
Submitted:
17 April 2023
Posted:
18 April 2023
You are already at the latest version
Abstract
The Yellow-legged Hornet (Vespa velutina nigrithorax) (YLH) is an invasive insect that arrived in Europe in 2004 and is now spread across nine countries. It is a threat to the native entomofauna and harmful to beekeeping and agriculture, as it is a ravenous predator of the European Honey Bee (Apis mellifera) and other pollinating species. Its expansion has been unstoppable and all resources are needed to fight against it, including native vertebrate predators. Among these, the European Bee-eater (Merops apiaster) (EBE) is a potential one, but little is known about its predation on YLH. In a study carried out in Portugal, remains of YHL were detected in EBE nesting sites, which to the best of our knowledge is the first such report. This means that this bird could be one more agent in the biological control of this pest (although research on predation intensity is still needed), in conjunction with other natural predators and other strategies. In the Iberian Peninsula both species are allopatric in vast regions, so the role of EBE may be more limited. However, in the rest of Europe, at a country or continent scale, the scenario may be different and sympatry may occur to a greater extent.
Keywords:
yellow-legged hornet
; European bee-eater
; biological control
; predation
; sympatry
1. Introduction
The Yellow-legged Hornet (Vespa velutina) (YLH), originates from Southern, Eastern and South-eastern Asia [1,2], is an insect that has become invasive in the European continent and is very concerning in terms of economic and biodiversity damages. It is represented by the subspecies V. v. nigrithorax, which is native to Nepal, India, Bhutan, Vietnam and China [2]. The YLH first arrived in Europe via France around 2004 and then expanded to several other European countries, namely Spain in 2010, Portugal and Belgium in 2011, Italy in 2012, Germany in 2014, UK in 2016, Netherlands and Switzerland in 2017 and more recently Luxembourg, in 2020 [3,4,5].
In Europe the YLH has been found to cause much negative economic impacts on beekeeping (predation on honey bees and losses of swarms, with consequent damages in the production of honey and other bee products), and on agriculture (decline of crops pollination, and damage in orcharding and other fruit plantations) [6,7,8,9,10]. Impacts on biodiversity (predation over many native insect species, including other Apidae, Vespidae and insect pollinators; capture and collateral destruction of dozens of non-target native insect species in non-selective traps), and possibly on ecosystem functions and services (e.g. pollination declining of wild and domestic plants, and changes in trophic chains) are also referred [6,8,9,10,11,12].
Several researchers claim that it is now impossible or nearly impossible to eradicate the YLH [7,9,13], which will continue to spread in the countries where it is already present [10,14,15], and that in the long term or even in a few years it will very likely spread over most of the rest of Central, Eastern and Southeastern Europe [1,13,16,17,18,19]. Even with the application of the current methods of destruction of the exotic hornet by the authorities and beekeepers (surveillance, use of baited traps, location and destruction of nests), the spread has been unstoppable [9,20,21]. Thus, all resources are needed to fight against this pest, in order to reduce the density of its populations in areas it has invaded and occupied, and this should include potential natural predators.
Several species of wild animals in Europe are known to feed on wasps and some even capture the large European Hornet (Vespa crabro). However, very often, published studies on the feeding habits of insect predators do not discriminate prey at the species or genus level, and sometimes not even at the family level. So, references to the capture of wasps of the genus Vespa by a specific predator are not very common.
Wasp predators are mostly birds, but there are also references to several mammal species preying on or opportunistically consuming wasps [7,22,23]. For example, in Europe, the European Badger (Meles meles) is known to dig the secondary nests of social wasps in the soil, eating larvae, adults, combs and the envelope of the nests [24,25], in summer or early autumn [26]. The Badger is probably the main wasp predator among mammals [24]. According to Edwards [25], moles (Talpa spp.) also destroy some wasp nests, although in small numbers, and some species of mice, such as the Wood Mouse (Apodemus sylvaticus) or the House Mouse (Mus musculus) may also eat or destroy wasp queens existing in the soil, when they are hibernating. In turn, van Bergen [27] detected European Pine Marten (Martes martes) destroying a wasp nest, possibly of Dolichovespula sylvestris, and Spradbery [24] also mentions other Mustelidae - Stoat (Mustela erminea) and Least Weasel (M. nivalis) -, which, as well as the Wood Mouse, disrupt colonies that are still incipient in mouse burrows in the ground.
Predation or destruction of European Hornet nests by the European Badger and Red Fox (Vulpes vulpes), and probably also by the Pine Marten or Beech Marten (Martes foina), was recorded by Nadolski [28]. The Common Wall Gecko (Tarentola mauritanica) has also been observed successfully hunting the European Hornet in Sardinia, Italy [29]. However, since the vast majority of YLH nests are located above ground, mostly in trees [30,31,32,33,34], the impact of mammals on this hornet is likely much smaller.
As far as birds are concerned, there is abundant scientific documentation attesting their predation on a large number of species of Hymenoptera and, among these, on Vespidae. The vast majority of species that consume wasps are generalists - insectivores or omnivores -, capturing them at random, but a very restricted number of species are specialized in Hymenoptera or even in Vespidae.
Regarding the bird species known to feed on the European Hornet, we found references to the European Bee-eater (Merops apiaster) (EBE) [35,36,37,38,39,40], European Honey Buzzard (Pernis apivorus) [27,41,42], Iberian Grey Shrike (Lanius meridionalis) [43], Red-backed Shrike (Lanius collurio) [44], Eurasian Jay (Garrulus glandarius) (Stachanoff cit. in Spradbery [24], and Eurasian Magpie (Pica pica) [45]. Since these birds are able to catch hornets such as the European Hornet, they are also likely potential predators of the YLH, as it is slightly smaller and its nests are mostly out in the open, in trees and bushes [30,31,32,33,34].
The Iberian Grey Shrike [46], the European Honey Buzzard [47,48,49,50] and the EBE [51] are among the bird species for which predation on the YLH has already been confirmed. Mollet & de la Torre [30], Villemant et al. [52] and Rome & Villemant [15] (authors from which most subsequent articles are based when the potential predators of the YLH are mentioned, although they do not refer to dates, locations or eventual original sources of information), also report that woodpeckers (e.g., Picus sp., Dryobates minor), Magpie (Pica pica) and tits such as the Eurasian Blue Tit (Cyanistes caeruleus) were observed in late autumn/early winter pecking the nest envelope to consume the last remaining individuals, larvae or torpid adults, of the respective declining colonies. Additionally, Bunker [5], quoting F. Muller (pers. comm.), states that "There have been rare records of more specialist enemies such as the European honey buzzard (Pernis apivorus) and the European bee-eater (Merops apiaster), but these have a negligible impact on the population". However, here again, information is not accurate, for the reasons mentioned immediately above and because, so far, we have found at least four articles that accurately refer or document YLH predation by the European Honey Buzzard - two of them published in scientific journals [48,49]. With the exception of our own work [51], there is a lack of information on EBE predation on this pest (Figure 1).
However, the opinion concerning predation importanceof the Honey Buzzard is not consensual, namely from the Spanish researchers who found YLH breeding combs in buzzard nests. In fact, Macià et al. [48] refer that this bird of prey should be considered as a potential biocontrol agent since it is capable of destroying active nests of YLH, and, at the same time; Rebollo et al. [49] report that the Honey Buzzard population in Galicia (the largest and second densest one in Spain [53]) could destroy thousands of YLH nests every year, that is equivalent to the number of nests destroyed by the authorities' control teams (S. Rebollo cit. in Anonymous [54].
