
Submitted:
24 March 2025
Posted:
25 March 2025
You are already at the latest version
Abstract
Background: Follicle stimulating hormone (FSH) is key regulator for follicular development, dif-ferentiation and maturation, the effects involve various intra follicular factors, such as members of FOXO subfamily. However, the specific role and mechanism of FOXO3 and FOXO4 in growth and development of hen follicles by affecting granulosa cell (GC) division and FSH response function are still unclear. Method: This study selected GCs from 6-8mm chicken follicles to detect FSH induced phosphorylation and nuclear rejection of FOXO3/4, and its regulatory effects on follicular GC pro-liferation, differentiation and apoptosis, using immunofluorescence, Western blotting, RT-qPCR and flow cytometry methods. Results: This study found that the expression level of FSHR mRNA was highest in GC treated with 10 ng/ml FSH for 8 and 12 hours. The level of p-FOXO3/4 protein sig-nificantly increased in cells treated with FSH for 12 hours, while the expression level of non phosphorylated FOXO3/4 significantly decreased. After co-treatment with 10ng/ml LMB, FOXO3/4 phosphorylation was effectively prevented. The immunofluorescence results showed that FOXO3 and FOXO4 were originally distributed in GC nucleus and cytoplasm, whereas they were almost accumulated in cytoplasm as treated with FSH for 12 h. Conversely, FOXO3/4 nuclear translocation was blocked by LMB. Moreover, RT-qPCR and flow cytometry results showed that FSH treatment significantly increased proliferation and differentiation of cell, but significantly reduced GCs apoptosis. However, LMB also eliminated these stimulating or inhibitory effects on cell proliferation. Conclusion: These findings provided new evidence that FSH induced FOXO3/4 nuclear rejection promoted GCs proliferation and reduced GCs apoptosis during hen follicular development.
Keywords:
chicken
; FOXO3/4
; nuclear exclusion
; cell proliferation
; granulosa cell
1. Introduction
Ovarian follicular development plays a pivotal role in poultry reproduction, affecting clutch traits and thus egg production [1], it primarily depends upon proliferation, differentiation and apoptosis of granulosa cells (GCs) and theca cells as well as oocyte growth and mature of hen ovary [2,3,4]. Among them, the GCs not only nurture the developing oocytes in follicles, but also play an essential role in folliculogenesis, follicle selection and a well-organized preovulatory hierarchy by governing steroidogenesis and release of the intra-ovarain factors, such as the steroidogenic acute regulatory protein (StAR) whose main function is to transport cholesterol from the outer membrane to the inner membrane of mitochondria for the synthesis and secretion of progesterone (P4) in granulosa cells of follicles [5]; cytochrome P450 family 11 subfamilies A member 1 (CYP11A1/P450scc) that plays a catalytic role in the hydrolysis of cholesterol to produce progesterone, is the rate-limiting enzymatic step within granulosa cells of the hen ovary [6,7]; Myelocytomatosis oncogene (c-MYC) is a well-known nuclear oncoprotein having multiple functions in cell proliferation, apoptosis and cellular transformation of several developing organs [8,9]; proliferating cell nuclear antigen (PCNA) is a central factor in cell cycle regulation and DNA replication, and shown to have a fundamental role in cellular proliferation [10,11]; B-cell lymphoma-2 (BCL-2) mainly serves as one anti-apoptotic member of the family by sequestering proforma of death-driving cysteine proteases [12], and the caspase-3 (CASP-3) acts as the main executioner of apoptosis in multicellular organisms and also in diverse biological processes, including cell differentiation, and proliferation, which is under the tight control of the BCL-2 family proteins [1,13]. More importantly, various members of gonadotropins and their receptors, such as follicle-stimulating hormone (FSH) and FSH receptor (FSHR) are also implicated in GC differentiation, maturation and steroidogenesis, and follicle development and selection [14,15,16].
FSH is a 35.5 kDa heterodimeric glycoprotein composed of two polypeptide units including alpha and beta subunits being synthesized and secreted by the anterior pituitary gonadotropic cells, acts as the primary hormonal inducer of ovarian follicle maturation and a critically important regulator of steroidogenesis in granulosa cells [17,18]. Its biological functions are activated through the interaction with its specific FSHR receptor in poultry [19]. FSH has been demonstrated to induce follicle selection marked by the highest expression levels of FSHR mRNA within the granulosa layer from individual prehierarchical follicles of 6-8 mm in diameter [15], and affect granulosa cells proliferation and differentiation which synthesize progesterone and estrogen crucial for the growth and maturation of preovulatory follicles promoted by the up-regulation of StAR, CYP11A1, and steroid 17-alpha-hydroxylase (CYP17) expression in hen ovary [3,20]. The function and regulatory mechanism of FSH in ovarian follicle growth, differentiation and maturation by its interaction with FSHR to activate a number of downstream effectors and intracellular signals have been well documented [2,21,22,23,24], but characterization of FSHR expression in the GCs of ovarian follicles induced by FSH remain to be further clarified. Furthermore, a wide variety of transcription factors, such as the members of forkhead box O (FOXO) subfamily were proved to be involved in ovarian follicle development and ovarian function by response to FSH signals [25,26,27], and the FOXO members mainly function to promote GC apoptosis and follicular atresia [25,28,29]. However, FSH plays the opposite role by facilitating GC proliferation and follicle growth, selection, and maturation [3,20,30,31]. Therefore, the contribution of the FOXO members to FSH signaling cascades has been attracting more and more researcher`s attention.
The members of FOXO family, including FOXO1 (also known as FOXO1A), FOXO3, FOXO4 and FOXO6 regulate many cellular programs, including cell proliferation, apoptosis, cell cycle transitions and DNA repair, autophagy, cell metabolism and responses to oxidative stress, which are of key importance in the decision over cell fate [32,33,34]. In mammalian ovaries, FOXO1 and FOXO3 were found to regulate follicular atresia by promoting GCs apoptosis [25,35]. Although the FSHR signals primarily via the cAMP/protein kinase A pathway [3,36], the previous studies showed that the phosphatidylinositol-3 kinase (PI3K)/3-PhosPhoinositide-dePendentProteinkinase-1 (PDK1) pathway is also activated by FSH in rat/mouse GCs, and the AKT activated downstream of PI3K in FSH-treated cells then phosphorylates FOXO1 to abrogate its transcriptional activity [34,35]. The FOXO3 is a proapoptotic molecule, and the expression levels of FOXO3 mRNA in GCs of porcine ovaries increased during atresia, while FOXO3 protein was abundant in granulosa cells of early atretic follicles [25]. In addition, the GC apoptosis was significantly and positively correlated with the total FoxO3 and negatively correlated with the phosphorylated FOXO3 protein (p-FOXO3) levels in the polycystic ovary syndrome (PCOS) patients [29]. Furthermore, the recent study has revealed that FOXO3 was expressed in the follicles at all stages, and the expression level in follicles increased with increasing follicle hierarchy, and promotes GC apoptosis of hen ovarian follicles [27].
Similar to other members of FOXOs, FOXO4 has been proved to play major roles in control of cellular proliferation, cell cycle arrest and apoptosis in mammals [37,38,39]. And the FOXO4 functions in a mechanism of cellular nuclear translocation and transcriptional regulation alike with its isoform FOXO3 factor although their molecular component and structure are divergent from each other [40,41]. FOXO4 can be phosphorylated by PI3K/PDK1 signaling on its two phosphorylation motif sites (Thr-32 and Ser-197) resulting in its inactivation and nuclear exclusion [42,43]. In the absence of PI3K/PDK1 activation, FOXO4 is located in the nucleus, where it functions as a transcription factor [44]. And in the presence of low levels of oxidative stress generated by treatment of cells with H2O2, FoxO4 becomes activated and remains in the nucleus. This effect appears to involve phosphorylation of FOXO4 by jun N-terminal kinase (JNK) at the two sites (thr- 447 and thr- 451) [45]. In addition, the monoubiquitylation on two conserved lysine residues (K199 and K211) of FOXO4 in response to increased cellular oxidative stress facilitates its re-localization to the nucleus and an increase in its transcriptional activity [46]. Recently, it has been revealed that the expression of FOXO4 is tightly controlled by microRNAs, and its subcellular localization is dependent on phosphorylation modification [37]. Thus, the nuclear re-localizations and functions of FOXO4 are firmly commanded by different signaling pathways that depended on the various physiological states or cellular processes. However, the exact roles and mechanisms of FOXO3 and FOXO4 (herein-after abbreviated as FOXO3/4) in prehierarchical follicular growth and development by controlling the GC proliferation and apoptosis under the FSH signals of chicken ovaries remain unclear.
In this study, we provide evidence to explain and support the claim that FSH induces phosphorylation and nuclear exclusion of the FOXO3/4 in granulosa cells of hen ovarian follicles. The FSH-induced nuclear exclusion of FOXO3/4 leads to the inhibition of cell apoptosis but increased proliferation and differentiation of granulosa cells. To the best of our knowledge, this study for the first time presents the novel data on the nuclear exclusion of FOXOs by involvement of FSH in hen ovarian granulosa cells. The current finding has laid a dependable foundation for further understanding the molecular regulatory mechanism of FOXO3/4 nuclear exclusion in follicular development and atresia of chicken ovary.
2. Materials and Methods
2.1. Ethics Statement
In this study, all procedures conducted in chickens were approved by the Institutional Animal Care and Use Committee (IACUC) of Jilin Agricultural University (Changchun, China). The animal experiment was performed in compliance with the ARRIVE guidelines [47]. Chickens were sacrificed before removing organs following the IACUC guidelines for experimental animals (Permission No. GR (J) 19-030). Euthanasia of the hens is fully compliant with the Chinese applicable laws and regulations concerning the care and use of laboratory animals, which was issued based on the Regulations for the Administration Affairs Concerning Experimental Animals of the State Council of the People’s Republic of China (2017 Revision). All of our efforts have been made to minimize the suffering of the animals.
