
Submitted:
22 April 2025
Posted:
22 April 2025
You are already at the latest version
Abstract
The invasive aquatic macrophyte Eichhornia crassipes (water hyacinth) exhibits exceptional adaptability across a wide range of light environments, yet the mechanistic basis of its photosynthetic plasticity under both high and low light stress remains poorly resolved. This study integrates chlorophyll fluorescence and gas exchange analyses to evaluate three photosynthetic models—rectangular hyperbola (RH), non-rectangular hyperbola (NRH), and the Ye mechanistic model—in capturing light-response dynamics in E. crassipes. The Ye model provided superior accuracy (R2 > 0.996) in simulating net photosynthetic rate (Pn) and electron transport rate (J), outperforming empirical models that overestimated Pnmax by 36–46% and Jmax by 1.5–24.7% and failed to predict saturation light intensity. Mechanistic analysis revealed that E. crassipes maintains high photosynthetic efficiency in low light (LUEmax = 0.030 mol mol−1 at 200 µmol photons m−2 s−1) and robust photoprotection under strong light (NPQmax = 1.375, PSII efficiency decline), supported by a large photosynthetic pigment pool (9.46 × 1016 molecules m−2) and high eigen-absorption cross-section (1.91 × 10−21 m2). Distinct thresholds for carboxylation efficiency (CEmax = 0.085 mol m−2 s−1) and water-use efficiency (WUEi-max = 45.91 μmol mol−1 and WUEinst = 1.96 μmol mmol−1) highlighted its flexible energy management strategies. These results establish the Ye model as a reliable tool for characterizing aquatic photosynthesis and reveal how E. crassipes balances light harvesting and dissipation to thrive in fluctuating environments. Insights gained have implications for both understanding invasiveness and managing eutrophic aquatic systems.
Keywords:
aquatic macrophytes
; Eichhornia crassipes
; photosynthetic plasticity
; light stress adaptation
; photoprotection
; chlorophyll fluorescence
; mechanistic modeling
1. Introduction
Aquatic macrophytes play pivotal roles in ecosystem functioning by mediating biogeochemical cycles and creating habitat heterogeneity within aquatic environments [1]. However, their photosynthetic traits fundamentally differ from terrestrial plants due to the unique environmental constraints, including rapid light attenuation in water [2], thermal fluctuations [3], and variable nutrient bioavailability [4]. Floating macrophytes such as Eichhornia crassipes (Mart.) Solms (water hyacinth) face a distinct challenge wherein their leaves, positioned at the air-water interface, are exposed to high light at the surface while simultaneously contending with self-shading and turbidity-driven light limitation [2]. This dual pressure necessitates adaptive strategies to balance photoprotection against intense sunlight with efficient light harvesting under low-light conditions—a dynamic critical to their ecological success.
E. crassipes, a free-floating perennial monocot native to South America, exemplifies a striking ecological paradox. Introduced to China as an ornamental species in the early 20th century, it has since become a pervasive invasive plant in ponds, reservoirs, and rivers [1]. Despite its notorious capacity for rapid proliferation, which often disrupts aquatic ecosystems, E. crassipes exhibits extraordinary physiological plasticity, enabling tolerance to a wide range of environmental stressors [5,6]. Central to this adaptability is its sophisticated photosynthetic apparatus, which integrates morphological traits (e.g., large floating leaves, vertical petioles) and biochemical adjustments to optimize light capture and carbon assimilation [6,7].
Plant leaves absorb, excite, transmit, and convert light energy based on the intrinsic properties of their light-harvesting pigments, including spatial structure and charge distribution [8]. The ecological dominance of E. crassipes appears closely associated with its ability to regulate photosynthesis [6], including strategies to tolerate or avoid light stress through increased leaf area, optimized photosynthetic efficiency via chloroplast arrangement and stomatal regulation, and vertical growth for enhanced light capture [6,9,10]. Under high light, its glossy leaves minimize photodamage through reflective surfaces and thermal dissipation mechanisms, while in shaded or turbid conditions, increased pigment density and chloroplast reconfiguration enhance light absorption efficiency [6,8]. These adaptations are governed by photochemical processes in photosystem II (PSII), where spatial organization of pigments and charge separation dynamics in reaction centers determine the quantum efficiency of electron transport [11,12]. Chlorophyll fluorescence analysis, a non-invasive method for probing PSII dynamics [13], has proven instrumental in characterizing these mechanisms in terrestrial plants. Recent advances integrating gas exchange, chlorophyll fluorescence, and photosynthetic modeling have clarified photoadaptive strategies in terrestrial systems by precisely characterizing photosynthetic capacity, electron transport dynamics, and PSII down-regulation [14,15,16,17,18]. However, analogous studies on aquatic macrophytes like E. crassipes are lacking, hindering our understanding of their unique photoadaptive physiology.
While terrestrial plants prioritize sustained light utilization, aquatic species like E. crassipes must balance efficient low-light harvesting with robust photoprotection. Previous studies highlight its capacity to maintain high photosynthetic efficiency across light gradients [6,9,19], yet the photophysical mechanisms enabling this plasticity remain unresolved. This study presents a comprehensive analysis of photosynthetic performance in E. crassipes using simultaneous measurements of chlorophyll fluorescence and gas exchange parameters using LI-6400 portable photosynthesis system. The purpose is to: a) quantify key photosynthetic light response characteristics; b) evaluate the applicability of three established photosynthetic models (rectangular hyperbola, non-rectangular hyperbola, and mechanistic models) for aquatic plants; c) investigate physiological foundation for its invasive success. Our findings provide novel insights into the photobiological adaptations of aquatic macrophytes and establish methodological frameworks for analyzing aquatic photosynthesis.
2. Materials and Methods
2.1. Plant Material
Mature E. crassipes plants were used for the experiment. They grew naturally a eutrophic pond adjacent to Jinggangshan University, Ji’an City, Jiangxi Province, China (27.09°N, 115.03°E; elevation 381.6 m). The plants were in a phase of vigorous vegetative growth, reaching heights of 31-52 cm and displaying 5-8 leaves with well-developed root system.
2.2. Gas Exchange and Chlorophyll Fluorescence Measurements
Measurements were taken on clear days in July 2019, from 8:30-11:30 and 14:00-17:30, at an average daytime temperature of 36°C. In this region, photosynthetically active radiation (PAR) typically reached around 2200 μmol photons m‒2 s‒1 in summer. Five to seven biologically independent plants with uniform growth were randomly selected, with fully expanded leaves from the upper canopy designated for measurement.
Prior to measurements, selected leaves were light-adapted for one hour under natural light. Simultaneous recordings of chlorophyll fluorescence and gas exchange parameters were obtained using a LI-6400 portable photosynthesis system equipped with a 6400-40 leaf chamber fluorometer (Li-Cor INC., USA). The open-path system maintained controlled conditions, including a CO2 concentration of 380 μmol mol‒1, relative humidity of 50–70%, and air temperature within ± 1°C. A 16-step light intensity (I) gradient (2400, 2200, 2000, 1800, 1600, 1400, 1200, 1000, 800, 600, 400, 200, 150, 100, 50, and 0 μmol photons m‒2 s‒1) was applied using the embedded “Flr Light Curve” automated protocol. Each light step included a 120-180 s equilibration period, followed by automated reference/sample cell matching. Steady-state measurements of net photosynthetic rate (Pn), electron transport rate (J), stomatal conductance (gs), transpiration rate (Tr), intercellular CO2 concentration (Ci), and PSII quantum efficiency (ΦPSII), and non-photochemical quenching (NPQ) were recorded.