The contribution of each of the species mentioned above in the natural limitation of the expansion of YLH, as well as of other birds that consume Vespidae, should not be significant. However, EBE and Honey Buzzard may be the exception, as they are specialists, or more specialized, in Hymenoptera and Vespidae, respectively [41,55,56,57,58,59]. Therefore, these ones are the most important potential enemies of YLH among wild vertebrates, and so they have the potential to contribute to the biological control of this invasive pest in a more significant way. For this reason, and because there was little information about the predation by these two species on the YLH, mainly in what concerns the EBE and for which, till then, nothing had been published on this matter, the National Institute for Agrarian and Veterinary Research (INIAV) started a study on the diet of this latter species to assess the relative importance of its predation on YLH.
The EBE's diet in Europe is largely composed of European Honeybee (Apis mellifera) and bumblebees (Bombus sp.), with prey captured usually in the air [58]. The European Hornet is also part of its diet, but it is an uncommon prey species [36,37,39,60]. In a set of 38 studies on the EBE diet, Vespidae represented, on average, around 8-9% of the total prey, only reaching 30-40% in 4 of these studies [58], whereas in Spain the percentages of Vespids do not go beyond 2% [61,62,63]. In Portugal, according to Costa et al. [64], the composition of the EBE's diet varies according to the country's regions, but is mainly composed of Hymenoptera (the majority) and Coleoptera, which totalize approximately 90% of the items consumed in the breeding season (adults and nestlings). Concerning only the nestlings, Hymenoptera constitute 68% to 85% of their diet wherein –European bees corresponding to 26-47%, wasps ~3-33% and other Hymenoptera 21-30% [64].
However, the importance of predation also depends on other factors, such as the geographic coexistence between both species, predator and prey, and although the potential area of occurrence of YLH in Europe (see Verdasca et al. [19] or GBIF Secretariat [65], among others), coincides very reasonably with the distribution range of the European Honey Buzzard [66,67], the same does not seem to happen with the EBE, at least in the present and in certain regions of Europe [67,68].
2. Materials and Methods
The occurrence areas of the YLH and the EBE in Portugal are somewhat disjointed [69,70], so the study area was located in the region where these areas overlap. The study area comprised the counties of the district of Setúbal, that are located on the left bank of the Tagus River in front of the city of Lisbon, and the counties of the districts of Santarém and Portalegre bordering the same river (Figure 2).
Search for nesting sites started in March of 2021, with many nesting sites located after information from fellow ornithologists. Pellets and prey remains were collected from May to September and once by month.
In nesting sites where it was difficult to collect food material, due to the inaccessibility of the nests or the absence of perches, artificial perches (dry branches) were placed nearby in order to increase samples pellet, which was successfully achieved. Most of the material was collected under perches, but some was also collected inside nest tunnels or on the ground below its entrances after fledging. After collection, each pellet was stored in an individual bag and, together with the remains of prey found (Figure 3), stored in a larger bag corresponding to the respective nesting site. This bag was later stored in a freezer until it was analyzed.
Since bee-eaters rarely take again a fallen prey that was carried in the beak (Koenig & Ursprung cit. in Bastian & Bastian [58], it is rather easy to find a fair number of whole insects captured by these birds near their nests and roosting sites.
The pellets were kept for further analysis by molecular genetic methods, in order to determine the frequency of occurrence of YLH as a food item and its importance in the EBE's diet, and therefore the impact that this bird may have in limiting this very harmful exotic hornet to the beekeeping activity.
Prey remains were identified with the aid of a 20X - 40X stereomicroscope, insect field guides (e.g. Chinery [71]) and, whenever necessary, with the expertise of fellow entomologists at INIAV. They were processed and identified considering the site and date they were collected.
3. Results
At present, only a first identification of prey remains collected in 2021 has been conducted. Thus, only preliminary results on prey captured by this bee-eater are presented here.
Most of the pellets and prey remains were collected in June and July, when the birds were feeding their young. Few of them were collected in May, when the birds were incubating their eggs, as well as in August/September, when juveniles and adults forage far from nesting sites and spend little time there and begin their autumnal migration.
Out of a total of 30 nesting sites located (Figure 2), only in 13 food material was collected. This is because some colonies did not become active that year, or because nesting sites with few nests were abandoned due to breeding failure early in the season, or because they were completely inaccessible.
Table 1 shows the families, genera and species of a first set of insects that were identified among the remains found on the ground below the nest tunnels and perches of the EBE nesting sites surveyed.
One of the main objectives of the present study was to study YLH as Bee-eater prey. To this end, we found prey remains parts of 5 specimens of YLH, collected in 4 different nesting sites (Figure 2). Except for the two nearby places in the region of Tomar, the other places where the YLH was found were more than 20 km (Abrantes region) and 50 km (Santarém region) away from the first two. The parts of the hornet that were found are a 1st abdominal segment, a complete individual, an individual without a head, an isolated head and finally a mandible and two abdominal segments (1st and 2nd).
In addition to the YLH, a complete specimen of European Hornet and more than a dozen specimens of the large Mammoth Wasp (Megascolia maculata) were also found (Table 1). The latter wasp and bumblebees (Bombus sp.) were the insects most frequently found among prey remains after the European Honey Bee (Apis mellifera) and, so far, also after some smaller wasp species such as Polistes sp. or Vespula sp.
Among the remains of prey, the Hymenoptera seem to be the most important prey group of the studied bee-eaters, followed by the Coleoptera and within this order, the genera Cetonia and Protaetia, of the Scarabaeidae family, are the most captured beetles.
The presence of the Red Palm Weevil (Rhynchophorus ferrugineus) among the remains of prey is noteworthy, as it is yet another exotic insect from tropical Asia and Oceania and has also become a pest in many countries, including Portugal. It is responsible for the destruction of thousands of palm trees [72].
4. Discussion
The findings of several Yellow-legged Hornet individuals caught by the Bee-eater in Central Portugal [51], provide evidence that this pest is also a prey of this bird in Europe. To understand the importance of this predatory activity is outside the scope of the present work but will be followed up on in due course. The capture of the YLH was expected, as the EBE has always hunted wasps and among these, although in small numbers, the great European Hornet [36,37,40,60,62] - an even larger hornet -, and also because this exotic hornet is expanding territorially [73]. The data presented here are still very incipient and caution is needed in extrapolating from them. In any case, it could be that the greater number of YLH individuals found among prey remains than that of European Hornet, may indicate either a greater availability of the first species or that it is easier to capture. In both cases, this could suggest that EBE may have a role in the biological control of YLH.
With regard to the specific effectiveness of the EBE in capturing and feeding on YLH, it is important to know how much this hornet contributes to the diet of the insectivorous bird. Work is currently underway to further understand this predation potential and this will require another year of collecting material on the ground. Yet, no matter how much YLH the EBE captures, this effort will only be effective in areas where both species, prey and predator, cohabit. Indeed, contrary to what happens with the European Honey-buzzard, a predator specialized in Vespidae and which in France, Spain and Portugal has already been confirmed to feed on combs from YLH nests [47,48,49,50], and whose distribution ranges in Europe, both in the present and in future predictions, seem to overlap reasonably [17,19,65,66,67], the same does not seem to occur always with EBE.
