
Submitted:
22 August 2023
Posted:
24 August 2023
Read the latest preprint version here
Abstract
Due to the variation of the body coloration and the similarity of the related species, there are still some unresolved problems and debates in the taxonomic study of the genus Sycanus in China. We conducted the inter-specific phylogenetic analyses and the species delimitation for Sycanus in China based on DNA barcoding dataset of the mitochondrial cytochrome c oxidase subunit I gene of 81 samples and partial 28S rDNA sequences of 31 samples. The result showed that all the samples were divided into twelve species by integrating the results of DNA barcoding and morphological comparison. In the present paper, the genus Sycanus in China was reviewed systematically, all species were described or re-described, keyed, illustrated and photographed, among them, Sycanus taiwanensis Zhao & Cai sp. nov., Sycanus hsiaoi Li & Cai sp. nov. and Sycanus hainanensis Wang & Cai sp. nov. were described as new to science. The biological informations of Sycanus falleni Stål, 1863 and Sycanus croceus Hsiao, 1979 were briefly recorded.
Keywords:
China
; Sycanus
; taxonomy
; DNA barcoding
; species delimitation
; phylogeny
; new species
; biology
1. Introduction
In the research of entomology taxonomy, there are sometimes challenges in distinguishing the differences between the close relative species and in identifying complex morphological changes among different geographical populations of a species, which pose a significant obstacle for entomologists who rely on the traditional taxonomy methods to address the issue of species identification or delimitation. Certain morphological changes can be attribute to variations within populations, while others are due to species differentiation, even if they are continuous and gradual [1,2,3,4]. This is very common in the field of biological taxonomy and often affects the ability of entomology taxonomic researchers to accurately determine species boundaries. The integration of molecular biology and morphology has revitalized the study of traditional taxonomy. The molecular phylogenetic analysis method, which is based on DNA data, provides an objective representation of actual genetic relationships and species differentiation, and offers entomology taxonomical researchers an effective method to gain insight into the laws and essence of biological evolution and speciation through the investigateion of phenotypic changes in insects. In particular, DNA barcoding technology that utilizes the mitochondrial cytochrome c oxidase subunit I gene (COI) has emerged as a pivotal tool in insect identification, species delimitation, and species divergence evolution [1,2,3,4,5,6,7,8,9,10,11,12,13,14,15]. One of the significent developments in the modern entomology taxonomy is the integrative taxonomical method which combines DNA barcoding molecular data and morphological evidence [5,16]. Although several studies have demonstrated that DNA barcoding technology may not always be effective in resolving species delimination, and in some cases, lead to an overestimation of species diversity or potential suppression to the traditional morphological entomology taxonomy [17,18,19,20]. Nevertheless, it is undeniable that DNA barcoding technology is important to entomology taxonomy. The contribution of integrative taxonomical method is crucial in providing clarity regarding the taxonomic status of many species that pose in both present and future studies [1,2,3,4,5,6,7,8,9,10,11,12,13,14,15,16,21]. The utilization of COI DNA barcoding is appropriate for species identification in closely related species [5], but not recommended for use in high taxonomotic goups due to the potential issue of parallel evolution.
The genus Sycanus was established by Amyot & Serville in 1843 for the oriental species, Reduvius collaris Fabricius, 1781 as the type species, with seventy-six Oriental and one Madagascar known species [22,23,24,25]. The beautiful assassin bugs are easily separated from other allied genera by the following characters: the head is much longer than the pronotum, the scutellum is apically armed with a spine or tubercle, and the connexivum of abdomen is extremely laterally dilated. Prior to this study, a total of twelve species belonging to the genus Sycanus have been recorded within China [24,26,27,28,29,30,31,32]. In the present paper, the fifteen species of the genus Sycanus in China were listed. With the exception of Sycanus collaris (Fabricius, 1781), Sycanus bifidus (Fabricius, 1787) and Sycanus fuscirostris Dohrn, 1859, the remaining twelve Chinese Sycanus species are described and re-described, morphologically compared and photographed in detail, among them, including three new species: Sycanus hainanensis Wang & Cai sp. nov., Sycanus hsiaoi Li & Cai sp. nov. and Sycanus taiwanensis Zhao & Cai sp. nov..
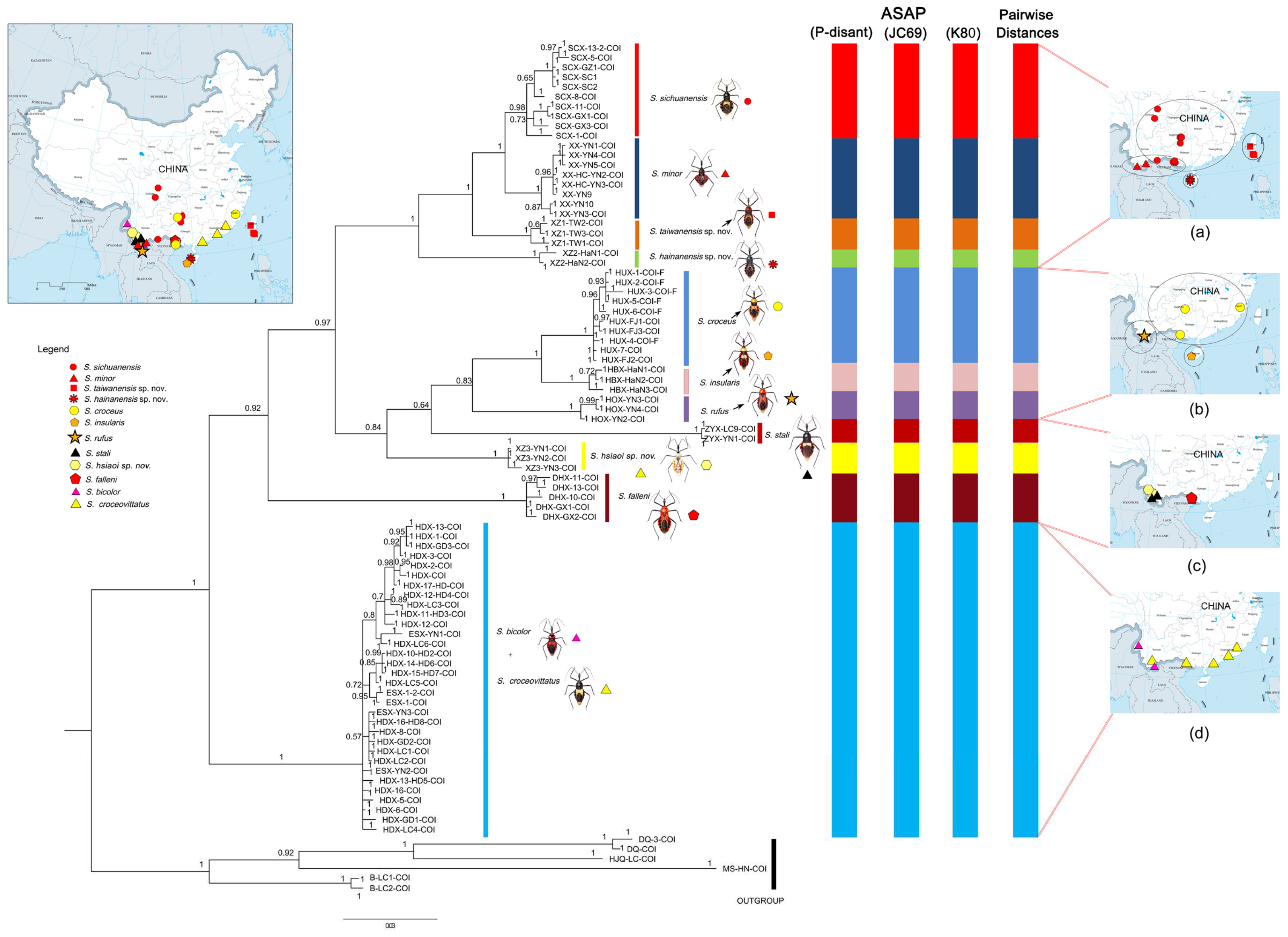
In this study, we conducted species delimitation and phylogeny analyses on the twelve Sycanus species found in China based on the COI DNA barcodes dataset of 81 Chinese Sycanus samples (Figure 1, S1, S2). The findings revealed that all samples used in this study were divided in 11 molecular operational taxonomic unites (MOTUs) and identified as eleven distinct species. Among these species, three were newly discovered, as confirmed by morphological analyses. However, two species, that is, Sycanus bicolor Hsiao, 1979 and Sycanus croceovittatus Dohrn, 1859, clould not be differentiated from each other based on their DNA barcodes sequences (Figure 1, S1, S2). To further investigate the differentiation between these two species in terms of body structure and body coloration, we added partial 28S rDNA sequences. The two species can be categorized into two distinct clades in the NJ tree obtaining from the phylogeny analyses of 28s rDNA dataset of 34 specimens belonging to five Sycanus species (Figure S3). However, they are still considered to be the most closely related species among the group. Due to their consistent and distinguishable morphological characteristics, we assigned the two species, S. bicolor Hsiao, 1979 and S. croceovittatus Dohrn, 1859, as two separate and distinct species.
It should be noted that although Sycanus collaris Fabricius, 1781, Sycanus bifidus Fabricius, 1787, and Sycanus fuscirostri Dohrn, 1859 have been recorded to be found within China [22,26,27], they are not included in this study. This is because we have not located the specimens of three species during the examination of the Chinese domestic insect specimen collection institutions and the recent field trip. It is also regrettable that we failed to examine these old type specimens. Therefore, the distribution of these species within China remains uncertain, thereby necessitating further investigation in subsequent studies.
2. Materials and Methods
2.1. Specimens and Acronyms
This study is based on the materials deposited in Entomological Museum of China Agricultural University, Institute of Zoology of Chinese Academy of Sciences and Nankai University and so on. The following acronyms for public collections are used:
BMNH, the Natural History Museum, London;
CAU, Entomological Museum of China Agricultural University, Beijing, China;
CATAS, Chinese Academy of Tropical Agricultural Sciences, Haikou, Hainan, China.
IOZ, Institut of Zoology, Chinese Academy of Sciences, Beijing, China;
NKU, College of Life Sciences, Nankai University, Tianjin, China;
NNU, Nanning Normal University, Nanning, Guangxi, China;
TJNHM, Tianjin Natural History Museum, Tianjin, China.
2.2. Taxonomy
The external structures were examined using a binocular dissecting microscope. Male genitalia was soaked in hot 90℃ lactic acid for approximately 10–15 minutes to remove soft tissue, then put it in hot boiling distilled water, a few minutes later endosoma extend naturally from the phallus, or pull it out of phallus using a pair of the pointed tweezers. The dissected parts of genitalia were placed in a plastic microvial with lactic acid under the corresponding specimen. All habitus photographs were taken by using the Canon D60 SLR camera (Canon Inc., Tokyo, Japan). All photographs of male genitalia were taken with the aid of the Research Stereo Microscope SMZ25 (Nikon Corporation, Tokyo, Japan). The drawings were traced with the aid of a camera lucida. All measurements were in millimeters. Body length was measured from the apex of head to the tip of the hemelytra in resting position. Maximum width of pronotum was measured across humeral angles. The classification system and morphological terminology mainly followed Hsiao and Ren [31]. The species names in the text were arranged alphabetically.
2.3. Integrative Molecular Analysis
2.3.1. DNA Extraction and Sequencing
We used thorax muscles of a single adult individual for DNA extraction. The total DNA was extracted using the Animal Tissue Extraction Kit (Golden EasyPure® Genomic DNA Kit, Transgenbiotech, Beijing, China) and stored at –20 °C. The COI DNA barcodes were amplified using the developed primers COI-F (5′-TGATCAGGTGTAGTG GGGAC-3′) and COI-R (5′-TCCCTAATGACCCGAAAGCT-3′), and the partial 28S rDNA sequences were obtained using the known primers 28S-F (5′-AGACTCCTTGGT CCGTGTTTC-3′) and 28S-R (5′-ATCACTCGGCTCGTGGATCG-3′) [33]. The PCR reaction system was 50 µL: 1 µL DNA template, 25 µL 2× EasyTaq®PCR SuperMix (Transgenbiotech, Beijing, China), 1 µL forward and reverse primers (10 µM), and 22 µL nuclease-free water. The reaction procedure was as follows: pre-denaturation at 94 °C for 3.5 min, denaturation at 94 °C for 30 s, annealing at 56 °C for 30 s , elongation at 72 °C for 1 min in COI DNA barcodes and 1.5 min in 28S rDNA, and 35 cycles and elongation at 72 °C for 8 min. The PCR products were detected by 2% agarose gel electrophoresis, then observed and photographed in the gel imaging analysis system. For all specimens, sequences were obtained and bidirectionally sequenced using the same PCR primer pairs. Qingke Biotechnology Co., Ltd. (Hangzhou,, China) and Shanghai Shenggong Biotechnology Co., Ltd. (Shanghai, China) completed the sequencing of PCR amplicons using the Sanger sequencing method.
2.3.2. Phylogenetic Analyses
A total of COI DNA barcodes of 81 individuals and partial 28S rDNA sequemces of 34 ones were successfully obtained from 188 Chinese Sycanus samples (Table A1, S1). In addition, COI DNA barcodes of 6 individuals and partial 28S rDNA sequences of 8 ones were obtained from the closed related genus as outgroups and used in the phylogenetic analyses (Table A1). The resultant sequences were 675 bp for COI DNA barcodes and 639 bp for partial 28S rDNA sequences, and were identified as the genus Sycanus members’s genes in the BLAST procedure search in GenBank. The DNA sequences were aligned using MEGA X [34].
Neighbor-joining (NJ) trees was used to estimate the topologies using MEGA X based on the Kimura-2-Parameter (K2P) model [35] and 1000 bootstrap replicates. The monophyletic for the delimited species was defined by NJ analyses.
The maximum likelihood (ML) tree was constructed using the IQ-TREE web server [36] with 1000 replicates using the ultrafast bootstrap approximation approach. The substitution model was automatically predicted in the IQ-TREE software and we used the best-fit model of HKY+F+I+G4 on COI DNA barcoding dataset according to the Bayesian information criterion (BIC).
The Bayesian inference (BI) was conducted using MrBayes 3.2.7 [37] with two simultaneous Markov Chain Monte Carlo (MCMC) runs of 2 million generations and tree sampling every 200000 generations. The GTR + I + G model was used in the BI analysis. The first 25% of trees were discarded as burn-in.
2.3.3. Species Delimitation
We used two different species-delimitation methods for the COI dataset of Sycanus. In the Estimates of Evolutionary Divergence over Sequence Pairs, according to the result of the twelve delimited species of 81 individuals based on the morphological characteristics, we used the Kimura-2 parameter (K2P) model [35] to compute intraspecific and interspecific pairwise genetic distances (p-distance) of twelve species and interspecific pairwise genetic distances of 81 individual using in MEGA X software (Table S2, 3).
2.4. Biological Study
Sycanus falleni Stål, 1863 and Sycanus croceus Hsiao, 1979 collected from Huashan, Ningming town, Guangxi province, China, were reared under laboratory conditions (26 ± 2 ℃, natural light) and fed on larvae of yellow mealworm, Tenebrio molitor. The annual life history was recorded. The egg and 1–5-instars nymphs were described and illustrated (Table S4, S5).
3. Results
3.1. Phylogenetic Relationships and Species Delimitation
3.1.1. Phylogenetic Relationships
In total, 81 ingroup individuals of twelve delimited species and 6 outgroup individuals of 4 related genera were included in the COI DNA barcodes dataset (Alignment S1), after trimming both ends of the alignment, 675 bp of the COI DNA barcoding sequences were used for the analyses. In the COI sequence matrix, number of constant sites is 433 (=64.1481% of all sites); number of invariant (constant or ambiguous constant) sites is 433 (=64.1481% of all sites); number of variant sites is 242 (=35.8519% of all sites); number of parsimony informative sites is 215 (=31.8519% of all sites).
The phylogenetic trees, constructed using three approaches (Bayesian Inference, BI; Maximum Likelihood, ML; Neighbor-Joining, NJ) based on the COI dataset, showed some differences in the topological structure of the clades. However, they provided strong support for both the species delimitation results and the monophyly of Sycanus that were based on the analysis of morphological characters (Figures 1, S1, S2, 2, 3, 4), with the exception of Sycanus bicolor Hsiao, 1979 and Sycanus croceovittatus Dohrn, 1859. The COI DNA barcodes sequence from Sycanus bicolor and Sycanus croceovittatus have been combined into a species in the phylogenetic trees and in the process of species delimitation.
Based on the BI, ML and NJ molecular phylogeny trees of COI DNA barcodes, Sycanus sichuanensis Hsiao 1979, Sycanus minor Hsiao, 1979, Sycanus taiwanensis Zhao & Cai sp. nov. and Sycanus hainanensis Wang & Cai sp. nov. form a clade, known as Sycanus sichuanensis species group; Sycanus croceus Hsiao, 1979, Sycanus insularis Hsiao, 1979, Sycanus rufus Hsiao, 1979 and Sycanus hsiaoi Li & Cai sp. nov. form a clade, known as Sycanus croceus species group. These two species groups are found to be sister groups (Figure 1, S1, S2). As for other species, Sycanus stali Dohrn, 1859 is a sister species of Sycanus sichuanensis species group and Sycanus croceus species group in the NJ trees, but it is categorized as part of the Sycanus croceus species group in BI and ML trees; Sycanus falleni is the basal part of ingroup in both ML and NJ trees, but it is considered a sister species of both the Sycanus sichuanensis species group and Sycanus croceus species group in the BI tree (Figure 1, S1, S2). Whereas the species complex consisting of Sycanus bicolor and Sycanus croceovittatus is the basal part of ingroup in the BI tree, but it is sister species of both the Sycanus sichuanensis species group and Sycanus croceus species group in the NJ tree.
The phylogeny and species delimitation based on COI DNA barcodes dataset failed to separate the two species, Sycanus bicolor Hsiao, 1979 and Sycanus croceovittatus Dohrn, 1859 (Figure 1, 2a,b, 3a,b, 4a,b, S1, S2,). So we added dataset of partial 28S rDNA sequences (639 bp, 34 ingroup individuals of five Sycanus species and 8 outgroup individuals of four related species) to solve the problem of the division of the two close related species and further built phylogeny trees (NJ) to analyze the relationship between the two species (Figure S3, Alignment S2). In the matrix of partial 28S rDNA sequences, number of constant sites is 536 (= 83.8811% of all sites); number of invariant (constant or ambiguous constant) sites is 536 (= 83.8811% of all sites); number of variant sites is 103 (= 16.12% of all sites); number of parsimony informative sites is 64 (= 10.0156% of all sites). The sequence matrix of partial 28S rDNA has a lower proportion of the parsimony informative sites than COI DNA barcodes dataset in the phylogeny analyses. In NJ phylogenetic trees based on partial 28S rDNA sequence, Sycanus croceus and Sycanus sichuanensis form a clade, and Sycanus croceovittatus and Sycanus bicolor form a clade, the two clades are sister group. Sycanus falleni is the basal part of ingroup (Figure S3, Alignment S2). The results showed that they are also two closely related species (Figure S3).
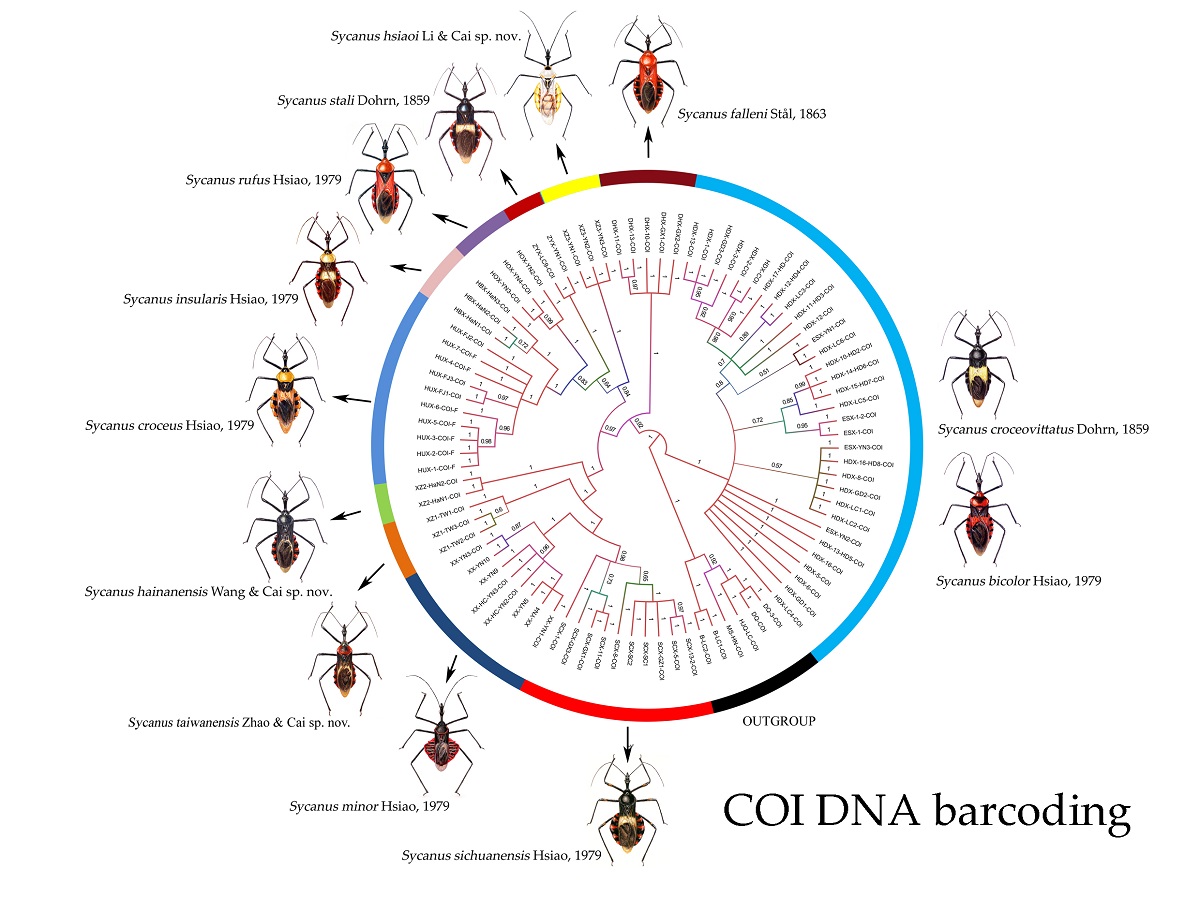
Figure 1.
Bayesian phylogenetic tree of cytochrome c oxidase subunit I (COI) sequence for 81 terminals of Sycanus; the results of Bayesian inference (BI) analyse agree with those of the two different species delimitation approaches, DNA barcoding gap and ASAP (The Assemble Species by Automatic Partitioning, P-distant, JC69, K80) analysis. The numbers above the branches are the posterior probabilities of the Bayesian inference (BI). The clades / species identified in this study are indicated in different colors. Distribution sites of materials used in molecular study for each Sycanus species in China were mapped.
Figure 1.
Bayesian phylogenetic tree of cytochrome c oxidase subunit I (COI) sequence for 81 terminals of Sycanus; the results of Bayesian inference (BI) analyse agree with those of the two different species delimitation approaches, DNA barcoding gap and ASAP (The Assemble Species by Automatic Partitioning, P-distant, JC69, K80) analysis. The numbers above the branches are the posterior probabilities of the Bayesian inference (BI). The clades / species identified in this study are indicated in different colors. Distribution sites of materials used in molecular study for each Sycanus species in China were mapped.
3.1.2. Species Delimitation
The ASAP analysis produced ten kinds of partition results when the ASAP score was 2.50–10.00 (10 group, 2.50; 7 groups, 3.00; 8 groups, 4.00; 11 groups, 6.50; 64 groups, 49 groups, 7.00; 37 groups, 8.00; 6 groups, 9 groups, 8.50; 47 groups, 10.00). The ASAP scale of the 10 groups partition is 2.50 and the lowest among them (The lower the score, the better the partition) [38]. However we selected and accepted the partition result of 11 groups (ASAP score: 6.50) by comparing it with the results of inter-specific pairwise genetic distances (Table S3) and morphological comparison. The 11 MOTUs delimited by ASAP were also well supported by NJ, BI, and ML trees. The difference between 11 groups and 10 groups partitions is mainly due to the close relationship of Sycanus sichuanensis Hsiao, 1979 and Sycanus minor Hsiao, 1979 (P-distant=3.02%). In addition, in the 9 groups partition, the relationship of Sycanus insularis Hsiao, 1979 and Sycanus croceus Hsiao, 1979 is too close to be divided in morphological classification, but P-distant between the two species is 4.28% (Table S2). So the partition result of 9 groups and 10 groups also is possible. But we integrated the results of morphological comparison and molecular phylogenetic analysis (p-distance >2%), we chose the partition result of 11 groups. The two species Sycanus bicolor Hsiao, 1979 and Sycanus croceovittatus Dohrn, 1859 are mixed together and cannot be distinguished by species delimitation of COI DNA barcodes dataset.
Figure 2.
Sycanus spp. in China, habitus. (a), Sycanus bicolor Hsiao, 1979; (b), Sycanus croceovittatus Dohrn, 1859; (c), Sycanus croceus Hsiao, 1979; (d), Sycanus falleni Stål, 1863; (e), Sycanus hsiaoi Li & Cai sp. nov.; (f), Sycanus hainanensis Wang & Cai sp. nov.; (g), Sycanus insularis Hsiao, 1979; (h), Sycanus minor Hsiao, 1979; (i), Sycanus rufus Hsiao, 1979; (j), Sycanus sichuanensis Hsiao, 1979; (k), Sycanus stali Dohrn, 1859; (l), Sycanus taiwanensis Zhao & Cai sp. nov.
Figure 2.
Sycanus spp. in China, habitus. (a), Sycanus bicolor Hsiao, 1979; (b), Sycanus croceovittatus Dohrn, 1859; (c), Sycanus croceus Hsiao, 1979; (d), Sycanus falleni Stål, 1863; (e), Sycanus hsiaoi Li & Cai sp. nov.; (f), Sycanus hainanensis Wang & Cai sp. nov.; (g), Sycanus insularis Hsiao, 1979; (h), Sycanus minor Hsiao, 1979; (i), Sycanus rufus Hsiao, 1979; (j), Sycanus sichuanensis Hsiao, 1979; (k), Sycanus stali Dohrn, 1859; (l), Sycanus taiwanensis Zhao & Cai sp. nov.