2.3. Photosynthesis Models and Calculations
2.3.1. Rectangular Hyperbola (RH) Model
The rectangular hyperbola model [20] is one of the classic models used to describe the photosynthetic light-response curve of plants. This model is characterized by its simplicity, minimal parameter requirements, and ease of computation. The mathematical representation of the RH model is given by:
(1)
where Pnmax represents the maximum net photosynthetic rate, α is the initial slope of Pn-I response curve, and Rd is the dark respiration rate. Additionally, this model has been employed to characterize the response of J to I.
2.3.2. Non-Rectangular Hyperbola (NRH) Model
The NRH model [21,22], by introducing a curvature parameter (θ) to correct the “convexity” between low and high light intensities, thereby improving the accuracy of light-response curve fitting. When combined with the Farquhar–von Caemmerer–Berry (FvCB) biochemical model [23], the NRH model has become one of the core frameworks for modeling plant photosynthesis. The relationship between Pn and I in the NRH model is expressed as:
(2)
where θ represents the convexity of curve, and other parameters are as previously defined. Similar to the RH model, this model has also been applied to describe the response of J to I.
2.3.3. Photosynthetic Mechanistic Model (Ye Model)
The Ye model [24,25] is a mechanistic representation of the photosynthetic process, explicitly incorporating primary photophysical and photochemical reactions, including light absorption, exciton resonance transfer, quantum energy level transitions, and de-excitation. The model integrates photochemical reactions, exciton transfer, and physiological heat dissipation mechanisms. Through statistical weighting parameters, it quantitatively describes the partitioning of absorbed energy among photochemical reactions, thermal dissipation, and fluorescence. Importantly, this model can directly account for photoinhibition effects and provides an accurate representation of photosynthesis across the entire range of light intensities, from low light to saturation and inhibition [26,27,28]. The mathematical formulation of the Ye model is given by:
(3)
where α′ represents the light energy distribution coefficient between PSII and PSI (dimensionless), β′ is the leaf light absorption coefficient (dimensionless), N0 is the number of light-harvesting pigment molecules, σik is the intrinsic light absorption cross-section of light-harvesting pigment molecules (m2), φ denotes the exciton utilization efficiency (dimensionless), η is the efficiency of photosynthetic electron transport (its reciprocal represents the number of electron required to assimilate one CO2 molecule), S represents the measured leaf area (m2). gi and gk are the energy level degeneracies of the light-harvesting pigment molecule in ground and excited states, respectively (dimensionless). ξ1, ξ2 and ξ3 are the statistical weighting factors for exciton transfer to photochemistry, heat dissipation, and fluorescence, respectively (dimensionless). kP is the photoreaction rate constant (s−1). kD is the heat dissipation rate constant (s−1). τ represents the average lifetime of the light-harvesting pigment molecule in the lowest excited state (s−1) [24,25].
According to Equation (3), Pn depends on multiple biophysical parameters, including α′, β′, N0, σik, φ, η, S, gi, gk, ξ1, ξ2, ξ3, kP, kD, and τ. Under steady-state conditions, these parameters are intrinsic to the plant species but may vary under different environmental conditions. To facilitate practical applications, three aggregate parameters are introduced to simplify Equation (3): (μmol electrons (μmol photons)−1), (m2 s (μmol photons)−1), and (m2 s (μmol photons)−1).
(4)
where αp is the initial slope of the Pn-I response curve, while βp and γp are parameters characterizing light limitation and light saturation, respectively.
The photosynthetic quantum efficiency (P′n), defined as the number of CO2 molecules fixed per photon absorbed at a given I, is derived from Equation (4) as:
(5)
The maximum Pn (Pnmax) is determined by:
(6)
while the saturation I (Isat) is given by:
(7)
Ye et al. (2013) also established the relationship between J and I in their photosynthetic mechanistic model using the following equation:
(8)
where αe is the initial slope of the J-I response curve, βe is PSII dynamics down-regulation coefficient, and γe are the saturation coefficient.
The maximum J (Jmax) is determined by:
(9)
while the saturation I (Ie-sat) is given by:
(10)
Additionally, by incorporating chlorophyll content (unit: mg m−2), Equation (8) can be used to simulate J-I curves, allowing the extraction of key traits characterizing light-harvesting pigment molecules, including the total photosynthetic pigment molecules (N0), the eigen-absorption cross-section of photosynthetic pigment molecules (σik), the minimum average lifetime of the lowest excited-state photosynthetic pigment molecules (τmin), the effective absorption cross-section of pigment molecules (σ′ik), and the total excited-state pigment molecules (Nk) [25]. Moreover, building upon the Pn-I and J-I mechanistic models, Ye and Yang et al. developed quantitative models describing the light response of light-use efficiency (LUE) [29], carboxylation efficiency (CE), intrinsic and instantaneous WUE (WUEi and WUEinst, respectively) [30], ΦPSII and NPQ [18]. These interconnected models provide a comprehensive framework for quantifying plant photosynthetic physiology, as detailed previously.
2.4. Statistical Analysis
Non-linear regression was performed to fit Pn-I, J-I, NPQ-I, ΦPSII-I, LUE-I, CE-I, WUEi-I, and WUEinst-I curves using the Photosynthesis Model Simulation Software (PMSS, Jinggangshan University) (http://photosynthetic.sinaapp.com/index.html-Chinese/English version). Goodness of fit of the three models was evaluated by the coefficient of determination (R2 = 1 - SSE/SST, where SST is the total sum of squares and SSE is the error sum of squares), Akaike’s information criterion (AIC), and Mean absolute Error (MAE = , where is the fitted values from model and is the measured values). A one-way analysis of variance (ANOVA) was conducted using SPSS Statistics 24.0 to compare differences between the model-fitted and measured values, with statistical significance set at p < 0.05. The ratio of Jmax to Pnmax can be used to estimate the apparent number of electrons required to assimilate one molecule of CO2 (na), providing an indicator of the photosynthetic electron utilization efficiency in plant leaves. Data are presented as mean ± SE (n = 4).
3. Results
3.1. Photosynthetic and Electron Transport Responses
The photosynthetic light-response dynamics of E. crassipes were rigorously assessed through coupled chlorophyll fluorescence and gas exchange analyses. Figure 1A–C illustrates the light-dependent progression of net photosynthetic rate (Pn), which exhibited a hyperbolic increase with increasing I, saturating at approximately 2000 μmol photons m‒2 s‒1 without photoinhibition. The Ye mechanistic model demonstrated superior goodness-of-fit (R2 = 0.9963, MAE = 0.40, AIC = 4.34) in replicating the Pn-I curve (Figure 1C), with no statistically significant deviation between modeled and observed Pnmax (24.64 ± 1.08 vs. 24.70 ± 1.01 μmol CO2 m−2 s−1, p > 0.05, Table 1). In contrast, both rectangular (RH) and non-rectangular hyperbola (NRH) models overestimated Pnmax by 45.7% and 36.4%, respectively (p < 0.05), and failed to yield saturation light intensity (Isat), highlighting their limitations in capturing high-light dynamics (Figure 1A-B, Table 1).
