactating mammary gland and triggers uterine contractions during labor, the release of oxytocin is mainly caused by parturition and suction reflex [22,23,24,25]. Vasopressin serves to regulate plasma osmolality and blood pressure, it acts on the kidney to increase water reabsorption and causes vasoconstriction. The release of vasopressin increases as a function of plasma osmolality and water status of the body [26,27,28]. Furthermore, oxytocin and vasopressin are also released via dendritic release [29,30] into the blood vessels inside the brain [21]. Through the actions at the level of the CNS, via their specific receptors on neurons, oxytocin and vasopressin are independently involved in facilitating of maternal and parental care, and other social behaviors [31,32,33,34]. The region that expresses the oxytocin receptor and controls maternal behavior, for example, is the medial preoptic area (MPOA). Oxytocin and vasopressin are also released within the SON itself, where they have local regulatory effects [35,36,37].
The secretion of oxytocin and vasopressin is dependent on the electrical activity of magnocellular neurons [38] which has an in vivo specific pattern for oxytocin and AVP neurons [22,23,25,27,28,39]. Both hormones are released in response to the activity-dependent Ca2+ influx, which primes large dense-cored vesicles for exocytosis [28,40,41,42,43]. Magnocellular neuron activity is controlled by various excitatory and inhibitory synaptic inputs [27,44,45,46]. These inputs use glutamate [47,48,49,50] and γ-aminobutyric acid (GABA) [24] as the main neurotransmitter. Other inputs use norepinephrine, serotonin, dopamine, histamine, acetylcholine, angiotensin II, nitric oxide, β endorphin and ATP (for review see [51]).
2.2. Paraventricular Nucleus (PVN)
Paraventricular nucleus (PVN) is located on the sides of the third ventricle of the hypothalamus, at the top of the ventricle (Figure 1). It is one of the most important neuroendocrine and autonomic regulatory brain center, with neurons playing an essential role in controlling stress, metabolism, growth, reproduction, immune, gastrointestinal, renal and cardiovascular functions. The PVN receives afferent inputs from many regions which are then reflected in changes in single specific outputs [52]. By function, the PVN contains three types of peptidergic neurons, predominantly neuroendocrine: magnocellular, parvocellular and preautonomic (for review see [53]. Anatomically, the PVN of the rat consists of three parts: the dorsal and medial parts contain mainly parvocellular neurons, and the posterior part contains magnocellular neurons [54,55]. Firstly, magnocellular neurons have large cell bodies due to their necessary large capacity for peptide synthesis and secretion, while parvocellular neurons are small, reflecting less need for production of less stored peptide [1]. Further, magnocellular PVN neurons project to the posterior pituitary, where they release oxytocin and vasopressin directly into the general circulation. These neurons receive similar afferent inputs as magnocellular oxytocin and vasopressin neurons in the SON [51]. Secondly, parvocellular neurosecretory cells synthesize hypophysiotropic hormones and project to the median eminence. These hormones are secreted into the hypothalamic-hypophysial portal system, and transported vascularly to the anterior pituitary gland, where they cause the immediate release of pituitary hormones. Parvocellular neurons produce and secrete corticotropin-releasing hormone (CRH), which regulates secretion of adrenocorticotropic hormone (ACTH); thyrotropin-releasing hormone (TRH), which controls secretion of thyroid-stimulating hormone (TSH); somatostatin and growth hormone-releasing hormone (GHRH), which both control secretion of growth hormone (GH). The majority of these neurons lie in the medial parts of PVN. In addition, parvocellular preautonomic neurons project to autonomic centers in the brain stem and spinal cord [54,56,57].
Each of these PVN neuron types, especially CRH, oxytocin, vasopressin, somatostatin and TRH neurons, are also centrally projecting neurons releasing peptides in hippocampus, locus coeruleus, lateral septum and nucleus accumbens, for example [1] and affecting many types of behavior in mice and humans [31,32]. For example, CRH plays a critical role in the brain's response to stress, emotional regulation and other physiological processes associated with sleep, learning, memory, cognition, food intake, and motor co-ordination. Dysregulation of the CRH system has been associated with neurological and psychiatric disorders including anxiety and depression [53,58]. Next, oxytocin neurons in the PVN are involved in the regulation of empathic [59,60] and feeding behavior [61]. Retrograde tracer analysis has shown that projection of vasopressin neurons from SCN to PVN links light perception to feeding behavior, so that light exposure acutely suppresses food intake and increases c-Fos expression in the oxytocin neurons of PVN; the light-induced suppression of food intake was mostly abolished by blockade of the oxytocin receptor [61]. Last, TRH plays a role in central control of food intake so that central administration of TRH may produce anorexigenic or orexigenic effects, depending on the injection site of TRH [62].
A group of preautonomous parvocellular neurons that do not belong to the endocrine neurons and are involved in the central control of the autonomic nervous system include neurons releasing angiotensin II (AngII) and nitric oxide, for overview see [63].
2.3. Suprachiasmatic Nucleus (SCN)
Suprachiasmatic nucleus (SCN) is located directly above the crossing of optic nerves, on both sides of the third ventricle (Figure 1). It represents a heterogeneous structure of ~10,000 GABA producing neurons accompanied by a large number of glial cells [64]. The SCN is a principal internal time-keeping system that synchronizes the behavior and physiology of an entire organism with a 24-hour cycle [65,66,67], and helps to establish the normal sleep-wake cycle by influencing the synthesis and release of melatonin from the pineal gland. Endogenous circadian rhythms are driven by the expression of clock genes in each SCN neuron [68]. It regulates the circadian rhythms of various physiological processes, including motor activity, body temperature, plasma hormone levels, renal secretion and metabolism
The SCN contains two types of nerons, and anatomically it can be divided into dorsomedial (shell) and ventrolateral (core) parts [69]. The light-responsive neurons producing vasoactive intestinal peptide (VIP) are found in ventrolateral part of SCN, while the endogenously rhythmic neurons producing AVP are found in the dorsomedial region of this nucleus, which does not receive a direct input from the retina [70,71,72]. Intercellular communication between endogenously rhythmic neurons and light-responsive neuron is likely responsible for the unique pacemaker properties of the SCN observed at a tissue and whole body level [69]. Finally, the circadian rhythm of AVP secretion runs parallel to the rhythm of electrical activity in SCN neurons [73,74,75]. Using AVP secretin, the SCN interacts with other regions of the brain to synchronize the circadian rhythms of various physiological processes. For example, retrograde tracer analysis has shown that the projection of AVP neurons from the SCN to PVN links light reception to feeding behavior [61].
2.4. Anteroventral Periventricular Nucleus (AVPV)
Anteroventral periventricular nucleus (AVPV) belongs to the preoptic region (POA), a broader area encompassing several nuclei, including periventricular nucleus (PeN), median preoptic nucleus (MnPO) and medial preoptic area (MPOA). The AVPV, specifically located in the hypothalamus (Figure 1), is a small cluster of cells that is characterized by its sexually dimorphic structure, being larger in females than in males. The AVPV is particularly important as the region containing neurons producing gonadotropin-releasing hormone (GnRH, also called luteinizing hormone-releasing hormone, LHRH) and neurons producing kisspeptin that are both involved in the regulation of fertility and production of gonadal steroid hormones [76].
Firstly, GnRH neurons regulate reproductive function in mammals by inducing synthesis and release of follicle stimulating hormone (FSH) and luteinizing hormone (LH) from the anterior pituitary gland. They are extremely rare, elongated, bipolar or unipolar, and in rodents, the cell bodies of GnRH neurons are diffusely distributed throughout the basal forebrain. The loosely organized population of GnRH cells receives neuronal inputs from many (> 50) different brain regions [77]. One or two very long dendritic processes of GnRH neurons are located primarily in the region around the organum vasculosum of the lamina terminalis (OVLT), and their nerve terminals are mainly found in the median eminence [78]. In the AVPV, GnRH is released in pulses at hourly intervals, and this pulsatility is essential for the maintenance of the reproductive function of adult animals [79]. In additin, GnRH is also released in various regions of the CNS and affects neurological disorders, neuroprotection, cognitive functions and mood regulation, [80].
Secondly, kisspeptin neurons are located near GnRH neurons and their dendrons, produce neuropeptide kisspeptin which activates the hypothalamic-pituitary-gonadal axis and influences neural circuits controlling sexual behavior [81]. Kisspeptin acts through specific kisspeptin receptors (Kiss1R, also known as GPR54) expressed by GnRH neurons [82] to induce the synthesis and release of GnRH, and consequently LH surge and ovulation [83]. Furthermore, kisspeptin neurons express estrogen receptors that play a well-established role in the estrogen-dependent regulation of reproduction [76]. AVPV kisspeptin neurons are primarily involved in the estrogen-induced positive feedback mechanism, which triggers the preovulatory luteinizing hormone (LH) surge in females [76]. Fore example, Kiss1 (kisspeptin gene)- and Gpr54 (kisspeptin receptor gene)-deficient mice, rats, and humans are infertile and suffering for hypogonadotropic hypogonadism [83]. AVPV kisspeptin neurons project to neuronal nitric oxide synthase (nNOS) producing neurons in the ventrolateral part of the ventromedial hypothalamus (VMH) to stimulate lordosis behavior. Linking of kisspeptins to metabolism and reproduction has been recently reviewed at [84]. In addition, kisspeptin is also produced by kisspeptin neurons in the ARC (see next chapter 2.5.).
2.5. Anterior Hypothalamic Nucleus (AHN)
Anterior hypothalamus (AHN) is a specific nucleus that controls body temperature. About 30% of AHN neurons are warm-sensitive, many of them receiving inputs from afferent pathways that communicate information from skin thermoreceptors [85].
2.6. Arcuate Nucleus (ARC)
Arcuate nucleus (ARC), located in the middle hypothalamic region, contains diverse populations of neurons that participate in the control of food intake, body weight [86,87], energy homeostasis [88], feeding behavior, blood pressure and cardiovascular function, particularly in the context of obesity and metabolic disorders (for review, see [89]). Neurons in the arcuate nucleus synthesize the neuropeptide Y (NPY), agouti-related peptide (AgRP), proopiomelanocortin (POMC), cocaine and amphetamine-regulated transcript (CART) and the α-melanocyte-stimulating hormone (α-MSH) according to the nutritional status of the organism. Their nerve endings in the PVN activate or inhibit neurons synthesizing TRH to control food intake [1]. ARC neurons also control the endocrine functions of the anterior pituitary gland by producing both releasing and inhibiting neurohormones secreted in the median eminence. These include growth hormone-releasing hormone (GHRH) and somatostatin, which regulate the hypothalamic-pituitary-growth hormone axis, and prolactin-inhibiting hormone (dopamine), which regulates the hypothalamic-pituitary-prolactin axis.
Arcuate nucleus contains GnRH neurons producing GnRH, which regulates hypothalamic-pituitary-gonadal axes in a sex-specific manner [1], and in rodents kisspeptin neurons producing kisspeptin [90]. ARC kisspeptin neurons mediate negative feedback regulation of GnRH and LH secretion by estrogen, and they are crucial for generating pulsatile GnRH/LH release in female mammals [91]. Simultaneously, kisspeptin neurons coexpress neurokinin B (NKB) and dynorphin A (DYN), therefore they are termed KNDy neurons that regulate rhythmic GnRH pulses essential for reproductive cyclicity. The pulsatility of GnRH release is markedly reduced in aged animals, suggesting that the malfunctions of ARC kisspeptin neurons may be responsible for reproductive aging and menopause-related disorders [92,93].
2.7. Ventromedial Hypothalamic Nucleus (VMH)
Ventromedial nucleus of the hypothalamus (VMH) is alsolocated in the middle hypothalamic region. It has been implicated in the regulation of a wide range of physiological and behavioral processes including feeding, sexual and maternity behavior, blood calcium homeostasis, circadian neuroendocrine control, hyper-running physical activity and emotional behavior (for review, see [1]). The VMH was considered the brain's center of satiety or fullness [96]. For example, lesions of VMH result in hyperphagia and obesity in a variety of species including humans (for review, see [97]. The majority of VMH neurons express steroidogenic factor 1 (SF-1), an orphan nuclear receptor, which was identified as a key regional marker of VMH neurons in the Hypomap Atlas [3]. Neurons producing SF-1 play a critical role in regulating energy homeostasis, glucose metabolism, and body weight [98,99,100](for reviews see [101]), which highlights SF-1 neurons as potential targets for anti-obesity therapies.
2.8. Dorsomedial Hypothalamic Nucleus (DMH)
Dorsomedial nucleus of the hypothalamus (DMH) belongs to complex neuronal network that integrates incoming nutrient and hormonal signals and gates neuronal output to maintain homeostasis. Among feeding circuits, the DMH is recognized as a key player in the regulation of appetite [102]. Further, the DMH is also involved in control of circadian rhythms, termoregulation, including control of brown adipose tissue thermogenesis, and autonomic functions (for review, see [1]), regulation of neuroendocrine stress responses [103] and the cardiovascular response associated with emotional stress [104]. Stimulation of this nucleus, in animal experiments, produced aggressive behavior that lasts only as long as the stimulus is applied. The DMH receives inputs predominantly from adjacent hypothalamic nuclei, notably the VMH and POA, and in turn projects into PVN, for example [102].
2.9. Tuberomammilary Nucleus (TMN)
Tuberomammillary nucleus (TMN) located in the posterior hypothalamus consists primarily of histaminergic neurons, which are involved in memory, changes in emotion, and heart failure. Activity of histaminergic TMN neurons is closely associated with the sleep-wake cycle. These neurons are tonically activated during wakefulness, just a little activated during slow wave sleep, and not at all activated during rapid-eye-movement (REM) sleep [105]. In addition, the TMN receives inputs from the pre-frontal cortex and the POA, while its efferents reach most parts of the brain [106].
2.10. Lateral Hypothalamic Area (LHA)
The lateral hypothalamic area (LHA) also called the lateral hypothalamus (LH), is primarily located in the lateral hypothalamic region and is considered part of the posterior hypothalamus. It is heterogeneous region, containing many molecularly defined neuronal populations that mediate distinct aspects of physiology, including energy balance and feeding, arousal and sleep, stress and reward, body temperature and autonomic functions [107]. The LHA contains three main types of neurons. Firstly, the dynorphin peptide (DYN) neurons [108] which are involved in the food intake, appetitive motivation and modulation of feeding-related behaviors such as arousal. DYN is also the endogenous ligand for the kappa opioid receptor (KOR), that regulate food intake [109]. Secondly, hypocretin/orexin neurons which project extensively throughout the CNS, influencing multiple brain regions involved in wakefulness [110,111]. LHA-derived orexinergic neurons also project into neuroendocrine centers, which are known to be important for the physiological response to stress [112]. Dysfunction of the orexin/hypocretin neurotransmitter system leads to the sleep disorder, narcolepsy. Thirdly, the LHA is responsive to noxious stimuli [113], and a large population of neurotensin-expressing neurons in the LHA project to brain regions that participate in descending control of pain processing [114].
3. Sources of Extracellular ATP in the Brain
In the brain, ATP is released during normal neuronal activity, but high extracellular ATP levels occur at pathological brain states, such as seizures [115], hypoxia [116] or Alzheimer's disease [117]. Large amounts of ATP are also released from damaged or dead cells. The main source of extracellular ATP in the brain are astrocytes [118,119,120,121,122] which play an important and active role in neural function as a participant in the "tripartite synapse" [123]. High frequency stimulation-induced elevations in extracellular glutamate could trigger a sufficient increase in intracellular astrocytic Ca2+ to stimulate ATP release [124]. ATP was detected in large amounts in dense-core vesicles and in lysosomes of astrocytes [125]. Consequently, ATP is released from astrocytes into the synaptic cleft, where it can act on presynaptic or postsynaptic P2 receptors [126,127,128] to modulate the release of neurotransmitters [129,130,131,132,133,134] or postsynaptic efficacy [135]. ATP is released from astrocytes by the mechanism of Ca2+-regulated exocytosis [136,137,138], or via Ca2+-independent non-vesicular pathway, for example through pore-forming molecules such as pannexin-1 hemichannels [139,140,141,142] or P2X7 channels [143]. Finally, ATP can also be released via astrocyte-selective vesicular nucleotide transporter (VNUT) [144].
4. A Brief Overview of Purinergic P2X1-7 Receptors
Purinergic P2X receptors (P2X1-7) are ligand-gated ion channels activated by extracellular ATP. Each P2X subunit has two transmembrane domains, a long cysteine-riched extracellular ligand-binding domain, and the N-terminal and C-terminal regions located intracellularly [145]. Three P2X subunits form homomeric and/or heteromeric ion channels [146]. P2X sometimes even interact with other ion channels, for example P2X7 interacts with pannexin-1 hemichannel [147], P2X4 with GABA receptor type A (GABAA receptor) [148] and P2X5 with acid-sensing ion channel 3 (ASIC3) signaling pathway [149]. P2X receptors present almost equal permeability to N+ and K+, and substantial calcium permeability [150,151]. When expressed in Xenopus oocytes or human embryonic kidney cells (HEK), the homomeric P2X1-7 subtypes differ in sensitivity to agonists, antagonists and allosteric modulators, ion selectivity and desensitization rate [145,152,153]. Most P2X exhibit sensitivity to the inhibitory effects of suramin and pyridoxalphosphate-6-azophenyl-2', 4'-disulfonic acid (PPADS) [145]. However, many new selective antagonists are currently available for different P2X subtypes (for review see [154,155,156]). Under physiological condition, P2X can be modulated by a wide array of native molecules and allosteric modulators such as protons, trace metals like zinc and copper, phosphoinositides, alcohols, reactive oxygen species [145,154,156] and neurosteroids [157].
Here is a brief overview of functional characteristics and pharmacology of P2X1-7 subtypes:
P2X1 has the highest affinity for ATP, desensitizes more quickly compared to other P2X subtypes, in the millisecond range, and can be activated by synthetic ligands such as α,β-methyleneadenosine 5'-triphosphate (αβmeATP) [145,158]. In addition, homomeric P2X1 belongs to P2X that exhibit the highest relative Ca2+ permeability among P2X [159]. P2X1 are expressed namely in urinary bladder and vas deferens, where they are known to have a role in contraction of smooth muscles, or in neutrophil chemotaxis or thromboinflammation [145,160].
P2X2 exhibits moderate sensitivity to ATP and is considered to be a non-desensitizing receptor channel selectively permeable to cations and large organic cations, such as N-methyl D-glucamine (NMDG+) [145,161]. P2X2 can be inhibited by PPADS and suramin [162], but up to date, no P2X2-specific inhibitor has been found. Studies on the P2rx2r reporter mouse models have shown that P2X2 is widely distributed in the CNS [163,164,165], where it is present in different splice variants including the P2X2b, P2X2c, P2X2d, P2X2e, P2X2f and P2X2g [166]. The presynaptic P2X2 have been shown to underlie an increase in GABA release in a subset of GABAergic interneurons in the spinal cord [167]. Further, experiments with knockout mice have shown that Ca2+ entry through presynaptic P2X2 increases the frequency of spontaneous glutamatergic currents mediated by α-amino-3-hydroxy-5-methylisoxazole propionic acid (AMPA) receptors in GABAergic hippocampal interneurons [168]. Multiple types of P2X can be co-expressed in native neurons and some of them can form heteromultimers. For example, it has been shown that P2X2 and P2X3 form heteromultimers in sensory neurons [169]. In addition, P2X2 are known to have function in inflammatory and neuropathic pain, and sense of taste [145,170]. It has been suggested that stimulating P2X2 in the medial prefrontal cortex could be a potential therapeutic strategy for depressive disorders [171].
P2X3 is a fast desensitizing receptor, almost like P2X1, and its recovery from desensitization can take longer than 15 minutes. It is better activated by αβmeATP or β,γ-methyleneadenosine 5′-triphosphate (βγmeATP) than by ATP [145], and the homomeric P2X3 has the lowest Ca2+ permeability among P2X subtypes [159]. Potential structural reasons for the desensitizing character of this receptor was discovered directly from X-ray structures of the P2X3 receptor [172]. Further, P2X3 is exclusively expressed in primary sensory neurons [173,174], and it is known to form P2X2/P2X3 heteromeric channels [169], which play a crucial role in pain signaling [175,176,177]. In addition, ATP-activated P2X3 is also involved in taste function [170,178] or bladder contraction [179] and play a pivotal role in chronic cough [180].
P2X4 exhibits a moderate rate of desensitization, approximately within a few seconds [145], and high Ca2+ permeability [159]. The first P2X structure solved was the zebrafish P2X4 crystal and its shape was compared to a dolphin [181]. Unlike other P2X, the P2X4 can be located intracellularly, specifically on lysosomal membranes, where its otherwise extracellular part is oriented into the lumen organelles [182,183]. ATP may also enter lysosomes, but the receptors cannot be activated due to inhibition at low pH. Moreover, P2X4 cannot be effectively activated by αβmeATP [184], is relatively resistant to the common P2X antagonists suramin and PPADS [185], and is selectively allosterically potentiated by ivermectin, a broad-spectrum antiparasitic drug of bacterial origin [152,186]. The P2X4 subunits may assemble as functional heterotrimer with P2X6 subunit [187]. In addition, P2X4 is widely distributed throughout the CNS [4,188], and is highly expressed on microglia and macrophages [189]. P2X4 has emerged as a potential target for CNS disorders such as epilepsy, ischemia, chronic pain, anxiety, multiple sclerosis and neurodegenerative diseases, for review, see [190].
P2X5 generates low amplitude non-desensitizing currents [145]. In humans, most hP2X5 variants lack exon 10, the critical region encoding the second transmembrane domain (TM2), thereby rendering the receptor non-functional [191]. The P2X5 is highly expressed in the mouse CNS during embryonic development [192]. Further, in adult mice, it is distributed across various brain regions, including the hypothalamus [193], but its role in the brain is not well understood. For example, activation of P2X5 on satellite cells regulates skeletal muscle differentiation [194]. A study in mice P2X5 KO demonstrated the role of P2X5 in the regulation of bone loss in periodontitis [195].
P2X6 lacks part of a binding site for ATP [196], the homomeric P2X6 is therefore non-functional and is not expressed on the cell surface [197]. However, P2X6 is thought to form functional heteromeric channels with P2X2 [198] and P2X4 [187,199]. P2X2/P2X6 heteromers are expressed in respiratory neurons of the brainstem, and recombinant P2X4/P2X6 is functional in the oocytes (Burnstock 2018).