The two species, the YLH and the EBE, seem to have somewhat different climate niches, with the YLH preferring wetter and fairly cooler regions [10,19,21,32,74,75], while the EBE, as a thermophilic species, prefers regions of greater aridity, that is, hotter and drier [56,58,76,77,78,79,80,81]. For this reason, on a country-wide scale, the current area of sympatry between the two species does not appear to be particularly large, namely in the Iberian Peninsula (IP), as illustrated in Figure 4, which, despite some flaws (e.g. in England), reasonably reflects the distribution of the invasive wasp [19,73,82] and the EBE [67] in Europe. Presently, in Portugal, the overlap between the areas of occurrence of the two species is not very large, as already mentioned, and in Spain the disjunction seems to be much more evident (cf. Figure 4). Indeed, most of risk, adequacy or general favorability models in general, which address the expansion of the YLH and cover Portugal, Spain or IP as a whole [4,13,16,17,19,32,73,74], suggest that almost half of the Portuguese territory (i.e., most of the provinces of Alentejo and Algarve, in the south, and some parts of the east, north of the Tagus River), and much of Spanish territory (e.g., the autonomous communities of Extremadura, Andalusia, Castilla-La Mancha and Murcia, located in the center and south of this country) [21,32], will have low or medium-low suitability for YLH. Paradoxically, these regions, where the probability of expansion or occurrence of the YLH is low, very low or even null due to climatic inadequacy [13,17,19,32,73,74], are exactly those where most of the EBE populations are traditionally located and where its populations are strongest [70,83,84,85,86,87]. It is true that in the future, in the medium and long term (2030 to 2100), the predictions point to an increase, not very accentuated, of the climate suitability for YLH in these regions of the IP (smaller in the referred areas of Spain) [16,17,20], and a slight expansion of the EBE northwards into the Eurosiberian region of the IP by 2040, 2050 and 2100 [81,88], an expansion that has been observed since the late 1990s [86,87]. However, it does not seem that there will be a significant increase in the overlapping of the ranges of the two species.
With regard to France, Figure 4 also shows that there seems to be some spatial segregation between YLH and EBE, except in the south in the regions of Occitania and Provence-Alpes-Côte d'Azur and in Rhône-Alpes, where coexistence it is more consistent. However, contrary to what happens in IP, both the YLH and the EBE are present in almost all the France, with the hornet occurring in all but one of its 96 departments (i.e., except Haut-Rhin) [15,89] and the EBE being present or seen in almost all French departments, breeding in all the 13 major administrative regions [67,90,91], although significantly more rare and irregular in the north and northwest of the country, in the Center region and the central Massif [58,92], as evidenced, moreover, in the cartography [67,90,91]. However, there is a greater spatial coincidence at the regional scale in France than in IP, or a greater probability of joint occurrence of the two species, and this overlap in YLH and EBE distribution ranges is expected to increase further in the future due to climate change. Indeed, Stiels et al. [81] forecast a significant increase in the range and on the probability of occurrence of the EBE in a large part of France in 2050, which is predicted to be medium to high in much of its territory, at the same time that the climate suitability models for the YLH, either for the short term [13,17,19], or for the medium-long term - 2030 to 2100 [16,17,20], predict equal or greater likelihood of YLH occurrence in almost all of France. That is, in the medium-long term, the probability of coexistence of the two species in France at the local scale will be much greater and greater than in the IP.
In the rest of Europe, the same climate suitability models point, in general, to an expansion of YLH in a large number of countries, such as in Germany, Belgium, the Netherlands, Switzerland and Luxembourg, the United Kingdom and Italy where it has already arrived or settled [15,82] or in the east and southeast of this continent, where it is expected to settle in the medium-long term [13,16,17,19,20]. In turn, the EBE, which since the 1980s and 1990s has been expanding to the north and its populations have been growing in several European countries [58,79], is present in all those countries, although in most of them their populations are in generally still small or nesting is still irregular (e.g. UK, The Netherlands, Luxembourg or Belgium) [58,67,93]. This northward expansion across the European continent, including France, has been largely attributed to climate changes and an increase in food availability as a result of these [58,77,78,79,80,81,82,83,84,85,86,87,88,89,90]. In Germany, where the population has grown unparalleled in Central Europe, the EBE is present in almost all country and federal states, i.e., with the exception of the 3 urban states of Berlin, Hamburg and Bremen [90,93]. In Italy, as is typical in a Mediterranean European country, the EBE population is even stronger and is distributed over most of the territory [58,67]. Currently, EBE populations in countries of Central and Northern Europe are far from having such a wide national distribution as in countries of Southern and Southeast Europe [67], but predictions for the future (2050) point to a significant increase in the probability of occurrence of this bird in Central Europe, Northern Europe and Eastern Europe [1,81], and consequently in its territorial and population expansion. That is, the probability of spatial coexistence of the two species, hornet and Bee-eater, in these regions is once again quite reasonable, at least on a regional scale, and, once again, higher than could happen in IP. On the other hand, since the availability of large insects is an important factor for the establishment of the EBE in new territories [90], expansion of YLH may contribute the expansion of the bird. In the eventuality, as is expected [19], that the YLH expands further eastwards, especially towards Southeastern Europe, where climate suitability for this insect is expected to be largely medium to high [17,19], the possibility of territorial coexistence of both species in this part of the European continent is quite reasonable, since the EBE is currently widely distributed across the territories of these countries [58,67], and tends to be even more common in the future [81].
However, despite the existence of correspondence between their distribution ranges on a continental or national scale, the potential contribution of the EBE in the biological control of YLH will only be effective when both species coexist locally, which depends, among other aspects, on the availability of nesting habitat for the EBE. During the nesting season, the geographic distribution and density of the EBE depends on the availability of soft geological substrates, such as sand, sandstone, clay, gypsum or loess, and, as a rule, on the existence of steep walls in these substrates where the species can dig galleries to build their nests [58,87]. Traditionally, this occurs mainly on the banks of rivers and lakes [58,78], which, according to some authors, the YLH seems to take advantage to expand, particularly the river valleys [31,34], or build large number of colonies, among other habitats [94]. However, nowadays, as happens mainly in Central Europe, EBE colonies are common in sand and gravel pits, quarries, open pit mines, mining subsidence areas, terraces of olive groves and vineyards, railway and road slopes, among other anthropogenic habitats [58,87], which could contribute to a greater territorial dispersion of the bird. On the other hand, the role of the EBE in YLH control will also be conditioned by its quite small home range, which rarely exceeds 1 km from the nesting site during the nestling period, with most foraging activity limited to the first 700 m [58]. Only after fledging and until departure for Africa does the foraging distances increase, reaching extents as far as 15-17 km [58]. From this time onwards and when they migrate southwards towards the wintering grounds of Africa (mainly from August to September/October, but also in November; Bastian & Bastian [58]), EBE predation on the YLH could even be as important as during the nesting season, if not even more. Indeed, this is exactly the time when the YLH colonies reach their maximum growth and activity (from September onwards, but mainly in October and November, concerning the number of workers and the emergence and mating flights of sexual adults [21,31,95]), thus, coinciding with the time when the birds are no longer “attached” to the nest site and form flocks of some dozen individuals (20 – 40 birds) [58], which can move anywhere searching for food and opportunistically taking advantage of this increased availability of YLH.
5. Conclusions
It is confirmed that the European Bee-eater captures the Yellow-legged Hornet and, although data are still scarce, this does not seem to be uncommon. This is not surprising as Hymenoptera traditionally make up a large part of its diet. However, the real importance of this predation remains to be assessed, in order to have an approximation to its contribution in the biological control of this pest, which considerably affects beekeeping and biodiversity.
In the Iberian Peninsula, the role of the EBE is limited by the relatively small overlapping of their distribution ranges (in vast regions where there are the most numerous and widespread populations of the EBE the YLH is not expected to reach there and in most of those where it is already invaded the first one does not occur), an aspect that does not seem to happen in the Western and Central Europe, despite the current lower EBE populations in this regions.