Parallel analysis of electron transport rate (J) revealed a biphasic response to I, peaking at 186.07 ± 10.04 μmol CO2 m−2 s−1, and then gradually declined, indicating dynamic downregulation of PSII activity (Figure 1D–F). The Ye model precisely simulated this trajectory (R2 = 0.9979, MAE = 2.12, AIC = 19.29), whereas RH and NRH models diverged markedly at supra-optimal light intensity, overestimating Jmax by 24.7% and 1.5%, respectively (Table 1). This also leaded to a significant overestimation of the value of apparent electron requirement for CO2 assimilation (na) in both models. However, the na value from the Ye model closely aligned with the measured (Table 1).
3.2. Quantum Yield and Photophysical Traits of Light-Harvesting Pigment Molecules
Photosynthetic quantum efficiency (P′n), reflecting the efficiency of light energy conversion, decreased from 0.023 ± 0.002 to –0.47×10−3 μmol CO2 μmol photons−1 with increasing I, indicating diminished light utilization (Figure 2A). The negative P′n at I = 2400 μmol photons m−2 s−1 indicates that respiratory CO2 release exceeded photosynthetic CO2 fixation, which was supported by the stabilization of respiratory rate in the light (RLight) at 2.97% of Pn under saturating light (Figure 2B). Mechanistic modeling of J-I curves resolved intrinsic photophysical parameters of light-harvesting pigments. As shown in Table 1, the total photosynthetic pigment pool (N0 = 9.46 ± 0.08 × 1016) and eigen-absorption cross-section (σik = 1.91 ± 0.04 × 10−21 m2) underscored robust light-capturing capacity. The minimum average lifetime of the lowest excited-state pigment molecules (τmin) was 11.53 ± 1.27 ms. The total excited-state pigment molecules (Nk) increased with I (Figure 2C), suggesting a gradual shift toward energy dissipation rather than abrupt saturation. Despite a 73.8% reduction in effective absorption cross-section (σ′ik) under high light (0.5 × 10−21 m2 at I = 2000 μmol photons m−2 s−1, Figure 2D), residual absorption capacity remained sufficient to sustain photosynthetic activity, reflecting adaptive plasticity in light harvesting.
3.3. Photoprotection and Metabolic Efficiency Dynamics
Non-photochemical quenching (NPQ) increased monotonically to 1.375 ± 0.062 at I = 2350 μmol photons m−2 s−1 (Figure 3A), consistent with enhanced thermal dissipation under high light. This photoprotective response coincided with a decline in PSII quantum efficiency (ΦPSII) from 0.762 ± 0.007 to 0.412 ± 0.011 (Figure 3B), suggesting reduced photochemical efficiency as light saturation intensified. Light-use efficiency (LUE) followed a typical peaked response, with a sharp rise at low light and a gradual decline beyond saturation at approximately 200 μmol photons m−2 s−1 (Figure 3C), while carboxylation efficiency (CE) reached its maximum 0.085 mol m−2 s−1 at I = 2200 μmol photons m−2 s−1 (Figure 3D), reflecting distinct thresholds for light capture and CO2 assimilation. Water-use metrics exhibited analogous saturation patterns, with intrinsic water-use efficiency (WUEi) plateauing at I = 1600 μmol photons m−2 s−1 (45.91 ± 6.28 μmol mol−1, Figure 3E) and instantaneous WUE (WUEinst) stabilizing earlier at I = 1400 μmol photons m−2 s−1 (1.96 ± 0.29 mmol−1, Figure 3F).
Critically, derivative models from the Ye mechanism framework (e.g., NPQ-I, ΦPSII-I, CE-I, WUEi-I, WUEinst-I) accurately simulated photoprotective and metabolic responses (R2 > 0.99) (Figure 3). Fitted NPQmax, CEmax, WUEi-max, and WUEinst-max, and their saturation light intensities, closely matched measured values (p > 0.05) (Table 2). While the ΦPSII-I model slightly underestimated ΦPSII-max, the LUE-I model only moderately captured LUE changes, exhibiting larger discrepancies compared to other parameters.
4. Discussion
4.1. Applicability of Ye Mechanistic Model in Aquatic Plants Photosynthesis
Our results demonstrate the superior performance of Ye mechanistic model in the simulating photosynthetic light-response curves of E. crassipes compared to traditional empirical models (RH, NRH), aligning with previous studies that have validated the Ye model in various plant species, including terrestrial plants [26,27,28,30,31], cyanobacteria [32], and eukaryotic algae [33]. The Ye model’s advantage lies in its detailed incorporation of both photophysical and photochemical processes. Specifically, it accounts for light absorption (σik), exciton transfer efficiency (φ), and energy dissipation dynamics (kD, τ), all critical for accurately representing aquatic photosynthesis in variable light conditions.
While the RH and NRH models provided reasonable approximations under low light, they fell short in capturing high-light saturation and the dynamic down-regulation of PSII. These models overestimated the Pnmax by 36–46% and the Jmax by 1.5–24.7% because they assume linear or hyperbolic light responses without accounting for the underlying physiological mechanisms of photosynthesis [16]. These findings align with prior critiques of empirical models in terrestrial systems [16,34,35,36], which lack the flexibility to simulate dynamic photoinhibition or species-specific photoprotective strategies. In contrast, the Ye model’s parameterization of Nk (total excited-state pigments), τmin (the minimum average lifetime of excited-state pigments), and σ′ik (effective absorption cross-section)—explains how E. crassipes rapidly dissipates excess energy as heat under high light, thereby avoiding photodamage. In addition, the close alignment between modeled and observed Jmax/Pnmax ratio (na = 7.46 vs. 7.52) in E. crassipes underscores the Ye model’s ability to quantify electron transport efficiency. This ratio reflects the balance between linear electron flow (driven by PSII activity) and carboxylation efficiency, a relationship inherently modulated by large N0 and adaptive NPQ [37]. Traditional models, which oversimplify electron transport as a static function of light, fail to resolve these interdependencies. The Ye model’s mechanistic basis thus provides a critical tool for studying aquatic plants, where environmental variability (e.g., light fluctuations, nutrient gradients) demands precise representation of energy allocation and stress responses.