P2X7 shows very low sensitivity to ATP and no apparent desensitization when stimulated with high doses of agonists [197]. This receptor is more sensitive to 3´-O-(4-benzoyl)benzoyladenosine-5´-triphosphate (BzATP) than to ATP, exhibits high permeability for Ca2+ and is permeable for large molecules such as N-methyl-D-glucamine ion (NMDG+), Yo-pro fluorescent dye or ethidium [145,200,201]. Within the P2X family, the P2X7 has the longest C-terminus, which is responsible for many of the specific properties of this receptor [202,203,204] including cytokine release from the brain's resident immune cells, microglia, primed with bacterial lipopolysaccharide [178]. The P2X7 can be inhibited by low pH, bivalent cations and Brilliant Blue-G (BBG), while being relatively insensitive to PPADS or suramin [197,200]. This is the most extensively investigated P2X subtype for drug development, and there are already many strong and selective, mainly allosteric, antagonists [205]. P2X7 is widely expressed in immune-related cells, various glial cells, Schwan cells, T-cells and macrophages [206] [207]. Its activation in glial cells leads to neuroinflammation, for a review, see [208,209], which could contribute to the development of various neurodegenerative diseases such as Alzheimer’s disease, Parkinson’s disease [210,211,212,213] and depression-related emotional disturbances, reviewed by [214,215].
A study performed using the P2X7 reporter mice showed that the expression of the P2X7 is selectively increased in CA1 pyramidal and dentate granule neurons and microglia in mice that developed a status epilepticus after intra-amygdala injection of kainic acid [216]. After neural tissue damage or following status epilepticus, no upregulation of P2X7 protein in neurons was observed [217]. The P2rx7-EGFP reporter mouse, which expresses enhanced green fluorescence protein (EGFP) immediately downstream of the P2rx7 proximal promoter, revealed that the P2X7 is detectable in the brain of a mouse already in early embryonic development, in E9, subsequently increasing between E13 and E18, and shows specific localization in hypothalamic ependymal cells [218].
5. Expression and Function of P2X in Hypothalamic Nuclei
5.1. SON
5.1.1. Expression of P2X mRNA and Protein in SON
Molecular methods proved mRNA of almost all P2X receptors in the supraoptic nucleus (SON). In situ hybridization analysis of six P2X (P2X1-P2X6) mRNAs showed that SON expresses a strong hybridization signal for three of them, P2X2, P2X4 and P2X6 [4]. Another in situ hybridization study demonstrated P2X3 and P2X4 mRNAs expression in the SON [219]. Real-time reverse-transcription (RT-PCR) analysis confirmed expression of P2X2 mRNA in SON [220]. A quantitative real-time reverse-transcription PCR (qRT-PCR) analysis of seven P2X (P2X1-P2X7) in circular punches of rat SON revealed the presence of P2X2, P2X3, P2X4, P2X6 and P2X7 mRNAs, with P2X3, P2X4 and P2X7 dominating [219]. Another qRT-PCR analysis of P2X1-P2X7 and three metabotrophic P2Y (P2Y1, P2Y2 and P2Y12) in circular punches of rat SON showed significant expressions of three of them, in the order: P2X2 > P2X7 > P2X4, moderate expression of P2X3, P2Y1 and P2Y2, and minimal expression of P2X1, P2X5, P2X6 and P2Y12 [134] (Figure 2A).
High level of P2X2 mRNA expression matched the high level of P2X2 protein in the SON. Ultrastructural studies have shown that P2X2 immunoreactivity-positive neurons and nerve fibres are localized in many hypothalamic nuclei, including those containing magnocellular neurons such as the SON [221,222,223,224]. In the coronal sections of the rat hypothalamus (30 µm thick), the SON neuronal cell bodies were strongly P2X2 immunopositive, but only a few nerve fibres were positive [222]. Strong P2X2 immunoreactivity was also found in synapses of rat hypothalamo-neurohypophysial explant [225] which contains SON and PVN with their intact synapses and axonal projections extending through the median eminence and terminating in the posterior lobe of the pituitary gland [225]. Both, SON and the magnocellular part of PVN, displayed P2X2 labelled somata and numerous neuronal processes-axons and dendrites [225]. Besides, P2X2 immunoreactivity was observed at the axo-dendritic and axo-somatic synapses in this system [225]. Axo-dendritic synapses showed either axon terminals (synaptic buttons) marked with P2X2 and unmarked dendrites or unmarked axon terminals and marked dendrites. Moreover, axo-somatic synapses involved P2X2-positive axons on both P2X2-positive and P2X2-negative soma [22550]. Heavy P2X5 immunostaining was also observed in mouse SON [193].
A double-labeling fluorescence study showed that P2X are differentially expressed on vasopressin- and oxytocin-containing neurons, both in SON and PVN [226,227]. Immunocytochemical labeling in isolated vasopressinergic terminals indicated the presence of P2X2, P2X3, P2X4, and P2X7, but not P2X6 [226], while P2X2, P2X3 and P2X4 were not found in terminals which labeled for oxytocin [226,228]. In the SON sections, vasopressin-containing neurons express P2X2, P2X4, P2X5 and P2X6, while oxytocin-containing neurons express P2X2, P2X4 and P2X5 [227]. Another double-labeling study revealed high colocalization of P2X5 protein and vasopressin in the SON (in 46-58% of AVP neurons) [229]. Furthermore, the distribution of immunoreactivity to P2X6 in the rat hypothalamo-neurohypophysial system studied at the electron microscope level revealed the presence of P2X6 in subpopulations of neurosecretory cell bodies, neurosecretory granules, endoplasmic reticulum, Golgi complex, neurosecretory and non-neurosecretory axons and dendrites in the SON and PVN, pituicytes and microvesicles of the neurohypophysis [230]. The authors hypothesized that P2X6 could be involved in membrane recycling of the granular vesicles and microvesicles at the neurohypophysial level [230]. In addition, immonohistochemistry indicated that P2X7 is located in both vasopressin and oxytocin terminals and soma [231,232].
5.1.2. P2X Activity in SON Cells Studied by Intracellular Calcium Measurements
In the first of a number of these studies on the hypothalamus, ATP was shown to induce a rapid increase in intracellular Ca2+ concentration ([Ca2+]i) in non-identified fura-2M loaded rat hypothalamic neurons in culture [233]. The hydrolysis-resistant ATP analogue, 2-methylthioadenosine-5'triphosphate (2MeSATP), produced a similar response, but the P2X1/P2X3 subtype agonist αβmeATP was much less effective, and ADP, adenosine-5'-monophosphate (AMP), GTP or adenosine had no effect [233]. The ATP-induced calcium response was inhibited by nifedipine (by 62%) and by suramin, general P2X antagonist, indicating that this response resulted from the activation of P2X and influx of extracellular Ca2+ largely through high voltage-gated Ca2+ channels [233].
Consequently, calcium imaging was used to identify functional P2X in fura-2M loaded nerve terminals of magnocellular neurons isolated from posterior pituitary [234], somata of acutely enzymatically dissociated SON neurons [219], SON neurons in hypothalamo-neurohypophyseal system explants [235] and SON neurons in freshly isolated rat brain slices [134]. ATP administration increased [Ca2+]i in approximately 40 % of nerve terminals isolated from posterior pituitary, and this effect was inhibited by suramin and PPADS [234]. Locally applied ATP increased [Ca2+]i also in somata of acutely enzymatically dissociated SON neurons [219] and SON neurons of hypothalamic slices [134]. The effects of ATP was mimicked by general purinoceptor agonists adenosine 5'-O-(2-thiotriphosphate (ATPγS) and 2MeSATP, but not by AMP, adenosine, UTP or UDP, suggesting that P2X2 was involved [134,219]. ATP-induced [Ca2+]i responses were prolonged in the presence of ivermectin, selective positive allosteric modulator of P2X4, suggesting the contribution of P2X4 subtype [134]. A relatively specific P2X7 agonist BzATP increased [Ca2+]i in enzymatically dissociated SON neurons, but the potency of BzATP was lower than potency of ATP, suggesting that P2X7 was not involved [219]. In contrast, BzATP caused a more prominent increase in [Ca2+]i than ATP in SON cells, which were considered non-neuronal because they were immunohistochemically negative for antibodies against vasopressin or oxytocin [219].
It should be emphasized here that both P2X and P2Y receptors are involved in the ATP-induced [Ca2+]i increases observed in SON cells [134,235,236,237]. In hypothalamo-neurohypophyseal explantate, the ATP application triggered an increase in [Ca2+]i by activating a P2Y1 subtype [235]. Also in SON slices, ATP-induced calcium responses were partially reduced (by 25 %) in the presence of specific P2Y1 antagonist N(6)-methyl-2'-deoxyadenosine-3',5'-bisphosphate (MRS2179), and the application of ADP stimulated [Ca2+]i increase in some cells [134], most likely astrocytes [236].
5.1.3. P2X Activity in SON Cells Studied by Using Electrophysiology
Extracellular recording in vivo from SON in anaesthetized rats showed that locally applied ATP excites vasopressinergic neurons that were antidromically identified by means of a bipolar stimulating electrode inserted into the posterior pituitary, and distinguished from oxytocin-containing cells based on the pattern of their electrical phasic activity and their inhibition by arterial baroreceptor activation [238]. This effect was mimicked by αβmeATP and inhibited by suramin, which also blocked the excitation of SON neurons produced by vagus nerve stimulation, a procedure that excites A1 neurons and thus vasopressin neurons [238].
The intracellular recording from SON neurons in superfused explants of rat basal hypothalamus revealed that ATP and ATP agonists (αβmeATP, βγmeATP and 2MeSATP), but not adenosine, evoked depolarization that was not inhibited by tetrodotoxin (TTX), which inhibits voltage-gated sodium channels, but was reversibly inhibited by PPADS, suggesting the involvement of P2X [239].
Whole-cell patch-clamp recording from single dissociated SON neurons showed that ATP dose-dependently evoked a depolarizing inward current, which was accompanied by extracellular Ca2+ entry into the cell [219]. ATP-induced current, inhibited by suramin, could be recorded from both the isolated terminals and somata of magnocellular neurons in explants of the hypothalamo-neurohypophyseal systém [226,228]. Electrophysiological evidence for the presence of P2X in AVP but not oxytocin neurons have also been obtained, as ATP-induced current was observed in 70% of isolated vasopressin-containing neurohypophyseal nerve terminals but not in terminals labeled for oxytocin [226]. ATP-induced somatic current was found in a subpopulation of 62% of SON neurons in acute rat brain slices [134]. The effect of ATP in slices was mimicked by 2MeSATP and ATPγS, but not by the P2X1/P2X3 agonist, αβmeATP [134]. In addition, suramin or PPADS inhibited and ivermectin and/or acid pH 6.5 potentiated the ATP-induced current, indicating the presence of both P2X2 and P2X4 in subpopulation of SON neurons, most likely AVP neurons (Figure 2) [134,240]. Finally, BzATP failed to induce membrane current in SON neurons of slices [134], while BzATP-induced current was observed in dislocated neurohypophysial terminals [232].
ATP application dramatically increases the frequency of glutamatergic miniature excitatory postsynaptic currents (mEPSCs) and GABAergic miniature inhibitory postsynaptic currents (mIPSCs) in 55% and 43% of SON neurons, respectively [134]. ATP increased the frequencies of mIPSC and mEPSC without changing their amplitude, indicating that P2X are present in presynaptic nerve terminals of neurons contacting SCN neurons and that binding of ATP to these receptors facilitated GABA release [134]. The effect of ATP on mIPSC frequency was inhibited by suramin and PPADS and potentiated by pH=6.5, indicating involvement of presynaptic P2X2 [134]. In addition, the effect of ATP on mIPSCs frequency decreased during prolonged (40-60 s) or repeated ATP application, indicating the presence of a desensitizing P2X4 in the presynaptic membrane [134]. The application of ADP, 2MeSAMP and 2MeSADP, agonists of P2Y1 and P2Y12 receptors, failed to modulate the frequency of mIPSC [134].
5.1.4. Functional Relevance of ATP Acting on P2X in the SON
Microinjection of ATP, but not ADP, AMP or adenosine, into the SON in water-loaded and ethanol-anesthetized rats decreased the outflow of urine, suggesting that ATP stimulated P2X on vasopressin-containing neurons, releasing vasopressin from nerve terminals in neurohypophysis and inducing an antidiuretic effect [241]. Also administration of ATP to isolated neurohypophysial nerve terminals induced vasopressin release, but no significant oxytocin release, and this effect was inhibited by suramin or PPADS [226,234]. These observations are in agreement with immonohistochemical [228,231,232] and electrophysiological [134,226] evidence indicating selective expression of P2X2 and P2X4 in subpopulation of vasopressin-containing, but not in oxytocin-containing neurons.
Endogenous sources of ATP in the SON are noradrenergic A1 neurons of the caudal ventrolateral medulla [238,242,243] and astrocytes [5]). In neurohypophysis, endogenous sources of ATP are nerve terminals of magnocellular neurons, which contain ATP in vasopressin and oxytocin secretory vesicles, and release ATP together with neuropeptides [234,244,245]. Electrical field stimulation of isolated neurohypophysis caused a rapid increase in extracellular accumulation of ATP and an increase in the release of vasopressin and oxytocin [244]. However, in the presence of suramin or PPADS, only an increase in the release of vasopressin was reduced, suggesting that endogenously released ATP activated selectively P2X present in vasopressin nerve terminals and potentiated the release of vasopressin, but not oxytocin [245]. Following specific P2X knockout (KO), suramin and PPADS significantly reduce the electrically stimulated release of AVP from intact neurohypophyses of wild-type, P2X3 KO and P2X7 KO, but not P2X2/3 KO mice, suggesting that endogenous ATP stimulated AVP release via P2X2 [246].
Refeeding after fasting represents a complex stimulation to hormone secretion involving the volume/baroreceptors and peripheral/central osmoreceptors. This experimental protocol is widely used to study osmotically-driven stimulation and the synthesis and release of neurohypophyseal peptides, particularly vasopressin and to smaller extend also oxytocin [247,248,249,250,251]. It has been found that fasting for 2 days and refeeding of rats for 2 h, causes an increase in the expression of AVP and P2X2 mRNAs but not oxytocin mRNA in the SON [240], suggesting that ATP via P2X2 likely plays a role during stimulation of vasopressin synthesis in the SON [240].
5.2. PVN
5.2.1. Expression of P2X mRNA and Protein in PVN
In situ hybridization analysis of six P2X (P2X1-X6) mRNAs showed that paraventricular nucleus (PVN) expresses a moderate signal for P2X2, P2X4 and P2X6 [4]. Hybridization analysis of the P2X3 and P2X4 mRNA signal showed that these receptors are expressed in all regions of the PVN, i.e. in the dorsal parvocellular, medial parvocellular and posterior magnocellular component [219].
Ultrastructural studies have shown strong P2X2 and P2X4 immunoreactivity signal in the PVN [108,221,224,225,227,252,253]. P2X2 immunoreactivity was found in every subnucleus, in subpopulations of endocrine neurons, and neurosecretory and non-neurosecretory axons and dendrites [221,222], the most abundant staining was observed in the lateral parvocellular sub-division of the PVN[254]. The majority of the P2X2 immunoreactivity-positive neurons in the PVN were multipolar neurons with long axons and dendrites [222]. Axons with strong P2X2 immunoreactivity were found in the organum vasculosum of the lamina terminalis and median eminence containing nerve endings of parvocellular PVN neurons [222]. In the PVN of the rat hypothalamo-neurohypophysial explants, P2X2 immunoreactivity was found in the subpopulation of magnocellular PVN neurons and their nerve terminals in the neurohypophysis, and axo-dendritic and axo-somatic synapses [225]. Immunohistochemical analysis of the rat hypothalamo-neurohypophysial system revealed the presence of P2X6 in both neurosecretory and non-neurosecretory somata, axons and dendrites in thePVN [230].
Double-labeling fluorescence immunohistochemistry has shown that P2X are expressed differentially on magnocellular vasopressin- and oxytocin-containing PVN neurons: vasopressin-containing neurons express P2X4, P2X5 and P2X6, and oxytocin-containing neurons express only P2X4 [227]. High colocalization of P2X5 and vasopressin was reported in PVN (in 70-90% of AVP neurons) [229].
P2X2 has been found also in parvocellular CRH, TRH and CART neurons that are associated with the regulation of food intake [108]. Injection of the retrogradely transported tracer, rhodamine-tagged microspheres, into the pressor region of the rostral ventrolateral medulla was used to identify non-neuroendocrine parvocellulr neurons in the PVN, and to determine P2X subtype on these neurons [253]. Consequently, the presence of P2X1-P2X6 subtypes was examined, and the proportions of double-labelled neurons for each of the P2X subtypes were quantified. All P2X1-P2X6 subtypes were detected throughout the PVN and it was found that more than one subtype of P2X may be present on a single neuron, the P2X3 and P2X5 being dominant [253]. In addition, double-immunofluorescence showed that P2X7 is co-localizated with the microglial marker in the PVN [255].
5.2.2. P2X Activity in Magnocellular PVN Neurons Studied by Using Electrophysiology
Whole-cell patch-clamp recordings obtained from rat magnocellular PVN neurons, which were identified by their distinct morphology and electrophysiological characteristics, showed that ATP failed to evoke somatic currents [135]. Application of norepinephrine or ATP or BzATP induced a sustained increase in the mEPSC amplitude, and the effect of norepinephrine was inhibited by BBG, the P2X7 antagonist [135]. Interestingly, some studies provided evidence that P2X7 can directly activate phosphatidylinositol-3-kinase (PI3K) a key mediator of AMPA receptor insertion into the cell membrane at hypothalamic glutamate synapses [256,257,258]. Therefore, it has been suggested that endogenously released ATP regulates synaptic plasticity in PVN through the increase in postsynaptic calcium and the insertion of new AMPA receptors into the postsynaptic membrane [135,258].
Another whole-cell patch-clamp recording obtained from magnocellular PVN neurons, identified by their anatomical location in rat brain slices and their unique electrophysiological properties during depolarizing current injection, showed that application of ATP increased the frequency of current-stimulated action potentials [259]. This effect of ATP was blocked by either PPADS and/or kynurenic acid (a glutamate receptor antagonist) [259]. It has been suggested that ATP triggered a shift in the holding current, which was caused by an ATP-induced increase in a tonic extrasynaptic glutamatergic excitatory current, [259]. As hyperosmotic stimulus also exhibits potentiating effect on N-methyl-D-aspartate (NMDA)-evoked currents and increases the frequency of action potentials in magnocellular neurons, it has been suggested that there is a functional excitatory coupling between P2 purinoreceptors and extrasynaptic N-methyl-D-aspartate (NMDA) receptors [259].
5.2.3. P2X Activity in Parvocellular PVN Neurons Studied by Using Electrophysiology
The presence of functional P2X2 on parvocellular PVN neurons was confirmed by single-channel recordings, obtained from outside-out patches in coronal whole-brain slices [260]. Analyses of the single-channel properties of native P2X have shown that at least two distinct P2X populations are present. The first showing “flickery” openings and the second showing “non-flickery” openings. These populations are likely to consist of mixtures of homomeric and heteromeric P2X2, possibly containing P2X2, P2X3, P2X4 and P2X6 subunits [260].
Whole-cell patch-clamp recordings from retrogradely labeled parvocellular neurons projecting to the rostral ventrolateral medulla in the brainstem region showed that ATP application increases the frequency of action potentials in identified neurons, and that this effect is blocked by PPADS and/or kynurenic acid [261]. ATP had no significant effect on the frequency or amplitude of spontaneous excitatory postsynaptic currents (sEPSC), but increased the amplitude of current induced by focal application of AMPA. This ATP effect was blocked by PPADS and chelation of intracellular Ca2+ with BAPTA [261]. Further, hyperosmotic stimulus potentiated the AMPA-evoked currents and this effect was also blocked by PPADS [261]. Thus, it has been suggested that there is a functional stimulatory coupling between extrasynaptically located P2 purinoreceptors and AMPA receptors, which plays a role in the response of parvocellular PVN neurons to acute hyperosmotic stimulus and which in turn could contribute to osmotically driven sympathoexcitatory reactions of PVN [261].
Another whole-cell patch-clamp recording was performed on identified PVN neurons in slices isolated from transgenic mice expressing tdTomato under the control of the P2X4 gene (P2rx4 tdTomato mice). These recordings showed that ATP application does not evoke inward currents that would indicate the presence of functional P2X4 in somata, but increased the frequency and amplitude of mIPSC in some PVN neurons [262]. The ATP-evoked increase in mIPSC frequency was due to the activation of presynaptic P2X4 expressed in the terminals of AgRP/NPY neurons originating from arcuate nucleus (ARC) and projecting to the PVN. These experiments showed that presynaptic P2X4 regulates the release of GABA onto parvocellular PVN neurons involved in sympathoexcitation [262].
5.2.4. Functional Relevance of ATP Acting at P2X in the PVN
Microinjection of ATP into the PVN of water-loaded and ethanol-anesthetized rats produced an antidiuretik effect, while ADP, AMP and adenosine reduced the outflow of urine much less than ATP [263]. The antidiuretic effect of ATP was blocked by prior injection of quinidine (a P2-purinoceptor antagonist) into the PVN, suggesting that ATP activated P2X on magnocellular vasopressin-containing neurons, releasing AVP in the posterior pituitary gland and stimulating AVP(V2) receptors in the kidney [263].
Microinjection of PPADS into the PVN increased blood pressure and heart rate [264,265], indicating that P2X are also involved in the modulation of autonomic function that control the cardiovascular system [264]. Microinjection of αβmeATP [266] or ATP [267] to PVN in vivo and/or in decorticated, unanesthetized, arterially perfused in situ preparation of rat caused pressor and tachycardiac responses [266,267]. Cardiovascular responses evoked by microinjection of αβmeATP or ATP were reduced by pretreatment with PPADS or the glutamate receptor antagonists [266,267], suggesting that ATP-mediated sympatho-excitation depends, at least in part, on the activation of ionotropic glutamate receptors and coupling between extrasynaptically located P2 purinoreceptors and AMPA receptors [261].