2.2. Sampling and Granulosa Cell Culture
A commercial strain of Lohmann Brown laying hens was raised in laying batteries according to the standard husbandry practices as previously reported [48]. 22 hens were procured from the population and euthanized at 21 weeks of age, and the follicles of 6-8mm diameter size were immediately collected. Follicle GCs were subsequently isolated from the follicles and cultured according to a protocol that was previously reported by us [4]. A cell suspension was then prepared from the isolated GCs before adding M199 culture medium (Biosharp Life Sciences, Anhui, China) containing 10% of newborn calf serum, 100 IU/ml of penicillin, 75 IU/ml of streptomycin, and 10 ng/ml of insulin. The GCs was then incubated in a 5% CO2 and 95% air at 37℃. Cells were treated with 10 ng/ml FSH (R&D Systems, Shanghai, China) for 0 h, 8 h, 12 h, and 24 h, and then expression levels of FSHR gene were quantified. To evaluate expression of the genes (such as FOXO3/4 et al.) directly induced by FSH, cells were cultured with or without 10 ng/ml FSH for 12 h in the presence or absence of 10 ng/ml Leptomycin B (LMB; Beyotime, Shanghai, China).
2.3. Immunofluorescence Staining
In the process of cell culture, six-well tissue culture plates were filled with circular glass coverslips, which were placed there to promote cell growth. Following this, cells were immobilized by use of 4% paraformaldehyde (PFA, Beyotime, Shanghai, China), and the ends of the coverslips were homogeneously coated with an immunostaining guard pen (Biosharp Life Sciences, Anhui, China) to generate a 1mm oily barrier. The cells were then incubated with primary antibodies: anti-FOXO3 (Proteintech, Wuhan, China, 1:500 dilution), anti-FOXO4 (Bioss, Beijing, China, 1:500 dilution) at 4°C for 12 hours. After washing with PBS, the cells were incubated with fluorescent-conjugated goat anti-rabbit IgG antibody (Epizyme Biotech, Shanghai, China, 1:1000 dilution). Nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI, Epizyme Biotech, Shanghai, China). All immunofluorescence images were captured by laser confocal microscope (Zeiss).
2.4. Western Blot Analysis
The procedure made use of a RIPA lysis buffer (Biosharp Life Sciences, Anhui, China) to treat the cells for purposes of protein extraction from the samples. This was then proceeded by the adding of protease inhibitors cocktail (Epizyme Biotech, Shanghai, China) and phosphatase inhibitor cocktail (Epizyme Biotech, Shanghai, China), with subsequent sonication for fragmentation. The concentration of the protein was then adjusted following the instructions provided in the BCA protein assay kit (Beyotime, Shanghai, China). Nitrocellulose membrane transfer was then completed after electrophoresis at 150 V in a 10% SDS PAGE gel (Epizyme Biotech, Shanghai, China). After blocking with 5% nonfat milk, the membranes were incubated to the primary antibodies: anti-FOXO3 (Proteintech, Wuhan, China, 1:2000 dilution) and p-FOXO3 (Bioss, Beijing, China, 1:1000 dilution), anti-FOXO4 (Bioss, Beijing, China, 1:1000 dilution) and p-FOXO4 (Bioss, Beijing, China, 1:1000 dilution), and anti-β-actin (Boster Biological Technology, Wuhan, China, 1:10000 dilution) at 4℃ overnight. Then washing with PBS, and the samples were incubated with a horseradish peroxidase-conjugated goat anti-mouse IgG antibody as secondary antibody for 1.5 hours. Images were taken and visualized by an ECL Plus western blotting detection system according to the manufacturer’s instructions.
2.5. Quantitative Real-Time RT-PCR
To assess the mRNA expression of the target genes in the GCs, real-time quantitative reverse transcriptase PCR (qRT-PCR) was conducted according to our previously described method [48]. The used in this study were shown in Table 1. The reaction system contained SYBR 10 μl, 0.4 μl upper- and 0.4 μl lower-primer, and cDNA 2 μl. The amplification conditions used were 95℃ for 10 minutes and 95℃ for 15 seconds with an annealing temperature of 60℃ for 60 seconds and a total of 40 cycles. The 18S rRNA was used as a normalizing gene.
2.6. Flow Cytometric Analysis
The cells processed as explained earlier were digested with trypsin without EDTA and the cells collected. After centrifugation, the supernatant was ridden, and the cells were washed with PBS, suspended again, and re-centrifuged to eradicate the supernatant. Annexin V/PI double staining was used to detect the level of apoptosis of granulosa cells in different treatment groups. The prepared cell samples were stained according to the instructions of the annexin V-FITC/PI apoptosis test kit which was instantly analyzed using the CytoFlex flow cytometer. In accordance to the scatter plot of the bivariate flow cystoscope, the proliferating live cells were displayed as (FITC-/PI-) in the lower left quadrant. Necrotic cells, or non-viable cells, were displayed in the upper right quadrant as (FITC+/PI+). Some believe these cells to be non-viable apoptotic cells, or dead cells, which are represented as (FITC-/PI+). Apoptotic cells were exhibited in the lower right quadrant as (FITC+/PI-).
2.7. Statistical Analysis
Statistical analysis was implemented d using the SPSS12.0 software package (Sun et al., 2021). All the experiments were repeated at least three times using different batches of sampled birds. The data were analyzed with a one-way ANOVA and Tukey’s multiple-comparison test when more than two groups were involved or using a Student’s t-test when treatment and control groups were compared after confirming normal distributions for parametric analysis. P < 0.01 or P < 0.05 was considered to be statistically significant.
3. Results
3.1. The Expression of FSHR mRNA in the GCs Regulated by FSH
To reveal the effects of FSH signaling on the expression and phosphorylation of FOXO3/4 in the undifferentiated GCs by its interaction with the FSHR in the hen ovaries, the expression levels of FSHR mRNA in the GCs induced by FSH were firstly determined in the prehierarchical follicles of 6-8 mm diameter. As shown in Figure 1, expression levels of FSHR mRNA were dramatically elevated in the GCs cultured with 10ng/ml FSH for for 8 h or 12 h (P < 0.01). However, the levels of FSHR mRNA were then markedly declined in the GCs treated with 10 ng/ml FSH for 24 h (P < 0.05) although the FSHR mRNA expression levels were significantly higher compared with the controls (P < 0.05). Unexpectedly, the FSHR mRNA expression levels presented unchanged in the GCs under treatment with 12ng/ml FSH from 0 h to 12 h compared with the control group (P > 0.05). These results provided an evidence for FSH signaling to activate its FSHR pathway that may be involved in the regulation and function of FOXO family members in chicken ovarian follicle development.
3.2. FoxO3/4 Phosphorylation and Nuclear Exclusion Induced by FSH
In order to assess the actions of FSH on the phosphorylation and nuclear exclusion of the FOXO3/4 in the GCs from the ovarian prehierarchical follicles, the levels of phosphorylated FOXO3/4 (p-FOXO3/4) were analyzed by the western blotting method and their subcellular localizations in the cultured GCs were concomitantly inspected by the immunofluorescence analysis. The results demonstrated that expression levels of FOXO3/4 protein were decreased significantly in the cultured GCs with the treatment of 10ng/ml FSH and co-treatment of 10ng/ml FSH and Leptomycin B (LMB) for 12h compared to the control group (P < 0.01, Figure 2A,B). However, the levels of the p-FOXO3/4 were dramatically increased in the GCs under the FSH treatment for 12 h and this stimulatory action of FSH was effectively prevented by the co-treatment with 10 ng/ml LMB (P < 0.01). Moreover, either between FOXO3 and FOXO4 proteins or p-FOXO3 and p-FOXO4 molecules exhibited the greatly similar expression patterns in the GCs with the FSH treatment, respectively. Furthermore, the double immunofluorescence staining revealed that total FOXO3/4 (including FOXO3/4 and p-FOXO3/4) were localized in the GC nucleus and cytoplasm, whereas the nuclear FOXO3/4 were almost failed to be visualized in nuclei but mainly accumulated in the cytoplasm of the GCs treated with FSH for 12 h. This indicated that the nuclear FOXO3/4 was re-localized to the cytoplasm of the cells upon induction of FSH signaling. Conversely, this FSH-induced nuclear translocation of the FOXO3/4 in the cultured GCs was greatly blocked by the LMB (P < 0.01; Figure 3A,B). These findings revealed that FSH promotes phosphorylation and concomitant nuclear exclusion of FOXO3/4 in the cultured GCs, which may bring directly about changes of the effects of FOXO3/4 on ovarian follicle development.
3.3. Expression of the Genes Associated with Cell Proliferation, Differentiation and Apoptosis of the GCs Induced by FSH
To further explore biological functions of the FSH-induced phosphorylation and nuclear exclusion of FoxO3/4 in ovarian follicle development and growth, expression of a battery of candidate genes including MYC, PCNA, STAR, CYP11A1, BCL2, and CASP3 that are tightly associated with cell proliferation, differentiation and apoptosis of the GCs were examined. It has now been proved that the mRNA expression levels of the MYC, PCNA and BCL2 genes that involved in follicle development by promoting cell proliferation and anti-apoptosis, STAR and CYP11A1 genes that serve as indicators for GC differentiation were dramatically enhanced in the GCs by the treatment of FSH, while the expression level of the CASP3 mRNA was significantly reduced. However, all of these stimulatory or inhibitory effects induced by FSH on expression of the genes were predominantly abolished by the LMB (P < 0.01; Figure 4). These results indicated that the FOXO3/4, critical factors in promoting cell apoptosis oppositely pay the roles in invigorating GCs proliferation and differentiation but reducing apoptosis in response to the FSH-induced phosphorylation and nuclear exclusion during ovarian follicle growth and development.