Figure 3.
Sycanus spp. in China, lateral view of head, pronotum and scutellum. (a), Sycanus bicolor Hsiao, 1979; (b), Sycanus croceovittatus Dohrn, 1859; (c), Sycanus croceus Hsiao, 1979; (d), Sycanus falleni Stål, 1863; (e), Sycanus hsiaoi Li & Cai sp. nov.; (f), Sycanus hainanensis Wang & Cai sp. nov.,; (g), Sycanus insularis Hsiao, 1979; (h), Sycanus minor Hsiao, 1979; (i), Sycanus rufus Hsiao, 1979; (j), Sycanus sichuanensis Hsiao, 1979; (k), Sycanus stali Dohrn, 1859; (l), Sycanus taiwanensis Zhao & Cai sp. nov..
Figure 3.
Sycanus spp. in China, lateral view of head, pronotum and scutellum. (a), Sycanus bicolor Hsiao, 1979; (b), Sycanus croceovittatus Dohrn, 1859; (c), Sycanus croceus Hsiao, 1979; (d), Sycanus falleni Stål, 1863; (e), Sycanus hsiaoi Li & Cai sp. nov.; (f), Sycanus hainanensis Wang & Cai sp. nov.,; (g), Sycanus insularis Hsiao, 1979; (h), Sycanus minor Hsiao, 1979; (i), Sycanus rufus Hsiao, 1979; (j), Sycanus sichuanensis Hsiao, 1979; (k), Sycanus stali Dohrn, 1859; (l), Sycanus taiwanensis Zhao & Cai sp. nov..
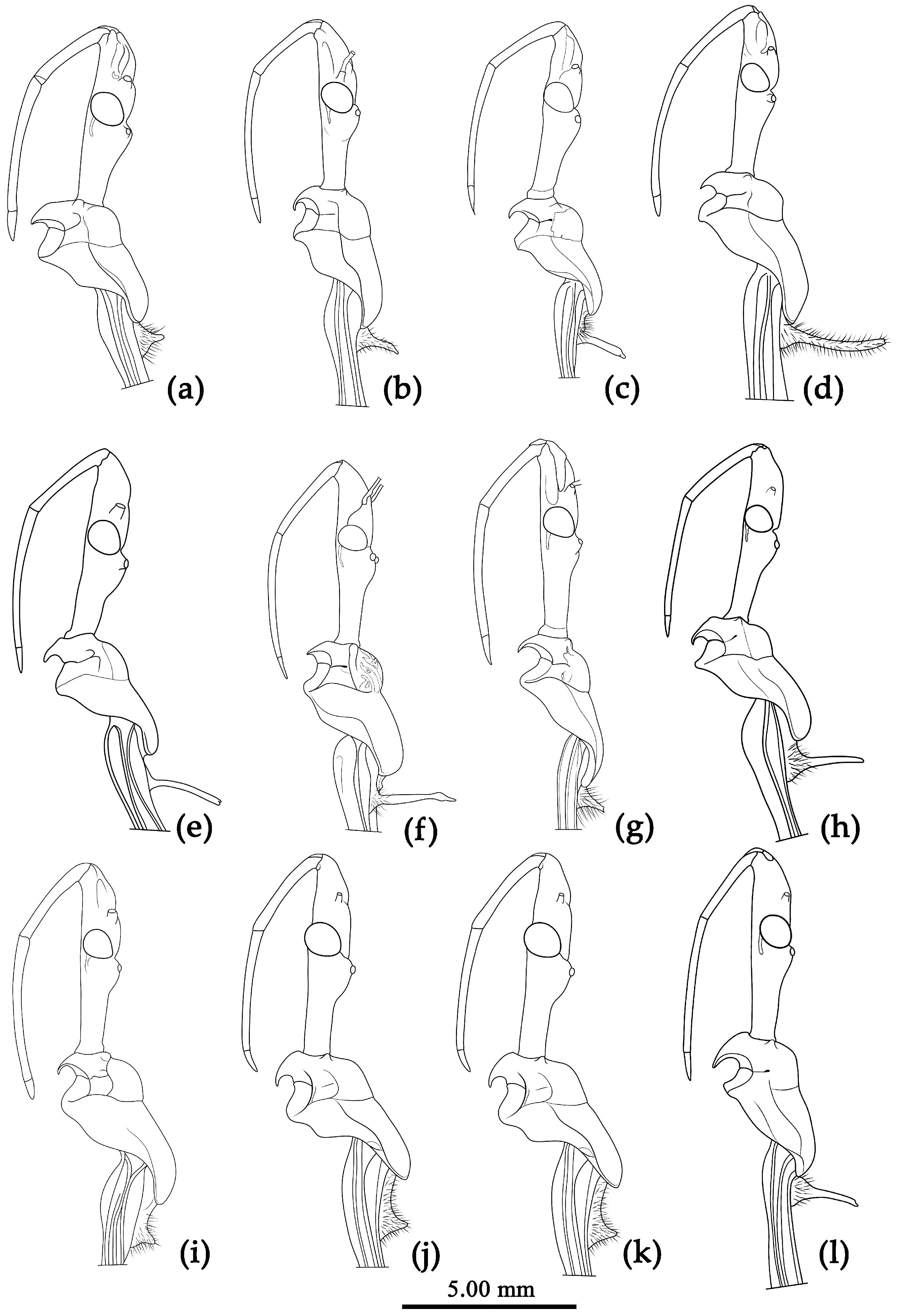
Figure 4.
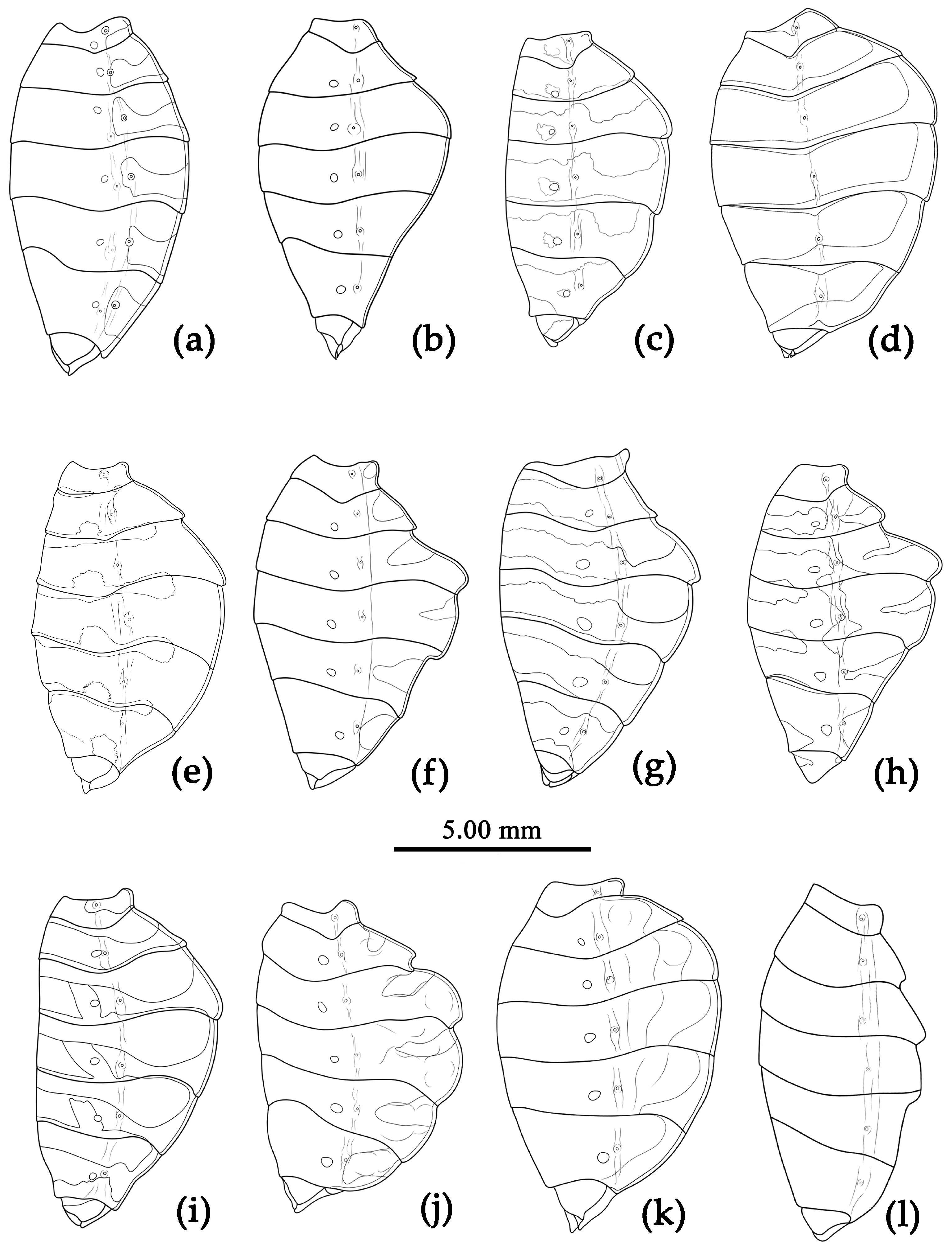
Sycanus spp. in China, lateral view of abdomen. (a), Sycanus bicolor Hsiao, 1979; (b), Sycanus croceovittatus Dohrn, 1859; (c), Sycanus croceus Hsiao, 1979; (d), Sycanus falleni Stål, 1863; (e), Sycanus hsiaoi Li & Cai sp. nov.; (f), Sycanus hainanensis Wang & Cai sp. nov.; (g), Sycanus insularis Hsiao, 1979; (h), Sycanus minor Hsiao, 1979; (i), Sycanus rufus Hsiao, 1979; (j), Sycanus sichuanensis Hsiao, 1979; (k), Sycanus stali Dohrn, 1859; (l), Sycanus taiwanensis Zhao & Cai sp. nov.
Figure 4.
Sycanus spp. in China, lateral view of abdomen. (a), Sycanus bicolor Hsiao, 1979; (b), Sycanus croceovittatus Dohrn, 1859; (c), Sycanus croceus Hsiao, 1979; (d), Sycanus falleni Stål, 1863; (e), Sycanus hsiaoi Li & Cai sp. nov.; (f), Sycanus hainanensis Wang & Cai sp. nov.; (g), Sycanus insularis Hsiao, 1979; (h), Sycanus minor Hsiao, 1979; (i), Sycanus rufus Hsiao, 1979; (j), Sycanus sichuanensis Hsiao, 1979; (k), Sycanus stali Dohrn, 1859; (l), Sycanus taiwanensis Zhao & Cai sp. nov.
3.2. Biology
3.2.1. Sycanus croceus Hsiao, 1979
We documented the annual life history of Sycanus croceus Hsiao, 1979 based on the live specimens distributed in Ningming, Guangxi, China (Figure 5; Table S4). They occur one generation per year. The overwintering adults occur in large numbers in May. The female in the experiment can lay an average of 60 eggs which are stuck to formed several egg mass. They prefer to lay eggs on the surface of plant stems (Figure 5a). There are five instars of nymphs (Figure 5b–f). Nymph stage is 156 ± 4.16 days from late June to early December (Table S4). The adults emerge from early October to late December. The adults or 5th nymph overwinter from early November to April of the following year, and adults peak in May (Figure 5g), then begin to reproduce in June and July. The adults generally die after oviposition in late June.
Egg. The egg is yellowish brown, 3.00 mm in length, 1.00 mm in width, and elongate oblong. The egg gradually changes from yellowish brown to reddish brown, with a metallic shine, operculum is white (Figure 5a).
Coloration of nymph. The nymph is orange with black markings (Figure 5b–f). Femora yellowish white, apically with a grep annular markings, medianly with black annular markings, and basally with an irregular black markings; tibiae and tarsus pale greyish brown, sub-basal part with a black annular markings, and apical parts of tibiae greyish brown. First to third antennal segments and base of fourth black, but two median annular markings of first, median annular markings of second greyish white, most of fourth reddish brown. Eyes black. Abdomen milk-white, with orange to red stripes, dorsally with black markings. The wing pad of third to fifth instar nymphs gradually from grey to black.
Structure of nymph. The body is covered with pale longer setae. First antennal segment and basal half of second sparsely covered with longer setae, apical half of second, third and fourth with yellowish brown short setae; legs sparsely covered with yellowish brown setae of various lengths. Body posteriorly widened. Leg and antennae slender. Third to fifth instar nymph with visible wing pad (Figure 5b–f).
Measurement of nymph [1st: 2nd: 3rd: 4th: 5th, in mm]. Body length 3.45:5.09:6.00:9.45:15.63; maximal width of abdomen 1.09:1.45:1.82:2.73:4.55; head length 1.09:1.64:2.18:2.73:4.55; length of antennae I–IV = 1.27:1.82:2.55:3.27:4.55 / 0.36:0.82:1.09:1.45:1.82 / 0.18:0.33:0.42:0.91:1.45 / 1.64:2.18:2.73:3.27:?; length of visible rostrum = 0.36:0.55:0.73:1.27:1.82 / 0.73: 1.09: 1.45: 2.18: 3.09 / 0.12:0.18: 0.36: 0.55:0.91; length of thorax 0.64:0.91:1.27:2.18:3.64; maximal width of pronotum 0.36:0.91:0.91:1.27:2.55; length of wing pad /:/:0.36:0.91:2.73.
3.2.2. Sycanus falleni Stål, 1863
We documented the part of the annual life history of Sycanus falleni Stål, 1863 based on the live specimens collected in Ningming, Guangxi, China (Table S5). They occur one generation per year in Ningming, Guangxi, China. Female adults lay eggs on the surface of leaves and stems of plants in an egg mass with more than 100 eggs (Figure 6a). There are five instars of nymphs (Figure 6b–f). The stage of five nymphs is 111.20 ± 3.35 days from early August to late December (Table S5). The adults emerge in December (Figure 6h).
Egg. The egg is yellowish brown, 3.50 mm in length, 1.00 mm in width, and elongate oblong. The egg gradually changes from yellowish brown to reddish brown, with a metallic shine, operculum is pale brown (Figure 6a).
Coloration of nymph. Body red with black markings (Figure 6). Leg black, femora with basal, median and subapical yellowish white annular markings, apical half and sub-basal part of tibiae blackish brown. First to third antennal segments and base of fourth black, two median annular markings of first, median annular markings of second greyish white, most of fourth brown. Eyes black. Rostrum black, apical part brown. Abdomen dorsally medially with black markings and dorsal abdominal gland, ventrally with black transversal stripes. The wing pad of third to fifth instar nymphs with black markings.
Structure of nymph. Body covered with pale setae. Body gradually posteriorly widened. Leg and antennae slender. Third to fifth instar nymph with visible wing pad.
Measurement of nymph [1st: 2nd: 3rd: 4th: 5th, in mm]. Body length 4.73:?:7.64:9.09:?; maximal width of abdomen 1.45:?:2.55:3.64:?; head length 1.45:?:2.18:3.27:?; length of antennae I–IV = 1.82:?:3.09:3.82:? / 0.73:?:1.00:1.45:? / 0.22:?:0.36:0.73:? / 2.18:?:3.27:4.55:?; length of visible rostrum = 0.55:?:0.82:1.09:? / 0.91:?: 1.64:1.64:?/0.36:?:0.36:0.55:?; length of thorax 1.27:?:2.36:2.73:?; maximal width of pronotum 073:?:1.45:1.82:?; length of wing pad /:/:0.40:1.45:4.20.
3.3. Systematics
- Genus Sycanus Amyot & Serville, 1843
- Hsiao, 1979: 154, 159, keyed nine Chinese species [31].
Diagnostic characters. Body of median to large size, mostly large in size. Head unarmed, spindle-shaped, slender and elongate, and longer than pronotum and scutellum together in length; rostrum slender, second rostral segment distinctly longer than first segment; first antennal segment sub-equal to fore femur in length. Pronotum unarmed and constricted before middle; apical angle of collar round; anterior pronotal lobe small and bulged, shorter than half of posterior lobe in length and width, its median longitudinal sulcus short and deeply depressed; posterior lobe wide, its surface rough or wrinkled; posterior and lateral angles obtuse and rounded; posterior margin nearly straight; scutellum round, apical part with a long spine or tubercle, apex of long spine bi-forked. Macropterous form, hemelytra long and passing abdominal apex; legs lender, femora sometimes sub-nodular; abdomen laterally dilated, connexivum strongly expanded dorsal-laterally, more or less undulated, lateral margins evenly curved.
Distribution. Oriental (79), Madagascar (1).
The key to the Chinese species in the genus Sycanus Amyot & Serville, 1843
- 1.
- Pronotum totally black...2
- -.
- Pronotum red or bicolor...8
- 2.
- Scutellum with short tubercle-shaped spine...Sycanus stali Dohrn, 1859
- -.
- Scutellum with long spine, apex bi-forked...3
- 3.
- Connexivum ventrally and dorsally with red markings…4
- -.
- Connexivum totally black…5
- 4.
- Corium black, posterior margin yellowish...Sycanus hainanensis Wang & Cai sp. nov.
- -.
- Corium yellowish, basal and apical parts black...Sycanus sichuanensis Hsiao, 1979
- 5.
- Basal half of hemelytron black, apical half yellow…6
- -.
- Most of hemelytron black...7
- 6.
- Apical half of corium pale golden-yellow...Sycanus croceovittatus Dohrn, 1859
- -.
- Apical half of corium pale stramineous...Sycanus collaris (Fabricius, 1781)#
- 7.
- Hemelytron black, apical margin yellow…Sycanus bifidus (Fabricius, 1787)#
- -.
- Hemelytron greyish yellow, only basal part black…Sycanus fuscirostris Dohrn, 1859#
- 8.
- Apical spine of scutellum long, apex biforked…9
- -.
- Apical spine of scutellum very short, apex un-biforke…14
- 9.
- Pronotum and corium of hemelytron totally red…Sycanus falleni Stål, 1863
- -.
- Pronotum and corium of hemelytron partly red…10
- 10.
- Posterior pronotal lobe red, median part with black markings; fourth and sixth abdominal connexiva laterally expended…11
- -.
- Posterior pronotal lobe yellow or greyish yellow; abdomen laterally roundly expended…12
- 11.
- Abdomen and connexivum red or yellow with black markings...Sycanus minor Hsiao, 1979
- -.
- Abdomen totally black, connexivum bicolor...Sycanus taiwanensis Zhao & Cai sp. nov.
- 12.
- Corium of hemelytron totally yellowish...Sycanus hsiaoi Li & Cai sp. nov.
- -.
- Basal part of corium of hemelytron black, apical half yellow...13
- 13.
- Posterior pronotal lobe and median transversal markings of hemelytron pale grayish yellow…Sycanus insularis Hsiao, 1979
- -.
- Posterior pronotal lobe orange, most of corium of hemelytron (except inner side and most of clavus black) yellow or orange…Sycanus croceus Hsiao, 1979
- 14.
- Pronotum and corium of hemelytron totally red; ventral surface of head paler; abdominal sterna yellow, inter-segment with black stripe and lateral side with black round markings; black markings of sixth and seventh connexival segments not extending to outer margin...Sycanus rufus Hsiao, 1979
- -.
- Pronotum and corium of hemelytron bicolor, black and red; ventral surface of head black; abdominal sterna black; black markings of connexivum totally extending to outer margin......Sycanus bicolor Hsiao, 1979
- #Notes. Sycanus collaris (Fabricius, 1781), Sycanus bifidus (Fabricius, 1787) and Sycanus fuscirostris Dohrn, 1859 have been recorded in China, but we have not examined the specimen for the three species. They are not included in this study.
- (1) Sycanus bicolor Hsiao, 1979二色犀猎蝽
- (Figures 2a, 3a, 4a , 7, 8, S4)
Redescription. Coloration. Body black, with red markings (Figure 7). Anterior pronotal lobe, propleural episternum, posterior 1/3 of posterior pronotal lobe, scutellum (except posterior margin), apical half of corium of hemelytron (except apical angle black), most of meso- and meta- pleura, coxae, posterior half of each connexival segment red; abdomen black, abdomen ventrally totally black,or sometimes laterally with red markings at segmental sutures; rostrum black, posterior half of second and third segments gradually paler, brown.
Structure. Body large-sized. Head beneath, pronotum, pleura and sterna of thorax, corium clothed with procumbent white short setae; head (except ventral surface), pronotum and propleuron, scutellum, legs, stetna of abdomen clothed with longer setae; first antennal segment sparsely with oblique short setae, second segment densely with short setae, third and fourth segments with procumbent short setae. Head longer than pronotum, constricted in middle, anterior lobe sub-equal to 1/2 of posterior lobe in length; post-ocular area of head round, then posteriorly thinned, and longer than anti-ocular area; ocelli separated, distance between ocelli longer than distance between ipsolateral ocellus and eyes. Anterior pronotal lobe hemispheric, smooth, median longitudinal sulcus short and deep; posterior pronotal lobe with irregular wrinkles; lateral pronotal angle round and obtuse; posterior and posterior-lateral margins of pronotum nearly straight; apical spine of scutellum situated at middle part, thick and short, produced upwards (Figure 3a, 7b). Hemelytron extending beyond tip of abdomen. Abdomen laterally dilated. Pygophore elliptic, median pygophore process shown in Figure 8a, b; paramere clavate, apical half swelled with thick setae, middle part bent (Figure 8c, d). Struts triangular (Figure 8e); phallosoma elliptic; dorsal phallothecal sclerite sclerotized; endosoma apically with a horned process and laterally with about 5 pairs of small spines (Figure 8f-h).
Measurement [♂(n=4)/♀(n=3), in mm]. Body length 17.21–18.47 / 20.75–23.09, maximal width of abdomen 5.70–7.41 / 7.41–7.70; Head length 4.56–4.85 / 5.13–5.42; length of ante-ocular part 1.60–1.80 / 1.82–2.00; length of post-ocular part 1.82–2.11 / 2.34–2.39; length of synthlipsis 0.68–0.80 / 0.86; distance between ocelli 0.40–0.46 / 0.46-; length of antennal segments I–IV = 4.62–5.70 / 4.85–5.24, 2.28–2.30 / 2.00, 1.03–1.10/ 0.97, 7.41–7.50/ 3.99–5.13; length of visible rostral segments I–III = 2.00–2.39 / 2.39–2.51, 3.14–3.71 / 3.99–5.42, 0.63–0.68 / 0.71–0.80; length of anterior pronotal lobe 1.14–1.25 / 1.31–1.37; length of posterior pronotal lobe 1.73–2.00 / 2.57–2.68; maximal width of pronotum 3.53–3.93 / 4.85; length of scutellum 0.68–1.03 / 1.43–1.48; length of hemelytron 10.83–11.40 / 13.00–13.68.
Type material. Holotype, ♂, CHINA, Yunnan, Xishuangbanna, Mengzhe, 1400m, 15-VI-1958, Zhu Zhibin leg.(IOZ). Allotype, ♀, same as holotype, 15-VI-1958, Cheng Hanhua leg. (TJNHM). Paratypes (Figure S1), 2♂, 3♀, CHINA, Yunan, Xishuangbanna, Xiaomengyang, Mengzhe, Damenglong, Luxi and river valleys in west area of Nujiang River (IOZ); 1♀, CHINA, Yunan, Ruili, 1400 m, 6-VI-196, Zhou Bentao leg. (IOZ).
Specimens examined. CHINA: 2♂, Yunan, Yuanjiang, 20-VII-1976, Xiong Shaoda leg. (CAU); 1♀, Yunnan, Yongdeng, Hongcheng, 4-X-1980, Xi Gengsi leg. (CAU); 3♂, 3♀, Yunan, Mengla, Yaoqu, 2005-V-9, 850 m, N21°43′, E101°32′, Bai Xiaoshuan & Cui Jianxin leg. (CAU); 1♀, Yunan, Mengla, Shangyong, 2005-V-18, Cui Jianxin leg. (CAU); 1♂, 1♀, Yunnan, Mengla, 1979-IX-18, 650 m, Zheng Leyi leg. (NKU); 1♂, Yunnan, Xishuangbanna, Xiaomengyang, 1957-IX-12, Wang Shuyong leg. (IOZ); 2♂, 1♀, Yunnan, Mangshi, 1000m, 1956-VI-3-4, 1959-VI-29, Huang Tianrong & Zhou Benshou leg. (NKU); 1♂, 1♀, Yunnan, Jingdong, 1170 m, 1958-VI-16, 1958-VI-14 (NKU); 1♂, Yunnan, Zhenyuan, 1400 m, VI-23 (IOZ); 1♂, 1♀,Yunnan, Xishuangbanna, Mengzhe, 1958-VI-13, 1200 m, Pu Zhelong leg. (IOZ); 2♂, Yunnan, Xishuangbanna, Mengzhe, 1400m, 1958-VI-14 (NKU); 1♀, Yunnan, Xishuangbanna, Menglong, 650 m (IOZ); 1♂, Yunnan, West Nujianghegu, 800 m, 1955-V-9 (NKU); 1♂, Yunnan, Longkou, Longjiang, Mengqiao, 1979-6-15, 1650 m (NKU); 1♂, Yunnan , Lianghe (NKU).
Distribution. China (Yunnan [Luxi, river valleys in west area of Nujiang River, Yuanjiang, Jingdong, Hongcheng, Mangshi, Mengla, Menglong, Mengzhe, Xiaomengyang, Damenglong, Lianghe, Zhenyuan, Yongdeng, Ruili]).
Remark. After we examined the syntype of Sycanus miles Walker, 1873 (Penang) kept in the Natural History Museum, London, we found that Sycanus bicolor Hsiao, 1979 is similar to Sycanus miles in body color and structure. However Distant have assigned Sycanus miles as a synonym of Sycanus versicolor Dohrn, 1859 (India) [27,44,45]. We haven’t examined the type of S. versicolor, so this species needs further study.
- (2) Sycanus bifidus (Fabricius, 1787)
- Reduvius bifidus Fabricius, 1787: 312; Zimsen, 1964: 339 [46]. Syntypes, 3 specimens, China, Hong Kong.
- Cimex bifidus: Gmelin, 1790: 2200 [47].
- Zelus bifidus: Fabricius, 1803: 285 [48].
- Harpactor bifidus: Westwood, 1842: 50.
Remark. We examined the specimens of Sycanus bifidus (Fabricius, 1787) collected from Malaya and kept in British Museum. But we didn’t examine type species of Sycanus bifidus. Syntypes of the species are kept in ZMUC (Zoological Museum. University of Copenhagen, Copenhagen. Denmark), 3 specimens, collected from China, Hong Kong. In the basis of Distant’s description [29], the three species, Sycanus bifidus Fabricius, 1787, Sycanus collaris (Fabricius, 1781) and Sycanus croceovittatus Dohrn, 1859 are similar in body coloration. We collected many specimens of Sycanus croceovittatus from Hong Kong, China, and doubted that the species distributed in Hong Kong is Sycanus croceovittatus. Then Sycanus croceovittatus Dohrn, 1859 may be synonym of Sycanus bifidus (Fabricius, 1787).
Distribution. China (Hong Kong); Java, Borneo, India, Bengal.
- (3) Sycanus collaris (Fabricius, 1781)
- Reduvius collaris Fabricius, 1781: 380[22]. India.
- Zelus collaris: Fabricius, 1803: 285[48].
- Arilus collaris: Burmeister, 1835: 229 [49].
- Sycanus leucomesus Walker, 1873: 84[44], Myanmar (Burma).
Distribution. Philippines, Malay Archipelago, Malay Peninsula, Sri Lanka,Thailand, Malacca, Sarawak, India, Assam, China?.
Remark. In the world catalogue of the assassin bugs [24], the type species of genus Sycanus, Sycanus collaris (Fabricius, 1781), has been recorded to be distributed within China, but we have consulted a large number of documents and has no records in China. They are mainly distributed in Southeast Asian countries. We have doubts about the distribution of this species in China, but listed them in the paper.
- (4) Sycanus croceovittatus Dohrn, 1859
- (Figures 2b, 3b, 4b, 9, 10, S8c,d)
Redescription. Coloration. Body black, slightly shiny; apical half of corium of hemelytron yellow or orange; membrane of hemelytron pale brown, semitransparent; spot between ipsolateral eye and ocellus yellowish brown; each sternum of abdomen laterally with two round white spots (Figure 9).
Structure. Body large-sized. Body mostly clothed with black long or short setae and yellow procumbent short setae; first antennal segment sparsely with long setae, second segment densely with short setae, third and fourth segments densely with procumbent short setae; anterior margin of hemelytron with brown bent short setae; legs with black long setae; each segment of sternum of abdomen laterally with two white setae floccus. Anterior lobe of head sub-equal to posterior lobe, anti-ocular part distinctly shorter than post-ocular part; rostrum incurved, long and slender. Collar indistinct; anterior pronotal lobe small, hemisphered and bulged, deeply depressed at base; posterior pronotal lobe rugose, lateral pronotal angle obtuse and round, posterior margin nearly straight, posterior angle nearly absent; scutellum with an erect upward, thick, bi-forked spine (Figure 9). Femora nearly of equal thickness, apical part somewhat thickened; hemelytron extending beyond tip of abdomen. Fourth and sixth connexival segments of abdomen laterally expended. Pygophore elliptic, median pygophore process shown in Figure 10a, b; paramere clavate, apical half swelled with thick setae, middle part bent (Figure 10c, d). Struts triangular (Figure 10e); endosoma apically with a horned process and laterally with about 7 pairs of small spines (Figure 10f–i).
Measurement [♂(n=21)/♀(n=19), in mm]. Body length 22.31–23.00 / 21.8–24.52; maximal width of abdomen 6.83–8.70 / 6.83–9.80. head length 5.25–5.41 / 5.36–6.14; length of ante-ocular part 1.84–1.94 / 2.00–2.21; length of post-ocular part 2.36–2.52 / 2.36–3.05; distance between ocelli 0.39–0.50 / 0.45–0.53; length of synthlipsis 0.79–0.89 / 0.84–0.95; length of antennal segments I–IV= 6.46–6.93 / 6.20–7.51, 2.21–2.52 / 2.26–2.36, 1.21–1.73 / 1.21–1.42, 7.35–7.46 / 7.88–9.19; length of visible rostral segments I–III= 2.52–2.68 / 2.84–3.36, 3.89–4.10 / 3.83–5.07, 0.74–0.79 / 0.74–0.89; length of anterior pronotal lobe 1.31–1.37 / 1.13–1.50; length of posterior pronotal lobe 2.31–2.78 / 2.63–3.57; maximal width of pronotum 4.31–4.78 / 4.73–6.46; length of scutellum 1.31–1.42 / 1.10–1.21; length of hemelytron 13.39–15.12 / 14.18–16.80.
Specimens examined. CHINA, Yunnan: 1♀, Yunan, Mengyang, 1991-VI-8, 800m, Cai Wanzhi leg.(CAU); 2♂, Yunnan, Menglun, 1991-V-18, Cai Wanzhi leg.(CAU); 1♀, Yunnan, Mengla, 1991-V-28, Cai Wanzhi leg.(CAU); 1♂, Yunnan, Menghai, 1991-V-30, Cai Wanzhi leg.(CAU); 1♀, Yunan, Mengla, Yaoqu, 2005-V-9, Bai Xiaoshuan leg.(CAU); 1♂, 1♀,Yunnan, Wenshan, Malipo, 2005-VIII-17(CAU); 4♂, 1♀, Yunnan, Lvchun, Huanglian mountain, 2012-V-10, Cai Wanzhi & Niu Xinwei leg.(CAU); 2♂, 1♀, Yunnan, Banna, Jinghong, Jinuo, 1053m, 2021-VII-25, Chen Zhaoyang & Liu Qinpeng leg.(CAU); 1♀, Yunnan, Honghe, Jinping, Fenshuiling National Nature Reserve, 2012-IX-24(CAU); 1♀, Yunnan, Honghe, Yuanjiang, 2014-VIII-18(CAU); 1♂, Yunnan, Puer, Mojiang, Sinanjiang, Dashaba, 2005-IX-5(CAU); 1♂, Yunnan, Yingpan Town, Fengqing County, Lincang, 2005-VI-20(CAU); 1♂, 1♀, Yunnan, Wenshan, Malipo, 2005-VIII-17(CAU); 2♂, 1♀, Yunnan, Banna, Jinghong, Jinuo, 1053m, 2021-VII-25(CAU); 6♂, 2♀, Yunnan, Hekou, 2011-V-20(CAU); 1♂, Yunnan, Xishuangbanna, Mengla, Yaoqu, 2013-III-24(CAU).
CHINA, Guangxi: 5♂, 8♀, Guangxi, Shangsi, Shiwandashanm, 2006-VIII-28, 1♂, 1♀, 2006-IX-2, Huang Xia leg.(CAU); 6♂, 4♀, Guangxi, Ningming, Longrui, 2006-V-13–22, Huang Xia & Shi Zhongting leg.(CAU); 1♂, Guangxi, Huaping, 1963-VI-6, Yang Jikun leg.(CAU); 2♀, Guangxi, Nanning, Fusui, 2004-VIII-18/20, 240m, Zhang Kuiyan leg.(CAU); 2♂, Gungxi, Longzhou, Nonggang, 1982-V-19, 240m, Wang Xinli leg.(CAU); 1♂, Guangxi, Jinxiu, 1982-V-10, 720 m, Wang Xinli leg.(CAU); 1♂, Guangxi, Ningming, Longrui, 1984-V-25, 180 m, Li Fasheng leg.(CAU); 1♂, Guangxi, Baise, Napo, Baisheng, 2020-V-26, Zhao Ping leg.(NNU); 5♂, 7♀, Guangxi, Baise, Napo, 2020-V-26(NNU); 1♀, Guangxi, Nanning, Wuming, Daming Mountain, 2016-VIII-8–10, Zheng Yuchen leg.(CAU); 1♂, Guangxi, Baise, Leye, 1200m, 2017-V-24(CAU); 4♂, 4♀, Guangxi, Ningming, Longrui Nature Reserve, 2006-V-17, Huang Xia & Shi Zhongting leg(CAU).; 1♀, Guangxi, Nanning, Fusui, 2004-VIII-18, 200m, Zhang Kuiyan leg.(CAU); 5♂, 7♀, Guangxi, Baise, Napo, 2020-V-26(NNU); 3♂, 1♀, Guangxi, Chongzuo, Longzhou, Zhubu, 2020-V-26(NNU); 4♂, Guangxi, Fangchenggang, Shangsi, Shiwandashan, 2006-VIII-28, Huang Xia leg.(CAU).
CHINA, Guangdong: 1♂, Guangdong, Meixian, 1981-IX-6 (CAU); 1 ♂, Guangdong, Yangjiang, Yangchun, Mashui, 2002-IV-30 (CAU); 1♂, Guangdong, Huizhou, 2004-VIII-19 (CAU); 1♂, Guangdong, Meixian, 1981-IX-6 (CAU); 2♀, Hong Kong, 2019-V-21–22 (CAU).
CHINA, Hainan: 1♂, 3♀, Hainan, Baisha, Yinggeling, 2008-X-8, Zhang W.J. leg.(CAU); 1♀, Hainan, Wuzhishan, Fanyang, Bulun, 2008-X-26, Zhang W.J. leg.(CAU); 3♂, Hainan, Baisha, Nankai, Mohao, 2008-V-30, Zhang W. J. leg.(CAU); 1♂, Hainan, Ledong, Jianfengling, 2007-V-10, Zhang W.J. leg.(CAU);1♂, Hainan, Baisha, Nankai, 2008-IV-28 Zhang W.J. leg.(CAU).
Distribution. CHINA (Guangdong <Baiyun Mountain, Gaoyao, Guangzhou, Meixian>, Guangxi <Fangcheng Fulong, Shangsi, Nanning, Napo, Ningming, Jinxiu, Longzhou, Huaping>, Yunnan <Jinping, Hekou, Xishuangbanna: Mengban, Menghai, Menglun, Mengyang, Mengla>, Guizhou <Maolan, Guiyang, Wangmo, Guiding>, Hunan, Fujian <Fuzhou, Hua’an>, Hainan<Jianfengling>, Hong Kong); MYANMAR, INDIA, VIETNAM, MALAYSIA ? (Federated Malay States).
Remark.Sycanus croceovittatus is set up by Dohrn in 1859 with Syntypes, 5 speceis (♀/♂) collected from China and kept in Zoologisches Museum, Humboldt University, Berlin, Germany. We have examined the type species of Sycanus leucomesus Walker, 1873, and Sycanus leucomesus is a synonym of Sycanus croceovittatus or Sycanus collaris (Fabricius, 1781) [24,32].
- (5) Sycanus croceus Hsiao, 1979
- (Figures 2c, 3c, 4c, 5, 11, 12, S5)
Redescription. Coloration. Body orange, with black markings (Figure 11). Head (except ventral surface and markings around ocellus orange), antennae, eyes, anterior pronotal lobe, scutellum (except apical spine and posterior margin orange), clavus (except basal part yellow), inner side and apical angle of corium, leg (except coxae yellow), meso-sternum (except anterior and posterior margins yellow), meta-sternum, anterior half of meso- and meta-pleuron, basal part of scutellum, segmental strip of abdomen, large markings of connexivum, discontinuous medial longitudinal strip and small round spots laterally on ventral surface of abdomen black; membrane of hemelytron pale brown, semitransparent, with metallic-shiny; first visible rostral segment and basal part of second black, most of second and third brown.
Structure. Body median to large-sized. Body clothed with yellowish short setae; legs with vertical long setae; thorax densely with short setae (Figure 11). Post-ocular area of head longer than anti-ocular area; antennae long and slender, first segment longest, third shortest, fourth slightly shorter than first; rostrum long, bent, first extending beyond middle part of eyes, second segment longest. Anterior angle of pronotum round; anterior pronotal lobe small, hemisphere, posterior part with short longitudinal sulcus; posterior pronotal lobe rough, lateral angle round; posterior margin of scutellum round, apical spine sub-vertical, apex bi-forked (Figure 11a); legs slender; hemelytron extending beyond apex of abdomen; fourth to sixth connexival segments roundly laterally dilated (Figure 11). Pygophore elliptic, median pygophore process “Y”-shaped (Figure 12a, b); paramere clavate, apical part with thick setae, middle part slightly bent (Figure 12a-c). Struts triangular (Figure 12d); middle part of endosoma with two horn-shaped processes (Figure 12e, f) and apical part laterally with about 11 pairs of small spines (Figure 12e-g).
Measurement [♂(n=3)/♀(n=3), in mm]. Body length 19.64–20.74 / 20.91–25.20; maximal width of abdomen 7.27 / 8.18. Head length 4.73 / 5.09–5.46; length of anteocular part 1.63–1.64 / 1.82–1.84; length of postocular part 2.36–2.52 / 2.55–2.84; distance between ocelli 0.47–0.55 / 0.47–0.55; length of synthlipsis 0.79 / 0.72–0.84; length of antennal segments I–IV = 6.73–6.83 / 6.18–7.30, 2.21 / 2.36–2.63, 1.64–1.94 / 1.64–2.10, 6.55 / 5.09–6.56; length of visible rostral segments I–III = 2.26–2.36 / 2.55–2.68, 3.41–3.64 / 3.04–4.04, 0.55–0.71 / 0.36–0.74; length of anterior pronotal lobe 1.21–1.27 / 1.27–1.37; length of posterior pronotal lobe 2.36–2.55 / 2.55–3.07; maximal width of pronotum 3.82–4.15 / 4.55–5.36; length of hemelytron 13.64–13.65 / 14.18–17.06.