Author Contributions
Conceptualization, N.O. and J.G.; methodology, N.O.; investigation, N.O. and M.I.P.C.; writing—original draft preparation, N.O.; writing—review and editing, M.I.P.C., A.N., I.S.P.C., M.F. and J.G.; project administration, J.G. and A.N.; funding acquisition, J.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the project "Atlantic-POSitiVE", INTERREG Atlantic Area programme (EAPA_800/2018).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to thank the colleagues Luís Filipe Ferreira, Joana Costa, Nuno Cabrita, Paulo Carmo and Carlos Noivo for providing information on the location of several Bee-eater colonies, and also to the Mr. Carlos Cruz for the facilities granted in accessing the Soarvamil sand quarries. We also thank the help of our institute (INIAV) colleagues Pedro Naves and Márcia Santos in identifying some insects or their parts and to José Freitas for the photograph of Bee-eater.
Conflicts of Interest
The authors declare no conflict of interest.
References
- UN (2022). Standard country or area codes for statistical use (M49). Statistics Division, Department of Economic and Social Affairs, United Nations: New York. Available online: https://unstats.un.org/unsd/methodology/m49/.
- Perrard, A.; Arca, M.; Rome, Q.; Muller, F.; Tan, J.; Bista, S.; Nugroho, H.; Baudoin, R.; Baylac, M.; Silvain, J.-F.; Carpenter, J.M.; Villemant, C. Geographic variation of melanisation patterns in a hornet species: Genetic differences, climatic pressures or aposematic constraints? PLoS ONE 2014, 9, e94162. [Google Scholar] [CrossRef] [PubMed]
- Laurino, D.; Lioy, S.; Carisio, L.; Manino, A.; Porporato, M. Vespa velutina: An alien driver of honey bee colony losses. Diversity 2020, 12, 5. [Google Scholar] [CrossRef]
- Barbet-Massin, M.; Salles, J.; Courchamp, F. The economic cost of control of the invasive yellow-legged Asian hornet. NeoBiota 2020, 55, 11–25. [Google Scholar] [CrossRef]
- Bunker, S. Vespa velutina (Asian hornet). CABI Compendium. Available online: https://www.cabidigitallibrary.org/doi/10.1079/cabicompendium.109164 (accessed on 28 February 2023).
- Rojas-Nossa, S.V.; Gil, N.; Mato, S.; Garrido, J. Vespa velutina: características e impactos de una exitosa especie exótica invasora. Ecosistemas 2021, 30, 2159. [Google Scholar] [CrossRef]
- Monceau, K.; Bonnard, O.; Thiéry, D. Vespa velutina: A new invasive predator of honeybees in Europe. J. Pest Sci. 2014, 87, 1–16. [Google Scholar] [CrossRef]
- Nave, A.; Fernandes, J.; Ferreira, M.A.; Rato, F.; Godinho, J. A vespa-asiática, Vespa velutina, espécie exótica e invasora em Portugal. Vida Rural 2022, 263, 58–64. [Google Scholar]
- Rodríguez-Flores, M.S. , Seijo-Rodríguez, A.; Escuredo, O.; Seijo-Coello, M. del C. Spreading of Vespa velutina in northwestern Spain: influence of elevation and meteorological factors and effect of bait trapping on target and non-target living organisms. J. Pest Sci. 2019, 92, 557–565. [Google Scholar] [CrossRef]
- Villemant, C.; Barbet-Massin, M.; Perrard, A.; Muller, F.; Gargominy, O.; Jiguet, F.; Rome, Q. Predicting the invasion risk by the alien bee-hawking Yellow legged hornet Vespa velutina nigrithorax across Europe and other continents with niche models. Biol. Conserv. 2011, 144, 2142–2150. [Google Scholar] [CrossRef]
- Verdasca, M.J.; Godinho, R.; Rocha, R.G.; Portocarrero, M.; Carvalheiro, L.G.; Rebelo, R.; Rebelo, H. A metabarcoding tool to detect predation of the honeybee Apis mellifera and other wild insects by the invasive Vespa velutina. J. Pest Sci. 2022, 95, 997–1007. [Google Scholar] [CrossRef]
- Rojas-Nossa, S.V.; Calviño-Cancela, M. The invasive hornet Vespa velutina affects pollination of a wild plant through changes in abundance and behaviour of floral visitors. Biol. Invasions 2020, 22, 2609–2618. [Google Scholar] [CrossRef]
- Villemant C.; Muller, F.; Rome, Q.; Thiery, D. Evolution des populations du Frelon Asiatique (Vespa velutina, Lepeletier 1836) en France: Conséquences écologiques et socio-économiques. Proposition de mise en œuvre d’un plan d’action. Convention MEEDM/MNHN 2009 - fiche 3. Rapport 2009/7. Muséum National d'Histoire Naturelle: Paris, France, Octobre 2009, 14 p. Available online: http://spn.mnhn.fr/spn_rapports/archivage_rapports/2009/SPN%202009%20-%207%20-%20Vespa%20velutina.pdf.
- Cerri, J.; Lioy, S.; Porporato, M.; Bertolino, S. Combining surveys and on-line searching volumes to analyze public awareness about invasive alien species: a case study with the invasive Asian yellow-legged hornet (Vespa velutina) in Italy. NeoBiota 2022, 73, 177–192. [Google Scholar] [CrossRef]
- Rome, Q.; Villemant, C. Le Frelon asiatique Vespa velutina - Inventaire national du Patrimoine naturel. Muséum national d’Histoire naturelle [Ed]: Paris, France. Available online: http://frelonasiatique.mnhn.fr (accessed on 28 February 2023).
- Ibáñez-Justicia, A.; Loomans, A.J.M. Mapping the potential occurrence of an invasive species by using CLIMEX: case of the Asian hornet (Vespa velutina nigrithorax) in The Netherlands. Proc Neth Entomol Soc Meet. 2011, 22, 39–46. Available online: https://www.researchgate.net/publication/260277540_Mapping_the_potential_occurrence_of_an_invasive_species_by_using_CLIMEX_case_of_the_Asian_hornet_Vespa_velutina_nigrithorax_in_The_Netherlands_Proc_Neth_Entomol_Soc_Meet/citation/download.