Salt-loading, in which drinking water is replaced with 2% NaCl solution for up to 4 days, is an experimental model known to cause a gradual increase in blood pressure and plasma osmolarity that activates oxytocin and vasopressin neurons to a similar extent, causing depletion of stores of both hormones in pituitary [43]. There are several mechanisms, by which salt-loading could influence the activity of vasopressin neurons [268,269](Sharma et al, 2017). These include potentiation of glutamatergic synaptic inputs observed in the SON [27], or norepinephrine-induced release of ATP from astrocytes that are positioned in close proximity of synapses to sense and modulate the afferent synaptic activity described in the PVN [135]. For example, immunohistochemical assessment of glial-fibrillary acidic protein (GFAP) revealed that salt-loading increased glial cell reactivity in the PVN, and measuremens of extracellular ATP and adenosine accumulation via biosensors in hypothalamic slices showed that baseline ATP release was increased 17-fold in the PVN while adenosine remained unchanged. Furthermore, disruption of Ca2+-dependent vesicular release mechanisms in PVN by virally-driven expression of a dominant-negative SNARE protein decreases the release of ATP, indicating that salt-loading stimulates the release of ATP in the PVN from glial cells through the mechanism of Ca2+-dependent exocytosis, and that astrocytic ATP likely contributes to the neural control of blood pressure and plasma osmolarity[270].
Systemic osmotic stimulation induced by intravenous administration of hypertonic solutions triggers a rapid neurohumoral response, which includes release of vasopressin from the posterior pituitary, activation of the sympathetic nervous system, and increase in blood pressure. It was observed that hyperosmotic stimulation increased the number of activated (Fos-positive) P2X2-expressing PVN neurons, and this effect was reduced after PPADS pre-treatment, indicating that a subset of P2X2 expressing PVN neurons might facilitate increased sympathetic activity [254].
Glial-derived ATP also contributes to the effect of ghrelin on vasopressin neurons in the PVN of rat brain slices [271]. Activation of P2X7 in PVN microglia mediating the production of proinflammatory cytokines resulted in increased activity of oxytocinergic and vasopressinergic neurons and activity of the renal sympathetic nervous system [255].
Endogenous source of ATP in the PVN is also blood [272]. PVN is characterized by high capillary density, thin blood vessels and complex vascular topology compared to other areas of the brain. The PVN is thus susceptible to penetration of ATP released from vasculature in response to hemodynamic disturbance after blood pressure increase, which promotes sympathetic overexcitation in hypertension [272].
5.3. SCN
5.3.1. Expression of P2X mRNA and Protein in the SCN
Strong signal for P2X2, P2X4 and P2X6 mRNA was found in the SCN using in situ hybridization [4,273]. Expression of seven P2X (P2X1-7) was studied in circular punches of rat SCN using qRT-PCR [223]. Transcripts for all P2X subunits were found in the SCN, and the relative expression levels of mRNA corresponded to the order: P2X2 > P2X7 > P2X4 > P2X5 > P2X1 = P2X3; the amount of the P2X6 mRNA was very low [223]. Compared to the SON which was examined in the same way in the same laboratory [134], the amount of P2X2 mRNA in the SCN was much lower, about half [223]. Next, spatial and time-of-day-dependent variations in expression of seven P2X (P2X1-7) were analyzed in mouse SCN using qRT-PCR and immunohistochemistry [224]. The relative expression levels of mRNA followed the order: P2X7 > P2X3 > P2X4 > P2X2 > P2X5 > P2X1 > P2X6. Among seven P2X receptors, the following five showed a time-of-day-dependent variation in the relative expression of mRNA: P2X2, P2X3, P2X4, P2X5 and P2X7. All of these rhythmically expressed receptors showed minimal levels of expression at the end of the dark phase (ZT22) and peak levels of expression at the early dark phase (ZT14) [224].
Quantitative immunohistochemical analysis showed a moderate P2X2, P2X4, P2X5 and P2X7 immunoreactivity and very weak P2X1, P2X3 and P2X6 immunoreactivity in the bodies and fibers of SCN neurons [224]. P2X4 immunoreactivity rises in neuronal somata during the dark phase (at ZT 14 - 22), especially in the ventrolateral region of the SCN, while P2X2 immunoreactivity displays no time-of-day-dependent variation [224]. Interestingly, this finding was not specific to SCN and time-of-day-dependent P2 receptor expression was also found in various hippocampal layers [274]. Immunohistochemical staining for P2X2 performed in rat brain coronal sections showed that P2X2 immunoreactivity was absent on the cell bodies of SCN neurons, but confocal microscopy revealed the colocalization of P2X2 with the presynaptic marker of synapsin in numerous, but not all, nerve terminals of SCN neurons in primary cell culture [223]. In addition, P2X2 immunoreactivity-positive neurons and nerve fibres were found in the retrochiasmatic area [222], a region located behind the SCN and in front of arcuate nucleus. This area receives projections from the SCN and sends outputs to control the daily production of melatonin in the pineal gland [275].
Interestingly, a double-labeling fluorescence study revealed that unlike SON and PVN, there is no collocalization of P2X5 and vasopressin in the SCN [229]. Another double-labeling study showed a relatively high signal of P2X7 colocalized with glial-fibrillary acidic protein (GFAP) throughout the SCN region in acutely isolated rat brain slices [6], and experiments performed on the P2rx7-EGFP reporter mouse confirmed positive signal for P2X7 in the SCN [218].
5.3.2. P2X Activity in SCN Cells Studied by Intracellular Calcium Measurements
Single cell calcium measurements in Fura-2AM loaded hypothalamic slices showed that ATP application induces [Ca2+]i increase in 44% of SCN cells, while application of BzATP, ADP, or αβme-ATP induced an increase in [Ca2+]i in only 28%, 26%, 12% and 11% of SCN cells, respectively [223]. ATP-induced [Ca2+]i increases were inhibited by non-selective P2X/P2Y receptor antagonists suramin and PPADS, and partly by tetrodotoxin (TTX), indicating that activation of P2X in neurons contributed to the ATP-stimulated [Ca2+]i responses in SCN slices [223]. Additionaly, in the primary culture of SCN astrocytes, the ATP- and BzATP-stimulated [Ca2+]i responses were inhibited by blockers of P2X7 (AZ 10606120 and A438079) and reduced by MRS2179, blocker of P2Y1, indicating that the ATP-stimulated [Ca2+]i responses in SCN astrocytes resulted from activation of P2X7 and P2Y receptors [6].
5.3.3. P2X Activity in SCN Neurons Studied by Using Electrophysiology
Whole-cell patch-clamp recordings from SCN neurons in rat brain slices revealed that ATP application induced very small somatic current in just 7 % of ventrolateral neurons producing vasoactive intestinal peptide (VIP) and no current in endogenously rhythmic neurons in the dorsomedial region producing AVP [223]. However, ATP application caused a dramatic increase in the frequency mIPSC, without changing their amplitude, in 40% of SCN neurons, both in the ventrolateral and dorsomedial regions, indicating that P2X are present in the nerve terminals of presynaptic neurons contacting SCN neurons [223]. In addition, the ATP-induced increase in mIPSC frequency was inhibited by PPADS and mimicked by 2MeS-ATP or ATPγS, but not by BzATP or ADP, suggesting that presynaptic neurons in the SCN express P2X2 [223].
5.3.4. Functional Relevance of ATP Acting at P2X in the SCN
The ATP content and extracellular ATP concentrations in the SCN show a time-of-day-dependent rhythm that negatively correlates with the rhythm of electrical and metabolic activity of SCN neurons, i.e. the extracellular ATP is high at night and low during the day [6,276,277]. The circadian rhythm in extracellular ATP accumulation matches the circadian rhythm of glia activity, which is also in antiphase with the daily rhythm of electrical activity of SCN neurons [278,279], indicating that ATP is released from SCN astrocytes [6], and the rhythm of ATP secretion is controlled by clock genes in astrocytes [280].
Furthermore, SCN2.2 cells, an SCN-derived cell culture containing 80 % astrocytes, generate circadian oscillations in both the production and extracellular accumulation of ATP with an average period of 23.7 h. [277,281]. Extracellular accumulation of ATP in SCN organotypic cultures depends on the time of day, peaking between 24:00 and 04:00 and minimal at ∼12:00 [6]. Several selective P2X7 antagonists (AZ10606120, A438079 and BBG) have been shown to inhibit extracellular ATP accumulation in organotypic cultures of SCN [6], suggesting that ATP may go out of the cell via P2X7 channels, and that extracellular ATP allows astrocytes to fine-tune their activity with autocrine/paracrine signaling via P2X7.
Interestingly, ATP levels (measured as ATP content in 2 mm-thick coronal slices) have been reported to increase in the initial hours of spontaneous sleep in wake-active regions (i.e. basal forebrain and lateral hypothalamus), but not in sleep-active region (i.e. ventrolateral preoptic region) of the rat [282]. This ATP increase depends on sleep, but not on the time of day, suggesting that the circadian rhythm of extracellular ATP accumulation in the SCN does not appear to be related to sleep, but rather represents an extracellular signal that could, for example, synchronize SCN neurons. Further studies are needed to uncover the role of extracellular ATP accumulation in the SCN.
5.4. AVPV
5.4.1. Expression of P2X mRNA and Protein in GnRH Neurons of AVPV
In situ hybridization showed that the median preoptic nucleus (MnPO), an area that belongs to the preoptic region (POA) and is located near the third ventricle, like the anteroventral periventricular nucleus (AVPV), expresses a strong signal for P2X4 mRNA, a moderate signal for P2X6, but the signal for P2X2 is absent [283].
GnRH neurons originate primarily from embryonic olfactory placode and migrate to specific regions in the brain during development. RT-PCR analysis identified P2X2 and P2X4 in embryonic GnRH neurons in culture from olfactory placode of rhesus monkeys [284]. In adult animals, the distribution of GnRH-positive neurons is diffuse in the brain, hindering efforts to monitor their P2X expression at the tissue level. A detailed analysis was therefore performed on the transcriptome of an entire set of GnRH neurons obtained from coronal brain section of intact, proestrous, and metestrous female GnRH-GFP transgenic mice [285]. About 1500 individual GnRH neurons were sampled from proestrous and metestrous groups, their transcriptome was analyzed using microarray hybridization and real-time PCR, and changes in mRNA expression of genes involved in neurotransmitter signaling during the estrous cycle were investigated. Among P2X, differential gene expression during the estrous cycle was most evident in P2rx7 gene, whose expression was significantly down-regulated by estrogen in proestrous GnRH neurons, suggesting that P2X7 contributes to the induction of the pre-ovulatory GnRH surge [285].
Immunohistochemical analysis showed that P2X2 immunoreactivity-positive neurons and nerve fibres [108,222], and ectonucleotide pyrophosphatase/phosphodiesterase 1 (NPP1) [286] are abundantly expressed in the periventricular nucleus (PeN). Double-labeling fluorescence immunohistochemistry, which examined colocalization between GnRH and P2X1–6 receptors in mice hypothalamus, showed that P2X2, P2X4, P2X5, and P2X6, but not P2X1 and P2X3 are expressed on the perikarya of GnRH neurons in the region around the organum vasculosum of the lamina terminalis (OVLT), and P2X2 and P2X6 are expressed on the axon terminals of GnRH neurons in the median eminence [78].
5.4.2. Expression of P2X mRNA and Protein in Kisspeptin Neurons of AVPV
RNA sequencing (RNA-seq) analysis performed on selectively isolated and actively translated mRNAs from just AVPV Kiss1 neurons of female mice showed that the P2X5 receptor gene (P2rx5) is expressed in AVPV kisspeptin neurons and that estrogen upregulated P2rx5 expression [76], suggesting that ATP-activated P2X5 signaling in AVPV kisspeptin neurons may be involved in GnRH/LH surge generation [76]. Another RNA-seq analysis revealed that the P2X2 receptor gene (P2rx2) is highly expressed in AVPV Kiss1 neurons of rats and is upregulated by estrogen [287]. However, this study failed to detect the expression of other purinergic receptor genes regardless of estrogen milieu, indicating that there may be a species difference in the type of P2X in AVPV kisspeptin neurons [287].
Immunohistochemical analysis showed that the proestrous level of estrogen significantly increased the number of P2X2-immunopositive AVPV kisspeptin neurons, visualized by tdTomato in Kiss1-tdTomato rats, suggesting that ATP-purinergic signaling is involved in estrogen-dependent regulation of reproduction [287].
5.4.3. P2X Activity in GnRH and Kisspeptin Neurons in AVPV Studied by Intracellular Calcium Measurements
Application of ATP, ATP-γS and 2MeSATP to GnRH neurons in fura-2 AM loaded culture derived from the olfactory placode stimulated an increase in [Ca2+]i, while the response to αβmeATP was significantly smaller, and ADP, AMP or BzATP elicited no response [284]. ATP-induced [Ca2+]i response was blocked by PPADS, indicating that functional P2X2 or P2X4 are present in embryonal GnRH neurons [284]. ATP also stimulated [Ca2+]i increases in non-neuronal cells of the olfactory culture, and these cells often responded with a small amplitude to ADP and AMP, but not to adenosine, indicating the presence of P2Y [284]. Cells in olfactory placode culture exhibit spontaneous [Ca2+]i oscillations that are supposed to be involved in synchronization between GnRH neurons and non-neuronal cells [284]. Inhibition of P2X by PPADS has no effect on spontaneous [Ca2+]i oscillations in cultured GnRH neurons from nasal explants of mouse embryos [288].
Intracellular calcium measurements in the immortalized cell line of GnRH neurons (GT1 cells), which have neuronal markers and secrete GnRH [289], did not confirm endogenous expression of any functional P2 purinergic receptor (P2X or P2Y) [290]. By contrast, functional P2X2 was identified in immortalized cell line of AVPV kisspeptin neurons (mHypoA-51 cells), in which ATP application increased [Ca2+]i and co-administration of PPADS blocked this effect [287].
5.4.4. P2X Activity in GnRH and Kisspeptin Neurons Studied by Using Electrophysiology
Although numerous neurotransmitters and gliotransmitters have been shown to directly modulate the electrical activity of GnRH neurons and kisspeptin neurons, for review, see [77], no direct effects of extracellular ATP are described yet.
5.4.5. Functional Relevance of ATP Acting at P2X in the AVPV
Infusion of ATP, and not ADP or AMP, over olfactory cultures from the rhesus monkey embryo stimulated the release of GnRH [284]. The application of ATP, ADP, and αβ-methylene ATP, but not AMP or adenosine, stimulated GnRH release also from explants of median eminence [291]. Furthermore, inhibition on E-NTPDases in the neuroendocrine hypothalamus increased the midcycle LH surge suggesting that elevated endogenous ATP concentration facilitates GnRH neuron secretory activity by activating P2X receptors [292]. Administration of ATP into the AVPV region in estrogen-treated ovariectomized rats stimulated a surge-like increase in LH release, but could not increase LH release in Kiss1 KO rats, suggesting that ATP triggeres GnRH/LH surge by activating P2X expressed selectively on kisspeptin neurons [287]. Since kisspeptin neurons, but not GnRH neurons, express also estrogen receptors, kisspeptin neurons are primarily responsible for the estrogen-depenednt upregulation of P2X expression observed in mice [76]and rats [287].
Immortalized mouse GnRH neurons spontaneously release ATP in culture [293], and ATP is also released along with GnRH from nerve terminals in the median eminence into the hypophyseal-portal system vasculature in ovariectomized sheep [294]. Therefore, elevated concentrations of endogenous ATP in the hypophyseal-portal system may facilitate LH release at the level of pituitary gonadotrophs [295,296], which selectively express P2X2 [297].
5.5. AHN
5.5.1. Expression of P2X in the AHN
While extracellular ATP has been found to be involved in the development of fever [298], there is no evidence of P2X expression in AHN warm-sensitive neurons.
5.5.2. Functional Relevance of ATP in the AHN
At the level of the anterior hypothalamus, fever is initiated by interleukin IL-1beta [299] [300]. There are many evidence showing that extracellular ATP stimulates release of cytokines, such as interleukin IL-1beta and tumour necrosis factor [301,302]. In human monocytes, this effect of ATP may be completely blocked by the P2X7 antagonist, oxidized ATP (oATP), and partly by the antibody against P2X7 [301], suggesting that P2X7 and changes in the level of extracellular ATP in the AHN play a role in the development of febrile response [298].
The possible involvedment of P2X in the central mechanisms of body temperature regulation was studied by injecting ATP into the third ventricle in conscious rats at various ambient temperatures and during a fever induced by bacterial endotoxin, lipopolysaccharide (LPS) [298]. Intracerebroventricular injection of αβmeATP, a stable ATP analogue, induced a decrease or increase in body temperature depending on ambient temperature, and this effect was regulated by general P2X antagonists suramin and PPADS [298], suggesting that the population of warm-sensitive AHN neurons could be the site of action of extracellular ATP [300]. Systemic application of Brilliant Blue G (BBG), specific inhibitor of P2X7, attenuated both fever and increases in plasma cytokine levels evoked by LPS in rats [303].
Next, ATP release was measured in anterior hypothalamus of conscious rabbits in real time with amperometric biosensors during systemic inflammation induced by intravenous injection of LPS. An increase in ATP concentration was observed, confrming that ATP release plays a crucial role in the development of thermoregulatory febrile response [304]. However, all animals treated with BBG, PPADS or oATP application into the anterior hypothalamus, remained febrile 6 hours after induction of systemic inflammation by intravenous administration of LPS [304].
5.6. ARC
5.6.1. Expression of P2X mRNA and Protein in ARC
In situ hybridization revealed that ARC expresses a strong signal for P2X2 mRNA and moderate signals for P2X4 and P2X6 mRNAs [4]. After that, RT-PCR analysis confirmed expression of P2X2 mRNA in ARC [273]. Finally, RNA-seq analysis revealed that unlike AVPV kisspeptin neurons, P2rx2 is not expressed in ARC kisspeptin neurons [287].
P2X2 immunostaining showed similar labeling of cell bodies and fibers in the ARC as described in the PVN and SON [221,222,252,273]. Particularly strong P2X2 immunoreactivity was found in the cellular bodies of orexigenic AgRP/NPY neurons and neural extensions located in the ventromedial part of the ARC, a subregion of the nucleus with a weak blood-brain barrier, involved in the regulation of food intake [108]. Furthermore, a double-labeling showed that P2X2 immunoreactivity was only occasionally detected on anorexigenic POMC/CART neurons and α-MSH- or DYN-containing neurons [108]. Neurons synthesizing somatostatin or nNOS do not appear to express P2X2 [108]. Additionally, iImmunohistochemistry and genetic crosses of P2rx4 tdTomato reporter mice with cell-type-specific GFP reporter lines showed that the tdTomato-expressing cells are primarily AgRP/NPY neurons and tanycytes, providing evidence that P2X4 is expressed inthese cells [262]. Finally, P2X7 was not found to be located in the ARC of P2rx7-EGFP reporter mouse [218].
5.6.2. P2X Activity in ARC Cells Studied by Intracellular Calcium Measurements
Application of ATP and 2-MeSATP, but not the P2X1/P2X3 agonist αβmeATP, induced [Ca2+]i increase in the primary neuroglial cell culture of rat ARC [307]. In the absence of extracellular calcium, the 2-MeSATP-stimulated [Ca2+]i increase was suppressed in all ARC neurons, but only in 25% of astrocytes. Highly P2Y1-selective agonists, MRS2365 and 2-methylthioadenosine-5'diphosphate (2-MeSADP), activated 75-85% of all ARC astrocytes responding to 2-MeSATP [307]. It means that ARC neurons express P2X and the majority of ARC astrocytes express P2Y.
5.6.3. P2X Activity in ARC Cells Studied Using Electrophysiology
Whole-cell patch-clamp recording from single dissociated ARC neurons showed that the application of ATP evokes a large inward current that was inhibited by PPADS [308]. Also ATPγS and 2-MeSATP, but not αβmeATP or βγmeATP, evoked inward currents, suggesting that ARC neurons express functional P2X2 and/or P2X4 [308]. No evidence was found for ATP-evoked inward currents that would be indicative of the presence of functional P2X on somata of identified AgRP/NPY neurons in slices of either wild-type or P2rx4 tdTomato mice [262]. However, ATP application dramatically increased the frequency, but not the amplitude, of mIPSC in POMC neurons contacted by AgRP/NPY neurons, indicating that facilitation of spontaneous GABA release was caused by activation of presynaptic P2X4 in nerve terminals of AgRP/NPY neurons [262].
Perforated whole-cell patch-clamp recording showed that activation of P2Y6 by UDP increased the firing rate of AgRP neurons in lean mice and subsequently promoted feeding [309].
5.6.4. Role of ATP Acting in ARC Physiology
Food-restriction for 3 or 10 days and refeedig of rats after a restriction revealed enhanced expression of hypothalamic P2Y1 and nNOS mRNAs in cell bodies and cellular processes of ARC neurons and astrocytes, indicating that restricted feeding may enhance the sensitivity of the hypothalamus to extracellular ADP/ATP by regulation of the expression of P2Y1 [305]. It has been also observed that hypothalamic UDP contents increases in obese and diabetic mice [309], and that inhibition of P2Y6 signaling in AgRP neurons reduces food intake and fat mass [306]. In addition, Khakh et al. did not find any evidence that ATP regulates feeding behavior, either when ATP was applied within the ARC in vivo or when P2X4 receptors were genetically deleted [262].
5.7. VMH
5.7.1. Expression of P2X mRNA and Protein in the VMH
In situ hybridization study revealed a strong signal for P2X2 and moderate signals for P2X4 and P2X6 mRNAs in the VMH [4]. RT-PCR analysis confirmed expression of P2X2 mRNA in VMH [273]. Immunoreactivity studies showed no or low number of P2X2-positive neurons in the VMH [221,222]. Experiments performed on hypothalamic slices of transgenic mice selectively expressing enhanced GFP in SF-1-positive neurons (SF-1-GFP-positive neurons, characterizing VMH) revealed that 51% of these neurons express P2X4, while only 3% express P2X2 [98]. Moreover, P2X2 and P2X4 receptors are also expressed by non-SF-1-GFP-positive neurons in VMH [98]. The intensity of P2X5 immunofluorescence in VMH is low [229]. In addition, a P2rx7-EGFP reporter mouse was found to have a positive P2X7 signal in the nearby ependymal hypothalamic region, but it was not located in VHM itself [218].