Holotype, ♀, CHINA, Yunnan, Hekou, Xiaonanxi, 200m, 11-VI-1956, Huang Keren et al. leg. Allotype, same as Holotype, 10-VI-1956. Paratype, 1 ♀, CHINA, Guangxi, Baishou; 1 ♀, Yunnan, Hekou (ZOI).
Specimens examined.
CHINA, Guangxi: 1♂, 1♀, Guangxi, Jinxiu, 1983-VI-13 (NKU); 1♀, Guangxi, Longsheng, Huaping, 1983-VI-13 (NKU); 1♂, 1♀, Guangxi, Jinxiu, 1990-VI-12, 800 m, Li Xinzheng leg. (NKU); 1♀, Guangxi, Jinxiu, 2005-VII-24, Huang Xia leg. (CAU); 1♂, Guangxi, Laibin, Jinxiu, Dayaoshan, 1250m, 2015-V-7 (CAU); 1♀, Guangxi, Jinxiu, Dayaoshan, Luoyingou, 1100m, 2016-V-24, Zhao Jintang leg. (CAU); 2♀, Guangxi, Laibin, Jinxiu, Dayaoshan, 1100m, 2016-V-31, Zhao Jintang leg. (CAU); 1♀ Guangxi, Jinxiu, Dayao Mountain, 1100m, 2017-VII-20, Zhao J. T. leg. (CAU); 2♀, Guangxi, Jinxiu, Dayaoshan, 600m, 2018-V-30 Zhao J. T. leg. (CAU); 1♂, 1♀, Guangxi, Longsheng, Huaping, 1963-V-1, Liu Sikong leg. (NKU); 1♂, Guangxi, Huaping, Cujiang, 1963-IV-8, Yang Jikun leg.(CAU); 1♀, Guangxi, Huaping, Tianping mountain, 1963-IV-5, Yang Jikun leg.(CAU); 1♂, 1♀, Guangxi, Guilin, Huaping, Tianping mountain, 1963-VI-5–8, Yang Jikun leg. (CAU); 1♂, Guangxi, Longsheng 1992-V-23, Liu Guang leg. (CAU); 1♀, Guangxi, Jinxiu, 2005-VII-23, Zhao Ping leg. (CAU); 2♂, 2♀, Guangxi, Laibin, Jinxiu, Dayaoshan, 900m, 2015-V-14, Lu Yanquan leg. (CAU); 1♂, Guangxi, Longzhou Nonggang, 2019-IV-22, He Zhuqing leg. (CAU); 1♀, Guangxi, Chongzuo, Longzhou, Nonggang Nature Reserve, 2020-V-27 (CAU); 2♀, Guangxi, Laibin, Jinxiu, Fenzhan, 810m, 2020-VI-24 (CAU); 3♂, 5♀, Guangxi, Ningming, Huashan, 2014-VIII-13, 2020-VI (NKU);1♂, 1♀, Guangxi, Baishou, 1957-VI-27 (NKU); 1♀, Guangxi, Xing’an, Jinshi, 2007-VII (CAU); 1♀, Guangxi, Baise, Jingxi, Bangliang, Renzhuan, 2010-VIII-1, Zhou S.Y. leg. (CAU); 1♂, Guangxi, Baise, Napo, 2020-V-26 (NNU); 3♀, Guangxi, Hezhou, Gupo mountain, 2011-VII (CAU);
CHINA, Yunnan: 1♂, 1♀, Yunnan, Honghe, Hekou, Nanxi, Huayudong, 200m, 2016-IV-24, Wang Yutang & Yang Xiaodong leg. (CAU); 1♂, Yunnan, Xishuangbanna, Mengla, Mengxing, 1981-V-23, Yang Pingzhi leg. (CAU); 1♀, Yunnan, Xishuangbanna, Menghai, 1984-VI-21, Xiong Jiang leg. (CAU); 1♀, Yunnan, Menghai, 1984-VI-21, Xiong Jiang leg (CAU).; 1♂, Yunnan, Mengxing, 1981-V-23, Yang Pingzhi leg.(CAU); 1♂, Guangxi, Ren Shuzhi leg. (NKU);
CHINA, Guangdong: 2♀, Guangdong, Guangzhou, Conghua, Liuxihe, 2004-VI-20 (CAU); 1♀, Guangdong, Shixing, Chebaling Nature Reserve, 2003-IX-15 (CAU); 1♂, Guangdong, Nanling Nature Reserve, 2017-VI-18, Zhao Yisheng leg. (CAU); 1♀, Guangdong, Shaoguan, Wujiang, 2016-V-28, Ge Zhentai leg. (CAU); 1♂, 1♀, Guangdong, Shaoguan, Qujiang, Xiaokeng National Forest Park, 2013-V-1, Zheng Chaowu leg. (CAU); 1♂, 3♀, Guangdong, Shaoguan, Qujiang, Xiaokeng National Forest Park, 2013-X-1, Zheng Chaowu leg. (CAU); 3♂, 2♀, Guangdong, Nanling Nature Reserve, 2017-VI-18, Zhao Yisheng leg. (CAU);
CHINA, Jiangxi: 1♀, Jiangxi, Ganzhou, Longnan, Hengkeng, Jiulian Mountain, 500m, 2020-VII-05, Zheng Yuchen leg. (CAU); 1♂, 1♀, Jiangxi, Ganzhou, Chongyi, Yangling National Forest Park, 2014-VI-18, Li Yanjing leg. (CAU);
CHINA, Guizhou: 1♂, Guizhou, Rongjiang, Pingyang, Xiaodanjiang, 2005-V-31, Song Yuehua leg. (CAU); 1♂, Guizhou, Rongjiang, Pingyang, 2005-VI-1, Song Yue Hua leg (CAU);
CHINA, Fujian: 2♂, 2♀, Fujian, Quanzhou, Dehua, Shangyong, 2014-VI-25 (CAU); 1♀ Fujian, Wuyi Mountain, 2010-VII-8, Luo X.Y. leg. (CAU); 1♂, 2♀, Fujian, Quanzhou, Dehua, Chishui , 2014-VI-20 (CAU).
Distribution. CHINA (Guizhou <Rongjiang>, Yunnan <Hekou Nanxi, Menghai, Mengxing>, Guangxi <Baishou, Jinxiu, Huaping, Rong’an, Longzhou, Ningming, Du’an, Longsheng, Napo, Fangcheng, Fulong, Longzhou Nonggang>, Guangdong <Conghua, Chebaling, Shaoguan, Nanling>, Jiangxi (Ganzhou), Fujian (Fuzhou, Sanming, Longyan, Dehua, Hua’an, Yongding, Longqishan, Quanzhou); VIETNAM.
- (6) Sycanus falleni Stål, 1863
- (Figures 2d, 3d, 4d, 13, 14, S6, S8a,b)
Redescription. Coloration. Body red, with black markings. Head, antennae, legs (except coxae), rostrum (except apical part brown), most of meso- and meta-pleuron (except posterior part red), sternum of meso- and meta-thorax, sterna and connexiva of abdomen (except inter-segmental stripe of abdominal sterna yellow, and inter-segmental stripe and lateral margins of connexiva red), posterior half of clavus of fore wing, basal part of scutellum black, and with blue shiny; pronotum and pro-pleuron, pro-sternum (except middle part), posterior half of meso- and meta-pleuron, scutellum, coxae, inter-segmental stripe and lateral margins of connexivum, apical part of abdomen red; corium of hemelytron red, clavus black (except basal part red), membrane pale brown and semitransparent (Figure 13).
Structure. Body large-sized. Body clothed densely with yellowish short setae and depressed short setae; legs with vertical longer setae; thorax, pronotum densely with short setae. Head longer; post-ocular area of head longer than anti-ocular area; antennae long and thin, fourth segment longest and sub-equal to first, third shortest and sub-equal to second; rostrum long, bent, first extending to middle part of eyes, second segment longest. Anterior angle of pronotum round and indistinct; anterior pronotal lobe small, hemisphere, posterior part with short longitudinal sulcus; posterior pronotal lobe rough, lateral angle round; posterior margin of scutellum round, apical spine sub-vertical, apex bi-forked; legs slender; hemelytron extending beyond apex of abdomen; fourth to sixth connexival segments dilated laterally (Figure 13). Pygophore elliptic, median pygophore process shown in Figure 14a, b; paramere clavate, apical half swelled with thick setae, middle part bent (Figure 14a-c). Struts triangular (Figure 14d); endosoma apically laterally with more than 30 pairs of small spines and medianly with a pair of sclerite (Figure 14e-g).
Measurement [♂(n=10)/♀(n=10),in mm]. Body length 23.63–24.46 / 24.94–26.62; maximal width of abdomen 6.56–8.66 / 8.14–8.40. head length 5.46–5.51 / 5.51–6.20; length of ante-ocular part 1.79–1.84 / 2.10–2.15; length of post-ocular part 2.78 / 2.36–2.89; distance between ocelli0.58 / 0.53–0.58; length of synthlipsis0.84–0.89 / 0.89–1.00; length of antennal segments I–IV = 7.09–7.35 / 6.83–7.25, 2.52–2.57 / 2.52–2.63, 2.15–2.21 / 2.00–2.31, 7.35–7.61 / 6.83–7.35; length of visible rostral segments I–III = 2.63–2.78 / 2.84–3.05, 4.10–4.41 / 4.46–4.73, 0.84–0.89 / 0.79–0.89; length of anterior pronotal lobe1.31–1.37 / 1.50–1.58; length of posterior pronotal lobe2.52–2.78 / 3.15–3.57; maximal width of pronotum4.67–4.99 / 5.57–6.41; length of scutellum1.42–1.47 / 1.47–1.58; length of hemelytron15.23–16.28 / 16.80.
Type species. The type species are kept in Swedish Museum of Natural History, and the photograph is from website: “http://www2.nrm.se/en/het_nrm/f/sycanus_falleni.html”.
Specimens examined. CHINA: 10♀, 2♂, Guangxi, Ningming, Longrui, 2006-V-18, Huang Xia & Shi Zhongting leg. (CAU); 1♀, Guangxi, Ningming, Longrui, 1984-V-20, Lu Xiaolin & Wu Zheng Liangcai leg.(CAU); 2♀, Guangxi, Longrui, 1984-V-23, Ren Shuzhi leg. (NKU); 1♀, Guangxi, Pingxiang, Daqingshan (NKU); 1♂, 1♀, Guangxi, Pingxiang, 1964-VII-26, Wang Liangchen leg.(NKU); 1♂, Yunan, Pingbian, Lincang, 800-1300 m, 1956-VI-27, Huang Keren leg. (NKU); 1♀, Guangxi, Ningming, Longrui, 1984-V-18, Ren Shuzhi leg. (NKU).
Distribution. CHINA (Guangxi <Longsheng, Ningming, Pingxiang, Napo, Fangcheng Fulong, Longzhou Nonggang, Longrui>, Yunnan <Pingbian>); MYANMAR, CAMBODIA, INDIA, VIETNAM.
- (6) Sycanus hsiaoi Li & Cai sp. nov.
- (Figures 2e, 3e, 4e, 15, 16)
Diagnosis. The new species resemble to Sycanus croceus in the body shape and the body coloration. However in the new species, the coxae of leg is yellow, and its basal part is black; the sterna of meso- and meta-thorax are yellow, the markings on lateral side of sternum of meso-thorax and posterior margin of sternum of meta-thorax are black; the black marking of connexivum is smaller, and only laterally extending to half of connexivun or nearly reaching to lateral margin; the posterior pronotal lobe and the corium of fore wing are yellowish white (vs. In Sycanus croceus, the coxae is yellow without black markings; the sterna of meso- and meta-thorax are black, the anterior and posterior margins of sterna of meso- and meta-thorax are yellow; the black marking of connexivum is larger, almost laterally extending to the lateral margin of connexivun; the posterior pronotal lobe and the corium of fore wing are yellow).
The male external genitalia of the new species is somewhat similar to that of Sycanus stali Dohrn, 1859 among these Chinese Sycanus species, but there are great differences in body coloration between the two species: the new species is yellowish with some black markings (vs. Sycanus stali is black with red stripes). In addition, the male external genitalia in new species, the median process of pygophore is more distinct, apical part of endosoma laterally is without small spines (vs. In Sycanus stali, the median process of pygophore is smaller, apical part of endosoma laterally is armed with 6 pairs of small spines).
Description. Coloration. Body yellowish with black markings (Figure 15). Head (except ventral surface yellowish), anterior pronotal lobe (except anterior angles and lateral sides yellowish), upper margin of pleura of meso- and meta-thorax, scutellum (except posterior margin and apical spine yellowish), round markings of two lateral sides and median longitudinal strip and inter-segmental transversal strips of ventral surface of abdomen, legs (except apical half of coxae), antennae, black; posterior pronotal lobe (except post-lateral margin yellowish), corium, clavus, epimeron of propleuron, sternum of thorax (except lateral margin of meso-sternum and anterior margin of meta-sternum black), sternum of abdomen (except black strips), milk-white to yellowish white; ventral surface of head, coxae (except basal part black), anterior angles, lateral margin of anterior lobe and sternum of pro-thorax, episternum of propleuron, pleura of meso- and meta-thorax (except upper margin black), posterior margin and apical spine of scutellum, connexivum (except apical part of post-lateral angle black) yellowish.
Structure. Body median-sized. Body clothed with white setae; legs clothed with longer vertical setae. Head long, anti-ocular area shorter than post-ocular; ocelli separated, distance between ocelli wider than that between ipsolateral ocellus and eyes. Anterior pronotal lobe hemispheric, basal median longitudinal sulcus short; posterior pronotal lobe rough with irregular wrinkles; lateral pronotal angle round; posterior margin of pronotum straight; spine of scutellum situated in middle part, thick and long, produced upwards then backward, apex bi-forked. Hemelytron extending beyond tip of abdomen. Abdomen laterally widely roundly dilated, especially fourth to sixth segmental connexiva, posterior-lateral angle of each connexival segment indistinct, round (Figure 15). Pygophore elliptic, median pygophore process shown in Figure 16a, b; paramere clavate, apical part with thick long setae, middle part bent (Figure 16a-c); strut triangular (Figure 16d); endosoma apically armed with many small processes (Figure 16e-g), but laterally without small spines, subapical part with two pieces of narrow and long sclerites.
Measurement [♂(n=2)/♀(n=2), in mm]. Body length 17.27/19.87, maximal width of abdomen 5.82/5.38; Head length 4.55/4.65; length of ante-ocular part 1.64/1.47 ; length of post-ocular part 2.00/2.03; length of synthlipsis 0.73/0.61; distance between ocelli 0.27/0.28; length of antennal segments I–IV =5.09/5.34, 2.18/2.10, 2.18/2.37, 5.45/5.55; length of visible rostral segments I–III =2.00/2.56, 2.73/3.13, 0.73/0.68; length of anterior pronotal lobe 1.09/1.14; length of posterior pronotal lobe 1.64/2.33; maximal width of pronotum 3.27/4.43; length of scutellum 1.27/1.61; length of hemelytron 11.45/14.22.
Holotype, ♀, CHINA, Yunnan, Lancang, Qianliuyizu town, Tianba village, 1375m, 2017-VII-20, Zhou Zhen& Leo Xiaolong leg. (CAU). Paratypes, 1♂, 1♀, CHINA, Yunnan, Lancang, Gengma, 600m, 2019-VII (CAU); 1♂, CHINA, Yunnan, Puer, 2022-VII-4, Zhang Guirong leg. (NNU).
Etymology: The species name is named after the late Professor Hsiao Tsai-Yu in Nankai University, the famous Chinese Entomologist, for his great contribution to the taxonomy of Heteroptera insects.
Distribution. China (Yunnan<Lancang, Puer>).
- (8) Sycanus fuscirostris Dohrn, 1859
- Sycanus fuscirostris Dohrn, 1859: 99[27]. China.
Note. We didn’t examine type species, and didn’t find the Chinese specimens of this species.
- (9) Sycanus hainanensis Wang & Cai sp. nov.
- (Figures 17, 18, 2f, 3f, 4f)
Diagnosis: The body coloration and the structure of abdominal connexivum in Sycanus hainanensis sp. nov. are somewhat different with Sycanus sichuanensis. In the new species, the body shape is thinner, the posterior margin of corium is yellowish brown (Figure 13) (vs. In Sycanus sichuanensis, the body shape is wider, especially the connexivum of abdomen is dilated laterally, most of corium is milk-white to yellowish-white). Sycanus hainanensis Zhao & Cai sp. nov. is so much similar to Sycanus sichuanensis Hsiao, 1979 in external morphological characteristics and male external genital structure, so that it is difficultly to distinguish each other. In COI DNA-barcodes molecular analyses, the result shows that the genetic distance between the two species supports two independent species (Table S3, 9.64) .
Description. Coloration. Body black. Posterior margin of corium of hemelytron yellowish brown, membrane pale brown and semitransparent; connexivum black, transversal markings of each segment of red (Figure 17).
Structure. Body median-sized. Head, thorax, ventral surface of abdomen, corium of hemelytron clothed with white procumbent short setae; legs with pale vertical setae of different length. Anti-ocular part distinctly shorter than post-ocular part; rostrum in-curved, long and slender. Collar indistinct, tuber-shaped; anterior pronotal lobe small, hemispherical and bulged; posterior pronotal lobe rugose, lateral angle obtuse and round, posterior margin nearly straight; scutellum sub-apically with a vertical long spine, apex of spine bi-forked. Femora nearly of equal thickness, apical part somewhat thickened; hemelytron extending beyond tip of abdomen. Fourth and fifth connexival segments of abdomen laterally distinctly expended (Figure 17). Pygophore elliptic, median pygophore process “T”-shaped (Figures 18a, b); paramere clavate, sub-basal part somewhat bent, apical part with short setae (Figure 18a–c). Struts triangular (Figure 18d); dorsal phallothecal sclerite sclerotized, tongue-shaped; apical part of endosoma with a horned process and laterally with about 5-6 pairs of small spines (Figure 18e–g).
Measurement [♂(n=3)/♀ (n=7), in mm] Body length 16.38–16.82/18.62–18.91; maximal width of abdomen 4.61–4.94/5.22–6.38; head length 4.25–4.30/4.63–4.79; length of ante–ocular part 1.55–1.56/1.51–1.71; length of post–ocular part 1.99–2.01/2.22–2.24; distance between ocelli 0.31–0.38/0.31–0.39; length of synthlipsis 0.56–0.57/0.52–0.54; length of antennal segments I–IV= 5.19–5.24/5.18–5.40, 1.80–2.05/1.90–2.09, 1.07–1.18/1.16/1.37, 6.05–6.87/6.00–6.02; length of visible rostral segments I–III= 1.81–1.86/1.94–2.29, 2.76–3.24/3.31–3.57, 0.36–0.41/0.42–0.52; length of anterior pronotal lobe 0.76–0.86/1.09; length of posterior pronotal lobe 1.54–1.71/1.89–1.97; maximal width of pronotum 3.23–3.29/3.83–3.89; length of scutellum 1.13–1.24/1.34; length of hemelytron 10.39–10.59/11.85–12.07.
Type species. Holotype, ♀, CHINA, Hainan, Ledong, Jianfengling, Yulin Valley, 680 m, 2022-V-11, Zheng Yuchen leg. (CAU). Paratypes, 3♂, 3♀ (CAU), 1♂, 1♀ (CATAS), CHINA, Hainan, Jianfengling, 2022.5.13, Wang Jianyun & Liu Yinyi leg.; 2♀, CHINA, Hainan, Diaoluoshan, Mengshuichang, 1981-IX-5 (CAU); 1♀, CHINA, Hainan, Wanning, Shimei, 1981-VI-10 (CAU).
Etymology: The specific name hainanensis is named after the type locality.
Distribution. CHINA (Hainan).
- (10) Sycanus insularis Hsiao, 1979
- (Figures 2g, 3g, 4g, 19, 20)
Redescription. Coloration. Body black, with yellowish and orange markings. Head (except ventral surface yellow and apical part of rostrum pale brown), antennae (except median two annular markings of first segment brown), legs (except coxae orange), anterior pronotal lobe, episterna of pleura of meso- and meta-thorax, sterna of thorax, segmental strip of abdomen and its expending round markings of connexivum, basal half and apical angle of corium of hemelytron black; pronotum (except anterior lobe), propleuron, anterior-lateral margin of corium, apical half of corium (except apical angle) yellowish; coxa, abdomen (except black inter-segmental stripes), epimera of pleuron of meso- and meta-thorax orange (Figure 19).
Structure. Body median to large-sized. Body clothed with white short setae; legs, pronotum, scutellum with longer vertical setae. Head longer, post-ocular area of head longer than anti-ocular area; antennae slender and thin, first segment longest and sub-equal to fourth, third shortest and a little shorter than second; rostrum long, bent, first extending to middle part of eyes, second segment longest. Anterior angle of pronotum round and indistinct; anterior pronotal lobe small, hemisphere, posterior part with short longitudinal sulcus; posterior pronotal lobe rough, lateral angle round; posterior margin of scutellum round, apical spine sub-vertical, apex bi-forked; legs slender; hemelytron extending beyond apex of abdomen; fourth to five connexival segments dilated laterally (Figure 19). Pygophore elliptic, median pygophore process shown in Figure 20a, b; paramere clavate, apical half swelled with thick setae, middle part somewhat bent (Figure 20c). Struts triangular (Figure 20d); phallosoma elliptic; dorsal phallothecal sclerite sclerotized, tongue-shaped; endosoma apically with a horned process and laterally with about 13 pairs of small spines (Figure 20e–g).
Measurement [♂ (n=4) / ♀ (n=3), in mm]. Body length 19.55 / 23.71–24.85, maximal width of abdomen 7.70 / 10.26–10.83; Head length 4.85 / 5.13–5.42; length of ante-ocular part 1.71/ 1.82–2.00; length of post-ocular part 2.28 / 2.28–2.85; length of synthlipsis 0.68 / 0.80–0.86; distance between ocelli 0.46 / 0.46–0.57; length of antennal segments I–IV = 6.16 / 6.84–7.13, 2.28 / 2.57–2.68, 1.71 / 1.82, 6.10 / 8.55–6.27; length of visible rostral segments I–III = 2.28 / 2.74–2.85, 3.14 / 3.82–3.93, 0.68 / 0.68–0.80; length of anterior pronotal lobe 1.14 / 1.20–1.31; length of posterior pronotal lobe 2.17 / 2.57–3.02; maximal width of pronotum 3.88 / 4.56–5.13; length of scutellum 1.37 / 1.43–1.82; length of hemelytron 12.08 / 15.39–16.82.
Type material. Holotype, ♂, CHINA, Hainan, Jianfengling, Tianchi, 1964-V-10, Liu Shengli leg. Allotype, ♀, same as Holotype, 1964-V-9. Paratype, 3♀, same as Holotpye (kept in NKU, NOT in TJNHM).
Specimens examined. CHINA, Hainan: 1♀, Hainan, Jianfengling Nature Reserve, 1980-IV-10, 900m, Xiong Jiang leg.; 1♂, 2♀, 2023-IV-21, Zhao Ping leg. (CAU); 1♀, Hainan, Jianfengling Nature Reserve, 1982-IV-10, Liu Yuanfu leg. (CAU); 1♂, 2♀, Hainan, Ledong, Jianfengling, Tianchi, 2015-V-07, Cai Nanyi leg. (CAU); 1♀, Hainan, Ledong County, Xingfengling, Fengming Valley, 2015-IV-2, Lu Qiu leg. (CAU); 1♂, Hainan, Ledong, Main peak of Jianfengling, 1412m, 2019-IV-14–16, Song Haitian leg. (CAU); 1♀, Hainan, Ledong, Jianfengling, 2013-IV-12, Sun & Zhang leg. (CAU); 1♀, Hainan, Jianfeng mountain, 1982-Ⅵ-10, Liu Yuanfu leg. (CAU); 1♂, 1♀, Hainan, Jianfengling (CAU); 1♀, Hainan, Lingshui County, Diaoluoshan, 1981-V-6 (CAU); 1♀, Hainan, Lingshui County, Diaoluoshan, 2009-IV-9–12, Hou Xiaohui leg., by light trap (CAU); 1♂, 1♀, Hainan, Wuzhishan Nature Reserve, 2023-IV-18, , Liu Xingyue leg. (CAU).
Distribution. CHINA (Hainan <Jianfengling, Diaoluoshan, Wuzhishan>).
- (10) Sycanus minor Hsiao, 1979
- (Figures 2h, 3h, 4h, 21, 22, S8e)
Redescription. Coloration. Body color changeable greatly, generally dark brown to black, with red markings, sometimes almost totally black (Figure 21). Head (except apical part of rostrum and markings on outside of ocelli yellowish brown), antennae, pronotum (except lateral and posterior margins of posterior lobe orange, or collar, lateral margin and posterior lobe altogether orange), thorax (except coxal cavity orange, or most of pro-pleuron, markings of meso- and meta-pleura, anterior margin of meso- and meta- sterna orange), scutellum (except apical spine orange, posterior margin greyish brown), legs (except coxae orange, sometimes a sub-apical annulation of femur and tibiae paler), corium of hemelytron (except sub-apical part yellowish or milk white) black; membrane of hemelytron semitransparent, black; abdomen red to yellowish, median longitudinal stripe, transversal strip of sternum and markings of each connexival segment black (Figure 21). Sometimes body almost totally black, except posterior margin or transversal tripe of corium of hemelytron yellowish, sub-apical annulations of tibiae and femora milk-white, and third and fourth antennal segments and subapical annulation of first segment milk-white to pale brown.
Structure. Median-sized. Body clothed with white setae and depressed curved short setae, legs with straight setae of different length. Head longer, post-ocular area of head longer than anti-ocular area; antennae slender and thin, first segment longest and sub-equal to fourth, third shortest and distinctly shorter than second; rostrum long, bent, first extending to middle part of eyes, second segment longest. Anterior angle of pronotum round and indistinct; anterior pronotal lobe small, hemispheric, posterior part with short longitudinal sulcus; posterior pronotal lobe rough, lateral angle round; posterior margin of scutellum round, apical spine sub-vertical and produced posteriorly, apex bi-forked; legs slender; hemelytron extending beyond apex of abdomen; fourth to sixth connexival segments dilated laterally (Figure 21). Pygophore elliptic, median pygophore process shown in Figure 22a, b; paramere clavate, apical part with setae, middle part somewhat bent (Figure 22c). Struts triangular (Figure 22d); phallosoma elliptic, dorsal phallothecal sclerite sclerotized, endosoma apically with a horned process and laterally with about 5 pairs of small spines (Figure 22e–g).
Measurement [♂ (n=3) / ♀ (n=1), in mm]. Body length 17.20–18.18 /19.27, maximal width of abdomen 8.00–8.80 / 8.18; Head length 4.46–4.73/4.73; length of ante-ocular part 1.47–1.64/1.64; length of post-ocular part 2.15–2.18/2.18; length of synthlipsis 0.43–0.56/0.58; distance between ocelli 0.36–0.55/0.36; length of antennal segments I–IV = 5.20–6.00/5.45, 2.00–2.36/2.00, 0.91–1.10/1.09, 6.00-6.18/5.55; length of visible rostral segments I–III = 1.92–2.18/2.36, 3.15–3.64/3.64, 0.55–0.58/0.55; length of anterior pronotal lobe 0.84–1.09/1.27; length of posterior pronotal lobe1.71–2.00/2.18; maximal width of pronotum 3.31–3.73/4.36; length of scutellum 1.45/1.64; length of hemelytron 9.87–11.82/12.36.
Type material. Holotype, ♂, Yunnan, Xishuangbanna, Banna Menglong, Manbing, 1958-IV-14, Cheng Hanhua leg., 650m, Hsiao Caiyu Det., 1972, kept in Tianjin Natural Museum (TJNHM).
Specimens examined. CHINA, Guangxi: 2♂, Guangxi, Ningming, Huashan, 26-VIII-2019, Zhao Ping leg., (NNU);
CHINA, Yunnan: 1♂, Yunnan, Xishuangbanna, Manbing, 1958-IV-14, Zheng leg., Ren Shuzhi Det., 1972, kept in Nankai University (NKU); 1♂, 1♀, Yunnan, Mengla, Menglu, Cui Jianxin leg. (CAU); 1♂, 1♀, Yunnan, Banna, Jinghong, N21.59955, E100.47813, 530m, 2009-V-3, Cao Liangming leg. (CAU); 5♂, Yunnan, Banna, Jinghong, Jinuo, Jinuo, 2021-VII-22, 1099m, Chen Zhaoyang & Liu Qinpen leg. (CAU); 1♀, Yunnan, Xishuangbanna, Menglun botanical garden, Wild Elephant Valley (CAU); 1♂, 1♀, Yunnan, Banna, Jinghong, Jinuo mountain, National Highway 213,1053 m, 2021-VII-25 (CAU).
Distribution. CHINA (Yunnan <Xishuangbanna>, Guangxi <Ningming>); VIETNAM.
Remark. We didn’t check the type specimen, but we found a male specimens kept in Nankai University with the same information as the type specimen and identified by Pro. Ren Shuzhi.
In addition, the body color of Sycanus minor is changeable from orange or red with black markings to almost completely black. The three specimens from Mengla and Jinghong, Xishuangbanna, Yunnan, China, the body color is black (except the posterior margin of corium of fore wing is yellow to milk-white, and the annular markings of sub-apical part of femur is milk-white), and through the DNA barcoding molecular analysis, the result showed that the three specimens should belong to the species Sycanus minor.
- (12) Sycanus rufus Hsiao 1979
- (Figures 2i, 3i, 4i, 23, 24)
Redescription. Coloration. Body red, with black markings. Head (except apical part of rostrum and markings around ocelli yellowish brown), antennae, sterna and pleuron of meso- and meta-thorax (except coxal cavity orange), scutellum (except apical spine orange, posterior margin greyish brown), markings of anterior pronotal lobe, legs (except coxae orange, sometimes a sub-apical annulaion of femur and tibiae paler), basal part of corium of hemelytron blackish brown; membrane of hemelytron semitransparent, blackish brown; abdomen red to yellowish red, median longitudinal stripe, basal transversal strip and lateral markings of sternum, and basal round large markings of each connexival segment formed by which basal transversal strip of 3rd o 7th sterna laterally extended (round markings of sixth and seventh not extending to margin of connexivum) black; ventral surface of head, pronotum (except markings of anterior lobe black) and pro-pleuron (except posterior margin black) , corium of hemelytron (except basal half), coxae, coxal cavity red , or orange (Figure 23).
Structure. Body large-sized. Ventral surface of head, pronotum, pleura and sterna of thorax, corium of hemelytron clothed with yellow procumbent short setae; head (except ventral surface), pronotum and propleuron, scutellum, legs, ventral surface of abdomen with pale setae of different length. Anticular part distinctly shorter than post-ocular part; rostrum in-curved, long and slender. Collar indistinct and round; anterior pronotal lobe small, hemisphered and bulged, deeply depressed at base; posterior pronotal lobe rugose and reticulated, lateral angle obtuse and round, posterior margin nearly straight, posterior angle nearly absent; scutellum sub-apically with an erect upward, thick short spine, tuber-shaped in male, short cone-shaped in female. Femora nearly of equal thickness, apical part somewhat thickened; hemelytron extending beyond tip of abdomen. Fourth to sixth connexival segments of abdomen moderately expended (Figure 23). Pygophore elliptic, median pygophore process shown in Figure 24a, b; paramere clavate, apical part with setae, middle part somewhat bent (Figure 24c, d). Struts triangular (Figure 24e); phallosoma elliptic, dorsal phallothecal sclerite sclerotized, endosoma apically with a horned process and laterally with about 5 pairs of small spines (Figure 24f–h).