- Barbet-Massin, M.; Rome, Q.; Muller, F.; Perrard, A.; Villemant, C.; Jiguet, F. Climate change increases the risk of invasion by the yellow-legged hornet. Biol. Conserv. 2013, 157, 4–10. [Google Scholar] [CrossRef]
- Monceau, K.; Maher, N.; Bonnard, O.; Thiéry, D. Predation pressure dynamics study of the recently introduced honeybee killer Vespa velutina: learning from the enemy. Apidologie 2013, 44, 209–221. [Google Scholar] [CrossRef]
- Verdasca, M.J.; Carvalheiro, L.; Aguirre Gutierrez, J.; Granadeiro, J.P.; Rome, Q.; Puechmaille, S.J.; Rebelo, R.; Rebelo, H. Contrasting patterns from two invasion fronts suggest a niche shift of an invasive predator of native bees. PeerJ. 2022, 10, e13269. [Google Scholar] [CrossRef] [PubMed]
- Robinet, C.; Darrouzet, E.; Suppo, C. Spread modelling: a suitable tool to explore the role of human-mediated dispersal in the range expansion of the yellow-legged hornet in Europe. Int. J. Pest Manag. 2019, 65, 258–267. [Google Scholar] [CrossRef]
- Balmori, A. Sobre el riesgo real de una expansión generalizada de la avispa asiática Vespa velutina Lepeletier, 1836 (Hymenoptera: Vespidae) en la Península Ibérica. Boletín de la S.E.A. 2015, 56, 283–289. [Google Scholar]
- Matsuura, M.; Yamane, S. Biology of Vespine Wasps, 1st ed.; Springer-Verlag: Berlin Heidelberg, Germany, 1990; 323p. [Google Scholar] [CrossRef]
- Kim, C.-J.; Choi, M.B. First Discovery of Vespa velutina nigrithorax du Buysson (Hymenoptera: Vespidae), an Invasive Hornet in the Feces of the Yellow-Throated Marten in South Korea. Insects 2021, 12, 296. [Google Scholar] [CrossRef] [PubMed]
- Spradbery, P.J. Wasps: An Account of the Biology and Natural History of Solitary and Social Wasps., 1st ed.; University of Washington Press: Seatle, USA, 1973; 408p. [Google Scholar]
- Edwards, R. Social Wasps. Their Biology and Control; 1st ed.; Rentokil Ltd.: East Grinstead, UK, 1980; 398p. [Google Scholar]
- Fox-Wilson, G. Factors affecting populations of social wasps, Vespula species, in England (Hymenoptera). Proc. R. Entomol. Soc. Lond. Ser. A Gen. Entomol. 1946, 21, 17–27. [Google Scholar] [CrossRef]
- van Bergen, V.S. The Honey-Buzzards of the Sensebezirk: First Findings on Density, Diet, Reproduction and Food Competition in a Swiss Population of Honey-Buzzard Pernis Apivorus. 2019, December 12. 12 December. [CrossRef]
- Nadolski, J. Factors restricting the abundance of wasp colonies of the European hornet Vespa crabro and the Saxon wasp Dolichovespula saxonica (Hymenoptera: Vespidae) in an urban area in Poland. Entomol. Fenn. 2013, 24, 204–215. [Google Scholar] [CrossRef]
- Pusceddu, M.; Lezzeri, M.; Cocco, A.; Floris, I.; Satta, A. Bio-Ethology of Vespa crabro in Sardinia (Italy), an Area of New Introduction. Biology 2022, 11, 518. [Google Scholar] [CrossRef]
- Mollet, T.; de la Torre, C. Vespa velutina. The Asian Hornet. Bull. Tech. Agric. 2006, 33, 203–208. [Google Scholar]
- Villemant, C.; Muller, F.; Haubois, S.; Perrard, A.; Darrouzet, E.; Rome, Q. Bilan des Travaux (MNHN et IRBI) sur l’invasion en France de Vespa velutina, le frelon asiatique prédateur d’abeilles. In Journée Scientifique Apicole; Barbançon J.-M., L’Hostis, M. Eds.; JSA, Arles, ONIRIS-FNOSAD: Nantes, France, 11 Février 2011, pp 3-12. Available online: http://inpn.mnhn.fr/fichesEspece/Vespa_velutina_fichiers/2011_02_11_Bilan_Invasion_Vespa_velutina_JSA.pdf.
- Bessa, A.S.; Carvalho, J.; Gomes, A.; Santarém, F. Climate and land-use drivers of invasion: predicting the expansion of Vespa velutina nigrithorax into the Iberian Peninsula. Insect Conserv. Divers. 2016, 9, 27–37. [Google Scholar] [CrossRef]
- Fournier, A.; Barbet-Massin, M.; Rome, Q.; Courchamp, F. Predicting species distribution combining multi-scale drivers. Glob. Ecol. Conserv. 2017, 12, 215–226. [Google Scholar] [CrossRef]
- Carvalho, J.; Hipólito, D.; Santarém, F.; Martins, R.; Gomes, A.; Carmo, P.; Rodrigues, R.; Grosso-Silva, J.; Fonseca, C. Patterns of Vespa velutina invasion in Portugal using crowdsourced data. Insect Conserv. Divers 2020, 13, 501–507. [Google Scholar] [CrossRef]
- Krebs, J.R.; Avery, M.I. Central place foraging in the European Bee-eater, Merops apiaster. J. Anim. Ecol. 1985, 54, 459–472. [Google Scholar] [CrossRef]
- Krištín, A. Breeding biology and diet of the bee-eater (Merops apiaster) in Slovakia. Biologia (Bratisl.) 1994, 49, 273–279. [Google Scholar]
- Kiss, J.B.; Rekasi, J. Date privind hrana în Delta Dunării a unor specii de păsări granivore şi insectivore strict protejate. Analele Ştiinţifice ale Institutului Delta Dunării 1995, 4, 123–129. [Google Scholar]
- Galeotti, P.; Inglisa, M. Estimating predation impact of Honeybees Apis mellifera L. by European Bee-eaters Merops apiaster. Rev. Ecol. (Terre Vie) 2001, 56, 373–388. [Google Scholar] [CrossRef]
- Krištín, A.; Kaňuch, P. Šíri sa včelárik zlatý (Merops apiaster) severným smerom? K výskytu a potrave v Podpoľaní a okolí Zvolena (stredné Slovensko). Tichodroma 2005, 17, 89–94. [Google Scholar]
- Lourenço, P.M. Internet photography forums as sources of avian dietary data: bird diets in Continental Portugal. Airo 2018, 25, 3–26. [Google Scholar]
- Gamauf, A. Der Wespenbussard (Pernis apivorus) ein Nahrungsspezialist? Der Einfluß sozialer Hymenopteren auf Habitatnutzung und Home Range-Größe. Egretta 1999, 12, 57–85. [Google Scholar]
- Roberts, S.J.; Lewis, J.M.S.; Williams, I.T. Breeding European Honey-buzzards in Britain. Br. Birds 1999, 92, 326–345. [Google Scholar]
- Lepley, M.; Thevenot, M.; Guillaume, C.-P.; Ponei, P.; Bayle, P. Diet of the nominate Southern Grey Shrike Lanius meridionalis meridionalis in the north of its range (Mediterranean France). Bird Study 2004, 51, 156–162. [Google Scholar] [CrossRef]
- Tryjanowski, P.; Karg, M.K.; Karg, J. Diet composition and prey choice by red-backed shrike Lanius collurio in western Poland. Belg. J. Zool. 2003, 133, 157–162. [Google Scholar]
- Owen, D.F. The Food of Nestling Jays and Magpies. Bird Study 1956, 4, 257–265. [Google Scholar] [CrossRef]
- Labouyrie, F. Winter larders of the Iberian Grey Shrike Lanius meridionalis in southern France. Alauda 2020, 88, 35–44. [Google Scholar]
- Vigneaud, J.P. Gironde: fait rarissime, un rapace dévore un nid de frelons asiatiques. Sud Ouest, 2013, 20 August. Available online: https://www.sudouest.fr/environnement/frelon-asiatique/gironde-fait-rarissime-un-rapace-devore-un-nid-de-frelons-asiatiques-8644366.php.
- Macià, F.X.; Menchetti. M.; Corbella, C.; Grajera, J.; Vila, R. Exploitation of the invasive Asian Hornet Vespa velutina by the European Honey Buzzard Pernis apivorus. Bird Study 2019, 66, 425–429. [Google Scholar] [CrossRef]
- Rebollo, S.; Rey-Benayas, J.M.; Villar-Salvador, P.; Pérez-Camacho, L.; Castro, J.; Molina-Morales, M.; Leverkus, A.B.; Baz, A.; Martínez-Baroja, L.; Quiles, P.; et al. Servicios de la avifauna (high-mobile link species) en mosaicos agroforestales: regeneración forestal y regulación de plagas. Ecosistemas 2019, 28, 32–41. [Google Scholar] [CrossRef]
- Sequeira, I. Novo documentário mostra a vida selvagem a conviver com os humanos em Lousada. Wilder. 2020, 19 Mars. Available online: https://www.wilder.pt/?s=Novo+document%C3%A1rio+mostra+a+vida+selvagem+a+conviver+com+os+humanos+em+Lousada.
- Onofre, N.; Portugal e Castro, M.I.; Nave, A.; Godinho, J. Os Predadores Naturais da Vespa-Asiática (Vespa velutina) e a Contribuição do Abelharuco (Merops apiaster) para o seu Controlo. Silva Lusit. 2022, 30, 1–16. [Google Scholar] [CrossRef]
- Villemant, C.; Rome, Q.; Haxaire, J. Le Frelon asiatique (Vespa velutina) – In Muséum National d'Histoire Naturelle [Ed]. Inventaire national du Patrimoine Naturelle [Ed]: Paris, France, 2010. Available online: https://inpn.mnhn.fr/fichesEspece/Vespa_velutina_en.html (accessed on 28 February 2023).