3.4. Effects of FSH-Induced Phosphorylation and Nuclear Exclusion of FoxO3/4 on GCs Proliferation and Apoptosis
Based upon the molecular roles of the FSH-induced phosphorylation and nuclear exclusion of FoxO3/4 in regulation of the aforementioned genes, flow cytometric analysis was used to scrutinize proliferation and apoptotic rate of the cultured GCs with or without FSH treatment. As shown in Figure 5, a significantly higher cell proliferation rate and lower apoptotic rate of the GCs were observed in the FSH treatment group compared to the control group (P < 0.05). Nevertheless, these FSH-induced effects on proliferation and apoptosis of the GCs were significantly abrogated by the LMB (P < 0.05). These results solidify the claim that the transcriptional factors FOXO3/4 function to promote GC proliferation and apoptosis via the FSH signaling induced phosphorylation and nuclear exclusion of FOXO3/4 in ovarian follicle growth and development of hens.
4. Discussion
The ovarian follicle growth and development in chicken is a highly intricate and coordinated process involving a multitude of biological events controlled by reproductive hormones in the ovary [48,49,50], which predominantly contribute to egg productivity and reproductive performance of laying hens [4]. In this process, FSH signaling has a fundamental role in ovarian follicle development, follicle selection, and growth and differentiation (presenting a well-organized preovulatory hierarchy) by interaction with its receptor FSHR, a member of the rhodopsin-like G protein-coupled receptor family, being specifically expressed on ovarian follicular GCs [2,23,51,52], which subsequently activates the cAMP/protein kinase A pathway and indirectly initiates the production of several intra-ovarian factors and endocrine hormones, including CYP11A and STAR expression [21,22,24,53]. Moreover, the higher expression levels of FSHR in the GCs are required for follicle selection and immediately initiate a rapid growth in size of the pre-ovulatory follicles companied by GC differentiation and steroidogenesis [3,15,54]. However, in the earlier studies it was found that levels of FSHR expression were not responsive to the FSH challenges in the GCs from the ovarian prehierarchal follicles of 6-8 mm in diameter following the treatment with FSH for 1 h or 3 h, and simultaneously the cells failing to initiate a significant increase in cAMP production in vitro [15,55]. The reported results appeared paradox with our current data that levels of FSHR mRNA expression were dramatically elevated in the cultured GCs with 10 ng/ml FSH treatment for 8 h or 12 h, and which concomitantly brought about a significant increase of the STAR and CYP11A1 expression as well as a memorable augment of c-MYC and PCNA expression in the GCs and cell proliferation in the hen ovarian follicles. These present results corroborate the previous findings that the FSHR expression levels were remarkably elevated in the GCs of ovarian follicles challenged by FSH in chicken, human and mammals in vitro [27,56,57,58,59]. Additionally, Upon FSH stimulation, the notably raised FSHR expression levels were examined in the human embryonic kidney (HEK) 293 cells. FSHR expression in the CHO-cell system, as detected by FSH binding to receptors, was approximately threefold greater than expression in primary granulosa cells. Therefore, these studies have provided convincing evidence that in response to FSH stimulation, FSHR expression is significantly up-regulated in the GCs from the ovarian follicles of 6-8 mm in diameter, which corresponds to that a higher level of FSHR expression in the GCs, as aforementioned, is required for initiating the rapid follicle growth, differentiation and steroidogenesis of the hen ovary [3,54].
The FOXO family members as the crucial downstream regulators of FSH-induced signaling pathways, including FSHR/PI3K/AKT path, have recently been highlighted as an essential transcriptional factors of the important effectors involved in ovarian follicle growth, maturation and atresia by modulating the GC proliferation, differentiation and apoptosis [60,61,62,63]. Previous studies have well documented that FOXO3/4 belong to the FOXO subfamily of transcription factors and have several biological functions by regulating expression of genes associated GC proliferation, apoptosis, cell cycle arrest, autophagy and oxidative stress resistance [25,39,43,64]. In comparison, there is limited information on the regulatory mechanism and their roles in chicken ovarian GC proliferation, differentiation, and apoptosis, particularly, of which was induced by FSH signaling [61]. Prior study revealed the FOXO3 can be negatively regulated by its phosphorylation via the PI3K/Akt signaling pathway and ultimately drives apoptosis when activated in mammals [27]. In this study, we have demonstrated that the expression levels of FOXO3/4 were significantly decreased in the cultured GCs with stimulation of FSH; conversely levels of the p-FOXO3/4 were dramatically elevated in the cells. Whereby, the increased phosphorylation levels of FOXO3/4 were likely mainly caused by the activation of PI3K/AKT signaling under the stimulation of FSH as the previously reported [34,35,42]. Furthermore, relevant studies have shown that apoptosis was significantly and positively correlated with the total FoxO3 and negatively correlated with the p-FoxO3 protein levels in PCOS patients [29], and the FOXO3 is expressed in ovarian follicles and GCs, and promotes apoptosis of hen ovarian GCs [27]. Considering the results currently identified in this study, of which a memorable increase in cell proliferation but a remarkable decrement in apoptosis of the GCs were determined in the GCs by the treatment of FSH, accordingly, it was suggested that the FSH-induced phosphorylation of FOXO3/4 may facilitate the ovarian follicle growth and prevent follicular atresia by attenuating GC apoptosis of the chicken ovary.
To further investigate the exact roles and molecular mechanism of the p-FOXO3/4 in controlling the ovarian GC apoptosis, the FSH-induced nuclear exclusion of FoxO3/4 in GCs was examined in this work. Structural analysis has shown that all FOXO proteins contains a nuclear export sequence domain (NES) and a C-terminal transactivation domain (TAD) as well as one or two of the nuclear localization sequences (NLS) that contains the PKB motifs to be phosphorylated and a forkhead winged helix-turn-helix DNA binding domain [65,66]. Of which, the NLS domain mainly functions to shift FOXO towards the nucleus [67]; whereas phosphorylation of the PKB site in the forkhead domain leads to disruption of the NLS function and cytoplasmic retention of FOXO3 [68]. On the other hand, the NES domain was found to interact with the chromosomal region maintenance protein 1(CRM1), an evolutionarily conserved exportin 1 homolog that was shown to be a nuclear export receptor for NES-dependent protein transport [69,70,71]. Moreover, previous studies have proven that the NES is required for the translocation of FOXO3/4 from nucleus to cytoplasm [40,72]. The C-terminus of FOXOs containing a PKB phosphorylation site and a 14-3-3 binding motif, serves as a nuclear localization signal and the phosphorylation of FOXO3/4 leads to their translocation from nucleus to cytoplasm [73,74,75]. In the present study, the result of double immunofluorescence microscopy further showed that both FoxO3 and FoxO4 were distributed in the GC nucleus and cytoplasm, while the FOXO3/4 were almost failed to be observed in nuclei but accumulated in the cytoplasm of the GCs treated with FSH for 12 h. According to our best knowledge, it is the first time that this study has identified the effect of FSH stimulation on nuclear exclusion of FoxO3/4 in the cultured GCs of hen ovarian follicles. However, as the FOXO proteins are constantly exported from the nucleus in a CRM1-dependent manner, CRM1 inhibitors, such as LMB, can be used to demolish the nuclear export [76,77]. Here, it was found that as the levels of FOXO3/4 phosphorylation induced by FSH were increased dramatically in the GCs, simultaneously the nuclear exclusion of FOXO3 and FOXO4 occurred. But this stimulatory effect was dramatically abrogated by LMB. Obviously, this data in chicken ovary has provided another new evidence for the phosphorylation of FOXO3/4 to bring about the translocation the FOXO3/4 proteins from nucleus to cytoplasm because of the phosphorylation directly leading the retention of FOXO3/4 factors, and meanwhile the cellular relocation of the FOXO3/4 towards to cytoplasm being abolished by LMB as the aforementioned [68,73,74,75]. And this subcellular localization of FOXO3/4 is important for their activities and functions [65,75].
Additionally, this study demonstrated that the expression levels of the cell proliferation-related factors, including MYC, PCNA and BCL2 transcripts were dramatically enhanced in the GCs by the treatment of FSH, whereas the expression level of the CASP3 mRNA was significantly reduced, and this concomitantly resulted in a remarkable decrement in apoptosis of the GCs. The current data corresponds to the view that FOXO factors function as a tumor suppressor in human and mammals, which has been shown to regulate expression of genes implicated in cell proliferation, cell cycle arrest and apoptosis [25,41,43], and phosphorylation or nuclear exclusion of the FOXO is critical to promote cell proliferation and differentiation, but to inhibit apoptosis [39,64,78]. Intriguingly, it has earlier been reported that FOXO3 is a substrate of caspase-3-like proteases [79]; but the expressions of the CASP3 and BCL2 genes in this work were found to be modulated by FOXO3 and FOXO4 proteins, which directly influence the GC proliferation and apoptosis. These findings give us a notion that there might be a potential feedback-regulatory loop involving interactions between the FOXO3/4 factors and the caspase-3-like proteases, by which the normal homeostasis, cellular proliferation and differentiation of the prehierarchical follicles were maintained during chicken ovary development. A further study of elucidating the feedback mechanism is going on.