Measurement [♂ (n=4) / ♀ (n=3), in mm]. Body length 20.29–20.63/24.85–25.88, maximal width of abdomen 5.81–5.99/8.44–10.15; Head length 4.67–4.96/5.24–5.47; length of ante-ocular part 1.71/1.94–2.05; length of post-ocular part 2.28/2.45–2.57; length of synthlipsis 0.68–0.73/0.80–0.86; distance between ocelli 0.46/0.51; length of antennal segments I–IV = 5.89–6.38/6.95–7.01, 2.51–2.85/2.85–3.02, 2.17–2.45/2.37–2.57, 5.53/5.87–6.67; length of visible rostral segments I–III = 2.28/2.85, 3.42/3.99, 0.68–0.71/0.74–0.80; length of anterior pronotal lobe 1.03–1.14/1.14–1.20; length of posterior pronotal lobe 2.11–2.17/2.25–3.08; maximal width of pronotum 3.93–3.99/5.47–5.76; length of scutellum 1.14–1.25/1.20–1.25; length of hemelytron 12.83–12.85/16.82–17.67.
Type material. Holotype, ♂, CHINA, Yunnan, Xishuangbanna, Menga, 1050–1080 m, 9-VI-1958, Fu Fuji leg. Allotype, ♀, same as Holotype, 6-VI-1958. Paratype, 2♂, 1♀, CHINA, Yunnan, Lancang, Xishuangbanna, kept in IOZ.
Specimens examined. CHINA, Yunnan: 1♂, Yunnan, Mengla, V-29, 650m (CAU); 1♀, Yunnan, Xishuangbanna, Mengla, Yaoqu, 2006-VI-5, Wang Hesheng leg. (CAU); 1♀, Yunnan, Banna, Jinghong, Jinuo, Yunfenggusi, 1070m, 2021-VII-20, Chen Zhaoyang & Liu Qinpeng leg. (CAU); 3♂, 1♀, Yunnan, Xishuangbanna, Mengla, Bubeng, 2012-VII-10, Zhao & Chen leg. (CAU); 1♀, Yunnan, Banna, Jinghong, Jinuo, 1100m, 2018-V-31 (CAU); 1♀, Yunnan, Pu’er, Simao, 1436m, 2018-VI-20 (CAU); 1♀, Yunnan, Xishuangbanna, Jinghong Guanping, 2008-V-27, Huang Xinyong leg. (CAU).
Distribution. CHINA (Yunnan <Lancang, Pu’er, Xishuangbanna: Mengla, Meng’a, Xiaomengyang, Mengzha>).
- (13) Sycanus sichuanensis Hsiao 1979
- (Figures 2j, 3j, 4j, 25, 26, S8f)
- Sycanus szechuanus (sic): Hsiao & Ren 1981: 520[31].
Redescription. Coloration. Body dark brown to black, shiny. Corium of hemelytron (except basal part and apical angle), basal of membrane, yellowish; transversal markings of 3rd to 7th connexival segment red; membrane semitransparent, brown; sub-apical par of femur paler brown o white; marking around ocelli; apical part of rostrum brown (Figure 25) .
Structure. Body medain to large-sized. Head, thorax, corium of hemelytron clothed with yellow procumbent short setae and vertical short setae of different length; legs with vertical setae of different length. Ante-ocular part distinctly shorter than post-ocular; rostrum in-curved, long and slender. Collar indistinct and round; anterior pronotal lobe small, hemisphered and bulged, deeply depressed at base; posterior pronotal lobe rugose and reticulated, lateral angle obtuse and round, posterior margin nearly straight, posterior angle nearly absent; scutellum sub-apically with an long spine, apical spine produced posteriorly, apex bi-forked. Femora nearly of equal thickness, apical part somewhat thickened; hemelytron extending beyond tip of abdomen. Abdomen distinctly roundly laterally expended (Figure 25). Pygophore elliptic, median pygophore process “T”-shaped shown in Figure 26a, b; paramere clavate, apical half somewhat swelled with thick setae, middle part somewhat bent (Figure 26c). Struts triangular (Figure 26d); phallosoma elliptic, dorsal phallothecal sclerite sclerotized, endosoma apically with a horned process and laterally with about 5 pairs of small spines (Figure 26e–g).
Measurement [♂ (n=8)/♀ (n=5), in mm] body length 16.55–18.73/16.73–22.58; maximal width of abdomen 6.36–7.27/6.36–11.16. head length 4.18–4.73/4.18–5.25; length of ante–ocular part 1.45–1.55/1.45–1.84; length of post–ocular part 1.82–2.18/2.00–2.68; distance between ocelli 0.25–0.55/0.29–0.47; length of synthlipsis 0.55–0.73/0.55–2.26; length of antennal segments I–IV= 5.45–6.00/5.82–6.62, /2.18–2.94, 2.27–2.36/1.64–2.26, /6.67–6.91; length of visible rostral segments I–III=1.64–1.82/1.64–2.63, 3.27–3.45/2.73–3.89, 0.64/0.55–0.74; length of anterior pronotal lobe 1.00–1.09/0.91–1.21; length of posterior pronotal lobe 1.82/1.82–2.57; maximal width of pronotum 3.09–4.00/3.36–4.41; length of hemelytron 10.18–11.45/10.36–14.70.
Type material. Holotype, ♂, CHINA, Sichuan, Ermeishan, 1955-VI-28, Huangjin leg. Allotype, ♀, same as Holotype, 1955-VI-23. Paratypes: 3♀, 1♂, CHINA, Sichuan, Ermeishan (IOZ); 1♀, 1♂, CHINA, Sichuan, Ermeishan, 1955-VI-27, 580m, Xie Dabin leg.(IOZ); 1♀, 3♂, CHINA, Sichuan, Er’meishan, Huang Keren & Jin Yintao leg. (IOZ).
Specimens examined. CHINA, Guangxi: 1♂, Guangxi, Longzhou, Longrui, 1984-V-20, Ren Shuzhi leg. (NKU); 1♂, Guangxi, Tianpingshan, 1963-VI-17 (NKU); 1♀, Guangxi, Nonggang, Longzhou, 2003-VIII-15, Zhou Zhihong leg. (CAU); 1♀, Guangxi, Huaping, Tianping mountain, 1963-VI-5, Yang Jikun leg. (NKU); 1♂, 1♀, Guangxi, Jinxiu, 1983-V-27 (NKU); 1♀, Guangxi, Lonhzhou, Nonggang, 1983-V-15 (NKU); 1♀, Guangxi, Pingxiang, Orchid Valley Park, 2014-X-8, Sun & Luo leg. (CAU); 1♀, Guangxi, Guilin, Huaping, Tianping mountain, 1963-VI-5, Yang Jikun leg. (NKU); 1♀, Guangxi, Longzhou, Nonggang, 2003-VIII-15, Zhou Zhihong leg. (CAU); 1♂, 1♀, Guangxi, Jinxiu, 1983-V-27 (NKU); 1♀, Guangxi, Longzhou, Nonggang, 1983-V-15. (NKU).
CHINA, Sichuan: 1♀, Sichuan, Ermeishan, 600m-1200m, 2019-VI-27 (CAU); 1♀, Sichuan, Ermeishan, 580 m, 19-VI, Huang Keren leg. (NKU);1 ♀, Sichuan, Chengdu, Pengzhou, Bailu Town, 800m, 2018-VII-21, Zhou Chao leg. (CAU); 1♀, Sichuan, Leshan, Emeishan, 2014-VII -3, Liang Si leg. (CAU).
CHINA, Guizhou: 1♀, Guizhou, Rongjiang, Pingyang, 693m, 2016-VI-20-24, Wu Shengsheng leg. (CAU); 1♀ Guizhou, Rongjiang, Pingyang, Xiaodanjiang, 2005-VI-3, 920-970m, Zhao Ping leg. (NNU).
CHINA, Yunnan: 1♀, Yunnan, Honghe, Hekou, Huayu cave, 334m, 2016- IV-23, Yang Xiaodong leg. (CAU); 1♂, Yunnan, Honghekou, Huayu cave, 200m, 2016- IV-24, Yu Tang Wang leg. (CAU).
CHINA, Hunan: 1♀, Hunan, Yongzhou, Ningyuan, Jiuyi mountain, 915m, 2021-VI-4–7, Peng Huoliang leg. (CAU);
Distribution. CHINA (Hunan <Xiangxi, Xiangnan, Yonhzhou>, Hubei, Sichuan <Chengdu, Ermeishan, Ya’nan, Xingjing>, Guizhou<Maolan, Rongjiang, Shiqian>, Guangxi<Longsheng, Longzhou, Guilin, Pingxiang, Jinxiu, Huaping>, Yunan <Gengma, Honghe>); VIETNAM.
- (14) Sycanus stali Dohrn, 1859
- (Figures 2k, 3k, 4k, 27, 28, S7)
- Sycanus stali Dohrn, 1859: 96[27].
- Sycanus generosus Stål, 1863: 58[28].
- Sycanus marginatus Hsiao, 1979: 143, 154[30].
- Sycanus marginellus Putshkov P.V.: Putshkov, V.G., Putshkov, P.V. & Štys, 1987: 104[43], new name for S. marginatus Hsiao 1979.
- Sycanus hsiaoi Maldonado-Capriles 1990: 312[24], new name for S. marginatus Hsiao 1979. new synonym.
Redescription. Coloration. Body black. Apical half of corium of hemelytron (apical angle) yellowish; margin of connexivum red; apical part of rostrum brown (Figure 27).
Structure. Body large-sized. Body clothed with black short setae; legs, pronotum, scutellum with longer vertical setae. Head longer, post-ocular area of head longer than anti-ocular area; antennae slender and thin, fourth segment longest and sub-equal to first, third shortest and a little shorter than second; rostrum long, bent, first extending to middle part of eyes, second segment longest. Anterior angle of pronotum round; anterior pronotal lobe small, hemisphere, posterior part with short longitudinal sulcus; posterior pronotal lobe rough, lateral angle round; posterior margin of scutellum round, apical spine with tubercular processes; legs slender; hemelytron extending beyond apex of abdomen; abdomen roundly dilated laterally, fourth to five connexival segments dilated laterally (Figure 27). Pygophore elliptic, median pygophore process shown in Figure 28a, b; paramere clavate, apical half swelled with thick setae, middle part distinctly bent (Figure 28c). Struts triangular (Figure 28d); phallosoma elliptic, dorsal phallothecal sclerite sclerotized, endosoma apically with a horned process and laterally with about 6 pairs of small spines (Figure 28e–g).
Measurement [♂ (n=4) / ♀ (n=3), in mm]. Body length 21.27–25.08/26.11–27.27, maximal width of abdomen 8.18–8.55/9.92–10.91; Head length 4.73–5.53/5.70–5.82; length of ante–ocular part 1.63–1.88/2.00; length of post–ocular part 2.18–2.74/2.73–2.74; length of synthlipsis 0.64/0.91; distance between ocelli 0.27/0.33–0.51; length of antennal segments I–IV= 5.82/6.91–6.95, 2.36/2.96–3.09, 2.85/2.28, 6.55/9.09; length of visible rostral segments I–III= 2.18–2.85/2.91–2.96, 3.27–4.39/4.10–4.18, 0.64–0.74/0.80–0.91; length of anterior pronotal lobe 1.09–1.31/1.27–1.43; length of posterior pronotal lobe 2.00–2.85/3.02–3.09; maximal width of pronotum 3.64–4.85/5.42–5.45; length of scutellum 1.43/1.71; length of hemelytron 13.09–16.53/ 17.67–18.18.
Specimens examined. 1♂, 1♀, Yunnan, Menghai, 20-V-1991, 600m, Cai Wanzhi leg. (CAU); 30♂, 30♀, China, Yunnan, Puer, Simao district, Nanping, Baizhi tamper, Wanmu tea Garden, 2022-VI-15, Zhang Guirong leg. (NNU).
Distribution. CHINA (Yunnan<Lancang, Xishuangbanna: Menga, Menghai, Mengzha, Puer>); PHILIPPINES.
Notes. Although we haven’t examined the type of Sycanus stali Dohrn, 1859[27], we found the syntype of Sycanus generosus Stål, 1863 [28] kept in Swedish Museum of Natural History on the website: http://www2.nrm.se/en/het_nrm/g/sycanus_generosus.html (Figure S7a,b). Stål in 1874 regarded Sycanus generosus Stål, 1863 and Sycanus generosus Stål, 1863 as two variants of Sycanus stali Dohrn, 1859[41]. The type photograph of Sycanus marginatus Hsiao, 1979 (Figure S7c–e) is morphologically consistent with the type photograph of Sycanus generosus Stål, 1863 (Figure S7a,b). Therefore, we use Sycanus stali Dohrn, 1859 as the specific name.
- (15) Sycanus taiwanensis Zhao & Cai sp. nov.
- (Figures 2l, 3l, 4l, 29, 30)
Diagnosis. The male external genitalia of the new species is the most similar to that of Sycanus sichuanensis, but there are differences in external body color and structure. In the new species, the body is slightly thin and long, the connexivum is laterally dilated, the posterior pronotal lobe is red (except the middle part is black) and posterior margin of corium of fore wing is milk-white (vs. In Sycanus sichuanensis, the body is posteriorly widen, and the connexivum is wide and round, and the middle part of each segment of connexivum is elevated, the pronotum is totally black and the most of corium of fore wing is white) (Figure 29).
Description. Coloration. Body black, with red markings. Head (except rostrum and markings around ocelli yellowish brown), antennae, thorax (except lateral and posterior margins of posterior lobe red or totally red), scutellum, legs, corium of hemelytron (except posterior margin yellowish), abdomen (except connexivum) black; membrane of hemelytron semitransparent, brown; connexivum red, markings of 4th to 7th segmental suture black (Figure 29).
Structure. Body large-sized. Head, thorax, ventral surface of abdomen, corium of hemelytron clothed with white procumbent short setae; legs with pale vertical setae of different length. Ante-ocular part distinctly shorter than post-oculart; rostrum in-curved, long and slender; antennae slender and thin, first segment longest and somewhat longer than fourth, third shortest and a little shorter than second. Collar indistinct; anterior pronotal lobe small, hemisphered and bulged, deeply depressed at base; posterior pronotal lobe rugose and reticulated, lateral angle obtuse and round, posterior margin nearly straight; scutellum sub-apically with an oblique spine, spine indistinctly bi-forked. Femora nearly of equal thickness, apical part somewhat thickened; hemelytron extending beyond tip of abdomen. Fourth and fifth connexival segments of abdomen moderately expended (Figure 29). Pygophore elliptic, median pygophore process shown in Figure 30a, b; paramere clavate, apical half somewhat swelled with thick setae, middle part somewhat bent (Figure 39c). Struts triangular (Figure 30d); phallosoma elliptic; dorsal phallothecal sclerite sclerotized; endosoma apically with a horned process and laterally with about 5 pairs of small spines (Figure 30e-g), and middle part dorsally with two short and slender sclerites.
Measurement [♂ (n=3) / ♀ (n=3), in mm]. Body length 18.73–20.41/22.8–20.00, maximal width of abdomen 5.24–6.73/6.56–6.91; Head length 4.55–5.13/5.64–6.91; length of ante–ocular part 1.73–1.77/1.88–2.00; length of post–ocular part 2.00–2.57/2.36–2.57; length of synthlipsis 0.69–0.80/0.73–0.80; distance between ocelli 0.36–0.57/0.51–0.57; length of antennal segments I–IV = 5.45–6.56/6.00–6.44, 2.36–2.62/2.62–2.73, 1.54/1.68, 5.00/5.80; length of visible rostral segments I–III = 2.00–2.45/2.18–2.34, 3.45–3.99/3.64–3.93, 0.55–0.74/0.55–0.86; length of anterior pronotal lobe 1.09–1.25/1.09–1.37; length of posterior pronotal lobe 1.82–2.22/2.39–2.45; maximal width of pronotum 3.82–3.99/4.36–4.39; length of scutellum 0.86/1.25; length of hemelytron 11.82–12.56 /12.73–13.97.
Holotype, ♀, CHINA, Taiwan, Pingtung County, Lilong Mountain, 650 m, 2015-I-26 (CAU). Paratypes. 1♂, CHINA, Taiwan, Pingtung County, Lilong mountain, 650 m, 2015-I-26 (CAU); 1♂, CHINA, Taiwan, Pingtung County, Manchu Township, Jialeshui, 2019-XI-08, Liu Xingyue leg. (CAU); 1♀, CHINA, Taiwan, Pingtung County, Manchu Township, Jialeshui, 2010-V-29, Zhou Wenyi leg. (CAU); 1♀, CHINA, Taiwan, Pingtung County, Heng-Chun, Ken-Ting, 400 m, 2016-VIII-23, Wu S.P. & Chung Y.T. leg. (CAU); 1♀, CHINA, Taiwan, Pingtung County, Heng-Chun, Ken-Ting, 2010-X-7, Cai Wanzhi leg. (CAU); 1♂, CHINA, Taiwan, Pingtung County, Fenggang, Lilong Moutain, 2015-I-26, 650m, Zhong Yiting leg. (CAU); 2♂, 4♀, CHINA, Taiwan, Pingtung County, Lilong mountain, 650m, 2015-Ⅷ-17, Y.-T. Chung leg. (CAU); 1♂, 2♀, CHINA, Taiwan, Pingtung County, Lilong mountain, 651m, 2015-Ⅷ-18, Y.-T.Chung leg. (CAU); 1♀, CHINA, Taiwan, Pingtung County, Manchu Township, Gangkou, 2010-VI-13, Zhou Wenyi leg. (CAU); 1♀, CHINA, Taiwan, Pingtung County, Neipu Township, 2001-Ⅵ-23, Wu Shuping leg. (CAU); 1♂, CHINA, Taiwan, Kaohsiung City, Taoyuan, Zhou W.Y. leg. (CAU); 1♀, CHINA, Taiwan, Kaohsiung City, Taoyuan District, Meilong Mountain, 1370m, 2016-Ⅵ-15, Po-Hsin Kuo leg. (CAU); 1♀, CHINA, Taiwan, Kaohsiung, Taoyuan, Xiaoguanshan forest road,1523m, 2016-V-15, Po-Hsin Kuo leg. (CAU); 1♀, CHINA, Taiwan, Kaohsiung, Maolin (CZ), 365m, 2012-X-26, (CAU); 1♀, CHINA, Taiwan, Chiayi County, Alishan Township, Danayi Valley, 2012-VI-23, Song Fan leg. (CAU); 1♀, CHINA, Taiwan, Xinbei City, Sanxia District, Beichatian Mountain, 2016-Ⅶ-19, Wu Shuping leg. (CAU); 1♀, CHINA, Nantou country, Endemic Species Research Institute, 2016-Ⅺ-4, Zhao Yisheng leg. (CAU).
Etymology: The specific name taiwanensis alludes to the locality of the type species in Taiwan province, the Peoples Republic of China.
Distribution. CHINA (Taiwan).
4. Discussion
4.1. Molecular Analysis Based on COI DNA Barcodes
In the present study, we conducted the species delimitation and the phylogenetic analyses based on the 81 COI DNA barcodes dataset of twelve Sycanus species in China. All samples in this study were identified as eleven species. The molecular and morphological analysis results are consistent (except Sycanus bicolor and Sycanus croceovittatus). The integrative taxonomy based on COI DNA barcodes and morphological evidence was suitable and effective for accurate identification of Chinese Sycanus species (Figure 1, S1, S2). The morphological differences between Sycanus bicolor and Sycanus croceovittatus are significant (Figure 7, Figure 8, Figure 9 and Figure 10). However we were also confused why the DNA barcodes of the two species are so similar. We speculated whether there has been a species hybridization event between Sycanus croceovittatus and Sycanus bicolor, then resulting in a very similar genetic material for the two close related species, and even it is difficult to be distinguished by each other. However, in view of the obvious morphological differences between the two species (Figure 7, Figure 8, Figure 9 and Figure 10), we will not do any treatment for the time being and will leave it for further study.
4.2. Discovery of Island Endemic and Cryptic Species
The long-time separation of islands from the mainland has led to the isolation of island biota. In the face of the challenges of island environment, the radiation adaptations swiftly occur, then ultimately speciation events occur. The continental islands, as natural laboratories of evolution, are distributed with many island endemic species that have been cut off from mainland relatives [50]. In China, Taiwan and Hainan provinces are two large continental islands and form the endemic biotas.
In the present study, we discovered two interesting speciation cases of the genus Sycanus in Taiwan and Hainan islands of China by the species delimitation and phylogeny analysis based on COI DNA barcoding. In one case, the molecular analysis results showed that the Sycanus sichuanensis Hsiao, 1979 group consists of four species, the two known continental species Sycanus sichuanensis Hsiao, 1979 and Sycanus minor Hsiao, 1979, as well as Sycanus taiwanensis Zhao & Cai sp. nov. in Taiwan island and Sycanus hainanensis Wang & Cai sp. nov. in Hainan island which are two new island endemic cryptic species (Figure 1a; Table S2, S3).
In the other case, the Sycanus croceus Hsiao, 1979 group includes three mainland species, Sycanus croceus Hsiao, 1979, Sycanus rufus Hsiao, 1979, and Sycanus hsiaoi Li & Cai sp. nov., and one island endemic known species Sycanus insularis Hsiao, 1979. Sycanus insularis is similar to Sycanus croceus in the body shape and body color. Through the species delimitation analysis using COI DNA barcodes dataset, the result showed that the genetic distance is 4.28 between Sycanus insularis and Sycanus croceus. Sycanus insularis has become an independent island endemic species due to long-term island isolation (Figure 1b). Sycanus hsiaoi Li & Cai sp. nov. in Yunnan Province, China is similar to Sycanus croceus Hsiao, 1979 in body shape and body color, but there is a significant difference in genitalia. Through molecular data analysis, the actual genetic relationship between Sycanus hsiaoi sp. nov. and Sycanus croceus is indeed far. Sycanus hsiaoi sp. nov. is similar to Sycanus stali Dohrn, 1859 in the structure of male genitalia, and the molecular phylogenical analysis result supported the closer relationship of Sycanus stali and Sycanus hsiaoi sp. nov. (Figure 1c). The phylogenetic statuses of Sycanus stali in BI, ML and NJ trees are different, so it was not included in the Sycanus croceus species group inhere.
In addition, we found that the island endemic species of Sycanus in China, Sycanus insularis Hsiao, 1979, Sycanus taiwanensis Zhao & Cai sp. nov., and Sycanus hainanensis Wang & Cai sp. nov., are generally at the base of the evolutionary clade of their species group in phylogenetic tree, which indicates that the island endemic species (Taiwan and Hainan) may be genetically closer to their common ancestor than the mainland relatives (Figure 1a, b).
The Taxonomic Status of Sycanus falleni Stål, 1863 is quite confusing. Both morphological and molecular data show that it is far away from other members of the genus Sycanus in China due to the genetic distance between 14.2-19.7 (Figures 1c; Tables 2, 3), and seems to be an independent group. Since our taxonomic study on the genus Sycanus was limited to Chinese species inhere, and didn’t cover all of the world’s species, the further research is needed.
4.3. Polymorphism and Integrative Taxonomy of Molecular and Morphological Evidences
Some of the Sycanus species show the polymorphism of body color and size, so it is hard for the correct identification of these species. In Sycanus falleni, there are two kinds of body color, bright red and dark red (Figure S8a, b); In Sycanus croceovittatus and Sycanus stali, the transverse strip on the corium of fore wing is yellow white or milky white (Figure 27a,d, S8c,d); In Sycanus taiwanensis sp. nov., the posterior pronotal lobe is with a large black markings on the basal half of middle part, or whole red; In Sycanus minor, the body color can become from orange with black markings to completely black (S8e); In Sycanus sichuanensis, the body size and the transverse strip on the corium of fore wing are changable, the body length is from 16.55 to 18.73 mm in male, from 16.73 to 22.58 mm in female, the apical half or the apical margin of corium is white. The smaller individual of Sycanus sichuanensis distributed in Yunnan and southwest of Guangxi provinces is similar to Sycanus minor in body color, size and shape, which make us fail to distinguish the two species during the experimental research process. We also tried to describe their body color variation of the Chinese Sycanus species in the above content, so as to facilitate the correct identification of the genus Sycanus in China.
Prior to the present study, the revision of the genus Sycanus in China faced many problems, due to the polymorphism phenomenon. It was difficult for us to distinguish whether these complicated phenotype changes were inter-species or intra-species differences. By the molecular analysis based on COI DNA barcodes, the morphological taxonomic results have been rectified and corrected objectively. The Integrative taxonomy method based on morphological and molecular evidences is very necessary for the research of modern entomology taxonomy.
4.4. Biology and natural enemies of pests
Assassin bugs in Sycanus considered in this study not only possess amazing appearance, they are the important enemy insects in forest ecosystems of South China [51,52,53]. Sycanus falleni is easy to raise and can be fed with Tenebrio molitor. The nymph stage time of Sycanus falleni (113 days) is shorter than that of Sycanus croceus (156 days) (Tables S4, S5). The predation amount of Sycanus falleni is larger than that of Sycanus croceus, average oviposition number per egg mass is more than 120 and several times that of Sycanus croceus. Sycanus falleni is an important natural enemy insect resource.
5. Conclusions
In the present study, we conducted the species identification, the species delimitation (ASAP, p-distance) and phylogenetic analyses (BI, ML, NJ) based on the COI DNA barcodes of 81 Sycanus samples in China, twelve valid species are redescribed or described, among them two China continental island cryptic species and one Southwest China species were described as new to science: Sycanus taiwanensis Zhao & Cai sp. nov., Sycanus hainanensis Wang & Cai sp. nov. and Sycanus hsiaoi Li & Cai sp. nov. The integrative taxonomy combining with COI DNA barcoding and the morphological evidence shows the advantages of efficiency, accuracy and convenience, make us through the biological phenotypic phenomena to reveal the genetic nature of biological evolution and solve the practical taxonomic problems of the assassin bug Sycanus in China.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: The maximum likelihood (ML) tree of cytochrome c oxidase subunit I (COI) sequences for 81 terminals of Sycanus, Figure S2: The neighbor-joining (NJ) tree of cytochrome c oxidase subunit I (COI) sequences for 81 terminals of Sycanus, Figure S3: The neighbor-joining (NJ) tree of partial 28S rDNA sequences for 34 terminals of Sycanus, Figure S4: Sycanus bicolor Hsiao, 1979, Paratype, habitus, female, Figure S5: Sycanus croceus Hsiao, 1979, habitus, Figure S6: Sycanus falleni Stål, 1863, type species (kept in Swedish Museum of Natural History), female, habitus, Figure S7: (a, b), Sycanus generosus Stål, 1863, type species (kept in Swedish Museum of Natural History), female, habitus; (c-e) Sycanus marginatus Hsiao, 1979, Holotype (kept in Institut of Zoology, Chinese Academy of Sciences, Beijing, China), female, habitus, Figure S8: (a, b) Sycanus falleni Stål, 1863; (c, b) Sycanus croceovittatus (Dohrn, 1859); (e) Sycanus minor Hsiao, 1979; (f) Sycanus sichuanensis Hsiao, 1979, Table S1: GenBank Accession number under every sample individual code (in PDF file of Supporting information), Table S2: Pairwise genetic divergence (Kimura two-parameter) within and between 12 sampling species of the genus Sycanus using cytochrome c oxidase subunit I gene sequences (Table S2 is uploaded separately), Table S3: Pairwise genetic divergence (Kimura two-parameter) between 81 individuals of 12 species of the genus Sycanus using cytochrome c oxidase subunit I gene sequence (Table S3 is uploaded separately), Table S4: Life history of Sycanus croceus Hsiao, 1979 (China, Guangxi, Ningming), Table S5: Life history of Sycanus falleni Stål, 1863 (China, Guangxi, Ningming), Alignment S1: Alignment of COI dataset (Alignment S1 is uploaded separately), Alignment S2: Alignment of partial 28S rDNA sequences dataset (Alignment S2 is uploaded separately).
Author Contributions
Conceptualization, methodology, P.Z., H.L. and W.C.; fieldwork, P.Z. J.W. and Z.C.; labwork on adult morphology, P.Z.; lab work on biology, P.Z.; molecular work, P.Z. and S.C.; writing—original draft, P.Z. and C.S.; supervision, W.C.; project administration and funding acquisition, P.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the National Natural Science Foundation of China (No. 32270474), the Guangxi Natural Science Foundation Program (2021GXNSFAA220106), Central Public-interest Scientific Institution Basal Research Fund of Chinese Academy of Tropical Agricultural Sciences (NO. 1630042020002), the Project of Biological Resources Survey in Wuyishan National Park (No. HXQT2020120701) and Sanya Yazhou Bay Science and Technology City (No. SYND-4222022-04). Key Laboratory Foundation of Environment Change and Resources Use in Beibu Gulf, Education Ministry (Nanning Normal University, NNNU-KLOP-X2002), and Research Foundation of Nanning Normal University (602021239295).
Data Availability Statement
All sequences were deposited in the GenBank under accession numbers of OP927069–OP927154 (part sequences of mtDNA COI) and OP923231–OP923272 (partial 28S rDNA sequences).
Acknowledgments
The authors sincerely thank Prof. Ren Shuzhi, Prof. Bu Wenjun for their helps in specimens eaxmination kept in Nankai University. We are also grateful to Xinwei Niu in China Agricultural University for his help in picture processing.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A