4.2. Evolutionary Adaptations of E. crassipes
The photosynthetic performance of E. crassipes reveals evolutionary adaptations that differentiate it from native aquatic macrophytes. As shown in Table 3, its Pnmax and Isat exceed values reported for other aquatic plants like Nymphoides peltate, Nelumbo nucifera, and Phragmites australis [38,39,40], which show Pnmax values < 20 μmol CO2 m−2 s−1 and earlier photoinhibition thresholds. Even its photosynthetic capacity exceeds that of some common C3 plants such as Oryza sativa [41], Tamarix ramosissima [42], Solanum lycopersicum L. [43], Malus pumila Mill. [44], and Glycine max L. (Merr.) [45]. The Isat is close to C4 maize [19] and exceeds C4 sorghum [16]. These traits suggest that E. crassipes has evolved more efficient mechanisms for light absorption and energy dissipation, making it particularly competitive in nutrient-rich, high-light environments. The plant’s large photosynthetic pigment pool (9.46 × 1016 molecules m−2) and high eigen-absorption cross-section (1.91 × 10−21 m2) enhance photon absorption efficiency, which is coupled with dynamic photoprotective responses, such as NPQ, to prevent photodamage under high light. Interestingly, regional variation in photosynthetic capacity, such as the higher Pnmax observed in E. crassipes populations in Nanjing, China (32.03°N, 118.88°E; elevation 10 m) [19] and in Federal University of Lavras, state of Minas Gerais, Brazil. (19.913°S, 43.941°W; elevation 830 m) [6], indicates the species’ ability to adapt to local light conditions. This adaptability contrasts sharply with native species like N. peltate [38], which exhibit more rigid physiological responses to environmental gradients, further emphasizing the ecological flexibility of E. crassipes.

Table 3.
E. crassipes compared to other plants in photosynthetic ability.
| Plants | Pnmax | Isat | Jmax | Ie-sat | Reference |
| E. crassipes | 23.1–30.8 | ― | ― | ― | [6] |
| E. crassipes | 34.5 ± 0.72 | 2358 ± 69 | ― | ― | [19] |
| E. crassipes | 24.70 ± 1.01 | 2200.0 ± 81.7 | 186.1 ± 10.0 | 1750.0 ± 170.8 | This study |
| Nymphoides peltate | 12.66 | 219.98 | ― | ― | [38] |
| Nelumbo nucifera | 7.1–9.2 | ― | ― | ― | [39] |
| Phragmites australis | 9.0~19.5 | 924.1–2186.3 | ― | ― | [40] |
| Oryza sativa L. | 17.51–27.89 | ≈ 2000 | ― | ― | [41] |
| Oryza sativa L. (Kitaake) | 19.56 ± 0.62 | 1641 ± 32.0 | ― | ― | [19] |
| Tamarix ramosissima | 17.2–24.4 | 957–1360 | ― | ― | [42] |
| Solanum lycopersicum L. | 6.34–17.82 | ― | ― | ― | [43] |
| Malus pumila Mill. | 15.25–20.29 | 1413.8–1874.9 | ― | ― | [44] |
| Glycine max L. (Merr.) | 19.73 | 1800 | 143.51 ± 5.21 | 1601.6 ± 0.64 | [45] |
| Zea mays L. (Nongda 108) | 30.36 ± 0.42 | 2550 ± 37.0 | ― | ― | [19] |
| Sorghum bicolor L. (KFJT-4) | 37.49 ± 0.90 | 1866.7 ± 33.3 | 170.15 ± 4.45 | 1640.0 ± 74.83 | [16,18] |
| Sorghum bicolor L. (KFJT-1) | ― | ― | 133.84 ± 5.52 | 1600.0 ± 63.24 | [18] |
4.3. Synergistic Efficiency Metrics Supports Invasiveness of E. crassipes
The integration of key efficiency metrics—light-use efficiency (LUE), water-use efficiency (WUE), and carboxylation efficiency (CE)—provides a comprehensive physiological framework for understanding the invasiveness of E. crassipes. The species’ high LUEmax (0.030 mol mol−1) at low light intensity (approximately 200 µmol photons m−2 s−1), coupled with a sharp decline in LUE at higher light levels, indicates its strategic shift toward photoprotection rather than carbon fixation as light intensity increases. This plasticity enables E. crassipes to thrive under variable light conditions while maintaining efficient resource utilization. Moreover, the species exhibits superior WUE, as reflected in both intrinsic WUE (WUEi) and instantaneous WUE (WUEinst). These metrics suggest that E. crassipes effectively regulates stomatal conductance to balance CO2 uptake and water conservation, a critical strategy for survival in eutrophic environments. This efficient use of light and water, combined with the species’ high Jmax, places E. crassipes in direct competition with native macrophytes like P. australis, which exhibit lower CE and less effective photoprotective mechanisms [40]. However, under high light (>1600 μmol photons m−2 s−1), as observed in most C3 plants, E. crassipes also exhibited significant PSII dynamic down-regulation [18,46].
5. Conclusions
In conclusion, this study highlights the utility of the Ye mechanistic model in elucidating the complex photobiological adaptations that contribute to the ecological success of E. crassipes. By integrating chlorophyll fluorescence and gas exchange data, we provide a holistic view of the species’ photosynthetic performance, linking molecular-level mechanisms to its ecosystem-level invasiveness. The findings underscore the critical role of efficient light-harvesting and energy dissipation in enabling E. crassipes to dominate eutrophic, high-light environments. Future research should extend this approach to other aquatic macrophytes, particularly invasive species, to identify universal physiological traits that predict ecological success and inform management strategies in aquatic ecosystems.
Author Contributions
L.L. and X.Y. conducted the experiment and drafted the manuscript. X.Y. and Z.Y. performed the statistical analysis. X.Y., Z.Y., P. R. and S.Z. performed model calculations and statistical analysis. Funding for the research was secured by X.Y. and Z.Y. All authors critically reviewed and revised the manuscript with new data sets and contributed substantially to the completion of the present study. All authors read and approved the final manuscript.
Funding
This work was supported by the Natural Science Foundation of China (Grant No. 32260063 and 31960054).
References
- Fan, D.; Schwinghamer, T.; Liu, S.; Xia, O.; Ge, C.; Chen, Q.; Smith, D. L. Characterization of endophytic bacteriome diversity and associated beneficial bacteria inhabiting a macrophyte Eichhornia crassipes. Front. Plant Sci. 2023, 14, 1176648. [Google Scholar] [CrossRef]
- López-Pozo, M.; Adams, W. W.; Polutchko, S. K.; Demmig-Adams, B. Terrestrial and floating aquatic plants differ in acclimation to light environment. Plants 2023, 12, 1928. [Google Scholar] [CrossRef] [PubMed]
- Pilon, J.; Santamaría, L. Clonal variation in the thermal response of the submerged aquatic macrophyte Potamogeton pectinatus. J. Ecol. 2002, 90, 141–152. [Google Scholar] [CrossRef]
- Pasos-Panqueva, J.; Baker, A.; Camargo-Valero, M. A. Unravelling the impact of light, temperature and nutrient dynamics on duckweed growth: A meta-analysis study. J. Environ. Manage. 2024, 366, 121721. [Google Scholar] [CrossRef] [PubMed]
- Meneguelli-Souza, A. C.; Vitória, A. P.; Vieira, T. O.; Degli-Esposti, M. S. O.; Souza, C. M. M. Ecophysiological responses of Eichhornia crassipes (Mart.) Solms to As5+ under different stress conditions. Photosynthetica.