5. Conclusions
In summary, the current study initially demonstrated that expression levels of the phosphorylated FOXO3 and FOXO4 were significantly increased in the cultured GCs from the ovarian follicles challenged by FSH in chicken, whereas the expression levels of non-phosphorylated FOXO3/4 markedly decreased. Simultaneously, the FSH-induced nuclear translocation of FOXO3/4 in GCs was found, suggesting the phosphorylation of FOXO3/4 leads to their nuclear exclusion. Furthermore, the nuclear exclusion of FOXO3 and FOXO4 dramatically enhanced the expression of the MYC, PCNA, BCL2, STAR and CYP11A1 transcript levels but significantly reduced the expression of the CASP3 mRNA level in a similar manner, which concomitantly results in a memorable augment in cell proliferation but a remarkable decrement in apoptosis of the GCs. These findings provided novel evidence that the FSH-induced nuclear exclusion of FOXO3/4 promotes GC proliferation while attenuates the apoptosis of GCs in hen ovarian follicles.
Author Contributions
Conceptualization, R.X.; methodology, R.X., J.Z. and S.L.; formal analysis, J.Z., Y.S., S.L., X.S., N.Q. and R.X. ; investigation, S.L., N.Q. and R.X.; data curation, Y.S. and S.L.; writing—original draft preparation, J.Z., Y.S., S.L.; writing—review and editing, Y.S., C.Y. and N.Q.; supervision, R.X.; project administration, H.W.; funding acquisition, R.X. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the National Natural Science Foundation of China (Grant No. 32172714 and 31902145); the Project of Science and Technology Development Plan of Jilin Province (Grant No. 20220101306JC); the Project of Education Development Plan of Jilin Province (Grant No. JJKH20220362KJ); and the China Agriculture Research System of MOF and MARA (Grant No. CARS-41).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding author.
Acknowledgments
The author thanks the experimental chicken farm of Jilin Agricultural University for providing all the chickens.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Sun, X.; Liswaniso, S.; Shan, X.; Zhao, J.; Chimbaka, I.M.; Xu, R.; et al. The opposite effects of VGLL1 and VGLL4 genes on granulosa cell proliferation and apoptosis of hen ovarian prehierarchical follicles. Theriogenology. 2022, 181, 95-104. [CrossRef]
- Johnson, A.L.; Woods, D.C. Dynamics of avian ovarian follicle development: cellular mechanisms of granulosa cell differentiation. Gen Comp Endocrinol. 2009, 163, 12-17. [CrossRef]
- Johnson, A.L. Ovarian follicle selection and granulosa cell differentiation. Poult Sci. 2015, 94, 781-785. [CrossRef]
- Sun, X.; Niu, X.; Qin, N.; Shan, X.; Zhao, J.; Ma, C.; et al. Novel insights into the regulation of LATS2 kinase in prehierarchical follicle development via the Hippo pathway in hen ovary. Poult Sci. 2021, 100, 101454. [CrossRef]
- Johnson, A.L.; Bridgham, J.T. Regulation of steroidogenic acute regulatory protein and luteinizing hormone receptor messenger ribonucleic acid in hen granulosa cells. Endocrinology. 2001, 142, 3116-24. [CrossRef]
- Tilly, J.L.; Kowalski, K.I.; Johnson, A.L. Cytochrome P450 side-chain cleavage (P450scc) in the hen ovary. II. P450scc messenger RNA, immunoreactive protein, and enzyme activity in developing granulosa cells. Biol Reprod. 1991, 45, 967-74. [CrossRef]
- Stocco, D.M. StAR protein and the regulation of steroid hormone biosynthesis. Annu Rev Physiol.2001, 63, 193-213. [CrossRef]
- Morgenbesser, S.D.; Schreiber-Agus, N.; Bidder, M.; Mahon, K.A.; Overbeek, P.A.; Horner, J.; et al. Contrasting roles for c-Myc and L-Myc in the regulation of cellular growth and differentiation in vivo. EMBO J. 1995, 14, 743-56. [CrossRef]
- Bhandari, D.R.; Seo, K.W.; Jung, J.W.; Kim, H.S.; Yang, S.R.; Kang, K.S. The regulatory role of c-MYC on HDAC2 and PcG expression in human multipotent stem cells. J Cell Mol Med. 2011, 15, 1603-14. [CrossRef]
- Hao, H.; Xin, T.; Nancai, Y.; Yanxia, W.; Qian, L.; Wei, M.; et al. Short-interfering RNA-mediated silencing of proliferating cell nuclear antigen inhibit proliferation and induce apoptosis in HeLa cells. Int J Gynecol Cancer. 2008, 18, 36-42. [CrossRef]
- Strzalka, W.; Ziemienowicz, A. Proliferating cell nuclear antigen (PCNA): a key factor in DNA replication and cell cycle regulation. Ann Bot. 2011, 107, 1127-40. [CrossRef]
- Tsujimoto, Y. Role of Bcl-2 family proteins in apoptosis: apoptosomes or mitochondria? Genes Cells. 1998, 3, 697-707. [CrossRef]
- Ola, M.S.; Nawaz, M.; Ahsan, H. Role of Bcl-2 family proteins and caspases in the regulation of apoptosis. Mol Cell Biochem. 2011, 351, 41-58. [CrossRef]
- Tilly, J.L.; LaPolt, P.S.; Hsueh, A.J. Hormonal regulation of follicle-stimulating hormone receptor messenger ribonucleic acid levels in cultured rat granulosa cells. Endocrinology. 1992, 130, 1296-302. [CrossRef]
- Woods, D.C.; Johnson, A.L. Regulation of follicle-stimulating hormone-receptor messenger RNA in hen granulosa cells relative to follicle selection. Biol Reprod. 2005, 72, 643-50. [CrossRef]
- de Souza, D.K.; Salles, L.P.; Camargo, R.; Gulart, L.V.M.; Costa, E. Silva, S.; de Lima, B.D.; et al. Effects of PI3K and FSH on steroidogenesis, viability and embryo development of the cumulus-oocyte complex after in vitro culture. Zygote. 2018, 26, 50-61. [CrossRef]
- Urban, R.J.; Garmey, J.C.; Shupnik, M.A.; Veldhuis, J.D. Follicle-stimulating hormone increases concentrations of messenger ribonucleic acid encoding cytochrome P450 cholesterol side-chain cleavage enzyme in primary cultures of porcine granulosa cells. Endocrinology. 1991, 128, 2000-2007. [CrossRef]
- Bhartiya, D.; Patel, H. An overview of FSH-FSHR biology and explaining the existing conundrums. J Ovarian Res. 2021, 14, 144. [CrossRef]
- Zhao, L.H.; Chen, J.L.; Xu, H.; Liu, J.W.; Xu, R.F. Cloning and Expression of FSHb Gene and the Effect of FSH on the mRNA Levels of FSHR in the Local Chicken. Asian-Australasian Journal of Animal Sciences. 2010, 23, 292-301.
- Johnson, A.L.; Lee, J. Granulosa cell responsiveness to follicle stimulating hormone during early growth of hen ovarian follicles. Poult Sci. 2016, 95, 108-14. [CrossRef]
- Sassone-Corsi, P. Coupling gene expression to cAMP signalling: role of CREB and CREM. Int J Biochem Cell Biol. 1998, 30, 27-38. [CrossRef]
- Francis, S.H.; Corbin, J.D. Cyclic nucleotide-dependent protein kinases: intracellular receptors for cAMP and cGMP action. Crit Rev Clin Lab Sci. 1999, 36, 275-328. [CrossRef]
- Das, N.; Kumar, T.R. Molecular regulation of follicle-stimulating hormone synthesis, secretion and action. J Mol Endocrinol. 2018, 60, R131-55. [CrossRef]
- Casarini, L.; Crépieux, P. Molecular Mechanisms of Action of FSH. Front Endocrinol (Lausanne). 2019, 10, 305. [CrossRef]
- Matsuda, F.; Inoue, N.; Maeda, A.; Cheng, Y.; Sai, T.; Gonda, H.; et al. Expression and function of apoptosis initiator FOXO3 in granulosa cells during follicular atresia in pig ovaries. J Reprod Dev. 2011, 57, 151-8. [CrossRef]
- Shen, M.; Lin, F.; Zhang, J.; Tang, Y.; Chen, W.K.; Liu, H. Involvement of the up-regulated FoxO1 expression in follicular granulosa cell apoptosis induced by oxidative stress. J Biol Chem. 2012, 287, 25727-40. [CrossRef]