Table A1.
Information on the samples used in the molecular study.
| species | Obtained sequences | Locality, Sampling Time, Elevation | Geographical Coordinates | |||
|---|---|---|---|---|---|---|
| mtDNA COI barcodes (675bp) |
Partial 28S rDNA sequence (639bp) | |||||
| Ingroup | Sample Code | Sample Size | Sample Code | Sample Size | ||
| Sycanus bicolor Hsiao, 1979 | ESX-1-2-COI ESX-1-COI |
2 | / | 0 | Yaoqu, Mengla, Yunnan, China, 2005-V-7 | 21°43′22′′ N 101° 32′41′′ E |
| ES-YN1-3-COI ESYN1-3-COI ESYN1-3-COI |
3 | ESYN13c ESYN1c ESYN2c ESYN4 ESYN9 |
5 | Dengmacaidi, Hehua, Tengchong, Baoshan, Yunnan, China, 2006-Ⅵ-2, 1328 m | 24°58′21′′N 98°23′48′′E |
|
| Sycanus croceovittatus (Dohrn, 1859) | HDX-COI HDX-1-COI HDX-2-COI HDX-3-COI HDX-5-COI HDX-6-COI HDX-8-COI HDX-10-HD2-COI HDX-11-HD3-COI HDX-12-HD4-COI HDX-13-HD5-COI HDX-14-HD6-COI HDX-15-HD7-COI HDX-16-HD8-COI HDX-17-HD-COI |
15 | HDX2 HDX6 HDX8 |
3 | Hudiegu, Huashan, Ningming, Guangxi, China, 2014-VIII-12, 127 m | 22°12′50″N 107°3′21″E |
| HDX-12-COI HDX-13-COI HDX-16-COI |
3 | HDX10 | 1 | Nonggang, Longzhou, Guangxi, China, 2014-VIII-15 | 22°28′24″N 106°57′11″E |
|
| HDX-GD1-COI | 1 | / | 0 | Mashui, Yangchun, Yangjiang, Guangdong, China, 2002-Ⅳ-30, 44 m | 22°7′29′′N 111°41′23′′E |
|
| HDX-GD2-COI | 1 | / | 0 | Huizhou, Guangdong, China, 2004-Ⅷ-19, 22 m | 23°6′44′′N 114°24′56′′E |
|
| HDX-GD3-COI | 1 | / | 0 | Meixian, Guangdong, China, 1981-Ⅸ-6, 79 m | 24°15′58′′N 116°4′53′′E |
|
| HDX-LC1-COI HDX-LC2-COI HDX-LC3-COI HDX-LC4-COI HDX-LC5-COI HDX-LC6-COI |
6 | / | 0 | Wanmu tea Garden, Nanping, Simao, Puer, Yunnan, China, 2022-VI-15 | 22°44′50″N 100°58′32″E |
|
| Sycanus croceus Hsiao, 1979 | HUX-1-COI-F HUX-2-COI-F HUX-3-COI-F HUX-4-COI-F HUX-5-COI-F |
5 | HX2 HX3 HX4 HX5 HX9HD1 |
5 | Hudiegu, Huashan, Ningming, Guangxi, China, 2014-VIII-12, 374 m | 22°12′29″N 107°3′18″E |
| HUX-6-COI-F HUX-7-COI-F |
2 | HX6 HX7 |
2 | Pingyang, Rongjiang, Guizhou, China, 2016-VI-1 | 26°22′59″N 108°5′9″E |
|
| HUX-FJ1-COI HUX-FJ2-COI HUX-FJ3-COI |
3 | HUGX HUGX2 HUGX3 |
3 | Shangyong, Dehua, Quanzhou, Fujian, China, 2014-Ⅵ-25, 761 m | 25°43′37′′N 118°9′46′′E |
|
| Sycanus falleni Stål,1863 | DHX-10-COI DHX-11-COI DHX-13-COI |
3 | DHX1 DHX3 DHX6 DHX11 DHX13 DHX14 |
6 | Huashan, Ningming, Guangxi, China, 2014-VI-4, 127 m | 22°12′50″N 107°3′21″E |
| DHX-GX1-COI | 1 | DHGX7c DHGX9c DHGX10c |
3 | Longrui, Ningming, Guangxi, China, 2006-Ⅴ-18 | 22°12′50″N 107°3′21″E |
|
| DHX-GX2-COI | 1 | / | 0 | Nonggang, Guangxi, China, 2020-Ⅴ-27 | 22°28′16′′N 106°57′40′′E |
|
| Sycanus hsiaoi Li & Cai sp. nov. | XZ3-YN1-COI | 1 | / | 0 | Tianba, Qianliu, Lancang, Yunnan, China, 2017-Ⅶ-20, 1375 m | 22°53′52′′N 100°10′25′′E |
| XZ3-YN2-COI XZ3-YN3-COI |
2 | / | 0 | Gengma, Lincang, Yunnan, China, 2019-Ⅶ, 1118 m | 23°32′17′′N 99°23′49′′E |
|
|
Sycanus hainanensis Wang & Cai sp. nov. |
XZ2-HaN1-COI | 1 | / | 0 | Hainan Tropical Botanical Garden, Nada, Danzhou, Hainan, China, 2016-Ⅳ-11, 120 m | 19°30′42′′N 109°30′13′′E |
| XZ2-HaN2-COI | 1 | / | 0 | Hongxin, Yuanmen, Baisha, Hainan, China, 2008-Ⅹ-11, 228 m | 19°9′27′′N 109°29′10′′E |
|
| Sycanus insularis Hsiao, 1979 | HBX-HaN1-COI | 1 | / | 0 | Jianfengling, Ledong, Hainan, China, 2013-Ⅳ-12 | 18°44′34′′N 108°50′54′′E |
| HBX-HaN2-COI HBX-HaN3-COI |
2 | / | 0 | Tianchi, Jianfengling, Ledong, Hainan, China, 2015-V-07, 951 m | 18°44′34′′N 108°50′54′′E |
|
| Sycanus minor Hsiao,1979 | XX-HC-YN2-COI XX-HC-YN3-COI |
1 | / | 0 | Jinghong, Banna, Yunnan, China, 2009-V-3, 530m | 21°35′58′′N 100°28′41′′E |
| XX-YN10 XX-YN1-COI XX-YN3-COI XX-YN4 XX-YN5 XX-YN9 |
6 | / | 0 | Jiruo, Jinuo, Jinghong, Xishuangbanna, Yunnan, China, 2021-Ⅶ-22, 1090 m | 22°3′22′′N 101°59′7′′E |
|
| Sycanus rufus Hsiao, 1979 | HOX-YN2-COI HOX-YN3-COI HOX-YN4-COI |
3 | / | 0 | Yunfenggushanfeng, Jingmeng, Jinuo, Jinghong, Xishuangbanna, Yunnan, China, 2021-Ⅶ-20, 1070 m | 22°0′33′′N 100°49′12′′E |
|
Sycanus sichuanensis Hsiao, 1979 |
SCX-1-COI | 1 | SCX2 | 1 | Nanxi, Hekou, Yunnan, China, 2014-V-30, 117 m | 22°37′32′′N 103°56′53′′E |
| SCX-5-COI | 1 | SCX5 SCX6 |
2 | Fangxiang, Leishan, Guizhou, China, 2014-VII-22, 900 m | 26°26′21′′N 108°16′42′′E |
|
| SCX-8-COI | 1 | SCX8 | 1 | Maolan, Guizhou, China, 2007-V-28, 535 m | 25°23′45″N 108°4′45″E |
|
| SCX-11-COI | 1 | SCX11 | 1 | Daqingshan,Pingxiang, Chongzuo, Guangxi, China, 2014-VIII-8, 1000 m | 22°18′23″N 106°42′21″E |
|
| / | 0 | SCX12 | 1 | Changpi, Leishan, Guizhou, China, 2012-VII-21, 1008 m | 26°19′51′′N 108°4′30′′E |
|
| SCX-13-2-COI | 1 | / | 0 | Huashang, Ningming, Guangxi, China, 262 m | 22°15′24″N 107°0′56″E |
|
| SCX-GX1-COI | 1 | / | 0 | Lanhuagu park, Pingxiang, Guangxi, China, 2014-Ⅹ-8, 127 m | 22°12′50″N 107°3′21″E |
|
| SCX-GZ1-COI | 1 | / | 0 | Xiaodanjiang, Pingyang, Rongjiang, Guizhou, China, 2016-Ⅵ-20-24, 693m | 26°20′14′′N 108°20′41′′E |
|
| SCX-GX3-COI | 1 | / | 0 | Nonggang, Longzhou, Guangxi, China, 2003-Ⅷ-15 | 22°30′14′′N 106°59′0′′E |
|
| SCX-SC1 | 1 | / | 0 | Bailu, Pengzhou, Chengdu, Sichuan, China, 2018-VII-21, 800m | 31°9′32′′N 103°51′54′′E |
|
| SCX-SC2 | 1 | / | 0 | Emei Mountain, Leshan, Sichuan, China, 2014-Ⅶ-3, 2215 m | 29°32′50′′N 103°20′21′′E |
|
| Sycanus stali Dohrn, 1859 | ZYX-YN1-COI | 1 | / | 0 | Fula, Huimin, Lancang, Yunnan, China, 2017-VII-11, 1055 m | 22°15′32′′N 100°4′45′′E |
| ZYX-LC9-COI | 1 | / | 0 | Wanmu tea Garden, Nanping, Simao, Puer, Yunnan, China, 2022-VI-15, 1396 m | 22°44′50″N 100°58′32″E |
|
| Sycanus taiwanensis Zhao & Cai sp. nov. | XZ1-TW1-COI | 1 | / | 0 | Danayi valley, Alishan, Jiayi, Taiwan, China, 2012-Ⅵ-23, 42 m | 23°28′42′′N 120°27′11′′E |
| XZ1-TW2-COI | 1 | / | 0 | Fengganglilong mountain, Pingdong, Taiwan, China, 2015-I-26, 650 m | 22°11′33′′ 120°41′10′′ |
|
| XZ1-TW3-COI | 1 | / | 0 | Jialeshui, Manzhou, Pingdong, Taiwan, China, 2019-XI-08, 56 m | 21°59′20′′N 120°50′46′′E |
|
| Outgroup | ||||||
| Agriosphodrus dohrni (Signoret, 1862) | B-LC1-COI B-LC2-COI |
2 | BLC1 B1 |
2 | Kaili, Guizhou, China, 2017-IV-10, 700 m | 26°33′58″N 107°58′53″E |
| / | 0 | B2 | 1 | Guiyan, Guizhou, China, 2017-V-30, 1200 m | 26°38′48″N 106°37′41″E |
|
| Maldonadocoris annulipes Zhao, Yuan & Cai, 2006 | MS-HN-COI | 1 | MSHN | 1 | Wuzhishan, Hainan, China, 2022-V-10, 666 m | 18°53′0′′N 109°39′45′′E |
| Panthous ruber Hsiao, 1979 | / | 0 | NDLC1 ND2 |
2 | Ningming, Guangxi, China, 2014-VI-12, 119 m | 22°12′10″N 107°2′47″E |
| Yolinus albopustulatus China, 1940 | DQ-3-COI | 1 | DQ3 | 1 | Heishiding, Fengkai ,Zhaoqing, Guangdong, China, 2016-VIII-14, 920 m | 23°26′9″N 111°53′4″E |
| DQ-COI | 1 | DQ | 1 | Guiyang, Guizhou,China, 2016-IX-8, 1200 m | 26°38′48″N 106°37′41″E |
|
| Yolinus annulicornis Hsiao, 1979 | HJQ-LC-COI | 1 | / | 0 | Puer, Yunnan, China, 2022-VI-15, 1350 m | 22°49′30′′N 100°57′57′′E |
References
- Khadem, M. Deep interisland genetic divergence in the Macaronesian endemic Mosquito Ochlerotatus eatoni (Diptera: Culicidae), indication of cryptic species. J. Med. Entomol. 2015, 52, 1175–1180. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Du, Z.; Zhao, Q.; Li, D.; Shao, X.; Li, H.; Cai, W. Integrative taxonomy of the spinous assassin bug genus Sclomina (Heteroptera: Reduviidae: Harpactorinae) reveals three cryptic species based on DNA barcoding and morphological evidence. Insects 2021, 12, 251. [Google Scholar] [CrossRef]
- Lavery, S.D.; Farhadi, A.; Farahmand, H.; Chan, T.Y.; Azhdehakoshpour, A.; Thakur, V.; Jeffs, A.G. Evolutionary divergence of geographic subspecies within the scalloped spiny lobster Panulirus homarus (Linnaeus, 1758). PLoS ONE 2014, 9, e97247. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Yi, W.; Zheng, C.; Zhu, X.; Wang, S.; Xue, H.; Ye, Z.; Bu, W. Species delimitation of rice seed bugs complex: insights from mitochondrial genomes and ddRAD-seq data. Zool. Scr. 2022, 51, 185–198. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Ratnasingham, S.; deWaard, J.R. Barcoding animal life: cytochrome c oxidase subunit I divergences among closely related species. Proc. R. Soc. Lond. 2003, 270, S96–S99. [Google Scholar] [CrossRef]
- Floyd, R.M.; Wilson, J.J.; Hebert, P.D.N. DNA barcodes and insect biodiversity. In Insect Biodiversity: Science and Society; Foottit, R.G., Adler, P.H., Eds.; Wiley-Blackwel: Oxford, The United Kingdom, 2009; pp. 417–432. [Google Scholar]
- Hajibabaei, M.; deWaard, J.R.; Ivanova, N.V.; Ranasingham, S.; Dooh, R.T.; Kirk, S.L.; Mackie, P.M.; Hebert, P.D. Critical factors for assembling a high volume of DNA barcodes. Philos. Trans. R. Soc. Lond., B, Biol. Sci. 2005, 360, 1959–1967. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.; Duwal, R.K.; Lee, S. COI barcoding of true bugs (Insecta, Heteroptera). Mol. Ecol. Resour. 2011, 11, 266–270. [Google Scholar] [CrossRef]
- Kim, J.; Jung, S. COI barcoding of plant bugs (Insecta: Hemiptera: Miridae). PeerJ 2018, 6, e6070. [Google Scholar] [CrossRef]
- Kim, S.; Lee, Y.; Mutanen, M.; Seung, J.; Lee, S. High functionality of DNA barcodes and revealed cases of cryptic diversity in Korean curved-horn moths (Lepidoptera: Gelechioidea). Sci. Rep. 2020, 10, 6208. [Google Scholar] [CrossRef]
- Park, D.S.; Foottit, R; Maw, E; Hebert, P. D.N. Barcoding bugs: DNA-based identification of the true bugs (Insecta: Hemiptera: Heteroptera). PLoS ONE 2011, 6, e18749. [Google Scholar] [CrossRef]
- Schmidt, S.; Schmid-Egger, C.; Morinière, J; Haszprunar, G. ; Hebert, P. DNA barcoding largely supports 250 years of classical taxonomy: identifications for Central European bees (Hymenoptera, Apoidea partim). Mol. Ecol. Resour. 2015, 15, 985–1000. [Google Scholar] [CrossRef]
- Weis, A.; Meyer, R.; Dietz, L.; Doemel, J.S.; Leese, F.; Melzer, R.R. Pallenopsis patagonica (Hoek, 1881)—A species complex revealed by morphology and DNA barcoding, with description of a new species of Pallenopsis Wilson, 1881. Zool. J. Linn. Soc. Lond. 2014, 170, 110–131. [Google Scholar] [CrossRef]
- Zittra, C.; Wöss, G.; Van der Vloet, L.; Bakran-Lebl, K.; Barogh, B.S.; Sehnal, P.; Fuehrer, H.P. Barcoding of the genus Culicoides (Diptera: Ceratopogonidae) in Austria—An update of the species inventory including the first records of three species in Austria. Pathogens 2020, 9, 406. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.G.; Ning, X.; Yu, X.; Bu, W.J. Integrative species delimitation based on COI, ITS, and morphological evidence illustrates a unique evolutionary history of the genus Paracercion (Odonata: Coenagrionidae). PeerJ 2021, 9, e11459. [Google Scholar] [CrossRef] [PubMed]
- Hubert, N.; Hanner, R. DNA barcoding, species delineation and taxonomy: a historical perspective. Dna Barcodes 2015, 3, 44–58. [Google Scholar] [CrossRef]
- Hausmann, A.; Haszprunar, G.; Hebert, P.D. DNA barcoding the geometrid fauna of Bavaria (Lepidoptera): successes, surprises, and questions. PLoS One 2011, 6, e17134. [Google Scholar] [CrossRef] [PubMed]
- Pfeiler, E.; Nazario-Yepiz, N.O. DNA-based taxonomy and potential suppression of long-established names: The case of Telegonus fulgerator (Lepidoptera: Hesperiidae). Syst. Biodivers. 2020, 1–9. [Google Scholar] [CrossRef]
- Rudolf, M.; Kwong, S.; Gaurav, V.; Ng, P. DNA barcoding and taxonomy in diptera: a tale of high intraspecific variability and low identification success. Syst. Biol. 2006, 55, 715–728. [Google Scholar] [CrossRef]
- Zhang, H.; Bu, W. Exploring large-scale patterns of genetic variation in the COI gene among Insecta: implications for DNA barcoding and threshold-based species delimitation studies. Insects 2022, 13, 425. [Google Scholar] [CrossRef]
- Planas, E.; Ribera, C. Description of six new species of Loxosceles (Araneae: Sicariidae) endemic to the Canary Islands and the utility of DNA barcoding for their fast and accurate identification. Zool. J. Linn. Soc. Lond. 2015, 174, 47–73. [Google Scholar] [CrossRef]
- Fabricius, J.C. Species Inscctorum, Exhibentes Eorum Differentias Specificas, Synonyma Auctorum, Loca Natalia, Metamorphosin, Adjectis Observationibus, Descriptionibus. Hamburgi et Kilonii, impensis C. E. Bohnii, 1781, 2: 1–517. [CrossRef]
- Amyot, C.J.B.; Serville, J.G.A. Histoire Naturelle des Insectes, Hémiptères; Libraire Encyclopédique de Roret: Paris, France: i–lxxvi, 1843; pp. 1–675. [Google Scholar]
- Maldonado-Capriles, J. Systematic catalogue of the Reduviidae of the world (Insecta: Heteroptera). Caribbean Journal of Science, Special Edition, 1990; 1–694. [Google Scholar]
- Ishikawa, T.; Toriumi, W.; Susila, W. & Okajima, S. Sycanus aurantiacus (Hemiptera: Heteroptera: Reduviidae), a new harpactorine species from bali, indonesia, with brief notes on its biology. Zootaxa 2007, 1615, 21. [Google Scholar] [CrossRef]
- Fabricius, J.C. Mantissa insectorum sistens species nuper detectas adjectis synonymis, observationibus, descriptionibus, emendationibus. Christian Gottlieb Proft. Hafniae 1787, 2, 1–382. [Google Scholar]
- Dohrn, F.A. Beitrag zur Kenntnis der Harpactoridae. Stet. Ent. Zeit. 1859, 20, 91–99. [Google Scholar]
- Stål, C. Formae speciesque novae reduviidum. Annales de la Société Entomologique de France 1863, 3, 25–58. [Google Scholar] [CrossRef]
- Distant, W.L. Rhynchota 2 (Heteroptera). In The Fauna of British India, Including Ceylon and Burma; Taylor and Francis: London, The United Kingdom, 1903; pp. 1–242; 1904; pp. 243–503. [Google Scholar]
- Hsiao, T.Y. New species of Harpactorinae from China. I. (Hemiptera: Reduviidae). Acta Zootaxonomica Sinica 1979, 4, 137–155, [in Chinese, English summary]. [Google Scholar]
- Hsiao, T.Y.; Ren, S.Z. Reduviidae. In A Handbook for the Determination of the Chinese Hemiptera-Heteroptera (II) (In Chinese with English Summary); Hsiao, T.Y., Ren, S.Z., Zheng, L.Y., Jing, X.L., Zou, H.G., Liu, S.L., Eds.; Science Press: Beijing, China, 1981; pp. 390–538. [Google Scholar]
- Putshkov, P.V.; Putshkov, V.G. Family Reduviidae Latreille, 1807, assassin-bugs. In Catalogue of the Heteroptera of the Palaearctic Region, Cimicomorpha I; Putshkov, P.V., Putshkov, V.G., Eds.; The Netherlands Entomological Society: Amsterdam, The Netherlands, 1996; volume 2, pp. 254. [Google Scholar]
- Zhang, L.J.; Li, H.; Li, S.J.; Zhang, A.B.; Kou, F.; Xun, H.Z.; Wang, p.; Wang, Y.; Song, F.; Cui, J.X.; Gouge, D.H.; Cai, W.Z. Phylogeographic structure of cotton pest Adelphocoris suturalis (Hemiptera: Miridae): strong subdivision in China inferred from mtDNA and rDNA ITS markers. Sci. Rep., 2015, 5, 14009. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: assemble species by automatic partitioning. Mol. Ecol. Resour. 2021, 21, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Stål, C. Bidrag till Reduviidernas kännedom. Öfversigt af Kungliga Vetenskapsakademiens Förhandlingar 1867, 23, 235–302. [Google Scholar]
- Putshkov, V.G.; Putshkov, P.V.; Štys, P. Type-species designation and new names in Reduviidae (Heteroptera). Acta Entomologica Bohemoslovaka 1987, 84, 102–105. [Google Scholar]
- Stål, C. Enurneratio Hemipterorum. 4. Kungliga Svenska Vetenskapsakademiens Handlingar (N.F.) 1874, 12, 1–186. [Google Scholar]
- Lethierry, L.; Severin, G. Catalogue général des Hémiptères, Tome III. Hétéroptères. R. Friedländer & Fils, Libraires-Éditeurs: Berlin, German, 1896; pp. 1–275. [CrossRef]
- Putshkov, V.G.; Putshkov, P.V. A Catalogue of Assassin-bugs Genera of the World (Heteroptera, Reduviidae); VINITI: Moskva, Russia, 1985; p. 138. [Google Scholar]
- Zimsen, E. The type material of J.C. Fabricius; Munksgaard: Copenhagen, Denmark, 1964; pp. 1–656. [Google Scholar]
- Walker, F. Catalogue of the specimens of Hemiptera Heteroptera in the collection of the British Museum. Part VIII. Trustees: London, The United Kingdom, 1873; pp. 1–220.
- Distant, W.L. Rhynchotal Notes. XVI. Heteroptera: Family Reduviidae, Apiomerinae, Harpactorinae, and Nabidae. Ann. Mag. Nat. Hist. 1903, 11, 203–213, 245–258. [Google Scholar] [CrossRef]
- Gmelin, J.F. Caroli a Linné Systema Naturae (13th ed.). Lipsiae, 1790, 1: 1517–2224.
- Fabricius, J.C. Systema Rhyngotorum: secundum ordines, genera, species adjectis synonymis, locis, observationibus, descriptionibus; Reichard, Brunsvigae, Romania, 1803; i-vi, pp. 1–314. [CrossRef]
- Burmeister, H. Schnabelkerfe, Rhynchota, Abt. 1. Hemiptera: i-xii. In Handbuch der Entomologie, Band 2.; Enslin, Berlin, German, 1835; pp. 1–400. [CrossRef]
- Grandcolas, P. The origin of diversity in insects: speciation, adaptation and the earth dynamics. Comptes Rendus Biologies 2019, 342, 252–253. [Google Scholar] [CrossRef]
- Hou, Z.R.; Sun, B.B.; Liu, X.J.; Yin, Z.; Li, J.P.; Guo, X.H. Predatory functional response of bug Sycanus falleni to the larvae of fall armyworm Spodoptera frugiperda. J. Plant. Prot. 2020, 47, 852–858. [Google Scholar] [CrossRef]
- Wang, Y.N. Studies on the Biological Characteristics of Sycanus croceovittatus and its Control Effect on Spodoptera frugiperda. Master thesis, Hebei Agricultural University, Baoding, China, 2021. [CrossRef]
- Wang, Y.N.; Zhao, S.Y.; He, Y.Z.; Wu, K.M.; Li, G.P.; Feng, H.Q. Predation of the Larvae of Spodoptera frugiperda (J. E. Smith) by Sycanus croceovittatus Dohrn. Chinese Journal of Biological Control 2020, 36, 525–529. [Google Scholar] [CrossRef]
Figure 5.
Sycanus croceus Hsiao, 1979, (a) egg; (b–f) nymph; (g) adult. (a) egg mass; (b) first instar, hatching; (c) second instar; (d) third instar; (e) fourth instar; (f) fifth instar.
Figure 5.
Sycanus croceus Hsiao, 1979, (a) egg; (b–f) nymph; (g) adult. (a) egg mass; (b) first instar, hatching; (c) second instar; (d) third instar; (e) fourth instar; (f) fifth instar.