- Pereira, F. J.; Castro, E. M.; Oliveira, C.; Pires, M. F.; Pereira, M. P.; Ramos, S. J.; Faquin, V. Lead tolerance of water hyacinth (Eichhornia crassipes Mart. - Pontederiaceae) as defined by anatomical and physiological traits. An. Acad. Bras. Cienc. 1433. [Google Scholar]
- Downing-Kunz, M.; Stacey, M. Flow-induced forces on free-floating macrophytes. Hydrobiologia 2011, 671, 121–135. [Google Scholar] [CrossRef]
- Kühlbrandt, W.; Wang, D. N.; Fujiyoshi, Y. Atomic model of plant light-harvesting complex by electron crystallography. Nature 1994, 367, 614–621. [Google Scholar] [CrossRef]
- Ripley, B. S.; Muller, E.; Behenna, M.; Whittington-Jones, G. M.; Hill, M. P. Biomass and photosynthetic productivity of water hyacinth (Eichhornia crassipes) as affected by nutrient supply and mirid (Eccritotarus catarinensis) biocontrol. Biol. Control 2006, 39, 392–400. [Google Scholar] [CrossRef]
- Maranho, L. T.; Gomes, M. P. Morphophysiological adaptations of aquatic macrophytes in wetland-based sewage treatment systems: strategies for resilience and efficiency under environmental stress. Plants 2024, 13, 2870. [Google Scholar] [CrossRef]
- Long, S. P.; Taylor, S. H.; Burgess, S. J.; Carmo-Silva, E.; Lawson, T.; De Souza, A. P.; Leonelli, L.; Wang, Y. Into the shadows and back into sunlight: Photosynthesis in fluctuating light. Ann. Rev. Plant Biol. 2022, 73, 617–648. [Google Scholar] [CrossRef]
- Ruan, M.; Li, H.; Zhang, Y.; Zhao, R.; Zhang, J.; Wang, Y.; Gao, J.; Wang, Z.; Wang, Y.; Sun, D.; Ding, W.; Weng, Y. Cryo-EM structures of LHCII in photo-active and photo-protecting states reveal allosteric regulation of light harvesting and excess energy dissipation. Nat. Plants 2023, 9, 1547–1557. [Google Scholar] [CrossRef]
- Baker, N. R. Chlorophyll fluorescence: a probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Sun, Z.; Struik, P. C.; Gu, J. Evaluating a new method to estimate the rate of leaf respiration in the light by analysis of combined gas exchange and chlorophyll fluorescence measurements. J. Exp. Bot. 2011, 62, 3489–3499. [Google Scholar] [CrossRef] [PubMed]
- Yin, X. Y.; Struik, P. C. Theoretical reconsiderations when estimating the mesophyll conductance to CO2 diffusion in leaves of C3 plants by analysis of combined gas exchange and chlorophyll fluorescence measurements. Plant Cell Environ. 2009, 32, 1513–1524. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.-L.; Ma, X.-F.; Ye, Z.-P.; Yang, L.-S.; Shi, J.-B.; Wang, X.; Zhou, B.-B.; Wang, F.-B.; Deng, Z.-F. Simulating short-term light responses of photosynthesis and water use efficiency in sweet sorghum under varying temperature and CO2 conditions. Front. Plant Sci. 2024, 15, 1291630. [Google Scholar] [CrossRef]
- Ye, Z. P.; Kang, H. J.; An, T.; Duan, H. L.; Wang, F. B.; Yang, X. L.; Zhou, S. X. Modeling light response of electron transport rate and its allocation for ribulose biphosphate carboxylation and oxygenation. Front. Plant Sci. 2020, 11, 581851. [Google Scholar] [CrossRef]
- Yang, X.-L.; An, T.; Ye, Z.-W.-Y.; Kang, H.-J.; Robakowski, P.; Ye, Z.-P.; Wang, F.-B.; Zhou, S.-X. Modeling light response of effective quantum efficiency of photosystem II for C3 and C4 crops. Front. Plant Sci. 2025, 16, 1478346. [Google Scholar] [CrossRef]
- Xia, L.; Jianchu, Z.; Shaohua, Y.; Chenggang, R.; Man, W.; Wei, C.; Jing, S.; Puping, Z. Dynamics of photosynthesis in Eichhornia crassipes dolms of Jiangsu of China and their influencing factors. African Journal of Biotechnology 2010, 9, 7302–7311. [Google Scholar]
- Baly, E. C. C. The kinetics of photosynthesis. Proceedings of the Royal Society of London 1935, 149, 596–596. [Google Scholar] [CrossRef]
- Farquhar, G. D.; Wong, S. C. An empirical model of stomatal conductance. Aust. J. Plant Physiol. 1984, 11, 191–210. [Google Scholar] [CrossRef]
- Thornley, J. H. M. Mathematical Models in Plant Physiology: A quantitative approach to problems in plant and crop physiology (Experimental botany) London, UK: Academic Press, 1976.
- Farquhar, G. D.; von Caemmerer, S.; Berry, J. A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef]
- Ye, Z. P.; Suggett, J. D.; Robakowski, P.; Kang, H. J. A mechanistic model for the photosynthesis-light response based on the photosynthetic electron transport of photosystem II in C3 and C4 species. New Phytol. 2013, 199, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z. P. Nonlinear optical absorption of photosynthetic pigment molecules in leaves. Photosynth. Res. 2012, 112, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Martínez-García, E.; Rubio, E.; García-Morote, F. A.; Andrés-Abellán, M.; Miettinen, H.; López-Serrano, F. R. Net ecosystem production in a Spanish black pine forest after a low burn-severity fire: Significance of different modelling approaches for estimating gross primary production. Agr. Forest Meteorol. 2017, 246, 178–193. [Google Scholar] [CrossRef]
- Atsushi, S.; Tatsuya, K.; Shigeto, T.; Masashi, Y. Effect of temperature on photosynthesis characteristics in the passion fruits ‘Summer Queen’ and ‘Ruby Star’. Horticult. J. 2017, 86, 194–199. [Google Scholar]
- Liu, S.; Liu, W.; Shi, X.; Li, S.; Hu, T.; Song, L.; Wu, C. Dry-hot stress significantly reduced the nitrogenase activity of epiphytic cyanolichen. Sci. Total Environ.
- Ye, Z. P.; Zhang, H. L.; Huang, Z. A.; Yang, X. L.; Kang, H. J. Model construction of light use efficiency and water use efficiency based on a photosynthetic mechanistic model of light response. J. Plant Physiol. 2017, 53, 1116–1122. [Google Scholar]
- Ye, Z. P.; Ling, Y.; Yu, Q.; Duan, H. L.; Kang, H. J.; Huang, G. M.; Duan, S. H.; Chen, X. M.; Liu, Y. G.; Zhou, S. X. Quantifying light response of leaf-scale water-use efficiency and its interrelationships with photosynthesis and stomatal conductance in C3 and C4 species. Front. Plant Sci. 2020, 11, 374. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Yin, H.; Xiong, P.; Wan, C.; Liu, Q. Short-term responses of Picea asperata seedlings of different ages grown in two contrasting forest ecosystems to experimental warming. Environ. Exp. Bot. 2012, 77, 1–11. [Google Scholar] [CrossRef]
- Yang, X. L.; Dong, W.; Liu, L. H.; Bi, Y. H.; Xu, W. Y.; Wang, X. Uncovering the differential growth of Microcystis aeruginosa cultivated under nitrate and ammonium from a pathophysiological perspective. ACS ES&T Water.