5.7.2. P2X Activity in VMH Cells Studied by Intracellular Calcium Measurements
ATP application induced [Ca2+]i increase in acutely dissociated VMH neurons of the rat [310]. The effect of ATP was mimicked by 2MeSATP and ATPγS, potentiated by Zn2+ and inhibited by suramin and PPADS [310]. ATP induced only a small [Ca2+]i increase when extracellular Na+ is removed, even in the presence of 10 mM Ca2+, suggesting that the [Ca2+]i rise occurred primarily as a result of membrane depolarization and activation of voltage-dependent Ca2+ channels, rather than by an influx via the P2X channel[310].
5.7.3. P2X Activity in VMH Cells Studied by Using Electrophysiology
Whole-cell patch-clamp recording from acutely dissociated VMH neurons of the rat showed that application of ATP evoked a large inward current indicating that ATP activated ionotropic P2X receptors [310]. Patch-clamp recording from SF-1-positive VMH neurons in slices from transgenic mice showed that application of ATP increases frequency of action potentials, but induces inward current of small amplitude [98], which may be due to low number of somatic P2X2 and/or expression of desensitizing P2X4 in these neurons.
Increased surface expression of P2X4, triggered by the use of a peptide competing with P2X4 intracellular endocytosis motif, decreased the frequency and amplitude of GABAergic postsynaptic currents and increased the excitability of SF-1-GFP-positive neurons [98]. Interestingly, co-immunopurification and pull down assays revealed that P2X4 makes a complex with GABAA receptor, suggesting that the formation of the P2X4/ GABAA complex at postsynaptic sites may regulate GABAergic synaptic input and neuronal output in the VMH [98].
5.7.4. Functional Role of ATP Acting at P2 Receptors in the VMH
Intracerebroventricular infusion of ATP/ADP analogues 2-MeSATP and adenosine 5'-O-(2-thiodiphosphate (ADPβS) increased food intake in 18-h food-deprived rats [311]. This effect was abolished by pretreatment with the non-selective P2X/P2Y antagonist PPADS or selective P2Y1 antagonist MRS2179, suggesting that ATP/ADP-induced regulation of food intake is dependent on P2Y1 activation [311]. P2Y1 has been also reported to be involved in obesity [312] and P2Y1 shows major expression in hypothalamic astrocytes of Zucker diabetic fatty (ZDF) rats, which innately develop obesity associated with increases expression of hypothalamic P2Y1 [313].
5.8. DMH
5.8.1. Expression of P2X mRNA and Protein in DMH
Available information on specific expression of P2X mRNA or protein in the dorsomedial nucleus of the hypothalamus (DMH) are still missing. On the other hand, the co-localization of P2Y1 and nNOS was demonstrated on some neurons in this region [305].
5.8.2. P2X Activity in DMH Cells Studied by Intracellular Calcium Measurements
Application of ATP, 2-MeS-ATP or ATPgS stimulated the rise in [Ca2+]i in neurons mechanically dissociated from slices containing DMH region [314]. This effect was reduced by suramin and PPADS, and similar potency of ATP, 2-MeS-ATP and ATPgS is consistent with the pharmacological profile of P2X2 subtypes [314]. The ATP-induced increase in [Ca2+]i was dependent on external Ca2+ and Na+, and was substantially inhibited by voltage-dependent Ca2+ channel blockers, meaning that the [Ca2+]i increase occurred primarily due to membrane depolarization and activation of voltage-dependent Ca2+ channels [314].
5.8.3. P2X Activity in DMH Cells Studied by Using Electrophysiology
Perforated whole-cell patch-clamp recording performed on mechanically dissociated DMH neurons of the rat showed that application of ATP induced an inward current [314]. Rapid rise-time and slow desensitization kinetics was consistent with the expression of functional P2X2 [314]. Furthermore, whole-cell patch-clamp recordings of evoked IPSCs from DMH neurons in rat brain slices showed that ATPγS, a non-hydrolyzable analog of ATP, increases the amplitude of IPSC and potentiates GABA release from inhibitory nerve terminals through activation of presynaptic P2X [124]. It has been suggested that the activation of neighboring astrocytes could trigger the release of a gliotransmitter (ATP) that acts on the presynaptic terminal to stimulate GABA release in the GABA synapses in the DMH [124].
5.8.4. Functional Relevance of ATP Acting on P2 Receptors in the DMH
The mechanisms involving activation of presynaptic P2X and increase in GABA-ergic transmission under basal conditions and following high frequency stimulation could potentially represent modulation of inhibition of orexigenic neurons in DMH, thus influencing feeding behavior. [124]. However, the functional relevance of presynaptic P2X in DMH has not yet been explained in detail.
It has been also suggested that extracellular ATP in DMH is involved in modulating fear and anxiety behavior in the rat [315]. Possible involvement of P2 receptors has been studied by investigating the influences of P2X1/P2X3 agonist αβmeATP, the unspecific P2 receptor antagonist PPADS, the P2Y1-, P2Y11- and P2Y12-specific agonist adenosine 5'-O-(2-thiodiphosphate) (ADPbetaS), and the specific P2Y1 antagonist MRS 2179 on rat behavior in elevated plus-maze [315]. The anxiolytic-like effects were observed after activation of P2Y1, and there was evidence of co-localization of P2Y1 and neuronal NOS (nNOS) on some neurons in this region, suggesting that P2Y1 is involved in anxiety modulation in rats in close association with related nitric oxide production in DMH. [315].
5.9. TMN
5.9.1. Expression of P2X mRNA and Protein in TMN
In situ hybridization showed that TMN expresses strong signal for P2X4 and P2X6 mRNAs, but the intensity of hybridization signal of P2X2 was very low [4]. However, RT-PCR analysis showed expression of P2X2 mRNA in TMN [273]. Single-cell RT–PCR (scRT–PCR) analysis revealed that P2X2 is expressed in the majority of histaminergic neurons in TMN, while each of P2X1, P2X3, P2X4, P2X5 and P2X6 mRNAs was detected in less than 35% of the cells [316]. Single-cell RT-PCR also revealed that most of the histaminergic neurons in TMN express more than one subtype of P2X on soma [317].
5.9.2. P2X Activity in TMN Cells Studied by Electrophysiology
Slowly desensitizing P2X-mediated ATP-evoked currents have been recorded in neurons freshly dissociated from the TMN using the nystatin-perforated patch clamp technique [318]. The potency of ATP analogues was in the order of ATP ≥ 2MeSATP >> αβmeATP. Neither ADP, adenosine nor AMP induced any response, suggesting that TMN neurons express functional somatic P2X2 [318]. In addition, the ATP-induced current recorded from histaminergic neurons of the TMN was potentiated by acidification of the extracellular solution (pH = 6.6) and inhibited by suramin and PPADS, Cibacron blue and trinitrophenyl) adenosine 5'-triphosphate (TNP), suggesting that P2X2 is the major purinoreceptor on the soma of TMN neurons [316,317].
5.9.3. Functional Relevance of ATP Acting at P2X in the TMN
TMN is an important histaminergic nucleus in the hypothalamus involved in arousal and wakefulness, but to our knowledge the functional significance of extracellular ATP and P2X in this region has not yet been studied. Since the homeostatic theory of sleep involves ATP depletion and adenosine accumulation in the brain [282], the role of extracellular ATP and P2X receptors in major sleep-controlling centers, such as TMN, should be examined.
5.10. LHA
5.10.1. Expression of P2X mRNA and Protein in the LHA
Immunohistochemical analysis showed a specific localizations of P2X2 at orexinergic neurons in the lateral hypothalamus [319]. In some LHA cells, P2X2 was co-localized with dynorphin peptides (DYN) [108], which are the endogenous ligands for the kappa opioid receptors (KOR), that regulate food intake [109]. Furthermore, double immunohistochemistry revealed that virtually all orexin-immunoreactive neurons are also P2X2-immunoreactive, whereas 80% of P2X2-immunoreactive neurons are also orexin-positive [320].
In addition, iImmunohistochemical staining also showed that P2Y1 receptors and neuronal nitric oxide synthase (nNOS) colocalize in neurons of the VMH and LH [311], two functionally antagonistic regions involved in the regulation of food intake, in which activation of VMH neurons inhibits feeding, whereas stimulation of LH neurons enhances food intake.
5.10.2. P2X Activity in LHA Cells Studied by Electrophysiology
Whole-cell patch-clamp recording from LHA neurons in hypothalamic slices of transgenic mice expressing green fluorescent protein (GFP) exclusively in cells producing hypocretin/orexin showed that application of ATP depolarizes the cells and induces an increase in action potential frequency, but not frequency of spontaneous miniature postsynaptic currents [319]. In voltage clamp mode, ATP evoked a small inward somatic current that was mimicked by ATPγS, but not by αβmeATP, inhibited by suramin and PPADS, and potentiated by a decrease in extracellular pH, suggesting P2X2 involvement [319]. In addition, suprathreshold activation of neurons of embryonic chick and postnatal mouse lateral hypothalamus in vitro triggered simultaneous release of ATP with GABA [321].
5.10.3. Functional Relevance of ATP Acting at P2X in the LHA
The depolarizing response of LHA neurons to extracellular ATP could lead to an increased activity of the hypocretin arousal system [319], but to our knowledge the functional relevance of ATP acting at P2X has not yet been investigated for this system.
It is interesting that in the LHA there is an increase in ATP release while reducing neuronal activity during sleep [282]. Analysis of ATP content using the luciferin–luciferase-based assay in weighed and homogenized samples of rat brain tissue showed that ATP levels in the lateral hypothalamus decreased after sleep deprivation compared to a time-matched undisturbed sleeping controls [282]. A significant positive correlation was observed between ATP release and EEG delta activity during non-rapid eye movements during spontaneous sleep [282].
In addition, a direct coupling between purinergic signaling and NOS activity during adaptive feeding processes was found in rats after microinjections of P2 agonists into both VMH and LH [311]. As mentioned above (see 5.6.4), the authors demonstrated that food intake is dependent on P2Y1 and nitric oxide production in both regions [311].
6. Conclusions
The aim of this review was to summarize studies on the identification of P2X receptor genes and pathways differentially expressed in hypothalamic nuclei involved in endocrine and nonendocrine functions. Among the seven P2X isoforms described in mammalian cells (P2X1–7), the P2X2, P2X4 and P2X7 receptors were most frequently detected in the hypothalamus at both mRNA and protein levels. The P2X2 and P2X4 are mainly functional in neurons, and P2X7 in astrocytes, ependymal cells and tanycytes. Interestingly, there may be difference in the number or type of the P2X not only between hypothalamic nuclei, but also between neuronal types within individual nuclei. For example, functional P2X are present on vasopressin neurons, but not on oxytocin neurons in the supraoptic nucleus (SON) [226], on kisspeptin neurons, but not on GnRH neurons in the anteroventral periventricular nucleus (AVPV) [287], on VIP neurons but less on AVP neurons in the suprachiasmatic nucleus (SCN) [223], and much more P2X are present on orexigenic AgRP/NPY neurons than on anorexigenic POMC/CART neurons and neurons containing α-MSH or DYN in the arcuate nucleus (ARC) [108]. We believe that future work will clarify these differences as well as the involvement of P2X in specific hypothalamic functions and hypothalamic disorders.
Author Contributions
Conceptualization, H.Z.; writing—original draft preparation, J.C., M.I. and H.Z.; writing—review and editing, H.Z.; visualization, H.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the Charles University Grant Agency (272122) .This publication is based upon work from PRESTO COST Action CA21130 supported by COST (European Cooperation in Science and Technology).
Abbreviations
The following abbreviations are used in this manuscript:
| ADP | adenosine-5'-diphosphate |
| AgRP | agouti-related peptide |
| AMP | adenosine-5'-monophosphate |
| ATP | adenosine-5'-triphosphate |
| ATPγS | adenosine 5'-O-(2-thiotriphosphate) |
| 2MeSATP | 2-methylthio-adenosine triphosphate |
| AMPA | α-amino-3-hydroxy-5-methylisoxazole propionic acid |
| αβmeATP | α,β-methyleneadenosine 5′-triphosphate |
| AHN | anterior hypothalamic nucleus |
| ARC | arcuate nucleus |
| AVP | arginine vasopressin |
| AVPV | anteroventral periventricular nucleus |
| BBG | brilliant blue G |
| BzATP | 3´-O-(4-benzoyl)benzoyladenosine-5´-triphosphate |
| CNS | central nervous system |
| DMH | dorsomedial hypothalamic nukleus |
| DYN | dynorphin peptide |
| GABA | γ-aminobutyric acid |
| GABAA | GABA receptor type A |
| GFP | green fluorescent protein |
| GnRH | gonadotropin-releasing hormone |
| Kiss1R | kisspeptin receptor |
| LHA | lateral hypothalamic area |
| mIPSC | miniature inhibitory postsynaptic current |
| mEPSC | iniature excitatory postsynaptic current |
| MnPO | median preoptic nukleus |
| nNOS | neuronal nitric oxide synthase |
| NMDA | N-methyl-D-aspartate |
| MPOA | medial preoptic area |
| NPP1 | ecto-nucleotide pyrophosphatase/phosphodiesterase 1 |
| NPY | neuropeptide Y |
| P1 | purinergic receptor type 1 |
| P2X | purinergic receptor type 2 (ionotropic) |
| P2Y | purinergic receptor type 2 (metabotropic) |
| POA | preoptic area |
| PPADS | pyridoxalphosphate-6-azophenyl-2', 4'-disulfonic acid |
| PVN | paraventricular nucleus |
| SCN | supraschiasmatic nucleus |
| SON | supraoptic nucleus |
| VMH | ventromedial hypothalamic nucleus |
| VTM | ventral tuberomammillary nucleus |
| TMN | tuberomammillary nucleus |
References
- Russell, J.A. Fifty Years of Advances in Neuroendocrinology. Brain Neurosci. Adv. 2018, 2. [CrossRef]
- Sanchez Jimenez, J.G. and O. De Jesus, Hypothalamic Dysfunction, in StatPearls. 2024: Treasure Island (FL) with ineligible companies. Disclosure: Orlando De Jesus declares no relevant financial relationships with ineligible companies.
- Steuernagel, L.; Lam, B.Y.H.; Klemm, P.; Dowsett, G.K.C.; Bauder, C.A.; Tadross, J.A.; Hitschfeld, T.S.; Martin, A.d.R.; Chen, W.; de Solis, A.J.; et al. HypoMap—a unified single-cell gene expression atlas of the murine hypothalamus. Nat. Metab. 2022, 4, 1402–1419. [CrossRef]
- Collo, G.; North, R.; Kawashima, E.; Merlo-Pich, E.; Neidhart, S.; Surprenant, A.; Buell, G. Cloning OF P2X5 and P2X6 receptors and the distribution and properties of an extended family of ATP-gated ion channels. J. Neurosci. 1996, 16, 2495–2507. [CrossRef]
- Tasker, J.G.; Oliet, S.H.R.; Bains, J.S.; Brown, C.H.; Stern, J.E. Glial Regulation of Neuronal Function: From Synapse to Systems Physiology. J. Neuroendocr. 2011, 24, 566–576. [CrossRef]
- Svobodova, I.; Bhattaracharya, A.; Ivetic, M.; Bendova, Z.; Zemkova, H. Circadian ATP Release in Organotypic Cultures of the Rat Suprachiasmatic Nucleus Is Dependent on P2X7 and P2Y Receptors. Front. Pharmacol. 2018, 9, 192. [CrossRef]
- Collo, G.; Neidhart, S.; Kawashima, E.; Kosco-Vilbois, M.; North, R.; Buell, G. Tissue distribution of the P2X7 receptor. Neuropharmacology 1997, 36, 1277–1283. [CrossRef]
- Genzen, J.R.; Platel, J.-C.; Rubio, M.E.; Bordey, A. Ependymal cells along the lateral ventricle express functional P2X7 receptors. Purinergic Signal. 2009, 5, 299–307. [CrossRef]
- Loesch, A., On P2X receptors in the brain: microvessels. Dedicated to the memory of the late Professor Geoffrey Burnstock (1929-2020). Cell Tissue Res, 2021. 384(3): p. 577-588.
- Caruso, V.; Zuccarini, M.; Di Iorio, P.; Muhammad, I.; Ronci, M. Metabolic Changes Induced by Purinergic Signaling: Role in Food Intake. Front. Pharmacol. 2021, 12. [CrossRef]
- Gao, Z.; Guan, J.; Yin, S.; Liu, F. The role of ATP in sleep-wake regulation: In adenosine-dependent and -independent manner. Sleep Med. 2024, 119, 147–154. [CrossRef]
- Lemos, J.R.; Custer, E.E.; Ortiz-Miranda, S. Purinergic receptor types in the hypothalamic-neurohypophysial system. J. Neuroendocr. 2018, 30. [CrossRef]
- Zemková, H.; Balík, A.; Jindřichová, M.; Vávra, V. Molecular structure of purinergic P2X receptors and their expression in hypothalamus and pituitary. Physiol. Res. 2008, 57 Suppl 3, S23–S38. [CrossRef]
- Stojilkovic, S.S. Purinergic regulation of hypothalamopituitary functions. Trends Endocrinol. Metab. 2009, 20, 460–468. [CrossRef]
- Bjelobaba, I.; Janjic, M.M.; Stojilkovic, S.S. Purinergic signaling pathways in endocrine system. Auton. Neurosci. 2015, 191, 102–116. [CrossRef]
- Stojilkovic, S.S.; Zemkova, H. P2X receptor channels in endocrine glands. Wiley Interdiscip. Rev. Membr. Transp. Signal. 2013, 2, 173–180. [CrossRef]
- Burnstock, G. Purinergic signalling in endocrine organs. Purinergic Signal. 2014, 10, 189–231. [CrossRef]
- Ali, A.A.; Avakian, G.A.; Von Gall, C. The Role of Purinergic Receptors in the Circadian System. Int. J. Mol. Sci. 2020, 21, 3423. [CrossRef]
- Wang, X., et al., The circadian regulation of extracellular ATP. Purinergic Signal, 2023. 19(1): p. 283-295.
- Chen, Y.-H.; Lin, S.; Jin, S.-Y.; Gao, T.-M. Extracellular ATP Is a Homeostatic Messenger That Mediates Cell–Cell Communication in Physiological Processes and Psychiatric Diseases. Biol. Psychiatry 2024, 97, 41–53. [CrossRef]
- Møller, M.; Busch, J.R.; Jacobsen, C.; Lundemose, S.B.; Lynnerup, N.; Rath, M.F.; Banner, J. The accessory magnocellular neurosecretory system of the rostral human hypothalamus. Cell Tissue Res. 2018, 373, 487–498. [CrossRef]
- Voisin, D.L.; E Herbison, A.; A Poulain, D. Central inhibitory effects of muscimol and bicuculline on the milk ejection reflex in the anaesthetized rat.. J. Physiol. 1995, 483, 211–224. [CrossRef]
- Moos, F.C. GABA-induced facilitation of the periodic bursting activity of oxytocin neurones in suckled rats.. J. Physiol. 1995, 488, 103–114. [CrossRef]
- Brussaard, A.B.; Devay, P.; Leyting-Vermeulen, J.L.; Kits, K.S. Changes in properties and neurosteroid regulation of GABAergic synapses in the supraoptic nucleus during the mammalian female reproductive cycle. J. Physiol. 1999, 516, 513–524. [CrossRef]
- Jourdain, P.; Israel, J.-M.; Dupouy, B.; Oliet, S.H.R.; Allard, M.; Vitiello, S.; Theodosis, D.T.; Poulain, D.A. Evidence for a Hypothalamic Oxytocin-Sensitive Pattern-Generating Network Governing Oxytocin NeuronsIn Vitro. J. Neurosci. 1998, 18, 6641–6649. [CrossRef]
- Oliet, S.H. and C.W. Bourque, Mechanosensitive channels transduce osmosensitivity in supraoptic neurons. Nature, 1993. 364(6435): p. 341-3.
- Israel, J.-M.; Poulain, D.A.; Oliet, S.H.R. Glutamatergic Inputs Contribute to Phasic Activity in Vasopressin Neurons. J. Neurosci. 2010, 30, 1221–1232. [CrossRef]
- Choe, K.Y.; Han, S.Y.; Gaub, P.; Shell, B.; Voisin, D.L.; Knapp, B.A.; Barker, P.A.; Brown, C.H.; Cunningham, J.T.; Bourque, C.W. High Salt Intake Increases Blood Pressure via BDNF-Mediated Downregulation of KCC2 and Impaired Baroreflex Inhibition of Vasopressin Neurons. Neuron 2015, 85, 549–560. [CrossRef]
- Pow, D.; Morris, J. Dendrites of hypothalamic magnocellular neurons release neurohypophysial peptides by exocytosis. Neuroscience 1989, 32, 435–439. [CrossRef]
- Sabatier, N.; Caquineau, C.; Dayanithi, G.; Bull, P.; Douglas, A.J.; Guan, X.M.M.; Jiang, M.; Van der Ploeg, L.; Leng, G. α-Melanocyte-Stimulating Hormone Stimulates Oxytocin Release from the Dendrites of Hypothalamic Neurons While Inhibiting Oxytocin Release from Their Terminals in the Neurohypophysis. J. Neurosci. 2003, 23, 10351–10358. [CrossRef]
- Taylor, J.H.; Intorre, A.A.; French, J.A. Vasopressin and Oxytocin Reduce Food Sharing Behavior in Male, but Not Female Marmosets in Family Groups. Front. Endocrinol. 2017, 8, 181–181. [CrossRef]
- Iovino, M.; Messana, T.; Tortora, A.; Giusti, C.; Lisco, G.; Giagulli, V.A.; Guastamacchia, E.; De Pergola, G.; Triggiani, V. Oxytocin Signaling Pathway: From Cell Biology to Clinical Implications. Endocrine, Metab. Immune Disord. - Drug Targets 2021, 21, 91–110. [CrossRef]
- Martucci, L.L.; Launay, J.-M.; Kawakami, N.; Sicard, C.; Desvignes, N.; Dakouane-Giudicelli, M.; Spix, B.; Têtu, M.; Gilmaire, F.-O.; Paulcan, S.; et al. Endolysosomal TPCs regulate social behavior by controlling oxytocin secretion. Proc. Natl. Acad. Sci. 2023, 120. [CrossRef]
- Hu, H., C.A. Zarate, Jr., and J. Verbalis, Arginine vasopressin in mood disorders: A potential biomarker of disease pathology and a target for pharmacologic intervention. Psychiatry Clin Neurosci, 2024.