- Palomino, D.; Valls, J. Las rapaces forestales en España. Población reproductora en 2009-2010 y método de censo; SEO/BirdLife: Madrid, España, 2011; 153 p.
- Anonymous. El abejero liquidó 24.500 nidos de velutina en Galicia. Faro de Vigo. 2019, 31 Mars. Available online: https://www.farodevigo.es/sociedad/2019/03/31/abejero-liquido-24-500-nidos-15733121.html.
- Purroy, J.; Purroy, F.J. Abejero europeo –Pernis apivorus. In Enciclopedia Virtual de los Vertebrados Españoles; Salvador, A., Morales, M.B., Eds.; Museo Nacional de Ciencias Naturales: Madrid, España, 2016. Available online: http://www.vertebradosibericos.org/. (accessed on 28 February 2023).
- Valera, F. Abejaruco europeo – Merops apiaster. In Enciclopedia Virtual de los Vertebrados Españoles; Salvador, A., Morales, M.B., Eds.; Museo Nacional de Ciencias Naturales: Madrid, España, 2016; Available online: http://www.vertebradosibericos.org/ (accessed on 28 February 2023).
- Orta, J.; Kirwan, G.M.; Garcia, E.F.J. European Honey-buzzard (Pernis apivorus), version 1.0. In Birds of the World; del Hoyo, J., Elliott, A., Sargatal, J., Christie, D.A., de Juana, E., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. Available online: https://birdsoftheworld.org/bow/species/euhbuz1/cur/introduction (accessed on 28 February 2023).
- Bastian, H.-V.; Bastian, A. European Bee-eater (Merops apiaster), version 2.0. Birds of the World. In Birds of the World; Billerman, S.M., Keeney, B.K., Eds.; Cornell Lab of Ornithology, Ithaca, NY, USA, 2022. Available online: https://birdsoftheworld.org/bow/species/eubeat1/cur/introduction (accessed on 28 February 2023).
- Birkhead, TR. Predation by birds on social wasps. Br. Birds 1974, 67, 221–229. [Google Scholar]
- Massa, B.; Rizzo, M.C. Nesting and feeding habits of the European Bee-eater (Merops apiaster L.) in a colony next to a beekeeping site. Avocetta 2002, 26, 25–31. [Google Scholar]
- Herrera, C.M.; Ramírez, A. Food of Bee-eaters in Southern Spain. Br. Birds 1974, 67, 158–164. [Google Scholar]
- Martínez, C. Notes sur l'alimentation du Guêpier (Merops apiaster L.) dans une colonie du centre de l'Espagne. Alauda 1984, 52, 45–50. [Google Scholar]
- GIC. La evaluacion del impacto de la ecologia del Abejaruco (Merops apiaster) sobre el sector apicola en el ambito rural Transfronteriza de Extremadura. Grupo de Investigacion en Biologia de la Conservacion (G.I.C.), Universidad de Extremadura, Junta Extremadura. Informe definitivo, 2007, December; 27 pp. Available online: http://extremambiente.juntaex.es/files/interreg/faunatrans/estudios_mejoras/impacto%20ecologia%20abejaruco%20definitivo.pdf.
- Costa, J.; Rocha, A.; Alves, J.A. Diet of adult and nestling Bee-eaters (Merops apiaster) in Portugal. In IX Congresso de Ornitologia da SPEA, Vila Real, Portugal, 23 – 25 April 2016. Available online: https://www.researchgate.net/publication/313882699_Diet_of_adult_and_nestling_Bee-eaters_Merops_apiaster_in_Portugal.
- GBIF Secretariat Vespa velutina subsp. nigrithorax Buysson, 1905. GBIF Backbone Taxonomy. Available online: https://www.gbif.org/species/6247411 (accessed on 28 February 2023).
- GBIF Secretariat. Pernis apivorus (Linnaeus, 1758). GBIF Backbone Taxonomy. Available online: https://www.gbif.org/species/2480420 (accessed on 28 February 2023).
- Keller, V.; Herrando, S.; Voříšek, P.; Franch, M.; Kipson, M.; Milanesi, P.; Martí, D.; Anton, M.; Klvaňová, A.; Kalyakin, M.V.; et al. European Breeding Bird Atlas 2: Distribution, Abundance and Change; European Bird Census Council & Lynx Edicions: Barcelona, Spain. https://ebba2.info/maps/species/Merops-apiaster/ebba2/breeding/ (accessed on 28 February 2023).
- GBIF Secretariat. Merops apiaster Linnaeus, 1758. GBIF Backbone Taxonomy. Available online: https://www.gbif.org/species/2475443 (accessed on 28 February 2023).
- STOPVespa. Mapa STOPvespa. Available online: https://stopvespa.icnf.pt/geovisualizador/ (accessed on 28 February 2023).
- Equipa Atlas. Atlas das Aves Nidificantes em Portugal (1999-2005); Instituto da Conservação da Natureza e da Biodiversidade, Sociedade Portuguesa para o Estudo das Aves, Parque Natural da Madeira e Secretaria Regional do Ambiente e do Mar, Assírio & Alvim: Lisboa, Portugal, 2008; 590 p.
- Chinery, M. Insects of Britain and Western Europe., 2nd ed.; A & C Black Publishers Ltd: London, UK, 2007. [Google Scholar]
- DGAV. Plano de Ação para o controlo de Rhynchophorus ferrugineus (Olivier). Direcção Geral da Alimentação e Veterinária. Ministério da Agricultura e do Mar: Lisboa, Portugal, 2013, October; 19 p. Available online: https://www.drapc.gov.pt/base/documentos/plano_accao_r%20_ferrugineus%20_2013_dgav.pdf. /.
- Verdasca, M.J.; Rebelo, H.; Carvalheiro, L.G.; Rebelo, R. Invasive hornets on the road: motorway-driven dispersal must be considered in management plans of Vespa velutina. NeoBiota 2021, 69, 177–198. [Google Scholar] [CrossRef]
- MAGRAMA. Estrategia de gestión, control y posible erradicación del Avispón asiático o Avispa negra (Vespa velutina ssp. nigrithorax) en España. Estratégias de control - Critérios orientadores. Espécies Exóticas Invasoras, Ministério de Agricultura, Alimentación y Medio Ambiente, Govierno de España: Madrid, España, 2015; 58 p. Available online: https://www.miteco.gob.es/es/biodiversidad/publicaciones/estrategia_vespavelutina_tcm30-69976.pdf.