Figure 6.
Sycanus falleni Stål, 1863, (a) egg; (b–f) nymph; (g) adult. (a) egg mass; (b, c) first instar, hatching; (d) second instar; (e) third instar; (f) fourth instar; (g) fifth instar.
Figure 6.
Sycanus falleni Stål, 1863, (a) egg; (b–f) nymph; (g) adult. (a) egg mass; (b, c) first instar, hatching; (d) second instar; (e) third instar; (f) fourth instar; (g) fifth instar.

Figure 7.
Sycanus bicolor Hsiao, 1979, 二色犀猎蝽, habitus, (a-c), male, (d–f), female. (a, d), dorsal view; (b, e), lateral view; (c, f), ventral view. Scale bar (a–f) = 5.00 mm.
Figure 7.
Sycanus bicolor Hsiao, 1979, 二色犀猎蝽, habitus, (a-c), male, (d–f), female. (a, d), dorsal view; (b, e), lateral view; (c, f), ventral view. Scale bar (a–f) = 5.00 mm.

Figure 8.
Sycanus bicolor Hsiao, 1979, 二色犀猎蝽, (a–h), male external genitalia. (a, b), pygophore with paramere; (c, d), paramere; (e), struct; (f–h), phallus. (f), dorsal view; (b, g), lateral view; (a, h), ventral view. Scale bar of (a–h) = 0.50 mm.
Figure 8.
Sycanus bicolor Hsiao, 1979, 二色犀猎蝽, (a–h), male external genitalia. (a, b), pygophore with paramere; (c, d), paramere; (e), struct; (f–h), phallus. (f), dorsal view; (b, g), lateral view; (a, h), ventral view. Scale bar of (a–h) = 0.50 mm.