- Yang, X. L.; Liu, L. H.; Yin, Z. K.; Wang, X. Y.; Wang, S. B.; Ye, Z. P. Quantifying photosynthetic performance of phytoplankton based on photosynthesis–irradiance response models. Environ. Sci. Eur. 2020, 32, 24. [Google Scholar] [CrossRef]
- Smyth, T. J.; Pemberton, K. L.; Aiken, J.; Geider, R. J. A methodology to determine primary production and phytoplankton photosynthetic parameters from Fast Repetition Rate Fluorometry. J. Plankton Res. 2004, 26, 1337–1350. [Google Scholar] [CrossRef]
- Silsbe, G. M.; Kromkamp, J. C. Modeling the irradiance dependency of the quantum efficiency of photosynthesis. Limnol. Oceanogr. - Meth.
- Buckley, T. N.; Diaz-Espejo, A. Reporting estimates of maximum potential electron transport rate. New Phytol. 2015, 205, 14–17. [Google Scholar] [CrossRef]
- Krall, J. P.; Edwards, G. E. Relationship between photosystem II activity and CO2 fixation in leaves. Physiologia Plantarum 1992, 86, 180–187. [Google Scholar] [CrossRef]
- Yu, H.; Niu, Y.; Hu, Y.; Du, D. Photosynthetic response of the floating-leaved macrophyte Nymphoides peltata to a temporary terrestrial habitat and its implications for ecological recovery of Lakeside zones. Knowl. Manag. Aquat. Ec. 2014, 412, 08. [Google Scholar] [CrossRef]
- Zhao, S.; Ruan, F.; Shen, W.; Deng, K.; Jiang, T.; Wu, P.; Feng, K.; Li, L. The effect of nitrogen fertilizer on rhizome quality and starch physicochemical properties in Nelumbo nucifera. In Agronomy 2022, 12, 794. [Google Scholar] [CrossRef]
- An, S.; Liu, X.; Wen, B.; Li, X.; Qi, P.; Zhang, K. Comparison of the photosynthetic capacity of Phragmites australis in five habitats in saline‒alkaline wetlands. Plants 2020, 9, 1317. [Google Scholar] [CrossRef] [PubMed]
- Xu, J. Z.; Yu, Y. M.; Peng, S. Z.; Yang, S. H.; Liao, L. X. A modified nonrectangular hyperbola equation for photosynthetic light-response curves of leaves with different nitrogen status. Photosynthetica 2014, 52, 117–123. [Google Scholar] [CrossRef]
- Tiemuerbieke, B.; Ma, J.-Y.; Sun, W. Differential eco-physiological performance to declining groundwater depth in Central Asian C3 and C4 shrubs in the Gurbantunggut Desert. Front. Plant Sci. 2024, 14. [Google Scholar] [CrossRef]
- Luo, J.; Yang, Z.; Zhang, F.; Li, C. Effect of nitrogen application on enhancing high-temperature stress tolerance of tomato plants during the flowering and fruiting stage. Front. Plant Sci. 2023, 14, 1172078. [Google Scholar] [CrossRef]
- Zheng, M.; Mu, W.; Wang, Q.; Zhang, J.; Bai, Y.; Sun, Y.; Lu, Z.; Wei, X. Case study on the effects of sodium carboxymethyl cellulose and biostimulants on physiological and photosynthetic characteristics, yield, and quality of apples. Agronomy 2024, 14, 1403. [Google Scholar] [CrossRef]
- Ye, Z. P.; Stirbet, A.; An, T.; Robakowski, P.; Kang, H. J.; Yang, X. L.; Wang, F. B. Investigation on absorption cross-section of photosynthetic pigment molecules based on a mechanistic model of the photosynthetic electron flow-light response in C3, C4 species and cyanobacteria grown under various conditions. Front. Plant Sci. 2023, 14, 1234462. [Google Scholar] [CrossRef]
- Ye, Z.-P.; An, T.; Govindjee, G.; Robakowski, P.; Stirbet, A.; Yang, X.-L.; Hao, X.-Y.; Kang, H.-J.; Wang, F.-B. Addressing the long-standing limitations of double exponential and non-rectangular hyperbolic models in quantifying light-response of electron transport rates in different photosynthetic organisms under various conditions. Front. Plant Sci. 2024, 15, 581851. [Google Scholar] [CrossRef]
Figure 1.
Light-response curves of the net photosynthetic rate (Pn) and electron transport rate (J) for E. crassipes. The curves were simulated by RH model, NRH model, and Ye Model, respectively.
Figure 1.
Light-response curves of the net photosynthetic rate (Pn) and electron transport rate (J) for E. crassipes. The curves were simulated by RH model, NRH model, and Ye Model, respectively.
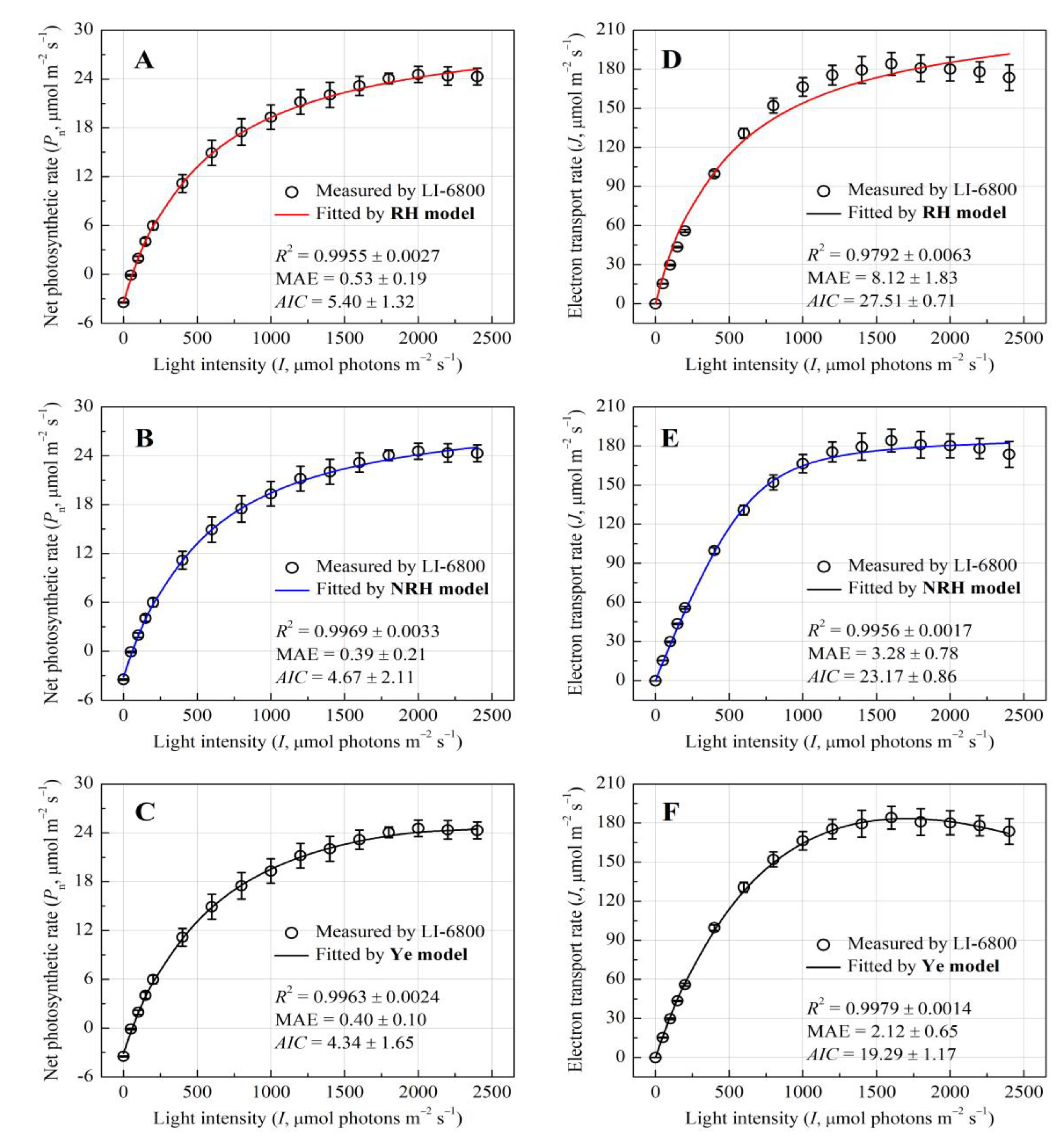
Figure 2.