- Cui, C.; Han, S.; Yin, H.; Luo, B.; Shen, X.; Yang, F.; et al. FOXO3 is expressed in ovarian tissues and acts as an apoptosis initiator in granulosa cells of chickens. Biomed Res Int. 2019, 6902906. [CrossRef]
- Shen, M.; Lin, F.; Zhang, J.; Tang, Y.; Chen, W.K.; Liu, H. Involvement of the up-regulated FoxO1 expression in follicular granulosa cell apoptosis induced by oxidative stress. J Biol Chem. 2012, 287, 25727-40. [CrossRef]
- Mikaeili, S.; Rashidi, B.H.; Safa, M.; Najafi, A.; Sobhani, A.; Asadi, E.; et al. Altered FoxO3 expression and apoptosis in granulosa cells of women with polycystic ovary syndrome. Arch Gynecol Obstet. 2016, 294, 185-92. [CrossRef]
- Chun, S.Y.; Eisenhauer, K.M.; Minami, S.; Billig, H.; Perlas, E.; Hsueh, A.J. Hormonal regulation of apoptosis in early antral follicles: follicle-stimulating hormone as a major survival factor. Endocrinology. 1996; 1447-56. [CrossRef]
- Hunzicker-Dunn, M.; Maizels, E.T. FSH signaling pathways in immature granulosa cells that regulate target gene expression: branching out from protein kinase A. Cell Signal. 2006, 18, 1351-9. [CrossRef]
- Huang, H.; Tindall, D.J. Dynamic FoxO transcription factors. J Cell Sci. 2007, 120, 2479-87. [CrossRef]
- de Keizer, P.L.; Burgering, B.M.; Dansen, T.B. Forkhead box o as a sensor, mediator, and regulator of redox signaling. Antioxid Redox Signal. 2011, 14, 1093-106. [CrossRef]
- Herndon, M.K.; Law, N.C.; Donaubauer, E.M.; Kyriss, B.; Hunzicker-Dunn, M. Forkhead box O member FOXO1 regulates the majority of follicle-stimulating hormone responsive genes in ovarian granulosa cells. Mol Cell Endocrinol. 2016, 434, 116-26. [CrossRef]
- Shen, M.; Liu, Z.; Li, B.; Teng, Y.; Zhang, J.; Tang, Y.; et al. Involvement of FoxO1 in the effects of follicle-stimulating hormone on inhibition of apoptosis in mouse granulosa cells. Cell Death Dis. 2014, 5, e1475. [CrossRef]
- Richards, J.S.; Russell, D.L.; Ochsner, S.; Hsieh, M.; Doyle, K.H.; Falender, A.E.; et al. Novel signaling pathways that control ovarian follicular development, ovulation, and luteinization. Recent Prog Horm Res. 2002, 57, 195-220. [CrossRef]
- Wang, W.; Zhou, P.H.; Hu, W. Overexpression of FOXO4 induces apoptosis of clear-cell renal carcinoma cells through downregulation of Bim. Mol Med Rep. 2016, 13, 2229-34. [CrossRef]
- Lu, C.; Yang, Z.; Jiang, S.; Yang, Y.; Han, Y.; Lv, J.; et al. Forkhead box O4 transcription factor in human neoplasms: Cannot afford to lose the novel suppressor. J Cell Physiol. 2019, 234, 8647-58. [CrossRef]
- Liu, W.; Li, Y.; Luo, B. Current perspective on the regulation of FOXO4 and its role in disease progression. Cell Mol Life Sci. 2020, 77, 651-63. [CrossRef]
- Van Der Heide, L.P.; Hoekman, M.F.; Smidt, M.P. The ins and outs of FoxO shuttling: mechanisms of FoxO translocation and transcriptional regulation. Biochem J. 2004, 380, 297-309. [CrossRef]
- Santos, B. F.; Grenho, I.; Martel, P. J.; Ferreira, B. I.; Link, W. FOXO family isoforms. Cell Death Dis. 2023, 14, 702. [CrossRef]
- Matsuzaki, H.; Ichino, A.; Hayashi, T.; Yamamoto, T.; Kikkawa, U. Regulation of intracellular localization and transcriptional activity of FOXO4 by protein kinase B through phosphorylation at the motif sites conserved among the FOXO family. J Biochem. 2005, 138, 485–491. [CrossRef]
- Lüpertz, R.; Chovolou, Y.; Unfried, K.; Kampkötter, A.; Wätjen, W.; Kahl, R. The forkhead transcription factor FOXO4 sensitizes cancer cells to doxorubicin-mediated cytotoxicity. Carcinogenesis. 2008, 29, 2045–2052. [CrossRef]
- Manning, B. D.; Cantley, L. C. AKT/PKB signaling: navigating downstream. Cell. 2007, 129, 1261–1274. [CrossRef]
- Essers, M. A.; Weijzen, S.; de Vries-Smits, A. M.; Saarloos, I.; de Ruiter, N. D.; Bos, J. L.; Burgering, B. M. FOXO transcription factor activation by oxidative stress mediated by the small GTPase Ral and JNK. EMBO J. 2004, 23, 4802–4812. [CrossRef]
- van der Horst, A.; de Vries-Smits, A. M.; Brenkman, A. B.; van Triest, M. H.; van den Broek, N.; Colland, F.; Maurice, M. M.; Burgering, B. M. FOXO4 transcriptional activity is regulated by monoubiquitination and USP7/HAUSP. Nat Cell Biol. 2006, 8, 1064–1073. [CrossRef]
- Percie du Sert, N.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M. T.; Baker, M.; Browne, W. J.; Clark, A.; Cuthill, I. C.; Dirnagl, U.; Emerson, M.; et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. PLoS Biol. 2020, 18, e3000410. [CrossRef]
- Xu, R.; Qin, N.; Xu, X.; Sun, X.; Chen, X.; Zhao, J. Inhibitory effect of SLIT2 on granulosa cell proliferation mediated by the CDC42-PAKs-ERK1/2 MAPK pathway in the prehierarchical follicles of the chicken ovary. Sci Rep. 2018, 8, 9168. [CrossRef]
- Gilchrist, R. B.; Ritter, L. J.; Myllymaa, S.; Kaivo-Oja, N.; Dragovic, R. A.; Hickey, T. E.; Ritvos, O.; Mottershead, D. G. Molecular basis of oocyte-paracrine signalling that promotes granulosa cell proliferation. J Cell Sci. 2006, 119, 3811-21. 3811–3821. [CrossRef]
- Woodruff, T. K.; Shea, L. D. The role of the extracellular matrix in ovarian follicle development. Reprod Sci. 2007, 14, 6-10. [CrossRef]
- Dias, J. A.; Campo, B.; Weaver, B. A.; Watts, J.; Kluetzman, K.; Thomas, R. M.; Bonnet, B.; Mutel, V.; Poli, S. M. Inhibition of follicle-stimulating hormone-induced preovulatory follicles in rats treated with a nonsteroidal negative allosteric modulator of follicle-stimulating hormone receptor. Biol Reprod. 2014, 90, 19. [CrossRef]
- Rimon-Dahari, N.; Yerushalmi-Heinemann, L.; Alyagor, L.; Dekel, N. Ovarian Folliculogenesis. Results Probl Cell Differ. 2016, 58, 167-90. [CrossRef]
- Ulloa-Aguirre, A.; Zariñán, T.; Pasapera, A. M.; Casas-González, P.; Dias, J. A. Multiple facets of follicle-stimulating hormone receptor function. Endocrine. 2007, 32, 251-63. [CrossRef]
- You, S.; Bridgham, J. T.; Foster, D. N.; Johnson, A. L. Characterization of the chicken follicle-stimulating hormone receptor (cFSH-R) complementary deoxyribonucleic acid, and expression of cFSH-R messenger ribonucleic acid in the ovary. Biol Reprod. 1996, 55, 1055-62. [CrossRef]
- Kim, D.; Ocón-Grove, O.; Johnson, A. L. Bone morphogenetic protein 4 supports the initial differentiation of hen (Gallus gallus) granulosa cells. Biol Reprod. 2013, 88, 161. [CrossRef]
- Li GY. Regulating effect of FSH on apoptosis, secretion of cytokines and genes expression by FSHR/cAMP/Foxo3a of yak follicular granulosa cells. Lanzhou: Doctoral Dissertation of Gansu Agricultural University. 2017 (in Chinese).
- Zheng MX. The research of follicle stimulating hormone regulates the expression of stem cell factor in granulosa cells to promote the follicular development. Yinchuan: Master Dissertation of Ningxia Medical University. 2017 (in Chinese).
- Wang YY. Effects of FSH on gene expression related to steroidogenesis in bovine follicles cultured in vitro. Beijing: Master Dissertation of the Institute of Animal Science of Chinese Academy of Agricultural Sciences. 2019 (in Chinese).
- Chen X. Study on difference of ovulation effect induced by FSH in sheep. Hohhot: Master Dissertation of Inner Mongolia University. 2021 (in Chinese).