- Kim, S.-H.; Kim, D.; Lee, H.; Jung, S.; Hee, W.-H. Ensemble evaluation of the potential risk areas of yellow-legged hornet distribution. Environmental Monitoring and Assessment 2021, 193, 601. [Google Scholar] [CrossRef] [PubMed]
- Voous, K.H. Atlas of European birds, 1st ed.; Thomas Nelson and Sons: New York, USA, 1960; 284p. [Google Scholar]
- Kinzelbach, R.; Nicolai, B.; Schlenker, R. Der Bienenfresser Merops apiaster als Klimazeiger: Zum Einflug in Bayern, der Schweiz und Baden im Jahr 1644. J. fur Ornithol. 1997, 138, 297–308. [Google Scholar] [CrossRef]
- Bastian, A.; Bastian, H.-V.; Fiedler, W.; Rupp, J.; Todte, I.; Weiss, J. Der Bienenfresser (Merops apiaster) in Deutschland – eine Erfolgsgeschichte. Fauna Flora Rheinl.-Pfalz 2013, 12, 861–894. [Google Scholar]
- Bastian, H.-V. Go North – Bienenfresser vom Mittelmeer bis zur Ostsee? Jahrb. Natschutz. Hessen 2014, 15, 138–141. [Google Scholar]
- Fiedler W. Chapter 17. Bird ecology. In Climate change – Observed impacts on earth, 3rd ed.; Letcher, T.M. Ed.; Elsevier: Amsterdam, Boston, Heidelberg, London, New York, Oxford, Paris, San Diego, San Francisco, Singapore, Sydney, Tokyo, 2021; pp. 373–388. Climate change – Observed impacts on earth. [CrossRef]
- Stiels, D.; Bastian, H.V.; Bastian, A.; Schidelko, K.; Engler, J.O. An iconic messenger of climate change? Predicting the range dynamics of the European Bee-eater (Merops apiaster). J. Ornithol. 2021, 162, 631–644. [Google Scholar] [CrossRef]
- Lioy, S.; Bergamino, C.; Porporato, M. The invasive hornet Vespa velutina: distribution, impacts and management options. CABI Reviews. 2022, 19 October. [CrossRef]
- Aymí, R. (2003). Abejaruco europeo Merops apiaster. In Atlas de las aves reproductoras de España; Martí, R., del Moral, J. C. Eds.; Dirección General de Conservación de la Naturaleza-Sociedad Española de Ornitología: Madrid, España; pp. 344–345.
- Carrascal, L. M.; Palomino, D. Las aves comunes reproductoras en España. Población en 2004-2006. SEO/BirdLife, Madrid, 2008; 202 p.
- Catry, P., Costa, H., Elias, G., Matias, R., Aves de Portugal - Ornitologia do Território Continental. Assírio & Alvim: Lisboa, Portugal, 2010; 944 p.
- Gordo, O.; Aymí, R.; Molina, B. Der Bienenfresser Merops apiaster in Spanien: seine Verbreitung, Häufigkeit und Bedrohung. Vogelwarte 2021, 59, 235–245. [Google Scholar]
- Aymí, R.; Gordo, O. Abejaruco europeo Merops apiaster. In III Atlas de las aves en época de reproducción en España, Molina, B., Nebreda, A., Muñoz, A.R., Seoane, J., Real, R., Bustamante, J., del Moral, J.C. Eds.; SEO/BirdLife: Madrid, España, 2022. Available online: https://atlasaves.seo.org/ave/abejaruco-europeo/ (accessed on 28 February 2023).
- Araújo, M.B.; Guilhaumon, F.; Neto, D.R.; Pozo, I.; Calmaestra, R. (2011). Impactos, Vulnerabilidad y Adaptación al Cambio Climático de la Biodiversidad Española. 2 Fauna de Vertebrados, 1st ed.; Dirección general de medio Natural y Política Forestal. Ministerio de Medio Ambiente, y Medio Rural y Marino: Madrid, España, 2011; 640p. [Google Scholar]
- MNHN & OFB (Ed) Fiche de Vespa velutina Lepeletier, 1836. Inventaire national du patrimoine naturel (INPN): France, 2003-2023. Available online: https://inpn.mnhn.fr/espece/cd_nom/433589 (accessed on 28 February 2023).
- Dellwisch, B; Bastian, A; Bastian, H.-V.; Schidelko, K.; Stiels, D.; Engler, J.O. Bienenfresser Merops apiaster in Deutschland – woher sie kamen, wohin sie gehen. Vogelwarte 2021, 59, 197–206.
- LPO. Guêpier d'Europe Merops apiaster. Le projet ODF, Répartitions, Tendances et Effectifs. Oiseaux de France. Ligue pour la Protection des Oiseaux (LPO). Available online: https://oiseauxdefrance.org/prospecting?species=3582 (accessed on 28 February 2023).
- Loury, P.; Puissauve, R. Fiches d’information sur les espèces aquatiques protégées: Guêpier d’Europe, Merops apiaster Linnaeus, 1758. Service du patrimoine naturel du MNHN & Onema, 2016; 4 p. Available online: https://inpn.mnhn.fr/fichesEspece/EspecesEauDouce/Guepier_europe-M.apiaster_2016.pdf (accessed on 28 February 2023).
- Bastian, H.-V. (2022). 10. Jahresbericht der FG “Bienenfresser” der DO-G. Fachgruppe “Bienenfresser” der Deutschen Ornithologen-Gesellschaft; 7 p. http://www.do-g.de/fileadmin/Jahresbericht_2022_-_Zehnter_Bericht_der_FG_Bienenfresser_DO-G.pdf. Available online: https://atlasaves.seo.org/ave/abejaruco-europeo/. /.
- Monceau, K.; Bonnard, O.; Thiéry, D. Chasing the queens of the alien predator of honeybees: A water drop in the invasiveness ocean. Open J. Ecol. 2012, 2, 183–191. [Google Scholar] [CrossRef]
- Rome, Q; Muller, F.; Touret-Alby, A.; Darrouzet, E.; Perrard, A.; Villemant, C. Caste differentiation and seasonal changes in Vespa velutina (Hym.: Vespidae) colonies in its introduced range. J. Appl. Entomol. 2015, 139, 771–782. [CrossRef]
Figure 1.
European Bee-eater (Merops apiaster) (Photo credit: José Freitas).

Figure 2.
Study area and Bee-eater nesting sites locations [Because the scale of the figure is very small, several locations/asterisks are overlapping; Administrative Units based on Oficial Cartography for Portugal (CAOP) from D.G.T. (Direção Geral do Território)].
Figure 2.
Study area and Bee-eater nesting sites locations [Because the scale of the figure is very small, several locations/asterisks are overlapping; Administrative Units based on Oficial Cartography for Portugal (CAOP) from D.G.T. (Direção Geral do Território)].