Figure 9.
Sycanus croceovittatus Dohrn, 1859, habitus, (a–c), male, (d–f), female. (a, d), dorsal view; (b, e), lateral view; (c, f), ventral view. Scale bar of (a–f)= 5.00 mm.
Figure 9.
Sycanus croceovittatus Dohrn, 1859, habitus, (a–c), male, (d–f), female. (a, d), dorsal view; (b, e), lateral view; (c, f), ventral view. Scale bar of (a–f)= 5.00 mm.

Figure 10.
Sycanus croceovittatus Dohrn, 1859, (a–h), male external genitalia. (a, b) pygophore with paramere; (c, d) paramere ; (e), struct; (f–i), phallus. (f), dorsal view; (b, g, i), lateral view; (a,h), ventral view. Scale bar of (a–h)= 1.00 mm.
Figure 10.
Sycanus croceovittatus Dohrn, 1859, (a–h), male external genitalia. (a, b) pygophore with paramere; (c, d) paramere ; (e), struct; (f–i), phallus. (f), dorsal view; (b, g, i), lateral view; (a,h), ventral view. Scale bar of (a–h)= 1.00 mm.
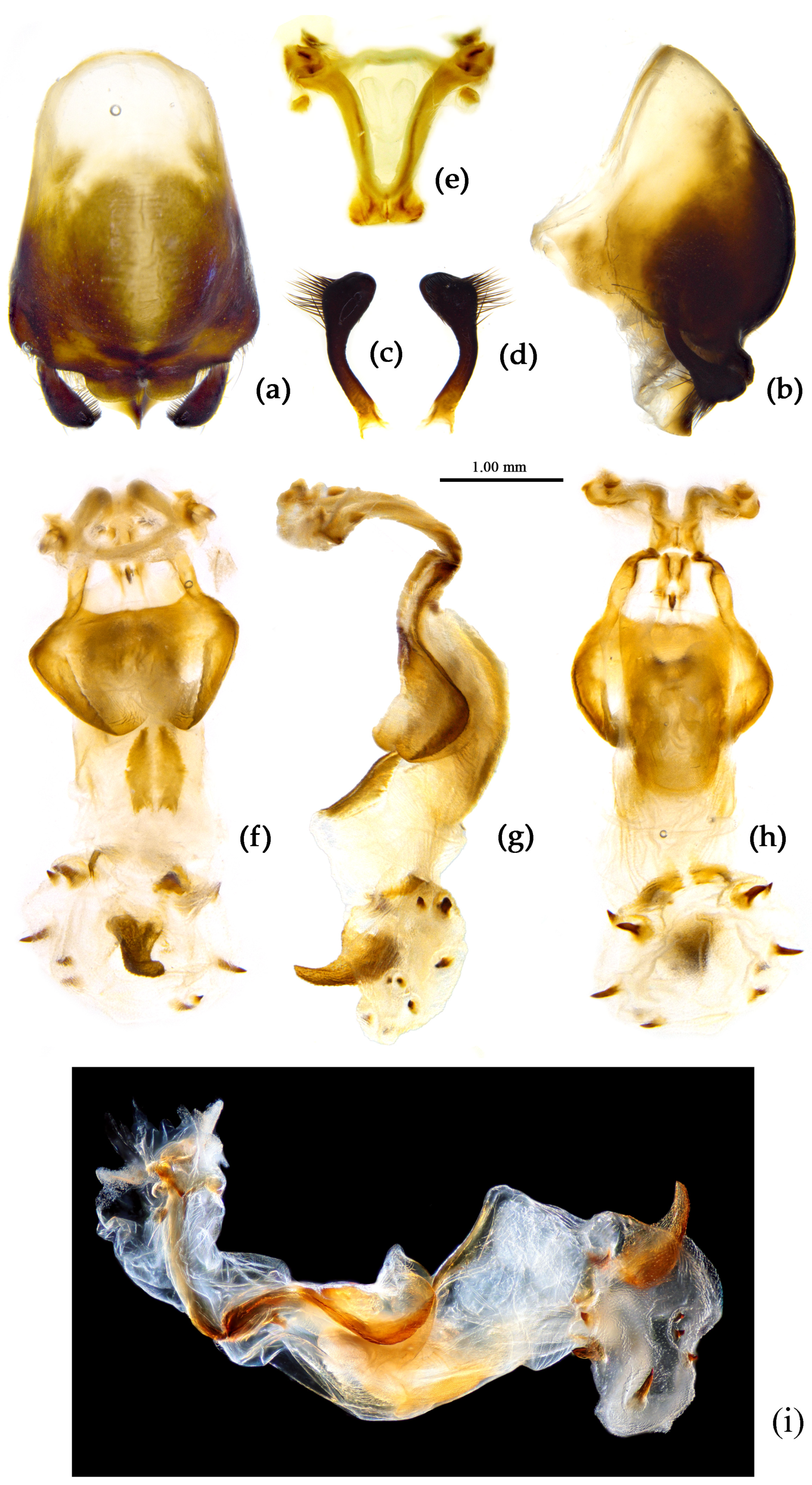
Figure 11.
Sycanus croceus Hsiao, 1979, Holotype, habitus, female. (a), dorsal view; (b), lateral view; (c), ventral view. Scale bar of (a–c)=5.00 mm.
Figure 11.
Sycanus croceus Hsiao, 1979, Holotype, habitus, female. (a), dorsal view; (b), lateral view; (c), ventral view. Scale bar of (a–c)=5.00 mm.

Figure 12.
Sycanus croceus Hsiao, 1979, (a–g), male external genitalia. (a, b) pygophore with paramere; (c) paramere; (d), struct; (e-g) phallus. (e) dorsal view; (b, f) lateral view; (a, g) ventral view. Scale bar of (a–g)= 0.50 mm.
Figure 12.
Sycanus croceus Hsiao, 1979, (a–g), male external genitalia. (a, b) pygophore with paramere; (c) paramere; (d), struct; (e-g) phallus. (e) dorsal view; (b, f) lateral view; (a, g) ventral view. Scale bar of (a–g)= 0.50 mm.
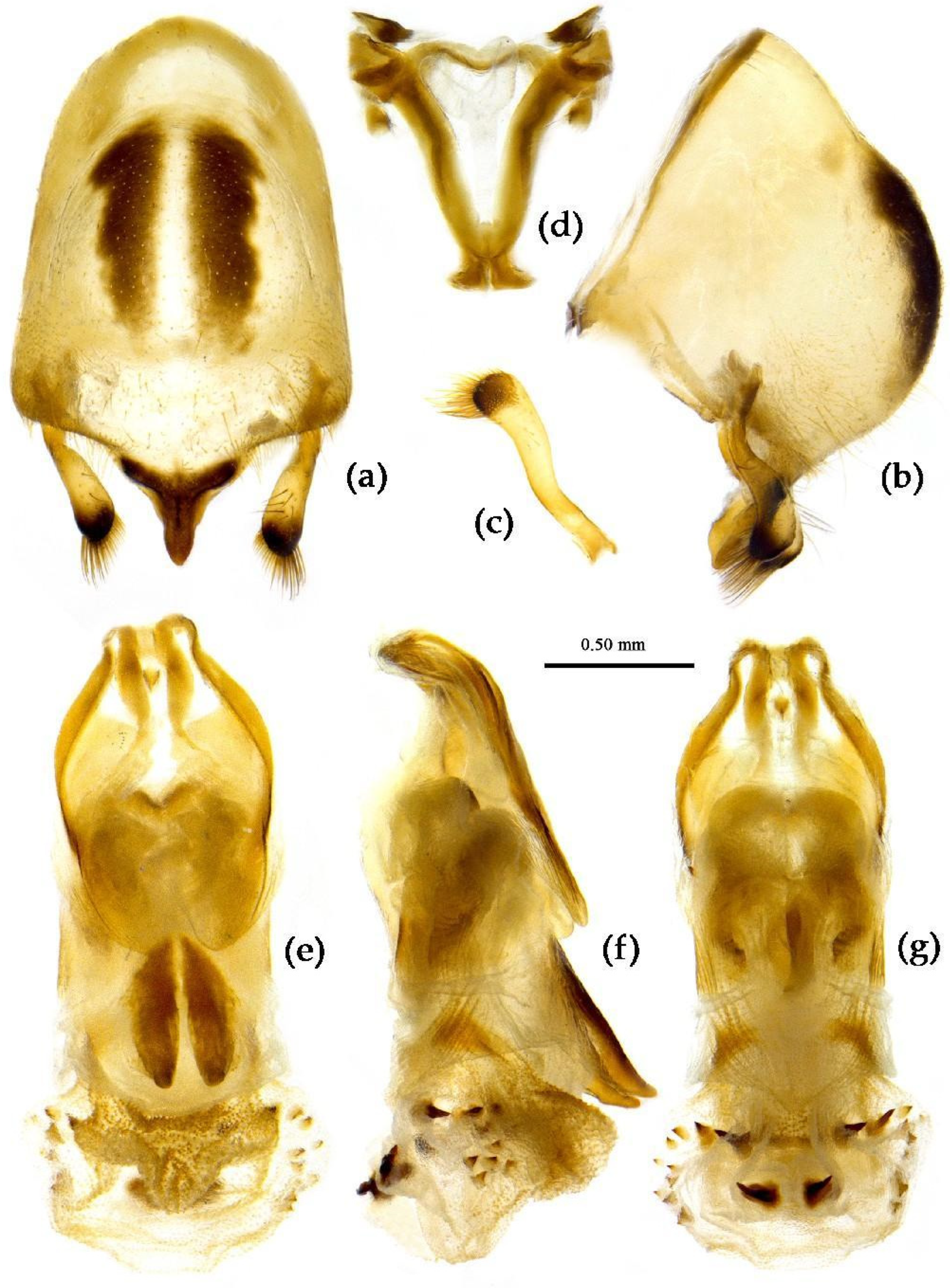
Figure 13.
Sycanus falleni Stål, 1863, habitus, (a–c), female; (d–f), male. (a, d), dorsal view; (b, e), ventral view; (c, f), lateral view. Scale bar of (a–f)= 5.00 mm.
Figure 13.
Sycanus falleni Stål, 1863, habitus, (a–c), female; (d–f), male. (a, d), dorsal view; (b, e), ventral view; (c, f), lateral view. Scale bar of (a–f)= 5.00 mm.

Figure 14.
Sycanus falleni Stål, 1863, (a–g), male external genitalia. (a, b) pygophore with paramere; (c) paramere; (d), struct; (e-g) phallus. (e) dorsal view; (b, f) lateral view; (a, g), ventral view. Scale bar of (a–g)= 0.50 mm.
Figure 14.
Sycanus falleni Stål, 1863, (a–g), male external genitalia. (a, b) pygophore with paramere; (c) paramere; (d), struct; (e-g) phallus. (e) dorsal view; (b, f) lateral view; (a, g), ventral view. Scale bar of (a–g)= 0.50 mm.
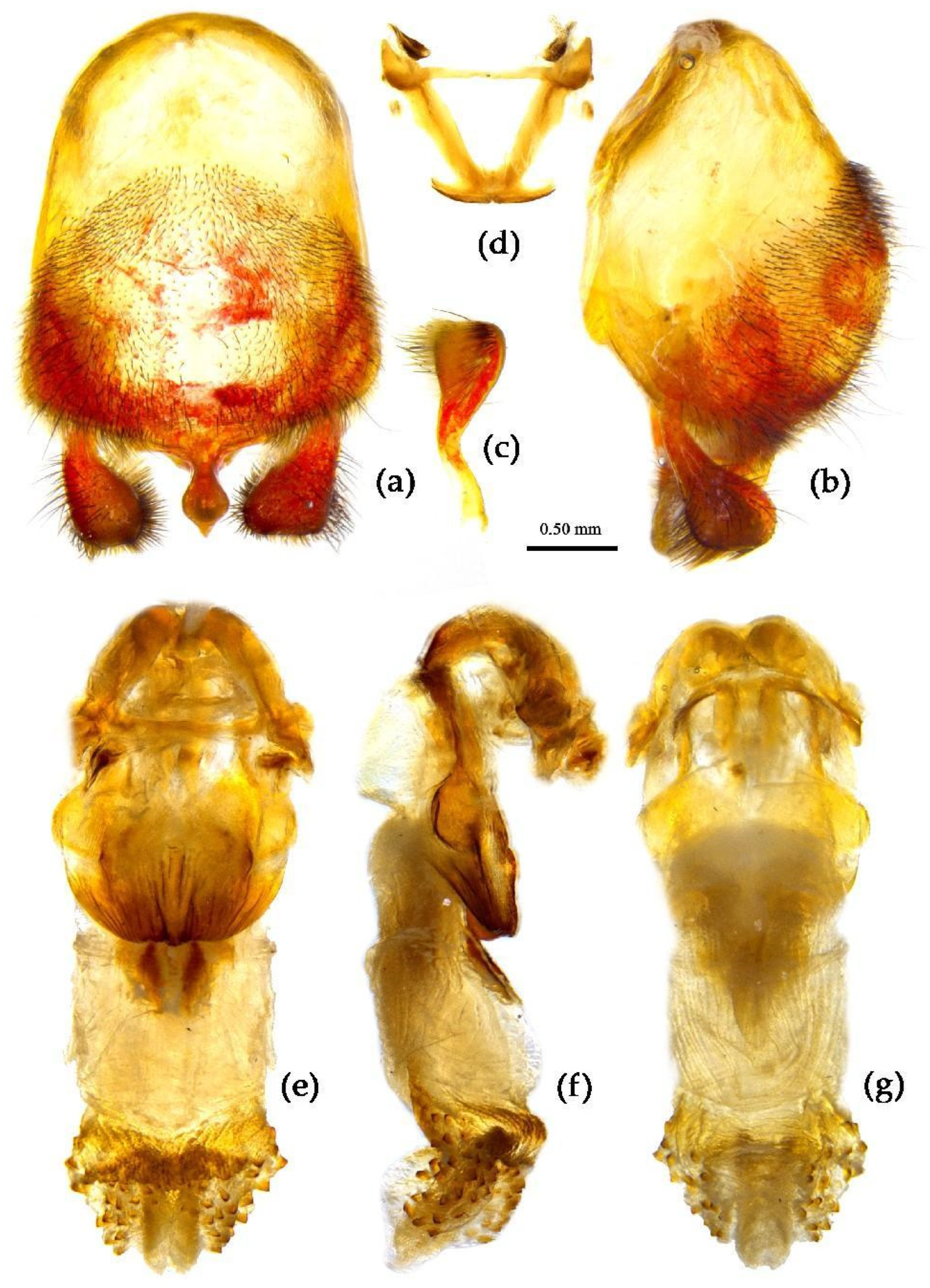
Figure 15.
Sycanus hsiaoi Li & Cai sp. nov., habitus, (a–c), Holotype, female, (d–f), Paratype, male. (a, d), dorsal view; (b, e), lateral view; (c, f), ventral view. Scale bar (a–f) = 5.00 mm.
Figure 15.
Sycanus hsiaoi Li & Cai sp. nov., habitus, (a–c), Holotype, female, (d–f), Paratype, male. (a, d), dorsal view; (b, e), lateral view; (c, f), ventral view. Scale bar (a–f) = 5.00 mm.
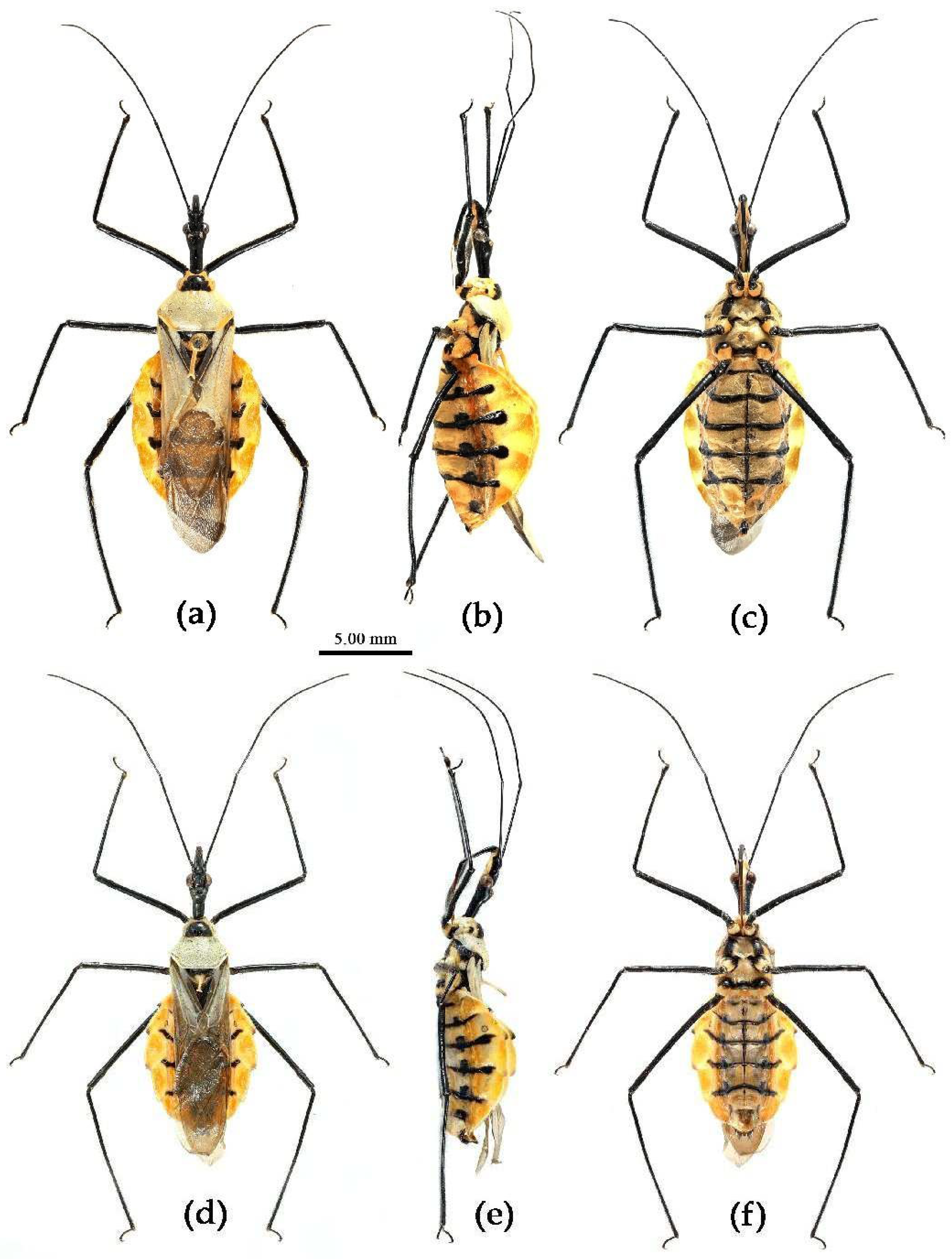
Figure 16.
Sycanus hsiaoi Li & Cai sp. nov., (a–g), Paratype, male external genitalia. (a, b), pygophore with paramere; (c), paramere; (d), strut; (e–g), phallus. (e) dorsal view; (b, f) lateral view; (a, g) ventral view. Scale bar of (a–g) = 0.50 mm.
Figure 16.
Sycanus hsiaoi Li & Cai sp. nov., (a–g), Paratype, male external genitalia. (a, b), pygophore with paramere; (c), paramere; (d), strut; (e–g), phallus. (e) dorsal view; (b, f) lateral view; (a, g) ventral view. Scale bar of (a–g) = 0.50 mm.

Figure 17.
Sycanus hainanensis Wang & Cai sp. nov., (a–c), Holotype, female; (d–f), Paratype, male. (a, d), dorsal view; (b, e), lateral view; (c, f), ventral view. Scale bar (a–f) = 5.00 mm.
Figure 17.
Sycanus hainanensis Wang & Cai sp. nov., (a–c), Holotype, female; (d–f), Paratype, male. (a, d), dorsal view; (b, e), lateral view; (c, f), ventral view. Scale bar (a–f) = 5.00 mm.