- Link W. Introduction to FOXO Biology. Methods Mol Biol. 2019, 1890, 1-9. [CrossRef]
- Zhu, M.; Miao, S.; Zhou, W.; Elnesr, S. S.; Dong, X.; Zou, X. Corrigendum to: “MAPK, AKT/FoxO3a and mTOR pathways are involved in cadmium regulating the cell cycle, proliferation and apoptosis of chicken follicular granulosa cells” [Ecotoxicol. Environ. Saf. 214 (2021) 112091]. Ecotoxicol Environ Saf. 2021, 214, 112091. [CrossRef]
- Chen, Y.; Xi, Y.; Li, M.; Wu, Y.; Yan, W.; Dai, J.; Wu, M.; Ding, W.; Zhang, J.; Zhang, F.; Zhou, S.; Wang, S. Maternal exposure to PM2.5 decreases ovarian reserve in neonatal offspring mice through activating PI3K/AKT/FoxO3a pathway and ROS-dependent NF-κB pathway. Toxicology. 2022, 481, 153352. [CrossRef]
- Kong, C.; Su, J.; Wang, Q.; Liu, K.; Fu, R.; Sui, S. Signaling pathways of Periplaneta americana peptide resist H2O2-induced apoptosis in pig-ovary granulosa cells through FoxO1. Theriogenology. 2022, 183, 108-19. [CrossRef]
- Ting, A. Y.; Zelinski, M. B. Characterization of FOXO1, 3 and 4 transcription factors in ovaries of fetal, prepubertal and adult rhesus macaques. Biol Reprod. 2017, 96, 1052-9. [CrossRef]
- Liu, Y.; Ao, X.; Ding, W.; Ponnusamy, M.; Wu, W.; Hao, X.; Yu, W.; Wang, Y.; Li, P.; Wang, J. Critical role of FOXO3a in carcinogenesis. Mol Cancer. 2018, 17, 104. [CrossRef]
- Sun, L.; Liu, J.; Bao, D.; Hu, C.; Zhao, Y.; Chen, S. Progress in the study of FOXO3a interacting with microRNA to regulate tumourigenesis development. Front Oncol. 2023, 13, 1293968. [CrossRef]
- Brownawell, A. M.; Kops, G. J.; Macara, I. G.; Burgering, B. M. Inhibition of nuclear import by protein kinase B (Akt) regulates the subcellular distribution and activity of the forkhead transcription factor AFX. Mol Cell Biol. 2001, 21, 3534-46. [CrossRef]
- Rena, G.; Woods, Y. L.; Prescott, A. R.; Peggie, M.; Unterman, T. G.; Williams, M. R.; Cohen, P. Two novel phosphorylation sites on FKHR that are critical for its nuclear exclusion. EMBO J. 2002, 21, 2263-71. [CrossRef]
- Fukuda, M.; Asano, S.; Nakamura, T.; Adachi, M.; Yoshida, M.; Yanagida, M.; Nishida, E. CRM1 is responsible for intracellular transport mediated by the nuclear export signal. Nature. 1997, 390, 308-11. [CrossRef]
- Kudo, N.; Wolff, B.; Sekimoto, T.; Schreiner, E. P.; Yoneda, Y.; Yanagida, M.; Horinouchi, S.; Yoshida, M. Leptomycin B inhibition of signal-mediated nuclear export by direct binding to CRM1. Exp Cell Res. 1998, 242, 540-47. [CrossRef]
- Watanabe, M.; Fukuda, M.; Yoshida, M.; Yanagida, M.; Nishida, E. Involvement of CRM1, a nuclear export receptor, in mRNA export in mammalian cells and fission yeast. Genes Cells.1999, 4, 291-97. [CrossRef]
- Vogt, P. K.; Jiang, H.; Aoki, M. Triple layer control: phosphorylation, acetylation and ubiquitination of FOXO proteins. Cell Cycle. 2005, 4, 908-13. [CrossRef]
- Obsilova, V.; Vecer, J.; Herman, P.; Pabianova, A.; Sulc, M.; Teisinger, J.; Boura, E.; Obsil, T. 14-3-3 Protein interacts with nuclear localization sequence of forkhead transcription factor FoxO4. Biochemistry. 2005, 44, 11608-17. [CrossRef]
- Nielsen, M. D.; Luo, X.; Biteau, B.; Syverson, K.; Jasper, H. 14-3-3 Epsilon antagonizes FoxO to control growth, apoptosis and longevity in Drosophila. Aging Cell. 2008, 7, 688-99. [CrossRef]
- Zanella, F.; Rosado, A.; García, B.; Carnero, A.; Link, W. Chemical genetic analysis of FOXO nuclear-cytoplasmic shuttling by using image-based cell screening. Chembiochem. 2008, 9, 2229-37. [CrossRef]
- Mutka, S. C.; Yang, W. Q.; Dong, S. D.; Ward, S. L.; Craig, D. A.; Timmermans, P. B.; Murli, S. Identification of nuclear export inhibitors with potent anticancer activity in vivo. Cancer Res. 2009, 69, 510-7. [CrossRef]
- Rahmani, K.; Dean, D. A. Leptomycin B alters the subcellular distribution of CRM1 (Exportin 1). Biochem Biophys Res Commun. 2017, 488, 253-8. [CrossRef]
- Li, Y.; Wang, Z.; Kong, D.; Li, R.; Sarkar, S. H.; Sarkar, F. H. Regulation of Akt/FOXO3a/GSK-3beta/AR signaling network by isoflavone in prostate cancer cells. J Biol Chem. 2008, 283, 27707-16. [CrossRef]
- Charvet, C.; Alberti, I.; Luciano, F.; Jacquel, A.; Bernard, A.; Auberger, P.; Deckert, M. Proteolytic regulation of Forkhead transcription factor FOXO3a by caspase-3-like proteases. Oncogene. 2003, 22, 4557-68. [CrossRef]
Figure 1.
The expression of FSHR mRNA in the undifferentiated granulosa cells regulated by FSH. The GCs from the hen ovarian prehierarchical follicles of 6-8 mm diameter were cultured at the varied concentrations of 0 ng/ml, 8 ng/ml, 10 ng/ml, and 12 ng/ml for 0 h, 8 h, 12 h, and 24 h, respectively. The data were presented as the means ± SEM from five replicate experiments. The bars with superscript symbol present the significant difference compared with control groups at **P < 0.01, * P < 0.05.
Figure 1.
The expression of FSHR mRNA in the undifferentiated granulosa cells regulated by FSH. The GCs from the hen ovarian prehierarchical follicles of 6-8 mm diameter were cultured at the varied concentrations of 0 ng/ml, 8 ng/ml, 10 ng/ml, and 12 ng/ml for 0 h, 8 h, 12 h, and 24 h, respectively. The data were presented as the means ± SEM from five replicate experiments. The bars with superscript symbol present the significant difference compared with control groups at **P < 0.01, * P < 0.05.
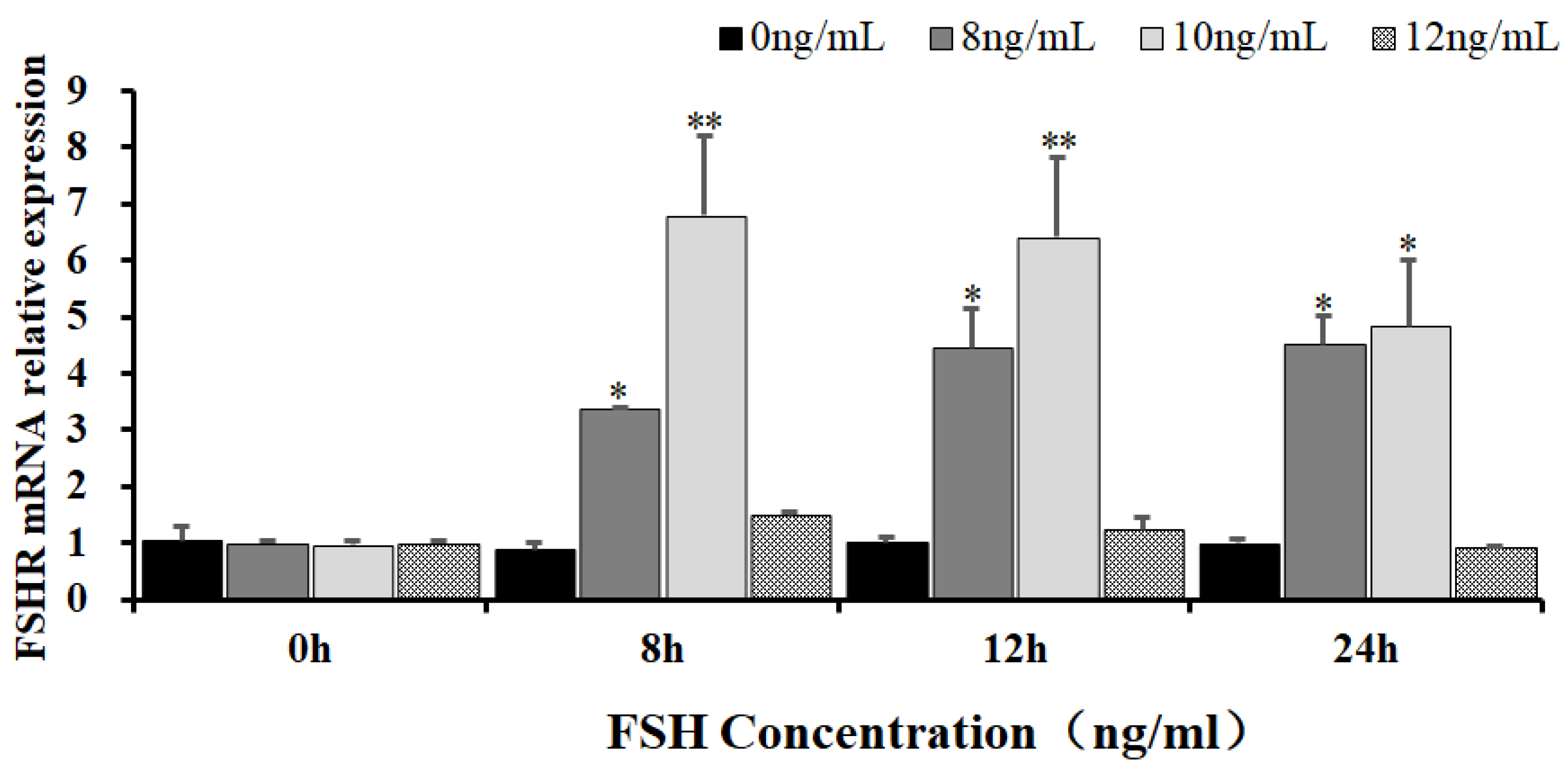
Figure 2.
Effects of FSH on the FoxO3/4 phosphorylation in the cultured GCs of the ovarian follicles. (A) The p-FOXO3 levels were determined by western blotting analysis in the GCs treated with or without the 10ng/ml FSH for 12 h in the presence or absence of LMB. (B) The p-FOXO4 levels were tested in the GCs under the treatment with or without the 10 ng/ml FSH for 12 h in the presence or absence of LMB. The data represent the mean ± SEM, (n = 5). Student’s t-test, **P < 0.01 and *P < 0.05.
Figure 2.
Effects of FSH on the FoxO3/4 phosphorylation in the cultured GCs of the ovarian follicles. (A) The p-FOXO3 levels were determined by western blotting analysis in the GCs treated with or without the 10ng/ml FSH for 12 h in the presence or absence of LMB. (B) The p-FOXO4 levels were tested in the GCs under the treatment with or without the 10 ng/ml FSH for 12 h in the presence or absence of LMB. The data represent the mean ± SEM, (n = 5). Student’s t-test, **P < 0.01 and *P < 0.05.