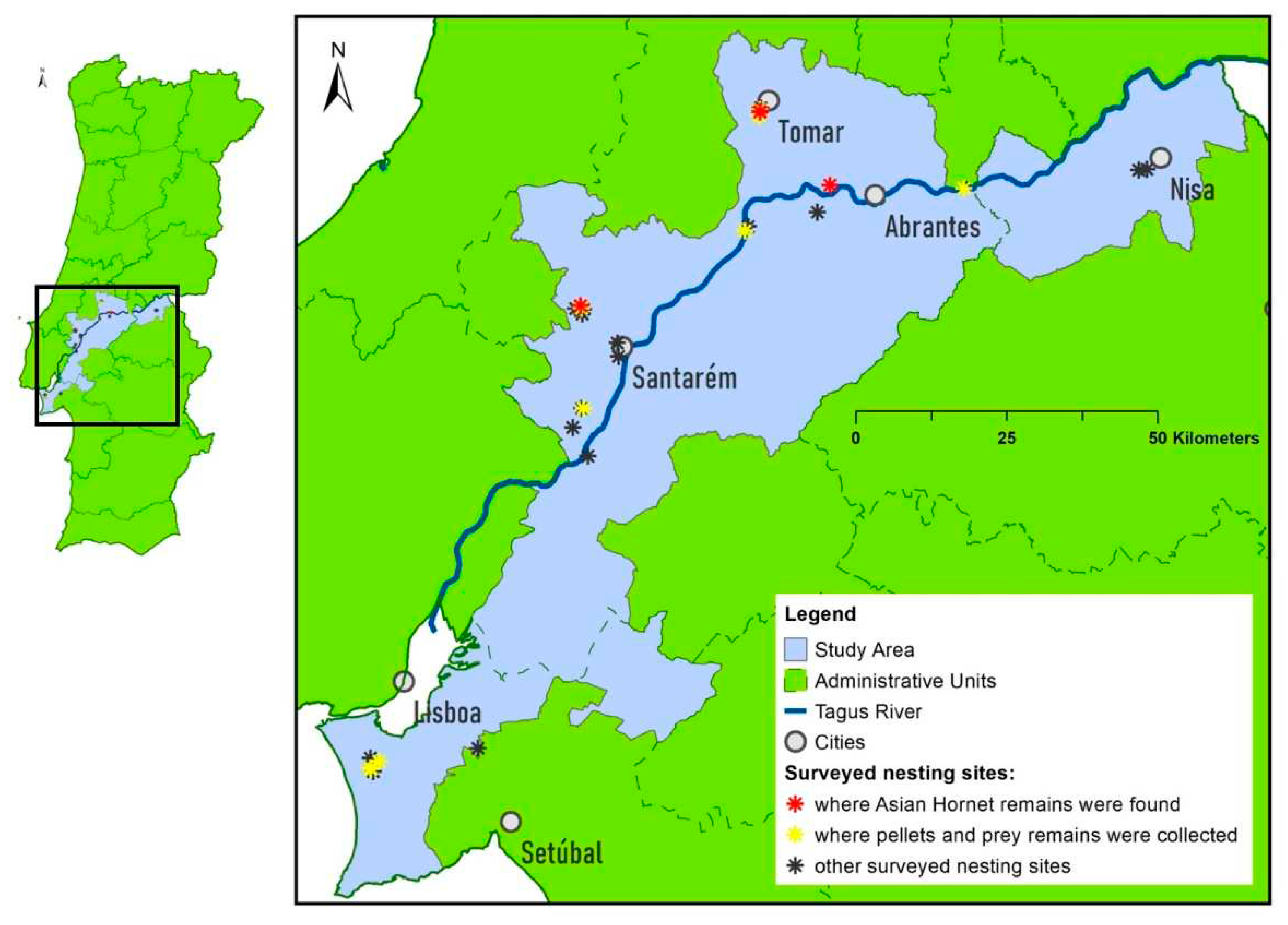
Figure 3.
Pellets and whole prey individuals [Yellow-legged Hornet (Vespa velutina) (bottom left) and a Mammoth Wasp (Megascolia maculata) (bottom right)] of European Bee-eater.
Figure 3.
Pellets and whole prey individuals [Yellow-legged Hornet (Vespa velutina) (bottom left) and a Mammoth Wasp (Megascolia maculata) (bottom right)] of European Bee-eater.

Figure 4.
Distribution of Yellow-legged Hornet (a) and Bee-eater (b) records in Europe according to GBIF Secretariat [65,68].
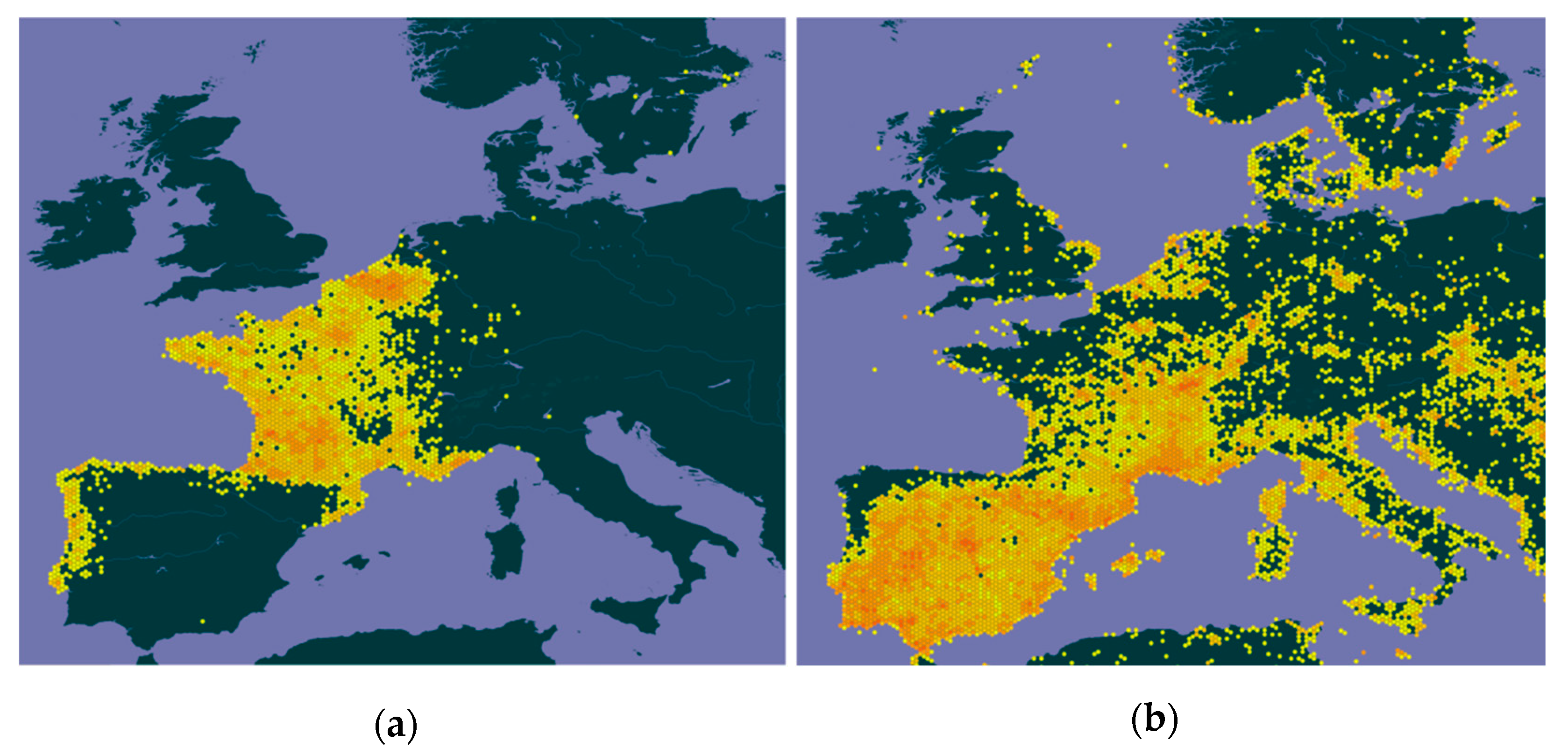

Table 1.
Nominal list of prey captured by the European Bee-eater, based on remains found under perches and nests at nesting sites visited in 2021.
Table 1.
Nominal list of prey captured by the European Bee-eater, based on remains found under perches and nests at nesting sites visited in 2021.
| Order | Family | Subfamily | Genus or Species |
|---|---|---|---|
| HEMIPTERA | Cicadellidae | ||
| DIPTERA | Syrphidae | Eristalinae | Volucella zonaria |
| HYMENOPTERA | Apidae | Apinae | Apis mellifera |
| Bombus sp. | |||
| Melecta sp. | |||
| Xylocopinae | Xylocopa violacea | ||
| Crabronidae | |||
| Scoliidae | Scoliinae | Megascolia maculata | |
| Vespidae | Polistinae | Polistes gallicus | |
| Polistes sp. | |||
| Vespinae | Vespa crabro | ||
| Vespa velutina | |||
| Vespula germanica | |||
| Vespula sp. | |||
| COLEOPTERA | Staphylinidae | ||
| Geotrupidae | |||
| Scarabaeidae | Cetoniinae | Cetonia sp. | |
| Protaetia sp. | |||
| Buprestidae | Buprestinae | Buprestis sp. | |
| Tenebrionidae | Tenebrioninae | Blaps sp. | |
| Curculionidae | Rhynchophorinae | Rhynchophorus ferrugineus |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