- Ludwig, M. Dendritic Release of Vasopressin and Oxytocin. J. Neuroendocr. 1998, 10, 881–895. [CrossRef]
- Leng, G.; Ludwig, M. Neurotransmitters and peptides: whispered secrets and public announcements. J. Physiol. 2008, 586, 5625–5632. [CrossRef]
- Kombian, S.B.; Hirasawa, M.; Mouginot, D.; Pittman, Q.J. Chapter 18 Modulation of synaptic transmission by oxytocin and vasopressin in the supraoptic nucleus. In Vasopressin and Oxytocin: From Genes to Clinical Applications; Elsevier: Amsterdam, The Netherlands, 2002; Volume, 139, pp. 235–246. [CrossRef]
- Kombian, S.B.; Hirasawa, M.; Mouginot, D.; Chen, X.; Pittman, Q.J. Short-Term Potentiation of Miniature Excitatory Synaptic Currents Causes Excitation of Supraoptic Neurons. J. Neurophysiol. 2000, 83, 2542–2553. [CrossRef]
- Brussaard, A.B.; Kits, K.S. Changes in GABAA Receptor-Mediated Synaptic Transmission in Oxytocin Neurons during Female Reproduction: Plasticity in a Neuroendocrine Context. Ann. New York Acad. Sci. 1999, 868, 677–680. [CrossRef]
- Li, C.; Tripathi, P.K.; Armstrong, W.E. Differences in spike train variability in rat vasopressin and oxytocin neurons and their relationship to synaptic activity. J. Physiol. 2007, 581, 221–240. [CrossRef]
- Sladek, C.D.; Kapoor, J.R. Neurotransmitter/Neuropeptide Interactions in the Regulation of Neurohypophyseal Hormone Release. Exp. Neurol. 2001, 171, 200–209. [CrossRef]
- Armstrong, W.E. The neurophysiology of neurosecretory cells. J. Physiol. 2007, 585, 645–647. [CrossRef]
- Leng, G.; Brown, C.H.; A Russell, J. Physiological pathways regulating the activity of magnocellular neurosecretory cells. Prog. Neurobiol. 1999, 57, 625–655. [CrossRef]
- Hu, B.; Bourque, C.W. Functional N-Methyl-D-Aspartate and Non-N-Methyl-D-Aspartate Receptors are Expressed by Rat Supraoptic Neurosecretory Cells in vitro. J. Neuroendocr. 1991, 3, 509–514. [CrossRef]
- Shibuya, I.; Kabashima, N.; Ibrahim, N.; Setiadji, S.V.; Ueta, Y.; Yamashita, H. Pre- and postsynaptic modulation of the electrical activity of rat supraoptic neurones. Exp. Physiol. 2000, 85, 145s–151s. [CrossRef]
- Wuarin, J.; Dudek, F. Patch-clamp analysis of spontaneous synaptic currents in supraoptic neuroendocrine cells of the rat hypothalamus. J. Neurosci. 1993, 13, 2323–2331. [CrossRef]
- Decavel, C.; Curras, M. Increased expression of the N-methyl-d-aspartate receptor subunit, NR1, in immunohistochemically identified magnocellular hypothalamic neurons during dehydration. Neuroscience 1997, 78, 191–202. [CrossRef]
- Boudaba, C.; Di, S.; Tasker, J.G. Presynaptic Noradrenergic Regulation of Glutamate Inputs to Hypothalamic Magnocellular Neurones. J. Neuroendocr. 2003, 15, 803–810. [CrossRef]
- Iremonger, K.J.; Benediktsson, A.M.; Bains, J.S. Glutamatergic synaptic transmission in neuroendocrine cells: Basic principles and mechanisms of plasticity. Front. Neuroendocr. 2010, 31, 296–306. [CrossRef]
- Vilhena-Franco, T.; Valentim-Lima, E.; Reis, L.C.; Elias, L.L.K.; Antunes-Rodrigues, J.; Mecawi, A.S. Role of AMPA and NMDA receptors on vasopressin and oxytocin secretion induced by hypertonic extracellular volume expansion. J. Neuroendocr. 2018, 30, e12633. [CrossRef]
- Brown, C.H., Magnocellular Neurons and Posterior Pituitary Function. Compr Physiol, 2016. 6(4): p. 1701-1741.
- Ferguson, A.V.; Latchford, K.J.; Samson, W.K. The paraventricular nucleus of the hypothalamus – a potential target for integrative treatment of autonomic dysfunction. Expert Opin. Ther. Targets 2008, 12, 717–727. [CrossRef]
- Rasiah, N.P., S.P. Loewen, and J.S. Bains, Windows into stress: a glimpse at emerging roles for CRH(PVN) neurons. Physiol Rev, 2023. 103(2): p. 1667-1691.
- Sawchenko, P.E., et al., The paraventricular nucleus of the hypothalamus and the functional neuroanatomy of visceromotor responses to stress. Prog Brain Res, 1996. 107: p. 201-22.
- Simmons, D.M.; Swanson, L.W. Comparison of the spatial distribution of seven types of neuroendocrine neurons in the rat paraventricular nucleus: Toward a global 3D model. J. Comp. Neurol. 2009, 516, 423–441. [CrossRef]
- Yang, Z.; Coote, J.H. Influence of the hypothalamic paraventricular nucleus on cardiovascular neurones in the rostral ventrolateral medulla of the rat. J. Physiol. 1998, 513, 521–530. [CrossRef]
- Shafton, A.D.; Ryan, A.; Badoer, E. Neurons in the hypothalamic paraventricular nucleus send collaterals to the spinal cord and to the rostral ventrolateral medulla in the rat. Brain Res. 1998, 801, 239–243. [CrossRef]
- Sanders, J.; Nemeroff, C. The CRF System as a Therapeutic Target for Neuropsychiatric Disorders. Trends Pharmacol. Sci. 2016, 37, 1045–1054. [CrossRef]
- Marsh, N.; Marsh, A.A.; Lee, M.R.; Hurlemann, R. Oxytocin and the Neurobiology of Prosocial Behavior. Neurosci. 2020, 27, 604–619. [CrossRef]
- Borroto-Escuela, D.O.; Cuesta-Marti, C.; Lopez-Salas, A.; Chruścicka-Smaga, B.; Crespo-Ramírez, M.; Tesoro-Cruz, E.; Palacios-Lagunas, D.A.; de la Mora, M.P.; Schellekens, H.; Fuxe, K. The oxytocin receptor represents a key hub in the GPCR heteroreceptor network: potential relevance for brain and behavior. Front. Mol. Neurosci. 2022, 15, 1055344. [CrossRef]
- Santoso, P.; Nakata, M.; Ueta, Y.; Yada, T. Suprachiasmatic vasopressin to paraventricular oxytocin neurocircuit in the hypothalamus relays light reception to inhibit feeding behavior. Am. J. Physiol. Metab. 2018, 315, E478–E488. [CrossRef]
- Vargas, Y.; Tron, A.E.C.; Rodríguez, A.R.; Uribe, R.M.; Joseph-Bravo, P.; Charli, J.-L. Thyrotropin-Releasing Hormone and Food Intake in Mammals: An Update. Metabolites 2024, 14, 302. [CrossRef]
- Barrett-Jolley, R.; Nunn, N.; Womack, M.; Dart, C. Function and Pharmacology of Spinally-Projecting Sympathetic Pre-Autonomic Neurones in the Paraventricular Nucleus of the Hypothalamus. Curr. Neuropharmacol. 2011, 9, 262–277. [CrossRef]
- Strecker, G.J.; Wuarin, J.-P.; Dudek, F.E. GABAA-Mediated Local Synaptic Pathways Connect Neurons in the Rat Suprachiasmatic Nucleus. J. Neurophysiol. 1997, 78, 2217–2220. [CrossRef]
- Moore, R.Y.; Eichler, V.B. Loss of a circadian adrenal corticosterone rhythm following suprachiasmatic lesions in the rat. Brain Res. 1972, 42, 201–206. [CrossRef]
- Stephan, F.K.; Zucker, I. Circadian Rhythms in Drinking Behavior and Locomotor Activity of Rats Are Eliminated by Hypothalamic Lesions. Proc. Natl. Acad. Sci. 1972, 69, 1583–1586. [CrossRef]
- Sumová, A.; Trávnícková, Z.; Peters, R.; Schwartz, W.J.; Illnerová, H. The rat suprachiasmatic nucleus is a clock for all seasons.. Proc. Natl. Acad. Sci. 1995, 92, 7754–7758. [CrossRef]
- Reppert, S.M., A clockwork explosion! Neuron, 1998. 21(1): p. 1-4.
- Lee, H.S.; Billings, H.J.; Lehman, M.N. The Suprachiasmatic Nucleus: A Clock of Multiple Components. J. Biol. Rhythm. 2003, 18, 435–449. [CrossRef]
- Moore, R.Y.; Card, J.P. Visual Pathways and the Entrainment of Circadian Rhythmsa. Ann. New York Acad. Sci. 1985, 453, 123–133. [CrossRef]
- Jacomy, H.; Burlet, A.; Bosler, O. Vasoactive intestinal peptide neurons as synaptic targets for vasopressin neurons in the suprachiasmatic nucleus. Double-label immunocytochemical demonstration in the rat. Neuroscience 1999, 88, 859–870. [CrossRef]
- Brancaccio, M.; Enoki, R.; Mazuski, C.N.; Jones, J.; Evans, J.A.; Azzi, A. Network-Mediated Encoding of Circadian Time: The Suprachiasmatic Nucleus (SCN) from Genes to Neurons to Circuits, and Back. J. Neurosci. 2014, 34, 15192–15199. [CrossRef]
- Inouye, S.T.; Kawamura, H. Persistence of circadian rhythmicity in a mammalian hypothalamic "island" containing the suprachiasmatic nucleus.. Proc. Natl. Acad. Sci. 1979, 76, 5962–5966. [CrossRef]
- Groos, G.; Hendriks, J. Circadian rhythms in electrical discharge of rat suprachiasmatic neurones recorded in vitro. Neurosci. Lett. 1982, 34, 283–288. [CrossRef]
- Pennartz, C.M.A.; de Jeu, M.T.G.; Bos, N.P.A.; Schaap, J.; Geurtsen, A.M.S. Diurnal modulation of pacemaker potentials and calcium current in the mammalian circadian clock. Nature 2002, 416, 286–290. [CrossRef]
- Stephens, S.B.Z.; Kauffman, A.S. Estrogen Regulation of the Molecular Phenotype and Active Translatome of AVPV Kisspeptin Neurons. Endocrinology 2021, 162. [CrossRef]
- Spergel, D.J. Modulation of Gonadotropin-Releasing Hormone Neuron Activity and Secretion in Mice by Non-peptide Neurotransmitters, Gasotransmitters, and Gliotransmitters. Front. Endocrinol. 2019, 10, 329. [CrossRef]
- Fu, J.; Yu, Q.; Guo, W.; He, C.; Burnstock, G.; Xiang, Z. P2X receptors are expressed on neurons containing luteinizing hormone-releasing hormone in the mouse hypothalamus. Neurosci. Lett. 2009, 458, 32–36. [CrossRef]
- Sarkar, D.K.; Chiappa, S.A.; Fink, G.; Sherwood, N.M. Gonadotropin-releasing hormone surge in pro-oestrous rats. Nature 1976, 264, 461–463. [CrossRef]
- Baca-Alonso, J.J.A. and J.L. Quintanar, The effect of gonadotropin-releasing hormone on the nervous system. Neuro Endocrinol Lett, 2024. 45(3): p. 188-196.
- Bakker, J., Can kisspeptin be a new treatment for sexual dysfunction? Trends Endocrinol Metab, 2025.
- Messager, S.; Chatzidaki, E.E.; Ma, D.; Hendrick, A.G.; Zahn, D.; Dixon, J.; Thresher, R.R.; Malinge, I.; Lomet, D.; Carlton, M.B.L.; et al. Kisspeptin directly stimulates gonadotropin-releasing hormone release via G protein-coupled receptor 54. Proc. Natl. Acad. Sci. 2005, 102, 1761–1766. [CrossRef]
- de Roux, N., et al., Hypogonadotropic hypogonadism due to loss of function of the KiSS1-derived peptide receptor GPR54. Proc Natl Acad Sci U S A, 2003. 100(19): p. 10972-6.
- Sliwowska, J.H.; Woods, N.E.; Alzahrani, A.R.; Paspali, E.; Tate, R.J.; Ferro, V.A. Kisspeptin a potential therapeutic target in treatment of both metabolic and reproductive dysfunction. J. Diabetes 2024, 16, e13541. [CrossRef]
- Boulant, J.A.; Hardy, J.D. The effect of spinal and skin temperatures on the firing rate and thermosensitivity of preoptic neurones. J. Physiol. 1974, 240, 639–660. [CrossRef]
- Bouret, S.G.; Draper, S.J.; Simerly, R.B. Formation of Projection Pathways from the Arcuate Nucleus of the Hypothalamus to Hypothalamic Regions Implicated in the Neural Control of Feeding Behavior in Mice. J. Neurosci. 2004, 24, 2797–2805. [CrossRef]
- Martins, A.B.; Brownlow, M.L.; Araújo, B.B.; Garnica-Siqueira, M.C.; Zaia, D.A.M.; Leite, C.M.; Zaia, C.T.B.V.; Uchoa, E.T. Arcuate nucleus of the hypothalamus contributes to the hypophagic effect and plasma metabolic changes induced by vasoactive intestinal peptide and pituitary adenylate cyclase-activating polypeptide. Neurochem. Int. 2022, 155, 105300. [CrossRef]
- Fioramonti, X.; Lorsignol, A.; Taupignon, A.; Pénicaud, L. A New ATP-Sensitive K+ Channel–Independent Mechanism Is Involved in Glucose-Excited Neurons of Mouse Arcuate Nucleus. Diabetes 2004, 53, 2767–2775. [CrossRef]
- Mehay, D.; Silberman, Y.; Arnold, A.C. The Arcuate Nucleus of the Hypothalamus and Metabolic Regulation: An Emerging Role for Renin–Angiotensin Pathways. Int. J. Mol. Sci. 2021, 22, 7050. [CrossRef]
- dos Santos, K.M.; Saunders, S.E.; Antunes, V.R.; Boychuk, C.R. Insulin activates parasympathetic hepatic-related neurons of the paraventricular nucleus of the hypothalamus through mTOR signaling. J. Neurophysiol. 2025, 133, 320–332. [CrossRef]
- Clarkson, J.; Han, S.Y.; Piet, R.; McLennan, T.; Kane, G.M.; Ng, J.; Porteous, R.W.; Kim, J.S.; Colledge, W.H.; Iremonger, K.J.; et al. Definition of the hypothalamic GnRH pulse generator in mice. Proc. Natl. Acad. Sci. 2017, 114, E10216–E10223. [CrossRef]
- Goto, T.; Hagihara, M.; Miyamichi, K.; Japan Dynamics of pulsatile activities of arcuate kisspeptin neurons in aging female mice. eLife 2023, 12. [CrossRef]
- Koysombat, K.; Tsoutsouki, J.; Patel, A.H.; Comninos, A.N.; Dhillo, W.S.; Abbara, A. Kisspeptin and neurokinin B: roles in reproductive health. Physiol. Rev. 2025, 105, 707–764. [CrossRef]
- Sawchenko, P.; Swanson, L. The organization of noradrenergic pathways from the brainstem to the paraventricular and supraoptic nuclei in the rat. Brain Res. Rev. 1982, 4, 275–325. [CrossRef]
- Mezey, E., et al., Distribution of the pro-opiomelanocortin derived peptides, adrenocorticotrope hormone, alpha-melanocyte-stimulating hormone and beta-endorphin (ACTH, alpha-MSH, beta-END) in the rat hypothalamus. Brain Res, 1985. 328(2): p. 341-7.
- Morgane, P.J.; Cho, Y.K.; Li, C.-S.; Smith, D.V. Electrophysiological studies of feeding and satiety centers in the rat. Am. J. Physiol. Content 1961, 201, 838–844. [CrossRef]
- King, B.M. The rise, fall, and resurrection of the ventromedial hypothalamus in the regulation of feeding behavior and body weight. Physiol. Behav. 2006, 87, 221–244. [CrossRef]
- Jo, Y.H., et al., Crosstalk between P2X4 and GABA-A receptors determines synaptic efficacy at central synapses. J Biol Chem, 2011.
- Cheung, C.C.; Kurrasch, D.M.; Liang, J.K.; Ingraham, H.A. Genetic labeling of steroidogenic factor-1 (SF-1) neurons in mice reveals ventromedial nucleus of the hypothalamus (VMH) circuitry beginning at neurogenesis and development of a separate non-SF-1 neuronal cluster in the ventrolateral VMH. J. Comp. Neurol. 2012, 521, 1268–1288. [CrossRef]
- Sapkota, S.; Roy, S.C.; Briski, K.P. Dorsomedial Ventromedial Hypothalamic Nucleus Growth Hormone-Releasing Hormone Neuron Steroidogenic Factor-1 Gene Targets in Female Rat. ASN Neuro 2024, 16, 2403345. [CrossRef]
- Fosch, A.; Zagmutt, S.; Casals, N.; Rodríguez-Rodríguez, R. New Insights of SF1 Neurons in Hypothalamic Regulation of Obesity and Diabetes. Int. J. Mol. Sci. 2021, 22, 6186. [CrossRef]
- Bellinger, L.L. and L.L. Bernardis, The dorsomedial hypothalamic nucleus and its role in ingestive behavior and body weight regulation: lessons learned from lesioning studies. Physiol Behav, 2002. 76(3): p. 431-42.
- Brasil, T.F.S.; Lopes-Azevedo, S.; Belém-Filho, I.J.A.; Fortaleza, E.A.T.; Antunes-Rodrigues, J.; Corrêa, F.M.A. The Dorsomedial Hypothalamus Is Involved in the Mediation of Autonomic and Neuroendocrine Responses to Restraint Stress. Front. Pharmacol. 2020, 10, 1547. [CrossRef]
- DiMicco, J.A., et al., The dorsomedial hypothalamus and the response to stress: part renaissance, part revolution. Pharmacol Biochem Behav, 2002. 71(3): p. 469-80.
- Sakai, K.; Takahashi, K.; Anaclet, C.; Lin, J.-S. Sleep-waking discharge of ventral tuberomammillary neurons in wild-type and histidine decarboxylase knock-out mice. Front. Behav. Neurosci. 2010, 4, 53. [CrossRef]
- Wada, H.; Inagaki, N.; Itowi, N.; Yamatodani, A. Histaminergic neuron system in the brain: Distribution and possible functions. Brain Res. Bull. 1991, 27, 367–370. [CrossRef]
- Mickelsen, L.E.; Bolisetty, M.; Chimileski, B.R.; Fujita, A.; Beltrami, E.J.; Costanzo, J.T.; Naparstek, J.R.; Robson, P.; Jackson, A.C. Single-cell transcriptomic analysis of the lateral hypothalamic area reveals molecularly distinct populations of inhibitory and excitatory neurons. Nat. Neurosci. 2019, 22, 642–656. [CrossRef]
- Colldén, G.; Mangano, C.; Meister, B. P2X2 purinoreceptor protein in hypothalamic neurons associated with the regulation of food intake. Neuroscience 2010, 171, 62–78. [CrossRef]
- Sandoval-Caballero, C.; Jara, J.; Luarte, L.; Jiménez, Y.; Teske, J.; Perez-Leighton, C. Control of motivation for sucrose in the paraventricular hypothalamic nucleus by dynorphin peptides and the kappa opioid receptor. Appetite 2024, 200, 107504. [CrossRef]
- Bernardis, L.L.; Bellinger, L.L. The lateral hypothalamic area revisited: Neuroanatomy, body weight regulation, neuroendocrinology and metabolism. Neurosci. Biobehav. Rev. 1993, 17, 141–193. [CrossRef]
- Bonnavion, P.; Mickelsen, L.E.; Fujita, A.; de Lecea, L.; Jackson, A.C. Hubs and spokes of the lateral hypothalamus: cell types, circuits and behaviour. J. Physiol. 2016, 594, 6443–6462. [CrossRef]
- Samson, W.K.; Taylor, M.M.; Follwell, M.; Ferguson, A.V. Orexin actions in hypothalamic paraventricular nucleus: physiological consequences and cellular correlates. Regul. Pept. 2001, 104, 97–103. [CrossRef]
- Lopez, R.; Young, S.L.; Cox, V.C. Analgesia for formalin-induced pain by lateral hypothalamic stimulation. Brain Res. 1991, 563, 1–6. [CrossRef]
- Khan, R.; Lee, B.; Inyang, K.; Bemis, H.; Bugescu, R.; Laumet, G.; Leinninger, G. Neurotensin-expressing lateral hypothalamic neurons alleviate neuropathic and inflammatory pain via neurotensin receptor signaling. Neurobiol. Pain 2024, 16, 100172. [CrossRef]
- Beamer, E.; Conte, G.; Engel, T. ATP release during seizures – A critical evaluation of the evidence. Brain Res. Bull. 2019, 151, 65–73. [CrossRef]
- Jurányi, Z.; Sperlágh, B.; Vizi, E. Involvement of P2 purinoceptors and the nitric oxide pathway in [3H]purine outflow evoked by short-term hypoxia and hypoglycemia in rat hippocampal slices. Brain Res. 1999, 823, 183–190. [CrossRef]
- Godoy, P.A.; Ramírez-Molina, O.; Fuentealba, J. Exploring the Role of P2X Receptors in Alzheimer’s Disease. Front. Pharmacol. 2019, 10, 1330. [CrossRef]
- Guthrie, P.B.; Knappenberger, J.; Segal, M.; Bennett, M.V.L.; Charles, A.C.; Kater, S.B. ATP Released from Astrocytes Mediates Glial Calcium Waves. J. Neurosci. 1999, 19, 520–528. [CrossRef]
- Fields, R.D. and B. Stevens, ATP: an extracellular signaling molecule between neurons and glia. Trends Neurosci, 2000. 23(12): p. 625-33.