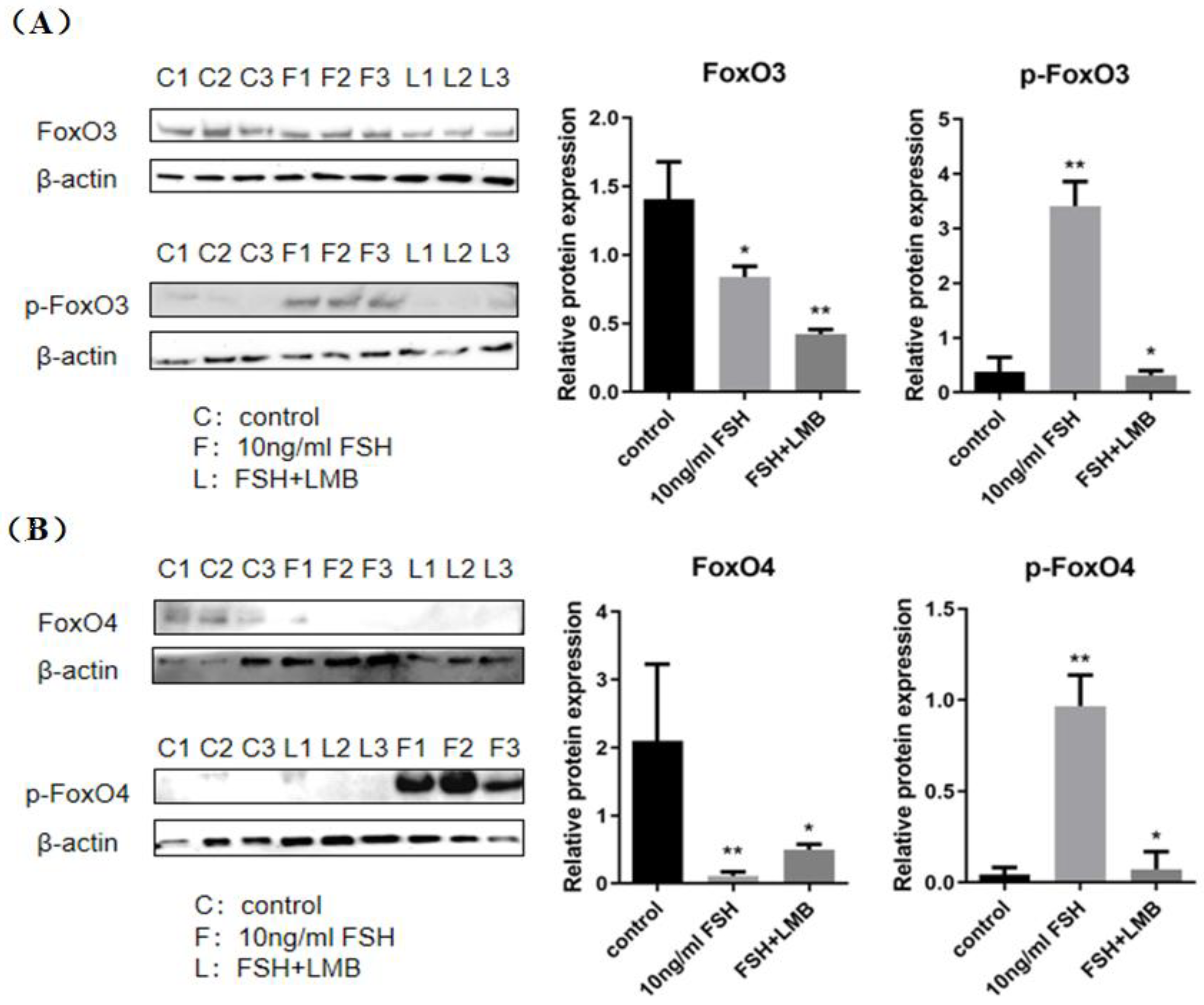
Figure 3.
FSH-induced the nuclear exclusion of FOXO3/4 in the cultured GCs of the ovarian follicles. (A) Immunofluorescence detection of FOXO3 subcellular localization in the GCs. The GCs were treated with or without the 10 ng/ml FSH for 12 h in the presence or absence of LMB, which were stained using goat anti-rabbit FOXO3 (green). The nucleus was stained with DAPI (blue). scale bar = 5 μm. (B) Immunofluorescence detection of FOXO4 subcellular localization in the GCs. FOXO4 (green) and nucleus stained with DAPI (blue). a. control, without treatment of FSH or LMB; b. treatment of the cells with 10 ng/ml FSH for 12 h; c. co-treatment of the cells with 10 ng/ml FSH plus LMB for 12 h. n = 3 independent experiments. Leica DMI8, 600X; scale bar = 2.5 μm.
Figure 3.
FSH-induced the nuclear exclusion of FOXO3/4 in the cultured GCs of the ovarian follicles. (A) Immunofluorescence detection of FOXO3 subcellular localization in the GCs. The GCs were treated with or without the 10 ng/ml FSH for 12 h in the presence or absence of LMB, which were stained using goat anti-rabbit FOXO3 (green). The nucleus was stained with DAPI (blue). scale bar = 5 μm. (B) Immunofluorescence detection of FOXO4 subcellular localization in the GCs. FOXO4 (green) and nucleus stained with DAPI (blue). a. control, without treatment of FSH or LMB; b. treatment of the cells with 10 ng/ml FSH for 12 h; c. co-treatment of the cells with 10 ng/ml FSH plus LMB for 12 h. n = 3 independent experiments. Leica DMI8, 600X; scale bar = 2.5 μm.
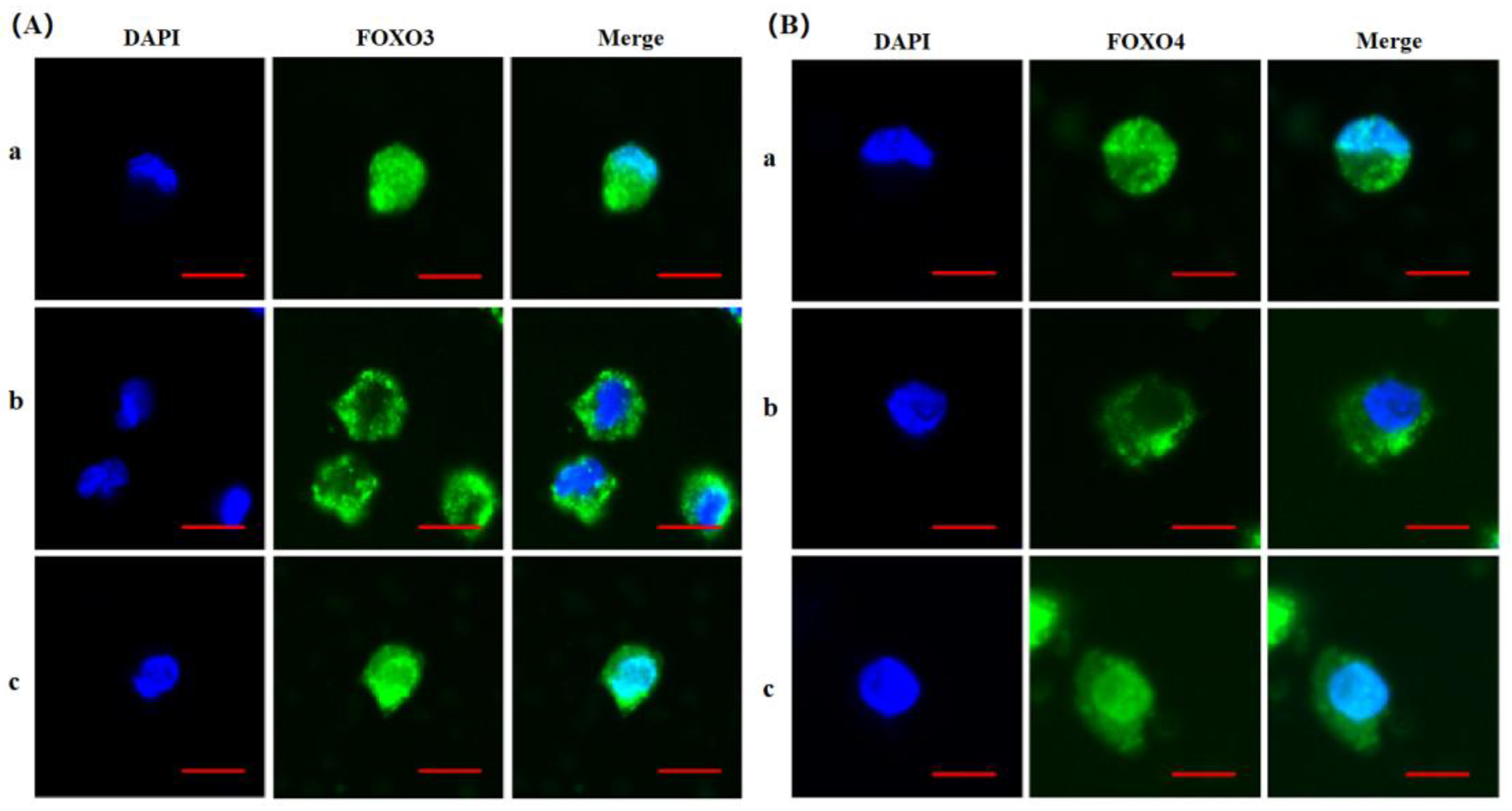
Figure 4.
FSH-induced expression of MYC, PCNA, StAR, CYP11A1, BCL-2, and CASP-3 genes in the cultured GCs. The data were presented as the means ± SEM from five replicate experiments, and bars with superscript symbol present the significant difference compared with control groups at **P < 0.01, * P < 0.05.
Figure 4.
FSH-induced expression of MYC, PCNA, StAR, CYP11A1, BCL-2, and CASP-3 genes in the cultured GCs. The data were presented as the means ± SEM from five replicate experiments, and bars with superscript symbol present the significant difference compared with control groups at **P < 0.01, * P < 0.05.