Light-response curves of the photosynthetic quantum efficiency (P′n), respiratory rate in the light (RLight), total excited-state pigment molecules (Nk) and effective absorption cross-section (σ′ik) for E. crassipes.
Figure 2.
Light-response curves of the photosynthetic quantum efficiency (P′n), respiratory rate in the light (RLight), total excited-state pigment molecules (Nk) and effective absorption cross-section (σ′ik) for E. crassipes.
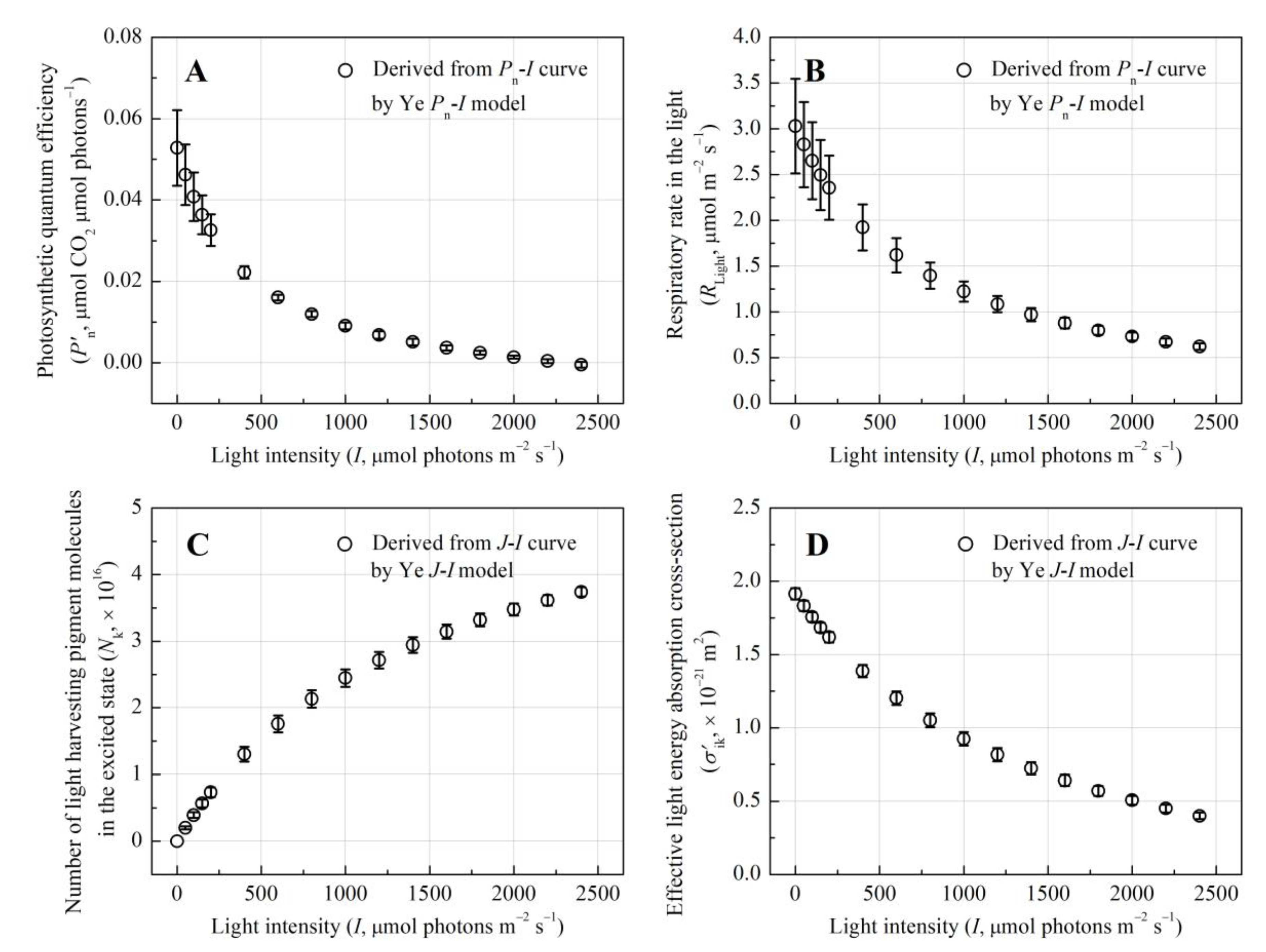
Figure 3.
Light-response curves of the non-photochemical quenching (NPQ), PSII quantum efficiency (ΦPSII), light-use efficiency (LUE), carboxylation efficiency (CE), and intrinsic and instantaneous water-use efficiency (WUEi and WUEinst, respectively) for E. crassipes.
Figure 3.
Light-response curves of the non-photochemical quenching (NPQ), PSII quantum efficiency (ΦPSII), light-use efficiency (LUE), carboxylation efficiency (CE), and intrinsic and instantaneous water-use efficiency (WUEi and WUEinst, respectively) for E. crassipes.