- Inoue, K.; Koizumi, S.; Tsuda, M. The role of nucleotides in the neuron–glia communication responsible for the brain functions. J. Neurochem. 2007, 102, 1447–1458. [CrossRef]
- Abbracchio, M.P.; Burnstock, G.; Verkhratsky, A.; Zimmermann, H. Purinergic signalling in the nervous system: an overview. Trends Neurosci. 2009, 32, 19–29. [CrossRef]
- Lalo, U.; Palygin, O.; Verkhratsky, A.; Grant, S.G.N.; Pankratov, Y. ATP from synaptic terminals and astrocytes regulates NMDA receptors and synaptic plasticity through PSD-95 multi-protein complex. Sci. Rep. 2016, 6, srep33609. [CrossRef]
- Araque, A.; Parpura, V.; Sanzgiri, R.P.; Haydon, P.G. Tripartite synapses: glia, the unacknowledged partner. Trends Neurosci. 1999, 22, 208–215. [CrossRef]
- Crosby, K.M.; Murphy-Royal, C.; Wilson, S.A.; Gordon, G.R.; Bains, J.S.; Pittman, Q.J. Cholecystokinin Switches the Plasticity of GABA Synapses in the Dorsomedial Hypothalamus via Astrocytic ATP Release. J. Neurosci. 2018, 38, 8515–8525. [CrossRef]
- Hong, Y.; Zhao, T.; Li, X.-J.; Li, S. Mutant Huntingtin Impairs BDNF Release from Astrocytes by Disrupting Conversion of Rab3a-GTP into Rab3a-GDP. J. Neurosci. 2016, 36, 8790–8801. [CrossRef]
- Lazarowski, E.R., et al., Molecular mechanisms of purine and pyrimidine nucleotide release. Adv Pharmacol, 2011. 61: p. 221-61.
- Lazarowski, E.R. Vesicular and conductive mechanisms of nucleotide release. Purinergic Signal. 2012, 8, 359–373. [CrossRef]
- Burnstock, G.; Fredholm, B.B.; Verkhratsky, A. Adenosine and ATP Receptors in the Brain. Curr. Top. Med. Chem. 2011, 11, 973–1011. [CrossRef]
- Jo, Y.-H.; Schlichter, R. Synaptic corelease of ATP and GABA in cultured spinal neurons. Nat. Neurosci. 1999, 2, 241–245. [CrossRef]
- Nörenberg, W.; Illes, P. Neuronal P2X receptors: localisation and functional properties. Naunyn-Schmiedeberg's Arch. Pharmacol. 2000, 362, 324–339. [CrossRef]
- Jang, I.; Rhee, J.S.; Kubota, H.; Akaike, N.; Akaike, N. Developmental changes in P2X purinoceptors on glycinergic presynaptic nerve terminals projecting to rat substantia gelatinosa neurones. J. Physiol. 2001, 536, 505–519. [CrossRef]
- Rodrigues, R.J., et al., Dual presynaptic control by ATP of glutamate release via facilitatory P2X1, P2X2/3, and P2X3 and inhibitory P2Y1, P2Y2, and/or P2Y4 receptors in the rat hippocampus. J Neurosci, 2005. 25(27): p. 6286-95.
- Burnstock, G., Purinergic cotransmission. Exp Physiol, 2009. 94(1): p. 20-4.
- Vavra, V.; Bhattacharya, A.; Zemkova, H. Facilitation of glutamate and GABA release by P2X receptor activation in supraoptic neurons from freshly isolated rat brain slices. Neuroscience 2011, 188, 1–12. [CrossRef]
- Gordon, G.R.J.; Baimoukhametova, D.V.; A Hewitt, S.; Rajapaksha, W.R.A.K.J.S.; E Fisher, T.; Bains, J.S. Norepinephrine triggers release of glial ATP to increase postsynaptic efficacy. Nat. Neurosci. 2005, 8, 1078–1086. [CrossRef]
- Fumagalli, M.; Brambilla, R.; D'Ambrosi, N.; Volonté, C.; Matteoli, M.; Verderio, C.; Abbracchio, M.P. Nucleotide-mediated calcium signaling in rat cortical astrocytes: Role of P2X and P2Y receptors. Glia 2003, 43, 218–230. [CrossRef]
- Pascual, O.; Casper, K.B.; Kubera, C.; Zhang, J.; Revilla-Sanchez, R.; Sul, J.-Y.; Takano, H.; Moss, S.J.; McCarthy, K.; Haydon, P.G. Astrocytic Purinergic Signaling Coordinates Synaptic Networks. Science 2005, 310, 113–116. [CrossRef]
- Pangršič, T.; Potokar, M.; Stenovec, M.; Kreft, M.; Fabbretti, E.; Nistri, A.; Pryazhnikov, E.; Khiroug, L.; Giniatullin, R.; Zorec, R. Exocytotic Release of ATP from Cultured Astrocytes. J. Biol. Chem. 2007, 282, 28749–28758. [CrossRef]
- Stout, C.E.; Costantin, J.L.; Naus, C.C.G.; Charles, A.C. Intercellular Calcium Signaling in Astrocytes via ATP Release through Connexin Hemichannels. J. Biol. Chem. 2002, 277, 10482–10488. [CrossRef]
- Schenk, U.; Westendorf, A.M.; Radaelli, E.; Casati, A.; Ferro, M.; Fumagalli, M.; Verderio, C.; Buer, J.; Scanziani, E.; Grassi, F. Purinergic control of T cell activation by ATP released through pannexin-1 hemichannels. Sci. Signal. 2008, 1, ra6. [CrossRef]
- Iglesias, R., et al., Pannexin 1: the molecular substrate of astrocyte "hemichannels". J Neurosci, 2009. 29(21): p. 7092-7.
- Li, S.; Bjelobaba, I.; Yan, Z.; Kucka, M.; Tomić, M.; Stojilkovic, S.S. Expression and Roles of Pannexins in ATP Release in the Pituitary Gland. Endocrinology 2011, 152, 2342–2352. [CrossRef]
- Khakh, B.S.; Sofroniew, M.V. Diversity of astrocyte functions and phenotypes in neural circuits. Nat. Neurosci. 2015, 18, 942–952. [CrossRef]
- Kinoshita, M.; Hirayama, Y.; Fujishita, K.; Shibata, K.; Shinozaki, Y.; Shigetomi, E.; Takeda, A.; Le, H.P.N.; Hayashi, H.; Hiasa, M.; et al. Anti-Depressant Fluoxetine Reveals its Therapeutic Effect Via Astrocytes. EBioMedicine 2018, 32, 72–83. [CrossRef]
- North, R.A. Molecular physiology of P2X receptors. Physiol. Rev. 2002, 82, 1013–1067. [CrossRef]
- Nicke, A.; Bäumert, H.G.; Rettinger, J.; Eichele, A.; Lambrecht, G.; Mutschler, E.; Schmalzing, G. P2X1 and P2X3 receptors form stable trimers: a novel structural motif of ligand-gated ion channels. EMBO J. 1998, 17, 3016–3028. [CrossRef]
- Pelegrin, P.; Surprenant, A. Pannexin-1 mediates large pore formation and interleukin-1β release by the ATP-gated P2X7 receptor. EMBO J. 2006, 25, 5071–5082. [CrossRef]
- Jo, Y.H., et al., Crosstalk between P2X4 and GABA-A receptors determines synaptic efficacy at central synapses. J Biol Chem, 2011. 256: p. 19993-20004.
- Queme, L.F.; Weyler, A.A.; Cohen, E.R.; Hudgins, R.C.; Jankowski, M.P. A dual role for peripheral GDNF signaling in nociception and cardiovascular reflexes in the mouse. Proc. Natl. Acad. Sci. 2019, 117, 698–707. [CrossRef]
- Khakh, B.S. Molecular physiology of p2x receptors and atp signalling at synapses. Nat. Rev. Neurosci. 2001, 2, 165–174. [CrossRef]
- Stojilkovic, S.S.; Tomić, M.; He, M.; Yan, Z.; Koshimizu, T.; Zemkova, H. Molecular Dissection of Purinergic P2X Receptor Channels. Ann. New York Acad. Sci. 2005, 1048, 116–130. [CrossRef]
- Khakh, B.S.; Proctor, W.R.; Dunwiddie, T.V.; Labarca, C.; Lester, H.A. Allosteric Control of Gating and Kinetics at P2X4Receptor Channels. J. Neurosci. 1999, 19, 7289–7299. [CrossRef]
- Coddou, C.; Yan, Z.; Obsil, T.; Huidobro-Toro, J.P.; Stojilkovic, S.S. Activation and Regulation of Purinergic P2X Receptor Channels. Pharmacol. Rev. 2011, 63, 641–683. [CrossRef]
- Coddou, C.; Stojilkovic, S.S.; Huidobro-Toro, J.P. Allosteric modulation of ATP-gated P2X receptor channels. Prog. Neurobiol. 2011, 22, 335–354. [CrossRef]
- Stokes, L.; Bidula, S.; Bibič, L.; Allum, E. To Inhibit or Enhance? Is There a Benefit to Positive Allosteric Modulation of P2X Receptors? Front. Pharmacol. 2020, 11, 627. [CrossRef]
- Illes, P.; Müller, C.E.; Jacobson, K.A.; Grutter, T.; Nicke, A.; Fountain, S.J.; Kennedy, C.; Schmalzing, G.; Jarvis, M.F.; Stojilkovic, S.S.; et al. Update of P2X receptor properties and their pharmacology: IUPHAR Review 30. Br. J. Pharmacol. 2020, 178, 489–514. [CrossRef]
- Sivcev, S.; Kudova, E.; Zemkova, H. Neurosteroids as positive and negative allosteric modulators of ligand-gated ion channels: P2X receptor perspective. Neuropharmacology 2023, 234, 109542. [CrossRef]
- Bianchi, B.R.; Lynch, K.J.; Touma, E.; Niforatos, W.; Burgard, E.C.; Alexander, K.M.; Park, H.S.; Yu, H.; Metzger, R.; Kowaluk, E.; et al. Pharmacological characterization of recombinant human and rat P2X receptor subtypes. Eur. J. Pharmacol. 1999, 376, 127–138. [CrossRef]
- Samways, D.S.K.; Li, Z.; Egan, T.M. Principles and properties of ion flow in P2X receptors. Front. Cell. Neurosci. 2014, 8, 6. [CrossRef]
- Oury, C.; Toth-Zsamboki, E.; Van Geet, C.; Thys, C.; Wei, L.; Nilius, B.; Vermylen, J.; Hoylaerts, M.F. A Natural Dominant Negative P2X1 Receptor Due to Deletion of a Single Amino Acid Residue. J. Biol. Chem. 2000, 275, 22611–22614. [CrossRef]
- Lynch, K.J., et al., Molecular and functional characterization of human P2X(2) receptors. Mol Pharmacol, 1999. 56(6): p. 1171-81.
- Zhong, Y.; Dunn, P.M.; Xiang, Z.; Bo, X.; Burnstock, G. Pharmacological and molecular characterization of P2X receptors in rat pelvic ganglion neurons. Br. J. Pharmacol. 1998, 125, 771–781. [CrossRef]
- Haustein, M.D.; Kracun, S.; Lu, X.-H.; Shih, T.; Jackson-Weaver, O.; Tong, X.; Xu, J.; Yang, X.W.; O’dell, T.J.; Marvin, J.S.; et al. Conditions and Constraints for Astrocyte Calcium Signaling in the Hippocampal Mossy Fiber Pathway. Neuron 2014, 82, 413–429. [CrossRef]
- Kim, S.-H.; Bahia, P.K.; Patil, M.; Sutton, S.; Sowells, I.; Hadley, S.H.; Kollarik, M.; Taylor-Clark, T.E. Development of a Mouse Reporter Strain for the Purinergic P2X2Receptor. eneuro 2020, 7. [CrossRef]
- Grohmann, M.; Schumacher, M.; Günther, J.; Singheiser, S.M.; Nußbaum, T.; Wildner, F.; Gerevich, Z.; Jabs, R.; Hirnet, D.; Lohr, C.; et al. BAC transgenic mice to study the expression of P2X2 and P2Y1 receptors. Purinergic Signal. 2021, 17, 449–465. [CrossRef]
- Kaczmarek-Hájek, K.; Lörinczi, É.; Hausmann, R.; Nicke, A. Molecular and functional properties of P2X receptors—recent progress and persisting challenges. Purinergic Signal. 2012, 8, 375–417. [CrossRef]
- Hugel, S.; Schlichter, R. Presynaptic P2X Receptors Facilitate Inhibitory GABAergic Transmission between Cultured Rat Spinal Cord Dorsal Horn Neurons. J. Neurosci. 2000, 20, 2121–2130. [CrossRef]
- Khakh, B.S.; Gittermann, D.; Cockayne, D.A.; Jones, A. ATP Modulation of Excitatory Synapses onto Interneurons. J. Neurosci. 2003, 23, 7426–7437. [CrossRef]
- Lewis, C.; Neidhart, S.; Holy, C.; North, R.A.; Buell, G.; Surprenant, A. Coexpression of P2X2 and P2X3 receptor subunits can account for ATP-gated currents in sensory neurons. Nature 1995, 377, 432–435. [CrossRef]
- Finger, T.E.; Danilova, V.; Barrows, J.; Bartel, D.L.; Vigers, A.J.; Stone, L.; Hellekant, G.; Kinnamon, S.C. ATP Signaling Is Crucial for Communication from Taste Buds to Gustatory Nerves. Science 2005, 310, 1495–1499. [CrossRef]
- Cao, X.; Li, L.-P.; Wang, Q.; Wu, Q.; Hu, H.-H.; Zhang, M.; Fang, Y.-Y.; Zhang, J.; Li, S.-J.; Xiong, W.-C.; et al. Astrocyte-derived ATP modulates depressive-like behaviors. Nat. Med. 2013, 19, 773–777. [CrossRef]
- Mansoor, S.E.; Lü, W.; Oosterheert, W.; Shekhar, M.; Tajkhorshid, E.; Gouaux, E. X-ray structures define human P2X3 receptor gating cycle and antagonist action. Nature 2016, 538, 66–71. [CrossRef]
- Chen, C.-C.; Akopian, A.N.; Sivilottit, L.; Colquhoun, D.; Burnstock, G.; Wood, J.N. A P2X purinoceptor expressed by a subset of sensory neurons. Nature 1995, 377, 428–431. [CrossRef]
- Cook, S.P. and E.W. McCleskey, Desensitization, recovery and Ca(2+)-dependent modulation of ATP-gated P2X receptors in nociceptors. Neuropharmacology, 1997. 36(9): p. 1303-8.
- North, R.A., P2X3 receptors and peripheral pain mechanisms. J Physiol, 2004. 554(Pt 2): p. 301-8.
- Ding, S.; Zhu, L.; Tian, Y.; Zhu, T.; Huang, X.; Zhang, X. P2X3 receptor involvement in endometriosis pain via ERK signaling pathway. PLOS ONE 2017, 12, e0184647–e0184647. [CrossRef]
- Zheng, X.-B.; Zhang, Y.-L.; Li, Q.; Liu, Y.-G.; Wang, X.-D.; Yang, B.-L.; Zhu, G.-C.; Zhou, C.-F.; Gao, Y.; Liu, Z.-X. Effects of 1,8-cineole on neuropathic pain mediated by P2X2 receptor in the spinal cord dorsal horn. Sci. Rep. 2019, 9, 7909. [CrossRef]
- North, R.A., P2X receptors. Philos Trans R Soc Lond B Biol Sci, 2016. 371(1700).
- Silva-Ramos, M., et al., Activation of Prejunctional P2x2/3 Heterotrimers by ATP Enhances the Cholinergic Tone in Obstructed Human Urinary Bladders. J Pharmacol Exp Ther, 2020. 372(1): p. 63-72.
- Khakh, B.S.; North, R.A. P2X receptors as cell-surface ATP sensors in health and disease. Nature 2006, 442, 527–532. [CrossRef]
- Kawate, T.; Michel, J.C.; Birdsong, W.T.; Gouaux, E. Crystal structure of the ATP-gated P2X4 ion channel in the closed state. Nature 2009, 460, 592–598. [CrossRef]
- Qureshi, O.S.; Paramasivam, A.; Yu, J.C.H.; Murrell-Lagnado, R.D. Regulation of P2X4 receptors by lysosomal targeting, glycan protection and exocytosis. J. Cell Sci. 2007, 120, 3838–3849. [CrossRef]
- Huang, P.; Zou, Y.; Zhong, X.Z.; Cao, Q.; Zhao, K.; Zhu, M.X.; Murrell-Lagnado, R.; Dong, X.-P. P2X4 Forms Functional ATP-activated Cation Channels on Lysosomal Membranes Regulated by Luminal pH. J. Biol. Chem. 2014, 289, 17658–17667. [CrossRef]
- Jelínkova, I.; Vávra, V.; Jindrichova, M.; Obsil, T.; Zemkova, H.W.; Stojilkovic, S.S. Identification of P2X4 receptor transmembrane residues contributing to channel gating and interaction with ivermectin. Pfl?gers Arch. Eur. J. Physiol. 2008, 456, 939–950. [CrossRef]
- Buell, G.; Lewis, C.; Collo, G.; North, R.A.; Surprenant, A. An antagonist-insensitive P2X receptor expressed in epithelia and brain.. EMBO J. 1996, 15, 55–62. [CrossRef]
- Jelínková, I.; Yan, Z.; Liang, Z.; Moonat, S.; Teisinger, J.; Stojilkovic, S.S.; Zemková, H. Identification of P2X4 receptor-specific residues contributing to the ivermectin effects on channel deactivation. Biochem. Biophys. Res. Commun. 2006, 349, 619–625. [CrossRef]
- Le, K.T., K. Babinski, and P. Seguela, Central P2X4 and P2X6 channel subunits coassemble into a novel heteromeric ATP receptor. J Neurosci, 1998. 18(18): p. 7152-9.
- Seguela, P.; Haghighi, A.; Soghomonian, J.; Cooper, E. A novel neuronal P2x ATP receptor ion channel with widespread distribution in the brain. J. Neurosci. 1996, 16, 448–455. [CrossRef]
- Srivastava, P.; Cronin, C.G.; Scranton, V.L.; Jacobson, K.A.; Liang, B.T.; Verma, R. Neuroprotective and neuro-rehabilitative effects of acute purinergic receptor P2X4 (P2X4R) blockade after ischemic stroke. Exp. Neurol. 2020, 329, 113308–113308. [CrossRef]
- Montilla, A.; Mata, G.P.; Matute, C.; Domercq, M. Contribution of P2X4 Receptors to CNS Function and Pathophysiology. Int. J. Mol. Sci. 2020, 21, 5562. [CrossRef]
- Kotnis, S.; Bingham, B.; Vasilyev, D.V.; Miller, S.W.; Bai, Y.; Yeola, S.; Chanda, P.K.; Bowlby, M.R.; Kaftan, E.J.; Samad, T.A.; et al. Genetic and Functional Analysis of Human P2X5 Reveals a Distinct Pattern of Exon 10 Polymorphism with Predominant Expression of the Nonfunctional Receptor Isoform. Mol. Pharmacol. 2010, 77, 953–960. [CrossRef]
- Guo, W.; Zhang, Z.; Liu, X.; Burnstock, G.; Xiang, Z.; He, C. Developmental expression of P2X5 receptors in the mouse prenatal central and peripheral nervous systems. Purinergic Signal. 2012, 9, 239–248. [CrossRef]
- Guo, W.; Xu, X.; Gao, X.; Burnstock, G.; He, C.; Xiang, Z. Expression of P2X5 receptors in the mouse CNS. Neuroscience 2008, 156, 673–692. [CrossRef]
- Ryten, M.; Dunn, P.M.; Neary, J.T.; Burnstock, G. ATP regulates the differentiation of mammalian skeletal muscle by activation of a P2X5 receptor on satellite cells. J. Cell Biol. 2002, 158, 345–355. [CrossRef]
- Kim, H.; Kajikawa, T.; Walsh, M.C.; Takegahara, N.; Jeong, Y.H.; Hajishengallis, G.; Choi, Y. The purinergic receptor P2X5 contributes to bone loss in experimental periodontitis. BMB Rep. 2018, 51, 468–473. [CrossRef]
- Soto, F.; Garcia-Guzman, M.; Karschin, C.; Stühmer, W. Cloning and Tissue Distribution of a Novel P2X Receptor from Rat Brain. Biochem. Biophys. Res. Commun. 1996, 223, 456–460. [CrossRef]
- North, R., P2X receptors: a third major class of ligand-gated ion channels. . Ciba Found Symp, 1996. 198: p. 91-105.
- King, B.F., et al., Coexpression of rat P2X2 and P2X6 subunits in Xenopus oocytes. J Neurosci, 2000. 20(13): p. 4871-7.
- Torres, G.E.; Egan, T.M.; Voigt, M.M. Identification of a Domain Involved in ATP-gated Ionotropic Receptor Subunit Assembly. J. Biol. Chem. 1999, 274, 22359–22365. [CrossRef]
- Surprenant, A., et al., The cytolytic P2Z receptor for extracellular ATP identified as a P2X receptor (P2X7). Science, 1996. 272(5262): p. 735-8.
- Virginio, C.; MacKenzie, A.; Rassendren, F.A.; North, R.A.; Surprenant, A. Pore dilation of neuronal P2X receptor channels. Nat. Neurosci. 1999, 2, 315–321. [CrossRef]
- Cheewatrakoolpong, B.; Gilchrest, H.; Anthes, J.C.; Greenfeder, S. Identification and characterization of splice variants of the human P2X7 ATP channel. Biochem. Biophys. Res. Commun. 2005, 332, 17–27. [CrossRef]
- Adinolfi, E.; Cirillo, M.; Woltersdorf, R.; Falzoni, S.; Chiozzi, P.; Pellegatti, P.; Callegari, M.G.; Sandonà, D.; Markwardt, F.; Schmalzing, G.; et al. Trophic activity of a naturally occurring truncated isoform of the P2X7 receptor. FASEB J. 2010, 24, 3393–3404. [CrossRef]
- Di Virgilio, F.; Schmalzing, G.; Markwardt, F. The Elusive P2X7 Macropore. Trends Cell Biol. 2018, 28, 392–404. [CrossRef]
- Gelin, C.F., A. Bhattacharya, and M.A. Letavic, P2X7 receptor antagonists for the treatment of systemic inflammatory disorders. Prog Med Chem, 2020. 59: p. 63-99.