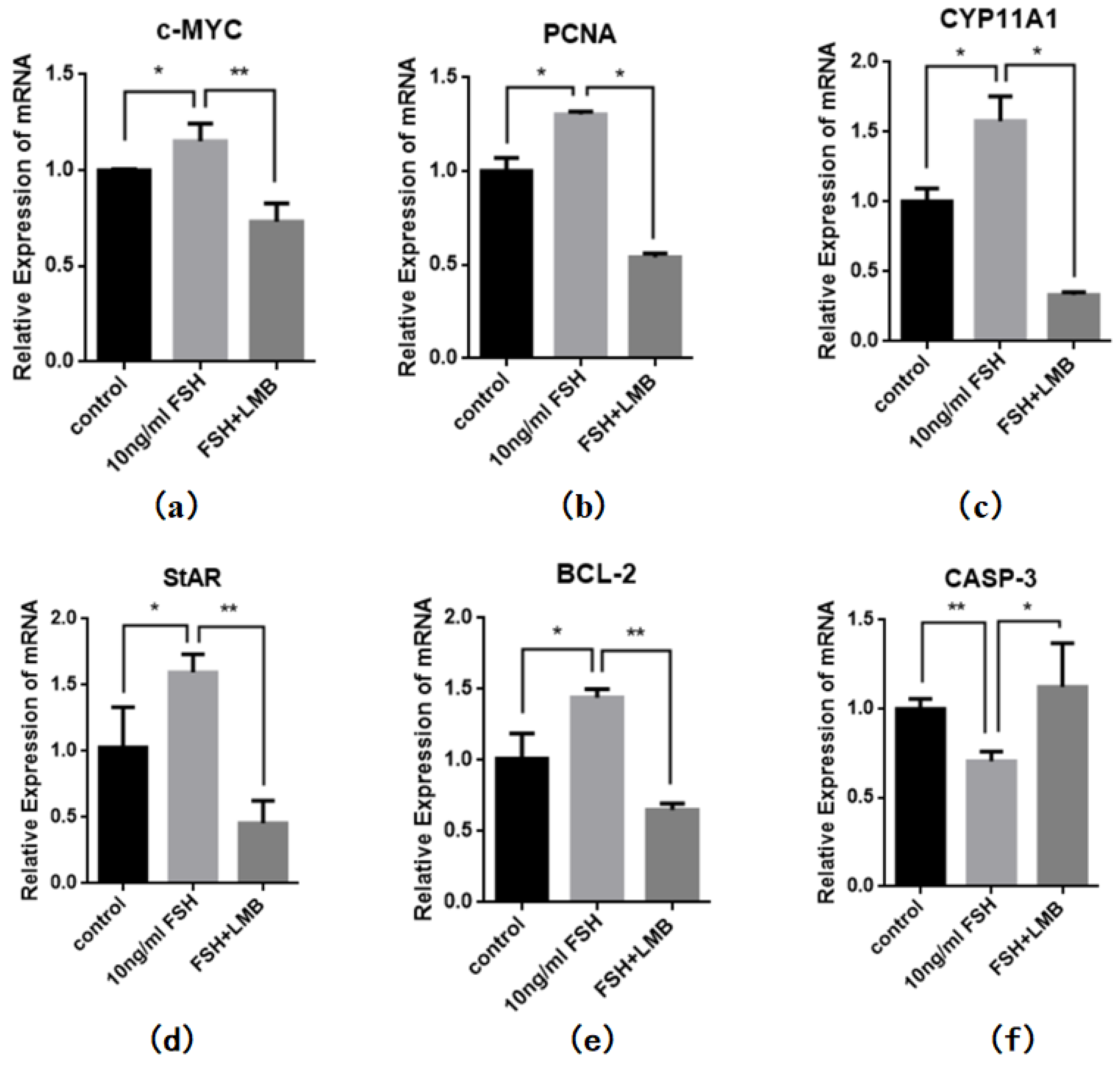
Figure 5.
FSH-induced nuclear exclusion of FoxO3/4 on proliferation and cell apoptosis of the cultured GCs. The data were presented as the means ± SEM from three replicate experiments, and bars with superscript symbol present the significant difference compared with control groups at * P < 0.05.
Figure 5.
FSH-induced nuclear exclusion of FoxO3/4 on proliferation and cell apoptosis of the cultured GCs. The data were presented as the means ± SEM from three replicate experiments, and bars with superscript symbol present the significant difference compared with control groups at * P < 0.05.
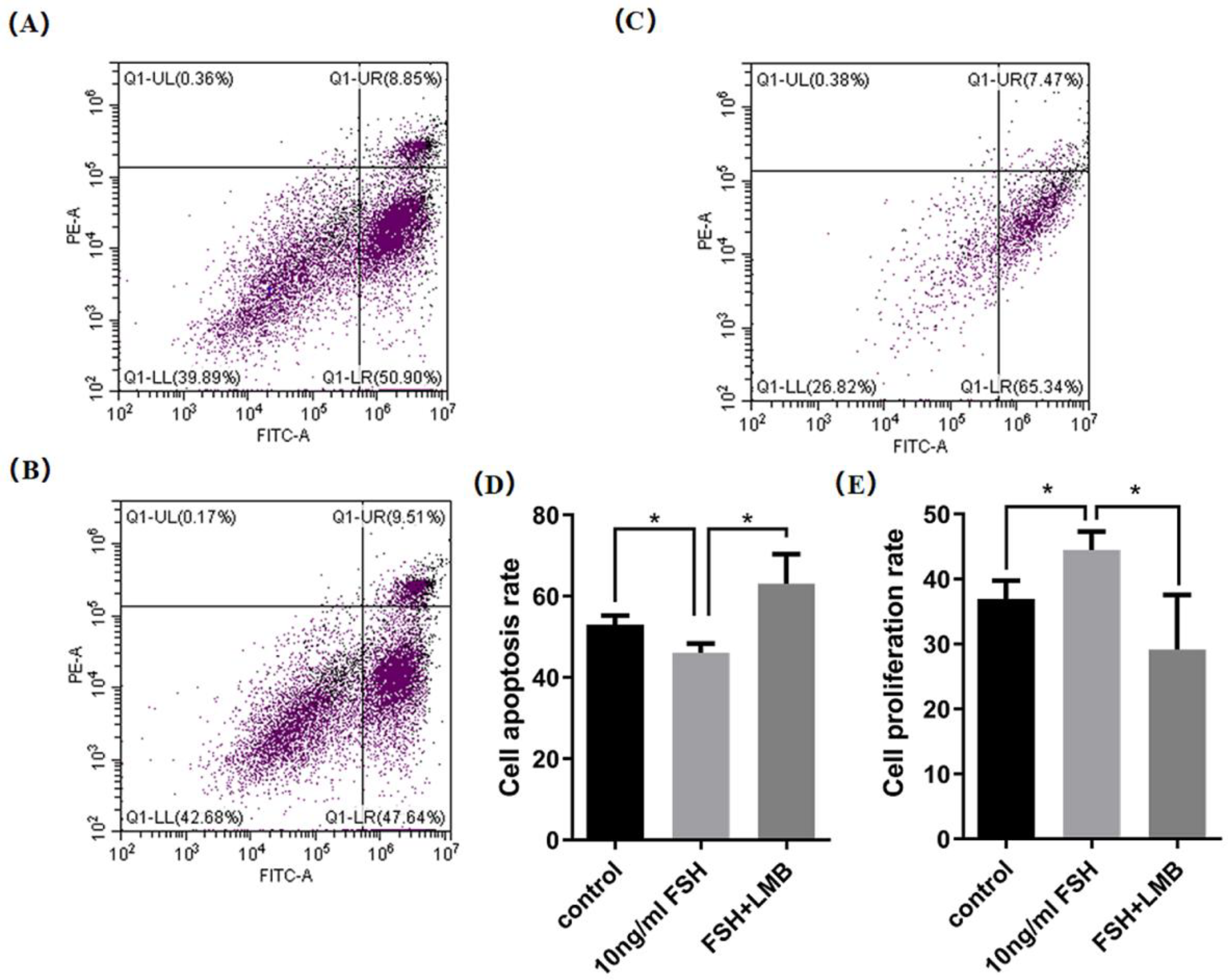

Table 1.
Primer pairs designed for quantitative real-time PCR analysis.
| Gene | Forward primer(5’-3’) | Reverse primer(5’-3’) | Accession No. | Size |
| FSHR | ATGTCTCCGGCAAAGCAAGA | AACGACTTCGTTGCACAAGC | NM_205079.1 | 147 bp |
| CASP-3 | ATTGAAGCAGACAGTGGACCAGATG | TGCGTTCCTCCAGGAGTAGTAGC | NM_204725.2 | 111 bp |
| PCNA | CTGAGGCGTGCTGGG | ATGGCGATGTTGCGG | NM_204170.3 | 133 bp |
| StAR | AGCAGATGGGCGACTGGAAC | GGGAGCACCGAACACTCACAA | NM_204686.2 | 147 bp |
| CYP11A1 | CCGCTTTGCCTTGGAGTCTGTG | ATGAGGGTGACGGCGTCGATGAA | NM_001001756.1 | 111 bp |
| c-MYC | GAGAACGACAAGAGGCGAAC | CGCCTCAACTGCTCTTTCTC | NM_001030952.2 | 211 bp |
| BCL-2 | CGCTACCAGAGGGAC | GAAGAAGGCGACGAT | NM_205339.3 | 135 bp |
| PDK1 | AGACATCCCGAGCTACACCT | CGCCTTGGAAGTATTGTGCG | NM_001031352.4 | 81bp |
| SGK3 | TGCGTCCAGGAATCAGTCTCAC | AAGTCTGCTTTGCCGATCTTTCTC | NM_001030940.2 | 74bp |
| 18s rRNA | TAGTTGGTGGAGCGATTTGTCT | CGGACATCTAAGGGCATCACA | AF173612.1 | 169 bp |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