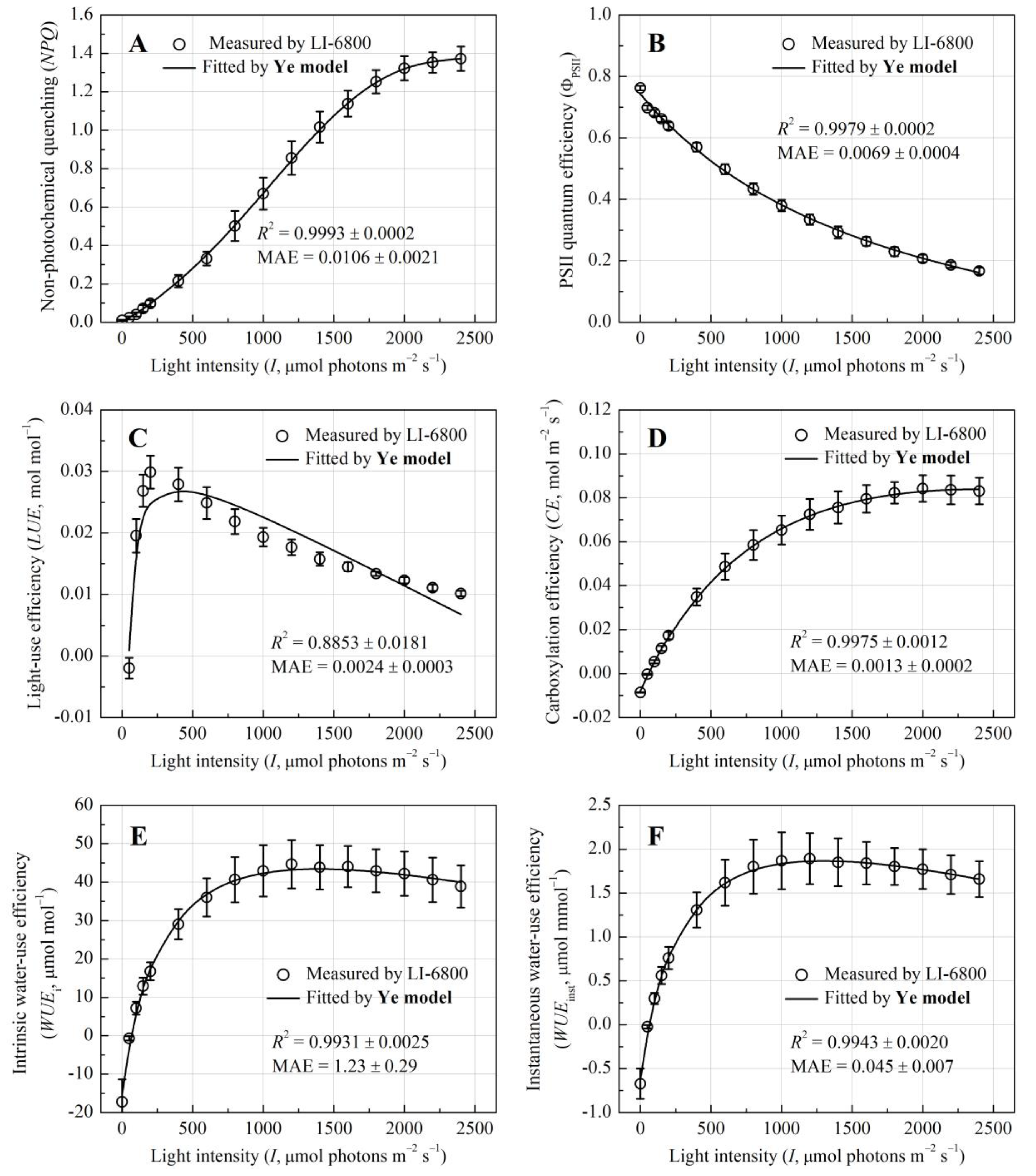
Table 1.
Fitted and measured values of traits defining Pn–I and J–I curves of E. crassipes. Values are means ± SE (n = 4). Within each row, values with different letters are significantly different (p < 0.05).
Table 1.
Fitted and measured values of traits defining Pn–I and J–I curves of E. crassipes. Values are means ± SE (n = 4). Within each row, values with different letters are significantly different (p < 0.05).
| Traits | Fitted value | Measured value | ||
| RH Model | NRH Model | Ye Model | ||
| αp (μmol mol−1) | 0.0659 ± 0.0111a | 0.0522 ± 0.0058a | 0.0528 ± 0.0107a | ― |
| Pnmax (μmol m−2 s−1) | 35.93 ± 1.33a | 33.70 ± 1.65a | 24.64 ± 1.08b | 24.70 ± 1.01b |
| Isat (μmol m−2 s−1) | ― | ― | 2520.41 ± 243.03a | 2200.00 ± 81.65a |
| Ic (μmol m−2 s−1) | 58.78 ± 3.59a | 60.02 ± 3.33a | 54.17 ± 2.45a | 53.06 ± 2.11a |
| Rd (μmol m−2 s−1) | 3.41 ± 0.46a | 2.91 ± 0.24b | 3.46 ± 0.16a | 3.46 ± 0.06a |
| αe (μmol mol−1) | 0.4658 ± 0.0109a | 0.2779 ± 0.0052c | 0.3424 ± 0.0076b | ― |
| Jmax (μmol m−2 s−1) | 232.08 ± 16.78a | 188.85 ± 11.66b | 184.10 ± 10.84b | 186.07 ± 10.04b |
| Ie-sat (μmol m−2 s−1) | ― | ― | 1699.64 ± 40.39a | 1750.00 ± 170.78a |
| na | 6.45 ± 0.34b | 5.62 ± 0.31b | 7.46 ± 0.12a | 7.52 ± 0.12a |
| σik (10−21 m2) | ― | ― | 1.91 ± 0.04 | ― |
| τmin (ms) | ― | ― | 11.53 ± 1.27 | ― |
| N0 (1016 m2) | ― | ― | 9.46 ± 0.08 | ― |
| Chl content (mg m−2) | ― | ― | ― | 707.34 ± 5.86 |
Table 2.
Fitted and measured values of photoprotection and physiological traits of E. crassipes. Values are means ± SE (n = 4). Within each row, values with different letters are significantly different (p < 0.05).
Table 2.
Fitted and measured values of photoprotection and physiological traits of E. crassipes. Values are means ± SE (n = 4). Within each row, values with different letters are significantly different (p < 0.05).
| Traits | Fitted value | Measured value |
| NPQmax | 1.366 ± 0.058a | 1.375 ± 0.062a |
| INPQ-sat (μmol m−2 s−1) | 2278.76 ± 41.25a | 2350.00 ± 50.00a |
| ΦPSIImax | 0.743 ± 0.007b | 0.762 ± 0.007a |
| LUEmax (mol mol−1) | 0.027 ± 0.002a | 0.030 ± 0.003a |
| ILUE-sat (μmol m−2 s−1) | 384.65 ± 17.60a | 250.00 ± 50.00b |
| CEmax (mol m−2 s−1) | 0.084 ± 0.006a | 0.085 ± 0.006a |
| ICE-sat (μmol m−2 s−1) | 2242.99 ± 82.55a | 2000.00 ± 81.65b |
| WUEi-max (μmol mol−1) | 44.17 ± 6.09a | 45.91 ± 6.28a |
| Ii-sat (μmol m−2 s−1) | 1621.82 ± 267.39a | 1500.00 ± 173.21a |
| WUEinst-max (μmol mmol−1) | 1.88 ± 0.28a | 1.96 ± 0.29a |
| Iinst-sat (μmol m−2 s−1) | 1391.42 ± 139.03a | 1300.00 ± 173.21a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