- Miras-Portugal, M.T.; Sebastián-Serrano, Á.; García, L.D.D.; Díaz-Hernández, M. Neuronal P2X7 Receptor: Involvement in Neuronal Physiology and Pathology. J. Neurosci. 2017, 37, 7063–7072. [CrossRef]
- Kopp, R.; Krautloher, A.; Ramírez-Fernández, A.; Nicke, A. P2X7 Interactions and Signaling – Making Head or Tail of It. Front. Mol. Neurosci. 2019, 12, 183. [CrossRef]
- Bartlett, R.; Stokes, L.; Sluyter, R. The P2X7 Receptor Channel: Recent Developments and the Use of P2X7 Antagonists in Models of Disease. Pharmacol. Rev. 2014, 66, 638–675. [CrossRef]
- Bhattacharya, A.; Lord, B.; Grigoleit, J.-S.; He, Y.; Fraser, I.; Campbell, S.N.; Taylor, N.; Aluisio, L.; O’connor, J.C.; Papp, M.; et al. Neuropsychopharmacology of JNJ-55308942: evaluation of a clinical candidate targeting P2X7 ion channels in animal models of neuroinflammation and anhedonia. Neuropsychopharmacology 2018, 43, 2586–2596. [CrossRef]
- Czamara, D.; Müller-Myhsok, B.; Lucae, S. The P2RX7 polymorphism rs2230912 is associated with depression: A meta-analysis. Prog. Neuro-Psychopharmacology Biol. Psychiatry 2018, 82, 272–277. [CrossRef]
- Deussing, J.M. and E. Arzt, P2X7 Receptor: A Potential Therapeutic Target for Depression? Trends Mol Med, 2018. 24(9): p. 736-747.
- Bhattacharya, A.; Biber, K. The microglial ATP-gated ion channel P2X7 as a CNS drug target. Glia 2016, 64, 1772–1787. [CrossRef]
- Bhattacharya, A.; Ceusters, M. Targeting neuroinflammation with brain penetrant P2X7 antagonists as novel therapeutics for neuropsychiatric disorders. Neuropsychopharmacology 2019, 45, 234–235. [CrossRef]
- Illes, P.; Verkhratsky, A.; Tang, Y. Pathological ATPergic Signaling in Major Depression and Bipolar Disorder. Front. Mol. Neurosci. 2020, 12, 331. [CrossRef]
- Liu, J., et al., P2X7 receptor: a potential target for treating comorbid anxiety and depression. Purinergic Signal, 2024.
- Jimenez-Pacheco, A.; Diaz-Hernandez, M.; Arribas-Blázquez, M.; Sanz-Rodriguez, A.; Olivos-Oré, L.A.; Artalejo, A.R.; Alves, M.; Letavic, M.; Miras-Portugal, M.T.; Conroy, R.M.; et al. Transient P2X7 Receptor Antagonism Produces Lasting Reductions in Spontaneous Seizures and Gliosis in Experimental Temporal Lobe Epilepsy. J. Neurosci. 2016, 36, 5920–5932. [CrossRef]
- Kaczmarek-Hajek, K.; Zhang, J.; Kopp, R.; Grosche, A.; Rissiek, B.; Saul, A.; Bruzzone, S.; Engel, T.; Jooss, T.; Krautloher, A.; et al. Re-evaluation of neuronal P2X7 expression using novel mouse models and a P2X7-specific nanobody. eLife 2018, 7. [CrossRef]
- Ortega, F.; Gomez-Villafuertes, R.; Benito-León, M.; de la Torre, M.M.; Olivos-Oré, L.A.; Arribas-Blazquez, M.; Gomez-Gaviro, M.V.; Azcorra, A.; Desco, M.; Artalejo, A.R.; et al. Salient brain entities labelled in P2rx7-EGFP reporter mouse embryos include the septum, roof plate glial specializations and circumventricular ependymal organs. Anat. Embryol. 2021, 226, 715–741. [CrossRef]
- Shibuya, I.; Tanaka, K.; Hattori, Y.; Uezono, Y.; Harayama, N.; Noguchi, J.; Ueta, Y.; Izumi, F.; Yamashita, H. Evidence that multiple P2X purinoceptors are functionally expressed in rat supraoptic neurones. J. Physiol. 1999, 514, 351–367. [CrossRef]
- Housley, G.D.; Kanjhan, R.; Raybould, N.P.; Greenwood, D.; Salih, S.G.; Järlebark, L.; Burton, L.D.; Setz, V.C.M.; Cannell, M.B.; Soeller, C.; et al. Expression of the P2X2Receptor Subunit of the ATP-Gated Ion Channel in the Cochlea: Implications for Sound Transduction and Auditory Neurotransmission. J. Neurosci. 1999, 19, 8377–8388. [CrossRef]
- Vulchanova, L.; Arvidsson, U.; Riedl, M.; Wang, J.; Buell, G.; Surprenant, A.; A North, R.; Elde, R. Differential distribution of two ATP-gated channels (P2X receptors) determined by immunocytochemistry.. Proc. Natl. Acad. Sci. 1996, 93, 8063–8067. [CrossRef]
- Xiang, Z.; Bo, X.; Oglesby, I.; Ford, A.; Burnstock, G. Localization of ATP-gated P2X2 receptor immunoreactivity in the rat hypothalamus. Brain Res. 1998, 813, 390–397. [CrossRef]
- Bhattacharya, A.; Vavra, V.; Svobodova, I.; Bendova, Z.; Vereb, G.; Zemkova, H. Potentiation of Inhibitory Synaptic Transmission by Extracellular ATP in Rat Suprachiasmatic Nuclei. J. Neurosci. 2013, 33, 8035–8044. [CrossRef]
- Lommen, J.; Stahr, A.; Ingenwerth, M.; Ali, A.A.H.; von Gall, C. Time-of-day-dependent expression of purinergic receptors in mouse suprachiasmatic nucleus. Cell Tissue Res. 2017, 369, 579–590. [CrossRef]
- Loesch, A.; Miah, S.; Burnstock, G. Ultrastructural localisation of ATP-gated P2X2 receptor immunoreactivity in the rat hypothalamo-neurohypophysial system.. J. Neurocytol. 1999, 28, 495–504. [CrossRef]
- Knott, T.K.; Velázquez-Marrero, C.; Lemos, J.R. ATP elicits inward currents in isolated vasopressinergic neurohypophysial terminals via P2X2 and P2X3 receptors. Pfl?gers Arch. Eur. J. Physiol. 2005, 450, 381–389. [CrossRef]
- Guo, W.; Sun, J.; Xu, X.; Bunstock, G.; He, C.; Xiang, Z. P2X receptors are differentially expressed on vasopressin- and oxytocin-containing neurons in the supraoptic and paraventricular nuclei of rat hypothalamus. Histochem. 2008, 131, 29–41. [CrossRef]
- Knott, T.K.; Hussy, N.; Cuadra, A.E.; Lee, R.H.; Ortiz-Miranda, S.; Custer, E.E.; Lemos, J.R. Adenosine Trisphosphate Appears to Act via Different Receptors in Terminals Versus Somata of the Hypothalamic Neurohypophysial System. J. Neuroendocr. 2012, 24, 681–689. [CrossRef]
- Xiang, Z.; He, C.; Burnstock, G. P2X5 receptors are expressed on neurons containing arginine vasopressin and nitric oxide synthase in the rat hypothalamus. Brain Res. 2006, 1099, 56–63. [CrossRef]
- Loesch, A.; Burnstock, G. Immunoreactivity to P2X6 receptors in the rat hypothalamo-neurohypophysial system: an ultrastructural study with extravidin and colloidal gold-silver labelling. Neuroscience 2001, 106, 621–631. [CrossRef]
- Gomes, D.A.; Song, Z.; Stevens, W.; Sladek, C.D. Sustained stimulation of vasopressin and oxytocin release by ATP and phenylephrine requires recruitment of desensitization-resistant P2X purinergic receptors. Am. J. Physiol. Integr. Comp. Physiol. 2009, 297, R940–R949. [CrossRef]
- Cuadra, A.E.; Custer, E.E.; Bosworth, E.L.; Lemos, J.R. P2X7 Receptors in Neurohypophysial Terminals: Evidence for their Role in Arginine-Vasopressin Secretion. J. Cell. Physiol. 2013, 229, 333–342. [CrossRef]
- Chen, Z.; Levy, A.; Lightman, S. Activation of specific ATP receptors induces a rapid increase in intracellular calcium ions in rat hypothalamic neurons. Brain Res. 1994, 641, 249–256. [CrossRef]
- Troadec, J.D., et al., ATP-evoked increases in [Ca2+]i and peptide release from rat isolated neurohypophysial terminals via a P2X2 purinoceptor. J Physiol, 1998. 511(Pt 1): p. 89-103.
- Song, Z.; Vijayaraghavan, S.; Sladek, C.D. ATP increases intracellular calcium in supraoptic neurons by activation of both P2X and P2Y purinergic receptors. Am. J. Physiol. Integr. Comp. Physiol. 2007, 292, R423–R431. [CrossRef]
- Espallergues, J.; Solovieva, O.; Técher, V.; Bauer, K.; Alonso, G.; Vincent, A.; Hussy, N. Synergistic activation of astrocytes by ATP and norepinephrine in the rat supraoptic nucleus. Neuroscience 2007, 148, 712–723. [CrossRef]
- Sladek, C.D.; Song, Z. Diverse Roles of G-Protein Coupled Receptors in the Regulation of Neurohypophyseal Hormone Secretion. J. Neuroendocr. 2011, 24, 554–565. [CrossRef]
- Day, T.A.; Sibbald, J.R.; Khanna, S. ATP mediates an excitatory noradrenergic neuron input to supraoptic vasopressin cells. Brain Res. 1993, 607, 341–344. [CrossRef]
- Hiruma, H.; Bourque, C.W. P2 purinoceptor-mediated depolarization of rat supraoptic neurosecretory cells in vitro.. J. Physiol. 1995, 489, 805–811. [CrossRef]
- Ivetic, M.; Bhattacharyya, A.; Zemkova, H. P2X2 Receptor Expression and Function Is Upregulated in the Rat Supraoptic Nucleus Stimulated Through Refeeding After Fasting. Front. Cell. Neurosci. 2019, 13, 284. [CrossRef]
- Mori, M.; Tsushima, H.; Matsuda, T. Antidiuretic Effects of ATP Induced by Microinjection into the Hypothalamic Supraoptic Nucleus in Water-Loaded and Ethanol-Anesthetized Rats. Jpn. J. Pharmacol. 1994, 66, 445–450. [CrossRef]
- Kapoor, J.R.; Sladek, C.D. Purinergic and Adrenergic Agonists Synergize in Stimulating Vasopressin and Oxytocin Release. J. Neurosci. 2000, 20, 8868–8875. [CrossRef]
- Buller, K.; Khanna, S.; Sibbald, J.; Day, T. Central noradrenergic neurons signal via atp to elicit vasopressin responses to haemorrhage. Neuroscience 1996, 73, 637–642. [CrossRef]
- Sperlagh, B.; Mergl, Z.; Juranyi, Z.; Vizi, E.; Makara, G. Local regulation of vasopressin and oxytocin secretion by extracellular ATP in the isolated posterior lobe of the rat hypophysis. J. Endocrinol. 1999, 160, 343–350. [CrossRef]
- Knott, T.K.; Marrero, H.G.; Custer, E.E.; Lemos, J.R. Endogenous ATP potentiates only vasopressin secretion from neurohypophysial terminals. J. Cell. Physiol. 2008, 217, 155–161. [CrossRef]
- Custer, E.E.; Knott, T.K.; Cuadra, A.E.; Ortiz-Miranda, S.; Lemos, J.R. P2X Purinergic Receptor Knockout Mice Reveal Endogenous ATP Modulation of Both Vasopressin and Oxytocin Release from the Intact Neurohypophysis. J. Neuroendocr. 2012, 24, 674–680. [CrossRef]
- Yoshimura, M.; Ohkubo, J.; Katoh, A.; Ohno, M.; Ishikura, T.; Kakuma, T.; Yoshimatsu, H.; Murphy, D.; Ueta, Y. A c-fos-Monomeric Red Fluorescent Protein 1 Fusion Transgene is Differentially Expressed in Rat Forebrain and Brainstem after Chronic Dehydration and Rehydration. J. Neuroendocr. 2013, 25, 478–487. [CrossRef]
- Edgar, R.S.; Green, E.W.; Zhao, Y.; van Ooijen, G.; Olmedo, M.; Qin, X.; Xu, Y.; Pan, M.; Valekunja, U.K.; Feeney, K.A.; et al. Peroxiredoxins are conserved markers of circadian rhythms. Nature 2012, 485, 459–464. [CrossRef]
- Gottlieb, H.B.; Ji, L.L.; Cunningham, J.T. Role of superior laryngeal nerve and Fos staining following dehydration and rehydration in the rat. Physiol. Behav. 2011, 104, 1053–1058. [CrossRef]
- Carreño, F.R.; Walch, J.D.; Dutta, M.; Nedungadi, T.P.; Cunningham, J.T. Brain-Derived Neurotrophic Factor-Tyrosine Kinase B Pathway Mediates NMDA Receptor NR2B Subunit Phosphorylation in the Supraoptic Nuclei Following Progressive Dehydration. J. Neuroendocr. 2011, 23, 894–905. [CrossRef]
- Lucio-Oliveira, F.; Traslaviña, G.; Borges, B.; Franci, C. Modulation of the activity of vasopressinergic neurons by estrogen in rats refed with normal or sodium-free food after fasting. Neuroscience 2015, 284, 325–336. [CrossRef]
- Yao, S.; Gourine, A.; Spyer, K.; Barden, J.; Lawrence, A. Localisation of p2x2 receptor subunit immunoreactivity on nitric oxide synthase expressing neurones in the brain stem and hypothalamus of the rat: a fluorescence immunohistochemical study. Neuroscience 2003, 121, 411–419. [CrossRef]
- Cham, J.L.; Owens, N.C.; Barden, J.A.; Lawrence, A.J.; Badoer, E. P2X purinoceptor subtypes on paraventricular nucleus neurones projecting to the rostral ventrolateral medulla in the rat. Exp. Physiol. 2006, 91, 403–411. [CrossRef]
- Ferreira-Neto, H.; Ribeiro, I.; Moreira, T.; Yao, S.; Antunes, V. Purinergic P2 receptors in the paraventricular nucleus of the hypothalamus are involved in hyperosmotic-induced sympathoexcitation. Neuroscience 2017, 349, 253–263. [CrossRef]
- Du, D.; Jiang, M.; Liu, M.; Wang, J.; Xia, C.; Guan, R.; Shen, L.; Ji, Y.; Zhu, D. Microglial P2X7 receptor in the hypothalamic paraventricular nuclei contributes to sympathoexcitatory responses in acute myocardial infarction rat. Neurosci. Lett. 2014, 587, 22–28. [CrossRef]
- Jacques-Silva, M.C.; Bernardi, A.; Rodnight, R.; Lenz, G. ERK, PKC and PI3K/Akt Pathways Mediate Extracellular ATP and Adenosine-Induced Proliferation of U138-MG Human Glioma Cell Line. Oncology 2004, 67, 450–459. [CrossRef]
- Bains, J.S. and S.H. Oliet, Glia: they make your memories stick! Trends Neurosci, 2007. 30(8): p. 417-24.
- Gordon, G.R.; Iremonger, K.J.; Kantevari, S.; Ellis-Davies, G.C.; MacVicar, B.A.; Bains, J.S. Astrocyte-Mediated Distributed Plasticity at Hypothalamic Glutamate Synapses. Neuron 2009, 64, 391–403. [CrossRef]
- Ferreira-Neto, H.C.; Antunes, V.R.; Stern, J.E. Purinergic P2 and glutamate NMDA receptor coupling contributes to osmotically driven excitability in hypothalamic magnocellular neurosecretory neurons. J. Physiol. 2021, 599, 3531–3547. [CrossRef]
- Whitlock, A.; Burnstock, G.; Gibb, A. The single-channel properties of purinergic P2X ATP receptors in outside-out patches from rat hypothalamic paraventricular parvocells. Pfl?gers Arch. Eur. J. Physiol. 2001, 443, 115–122. [CrossRef]
- Ferreira-Neto, H.C.; Antunes, V.R.; Stern, J.E. ATP stimulates rat hypothalamic sympathetic neurons by enhancing AMPA receptor-mediated currents. J. Neurophysiol. 2015, 114, 159–169. [CrossRef]
- Xu, J.; Bernstein, A.M.; Wong, A.; Lu, X.-H.; Khoja, S.; Yang, X.W.; Davies, D.L.; Micevych, P.; Sofroniew, M.V.; Khakh, B.S. P2X4 Receptor Reporter Mice: Sparse Brain Expression and Feeding-Related Presynaptic Facilitation in the Arcuate Nucleus. J. Neurosci. 2016, 36, 8902–8920. [CrossRef]
- Mori, M.; Tsushima, H.; Matsuda, T. Antidiuretic effects of purinoceptor agonists injected into the hypothalamic paraventricular nucleus of water-loaded, ethanol-anesthetized rats. Neuropharmacology 1992, 31, 585–592. [CrossRef]
- Cruz, J.C.; Bonagamba, L.G.; Machado, B.H. Modulation of arterial pressure by P2 purinoceptors in the paraventricular nucleus of the hypothalamus of awake rats. Auton. Neurosci. 2010, 158, 79–85. [CrossRef]
- Mińczuk, K.; Schlicker, E.; Krzyżewska, A.; Malinowska, B. Angiotensin 1-7 injected into the rat paraventricular nucleus of hypothalamus increases blood pressure and heart rate via various receptors. Neuropharmacology 2024, 266, 110279. [CrossRef]
- Busnardo, C.; Ferreira-Junior, N.C.; Cruz, J.C.; Machado, B.H.; Correa, F.M.A.; Resstel, L.B.M. Cardiovascular responses to ATP microinjected into the paraventricular nucleus are mediated by nitric oxide and NMDA glutamate receptors in awake rats. Exp. Physiol. 2013, 98, 1411–1421. [CrossRef]
- Ferreira-Neto, H.C.; Yao, S.T.; Antunes, V.R. Purinergic and glutamatergic interactions in the hypothalamic paraventricular nucleus modulate sympathetic outflow. Purinergic Signal. 2013, 9, 337–349. [CrossRef]
- Barad, Z.; Jacob-Tomas, S.; Sobrero, A.; Lean, G.; Hicks, A.-I.; Yang, J.; Choe, K.Y.; Prager-Khoutorsky, M. Unique Organization of Actin Cytoskeleton in Magnocellular Vasopressin Neurons in Normal Conditions and in Response to Salt-Loading. eneuro 2020, 7. [CrossRef]
- Balapattabi, K.; Little, J.T.; Farmer, G.E.; Cunningham, J.T. High salt loading increases brain derived neurotrophic factor in supraoptic vasopressin neurones. J. Neuroendocr. 2018, 30, e12639–e12639. [CrossRef]
- Sá, R.W.M.; Theparambil, S.M.; dos Santos, K.M.; Christie, I.N.; Marina, N.; Cardoso, B.V.; Hosford, P.S.; Antunes, V.R. Salt-loading promotes extracellular ATP release mediated by glial cells in the hypothalamic paraventricular nucleus of rats. Mol. Cell. Neurosci. 2022, 124, 103806. [CrossRef]
- Haam, J.; Halmos, K.C.; Di, S.; Tasker, J.G. Nutritional State-Dependent Ghrelin Activation of Vasopressin Neurons via Retrograde Trans-Neuronal–Glial Stimulation of Excitatory GABA Circuits. J. Neurosci. 2014, 34, 6201–6213. [CrossRef]
- Wei, B.; Cheng, G.; Bi, Q.; Lu, C.; Sun, Q.; Li, L.; Chen, N.; Hu, M.; Lu, H.; Xu, X.; et al. Microglia in the hypothalamic paraventricular nucleus sense hemodynamic disturbance and promote sympathetic excitation in hypertension. Immunity 2024, 57, 2030–2042.e8. [CrossRef]
- Kanjhan, R.; Housley, G.D.; Burton, L.D.; Christie, D.L.; Kippenberger, A.; Thorne, P.R.; Luo, L.; Ryan, A.F. Distribution of the P2X2 receptor subunit of the ATP-gated ion channels in the rat central nervous system. J. Comp. Neurol. 1999, 407, 11–32. [CrossRef]
- Lommen, J.; Detken, J.; Harr, K.; von Gall, C.; Ali, A.A.H. Analysis of Spatial and Temporal Distribution of Purinergic P2 Receptors in the Mouse Hippocampus. Int. J. Mol. Sci. 2021, 22, 8078. [CrossRef]
- Cipolla-Neto, J.; Skorupa, A.; Ribeiro-Barbosa, É.; Bartol, I.; Mota, S.R.; Afeche, S.; Delagrange, P.; Guardiola-Lemaitre, B.; Canteras, N. The Role of the Retrochiasmatic Area in the Control of Pineal Metabolism. Neuroendocrinology 1999, 69, 97–104. [CrossRef]
- Yamazaki, S., Y. Ishida, and S. Inouye, Circadian rhythms of adenosine triphosphate contents in the suprachiasmatic nucleus, anterior hypothalamic area and caudate putamen of the rat--negative correlation with electrical activity. Brain Res, 1994. 664(1-2): p. 237-40.