Figure 18.
Sycanus hainanensis Wang & Cai sp. nov., (a-g), Paratype, male external genitalia. (a, b), pygophore with paramere; (c), paramere; (d), strut; (e-g), phallus. (e) dorsal view; (b, f) lateral view; (a, g) ventral view. Scale bar of (a–g) = 0.50 mm.
Figure 18.
Sycanus hainanensis Wang & Cai sp. nov., (a-g), Paratype, male external genitalia. (a, b), pygophore with paramere; (c), paramere; (d), strut; (e-g), phallus. (e) dorsal view; (b, f) lateral view; (a, g) ventral view. Scale bar of (a–g) = 0.50 mm.
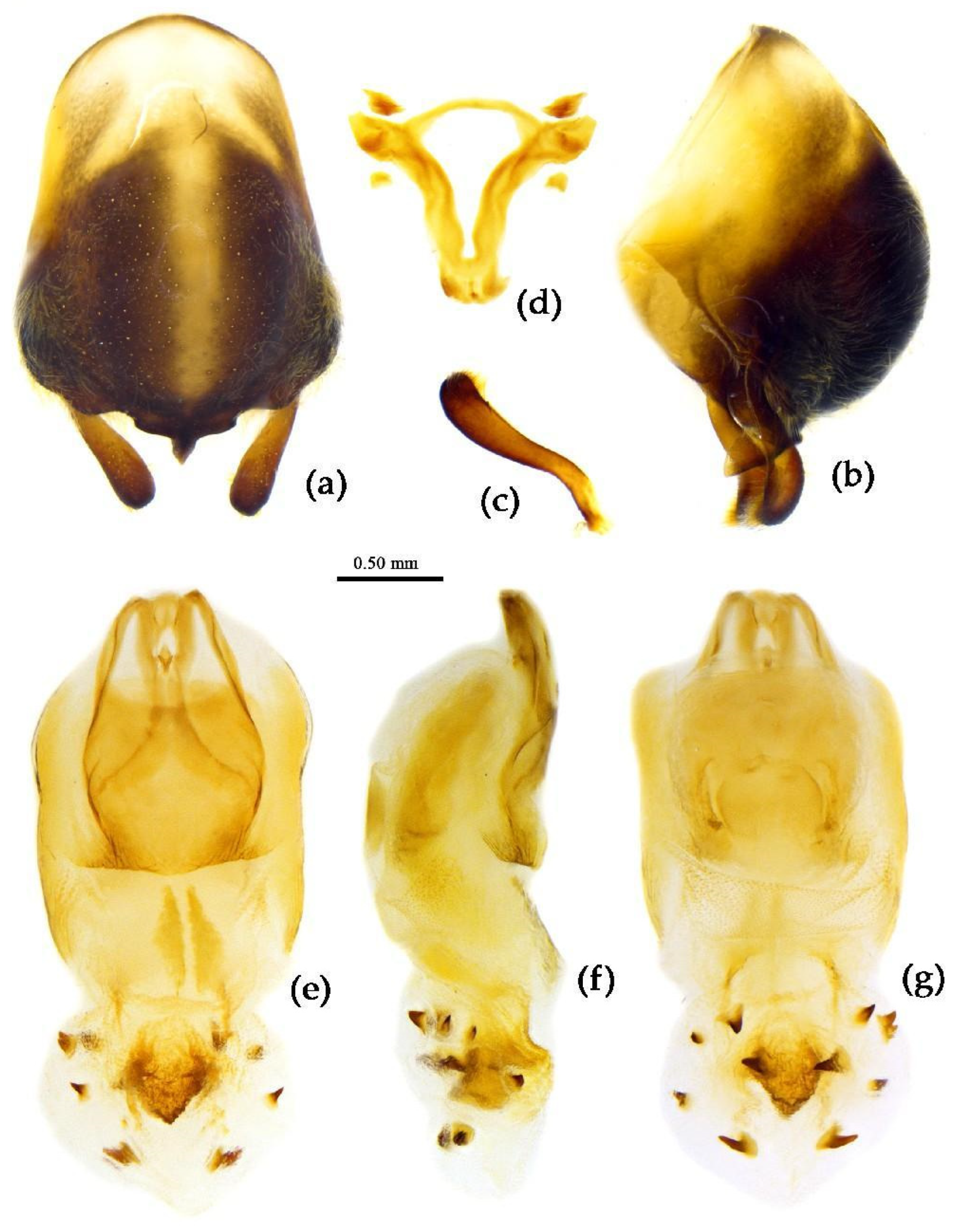
Figure 19.
Sycanus insularis Hsiao, 1979, ( a, b), male, Holotype; (c, d), female, Allotype, habitus. (a, c), dorsal view; (b, d), lateral view. Scale bar of (a, b)= 5.00 mm.
Figure 19.
Sycanus insularis Hsiao, 1979, ( a, b), male, Holotype; (c, d), female, Allotype, habitus. (a, c), dorsal view; (b, d), lateral view. Scale bar of (a, b)= 5.00 mm.

Figure 20.
Sycanus insularis Hsiao, 1979, (a–g), male external genitalia. (a, b) pygophore with paramere; (c) paramere; (d) struct; (e–g), phallus. (e), dorsal view; (b, f), lateral view; (a, g), ventral view. Scale bar of (a–g)= 0.50 mm.
Figure 20.
Sycanus insularis Hsiao, 1979, (a–g), male external genitalia. (a, b) pygophore with paramere; (c) paramere; (d) struct; (e–g), phallus. (e), dorsal view; (b, f), lateral view; (a, g), ventral view. Scale bar of (a–g)= 0.50 mm.

Figure 21.
Sycanus minor Hsiao, 1979, habitu, (a–c), female, (d-f), males. (a, d), dorsal view; (b, e), lateral view; (c, f), ventral view. Scale bar of (a–f)= 5.00 mm.
Figure 21.
Sycanus minor Hsiao, 1979, habitu, (a–c), female, (d-f), males. (a, d), dorsal view; (b, e), lateral view; (c, f), ventral view. Scale bar of (a–f)= 5.00 mm.
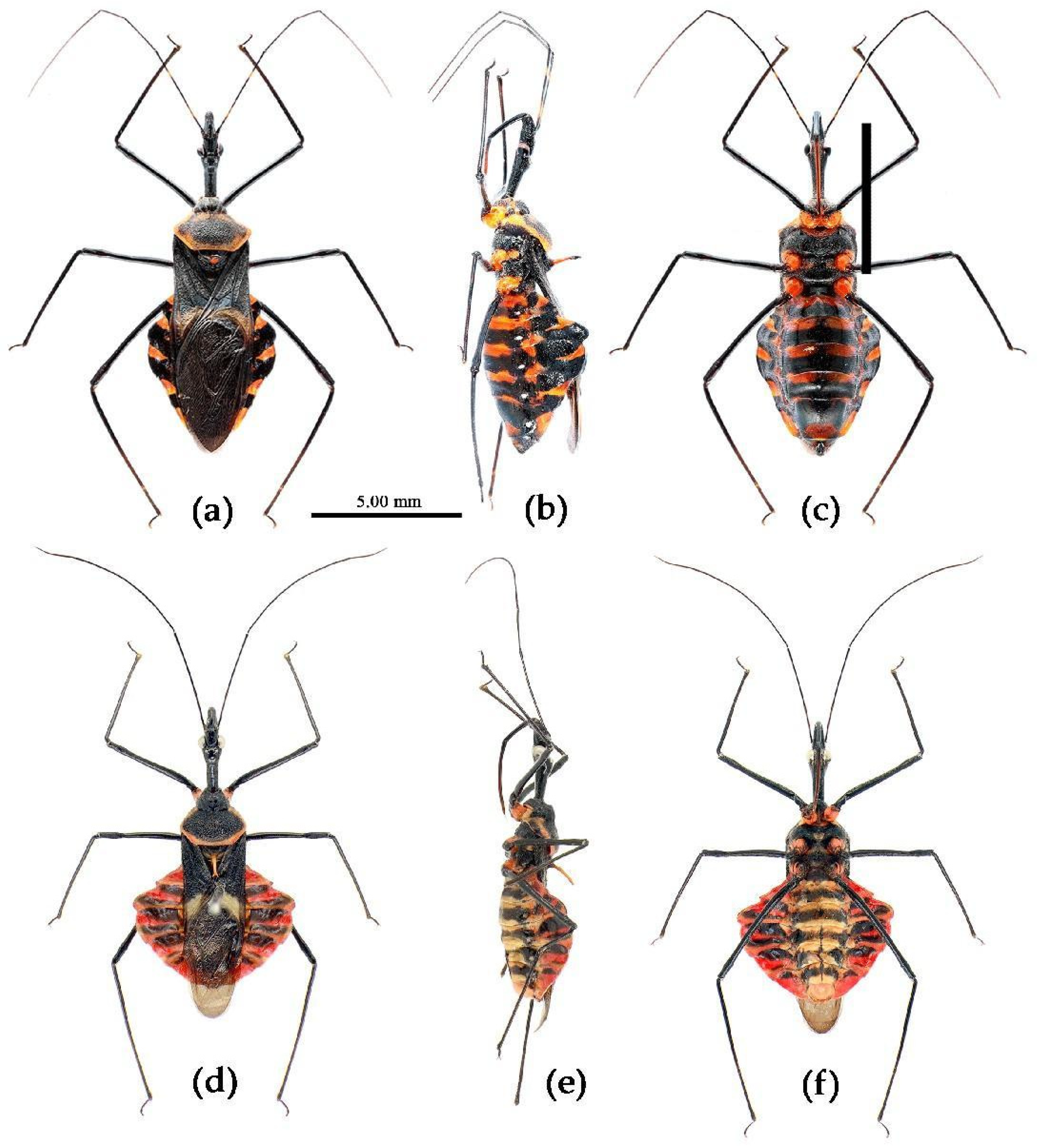
Figure 22.
Sycanus minor Hsiao, 1979, (a–g), male external genitalia. (a, b) pygophore with paramere; (c) paramere; (d), struct; (e–g), phallus. (e) dorsal view; (b, f) lateral view; (a, g) ventral view. Scale bar of (a–g)= 0.50 mm.
Figure 22.
Sycanus minor Hsiao, 1979, (a–g), male external genitalia. (a, b) pygophore with paramere; (c) paramere; (d), struct; (e–g), phallus. (e) dorsal view; (b, f) lateral view; (a, g) ventral view. Scale bar of (a–g)= 0.50 mm.
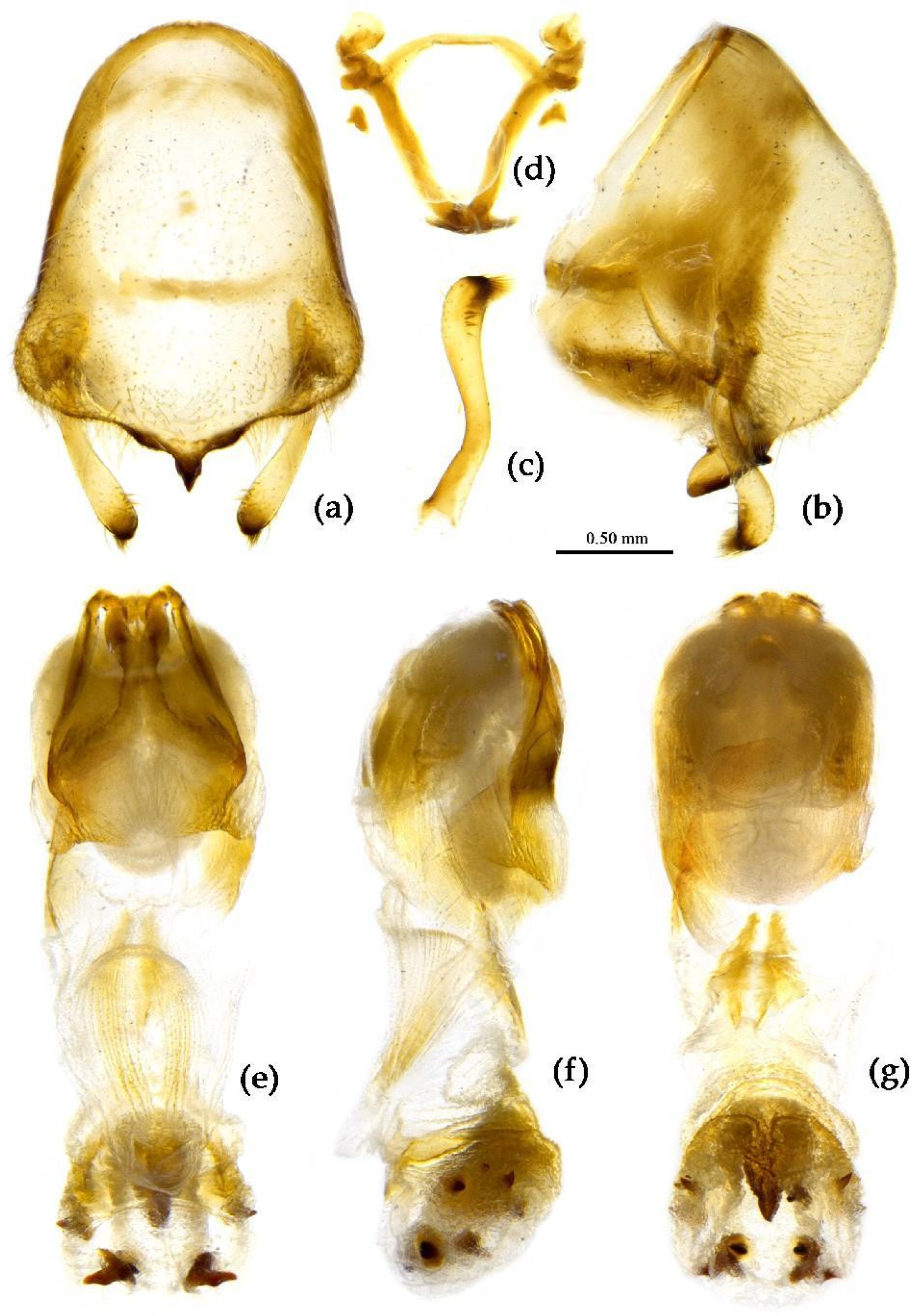
Figure 23.
Sycanus rufus Hsiao, 1979, Holotype, habitus, male. (a), dorsal view; (b), lateral view; (c), ventral view. Scale bar of (a–c)= 5.00 mm.
Figure 23.
Sycanus rufus Hsiao, 1979, Holotype, habitus, male. (a), dorsal view; (b), lateral view; (c), ventral view. Scale bar of (a–c)= 5.00 mm.

Figure 24.
Sycanus rufus Hsiao, 1979, (a–h), male external genitalia. (a, b), pygophore with paramere; (c, d), paramere; (e), struct; (f–h), phallus. (f), dorsal view; (b, g), lateral view; (a, h), ventral view. Scale bar of (a–h)= 0.50 mm.
Figure 24.
Sycanus rufus Hsiao, 1979, (a–h), male external genitalia. (a, b), pygophore with paramere; (c, d), paramere; (e), struct; (f–h), phallus. (f), dorsal view; (b, g), lateral view; (a, h), ventral view. Scale bar of (a–h)= 0.50 mm.
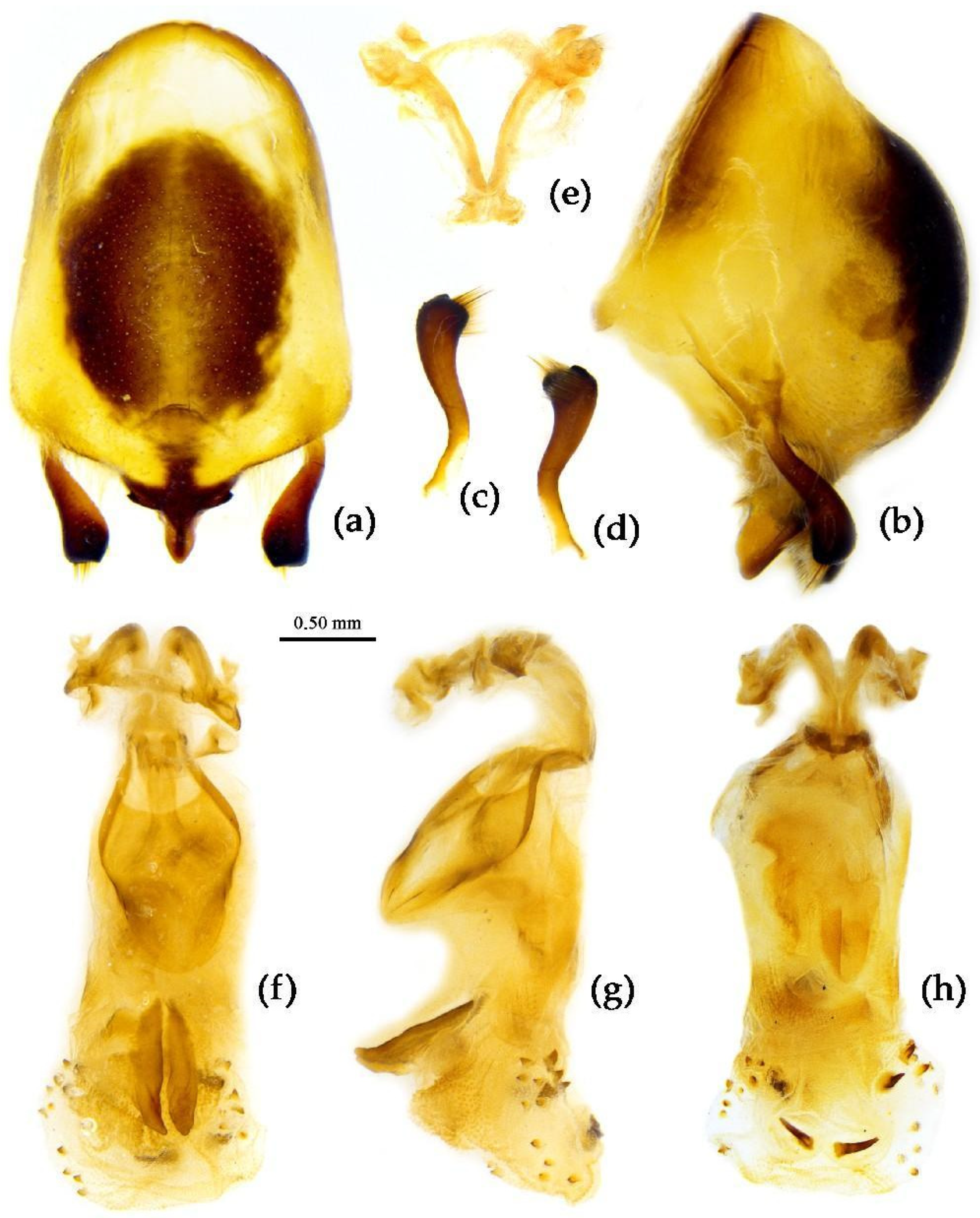
Figure 25.
Sycanus sichuanensis Hsiao, 1979, Holotype, habitus, male. (a), dorsal view; (b), lateral view; (c), ventral view. Scale bar of (a–c)= 5.00 mm.
Figure 25.
Sycanus sichuanensis Hsiao, 1979, Holotype, habitus, male. (a), dorsal view; (b), lateral view; (c), ventral view. Scale bar of (a–c)= 5.00 mm.

Figure 26.
Sycanus sichuanensis Hsiao, 1979, (a-g), male external genitalia. (a, b), pygophore with paramere; (c), paramere; (d), struct; (e–g), phallus. (e), dorsal view; (b, f), lateral view; (a, g), ventral view. Scale bar of (a–g)= 0.50 mm.
Figure 26.
Sycanus sichuanensis Hsiao, 1979, (a-g), male external genitalia. (a, b), pygophore with paramere; (c), paramere; (d), struct; (e–g), phallus. (e), dorsal view; (b, f), lateral view; (a, g), ventral view. Scale bar of (a–g)= 0.50 mm.
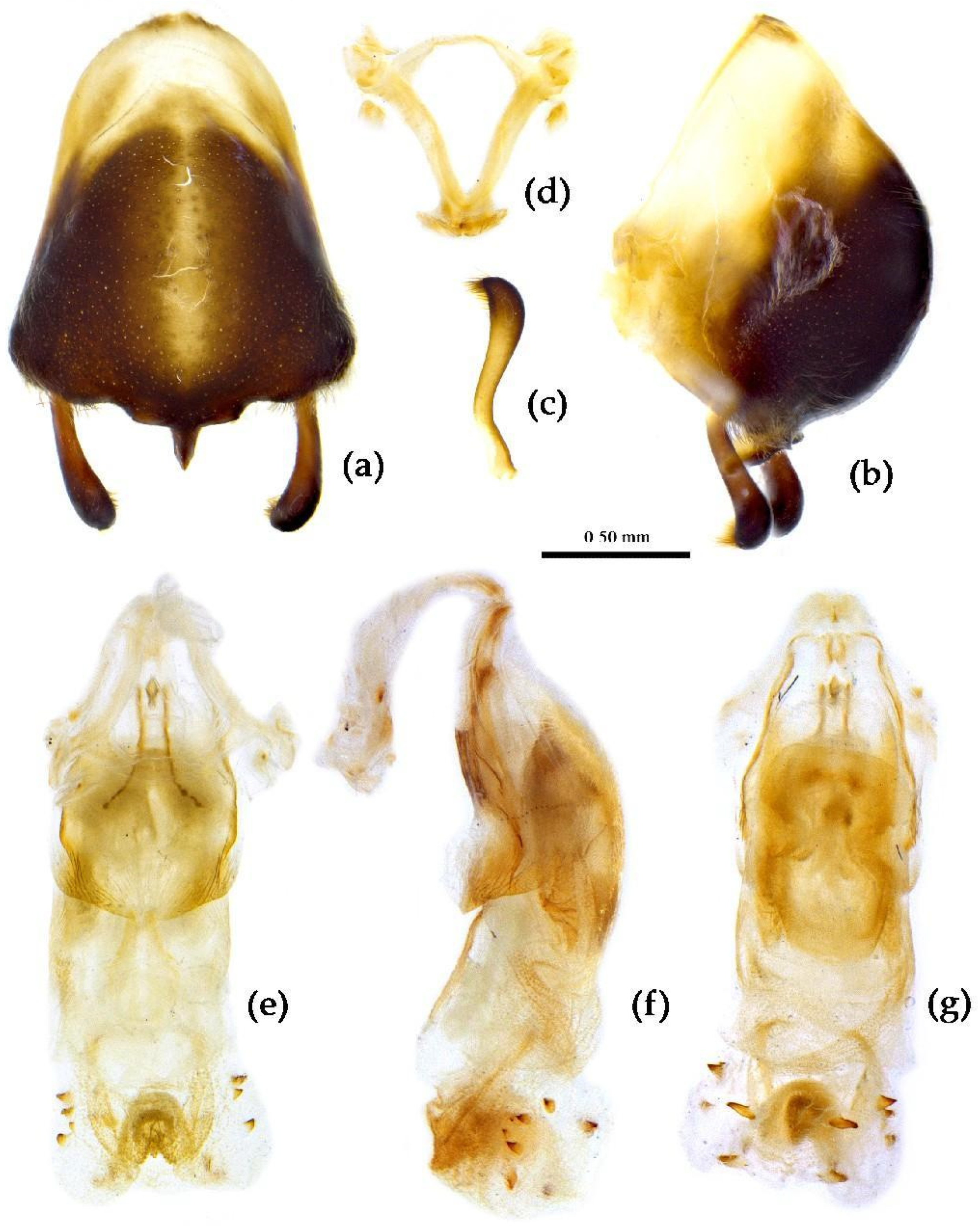
Figure 27.
Sycanus stali Dohrn, 1859, habitus, (a–c), female, (d–f), male. (a, d), dorsal view; (b, e), lateral view; (c, f), ventral view. Scale bar of (a–f)= 5.00 mm.
Figure 27.
Sycanus stali Dohrn, 1859, habitus, (a–c), female, (d–f), male. (a, d), dorsal view; (b, e), lateral view; (c, f), ventral view. Scale bar of (a–f)= 5.00 mm.

Figure 28.
Sycanus stali Dohrn, 1859, (a–g), male external genitalia. (a, b), pygophore with paramere; (c), paramere; (d), struct; (e–g), phallus. (e) dorsal view; (b, f) lateral view; (a, g) ventral view. Scale bar of (a–g)= 0.50 mm.
Figure 28.
Sycanus stali Dohrn, 1859, (a–g), male external genitalia. (a, b), pygophore with paramere; (c), paramere; (d), struct; (e–g), phallus. (e) dorsal view; (b, f) lateral view; (a, g) ventral view. Scale bar of (a–g)= 0.50 mm.

Figure 29.
Sycanus taiwanensis Zhao & Cai sp. nov., habitus, (a–c), Holotype, female, (d–f), Paratype, male. (a, d), dorsal view; (b, e), lateral view; (c, f), ventral view. Scale bar of (a–f)= 5.00 mm .
Figure 29.
Sycanus taiwanensis Zhao & Cai sp. nov., habitus, (a–c), Holotype, female, (d–f), Paratype, male. (a, d), dorsal view; (b, e), lateral view; (c, f), ventral view. Scale bar of (a–f)= 5.00 mm .

Figure 30.
Sycanus taiwanensis Zhao & Cai sp. nov., (a–g), Paratype, male external genitalia. (a, b), pygophore with paramere; (c), paramere; (d), struct; (e-g), phallus. (e) dorsal view; (b, f) lateral view; (a, g), ventral view. Scale bar of (a–g) = 0.50 mm.
Figure 30.
Sycanus taiwanensis Zhao & Cai sp. nov., (a–g), Paratype, male external genitalia. (a, b), pygophore with paramere; (c), paramere; (d), struct; (e-g), phallus. (e) dorsal view; (b, f) lateral view; (a, g), ventral view. Scale bar of (a–g) = 0.50 mm.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