- Womac, A.D.; Burkeen, J.F.; Neuendorff, N.; Earnest, D.J.; Zoran, M.J. Circadian rhythms of extracellular ATP accumulation in suprachiasmatic nucleus cells and cultured astrocytes. Eur. J. Neurosci. 2009, 30, 869–876. [CrossRef]
- Hastings, M.H.; Maywood, E.S.; Brancaccio, M. The Mammalian Circadian Timing System and the Suprachiasmatic Nucleus as Its Pacemaker. Biology 2019, 8, 13. [CrossRef]
- McArthur, A.J.; Hunt, A.E.; Gillette, M.U. Melatonin Action and Signal Transduction in the Rat Suprachiasmatic Circadian Clock: Activation of Protein Kinase C at Dusk and Dawn*. Endocrinology 1997, 138, 627–634. [CrossRef]
- Marpegan, L.; Swanstrom, A.E.; Chung, K.; Simon, T.; Haydon, P.G.; Khan, S.K.; Liu, A.C.; Herzog, E.D.; Beaulé, C. Circadian Regulation of ATP Release in Astrocytes. J. Neurosci. 2011, 31, 8342–8350. [CrossRef]
- Burkeen, J.F.; Womac, A.D.; Earnest, D.J.; Zoran, M.J. Mitochondrial Calcium Signaling Mediates Rhythmic Extracellular ATP Accumulation in Suprachiasmatic Nucleus Astrocytes. J. Neurosci. 2011, 31, 8432–8440. [CrossRef]
- Dworak, M.; McCarley, R.W.; Kim, T.; Kalinchuk, A.V.; Basheer, R. Sleep and Brain Energy Levels: ATP Changes during Sleep. J. Neurosci. 2010, 30, 9007–9016. [CrossRef]
- Buell, G.; Collo, G.; Rassendren, F. P2X Receptors: An Emerging Channel Family. Eur. J. Neurosci. 1996, 8, 2221–2228. [CrossRef]
- Terasawa, E.; Keen, K.L.; Grendell, R.L.; Golos, T.G. Possible Role of 5′-Adenosine Triphosphate in Synchronization of Ca2+ Oscillations in Primate Luteinizing Hormone-Releasing Hormone Neurons. Mol. Endocrinol. 2005, 19, 2736–2747. [CrossRef]
- Vastagh, C.; Rodolosse, A.; Solymosi, N.; Liposits, Z. Altered Expression of Genes Encoding Neurotransmitter Receptors in GnRH Neurons of Proestrous Mice. Front. Cell. Neurosci. 2016, 10, 230. [CrossRef]
- Bjelobaba, I.; Nedeljkovic, N.; Subasic, S.; Lavrnja, I.; Pekovic, S.; Stojkov, D.; Rakic, L.; Stojiljkovic, M. Immunolocalization of ecto-nucleotide pyrophosphatase/phosphodiesterase 1 (NPP1) in the rat forebrain. Brain Res. 2006, 1120, 54–63. [CrossRef]
- Inoue, N.; Hazim, S.; Tsuchida, H.; Dohi, Y.; Ishigaki, R.; Takahashi, A.; Otsuka, Y.; Yamada, K.; Uenoyama, Y.; Tsukamura, H. Hindbrain Adenosine 5-Triphosphate (ATP)-Purinergic Signaling Triggers LH Surge and Ovulation via Activation of AVPV Kisspeptin Neurons in Rats. J. Neurosci. 2023, 43, 2140–2152. [CrossRef]
- Constantin, S.; Klenke, U.; Wray, S. The Calcium Oscillator of GnRH-1 Neurons Is Developmentally Regulated. Endocrinology 2010, 151, 3863–3873. [CrossRef]
- Bosma, M.M. Ion channel properties and episodic activity in isolated immortalized gonadotropin-releasing hormone (GnRH) neurons. J. Membr. Biol. 1993, 136, 85–96. [CrossRef]
- Koshimizu, T.-A.; Tomic, M.; Koshimizu, M.; Stojilkovic, S.S. Identification of Amino Acid Residues Contributing to Desensitization of the P2X2 Receptor Channel. J. Biol. Chem. 1998, 273, 12853–12857. [CrossRef]
- Barnea, A.; Cho, G.; Katz, B.M. A putative role for extracellular ATP: Facilitation of67copper uptake and of copper stimulation of the release of luteinizing hormone-releasing hormone from median eminence explants. Brain Res. 1991, 541, 93–97. [CrossRef]
- Zsarnovszky, A.; Bartha, T.; Frenyo, L.V.; Diano, S. NTPDases in the neuroendocrine hypothalamus: Possible energy regulators of the positive gonadotrophin feedback. Reprod. Biol. Endocrinol. 2009, 7, 63–63. [CrossRef]
- He, M.-L.; Gonzalez-Iglesias, A.E.; Tomic, M.; Stojilkovic, S.S. Release and extracellular metabolism of ATP by ecto-nucleotidase eNTPDase 1–2 in hypothalamic and pituitary cells. Purinergic Signal. 2005, 1, 135–144. [CrossRef]
- Allen-Worthington, K.; Xie, J.; Brown, J.L.; Edmunson, A.M.; Dowling, A.; Navratil, A.M.; Scavelli, K.; Yoon, H.; Kim, D.-G.; Bynoe, M.S.; et al. The F0F1 ATP Synthase Complex Localizes to Membrane Rafts in Gonadotrope Cells. Mol. Endocrinol. 2016, 30, 996–1011. [CrossRef]
- Chen, Z.P.; Kratzmeier, M.; Levy, A.; A McArdle, C.; Poch, A.; Day, A.; Mukhopadhyay, A.K.; Lightman, S.L. Evidence for a role of pituitary ATP receptors in the regulation of pituitary function.. Proc. Natl. Acad. Sci. 1995, 92, 5219–5223. [CrossRef]
- Tomić, M.; Jobin, R.M.; Vergara, L.A.; Stojilkovic, S.S. Expression of Purinergic Receptor Channels and Their Role in Calcium Signaling and Hormone Release in Pituitary Gonadotrophs. 1996, 271, 21200–21208. [CrossRef]
- Zemkova, H.; Balik, A.; Jiang, Y.; Kretschmannova, K.; Stojilkovic, S.S. Roles of Purinergic P2X Receptors as Pacemaking Channels and Modulators of Calcium-Mobilizing Pathway in Pituitary Gonadotrophs. Mol. Endocrinol. 2006, 20, 1423–1436. [CrossRef]
- Gourine, A.V.; Melenchuk, E.V.; Poputnikov, D.M.; Gourine, V.N.; Spyer, K.M. Involvement of purinergic signalling in central mechanisms of body temperature regulation in rats. Br. J. Pharmacol. 2002, 135, 2047–2055. [CrossRef]
- Klir, J.J.; McClellan, J.L.; Kluger, M.J. Interleukin-1 beta causes the increase in anterior hypothalamic interleukin-6 during LPS-induced fever in rats. Am. J. Physiol. Integr. Comp. Physiol. 1994, 266, R1845–R1848. [CrossRef]
- Alexander, V.G.; Gourine, A.V.; Dale, N.; Gourine, V.N.; Spyer, K.M. Fever in systemic inflammation: roles of purines. Front. Biosci. 2004, 9, 1011–22. [CrossRef]
- Mehta, V.B.; Hart, J.; Wewers, M.D. ATP-stimulated Release of Interleukin (IL)-1β and IL-18 Requires Priming by Lipopolysaccharide and Is Independent of Caspase-1 Cleavage. J. Biol. Chem. 2001, 276, 3820–3826. [CrossRef]
- Hide, I., et al., Extracellular ATP triggers tumor necrosis factor-alpha release from rat microglia. J Neurochem, 2000. 75(3): p. 965-72.
- Gourine, A.V.; Poputnikov, D.M.; Zhernosek, N.; Melenchuk, E.V.; Gerstberger, R.; Spyer, K.M.; Gourine, V.N. P2 receptor blockade attenuates fever and cytokine responses induced by lipopolysaccharide in rats. Br. J. Pharmacol. 2005, 146, 139–145. [CrossRef]
- Gourine, A.V.; Dale, N.; Llaudet, E.; Poputnikov, D.M.; Spyer, K.M.; Gourine, V.N. Release of ATP in the central nervous system during systemic inflammation: real-time measurement in the hypothalamus of conscious rabbits. J. Physiol. 2007, 585, 305–316. [CrossRef]
- Seidel, B.; Bigl, M.; Franke, H.; Kittner, H.; Kiess, W.; Illes, P.; Krügel, U. Expression of purinergic receptors in the hypothalamus of the rat is modified by reduced food availability. Brain Res. 2006, 1089, 143–152. [CrossRef]
- Steculorum, S.M.; Timper, K.; Ruud, L.E.; Evers, N.; Paeger, L.; Bremser, S.; Kloppenburg, P.; Brüning, J.C. Inhibition of P2Y6 Signaling in AgRP Neurons Reduces Food Intake and Improves Systemic Insulin Sensitivity in Obesity. Cell Rep. 2017, 18, 1587–1597. [CrossRef]
- Pollatzek, E.; Hitzel, N.; Ott, D.; Raisl, K.; Reuter, B.; Gerstberger, R. Functional expression of P2 purinoceptors in a primary neuroglial cell culture of the rat arcuate nucleus. Neuroscience 2016, 327, 95–114. [CrossRef]
- Wakamori, M.; Sorimachi, M. Properties of native P2X receptors in large multipolar neurons dissociated from rat hypothalamic arcuate nucleus. Brain Res. 2004, 1005, 51–59. [CrossRef]
- Steculorum, S.M.; Paeger, L.; Bremser, S.; Evers, N.; Hinze, Y.; Idzko, M.; Kloppenburg, P.; Brüning, J.C. Hypothalamic UDP Increases in Obesity and Promotes Feeding via P2Y6-Dependent Activation of AgRP Neurons. Cell 2015, 162, 1404–1417. [CrossRef]
- Sorimachi, M.; Ishibashi, H.; Moritoyo, T.; Akaike, N. Excitatory effect of ATP on acutely dissociated ventromedial hypothalamic neurons of the rat. Neuroscience 2001, 105, 393–401. [CrossRef]
- Kittner, H.; Franke, H.; Harsch, J.I.; El-Ashmawy, I.M.; Seidel, B.; Krügel, U.; Illes, P. Enhanced food intake after stimulation of hypothalamic P2Y1receptors in rats: modulation of feeding behaviour by extracellular nucleotides. Eur. J. Neurosci. 2006, 24, 2049–2056. [CrossRef]
- Burnstock, G.; Gentile, D. The involvement of purinergic signalling in obesity. Purinergic Signal. 2018, 14, 97–108. [CrossRef]
- Yu, Y.; He, X.; Zhang, J.; Tang, C.; Rong, P. Transcutaneous auricular vagal nerve stimulation inhibits hypothalamic P2Y1R expression and attenuates weight gain without decreasing food intake in Zucker diabetic fatty rats. Sci. Prog. 2021, 104. [CrossRef]
- Matsumoto, N.; Sorimachi, M.; Akaike, N. Excitatory effects of ATP on rat dorsomedial hypothalamic neurons. Brain Res. 2004, 1009, 234–237. [CrossRef]
- Kittner, H.; Franke, H.; Fischer, W.; Schultheis, N.; Krügel, U.; Illes, P. Stimulation of P2Y1 Receptors Causes Anxiolytic-like Effects in the Rat Elevated Plus-maze: Implications for the Involvement of P2Y1 Receptor-Mediated Nitric Oxide Production. Neuropsychopharmacology 2002, 28, 435–444. [CrossRef]
- Vorobjev, V.S.; Sharonova, I.N.; Haas, H.L.; A Sergeeva, O. Expression and function of P2X purinoceptors in rat histaminergic neurons. Br. J. Pharmacol. 2003, 138, 1013–1019. [CrossRef]
- Vorobjev, V.S.; Sharonova, I.N.; A Sergeeva, O.; Haas, H.L. Modulation of ATP-induced currents by zinc in acutely isolated hypothalamic neurons of the rat. Br. J. Pharmacol. 2003, 139, 919–926. [CrossRef]
- Furukawa, K.; Ishibashi, H.; Akaike, N. ATP-induced inward current in neurons freshly dissociated from the tuberomammillary nucleus. J. Neurophysiol. 1994, 71, 868–873. [CrossRef]
- Wollmann, G.; Acuna-Goycolea, C.; Pol, A.N.v.D. Direct Excitation of Hypocretin/Orexin Cells by Extracellular ATP at P2X Receptors. J. Neurophysiol. 2005, 94, 2195–2206. [CrossRef]
- Florenzano, F.; Viscomi, M.T.; Mercaldo, V.; Longone, P.; Bernardi, G.; Bagni, C.; Molinari, M.; Carrive, P. P2X2R purinergic receptor subunit mRNA and protein are expressed by all hypothalamic hypocretin/orexin neurons. J. Comp. Neurol. 2006, 498, 58–67. [CrossRef]
- Jo, Y.-H.; Role, L.W. Coordinate Release of ATP and GABA at In VitroSynapses of Lateral Hypothalamic Neurons. J. Neurosci. 2002, 22, 4794–4804. [CrossRef]
Figure 1.
Structure of anterior hypothalamus showing anterior hypothalamic nucleus (AHN), paraventricular nucleus (PVN), anteroventral periventricular nucleus (AVPV), suprachismatin nucleus (SCN), supraoptic nucleus (SON), optic chiasm (OX) and the third ventricle (3V).
Figure 1.
Structure of anterior hypothalamus showing anterior hypothalamic nucleus (AHN), paraventricular nucleus (PVN), anteroventral periventricular nucleus (AVPV), suprachismatin nucleus (SCN), supraoptic nucleus (SON), optic chiasm (OX) and the third ventricle (3V).
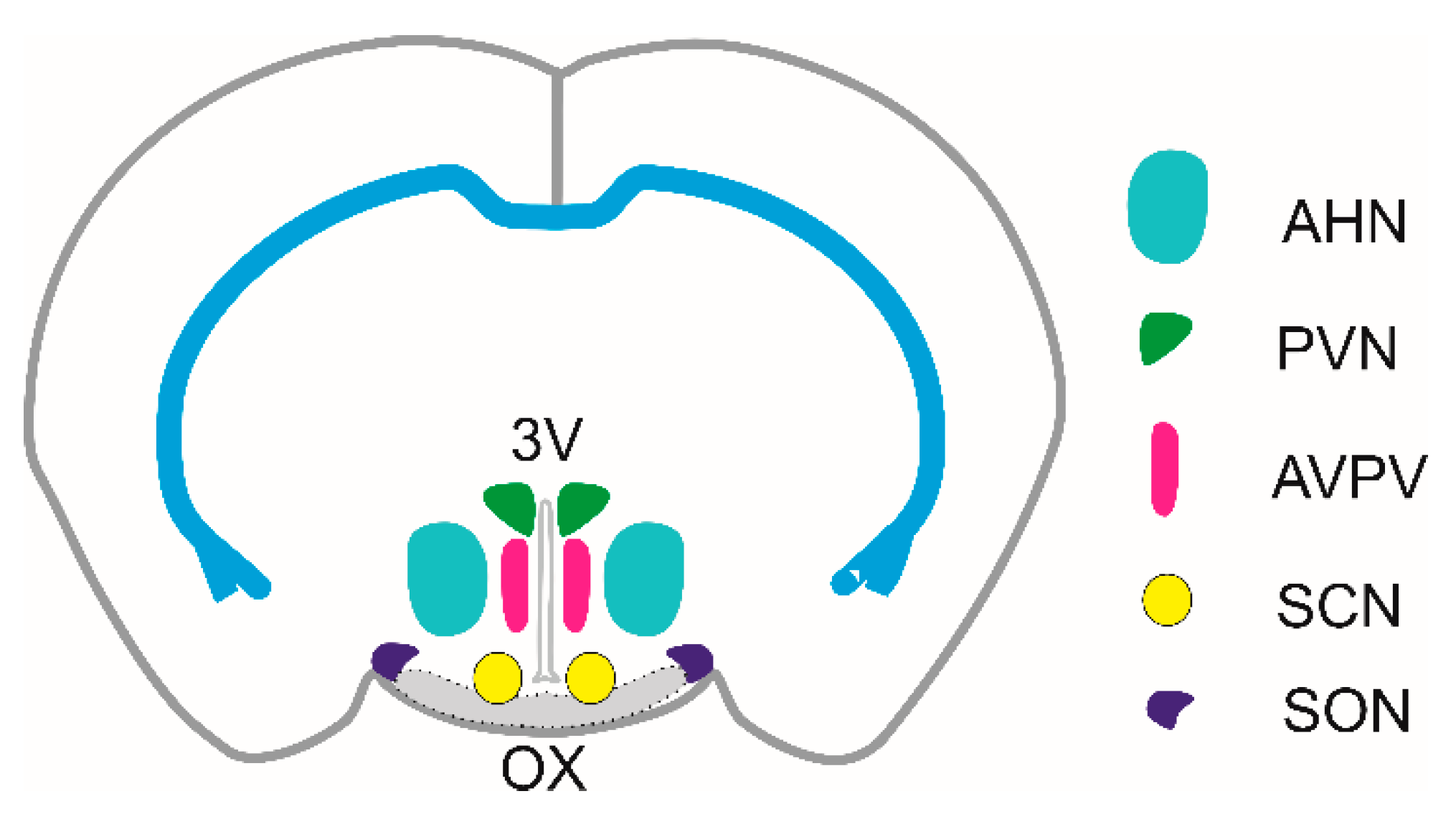
Figure 2.
A summary of the identified mechanisms of action of ATP in magnocellular AVP neurons in the SON and posterior pituitary gland. The ATP action is based on activation of P2X2 and P2X4 receptors in somata and nerve termines of AVP neurons, and P2X7 in hypothalamic astrocytes. (A) Quantitative RT-PCR analysis of seven P2X (blue columns) and three P2Y (pink columns) mRNA transcripts in hypothalamic SON tissue isolated from 16 day-old rats. P2 receptor expression was related to the expression of GAPDH as a housekeeping gene [134]. (B) Voltage clamp whole-cell recordings of ATP- induced inward current and increase in the frequency of mIPSC (upper trace) and ATP- induced increase in the frequency of mEPSCs (lower trace). (C) Current clamp whole-cell recordings of action potentials before, during and after the application of ATP. (D) Fluorescent calcium imaging of ATP-induced response in SON neuron. (E) ATP release along with AVP in neurohypophysis and subsequent P2X2 stimulation. (F) ATP- and BzATP-induced calcium response of hypothalamic astrocyte. Adatted from [134].
Figure 2.
A summary of the identified mechanisms of action of ATP in magnocellular AVP neurons in the SON and posterior pituitary gland. The ATP action is based on activation of P2X2 and P2X4 receptors in somata and nerve termines of AVP neurons, and P2X7 in hypothalamic astrocytes. (A) Quantitative RT-PCR analysis of seven P2X (blue columns) and three P2Y (pink columns) mRNA transcripts in hypothalamic SON tissue isolated from 16 day-old rats. P2 receptor expression was related to the expression of GAPDH as a housekeeping gene [134]. (B) Voltage clamp whole-cell recordings of ATP- induced inward current and increase in the frequency of mIPSC (upper trace) and ATP- induced increase in the frequency of mEPSCs (lower trace). (C) Current clamp whole-cell recordings of action potentials before, during and after the application of ATP. (D) Fluorescent calcium imaging of ATP-induced response in SON neuron. (E) ATP release along with AVP in neurohypophysis and subsequent P2X2 stimulation. (F) ATP- and BzATP-induced calcium response of hypothalamic astrocyte. Adatted from [134].
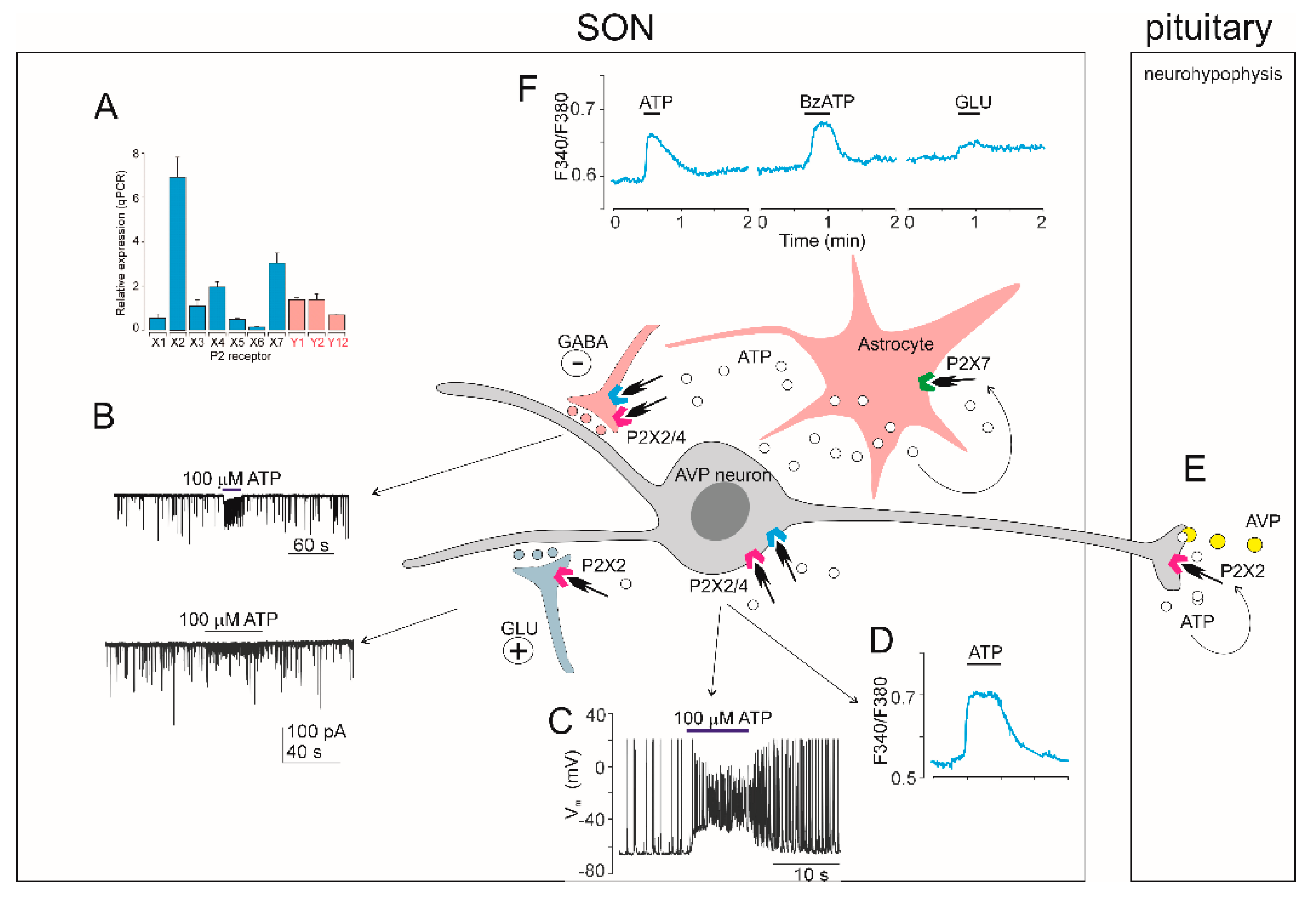
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



